![]()
![]()
![]() Parsimony analysis of endemicity was applied to analyze the distribution of enchodontoid fishes occurring strictly in the
Cenomanian. The analysis was carried out using the computer program PAUP* 4.0b10, based on a data matrix built with 17 taxa and 12 areas. The rooting was made on an hypothetical all-zero
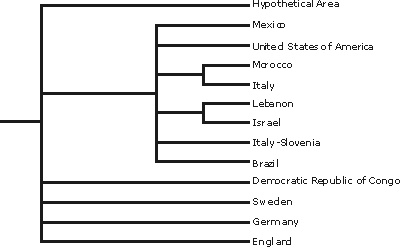
outgroup. Applying the exact algorithm branch and bound, 47 trees were obtained with 26 steps, a consistency index of 0.73, and a retention index of 0.50. The topology found with a majority rule consensus was: [(Mexico) + (United States) + (Morocco + Italy) + (Lebanon + Israel) + (Italy-Slovenia) + (Brazil)] +
(D.R. Congo) + (Sweden) + (Germany) + (England). The procedure delimited two areas of endemism in the Tethys Ocean. They are Morocco and southern Italy and Lebanon and Israel. The area of endemism formed by Morocco + Italy represents the North African region of the Tethys Ocean, and that formed by Lebanon + Israel is in the
mid-Tethyan Ocean. Our results are in partial agreement with the patterns of geographical distribution of certain invertebrate biota.
Parsimony analysis of endemicity was applied to analyze the distribution of enchodontoid fishes occurring strictly in the
Cenomanian. The analysis was carried out using the computer program PAUP* 4.0b10, based on a data matrix built with 17 taxa and 12 areas. The rooting was made on an hypothetical all-zero
outgroup. Applying the exact algorithm branch and bound, 47 trees were obtained with 26 steps, a consistency index of 0.73, and a retention index of 0.50. The topology found with a majority rule consensus was: [(Mexico) + (United States) + (Morocco + Italy) + (Lebanon + Israel) + (Italy-Slovenia) + (Brazil)] +
(D.R. Congo) + (Sweden) + (Germany) + (England). The procedure delimited two areas of endemism in the Tethys Ocean. They are Morocco and southern Italy and Lebanon and Israel. The area of endemism formed by Morocco + Italy represents the North African region of the Tethys Ocean, and that formed by Lebanon + Israel is in the
mid-Tethyan Ocean. Our results are in partial agreement with the patterns of geographical distribution of certain invertebrate biota.
Biogeography; parsimony analysis of endemicity; Late Cretaceous; Enchodontoidei
H.M.A. da & V. (2007).- Parsimony analysis of endemicity of enchodontoid fishes from the Cenomanian.- Carnets de Géologie / Notebooks on Geology, Brest, Letter 2007/01 (CG2007_L01)
![]() Analyse parcimonieuse d'endémicité des poissons Enchodontoidei du Cénomanien.- L'analyse parcimonieuse d'endémicité est réalisée pour expliquer la distribution cénomanienne des poissons
Enchodontoidei. Le programme PAUP* 4.ob10 a été utilisé. L'analyse est basée sur une matrice construite à partir de 17 taxons provenant de 12 régions. L'enracinement se fait sur un groupe externe hypothétique. Par application de l'algorithme du "branch and bound", 47 arbres ont été obtenus à 26 pas, avec un index de consistance de 0,73 et un index de rétention de 0,50. Le consensus donne la topologie suivante : [(Mexique) +
(Etats-Unis) + (Maroc + Italie) + (Liban + Israël) + (Italie-Slovénie) + (Brésil)] + (R.D. du Congo) +
(Suède) + (Allemagne) + (Angleterre). Deux aires d'endémismes ont été identifiées en Téthys
(Maroc et Italie du Sud; Liban et Israël). L'aire d'endémisme formée par le Maroc et l'Italie représente la région nord-africaine de la Téthys et celle formée par le Liban et Israël le
centre. Nos résultats s'accordent partiellement avec la répartition géographique de certains biotopes invertébrés.
Analyse parcimonieuse d'endémicité des poissons Enchodontoidei du Cénomanien.- L'analyse parcimonieuse d'endémicité est réalisée pour expliquer la distribution cénomanienne des poissons
Enchodontoidei. Le programme PAUP* 4.ob10 a été utilisé. L'analyse est basée sur une matrice construite à partir de 17 taxons provenant de 12 régions. L'enracinement se fait sur un groupe externe hypothétique. Par application de l'algorithme du "branch and bound", 47 arbres ont été obtenus à 26 pas, avec un index de consistance de 0,73 et un index de rétention de 0,50. Le consensus donne la topologie suivante : [(Mexique) +
(Etats-Unis) + (Maroc + Italie) + (Liban + Israël) + (Italie-Slovénie) + (Brésil)] + (R.D. du Congo) +
(Suède) + (Allemagne) + (Angleterre). Deux aires d'endémismes ont été identifiées en Téthys
(Maroc et Italie du Sud; Liban et Israël). L'aire d'endémisme formée par le Maroc et l'Italie représente la région nord-africaine de la Téthys et celle formée par le Liban et Israël le
centre. Nos résultats s'accordent partiellement avec la répartition géographique de certains biotopes invertébrés.
