![]()
Sommaire
[Introduction] [Étude paléontologique]
[Diversité et évolution ...]
[Conclusion]
[Références bibliographiques]
8 rue de Garambault, 45190 Beaugency (France)
La Guille, 38700 Sarcenas (France)
Manuscrit en ligne depuis le 28 Septembre 2008
![]()
L'étude de plus de 2200 spécimens de Phylloceratoidea (Cephalopoda, Ammonoidea) récoltés dans les dépôts aptiens et albiens du bassin vocontien (Sud-Est de la France) a permis l'identification et la description de 28 taxons repérés stratigraphiquement à l'échelle de la zone d'ammonite. Quatre nouveaux taxons sont décrits : Phylloceras (Hypophylloceras) moriezense , 1920 tenuicostulata nov. subsp. (base de l'Aptien supérieur), Phylloceras (Goretophylloceras) vocontium nov. sp. (sommet de l'Aptien moyen à base de l'Albien inférieur), Salfeldiella (Gyrophyllites) falloti nov. sp. (sommet de l'Aptien inférieur à base de l'Aptien supérieur), Phyllopachyceras brehereti nov. sp. (base de l'Aptien moyen à base de l'Aptien supérieur).
Ammonites ; Phylloceratoidea ; Crétacé ; Aptien ; Albien ; France ; bassin vocontien.
B. & M. (2008).- Les Phylloceratoidea (Ammonoidea) aptiens et albiens du bassin vocontien (Sud-Est de la France).- Carnets de Géologie / Notebooks on Geology, Brest, Mémoire 2008/04 (CG2008_M04)
Aptian and Albian Phylloceratids (Ammonoidea) from the Vocontian Basin (SE France).- More than 2,200 pyritized ammonites of the Superfamily Phylloceratatoidea have been collected in consonance with the scale of the ammonite zonation of the expanded Marnes Bleues Fm in the Vocontian Basin (SE France). This abundant material allows the description and the figuration (specimens and sutures) of 28 taxa of which 4 are new:
Owing to the abundance of the Phylloceratoidea and the variations of specific diversity in 9 fossiliferous levels ranging in age from the late Early Aptian to the end of the Albian it is possible to identify 3 peaks of abundance:
Of these 3 peaks the first is the most important probably because the Vocontian Basin attained its maximum depth during this time. The peaks appear to be correlated with the maximum flooding intervals of depositional sequences. But high sea-levels were not always favourable for the Phylloceratoidea. The very fossiliferous level, although it took place during a high in sea-level, is characterised by the virtual absence of the deep-water ammonites, e.g. Phylloceratids, Tetragonids and Lytoceratids. This fact is explained by the presence of a deep-water anoxia. Another depletion of Phylloceratoidea in conjonction with a great abundance of Desmoceratid and Mortoniceratid ammonites and benthic fauna (Inoceramids Bivalves) occurred during the early Late Albian. At that time, the Vocontian area appears to have reached a minimal depth resulting from a pause in the rate of subsidence associated with an influx of siliciclastic material that attained the center of the basin.
After a last episode during the Early Cenomanian of the Vocontian Basin when the Phylloceratoidea occurred in significant numbers, this group of ammonites shows a drastic decrease in abundance. This evolution is correlated with the filling of the basin from west to east, thus restricting the Phylloceratoidea and the other deep-water ammonites to a small eastern area (Leiostraca area of , 1965, 1980).
Ammonites; Phylloceratoidea; Cretaceous; Aptian; Albian; France; Vocontian Basin.
Dans le Sud-Est de la France (Drôme, Alpes-de-Haute-Provence, Hautes-Alpes et
Alpes-Maritimes), les étages Aptien et Albien sont représentés par des marnes à ammonites pyriteuses donnant des paysages de "bad-lands" caractéristiques (Fig. 1 ![]() ). Afin de distinguer ces marnes crétacées des marnes jurassiques
("Terres Noires" du Bathonien-Oxfordien), il est d'usage, suite aux travaux de cartographie (,
1963), de les
nommer, de manière informelle, "formation des Marnes
Bleues Vocontiennes" (abrégé dans la suite du texte en Marnes
Bleues).
). Afin de distinguer ces marnes crétacées des marnes jurassiques
("Terres Noires" du Bathonien-Oxfordien), il est d'usage, suite aux travaux de cartographie (,
1963), de les
nommer, de manière informelle, "formation des Marnes
Bleues Vocontiennes" (abrégé dans la suite du texte en Marnes
Bleues).
Relativement peu étudiées sur le plan stratigraphique et paléontologique lorsqu'on les compare aux alternances marno-calcaires de l'intervalle Berriasien-Barrémien car longtemps considérées comme monotones et pauvres en horizons fossilifères, les Marnes Bleues renferment, outre des niveaux à ammonites pyriteuses surtout présents dans leur partie aptienne, de nombreux niveaux riches en empreintes d'ammonites et plus rarement en nodules calcaréo-phosphatés (, 1983, 1988, 1995 ; et alii, 1986).
Conséquence du caractère pélagique dominant de la sédimentation à certaines périodes (Aptien moyen, limite Aptien-Albien, fin de l'Albien supérieur), l'ammonitifaune des Marnes Bleues est dominée par une association de formes de haute mer (Lytoceratoidea, Phylloceratoidea et Tetragonitoidea) caractéristiques du biome pélagique téthysien.
À l'aide d'un abondant matériel repéré stratigraphiquement à l'échelle de la zone d'ammonite, cette étude s'attachera à faire connaître la diversité des Phylloceratoidea des Marnes Bleues et leur répartition stratigraphique.

