![]()
![]()
![]() Abundant worm borings were found in
some brachiopod shells (Clitambonites, Estlandia, Nicolella)
from the Ordovician (Caradoc) oil shale in North Estonia. 9 of 21 brachiopod genera
(43 %) have been bored. Excluding the size and thickness of valves, no common morphological feature discriminates the brachiopods with borings from those without them. The Trypanites are host-specific, and the frequency of bored valves varies from 6.5 % in Bekkerina to 51 % in Estlandia. The worm larvae preferred hosts with thick lamellose shells, such as those of the clitambonitids, especially Clitambonites schmidti. The boring organisms were size-selective; they preferred large adult specimens. The majority of the borings are oriented, and living hosts were preferred to dead shells. Some clitambonitidine brachiopods,
like C. schmidti tolerated a large number of the borer's shafts in their valves. Few bored valves have the blister-like shell-repair structures in their interior.
Abundant worm borings were found in
some brachiopod shells (Clitambonites, Estlandia, Nicolella)
from the Ordovician (Caradoc) oil shale in North Estonia. 9 of 21 brachiopod genera
(43 %) have been bored. Excluding the size and thickness of valves, no common morphological feature discriminates the brachiopods with borings from those without them. The Trypanites are host-specific, and the frequency of bored valves varies from 6.5 % in Bekkerina to 51 % in Estlandia. The worm larvae preferred hosts with thick lamellose shells, such as those of the clitambonitids, especially Clitambonites schmidti. The boring organisms were size-selective; they preferred large adult specimens. The majority of the borings are oriented, and living hosts were preferred to dead shells. Some clitambonitidine brachiopods,
like C. schmidti tolerated a large number of the borer's shafts in their valves. Few bored valves have the blister-like shell-repair structures in their interior.
![]() Ordovician;
Estonia; oil shale; Brachiopoda; Trypanites; palaeoecology.
Ordovician;
Estonia; oil shale; Brachiopoda; Trypanites; palaeoecology.
O. (2005).- The distribution of worm borings in brachiopod shells from the Caradoc Oil Shale of Estonia.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2005/03 (CG2005_A03)
![]() Distribution des perforations de vers dans les coquilles de brachiopodes des
schistes bitumineux du Caradoc d'Estonie.- Des perforations dues à des vers
sont observées en abondance sur les coquilles de quelques brachiopodes (Clitambonites,
Estlandia, Nicolella) des schistes bitumineux caradociens du Nord
de l'Estonie. Ainsi, 9 des 21 genres de brachiopodes (43 %) étudiés ont des
valves perforées. La présence ou l'absence des perforations est liée à
taille et l'épaisseur des valves. Les autres traits morphologiques semblent
n'avoir aucune importance. Dans l'ensemble, en effet, les larves des vers
marquent une préférence pour des coquilles épaisses et lamelleuses, comme
celles des clitambonitidés, particulièrement Clitambonites schmidti. On
note, cependant que les représentants de l'ichnogenre Trypanites
marquent quelques préférences spécifiques, ainsi leur fréquence varie de 6.5
% chez Bekkerina à 51 % chez Estlandia. La majorité des perforations
sont orientées et les coquilles d'individus vivants étaient préférées aux
coquilles vides. Certains brachiopodes clitambonitidés, comme Clitambonites
schmidti, ont toléré un grand nombre de perforations dans leurs valves. Seul
un petit nombre de valves trouées ont révélées des cals de réparation de la
coquille sur leur face interne.
Distribution des perforations de vers dans les coquilles de brachiopodes des
schistes bitumineux du Caradoc d'Estonie.- Des perforations dues à des vers
sont observées en abondance sur les coquilles de quelques brachiopodes (Clitambonites,
Estlandia, Nicolella) des schistes bitumineux caradociens du Nord
de l'Estonie. Ainsi, 9 des 21 genres de brachiopodes (43 %) étudiés ont des
valves perforées. La présence ou l'absence des perforations est liée à
taille et l'épaisseur des valves. Les autres traits morphologiques semblent
n'avoir aucune importance. Dans l'ensemble, en effet, les larves des vers
marquent une préférence pour des coquilles épaisses et lamelleuses, comme
celles des clitambonitidés, particulièrement Clitambonites schmidti. On
note, cependant que les représentants de l'ichnogenre Trypanites
marquent quelques préférences spécifiques, ainsi leur fréquence varie de 6.5
% chez Bekkerina à 51 % chez Estlandia. La majorité des perforations
sont orientées et les coquilles d'individus vivants étaient préférées aux
coquilles vides. Certains brachiopodes clitambonitidés, comme Clitambonites
schmidti, ont toléré un grand nombre de perforations dans leurs valves. Seul
un petit nombre de valves trouées ont révélées des cals de réparation de la
coquille sur leur face interne.
![]() Ordovicien ;
Estonie ; schistes bitumineux ; Brachiopoda; Trypanites ; paléoécologie.
Ordovicien ;
Estonie ; schistes bitumineux ; Brachiopoda; Trypanites ; paléoécologie.
![]()
![]() It appears that the oldest macroborings in the world are specimens of Trypanites: small, simple holes reported in localized Early Cambrian archeocyathid reefs in Labrador ( et alii,
1977; et alii, 1978). Essentially no macroborings of any kind are known during the remainder of the Cambrian Period. The next oldest macroborings, found in carbonate hardgrounds, are of Early Ordovician age ( & ,
1993; & ,
2001; et alii, 2002; & ,
2003). Macroborings in brachiopod shells are known from the Ordovician (,
1969; , 1976; & ,
1980; , 2004).
It appears that the oldest macroborings in the world are specimens of Trypanites: small, simple holes reported in localized Early Cambrian archeocyathid reefs in Labrador ( et alii,
1977; et alii, 1978). Essentially no macroborings of any kind are known during the remainder of the Cambrian Period. The next oldest macroborings, found in carbonate hardgrounds, are of Early Ordovician age ( & ,
1993; & ,
2001; et alii, 2002; & ,
2003). Macroborings in brachiopod shells are known from the Ordovician (,
1969; , 1976; & ,
1980; , 2004).
![]() In Baltoscandia borings in brachiopods of medial Late Ordovian age have been recorded from the Mjøsa Limestone (Rakvere to Nabala stages) of Southern Norway, but the borings of Vermiforichnus occur very rarely there. The Mjøsa Limestone was deposited during the middle Upper Ordovician (Upper Caradoc) at the western margin of the extensive Lower Palaeozoic epicontinental sea of Baltoscandia
(Fig. 1
In Baltoscandia borings in brachiopods of medial Late Ordovian age have been recorded from the Mjøsa Limestone (Rakvere to Nabala stages) of Southern Norway, but the borings of Vermiforichnus occur very rarely there. The Mjøsa Limestone was deposited during the middle Upper Ordovician (Upper Caradoc) at the western margin of the extensive Lower Palaeozoic epicontinental sea of Baltoscandia
(Fig. 1 ![]() ). It consists predominantly of carbonate sediments deposited in shallow to very shallow marine environments ( and ,
1980).
). It consists predominantly of carbonate sediments deposited in shallow to very shallow marine environments ( and ,
1980).
![]() In the Oil Shale basin worm borings are known in the bryozoan colonies of the Leningrad area (Russia). These borings were originally identified as
Hicetes (, 1928), but they may be Trypanites (personal observations; see & ,
1982).
In the Oil Shale basin worm borings are known in the bryozoan colonies of the Leningrad area (Russia). These borings were originally identified as
Hicetes (, 1928), but they may be Trypanites (personal observations; see & ,
1982).