Biogéographie ; analyse parcimonieuse d'endémicité ; Crétacé supérieur ; Enchodontoidei
![]() Parsimony Analysis of Endemicity (PAE) was originally developed by
(1988) in a paleontological setting. According to &
(1995), PAE is a cladistic method that groups areas by their shared taxa based on the most parsimonious solution. This methodology allows an interpretation of the occupation of an area by taxa, to verify the relationships among areas, and to identify putative areas of endemism using a matrix built with taxa versus areas (or localities). Despite some criticisms (i.e., & ,
1999; & ,
2003), PAE is a tool potentially useful in historical biogeography, because it allows an analysis of historical information from the geographic distribution of taxonomic groups, even if phylogenetic analyses are lacking. Additionally, in a recent paper of PAE review,
(2006) claimed that it remains as an insightful method for inferring historical patterns.
Parsimony Analysis of Endemicity (PAE) was originally developed by
(1988) in a paleontological setting. According to &
(1995), PAE is a cladistic method that groups areas by their shared taxa based on the most parsimonious solution. This methodology allows an interpretation of the occupation of an area by taxa, to verify the relationships among areas, and to identify putative areas of endemism using a matrix built with taxa versus areas (or localities). Despite some criticisms (i.e., & ,
1999; & ,
2003), PAE is a tool potentially useful in historical biogeography, because it allows an analysis of historical information from the geographic distribution of taxonomic groups, even if phylogenetic analyses are lacking. Additionally, in a recent paper of PAE review,
(2006) claimed that it remains as an insightful method for inferring historical patterns.
![]() Recent papers have applied PAE to the study of faunal distributions, mainly those of extant taxa (i.e., & ,
1999; et alii, 1999; et alii,
2001; , 2001; & ,
2004; & ,
2005; et alii, 2006; et alii,
2006). Biogeographical studies using only fossil taxa are very scarce (i.e., ,
1996; , 2003; et alii,
2005; et alii, in
press) and PAE has been applied to the analysis of fossil assemblages in few studies (i.e., ,
1992; , 1998; & ,
2006). Generally, fossils in Biogeography are a very useful tool in the determination of the minimum age of a taxon, as well as for stratigraphic correlation, the more common objective. In some cases, the stratigraphic distribution seems to be more relevant than the geographic distribution. Certain papers with a biostratigraphic approach allude to the paleobiogeography of taxa over a short interval of time, using quantitative analyses based on similarity to infer the dynamics of their areas of distribution. However, these studies often do not emphasize historical relationships among taxa nor the areas of endemism. Their point of view is merely descriptive insofar as the recognition of spatial distribution patterns is concerned. The determination of the subjacent historical processes is intuitively proposed and depends on the geological data.
Recent papers have applied PAE to the study of faunal distributions, mainly those of extant taxa (i.e., & ,
1999; et alii, 1999; et alii,
2001; , 2001; & ,
2004; & ,
2005; et alii, 2006; et alii,
2006). Biogeographical studies using only fossil taxa are very scarce (i.e., ,
1996; , 2003; et alii,
2005; et alii, in
press) and PAE has been applied to the analysis of fossil assemblages in few studies (i.e., ,
1992; , 1998; & ,
2006). Generally, fossils in Biogeography are a very useful tool in the determination of the minimum age of a taxon, as well as for stratigraphic correlation, the more common objective. In some cases, the stratigraphic distribution seems to be more relevant than the geographic distribution. Certain papers with a biostratigraphic approach allude to the paleobiogeography of taxa over a short interval of time, using quantitative analyses based on similarity to infer the dynamics of their areas of distribution. However, these studies often do not emphasize historical relationships among taxa nor the areas of endemism. Their point of view is merely descriptive insofar as the recognition of spatial distribution patterns is concerned. The determination of the subjacent historical processes is intuitively proposed and depends on the geological data.
![]() The Enchodontoidei are assigned to the Order Aulopiformes and are represented by extinct marine teleostean fishes, generally with an elongate body and a long and narrow strut maxilla included in the mouth gape (,
1994). The Order Aulopiformes consists of fifteen living (Alepisauridae, Aulopidae, Bathysauridae, Bathysauroididae, Bathysauropsidae, Chlorophthalmidae, Evermannellidae, Giganturidae, Ipnopidae, Notosudidae, Paralepididae, Paraulopidae, Pseudotrichonotidae, Scopelarchidae, and Synodontidae) and nine extinct (Apateopholidae, Cimolichthyidae, Dercetidae, Enchodontidae, Eurypholidae,
Halecidae, Ichthyotringidae, Prionolepididae, and Serrilepidae) families of marine fish (,
1994, 2006). They form a morphologically very diversified group known as "lizardfishes", with both benthonic and pelagic habits. They are found in habitats ranging from estuaries to abyssal zones ( & ,
1996; , 2006).
The Enchodontoidei are assigned to the Order Aulopiformes and are represented by extinct marine teleostean fishes, generally with an elongate body and a long and narrow strut maxilla included in the mouth gape (,
1994). The Order Aulopiformes consists of fifteen living (Alepisauridae, Aulopidae, Bathysauridae, Bathysauroididae, Bathysauropsidae, Chlorophthalmidae, Evermannellidae, Giganturidae, Ipnopidae, Notosudidae, Paralepididae, Paraulopidae, Pseudotrichonotidae, Scopelarchidae, and Synodontidae) and nine extinct (Apateopholidae, Cimolichthyidae, Dercetidae, Enchodontidae, Eurypholidae,
Halecidae, Ichthyotringidae, Prionolepididae, and Serrilepidae) families of marine fish (,
1994, 2006). They form a morphologically very diversified group known as "lizardfishes", with both benthonic and pelagic habits. They are found in habitats ranging from estuaries to abyssal zones ( & ,
1996; , 2006).