Cliquer sur la vignette pour agrandir l'image.
Figure 1 : Expression géomorphologique des Marnes Bleues du bassin vocontien d'après l'affleurement des
Briers
(n° 16 sur Fig. 2 ![]() - Abréviations : Fig. 5
- Abréviations : Fig. 5 ![]() ).
).
Situé sur la marge européenne de l'océan alpin, appendice de la Téthys
occidentale, le Bassin vocontien (anciennement nommé "fosse vocontienne")
constitue, du Lias au début du Crétacé supérieur, une sorte de golfe pélagique d'orientation ouest-est entouré sur trois côtés de
plates-formes ( & ,
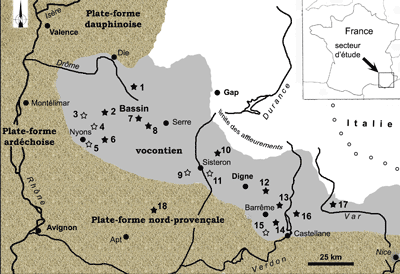
1977) : Fig. 2 ![]() .
.
Cliquer sur la vignette pour agrandir l'image.
Figure 2 : Situation géographique et cadre paléogéographique du bassin
vocontien
(Étoiles = affleurements avec Phylloceratoidea avec en noir matériel figuré dans cette
étude ; les numéros renvoient à la liste des affleurements de la Fig. 4 ![]() ).
).
Réputé pour ces alternances marno-calcaires du Crétacé inférieur souvent riches en ammonites présentes dans de nombreux musées de par le monde, le Bassin vocontien accueille, du sommet de l'Aptien inférieur (Zone à Deshayesi) à la base du Cénomanien (Zone à Mantelli), une puissante formation de marnes sombres (300 à 700 m).
Cette sédimentation détritique argilo-carbonatée, parfois rythmée, est entrecoupée de niveaux riches en matière organique (black shales) et de faisceaux marno-calcaires (, 1995). Des resédimentations silicoclastiques en provenance des plates-formes environnantes et des glissements synsédimentaires liés à des pentes circa- et intra-bassinales perturbent l'enregistrement pélagique (, 1982, 1988 ; , 1987 ; , 2002 ; & , 2003).
L'existence d'ammonites pyriteuses dans les Marnes Bleues est reconnue depuis plus de deux siècles (,
1779) mais les premières figurations paléontologiques correctes semblent être celles de
(1831) avec création d'une nouvelle espèce de Phylloceratoidea (Salfeldiella guettardi :
Fig. 3 ![]() ).
Cependant, c'est dans le célèbre ouvrage "Paléontologie française" que d'
(1840-1842) aborde véritablement
l'ammonitifaune des Marnes
Bleues. Dans cet ouvrage, réédité récemment en édition critique (,
2006), d' décrit et figure 21 ammonites pyriteuses aptiennes dont 2 nouvelles espèces de
Phylloceratoidea. Pour les ammonites albiennes, il faut attendre le travail de
(1885) qui
décrit, en annexe de son travail stratigraphique, trois espèces de Tetragonitoidea rattachées actuellement aux genres Kosmatella et Zelandites.
).
Cependant, c'est dans le célèbre ouvrage "Paléontologie française" que d'
(1840-1842) aborde véritablement
l'ammonitifaune des Marnes
Bleues. Dans cet ouvrage, réédité récemment en édition critique (,
2006), d' décrit et figure 21 ammonites pyriteuses aptiennes dont 2 nouvelles espèces de
Phylloceratoidea. Pour les ammonites albiennes, il faut attendre le travail de
(1885) qui
décrit, en annexe de son travail stratigraphique, trois espèces de Tetragonitoidea rattachées actuellement aux genres Kosmatella et Zelandites.
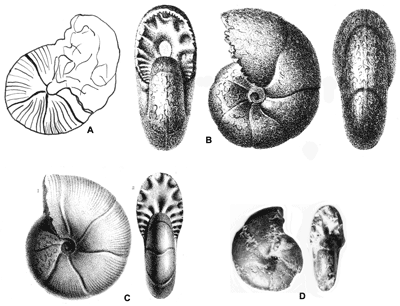
Cliquer sur la vignette pour agrandir l'image.
Figure 3 : Figurations historiques d'un Phylloceratoidea des Marnes Bleues : Salfeldiella guettardi.
A : , 1779, Pl. 10, dessin schématique d'après fig. 2.
B : , 1830, Pl. 2, fig. 8.
C : d', 1841, Pl. 53, fig. 1-2.
D : néotype désigné par in ,
2006, p. 61,
Pl. 31, fig. 7.a-b.
Bien qu'ayant effectué la première étude stratigraphique détaillée, (1900) n'a figuré aucune ammonite des Marnes Bleues. C'est à (1892) que l'on doit la publication des premières photographies [Phylloceras (Goretophylloceras) fortunei (= goreti) et Argonauticeras depereti] suivie une décennie plus tard par les figurations de la monographie de (1908) surtout consacrée aux faunes phosphatées du "Crétacé moyen" des plates-formes environnantes.
Durant plus d'un demi siècle, malgré les travaux paléontologiques thématiques de (1920), (1920a) et les études stratigraphiques ponctuelles de (1937, 1939), & (1954), l'ammonitifaune des Marnes Bleues ne fait l'objet d'aucun travail liant stratigraphie et études paléontologiques.