![]() In the oil shales of Estonia abundant borings in brachiopods are slightly older (lowermost Caradoc, see
Fig. 2
In the oil shales of Estonia abundant borings in brachiopods are slightly older (lowermost Caradoc, see
Fig. 2 ![]() )
than the Trypanites described from Wales and Norway, but closely resemble them. During Kukruse (Early Caradoc) time in the Baltoscandia Basin
(Fig. 1
)
than the Trypanites described from Wales and Norway, but closely resemble them. During Kukruse (Early Caradoc) time in the Baltoscandia Basin
(Fig. 1 ![]() ), light-brown organic-rich muds formed oil shale interbeds in calcareous sediments ( & ,
1997). The oil shale is composed of almost equal portions of carbonates, clayey siliciclastics and organic matter. The existence in the Estonian oil shales of a rich and diverse marine bottom fauna (,
1970, p. 172) together with a very low pyrite content suggest that conditions in the bottom waters were fully oxic ( & ,
1990). Most of the organic matter is conjectured to have been derived from algal mats ( & ,
1994), that covered extensive tidal flat areas. Their organic matter was transported to shallow subtidal environments ( et alii,
1988) during the Late Llanvirn-Early Caradoc regression ( et alii,
1988).
), light-brown organic-rich muds formed oil shale interbeds in calcareous sediments ( & ,
1997). The oil shale is composed of almost equal portions of carbonates, clayey siliciclastics and organic matter. The existence in the Estonian oil shales of a rich and diverse marine bottom fauna (,
1970, p. 172) together with a very low pyrite content suggest that conditions in the bottom waters were fully oxic ( & ,
1990). Most of the organic matter is conjectured to have been derived from algal mats ( & ,
1994), that covered extensive tidal flat areas. Their organic matter was transported to shallow subtidal environments ( et alii,
1988) during the Late Llanvirn-Early Caradoc regression ( et alii,
1988).
![]() The Estonian oil shale taphocoenosis includes a mixture of invertebrate remains from several different but adjacent parts of the basin. However, the extraordinarily well-preserved colonies of large, fragile bryozoa (e.g.,
Chasmatoporella) along with unweathered brachiopod shells, and rare, almost pristine
trilobites, were neither transported nor affected by post mortem hydrodynamic movements. Nevertheless, many brachiopods (e.g.,
Clitambonites, Estlandia, Nicolella, Cyrtonotella, Kullervo, Glossorthis) are preserved as discrete valves and presumably were not buried
in situ. Strophomenids and Porambonites are usually preserved as complete shells
presumably because of the structures of their articulations. The taphocoenosis here is comprised mainly of the remains of brachiopods from nearby areas, and a small amount of weathered, obviously allochthonous shell material.
The Estonian oil shale taphocoenosis includes a mixture of invertebrate remains from several different but adjacent parts of the basin. However, the extraordinarily well-preserved colonies of large, fragile bryozoa (e.g.,
Chasmatoporella) along with unweathered brachiopod shells, and rare, almost pristine
trilobites, were neither transported nor affected by post mortem hydrodynamic movements. Nevertheless, many brachiopods (e.g.,
Clitambonites, Estlandia, Nicolella, Cyrtonotella, Kullervo, Glossorthis) are preserved as discrete valves and presumably were not buried
in situ. Strophomenids and Porambonites are usually preserved as complete shells
presumably because of the structures of their articulations. The taphocoenosis here is comprised mainly of the remains of brachiopods from nearby areas, and a small amount of weathered, obviously allochthonous shell material.
![]() The borings were cleaned using ultrasound and brushes. Casts of some of them were made using the universal instant adhesive "Super Attak", Loctite Ltd. All borings were counted in relation to their occurrence on a single valve. They were measured and digitally photographed using a binocular light microscope. To determine the importance of the characteristics of the valve in relation to its role as a substrate for boring organisms the following measurements were made on each specimen: size (length and width), thickness, structure (lamellose or homogeneous), and surface relief. Specimens of Estlandia marginata, Clitambonites schmidti and C. squamatus were studied to determine the average frequency of borings per discrete valve (pedicle and brachial valves counted separately) and their orientation noted. Conclusions are based on measurements of least six valves (both ventral and dorsal) of a particular taxon.
The borings were cleaned using ultrasound and brushes. Casts of some of them were made using the universal instant adhesive "Super Attak", Loctite Ltd. All borings were counted in relation to their occurrence on a single valve. They were measured and digitally photographed using a binocular light microscope. To determine the importance of the characteristics of the valve in relation to its role as a substrate for boring organisms the following measurements were made on each specimen: size (length and width), thickness, structure (lamellose or homogeneous), and surface relief. Specimens of Estlandia marginata, Clitambonites schmidti and C. squamatus were studied to determine the average frequency of borings per discrete valve (pedicle and brachial valves counted separately) and their orientation noted. Conclusions are based on measurements of least six valves (both ventral and dorsal) of a particular taxon.
![]() A total of 29 articulate brachiopod genera (about 90 species) are recorded in the oil shales of the Kukruse Stage in northeastern Estonia ( & ,
1997); 21 genera (about 30 species) were examined for this study of the distribution of borings (Table 1
A total of 29 articulate brachiopod genera (about 90 species) are recorded in the oil shales of the Kukruse Stage in northeastern Estonia ( & ,
1997); 21 genera (about 30 species) were examined for this study of the distribution of borings (Table 1 ![]() ). The following taxa had one or more borings in at least a single valve: Estlandia marginata, Clitambonites schmidti, Clitambonites squamatus, Kullervo ssp., Glossorthis ssp., Nicolella ssp., Bekkerina ssp., Bilobia ssp., Cyrtonotella ssp., and Septomena ssp. Borings were not seen in Oxoplecia ssp., Platystrophia ssp., Paucicrura ssp., Porambonites ssp., Leptestia ssp., Leptelloidea ssp., Estonomena ssp., Sowerbyella ssp., Actinomena ssp., Bicuspina ssp. and Kierulfina ssp. A craniate brachiopod
Orthisocrania was studied too, but no borings were discovered
(Table 1
). The following taxa had one or more borings in at least a single valve: Estlandia marginata, Clitambonites schmidti, Clitambonites squamatus, Kullervo ssp., Glossorthis ssp., Nicolella ssp., Bekkerina ssp., Bilobia ssp., Cyrtonotella ssp., and Septomena ssp. Borings were not seen in Oxoplecia ssp., Platystrophia ssp., Paucicrura ssp., Porambonites ssp., Leptestia ssp., Leptelloidea ssp., Estonomena ssp., Sowerbyella ssp., Actinomena ssp., Bicuspina ssp. and Kierulfina ssp. A craniate brachiopod
Orthisocrania was studied too, but no borings were discovered
(Table 1 ![]() ).
).
![]() Borings occur in specimens of nine of the 21 brachiopod genera examined. Their frequencies range from 6.5 % in Bekkerina ssp. to 51 % in Estlandia marginata. The two genera that occur in large numbers in oil shale, Estlandia and Clitambonites, have the highest frequency of borings. The two were selected for detailed studies of site specificity and the morphology of
borings, and were chosen as a control group for the comparison of the boring frequency in the brachiopod fauna of the oil shale with that of stratigraphically older and younger brachiopods from the same area in the Baltoscandian Ordovician Basin (i.e., the North Estonian confacies belt of ,
1973).
Borings occur in specimens of nine of the 21 brachiopod genera examined. Their frequencies range from 6.5 % in Bekkerina ssp. to 51 % in Estlandia marginata. The two genera that occur in large numbers in oil shale, Estlandia and Clitambonites, have the highest frequency of borings. The two were selected for detailed studies of site specificity and the morphology of
borings, and were chosen as a control group for the comparison of the boring frequency in the brachiopod fauna of the oil shale with that of stratigraphically older and younger brachiopods from the same area in the Baltoscandian Ordovician Basin (i.e., the North Estonian confacies belt of ,
1973).
![]() Valves are considered to be strongly weathered if their shape is obviously rounded by wear and they lack the original sculpture and articulation. Rounded cardinal extremities or flattened ribs are considered to be the result of weathering. About 8 per cent of valves with borings are strongly weathered (most of these are the resistant ventral valves of Clitambonites squamatus). Borings in weathered valves are sparse; the majority were probably made on undamaged fresh valves. However, strongly weathered valves of Clitambonites and Estlandia have never been found without borings, but about one-third of these strongly weathered valve were not rebored after having been weathered. Large thin-shelled strophomenids (Leptestia and Estonomena), Leptelloidea and Sowerbyella do not
contain borings. They too were unweathered.