![]() The Enchodontoidei have a long temporal range: from the Early Cretaceous to the Early Eocene, and were most numerous in the Late Cretaceous. Geographically, they occur in the deposits of Tethyan Europe (Belgium, Czech Republic, England, France, Germany, Holland, Italy, Netherlands, Slovenia, and Sweden); Asia (Arabian Peninsula, India, Israel, Japan, Jordan, Lebanon, and Syria); Africa (Angola, Democratic Republic of Congo, Egypt, and Morocco); North America (Canada, Mexico, and United States), and South America (Bolivia and Brazil). In Brazil, enchodontoids are present in strata ranging in age from Cenomanian to Maastrichtian (i.e., ,
1969; , 1996; ,
2004; & ,
2006).
The Enchodontoidei have a long temporal range: from the Early Cretaceous to the Early Eocene, and were most numerous in the Late Cretaceous. Geographically, they occur in the deposits of Tethyan Europe (Belgium, Czech Republic, England, France, Germany, Holland, Italy, Netherlands, Slovenia, and Sweden); Asia (Arabian Peninsula, India, Israel, Japan, Jordan, Lebanon, and Syria); Africa (Angola, Democratic Republic of Congo, Egypt, and Morocco); North America (Canada, Mexico, and United States), and South America (Bolivia and Brazil). In Brazil, enchodontoids are present in strata ranging in age from Cenomanian to Maastrichtian (i.e., ,
1969; , 1996; ,
2004; & ,
2006).
![]() (2004) proposed a hypothesis on the phylogenetic relationships of Enchodontoidea, that includes some living and extinct members of Aulopiformes. He concluded that Enchodontoidea is a monophyletic group and sister group of Cimolichthyidae, and this clade in turn is the sister group of the living Alepisauridae. In addition, he furnished an area cladogram of Enchodontoidea and suggested generalizations about the paleobiogeography of one of its genera, Enchodus. According to him, Enchodus originated in the mid-Tethyan Ocean (nowadays the Middle East), with some endemic and widespread species recorded in this area. Moreover, one clade comprising three other species is from North America, which is the sister group to a species from the northwestern region of the Tethys Ocean (nowadays western Germany), and this in turn is the sister group of species from the western edge of the Tethys Ocean (nowadays Morocco) and North America.
(2004) proposed a hypothesis on the phylogenetic relationships of Enchodontoidea, that includes some living and extinct members of Aulopiformes. He concluded that Enchodontoidea is a monophyletic group and sister group of Cimolichthyidae, and this clade in turn is the sister group of the living Alepisauridae. In addition, he furnished an area cladogram of Enchodontoidea and suggested generalizations about the paleobiogeography of one of its genera, Enchodus. According to him, Enchodus originated in the mid-Tethyan Ocean (nowadays the Middle East), with some endemic and widespread species recorded in this area. Moreover, one clade comprising three other species is from North America, which is the sister group to a species from the northwestern region of the Tethys Ocean (nowadays western Germany), and this in turn is the sister group of species from the western edge of the Tethys Ocean (nowadays Morocco) and North America.
![]() In the absence of more methodologically sound biogeographical studies of Cretaceous enchodontoids, we applied PAE in order to analyze the distribution of these fishes occurring only in the Cenomanian (the first stage of the Late Cretaceous, with an estimated duration of about 4-5.4 My BP). The present paper is the first step in the paleobiogeographic reconstruction of the Late Cretaceous through the use of this methodology to analyze paleoichthyological assemblages.
In the absence of more methodologically sound biogeographical studies of Cretaceous enchodontoids, we applied PAE in order to analyze the distribution of these fishes occurring only in the Cenomanian (the first stage of the Late Cretaceous, with an estimated duration of about 4-5.4 My BP). The present paper is the first step in the paleobiogeographic reconstruction of the Late Cretaceous through the use of this methodology to analyze paleoichthyological assemblages.