La première étude stratigraphique moderne avec figurations de coupes (mais sans figuration des ammonites déterminées par ) apparaît dans le travail micropaléontologique de (1966). À la même époque, sur la base d'abondantes récoltes effectuées pour l'essentiel dans la partie orientale du bassin vocontien (Alpes-de-Haute-Provence et Alpes-Maritimes), (1960, 1963, 1964) donne des listes de faunes et propose un découpage zonal de l'Aptien mais toujours sans figuration de matériel (à l'exception, en 1968, du rare Lytoceratidae Argonauticeras). Ce même auteur proposera en 1986 une nouvelle zonation pour l'Aptien moyen ( & , 1986) et publiera en 1987 (, 1987b) une courte monographie illustrant 7 espèces de Phylloceratoidea de l'Aptien inférieur et moyen.
Suite aux recherches sédimentologiques conduites par entre 1983 et 1995 (avec détermination des ammonites par ), les Marnes Bleues se sont révélées beaucoup plus fossilifères que ce que ne le laissaient supposer les quelques niveaux pyriteux connus depuis longtemps. Grâce au lever détaillé de nombreuses coupes, ce chercheur repère et nomme un grand nombre d'horizons fossilifères tel le célèbre black shale du "niveau " (, 1983 ; et alii, 1986). Ce remarquable niveau fossilifère singulièrement presque dépourvu de Phylloceratoidea (voir cependant fig. 45-i in et alii, 2000) et dont l'extension dépasse largement le cadre du bassin vocontien (, 1985), avait été reconnu dès 1900 par et sa faune décrite partiellement par (1937).
Cette richesse paléontologique révélée par impulsa un très net regain d'intérêt pour l'ammonitifaune des Marnes Bleues avec notamment la recherche de coupes de références pour les limites d'étages Albien/Cénomanien ( et alii, 1996) et Aptien/Albien ( et alii, 2000). Actuellement, les dernières recherches se concentrent sur la signification environnementale des variations de populations d'ammonites ( et alii, 2005 ; et alii, 2005) ainsi que sur la stratigraphie de l'Aptien ( & , 2002, 2005 ; , 2005).
L'accompagnement des recherches sédimentologiques de par l'un de nous (MD) a permis la récolte dans les Marnes Bleues d'un peu plus de 6000 spécimens d'ammonites bien repérées stratigraphiquement (Collection ). Nous donnons dans la figure 4 la liste des affleurements du matériel étudié ici et nous renvoyons, pour la localisation géographique précise des affleurements, à la liste figurant dans les annexes de la thèse de (1995). Le matériel de cette étude (ainsi que l'ensemble des ammonites des Marnes Bleues en cours d'étude) sera déposé ultérieurement dans un Muséum d'Histoire naturelle (Genève ou Grenoble).
| N° sur Fig. 2 | Gisement | Matériel figuré | Commune | Département |
| 1 | Serre-Chaïtieu | X | Lesches-en-Diois | Drôme |
| 2 | Bâtie | Arnayon | Drôme | |
| Bert | X | Arnayon | Drôme | |
| Châtelat | X | Arnayon | Drôme | |
| Pré-Guittard | Arnayon | Drôme | ||
| 3 | Pissourier | Valouse | Drôme | |
| 4 | Pouet | Saint-Ferréol-Trente-Pas | Drôme | |
| 5 | Saint-Quénin | Châteauneuf-de-Bordette | Drôme | |
| 6 | Bartalasse | X | Bellecombe-Tarendol | Drôme |
| Terrois | X | Bellecombe-Tarendol | Drôme | |
| 7 | Folie | X | Bruis | Hautes-Alpes |
| 8 | Risou | X | Moydans | Hautes-Alpes |
| 9 | Houlettes | Sisteron | Alpes-de-Haute-Provence | |
| Sous-Les-Roches | Saint-Vincent-sur-Jabron | Alpes-de-Haute-Provence | ||
| 10 | Gaubert | X | Bayons | Alpes-de-Haute-Provence |
| 11 | Coignets | Salignac | Alpes-de-Haute-Provence | |
| 12 | Chênebuis | X | Tartonne | Alpes-de-Haute-Provence |
| Condamine | Tartonne | Alpes-de-Haute-Provence | ||
| Crouas | X | Clumanc | Alpes-de-Haute-Provence | |
| Fumiours | Clumanc | Alpes-de-Haute-Provence | ||
| Pillart | X | Tartonne | Alpes-de-Haute-Provence | |
| Sauzeries | X | Clumanc | Alpes-de-Haute-Provence | |
| Vallons | X | Tartonne | Alpes-de-Haute-Provence | |
| 13 | Aps | X | Moriez | Alpes-de-Haute-Provence |
| 14 | Pin | X | Senez | Alpes-de-Haute-Provence |
| 15 | Bourbens | Blieux | Alpes-de-Haute-Provence | |
| 16 | Briers | X | Saint-André-Les-Alpes | Alpes-de-Haute-Provence |
| 17 | Prés | X | Saint-Léger | Alpes-Maritimes |
| Rivet | X | Croix-sur-Roudoule | Alpes-Maritimes | |
| 18 | Montsalier | X | Montsalier | Vaucluse |
Figure 4 : Liste des affleurements du matériel étudié et figuré.