Valves are considered to be strongly weathered if their shape is obviously rounded by wear and they lack the original sculpture and articulation. Rounded cardinal extremities or flattened ribs are considered to be the result of weathering. About 8 per cent of valves with borings are strongly weathered (most of these are the resistant ventral valves of Clitambonites squamatus). Borings in weathered valves are sparse; the majority were probably made on undamaged fresh valves. However, strongly weathered valves of Clitambonites and Estlandia have never been found without borings, but about one-third of these strongly weathered valve were not rebored after having been weathered. Large thin-shelled strophomenids (Leptestia and Estonomena), Leptelloidea and Sowerbyella do not
contain borings. They too were unweathered.
![]() The shells of molluscs may have been substrates for boring organisms, but in the strata studied their fossils are mainly internal moulds. Several well preserved echinoderm specimens (Echinosphaerites) were examined for borings, but none were found. Bryozoans too lack borings like those in brachiopods,
but some bryozoans are covered with scratch marks that obviously were created differently
from the borings in brachiopods. Borings similar to Trypanites are recorded in the Monticulipora bryozoans of the Haljala and Kukruse stages of the Leningrad area of the Oil Shale Basin of northeastern Russia (,
1928).
The shells of molluscs may have been substrates for boring organisms, but in the strata studied their fossils are mainly internal moulds. Several well preserved echinoderm specimens (Echinosphaerites) were examined for borings, but none were found. Bryozoans too lack borings like those in brachiopods,
but some bryozoans are covered with scratch marks that obviously were created differently
from the borings in brachiopods. Borings similar to Trypanites are recorded in the Monticulipora bryozoans of the Haljala and Kukruse stages of the Leningrad area of the Oil Shale Basin of northeastern Russia (,
1928).
![]() The percentage of borings in brachiopod populations both preceding and succeeding the oil shale fauna seem to be remarkably lower. The collection of Estlandia (13 specimens) from the Uhaku Stage (in the shallow water [near-shore] area of the Baltic Basin), stratigraphically older than the Oil Shales, found no specimens with borings
(Figs. 1
The percentage of borings in brachiopod populations both preceding and succeeding the oil shale fauna seem to be remarkably lower. The collection of Estlandia (13 specimens) from the Uhaku Stage (in the shallow water [near-shore] area of the Baltic Basin), stratigraphically older than the Oil Shales, found no specimens with borings
(Figs. 1 ![]() - 2
- 2 ![]() ). The collection of Clitambonites schmidti from North Estonia yielded only one specimen with borings. Collections of Estlandia (ten specimens), Clitambonites (seven specimens) and Bekkerina (six specimens) from the stratigraphically younger Haljala Stage did not yield even one specimen with borings
(Fig. 2
). The collection of Clitambonites schmidti from North Estonia yielded only one specimen with borings. Collections of Estlandia (ten specimens), Clitambonites (seven specimens) and Bekkerina (six specimens) from the stratigraphically younger Haljala Stage did not yield even one specimen with borings
(Fig. 2 ![]() ). So the frequencies of boring are certainly lower in sites other than the oil shale milieu.
). So the frequencies of boring are certainly lower in sites other than the oil shale milieu.
![]() The oldest specimens of articulate brachiopods (Antigonambonites and Iru) with sporadic borings
are from the Volkhov Stage (Arenig) of the Baltic Ordovician (Fig. 2
The oldest specimens of articulate brachiopods (Antigonambonites and Iru) with sporadic borings
are from the Volkhov Stage (Arenig) of the Baltic Ordovician (Fig. 2 ![]() ). Three borings in Antigonambonites, one large (0.7 mm wide) and two small ones (0.1 mm wide), resemble those of Trypanites in
the oil shale brachiopods (see
Plate 1.a
). Three borings in Antigonambonites, one large (0.7 mm wide) and two small ones (0.1 mm wide), resemble those of Trypanites in
the oil shale brachiopods (see
Plate 1.a ![]() ) (,
2004). The others are morphologically different from those seen in the brachiopods of the Kukruse Stage for they penetrate the shell at right angles to the outer surface (see & ,
1969; & ,
1968). Trypanites borings occur sporadically in stratigraphical younger brachiopods, as in Ilmarinia dimorpha from the Oandu Stage (Late Caradoc), of the Ordovician of the Baltic.
) (,
2004). The others are morphologically different from those seen in the brachiopods of the Kukruse Stage for they penetrate the shell at right angles to the outer surface (see & ,
1969; & ,
1968). Trypanites borings occur sporadically in stratigraphical younger brachiopods, as in Ilmarinia dimorpha from the Oandu Stage (Late Caradoc), of the Ordovician of the Baltic.
![]() Ichnogenus: Trypanites ,
1932
Ichnogenus: Trypanites ,
1932
![]() Type ichnospecies: T. wesei ,
1932
Type ichnospecies: T. wesei ,
1932
![]() Remarks: The genus as redefined by
(1972) and later by &
(1987), includes simple, unbranched cylindrical or sub-cylindrical borings in a hard substrate (rock, shell, wood) with a single opening to the surface.
Remarks: The genus as redefined by
(1972) and later by &
(1987), includes simple, unbranched cylindrical or sub-cylindrical borings in a hard substrate (rock, shell, wood) with a single opening to the surface.
Trypanites aff. fimbriatus
Figs. 3 ![]() -
4
-
4 ![]() - 5
- 5 ![]() ;
Pls. 1
;
Pls. 1 ![]() -
2
-
2 ![]()
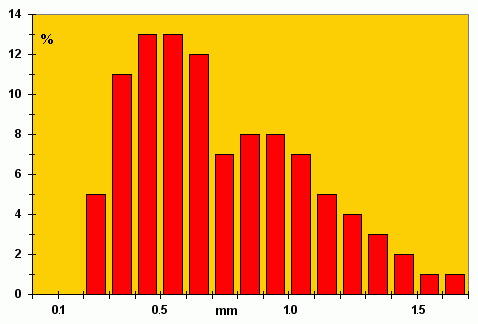
![]() Description: Single, elongate and cylindrical borings. Shafts smooth, commonly straight to slightly curved, more rarely sinusoidal, with few irregularities; cross-section generally circular, but may be oval or flattened. Majority of borings never intersect, and are almost always inclined at a low angle to or are parallel to their substrate. Diameter ranges from 0.2 mm to 1.6 mm, commonly 0.2 - 1.0 mm, and may be as long as 8 mm
(Fig. 3
Description: Single, elongate and cylindrical borings. Shafts smooth, commonly straight to slightly curved, more rarely sinusoidal, with few irregularities; cross-section generally circular, but may be oval or flattened. Majority of borings never intersect, and are almost always inclined at a low angle to or are parallel to their substrate. Diameter ranges from 0.2 mm to 1.6 mm, commonly 0.2 - 1.0 mm, and may be as long as 8 mm
(Fig. 3 ![]() ). A few borings are joined, situated side by side (Plate 1.c
). A few borings are joined, situated side by side (Plate 1.c ![]() ). In some of the intersecting borings the younger ones cross older borings, which may have been abandoned then (Plate 1.f
). In some of the intersecting borings the younger ones cross older borings, which may have been abandoned then (Plate 1.f ![]() ). Terminations of borings are rounded (Figs.
4
). Terminations of borings are rounded (Figs.
4 ![]() - 5
- 5 ![]() ;
Plate 1.e
;
Plate 1.e ![]() ). About half of the borings are oriented with their apertures towards the brachiopod commissure.
). About half of the borings are oriented with their apertures towards the brachiopod commissure.
![]() Discussion: T. aff. fimbriatus resembles T. fimbriatus
1952 (p. 51, pl. 8, figs. 4-6, & ,
1987, pl. 42, fig. 5) in diameter and shape. In contrast to T. fimbriatus the specimens of T. aff. fimbriatus sometimes cross and interpenetrate each other. Neither the shape nor the size of the shafts differ in the non-oriented borings so they cannot be assigned a discrete ichnospecies from those directed toward the brachiopod commissure. However, it is possible that discrete species of borers created the oriented and randomly situated shafts.