![]() The analysis is based on a data matrix (Appendix 1
The analysis is based on a data matrix (Appendix 1
![]() ) built with taxa (columns) versus areas (rows). The areas are treated as taxa and the taxa as characters (,
1988); character states are the presence/absence of taxa in the "terminal areas". The dataset consists of 16 genera distributed in eight families (i.e., Ichthyotringidae, Apateopholidae, Dercetidae, Prionolepididae, Enchodontidae, Eurypholidae,
Halecidae, and Serrilepidae), and one genus incertae sedis (Yabrudichthys), with most data obtained from the current literature. The genera are
(Appendix 1
) built with taxa (columns) versus areas (rows). The areas are treated as taxa and the taxa as characters (,
1988); character states are the presence/absence of taxa in the "terminal areas". The dataset consists of 16 genera distributed in eight families (i.e., Ichthyotringidae, Apateopholidae, Dercetidae, Prionolepididae, Enchodontidae, Eurypholidae,
Halecidae, and Serrilepidae), and one genus incertae sedis (Yabrudichthys), with most data obtained from the current literature. The genera are
(Appendix 1
![]() ): Ichthyotringa (1), Apateopholis (2), Cyranichthys (3), Dercetoides (4), Hastichthys (5), Rhynchodercetis (6), Prionolepis (7), Rharbichthys (8), Palaeolycus (9), Eurypholis (10), Saurohamphus (11), Enchodus (12), Halec (13), Phylactocephalus (14), Hemisaurida (15), Serrilepis (16), Yabrudichthys (17). The twelve areas are: Brazil, Mexico, United States, Democratic Republic of Congo, Morocco, Italy, Italy-Slovenia, England, Germany, Sweden, Lebanon, and Israel. The occurrences of the Messina and Lesina localities in southern Italy are referred to as "Italy". The occurrences in the Trieste-Komen Plateau are referred to Italy-Slovenia. Localities are listed in the
Appendix 2
): Ichthyotringa (1), Apateopholis (2), Cyranichthys (3), Dercetoides (4), Hastichthys (5), Rhynchodercetis (6), Prionolepis (7), Rharbichthys (8), Palaeolycus (9), Eurypholis (10), Saurohamphus (11), Enchodus (12), Halec (13), Phylactocephalus (14), Hemisaurida (15), Serrilepis (16), Yabrudichthys (17). The twelve areas are: Brazil, Mexico, United States, Democratic Republic of Congo, Morocco, Italy, Italy-Slovenia, England, Germany, Sweden, Lebanon, and Israel. The occurrences of the Messina and Lesina localities in southern Italy are referred to as "Italy". The occurrences in the Trieste-Komen Plateau are referred to Italy-Slovenia. Localities are listed in the
Appendix 2
![]() .
.
![]() The presence of the taxa in the several areas was coded as (1) and their absence as (0). A hypothetical area to root the tree with all the taxa absent was added to the data matrix. The data were analyzed using the computer program PAUP*4.0b10 (,
2001), applying the branch and bound algorithm (BandB) to perform a search of the most parsimonious of all possible tree topologies.
The presence of the taxa in the several areas was coded as (1) and their absence as (0). A hypothetical area to root the tree with all the taxa absent was added to the data matrix. The data were analyzed using the computer program PAUP*4.0b10 (,
2001), applying the branch and bound algorithm (BandB) to perform a search of the most parsimonious of all possible tree topologies.
![]() As a result, 47 trees were obtained with 26 steps, consistency index 0.73, and retention index 0.50. The majority rule consensus tree shows the topology
(Fig. 1
As a result, 47 trees were obtained with 26 steps, consistency index 0.73, and retention index 0.50. The majority rule consensus tree shows the topology
(Fig. 1
![]() ): [(Mexico) + (United States) + (Morocco + Italy) + (Lebanon + Israel) + (Italy-Slovenia) + (Brazil)] + (D.R. Congo) + (Sweden) + (Germany) + (England).
): [(Mexico) + (United States) + (Morocco + Italy) + (Lebanon + Israel) + (Italy-Slovenia) + (Brazil)] + (D.R. Congo) + (Sweden) + (Germany) + (England).
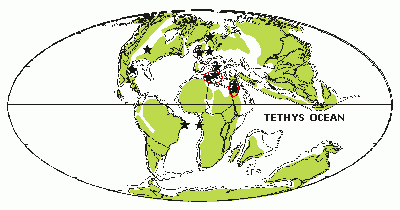
![]() Two areas of endemism can be delimited in the Tethys Ocean: Morocco and southern Italy, Lebanon and Israel
(Figs. 1
Two areas of endemism can be delimited in the Tethys Ocean: Morocco and southern Italy, Lebanon and Israel
(Figs. 1
![]() - 2
- 2 ![]() ). The first group appears in all trees, whereas the second one is present in 60 per cent of them. A unique clade found in 66 per cent of the trees contains these two areas of endemism, as well as four areas with unresolved relationships (i.e., Mexico, United States, Italy-Slovenia, and Brazil). The relationships among the four other areas remain unknown (i.e., D.R. Congo, Sweden, Germany, and England).
). The first group appears in all trees, whereas the second one is present in 60 per cent of them. A unique clade found in 66 per cent of the trees contains these two areas of endemism, as well as four areas with unresolved relationships (i.e., Mexico, United States, Italy-Slovenia, and Brazil). The relationships among the four other areas remain unknown (i.e., D.R. Congo, Sweden, Germany, and England).
![]() The area of endemism formed by Morocco and Italy represents the North African region of the Tethys Ocean, and that formed by Lebanon and Israel is in the mid-Tethyan Ocean
(Fig. 2
The area of endemism formed by Morocco and Italy represents the North African region of the Tethys Ocean, and that formed by Lebanon and Israel is in the mid-Tethyan Ocean
(Fig. 2 ![]() ). These areas are congruent with those we obtained in the application of PAE to only one family of the Enchodontoidei, the Dercetidae, but over a wider temporal range, from the Late Cretaceous to the Paleocene ( & ,
2006).
). These areas are congruent with those we obtained in the application of PAE to only one family of the Enchodontoidei, the Dercetidae, but over a wider temporal range, from the Late Cretaceous to the Paleocene ( & ,
2006).