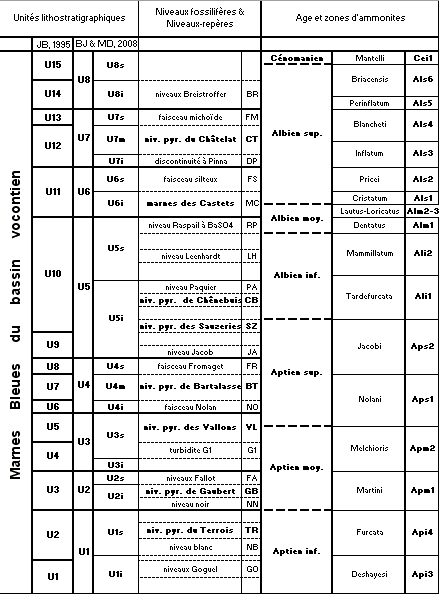
Cet abondant matériel joint aux levers lithostratigraphiques a conduit à l'adoption du cadre stratigraphique pour les Marnes Bleues vocontiennes présenté dans le tableau de la
Figure 5 ![]() .
.
Cliquer sur la vignette pour agrandir l'image.
Figure 5 : Cadre stratigraphique des Marnes Bleues du bassin vocontien (découpage zonal adapté d'après , 2005 et , 2002, 2008). NPy : niveaux à ammonites pyriteuses.
Quelques représentants d'ammonites caractéristiques des huit unités lithostratigraphiques informelles distinguées dans le tableau de la
Figure 5 ![]() sont illustrés dans les Figures 6
sont illustrés dans les Figures 6 ![]() ,
7
,
7 ![]() et 8
et 8 ![]() .
.

Cliquer sur la vignette pour agrandir l'image.
Figure 6 : Ammonites caractéristiques des unités U1, U2 et U3
(Aptien inférieur-Aptien moyen).
(Abréviations stratigraphiques : Fig. 5 ![]() ; barre échelle : 1 cm)
; barre échelle : 1 cm)
A : Eogaudryceras intermedium (, 1920b) - MD478 - U1s/TR : Zone à
Furcata.
B : Macroscaphites striatisulcatus (d',
1841) - MD1161 - U1s/TR : Zone à
Furcata.
C : Chelinoceras crassum , 1930 - MD283 - U1s/TR : Zone à
Furcata.
D : Gargasiceras aptiense (, 1927) - MD766 - U1s : base Zone à Martini.
E : Eotetragonites kossmatelliformis (,
1920b) - MD686 - U2i/GB : Zone à Martini.
F : Vergunniceras pretiosum (d', 1841) - MD2161 - U2i/GB : base de la Zone à Martini.
G : Epichelinoceras cf. tschernyschewi (, 1906) - MD303 - U2s/FA : sommet de la Zone à Martini.
H : Parahoplites sp. juv.- MD1393 - U3s/VL : Zone à Melchioris.
I : Eotetragonites raspaili (,
1947) - MD527 - U3s/VL : base de la Zone à
Melchioris.
J : Eotetragonites duvalianum (d',
1841) - MD742 - U3s/G1-NO : Zone à
Melchioris.
K : Valdedorsella angladei (, 1890) - MD2128 - U2i/GB : Zone à Martini.
L : Zurcherella zurcheri ( & , 1906) - MD2235 - U3s/VL : sommet de la Zone à
Melchioris.

Cliquer sur la vignette pour agrandir l'image.
Figure 7 : Ammonites caractéristiques des unités U4, U5 et U6
(Aptien supérieur-Albien supérieur).
(Abréviations stratigraphiques : Fig. 5 ![]() ; barre échelle : 1 cm)
; barre échelle : 1 cm)
A : Ephamulina aff. trituberculata , 1962 - MD466 - U4m/BT : Zone à
Nolani.
B : Valdedorsella akuskaensis (, 1899) - MD2265 - U4m/BT : Zone à
Nolani.
C : Chaschupseceras aff. caucasicum ( & , 1983) - MD282 - U4m/BT : Zone à
Nolani.
D : Nolaniceras nolani (, 1887) - MD122 - U4m/BT : Zone à
Nolani.
E : Neosilesites balearensis (, 1920b) - MD1355 -
U5i/SZ : sommet de la Zone à Jacobi.
F : Pseudosilesites aff. seranoniformis ,
1969 - MD1816 -
U5i/SZ : sommet de la Zone à Jacobi.
G : Hypacanthoplites clavatus (, 1906) - MD991 - U5i/CB : base de la Zone à
Tardefurcata.
H : Hypacanthoplites elegans (, 1906) - MD2368 -
U5i/SZ : sommet de la Zone à Jacobi.
I : Eotetragonites cf. heterosulcatus (,
1899) - MD2074 - U5i/PL : Zone à Jacobi.
J : Beudanticeras convergens (, 1908) - MD261 - U5i/CB : base de la Zone à
Tardefurcata.
K : Leymeriella tardefurcata (d',
1841) - MD1148 - base U5s : milieu de la Zone à
Tardefurcata.
L : Neoleymeriella cf. renascens (, 1930) - MD1145 - base U5s : sommet de la Zone à
Tardefurcata.
M : Metahamites sablieri (d', 1842) - MD1299 - U5s/RP : Zone à
Dentatus.
N : Turrilitoides densicostatus (, 1930) - MD2117 - U6i/MC : base de la Zone à
Lautus.
O : Mojsisovicsia cf. equicostata (,
1931) - MD1303 - U6i/MC : base de la Zone à
Lautus.
P : Eoscaphites aff. circularis (J.C. , 1836) - MD523 - U6s/FS : Zone à
Pricei.
Q : Dipoloceras (Rhytidoceras) elegans (, 1933) - MD779 - U6s/FS : Zone à
Pricei.

Cliquer sur la vignette pour agrandir l'image.
Figure 8 : Ammonites caractéristiques des unités U7 et U8
(Albien supérieur).
(Abréviations stratigraphiques : Fig. 5 ![]() ; barre échelle : 1 cm)
; barre échelle : 1 cm)
A : Beudanticeras beudanti (, 1822) - MD249 - U7i/ DP : Zone à
Inflatum.