Discussion: T. aff. fimbriatus resembles T. fimbriatus
1952 (p. 51, pl. 8, figs. 4-6, & ,
1987, pl. 42, fig. 5) in diameter and shape. In contrast to T. fimbriatus the specimens of T. aff. fimbriatus sometimes cross and interpenetrate each other. Neither the shape nor the size of the shafts differ in the non-oriented borings so they cannot be assigned a discrete ichnospecies from those directed toward the brachiopod commissure. However, it is possible that discrete species of borers created the oriented and randomly situated shafts.
![]() Remarks: Structures associated with the apertures of a few borings in a single Glossorthis sp. valve could be either the calcareous linings produced by boring worms or were secreted by the brachiopod in reaction to the boring worm (Plate 1.c
Remarks: Structures associated with the apertures of a few borings in a single Glossorthis sp. valve could be either the calcareous linings produced by boring worms or were secreted by the brachiopod in reaction to the boring worm (Plate 1.c ![]() ).
).
![]() Occurrence: Abundant borings in hundreds of brachiopods from the oil shale strata of the Kukruse and Kohtla localities NE Estonia, Kukruse Stage, Caradoc, Upper Ordovician.
Occurrence: Abundant borings in hundreds of brachiopods from the oil shale strata of the Kukruse and Kohtla localities NE Estonia, Kukruse Stage, Caradoc, Upper Ordovician.
![]() Brachiopod characteristics that may have influenced the intensity of boring include both shell morphology and the ecological aspects of the relationship (e.g., orientation, valve preference: see Tables
2
Brachiopod characteristics that may have influenced the intensity of boring include both shell morphology and the ecological aspects of the relationship (e.g., orientation, valve preference: see Tables
2 ![]() -
3
-
3 ![]() -
4
-
4 ![]() ).
Morphological characteristics of the host that may have affected the borer larva's choice of substrate include the thickness of the shell, the shell macrostructure as it relates to ease of penetration, and the relief of the shell surface.
).
Morphological characteristics of the host that may have affected the borer larva's choice of substrate include the thickness of the shell, the shell macrostructure as it relates to ease of penetration, and the relief of the shell surface.
![]() The organisms were able to bore into valves that were at a minimum 0.5 mm thick (Septomena sp.). However, the least possible thickness of a valve to be attacked was determined by the diameter of the smallest borer (0.15 mm recorded in Septomena). In the brachiopod genera most subject to boring (Estlandia and Clitambonites), the majority of the shafts range in diameter between 0.9 mm to 3.6 mm (recorded maximum width in C. squamatus). The wall thickness of most thin-shelled brachiopods without borings (e.g., Estonomena) ranges from 0.4-1.1 mm, so shell thickness may have been a limiting factor for the borers.
The organisms were able to bore into valves that were at a minimum 0.5 mm thick (Septomena sp.). However, the least possible thickness of a valve to be attacked was determined by the diameter of the smallest borer (0.15 mm recorded in Septomena). In the brachiopod genera most subject to boring (Estlandia and Clitambonites), the majority of the shafts range in diameter between 0.9 mm to 3.6 mm (recorded maximum width in C. squamatus). The wall thickness of most thin-shelled brachiopods without borings (e.g., Estonomena) ranges from 0.4-1.1 mm, so shell thickness may have been a limiting factor for the borers.
![]() Brachiopod specimens in the examined collection range in size from a 4.5 mm long juvenile Estlandia marginata to a
47 mm Estonomena sp. The smallest brachiopod with a boring (0.3 mm in diameter) is a juvenile Nicolella (6.0 mm long, 7.2 mm wide), and the largest is Cyrtonotella sp. (35 mm long, 47 mm wide). Immature brachiopods have thinner shells. The possible size specificity of Trypanites borings was examined on 14 small specimens (<10
mm long, <15 mm wide, and <1.0 mm thick) of Estlandia marginata. Although for the whole population of specimens of Estlandia marginata in the collection the boring is 51 %, only 28 % of these 14 small specimens are bored, and commonly just once. Thus, the borers appear to have been size-selective; they preferred the larger and thicker shells among the population of the same species. Large hosts have borings of a greater diameter; the valves of smaller hosts are commonly too thin to accommodate large shafts.
Brachiopod specimens in the examined collection range in size from a 4.5 mm long juvenile Estlandia marginata to a
47 mm Estonomena sp. The smallest brachiopod with a boring (0.3 mm in diameter) is a juvenile Nicolella (6.0 mm long, 7.2 mm wide), and the largest is Cyrtonotella sp. (35 mm long, 47 mm wide). Immature brachiopods have thinner shells. The possible size specificity of Trypanites borings was examined on 14 small specimens (<10
mm long, <15 mm wide, and <1.0 mm thick) of Estlandia marginata. Although for the whole population of specimens of Estlandia marginata in the collection the boring is 51 %, only 28 % of these 14 small specimens are bored, and commonly just once. Thus, the borers appear to have been size-selective; they preferred the larger and thicker shells among the population of the same species. Large hosts have borings of a greater diameter; the valves of smaller hosts are commonly too thin to accommodate large shafts.
![]() The size specificity of substrates for Ordovician Vermiforichnus (=
Trypanites, personal communication with Mark ) was recorded by
(1976) in assemblages of Macrocoelia and Heterorthis; the borings are invariably associated with the larger individuals. Here again it is probable that the small shells are too thin to accommodate boring and large shells offer an increased surface area for a larva to settle (see ,
1974). However for Trypanites this areal advantage is valid only within the same host taxon. Dorsal valves of C. schmidti with the highest recorded density of borings have less than half the area of C. squamatus
(Table 4
The size specificity of substrates for Ordovician Vermiforichnus (=
Trypanites, personal communication with Mark ) was recorded by
(1976) in assemblages of Macrocoelia and Heterorthis; the borings are invariably associated with the larger individuals. Here again it is probable that the small shells are too thin to accommodate boring and large shells offer an increased surface area for a larva to settle (see ,
1974). However for Trypanites this areal advantage is valid only within the same host taxon. Dorsal valves of C. schmidti with the highest recorded density of borings have less than half the area of C. squamatus
(Table 4 ![]() ) but in area have a boring density more than twice as great.
) but in area have a boring density more than twice as great.
![]() In Clitambonites schmidti an especially large number of borings occur in the fibrous secondary shell layer
(Plate 2.a
In Clitambonites schmidti an especially large number of borings occur in the fibrous secondary shell layer
(Plate 2.a ![]() ) of the lamellose anterior region of the shell: the thickest portion of the both valves. In Estlandia
(Plate 2.b
) of the lamellose anterior region of the shell: the thickest portion of the both valves. In Estlandia
(Plate 2.b ![]() ) and C. squamatus, which are thin anteriorly and have a less lamellose structure, boring frequency is highest slightly farther back from the anterior margin and the shafts are distributed more equally over the anterior two thirds of the shell. However, boring density per square mm is about half that of C. schmidti
(Table 4
) and C. squamatus, which are thin anteriorly and have a less lamellose structure, boring frequency is highest slightly farther back from the anterior margin and the shafts are distributed more equally over the anterior two thirds of the shell. However, boring density per square mm is about half that of C. schmidti
(Table 4 ![]() ). In the two most bored genera, Estlandia and Clitambonites, the structure of the shell is respectively fibrous pseudopunctate and fibrous unpunctate.
). In the two most bored genera, Estlandia and Clitambonites, the structure of the shell is respectively fibrous pseudopunctate and fibrous unpunctate.
![]() All types of shell surfaces have been bored, from the entirely smooth Bilobia
(Plate 2.d
All types of shell surfaces have been bored, from the entirely smooth Bilobia
(Plate 2.d ![]() ) to the strongly ribbed Glossorthis
(Plate 1.c
) to the strongly ribbed Glossorthis
(Plate 1.c ![]() ), and in a unique case even within a rib. This suggests that the borers' larvae were able to attach to and bore all of the types of surfaces represented by brachiopod valves.
), and in a unique case even within a rib. This suggests that the borers' larvae were able to attach to and bore all of the types of surfaces represented by brachiopod valves.