![]() (1992) accomplished a historical biogeography study of Cenomanian echinoids, applying PAE and cladistic vicariance analysis. Three regional groupings emerged: a North European grouping (England, Belgium, and Hungary), a Tethyan grouping (Algeria, Tunisia, Egypt, the Middle East, and Oman), and a Texas-Sergipe-Angola grouping. Morocco, Portugal, and the Charente remained with variable positions in the area cladograms, possibly explained by a vicariance event: the development of oceanic rifting along the line of the Bay of Biscay. Moreover, during the Cenomanian, an "Atlantic" biota was already differentiated from the North African biota; for the echinoid assemblages of the South Atlantic localities (Sergipe and Angola) are more closely related to those of Texas than to those of North Africa. These data are supported to some extent by our analysis, because Sergipe does not appear related to North Africa (i.e., Morocco), which in our analysis is included in an area of endemism with southern Italy. Furthermore, according to et alii
(1989) and (1992), two subgroups can be separated within North Africa, the eastern and western echinoid faunas. This eastern subgroup may be related to the area of endemism Lebanon + Israel as delimited herein.
(1992) accomplished a historical biogeography study of Cenomanian echinoids, applying PAE and cladistic vicariance analysis. Three regional groupings emerged: a North European grouping (England, Belgium, and Hungary), a Tethyan grouping (Algeria, Tunisia, Egypt, the Middle East, and Oman), and a Texas-Sergipe-Angola grouping. Morocco, Portugal, and the Charente remained with variable positions in the area cladograms, possibly explained by a vicariance event: the development of oceanic rifting along the line of the Bay of Biscay. Moreover, during the Cenomanian, an "Atlantic" biota was already differentiated from the North African biota; for the echinoid assemblages of the South Atlantic localities (Sergipe and Angola) are more closely related to those of Texas than to those of North Africa. These data are supported to some extent by our analysis, because Sergipe does not appear related to North Africa (i.e., Morocco), which in our analysis is included in an area of endemism with southern Italy. Furthermore, according to et alii
(1989) and (1992), two subgroups can be separated within North Africa, the eastern and western echinoid faunas. This eastern subgroup may be related to the area of endemism Lebanon + Israel as delimited herein.
![]() Our results are in agreement with the propositions of &
(1993) and (2000,
2001) regarding gradients of taxonomic diversity. According to these authors, during the Late Cretaceous, peak values of diversity occurred at the 30° 40° N paleolatitudes. The high values of diversity were maintained at 40° 50° N paleolatitudes, which may be explained by the existence of a reef belt there during Late Cretaceous times (,
1995). The areas of endemism delimited herein are situated between the 20° - 30° N paleolatitudes, that is, near one of the paleolatitudes with higher values of diversity as pointed out by &
(1993) and (2001). In this context, Italy-Slovenia could form an area of endemism, because of its latitudinal position (30° N). If data regarding the southern hemisphere obtained during our current study are taken into account, high values of diversity could be placed in paleolatitudes 20° - 30° S. The high values that the above-cited authors found mainly in the northern hemisphere are probably nothing but only a taxonomic artifact produced by the greater emphasis on studies of North American and European taxa (ibid.).
Our results are in agreement with the propositions of &
(1993) and (2000,
2001) regarding gradients of taxonomic diversity. According to these authors, during the Late Cretaceous, peak values of diversity occurred at the 30° 40° N paleolatitudes. The high values of diversity were maintained at 40° 50° N paleolatitudes, which may be explained by the existence of a reef belt there during Late Cretaceous times (,
1995). The areas of endemism delimited herein are situated between the 20° - 30° N paleolatitudes, that is, near one of the paleolatitudes with higher values of diversity as pointed out by &
(1993) and (2001). In this context, Italy-Slovenia could form an area of endemism, because of its latitudinal position (30° N). If data regarding the southern hemisphere obtained during our current study are taken into account, high values of diversity could be placed in paleolatitudes 20° - 30° S. The high values that the above-cited authors found mainly in the northern hemisphere are probably nothing but only a taxonomic artifact produced by the greater emphasis on studies of North American and European taxa (ibid.).
![]() (2000) proposed a reconstruction of the historical biogeography of an extant taxon of Gastropoda, and so delimited the ancestral areas of the clades by using the method of Weighted Ancestral Area Analysis (WAAA). This author correlated the areas he found with the paleogeography postulated for Cretaceous and Tertiary times. Regarding Cenomanian paleogeography (,
2000, fig. 2), two areas are split in the northern continents as a result of a vicariance event. The western area, delimited by the presence of the taxon Helicoidea, is partially congruent with the area of endemism Morocco + Italy described herein.
(2000) proposed a reconstruction of the historical biogeography of an extant taxon of Gastropoda, and so delimited the ancestral areas of the clades by using the method of Weighted Ancestral Area Analysis (WAAA). This author correlated the areas he found with the paleogeography postulated for Cretaceous and Tertiary times. Regarding Cenomanian paleogeography (,
2000, fig. 2), two areas are split in the northern continents as a result of a vicariance event. The western area, delimited by the presence of the taxon Helicoidea, is partially congruent with the area of endemism Morocco + Italy described herein.