B : Goodhallites goodhalli (, 1934) - MD773 -
U7i/DP : Zone à Inflatum.
C : Hysteroceras orbignyi (, 1922) - MD1003 -
U7i/DP : Zone à Inflatum.
D : Hysteroceras binum (J. , 1815) - MD1027 -
U7m/CT : Zone à Blancheti.
E : Anisoceras saussureanum (, 1848) - MD198 -
U7m/CT : Zone à Pricei.
F : Cantabrigites minor , 1933 - MD280 -
U7m/CT : Zone à Blancheti.
G : Scaphites meriani & , 1861 - MD2042 -
U8i/BR : Zone à Perinflatum.
H : Hemiptychoceras subgaultinum , 1940 - MD860 -
U8i/BR : Zone à Perinflatum.
I : Turrilitoides hugardianus (d',
1842) - MD2116 -
U7m/CT : Zone à Blancheti.
J : Turrilitoides hugardianus (d',
1842) - MD2120 -
U7m/CT : Zone à Blancheti.
K : Paraturrilites gresslyi ( & , 1861) - MD1181 - U7s/FM : Zone à
Blancheti.
L : Ostlingoceras puzosianum (d',
1842) - MD1371 -
U8i/BR : Zone à Perinflatum.
M : Stolickaia dispar (d', 1841) - MD2061 (flèche en fin de loge) - U8i : Zone à
Perinflatum.
N : Zelandites dozei (, 1885) - MD2182 -
U7m/CT : Zone à Blancheti.
O : Tetragonites collignoni (, 1940) - MD2080 -
U7m/CT : Zone à Blancheti.
P : Tetragonites rectangularis (, 1962) - MD2115 -
U7m/CT : Zone à Blancheti.
Q : Tetragonites timotheanum (, 1848) - MD2100 -
U7m/CT : Zone à Blancheti.
R : Tetragonites jurinianus (, 1848) - MD2077 -
U8s/CT : Zone à Perinflatum.
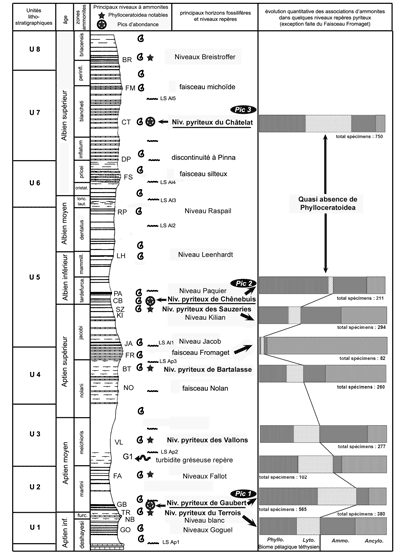
La situation stratigraphique des Phylloceratoidea étudiés ici est donnée dans la colonne lithologique synthétique de la série pélagique empruntée à
(1995, p. 114, fig. 48). En regard de cette colonne est figurée l'évolution quantitative des peuplements au cours du temps
(Fig. 9 ![]() ).
).
Cliquer sur la vignette pour agrandir l'image.
Figure 9 : Série synthétique des Marnes Bleues : principaux niveaux à Phylloceratoidea et évolution quantitative des populations d'ammonites (adaptée d'après , 1995 et & , 2003).
La classification adoptée a été développée au cours d'une récente étude sur les Phylloceratoidea de France au Jurassique et au Crétacé (, 2000a). L'auteur a mis en évidence l'importance des processus d'altération du développement à l'origine d'au moins deux rameaux à deux périodes différentes. Le premier rameau, connu essentiellement au Jurassique, comprend deux sous-familles, les Calliphylloceratinae et les Ptychophylloceratinae, les processus d'altération du développement étant plus prononcés chez les Calliphylloceratinae que chez les Ptychophylloceratinae. Le second épisode d'altération du développement, plus récent, concerne principalement un autre rameau, la sous-famille des Neocalliphylloceratinae au Crétacé. Cette sous-famille est représentée par de nombreux taxons dans les Marnes Bleues vocontiennes.
La prise en compte des phénomènes d'altération de la croissance permet de comprendre pourquoi certains taxons conservent des constrictions sur les tours adultes et d'autres non. Les premiers sont issus des seconds par processus pédomorphique. Ainsi les Salfeldiella sont issus des Phylloceras (Goretophylloceras) par altération du développement. Par leur origine on ne peut les assimiler à des Calliphylloceras ou encore à des Holcophylloceras, voire même à des Sowerbyceras comme le font encore parfois certains auteurs. Il n'est d'ailleurs pas toujours aisé de distinguer les petits spécimens du genre Salfeldiella de ceux du sous-genre Goretophylloceras, voire du sous-genre Hypophylloceras, si l'on ne tient compte que des caractères morphologiques. La connaissance du tracé des lignes cloisonnaires apporte de précieux arguments pour résoudre le délicat problème des affinités entre taxons.