![]() (1976) suggests that the host specificity of Ordovician Vermiforichnus borings (=
Trypanites) is related to the delicate and parvicostellate nature of its hosts. Several authors have suggested (,
1972; , 1974) that ectozoa avoided coarse-ribbed and angular-ribbed brachiopods. The brachiopod genera examined in this study do not support this idea. The high frequency of borings (20.8 %) in the strongly costate, angular-ribbed Nicolella demonstrates
(Table 1
(1976) suggests that the host specificity of Ordovician Vermiforichnus borings (=
Trypanites) is related to the delicate and parvicostellate nature of its hosts. Several authors have suggested (,
1972; , 1974) that ectozoa avoided coarse-ribbed and angular-ribbed brachiopods. The brachiopod genera examined in this study do not support this idea. The high frequency of borings (20.8 %) in the strongly costate, angular-ribbed Nicolella demonstrates
(Table 1 ![]() ;
Plate 2.c
;
Plate 2.c ![]() ) that high costa were not a limiting constraint for that borer. Moreover, borings are relatively infrequent on flat surfaces, with the exception of Septomena and a single boring in Bilobia
(Plate 2.d
) that high costa were not a limiting constraint for that borer. Moreover, borings are relatively infrequent on flat surfaces, with the exception of Septomena and a single boring in Bilobia
(Plate 2.d ![]() ). Borings usually occur between the ribs or in any type of depression, including the shell interior or the hinge line cavity. This suggests that cavities were preferred to flat surfaces., for after attachment these niches may have provided the borer larva with a safer place to start operations. The moderately costate brachiopods Clitambonites and Estlandia have the highest boring frequences
(Table 1
). Borings usually occur between the ribs or in any type of depression, including the shell interior or the hinge line cavity. This suggests that cavities were preferred to flat surfaces., for after attachment these niches may have provided the borer larva with a safer place to start operations. The moderately costate brachiopods Clitambonites and Estlandia have the highest boring frequences
(Table 1 ![]() ), but this may be ascribed to various ecological factors or to their relatively thick and lamellose shell.
), but this may be ascribed to various ecological factors or to their relatively thick and lamellose shell.
![]() Most borings are on the outer surfaces of the valves. Only in intensely bored shells of Estlandia
(Plate 2.b
Most borings are on the outer surfaces of the valves. Only in intensely bored shells of Estlandia
(Plate 2.b ![]() ) and Clitambonites
(Plate 2.a
) and Clitambonites
(Plate 2.a ![]() ) are borings found in the interior
(Plate 2.e
) are borings found in the interior
(Plate 2.e ![]() ), and they occur in only 3.3 % of all bored shells (see also
Table 2
), and they occur in only 3.3 % of all bored shells (see also
Table 2 ![]() ). The valves with interior borings are always strongly weathered.
). The valves with interior borings are always strongly weathered.
![]() A selective orientation of borings with respect to the host has been documented for Vermiforichnus (= Trypanites) by
(1976, see p. 161-162), and was confirmed for Trypanites in this study. A majority of borings were located at the anterior margin and oriented perpendicularly to the commissure
(Plate 2.a
A selective orientation of borings with respect to the host has been documented for Vermiforichnus (= Trypanites) by
(1976, see p. 161-162), and was confirmed for Trypanites in this study. A majority of borings were located at the anterior margin and oriented perpendicularly to the commissure
(Plate 2.a ![]() ). Presumably these borings were made in the shells of living brachiopods (,
1976).
). Presumably these borings were made in the shells of living brachiopods (,
1976).
![]() Similar observations were made by
(1957), who found a concentration of recent spionid polychaetes (Polydora ciliata) at the valve margins of Mytilus. Polydora hoplura has been described as oriented perpendicularly to the edge of oyster shells, and
(1964) discovered that larvae of Polydora websteri settled in a similar orientation. These Recent polychaetes appear to have no detrimental effect on the host, and in the same way the boring worms of the Ordovician may have benefited from the feeding currents produced by the host (,
1976).
Similar observations were made by
(1957), who found a concentration of recent spionid polychaetes (Polydora ciliata) at the valve margins of Mytilus. Polydora hoplura has been described as oriented perpendicularly to the edge of oyster shells, and
(1964) discovered that larvae of Polydora websteri settled in a similar orientation. These Recent polychaetes appear to have no detrimental effect on the host, and in the same way the boring worms of the Ordovician may have benefited from the feeding currents produced by the host (,
1976).
![]() An increased density of Trypanites borings has been ascribed to its association with the shells of living hosts. In addition to the advantages of feeding mentioned, living hosts would provided better protection from sedimentation, overturning, breakage or abrasion than empty shells could offer (,
1976). When worms in empty shells were buried by sediment, they probably succumbed.
An increased density of Trypanites borings has been ascribed to its association with the shells of living hosts. In addition to the advantages of feeding mentioned, living hosts would provided better protection from sedimentation, overturning, breakage or abrasion than empty shells could offer (,
1976). When worms in empty shells were buried by sediment, they probably succumbed.
![]() In Clitambonites and Estlandia, the great majority (about 90 %) of multiple borings in brachial valves and in the anterior two-thirds of pedicle valves are oriented perpendicularly to the commissure or inclined towards it (when not situated at the anterior margin itself). This pattern is best exhibited in Clitambonites schmidti
(Plate 2.a
In Clitambonites and Estlandia, the great majority (about 90 %) of multiple borings in brachial valves and in the anterior two-thirds of pedicle valves are oriented perpendicularly to the commissure or inclined towards it (when not situated at the anterior margin itself). This pattern is best exhibited in Clitambonites schmidti
(Plate 2.a ![]() ; see also
Table 2
; see also
Table 2 ![]() )
and is apparent in other taxa when the density of borings on the host is high (Nicolella, Glossorthis, and Septomena). The particularly high concentration of oriented borings near the commissure of both valves in C. schmidti (up to a maximum of 24 on a brachial valve) presumably owes its existence to two favoring factors in that location: the specific lamellose structure of the shell and the extraordinary thickness of the anterior margin, both of which facilitated the successful attachment of borer's larva. The density of borings in C. schmidti is twice as high as in the other two species C. squamatus and E. marginata
(Table 4
)
and is apparent in other taxa when the density of borings on the host is high (Nicolella, Glossorthis, and Septomena). The particularly high concentration of oriented borings near the commissure of both valves in C. schmidti (up to a maximum of 24 on a brachial valve) presumably owes its existence to two favoring factors in that location: the specific lamellose structure of the shell and the extraordinary thickness of the anterior margin, both of which facilitated the successful attachment of borer's larva. The density of borings in C. schmidti is twice as high as in the other two species C. squamatus and E. marginata
(Table 4 ![]() ), and this may be due to the attachment advantages of offered by the thick lamellose anterior part of the shell, made even more attractive by the improved feeding associated with a location close to the commissure.
), and this may be due to the attachment advantages of offered by the thick lamellose anterior part of the shell, made even more attractive by the improved feeding associated with a location close to the commissure.
![]() The number of borings on the brachial valves is about twice that on the pedicle valves in all three of the species studied (C. schmidti, C. squamatus and E. marginata). So the brachial valve was a preferred substrate for larvae when they attached to a living brachiopod
(Table 3
The number of borings on the brachial valves is about twice that on the pedicle valves in all three of the species studied (C. schmidti, C. squamatus and E. marginata). So the brachial valve was a preferred substrate for larvae when they attached to a living brachiopod
(Table 3 ![]() ) with its brachial valve up. The living adult brachiopods C. schmidti and C. squamatus presumably lay on the pedicle valve, with the posterior third
of that valve either on or slightly below the sediment line. There are only a
few unorientated borings of a clearly post-mortem origin. It is possible that young specimens of Estlandia, that may have retained a functional pedicle longer, were oriented on the substrate with their anterior ends upwards. That would explain the presence of a few oriented borings in the center of some pedicle valves (see
Plate 2.b
) with its brachial valve up. The living adult brachiopods C. schmidti and C. squamatus presumably lay on the pedicle valve, with the posterior third
of that valve either on or slightly below the sediment line. There are only a
few unorientated borings of a clearly post-mortem origin. It is possible that young specimens of Estlandia, that may have retained a functional pedicle longer, were oriented on the substrate with their anterior ends upwards. That would explain the presence of a few oriented borings in the center of some pedicle valves (see
Plate 2.b ![]() ). The pedicle valve of Clitambonites is never so attacked.