![]() Comparing our data with those obtained by
(2004) concerning the paleobiogeography of Enchodus above-cited, we confirm that in strata of Cenomanian age the highest number of occurrences of this genus is in the Middle East (Israel and Lebanon) (see
Appendix 2
Comparing our data with those obtained by
(2004) concerning the paleobiogeography of Enchodus above-cited, we confirm that in strata of Cenomanian age the highest number of occurrences of this genus is in the Middle East (Israel and Lebanon) (see
Appendix 2
![]() ). He concluded that Enchodus originated in the mid-Tethyan Ocean, which is represented now by the Middle East. Only one report of this genus is recorded in the North America and Morocco region, and there are none in Germany during this time. On the other hand, (ibid.) did not include in his analysis the records of the genus in Italy, Italy-Slovenia, and Brazil.
). He concluded that Enchodus originated in the mid-Tethyan Ocean, which is represented now by the Middle East. Only one report of this genus is recorded in the North America and Morocco region, and there are none in Germany during this time. On the other hand, (ibid.) did not include in his analysis the records of the genus in Italy, Italy-Slovenia, and Brazil.
Acknowledgments
![]() This was a contribution to the
"Vth Simpósio Brasileiro de Paleontologia de Vertebrados" (5th Brazilian Symposium on Vertebrate Paleontology), 2-4 August 2006, Santa Maria (RS, Brazil).
This was a contribution to the
"Vth Simpósio Brasileiro de Paleontologia de Vertebrados" (5th Brazilian Symposium on Vertebrate Paleontology), 2-4 August 2006, Santa Maria (RS, Brazil).
![]() We especially want to thank Juan (UNAM) for reviewing the first draft of this article. We are indebted to Mauro J. (UERJ) for critically reading the manuscript and contributing to its improvement. This research has financial support from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and the Fundação Carlos Chagas de à Pesquisa do Estado do Rio de Janeiro (FAPERJ). VG has research fellowship grants from the CNPq (Brazilian Federal Government) and from the "PROCIÊNCIA" (Rio de Janeiro State Government). H.M.A. da has a master fellowships grant from the CNPq. Thanks also to Mireille for the review of the paper and the French translation of the English abstract.
We especially want to thank Juan (UNAM) for reviewing the first draft of this article. We are indebted to Mauro J. (UERJ) for critically reading the manuscript and contributing to its improvement. This research has financial support from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and the Fundação Carlos Chagas de à Pesquisa do Estado do Rio de Janeiro (FAPERJ). VG has research fellowship grants from the CNPq (Brazilian Federal Government) and from the "PROCIÊNCIA" (Rio de Janeiro State Government). H.M.A. da has a master fellowships grant from the CNPq. Thanks also to Mireille for the review of the paper and the French translation of the English abstract.
C. (1954).- Les poissons crétacés du Jebel Tselfat (Maroc).- Notes du Service Géologique du Maroc, Notes et Mémoires, Rabat, vol. 118, 188 p.
C.C. & G.D. (1996).- Interrelationships of Aulopiformes. In: M.L.J.; L.R. & G.D. (eds.), Interrelationships of fishes.- Academic Press, San Diego, p. 335-404.
M., W., G., G., G., L. & G. (2001).- Biogeographic relationships of the Galapagos terrestrial biota: parsimony analysis of endemicity based on reptiles, land birds and Scalesia land plants.- Journal of Biogeography, Oxford, vol. 28, p. 495510.
D.R. & M.G.P. (2003).- Critique of parsimony analysis of endemicity as a method of historical biogeography.- Journal of Biogeography, Oxford, vol. 30, p. 819825.
L. & D.B. (1999).- A new Cenomanian ichthyofauna from southeastern Morocco and its relationships with other early Late Cretaceous Moroccan faunas.- Geologie en Mijnbouw, Amsterdam, vol. 78, p. 261-266.
F., B., T. & H. (2005).- Changes in the ammonite taxonomical diversity gradient during the Late Jurassic-Early Cretaceous.- Journal of Biogeography, Oxford, vol. 32, p. 535-547.
Y. (1985).- Saurorhamphus judeaensis (Salmoniformes: Enchodontidae), a new longirostrine fish from the Cretaceous (Cenomanian) of Ein-Yabrud, near Jerusalem.- Journal of Vertebrate Paleontology, Northbrook, vol. 5, n° 3, p. 181-193.
Y. (1989).- Yabrudichthys and Serrilepis, two new genera of enchodontoids (Teleostei) from Lower Cenomanian beds of Ein-Yabrud, Israel.- Israel Journal of Zoology, Jerusalem, vol. 36, p. 11-38.
Y. (1996).- New species of Enchodus (Aulopiformes: Enchodontidae) from the Northern Negev, Israel, with comments on evolutionary trends in the Enchodontoidei. In: G. & H.-P. (eds.), Mesozoic fishes - Systematics and paleoecology, 1.- Verlag Dr. F. Pfeil, München, p. 349-367.
J.A. (2000).- Evolution of taxonomic diversity gradients in marine realm: evidence of Recent bivalve faunas.- Paleobiology, Chicago, vol. 26, p. 188-214.
J.A. (2001).- Taxonomic diversity gradients through geological time.- Diversity and Distributions, Oxford, vol. 7, p. 175-189.
C. (1996).- A Late Cretaceous (Turonian) ichthyofauna from Lac des Bois, Northwest Territories, Canada, with paleobiogeographic comparisons with Turonian ichthyofaunas of the Western Interior Seaway.- Canadian Journal of Earth Sciences, Ottawa, vol. 33, p. 1375-1389.