L'abondance du matériel étudié (plus de 2200 spécimens) a rendu possible une étude statistique des principaux taxons, surtout précieuse lorsque les populations paraissent
proches. Le choix a été fait de présenter les résultats sous forme d'histogrammes des fréquences des paramètres dimensionnels
relatifs. En effet, l'expérience montre que ce sont les meilleurs critères pour l'étude des populations, au moins chez les
Phylloceratoidea. En toute rigueur, il faudrait constituer des échantillons avec des individus
contemporains. Il est bien évident que respecter cette condition est le plus souvent impossible et les échantillons étudiés sont constitués d'individus d'âges divers. Ce qui rend tout de même utilisables les résultats de l'étude statistique est la stabilité de l'espèce dans le temps (,
2000b). Un exemple pris chez les Phyllopachyceratinae (Fig. 10 ![]() ) montre qu'une population de P. baborense de
l'Aptien moyen représente correctement la population baborense
globale. C'est évidemment une conséquence de la stabilité de l'espèce dans le temps pouvant expliquer pourquoi il est difficile de mettre en évidence des différences morphologiques entre individus d'âge différent (,
2000b, équilibres
ponctués, p. 715). Nous verrons un certain nombre d'exemples d'histogrammes des fréquences des paramètres relatifs mettant bien en évidence l'existence de taxons différents
(espèces ou sous-espèces).
) montre qu'une population de P. baborense de
l'Aptien moyen représente correctement la population baborense
globale. C'est évidemment une conséquence de la stabilité de l'espèce dans le temps pouvant expliquer pourquoi il est difficile de mettre en évidence des différences morphologiques entre individus d'âge différent (,
2000b, équilibres
ponctués, p. 715). Nous verrons un certain nombre d'exemples d'histogrammes des fréquences des paramètres relatifs mettant bien en évidence l'existence de taxons différents
(espèces ou sous-espèces).
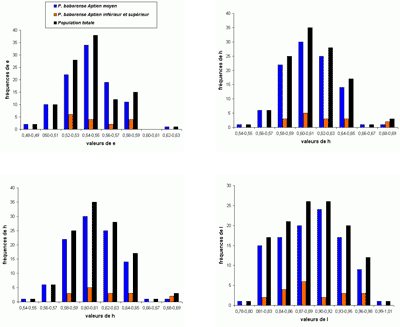
Cliquer sur la vignette pour agrandir l'image.
Figure 10 : Histogrammes des fréquences de e, h, o et l chez Phyllopachyceras baborense. Les dimensions des spécimens sont exprimées en mm. Les paramètres mesurés sont : D = diamètre, H = hauteur du tour (h = H/D), E = épaisseur du tour (e = E/D), O = diamètre de l'ombilic (o = O/D), l = E/H.
Famille des Phylloceratidae , 1884
Sous-famille des Phylloceratinae , 1884
Genre Phylloceras , 1865
Sous-genre Hypophylloceras , 1924
Le genre Euphylloceras , 1953, repris par & (2006, p. 26), est généralement considéré comme synonyme de Phylloceras (Hypophylloceras), point de vue adopté dans cette note. En effet, les Phylloceratidae du Crétacé ne sont pas très différents des Phylloceras sensu stricto du Lias et du Jurassique. Il est préférable de conserver le genre Phylloceras au Crétacé afin de tenir compte de l'uniformité de ce rameau de la famille. L'événement évolutif le plus important, au Crétacé, a été la profonde modification de la ligne suturale qui justifie la création de la famille des Neophylloceratidae , 1993.
Phylloceras (Hypophylloceras) aptiense , 1920
| 1920 | Phylloceras aptiense , p. 195, Pl. 1, fig. 4-4a. | |
| 2000a | Phylloceras (Hypophylloceras) aptiense .- , p. 127, Pl. 31, figs. 7.a-b & 8.a-b ; Pl. 32, fig. 1 ; figs. 265-269 dans le texte. | |
| 2005 | Phylloceras (Hypophylloceras) aptiense .- , p. 44, Pl. 1, figs. 8-11 ; Pl. 2, figs. 1-2. | |
| Synonymie plus complète, holotype, localité-type, âge de l'holotype : , 2000a, p. 127 et 129. | ||
Matériel étudié : 13 spécimens collection MD1506, 1507, 1508, 1509, 1510, 1511 et 1512.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1506 | 27,5 | 15,6 | 10,4 | 2,2 | 0,57 | 0,38 | 0,08 | 0,67 |
| 1509 | 31,6 | 16,3 | 11,9 | 2,3 | 0,52 | 0,38 | 0,07 | 0,73 |
| 1512a | 13,2 | 6,8 | 5,4 | 2,2 | 0,51 | 0,41 | 0,17 | 0,79 |
| 1512b | 14,4 | 8,1 | 5,8 | 2,4 | 0,56 | 0,40 | 0,17 | 0,72 |
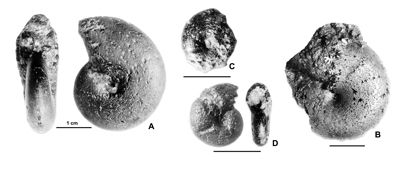
Cliquer sur la vignette pour agrandir l'image.
Figure 11 : Phylloceras (Hypophylloceras) aptiense ,
1920.
A : MD1509 - U2i/GB : Aptien moyen (Zone à Martini) - Gaubert, Hautes-Alpes.
B : MD1506 - U7m/CT : Albien supérieur (Zone à Blancheti) Ravine de Bert, Drôme.
C : MD1512a - U5i/SZ : Aptien supérieur (Zone à Jacobi) Les Sauzeries, Alpes-de-Haute-Provence.
D : MD1512b - U5i/SZ : Aptien supérieur (Zone à Jacobi) - Les Sauzeries, Alpes-de-Haute-Provence.
Description : les spécimens étudiés sont tous des moules internes pyritisés ne portant aucune trace d'ornementation, involutes, à flancs convexes et section du tour ovale.