). The pedicle valve of Clitambonites is never so attacked.
![]() The concavo-convex strophomenids rested on the pedicle valve while boring took place, but presumably all borings were made after death, for the boring on Bilobia penetrates the brachial valve of the tiny shell
(Plate 2.d
The concavo-convex strophomenids rested on the pedicle valve while boring took place, but presumably all borings were made after death, for the boring on Bilobia penetrates the brachial valve of the tiny shell
(Plate 2.d ![]() ), and all the the borings in Bekkerina are on the hinge line, a phenomenon functionally impossible in a living brachiopod.
), and all the the borings in Bekkerina are on the hinge line, a phenomenon functionally impossible in a living brachiopod.
![]() The
relatively short length of Trypanites borings and their large apertures suggest suspension feeding using to advantage the feeding currents produced by the living brachiopod.
The
relatively short length of Trypanites borings and their large apertures suggest suspension feeding using to advantage the feeding currents produced by the living brachiopod.
![]() There are few shells with repaired borings showing that the borings had disturbed the living host
(Plate 2.f
There are few shells with repaired borings showing that the borings had disturbed the living host
(Plate 2.f ![]() ). The few blisters (see also & ,
1992, p. 253-260) found in the interiors of bored brachiopods (see
Plate 2.g
). The few blisters (see also & ,
1992, p. 253-260) found in the interiors of bored brachiopods (see
Plate 2.g ![]() ) obviously formed when the boring organism approached soft tissues. These features are rare but they indicate clearly that the relationship of the borer and its host was not always neutral. To some extent boring animals may have benefited as parasites by taking nutrients from the shell material of a living brachiopod. However, if that were the case they were presumably immobile. It is certain that in dead shells, the borer's larva attached and made borings only as habitations.
) obviously formed when the boring organism approached soft tissues. These features are rare but they indicate clearly that the relationship of the borer and its host was not always neutral. To some extent boring animals may have benefited as parasites by taking nutrients from the shell material of a living brachiopod. However, if that were the case they were presumably immobile. It is certain that in dead shells, the borer's larva attached and made borings only as habitations.
![]() Brachiopods with borings are divided into two groups: those with many
borings, the clitambonitids Estlandia marginata, Clitambonites squamatus, and Clitambonites schmidti; and the moderately bored Nicolella, Glossorthis, Kullervo, Cyrtonotella and Bekkerina. Similar host-selective-distribution patterns has been recorded for Vermiforichnus (= Trypanites) borings in Middle Ordovician brachiopods from Wales (,
1976). The moderately costate and relatively thick-shelled clitambonitids may have been preferred by the borers because of their thick shell, lamellose structure, and large size. Depressions such as the areas between costa and between the shell lamellae could have protected newly attached larvae from possible predators, such as grazing snails.
Brachiopods with borings are divided into two groups: those with many
borings, the clitambonitids Estlandia marginata, Clitambonites squamatus, and Clitambonites schmidti; and the moderately bored Nicolella, Glossorthis, Kullervo, Cyrtonotella and Bekkerina. Similar host-selective-distribution patterns has been recorded for Vermiforichnus (= Trypanites) borings in Middle Ordovician brachiopods from Wales (,
1976). The moderately costate and relatively thick-shelled clitambonitids may have been preferred by the borers because of their thick shell, lamellose structure, and large size. Depressions such as the areas between costa and between the shell lamellae could have protected newly attached larvae from possible predators, such as grazing snails.
![]() The absence of borings in several brachiopod taxa (Porambonites, Oxoplecia, Leptestia, Leptelloidea, Estonomena, Sowerbyella, Actinomena, Orthisocrania) has two possible explanations: either the brachiopods with borings lived in environments other than those occupied by brachiopods without borings, or the boring animals avoided certain taxa completely. With the exception of size and valve thickness no common morphological feature distinguishes brachiopods with borings from those without them. Even if the articulates in the oil shale taphocoenosis had a discrete autoecology that does not explain the lack of borings in the dead shells. Even if one were to assume that the life style of certain brachiopods kept the borers away, some of their dead shells would have been accessible to them. In the concavo-convex Bekkerina
a lack of oriented borings may have been caused by the unfavourable autoecology of the living brachiopod. It is possible that some brachiopods repelled the larvae of borers by some chemical means or protected their shells against larval settlings by poisonous secretions on their surface.
The absence of borings in several brachiopod taxa (Porambonites, Oxoplecia, Leptestia, Leptelloidea, Estonomena, Sowerbyella, Actinomena, Orthisocrania) has two possible explanations: either the brachiopods with borings lived in environments other than those occupied by brachiopods without borings, or the boring animals avoided certain taxa completely. With the exception of size and valve thickness no common morphological feature distinguishes brachiopods with borings from those without them. Even if the articulates in the oil shale taphocoenosis had a discrete autoecology that does not explain the lack of borings in the dead shells. Even if one were to assume that the life style of certain brachiopods kept the borers away, some of their dead shells would have been accessible to them. In the concavo-convex Bekkerina
a lack of oriented borings may have been caused by the unfavourable autoecology of the living brachiopod. It is possible that some brachiopods repelled the larvae of borers by some chemical means or protected their shells against larval settlings by poisonous secretions on their surface.
![]() As a rule, brachiopods without borings are thin-shelled, but Porambonites is an exception. The thickest parts of the shell of large thin-shelled strophomenids and that of all large individuals of other genera provide adequate space for the small borings discovered in young Estlandia and Nicolella and in adult specimens of thin-shelled Septomena. Borers avoided flat surfaces, but could bore in them. Flat surfaces are bored in weathered shells of Clitambonites and Estlandia (about 2 % of the total number of borings). Some borings have been found in the interior of shells, and an exceptional boring exists in the middle of a flat brachial valve of Bilobia
(Plate 2.d
As a rule, brachiopods without borings are thin-shelled, but Porambonites is an exception. The thickest parts of the shell of large thin-shelled strophomenids and that of all large individuals of other genera provide adequate space for the small borings discovered in young Estlandia and Nicolella and in adult specimens of thin-shelled Septomena. Borers avoided flat surfaces, but could bore in them. Flat surfaces are bored in weathered shells of Clitambonites and Estlandia (about 2 % of the total number of borings). Some borings have been found in the interior of shells, and an exceptional boring exists in the middle of a flat brachial valve of Bilobia
(Plate 2.d ![]() ). The strophomenid Septomena has several oriented borings, although its sculpture is very
fine.
). The strophomenid Septomena has several oriented borings, although its sculpture is very
fine.
![]() Presumably the relief of the substrate was not a controlling factoring in the placement of borings. Recent boring polychaetes, such as Polydora, bore into diverse substrates including limestone pebbles (see ,
1966, p. 352-354). Available data on the specificity of the relationships between boring polychaetes and their hosts seems to display a generic monoxenous pattern. About 37 % of polychaetes infest a single host species, 63 % infest three or fewer host species ( & ,
1998). The host specificity of Trypanites borings in the oil shale brachiopods may reflect a similar pattern. The biological affinities of the organisms that bored the oil shale brachiopods are unknown. These Trypanites borings could have been produced by polychaetes, but they may well have been made by some kind of boring lophophorate, for example, the phoronids. Borings by representatives of this soft-bodied worm-like phylum are known from the Palaeozoic (,
1982). However, other soft-bodied, suspension-feeding invertebrates can not be ruled out as possible borers.
Presumably the relief of the substrate was not a controlling factoring in the placement of borings. Recent boring polychaetes, such as Polydora, bore into diverse substrates including limestone pebbles (see ,
1966, p. 352-354). Available data on the specificity of the relationships between boring polychaetes and their hosts seems to display a generic monoxenous pattern. About 37 % of polychaetes infest a single host species, 63 % infest three or fewer host species ( & ,
1998). The host specificity of Trypanites borings in the oil shale brachiopods may reflect a similar pattern. The biological affinities of the organisms that bored the oil shale brachiopods are unknown. These Trypanites borings could have been produced by polychaetes, but they may well have been made by some kind of boring lophophorate, for example, the phoronids. Borings by representatives of this soft-bodied worm-like phylum are known from the Palaeozoic (,
1982). However, other soft-bodied, suspension-feeding invertebrates can not be ruled out as possible borers.