C. (2004).- The phylogenetic relationships of the Enchodontidae (Teleostei: Aulopiformes). In: G., M.V.H. & R. (eds.), Recent advances in the origin and early radiation of vertebrates.- Verlag Dr. F. Pfeil, München, p. 619-634.
C. & K.A. (2005).- A new ichthyotringoid from the El Doctor Formation (Cretaceous: Albian-Cenomanian), Zimapán, Hidalgo, Mexico. In: F.J. (ed.), Fourth International Meeting on Mesozoic Fishes. Systematics, homology, and nomenclature, extended abstracts.- Ediciones Universidad Autónoma de Madrid, p. 91-93.
F.J. & V. (2006).- A new dercetid fish (Neoteleostei: Aulopiformes) from the Turonian of the Pelotas basin, southern Brazil.- Palaeontology, London, vol. 49, n° 2, p. 445-456.
P.L., L., C. & C.E. (2003).- Fossil fishes from the Cenomanian (Upper Cretaceous) of Namoura, Lebanon.- Journal of Systematic Palaeontology, Cambridge, vol. 1, n° 4, p. 227330.
V. & H.M.A. (2006).- Análise de parcimônia de endemismo para a família Dercetidae (Teleostei, Aulopiformes).- Paleontologia em Destaque, Porto Alegre, v. Ano 21, n° 53, p. 33-34.
V., M.J. & H.M.A. (2007).- Track analysis of the marine palaeofauna from the Turonian (Late Cretaceous).- Journal of Biogeography, Oxford.
D. (1998).- Biogeography of circum-Mediterranean Miocene-Pliocene rodents; a revision using factor analysis and parsimonious analysis of endemicity.- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, vol. 137, p. 273288.
C.J. & B. (1999).- Distribution patterns and biogeographic analysis of Austral Polychaeta (Annelida).- Journal of Biogeography, Oxford, vol. 26, p. 507533.
P.C. (1969).- The relationships of certain Upper Cretaceous teleosts with special reference to the myctophoids.- Bulletin of British Museum (Natural History), Geology, London, Supplement 7, 255 p.
P.C. (1976).- Enchodus (Teleostei: Enchodontidae) from the Upper Cretaceous Pierre Shale of Wyoming and South Dakota with an evaluation of the North American enchodontid species.- Palaeontographica, Abteilung A, Stuttgart, vol. 152, p. 91-112.
S. de (2001).- Biogeography of Indo-Pacific Pontoniinae (Crustacea, Decapoda): a PAE analysis.- Journal of Biogeography, Oxford, vol. 28, p. 1239-1253.
B. (2000). Biogeography of the Limacoidea sensu lato (Gastropoda: Stylommatophora): vicariance events and long-distance dispersal.- Journal of Biogeography, Oxford, vol. 27, p. 379390.
C.J. & L.R. (1999).- Cladistic biogeography: Interpreting patterns of plant and animal distributions. 2nd ed.- Oxford Biogeography Series, Oxford, n° 12, 187 p.
A. (1966).- L'ittiofauna cenomaniana di Floresta-Messina.- Palaeontographia Italica, Pisa, vol. 60, n° 30, p. 33-67.
B.S. (2003).- Paleobiogeography: the relevance of fossils to biogeography.- Annual Review of Ecology and Systematics, Palo Alto, vol. 34, p. 51-69.
B. (1889).- Om kritfaunan vid Tormarp i Halland och de halländska kritbildningarnes förhallande till öfriga svenska.- Geologiska Föreningens i Stockholm Förhandlingar, Stockholm, vol. 11, n° 121, p. 63-72.
R.A., C., M.M., M.A. & N. (2006).- Patterns of endemism in south-eastern Pacific benthic polychaetes of the Chilean coast.- Journal of Biogeography, Oxford, vol. 33, p. 750759.
J.J. & J.V. (1995).- Historical biogeography: Introduction to methods.- Annual Review of Ecology and Systematics, Palo Alto, vol. 26, p. 373-401.
J.J., D., C. & J. (1999).- Preliminary classification of the Mexican biogeographic provinces: a parsimony analysis of endemicity based on plant, insect, and bird taxa.- Southwestern Naturalist, San Angelo, vol. 44, p. 508515.
A.M. (2000).- The Palaeozoic, Mesozoic and Early Cenozoic fishes of Africa.- Fish and Fisheries, Edinburgh, vol. 1, p. 111-145.
J.S. (1994).- Fishes of the World.- 3rd ed., John Wiley and Sons, New York, 600 p.
J.S. (2006).- Fishes of the World.- 4th ed., John Wiley and Sons, New Jersey, 601 p.
S.S. (2006).- Misconceptions about parsimony analysis of endemicity.- Journal of Biogeography, Oxford, vol. 33, p. 2099-2106.
M.A., R. & D.R. (2006).- Areas of endemism and distribution patterns for Neotropical Piper species (Piperaceae).- Journal of Biogeography, Oxford, vol. 33, p. 12661278.
M. & Y. (1987).- A new enchodontid fish genus from the Upper Cenomanian of Jerusalem, Israel.- Palaeontology, London, vol. 30, n° 4, p. 717-731.