Plusieurs spécimens portent des constrictions : l'un des plus grands (MD1507 de l'Aptien supérieur) a une constriction au début du tour, effacée, presque radiale sur les flancs, un peu proverse sur le bord ventral. Les autres, plus petits (Fig. 11.C-D ![]() ), montrent environ trois fortes constrictions radiales au début du dernier tour. Leur aspect et leurs dimensions sont très proches du spécimen figuré par
(1964, Pl. 12, fig. 3). Les autres spécimens vocontiens ont les dimensions de l'espèce P. (H.) aptiense révisée récemment (,
2000a, p. 129).
), montrent environ trois fortes constrictions radiales au début du dernier tour. Leur aspect et leurs dimensions sont très proches du spécimen figuré par
(1964, Pl. 12, fig. 3). Les autres spécimens vocontiens ont les dimensions de l'espèce P. (H.) aptiense révisée récemment (,
2000a, p. 129).
La ligne cloisonnaire (Fig. 12 ![]() ) montre des selles S1 diphylles et S2 triphylles avec un développement assez notable des phyllites terminaux internes pour les selles S1 et même S2. Ces caractères s'observent également chez le spécimen le plus récent (Fig. 11.B
) montre des selles S1 diphylles et S2 triphylles avec un développement assez notable des phyllites terminaux internes pour les selles S1 et même S2. Ces caractères s'observent également chez le spécimen le plus récent (Fig. 11.B ![]() ) et ses dimensions sont très proches de celles de l'holotype (,
2000a, p. 129).
) et ses dimensions sont très proches de celles de l'holotype (,
2000a, p. 129).
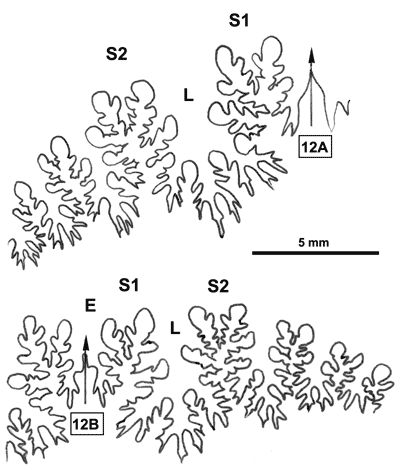
Cliquer sur la vignette pour agrandir l'image.
Figure 12 : Lignes cloisonnaires de Phylloceras (Hypophylloceras) aptiense.
A : MD1509 U2i : Aptien moyen (Zone à Martini) Gaubert, Hautes-Alpes.
B : MD1510a U2s : Aptien moyen (Zone à Martini) Serre Chaïtieu, Drôme.
Affinités : l'espèce la plus proche de P. (H.) aptiense est sans doute P. (H.) morelianum. Elle en diffère cependant par sa plus grande épaisseur et le tracé de sa ligne cloisonnaire dont les selles S1 et S2 sont plus élancées que chez P. (H.) morelianum (voir par exemple d', 1841, Pl. 54, fig. 3, et , 2000a, fig. 280, p. 133). Il n'en est pas moins vrai que les deux espèces sont très proches, peut-être la première n'est-elle qu'une sous-espèce de la seconde ? C'est peu vraisemblable étant donné l'allure différente des lignes cloisonnaires.
P. (H.) morelianum est plus rare que P. (H.) aptiense et la plupart des mentions de l'espèce morelianum doivent en réalité se rattacher à l'espèce aptiense.
Âge : les spécimens vocontiens ont été récoltés de la base de l'Aptien moyen (Zone à Martini) à l'Albien supérieur (Zone à Blancheti). (2005) la signale avec doute dans la fin de l'Aptien inférieur (Zone à Deshayesi). Jusqu'à présent, l'espèce était connue du "Gargasien" à l'Albien moyen (, 2000a, p. 129).
Phylloceras (Hypophylloceras) cypris & , 1923
| 1923 | Phylloceras cypris & , p. 22, Pl. 3, fig. 1, 2.c, non 2.a-b ; fig. 4 dans le texte. | |
| 2000a | Phylloceras (Hypophylloceras) cypris & .- , p. 129, Pl. 32, fig. 3.a-b & 4.a-b ; figs. 270-273 dans le texte. | |
| 2005 | Phylloceras (Hypophylloceras) cypris & .- , p. 52, Pl. 3, figs. 1-4 | |
| Synonymie plus complète, lectotype, localité-type, âge du lectotype : , 2000a, p. 129. | ||
Matériel étudié : 34 spécimens collection MD1513, 1514, 1515, 1516, 1517, 1518, 1519, 1520, 1521 (non mesuré), 1522, 1523, 1524, 1525, 2469, 2474, 2475a-f, 2476 et 2477.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1513 | 12,7 | 7,0 | 4,9 | 2,2 | 0,55 | 0,39 | 0,17 | 0,70 |
| 1522 | 16,9 | 10,0 | - | 1,2 | 0,59 | - | 0,07 | - |
| 15,0 | 8,0 | 6,0 | 1,1 | 0,53 | 0,40 | 0,07 | 0,75 |
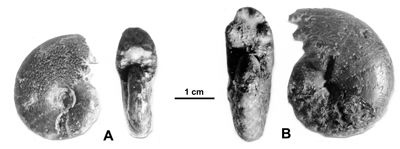
Cliquer sur la vignette pour agrandir l'image.
Figure 13 : Phylloceras (Hypophylloceras) cypris & ,
1923.
A : MD1513 U4m/BT : Aptien supérieur (Zone à Nolani) - Les Vallons, Alpes-de-Haute-Provence.
B : MD1522 U5i/CB : Albien inférieur (Zone à Tardefurcata) - Aps sud, Alpes-de-Haute-Provence.