![]() Taphonomic analyses of an assemblage of the bored Ordovician brachiopod Onniella meeki performed by and
(2000) suggested that 10-15 % of the borings in the
assemblage were predatory. Thus, some borings discussed here may represent predation: those that penetrate the valves. However, it is more likely that all borings in Ordovician brachiopods are simply habitations ( & ,
2001).
Taphonomic analyses of an assemblage of the bored Ordovician brachiopod Onniella meeki performed by and
(2000) suggested that 10-15 % of the borings in the
assemblage were predatory. Thus, some borings discussed here may represent predation: those that penetrate the valves. However, it is more likely that all borings in Ordovician brachiopods are simply habitations ( & ,
2001).
![]() I would like to thank V. for valuable discussions and data on the genesis of Estonian oil shale. T.K. , R.G. and M. are thanked for constructive remarks on the manuscript. A. , C.C. and R. provided helpful comments on the manuscript. Special thanks to N.J. for his assistance in improving the English text. I am grateful to A. for the opportunity to study his collection of strophomenid brachiopods from the oil shale.
I would like to thank V. for valuable discussions and data on the genesis of Estonian oil shale. T.K. , R.G. and M. are thanked for constructive remarks on the manuscript. A. , C.C. and R. provided helpful comments on the manuscript. Special thanks to N.J. for his assistance in improving the English text. I am grateful to A. for the opportunity to study his collection of strophomenid brachiopods from the oil shale.
![]()
H. & V. (1990).- Geology of the Baltic oil shale basin. In: D. and H. (eds.) Field Meeting Estonia 1990, An excursion guidebook.- Estonian Academy of Sciences, Tallinn, p. 40-45.
G.J. (1966).- Shell borings of sessile epibiontic organisms as palaeoecological guides.- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, v. 3, p. 311-362.
D. & H.F. (1992).- "Blisters" in a Famennian cyrtospiriferid brachiopod from Hunan (South China).- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, v. 94, p. 253-260.
R.G. (1972).- On some ichnotaxa in hard substrates, with a redefinition of Trypanites .- Paläontologische Zeitschrift, Stuttgart, Band 46, Heft 1/2, p. 93-98.
R.G. & A. (1987).- Bioerosion of the Plio-Pleistocene transgression of southern Italy.- Rivista Italiana di Paleontologica e Stratigrafia, Milano, v. 93, p. 379-442.
B. (1969).- Paleozoic shell-boring annelids and their trace fossils.- American Zoologist, Seattle, v. 9, p. 689-703.
M.R. & E.L. (1968).- Recent gastropod boreholes and Ordovician cylindrical borings.- U.S. Geological Survey, Professional Paper, Washington, 593-B, p. B1-B26.
A.V., R. & M. (2002).- Trace fossils and ichnofabrics across the Volkhov depositional sequence (Ordovician, Arenigian of St. Petersburg Region, Russia).- Journal of the Czech Geological Society, Praha, v. 47, p. 133146.
A.A. & R.G. (2001).- Bioerosional innovation for living in carbonate hardgrounds in the Early Ordovician of Sweden.- Lethaia, Oslo, v. 34, p. 1-12.
C.C. (1982).- The biology of Phoronida.- Advances in Marine Biology, London, v. 19, p. 1-89.
P.S. (1964).- The American oyster.- U.S. Fish and Wildlife Service Fishery Bulletin, Washington, v. 64, 480 pp.
C. (1957).- Über den Röhrenbau und die Nahrungsaufnahme einiger Spinoiden der Deutschen Küsten.- Helgoländer wissenschaftliche Meeresunters, Hamburg, v. 6, p. 100-135.
R. (1928).- Paleobiologicheskye nablyudenya nad nizhnesiluryiskimi bespozvonochnymi, I.- Yezhegodnik Russkogo Paleontologicheskogo Obschestva, T. VII (1927), p. 1-86 (in Russian)
L. & A. (1997).- Ordovician articulate brachiopods. In: and (eds.) Geology and Mineral Resources of Estonia.- Estonian Academy Publishers, Tallinn 1997, p. 226-228.
J.M. (1974).- Selective epizoan encrustation of some Silurian Brachiopods from Gotland.- Palaeontology, London, v. 17, p. 423-429.
V. (1973).- Aspects of carbonate sedimentation in the Ordovician of Baltoscandia.- Lethaia, Oslo, v. 6, p. 11-34.
N.P., D.R. & S.G. (1977).- The oldest macroborers: Lower Cambrian of Labrador.- Science, Washington, v. 197, p. 980-983.
P. & K.T. (2000).- Taphonomic inferences on boring habit in the Richmondian Onniella meeki epibole.- Palaios, Reno, v. 15, p. 499510.
D.R., N.P. & S.G. (1978).- Initial diversification of macroboring ichnofossils and exploitation of the macroboring niche in the lower Paleozoic.- Paleobiology, Chicago, v. 4, p. 163-170.
D.R. & S. (1982).- The macroboring ichnofossil Trypanites in colonies of the Middle Ordovician bryozan Prasopora: population behaviour and reaction to environmental influences.- Canadian Journal of Earth Sciences, Ottawa, v. 19, p. 679-688.
A. & R. (1994).- Scanning electron microscopy of Gloeocapsamorpha as produced from kerogen oxidation.- Oil Shale, Tallinn, v. 11, p. 293-303.
K. (1932).- Über einige Bohrgänge aus dem unteren Muschelkalk von Jena.- Paläontologische Zeitschrift, Stuttgart, Band 14, p. 150-160.
D. & T.A. (1998).- Symbiotic polychaetes: Review of known species.- Oceanography and Marine Biology: an Annual Review, London, v. 36, p. 217-340.
H. & R. (1997).- Ordovician and Silurian carbonate sedimentation basin. In: A. & A. (Eds.) Geology and Mineral Resources of Estonia.- Estonian Academy Publishers, Tallinn 1997, p. 192-205.
P.R. & T.L. (1980).- Macroborings of Middle Ordovician age from southern Norway.- Journal of Paleontology, Tulsa, v. 54, p. 1352-1355.
T.J. & C.R. (1993).- Borings and bioerosion in the fossil record.- Geology Today, London, v. 9, p. 138-142.
R.K. (1976).- Vermiforichnus borings from the Ordovician of central Wales.- Geological Magazine, Cambridge, v. 113, p. 159-164.
V., H. & R. (1988).- The condition of kukersite deposition.- Proceedings of the International Conference on Oil Shale and Shale Oil, Beijing (May 16-19, 1988), p. 42-50.
R.P. & C.W. (1969).- Cylindrical living burrows in Ordovician Dalmanellid brachiopod beds.- Journal of Paleontology, Tulsa, v. 43, p. 838-841.
R.P. (1972).- Autoecology of Richmondian Brachiopods (Late Ordovician of Indiana and Ohio).- Journal of Paleontology, Tulsa, v. 46, p. 386-406.
A. (1970).- Stratigrafia viruskoy i kharjuskoy seriy (ordovik) Severnoy Estonii [Stratigraphy of the Viruan Series (Middle Ordovician) in Northern Estonia].- Tallinn, Valgus, p. 1-346.
L. (1952).- Larger invertebrate fossils of the Woodbine Formation (Cenomanian) of Texas.- U.S. Geological Survey, Professional Paper, Washington, 242 iv, 226 pp.
P.D. & M.A. (2003).- Palaeontology and evolution of marine hard substrate communities.- Earth Science Reviews, Amsterdam, v. 62, p. 1-103.
C.W. (1974).- Substrate specificity of Devonian epizoa.- Journal of Paleontology, Tulsa, v. 48, p. 881-894.
O. (2004).- Trypanites borings in the shells of brachiopod Antigonambonites from Arenig of Baltica.- Proceedings of the Estonian Academy of Sciences, Geology, Tallinn, v. 53, p. 257-266.