L. & T. (2004).- Patterns of Amazonian area relationships based on raw distributions of papilionid butterflies (Lepidoptera: Papilioninae).- Biological Journal of the Linnean Society, London, vol. 82, p. 345357.
D.M. & D. (1993).- Geography of end-Cretaceous marine bivalve extinctions.- Science, Washington, vol. 260, p. 971-973.
J., J., J.-P. & C. (1989).- Les échinoides du Crétacé et du Paléogene du Dhofar (Sultanat d'Oman) et les relations entre les bassins de l'Océan Indien et de la Méditerranée.- Bulletin de la Société Géologique de France, Paris, 8e série, t. V, n° 2, p. 279-286.
B.R. (1988).- From fossils to earth history: applied historical biogeography. In: A.A. & P.S. (eds.), Analytical biogeography.- Chapman and Hall, London, p. 437-481.
Y. & I. (2005).- Zoogeography of the shallow-water holothuroids of the western Indian Ocean.- Journal of Biogeography, Oxford, vol. 32, p. 15231538.
C.R. (2002).- PALEOMAP website: http://www.scotese.com/
A.B. (1992).- Echinoid distribution in the Cenomanian: an analytical study in biogeography.- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, vol. 92, p. 263276.
S.M. (1995).- New horizons for paleontology, with two examples: the rise and fall of Cretaceous Supertethys and the cause of the modern ice age.- Journal of Paleontology, Tulsa, vol. 69, p. 999-1007.
D.L. (2001).- PAUP* Phylogenetic Analysis Using Parsimony (* and other methods), version 4.0b10.- Sinauer Associated, Sunderland.
L. (1987).- Ostéologie de Cyranichthys ornatissimus nov. gen. du Cénomanien du Zaïre et de Rhynchodercetis yovanovitchi du Cénomanien de l'Afrique du Nord. Les relations intergénériques et la position systématique de la famille néocrétacique marine des Dercetidae (Pisces, Teleostei).- Annales du Musée Royal de l'Afrique Centrale, Tervuren, Rapport annuel, vol. 1985-1986, p. 93-112.
Click on thumbnail to enlarge the image.
Figure 1: Majority rule consensus tree of the 47 most parsimonious cladograms obtained in the PAE from the enchodontoid distribution during Cenomanian.
Click on thumbnail to enlarge the image.
Figure 2: Paleocontinental map of the Cenomanian showing the localities used in the analysis (represented by stars) and the areas of endemism delimited in the PAE (encircled stars). Base map modified from (2002).
Taxa |
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 |
|
Areas |
|||||||||||||||||
|
Hypothetical |
0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
|
Brazil |
0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
|
Mexico |
1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
|
United States of America |
1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
|
Democratic Republic of Congo |
0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
|
Morocco |
1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
|
Italy |
0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 |
|
Italy-Slovenia |
0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 |
|
England |
0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
|
Germany |
0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
|
Sweden |
0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
|
Lebanon |
1 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 0 |
|
Israel |
0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 1 |
Appendix 1: Data matrix of taxa (columns) x areas (rows) of Enchodontoidei from the Cenomanian used in Parsimony Analysis of Endemicity (PAE).
| Taxa | Provenance | Country |
| Apateopholis | Haqel and En Nammoura | Lebanon |
| Cyranichthys | Kipala | Democratic Republic of Congo |
| Dercetoides | Ein-Yabrud | Israel |
| Enchodus | Ein-Yabrud and Givat-Shaul | Israel |
| Haqel, Hgula, and En Nammoura | Lebanon | |
| Sage Creek | United States | |
| Messina | Italy | |
| Trieste-Komen | Italy-Slovenia | |
| Sergipe | Brazil | |
| Jbel Tselfat | Morocco | |
| Eurypholis | Haqel, Hgula, and En Nammoura | Lebanon |
| English Chalk | England | |
| Halec | Lesina | Italy |
| Trieste-Komen | Italy-Slovenia | |
| Hastichthys | Ein-Yabrud | Israel |
| En Nammoura | Lebanon | |
| Hemisaurida | Trieste-Komen | Italy-Slovenia |
| Haqel | Lebanon | |
| Ichthyotringa | Haqel and Hgula | Lebanon |
| Jbel Tselfat | Morocco | |
| Niobrara | United States | |
| Hidalgo | Mexico | |
| Palaeolycus | Sedenhorst | Germany |
| Phylactocephalus | Haqel and Hgula | Lebanon |
| Prionolepis | Haqel and Hgula | Lebanon |
| Tormarp | Sweden | |
| Rharbichthys | Jbel Tselfat | Morocco |
| Messina | Italy | |
| Rhynchodercetis | Jbel Tselfat and Daoura | Morocco |
| Messina | Italy | |
| Haqel, Hgula, and En Nammoura | Lebanon | |
| Trieste-Komen | Italy-Slovenia | |
| Saurohamphus | Trieste-Komen | Italy-Slovenia |
| Ein-Yabrud | Israel | |
| Serrilepis | Ein-Yabrud | Israel |
| En Nammoura | Lebanon | |
| Yabrudichthys | Ein-Yabrud | Israel |
Appendix 2: Occurrences of enchodontoids in the Cenomanian included in the analysis. Most of data from: (1889); (1954); (1966); (1969, 1976); (1985, 1989, 1996); & (1987); (1987); & (1999); (2000); et alii (2003); & (2005).