Fósiles de rastro mandados a foronídeos |
|
Ni Diorygma ni Eodiorygma son foronídeos ** by Christian C. Emig The trace fossil Diorygma1 Biernat, 1961 manifested as a tubular protuberance formed by deposition of the secondary shell around a vermifor Figure from Biernat (1961) >> Each protuberance in Diorygma encloses two contiguous tubes interpreted by MacKinnon & Biernat (1970) as the separated anal and oral tubes of the U-shaped digestive tract of phoronids : in fact such a construction has never been described nor considered because anatomically impossible according to the larval metamorphosis (see Emig, 1982 and references herein). Thus, if right that must be demonstrated, the reconstruction of Mackinnon & Biernat (1970) would appear as an anatomical novelty. Bassett et al. (2004) write about the phoronids in relation with a similar structure by Eodiorygma Bassett, Popov & Holmer, 2004: “in some groups are protected distally around the lophophore by a pair of collar folds (Emig, 1979)” but this is a misinterpretation of the single invagination occurring in Phoronopsis as originally described by Gilchist (1907) – see Taxonomy.
Figure from MacKinnon & Biernat (1970) >> Furthermore, some phoronid species are boring or encrusting, but their tube as in the dwelling phoronid species is chitinous and covered with various detritic particles (see Emig, 1973, 1982; Pourreau, 1979). One must pointed again that to refer a fossil tube or boring to a phoronid is very difficult when not impossible to ascertain. A consequence is that the geological origin of the Phoronida remains unknown, perhaps before the Cambrian. The Phoronida web site provides answer to all questions about this zoological group and an exhaustive bibliography year after year and for each species, concerning adult and its actinotroch larva, as well as drawing and photographies of many of the species. Nevertheless, there are, at least among Recent marine groups, several organisms able to create such trace with double aperture as figured for the trace fossil Eodiorygma or Diorygma. By the way if it is a suspension-feeding organism a tentacular apparatus is not need to explain the occurrence within a brachiopod shell. Suspension feeders or sestonophages require either a sufficiently high density of digestible suspended particles or a sufficient supply through water movement. They are grossly classified into two major categories: (i) species which exploit existing water movement, i.e., which are unable to generate current on their own; (ii) species which actively produce water movement (Pérès, 1982). Thus, widespread mechanisms of suspension feeding exist among the zoological groups. 1 Diorygma is also a genus of lichenized fungi in the family Graphidaceae.
** Publicado en: Emig C. C., 2010. Fossil Phoronida and their inferred ichnotaxa. Carnets de Géologie / Notebooks on Geology, Letter 2010/03 (CG2010_L03), 5 p., 4 fig.
|
|
|
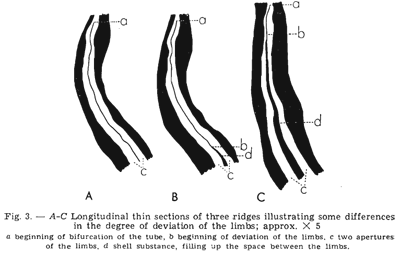 m organism settled inside the mantle cavity of the atrypide brachiopod Desquamatia subzonata Biernat, 1961. Although interpreted originally as a result of parasitic boring through the outside of the shell (Biernat, 1961), the relationships between Desquamatia Biernat, 1961 and Diorygma Biernat, 1961 were interpreted subsequently as being strictly host-specific, suggesting instead that Diorygma was probably a phoronid inhabiting the mantle cavity of the brachiopod and sharing its filter-feeding strategy (MacKinnon & Biernat, 1970).
m organism settled inside the mantle cavity of the atrypide brachiopod Desquamatia subzonata Biernat, 1961. Although interpreted originally as a result of parasitic boring through the outside of the shell (Biernat, 1961), the relationships between Desquamatia Biernat, 1961 and Diorygma Biernat, 1961 were interpreted subsequently as being strictly host-specific, suggesting instead that Diorygma was probably a phoronid inhabiting the mantle cavity of the brachiopod and sharing its filter-feeding strategy (MacKinnon & Biernat, 1970).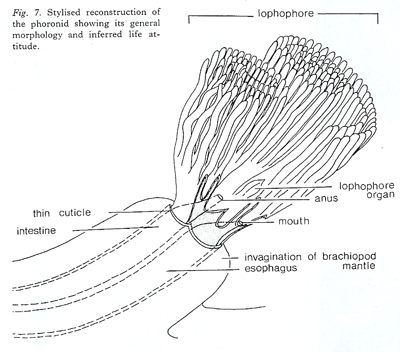 I have in mind that most of the authors, perhaps all, using the Phoronida as examples have never seen a living individual. The stylised reconstruction of the phoronid proposed by MacKinnon & Biernat (1970) sets a lophophore in two parts the coils being anal and the tentacles at mouth level ventral. More the lophophore is represented by a double row of tentacles that is completely wrong because the lophophore of all lophophorates (Phoronida, Bryozoa and Brachiopoda) shows a single row of tentacles. Both interpretations do not fit with the morphology and anatomy in the Phoronida. By the way this reconstruction does not appear confident with longitudinal sections figured by Biernat (1961).
I have in mind that most of the authors, perhaps all, using the Phoronida as examples have never seen a living individual. The stylised reconstruction of the phoronid proposed by MacKinnon & Biernat (1970) sets a lophophore in two parts the coils being anal and the tentacles at mouth level ventral. More the lophophore is represented by a double row of tentacles that is completely wrong because the lophophore of all lophophorates (Phoronida, Bryozoa and Brachiopoda) shows a single row of tentacles. Both interpretations do not fit with the morphology and anatomy in the Phoronida. By the way this reconstruction does not appear confident with longitudinal sections figured by Biernat (1961).