M.A. & T.J. (2001).- Domiciles, Not Predatory Borings: A simpler explanation of the holes in Ordovician shells analyzed by and , 2000.- Palaios, Reno, v. 16, Nº 5, p. 524-525.
![]()
| Species/genus | Number of studied valves |
Number of bored valves |
Boring frequency |
| Estlandia marginata | 267 | 131 | 51 % |
| Clitambonites squamatus | 271 | 130 | 48 % |
| Clitambonites schmidti | 397 | 180 | 45.3 % |
| Nicolella ssp. | 29 | 5 | 20.8 % |
| Glossothis ssp. | 6 | 1 | 16.7 % |
| Bilobia ssp. | 7 | 1 | 14.3 % |
| Kullervo ssp. | 9 | 1 | 11.1 % |
| Cyrtonotella ssp. | 11 | 1 | 9.1 % |
| Septomena ssp. | 29 | 2 | 6.9 % |
| Bekkerina ssp. | 46 | 3 | 6.5 % |
Table 1: List of studied brachiopods from oil shale (Kukruse Stage) and their boring frequency with Trypanites aff. fimbriatus.
| Species | Number of ventral valves | Boring frequency of ventral valves % | Number of dorsal valves | Boring frequency of dorsal valves % | Boring frequency of shell interiors % |
| Estlandia marginata | 105 | 47.6 % | 162 | 53.1 % | 6.7 % |
| Clitambonites squamatus | 176 | 40.3 % | 95 | 62.1 % | 1.8 % |
| Clitambonites schmidti | 193 | 43.5 % | 204 | 47.1 % | 1.5 % |
Table 2: Trypanites aff. fimbriatus boring frequency of the ventral and dorsal valves, and shell interiors.
| Species | Number of dorsal valves studied (no. of studied dorsal borings) | Borings on average per a dorsal valve | Number of ventral valves studied (no. of studied ventral borings) | Borings on average per a ventral valve |
| Estlandia marginata | 44 (250) | 5.7 | 21 (67) | 3.2 |
| Clitambonites squamatus | 19 (207) | 10.9 | 14 (83) | 5.9 |
| Clitambonites schmidti | 33 (368) | 11.2 | 28 (173) | 6.2 |
Table 3: Average number of Trypanites aff. fimbriatus borings in dorsal and ventral valves of Estlandia marginata, Clitambonites squamatus and C. schmidti.
| Species | Number of dorsal valves | Average area of dorsal valve | Borings per 10 mm2 |
| Estlandia marginata | 44 | 26.8 mm2 | 2.1 |
| Clitambonites squamatus | 19 | 43.5 mm2 | 2.5 |
| Clitambonites schmidti | 33 | 20.5 mm2 | 5.5 |
Table 4: Boring density on the dorsal valves in Estlandia marginata, Clitambonites squamatus and C. schmidti.
![]()
|
Click on thumbnail to enlarge the image.
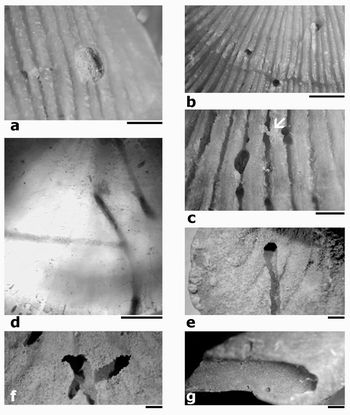
Plate 1: Trypanites ssp. borings (scale bars 1 mm):
(a) in exterior of ventral valve of Arenig Antigonambonites planus, Volkhov Stage, Lava River, NW Russia, TUG 1121/1;
(b) in dorsal exterior of Antigonambonites planus, Volkhov Stage (Arenig), Volkhov, NW Russia, TUG 1121/2;
(c) in ventral exterior of Glossorthis sp., note the two joined borings, and the possible mud constructions associated with the aperture of a boring in the central upper part of figure, TUG 1121/3;
(d) TUG 1121/2 translucent view;
(e) longitudinal section of a broken out boring in ventral interior of Estlandia marginata, TUG 1121/4;
(f) broken out crossing borings in dorsal interior of E. marginata, TUG 1121/5;
(g) longitudinal section in a ventral valve of Clitambonites squamatus, TUG 1121/6.
|
Click on thumbnail to enlarge the image.
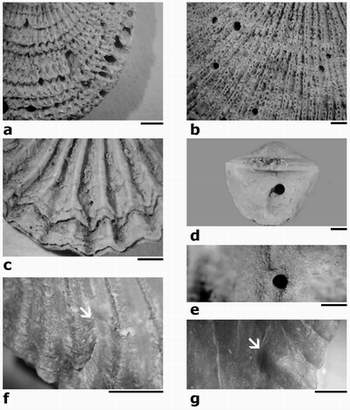
Plate 2: Trypanites aff. fimbriatus borings, Caradoc, Kukruse Stage, Kohtla, Estonia (scale bars 1 mm):
(a) oriented borings in anterior commissure of dorsal valve of Clitambonites schmidti, TUG 1121/7;
(b) oriented external borings in anterior half of ventral valve of Estlandia marginata, TUG 1121/8;
(c) oriented borings in dorsal exterior of Nicolella sp., TUG 1121/9;
(d) in dorsal valve exterior of Bilobia sp., TUG 1121/10;
(e) in ventral interior of Estlandia marginata, TUG 1121/11;
(f) repaired boring in ventral exterior of Glossorthis sp., TUG 1121/3;
(g) 'blister'-like structure in ventral interior of Glossorthis sp., below the repaired boring, TUG 1121/3.
![]()
|
Click on thumbnail to enlarge the image.
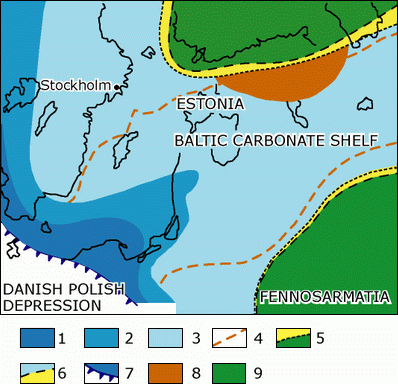
Figure 1: Setting of the oil shale-bearing deposits in the Upper Ordovician (Caradoc) of Baltoscandia (modified after et alii, 1988). 1 - black shales; 2 - argillaceous detrital limestones; 3 - detrital limy marls; 4 - supposed boundary of oil shale occurences; 5 - erosion boundary; 6 - boundary of the oil shale accumulation area; 7 - lineament; 8 - rich oil shale occurrences (with bored brachiopods); 9 exposed land during Late Ordovician.
|
Click on thumbnail to enlarge the image.
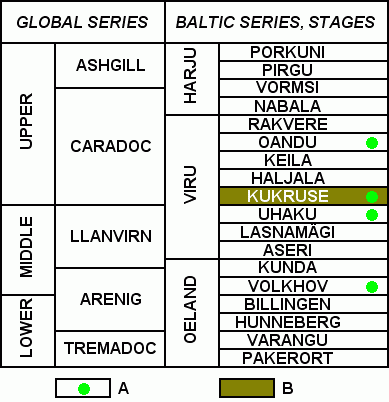
Figure 2: Stratigraphical subdivisions of Baltoscandian Ordovician. A: Worm borings in brachiopod shells; B: Main oil shale deposits.
|
Figure 3: Frequency histograms of maximum diameter of Trypanites aff. fimbriatus borings in 20 valves (100 borings examined) of the brachiopods Estlandia marginata, Clitambonites squamatus, C. schmidti and Septomena sp.

|
Figure 4: Trypanites aff. fimbriatus, Caradoc, Kukruse Stage, Kohtla, Estonia. Artificial cast of the boring preserved in Estlandia marginata shell, TUG 1121/20, scale bar 1mm.

|
Figure 5: Trypanites aff. fimbriatus, Caradoc, Kukruse Stage, Kohtla, Estonia. Artificial cast of the boring preserved in Estlandia marginata shell, TUG 1121/21, scale bar 1mm.
![]()