Description : les spécimens étudiés sont des moules internes pyritisés, de petite taille (D de 15 à 20 mm environ), ne présentant aucun des replis observables chez les plus grands spécimens (D de 20 à 25 mm environ). Ils ont l'allure des petits spécimens de P. (H.) aptiense, en particulier par la présence de constrictions au début du tour. Mais chez les spécimens rattachés à P. (H.) cypris les constrictions les plus nettes disparaissent plus tôt, vers 12 mm de diamètre et l'ombilic est moins ouvert, à diamètre égal, que chez P. (H.) aptiense. Quelques spécimens ont été récemment étudiés (, 2000a, p. 129). Quand le test est conservé il est orné de fines costules en faisceaux. Quant aux dimensions des spécimens de la présente étude, elles sont proches de celles du lectotype avec en particulier un ombilic modérément ouvert. Cela s'explique par leur petite taille du même ordre que celle du lectotype (désigné par , 1964, p. 217, D = 11,3 mm). Comme on l'observe chez beaucoup de Phylloceras l'ombilic tend à se fermer au cours de la croissance.
La ligne cloisonnaire (Fig. 14 ![]() ) est reconnaissable mais les selles n'ont pas toujours atteint le stade tétraphylle (voir aussi & ,
1923, fig. 4, p. 22), ce stade est atteint chez un fragment de grand spécimen (MD1516).
) est reconnaissable mais les selles n'ont pas toujours atteint le stade tétraphylle (voir aussi & ,
1923, fig. 4, p. 22), ce stade est atteint chez un fragment de grand spécimen (MD1516).
Cliquer sur la vignette pour agrandir l'image.
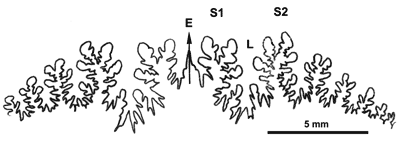
Figure 14 : Ligne cloisonnaire de Phylloceras (Hypophylloceras) cypris.
MD1518a U3s/G1-NO : Aptien moyen (Zone à Melchioris) Gaubert, Alpes-de-Haute-Provence.
Affinités : les petits spécimens de P. (H.) cypris se distinguent difficilement des petits spécimens de l'espèce P. (H.) moriezense dont ils ont la morphologie et la ligne cloisonnaire. En revanche, les individus de plus grande taille se différencient aisément par leur ornementation (costules en faisceaux sur les flancs pour P. (H.) cypris, costules régulières, toutes égales, localisées sur le bord ventral pour P. (H.) moriezense) et la forme de la section (flancs convexes chez P. (H.) cypris, méplat sur les flancs chez P. (H.) moriezense).
Âge : les récoltes vocontiennes de P. (H.) cypris s'échelonnent du sommet de l'Aptien inférieur (Zone à Furcata) à l'Albien supérieur (limite Zone à InflatumZone à Blancheti). L'espèce était connue auparavant du "Gargasien" à l'Albien moyen (, 2000a, p. 130).
Phylloceras (Hypophylloceras) cypris cytherae , 1964
| 1964 | Phylloceras (Hypophylloceras) cypris & , 1923, cytherae , 1964, p. 218, Pl. 12, figs. 2 & 7 ; Pl. 15, fig. 8 ; fig. 51.a-b dans le texte. | |
| 2000a | Phylloceras (Hypophylloceras) cypris & , 1923, cytherae , 1964. , p. 130, Pl. 32, fig. 2.a-b ; figs. 274-275 dans le texte. | |
| Synonymie plus complète, holotype, localité-type, âge de l'holotype : , 2000a, p. 130. | ||
Matériel étudié : 8 spécimens collection MD1526, 1527, 1528, 2443, 2478 et 2479.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1526a | 22,7 | 13,6 | - | - | 0,60 | - | - | - |
| 20,8 | 12,6 | 7,0 ? | 1,5 | 0,61 | 0,34 ? | 0,07 | 0,56 ? | |
| 1528 | 19,9 | 13,0 | 7,7 | 1,7 | 0,65 | 0,39 | 0,08 | 0,59 |
| 2443 | 18,0 | - | - | - | - | - | - | - |
| 14,7 | 8,6 | 5,3 | 2,1 | 0,58 | 0,36 | 0,14 | 0,61 |
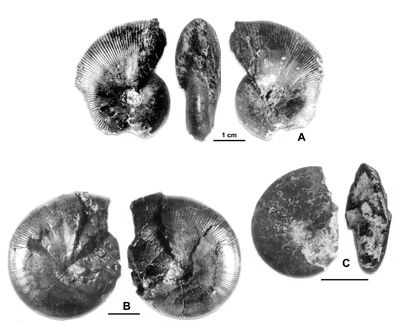
Cliquer sur la vignette pour agrandir l'image.
Figure 15 : Phylloceras (Hypophylloceras) cypris cytherae ,
1964.
A : MD1528 U3s/FA-NO, Aptien moyen (Zone à Melchioris) - Aps nord, Alpes-de-Haute-Provence.
B : MD1526a U1s/TR, Aptien inférieur (Zone à Furcata) - Ravin du Rivet,
Alpes-Maritimes.
C : MD2443 U3s, Aptien moyen (Zone à Melchioris) - Ravin des Prés, Alpes-de-Haute-Provence.
Description : le spécimen MD1527 est un moule interne lisse ; les deux autres (Fig. 15.A-B ![]() ) ont encore leur test bien conservé présentant une belle ornementation. Les costules partent de l'ombilic réunies en faisceaux d'une dizaine, le départ des faisceaux est souligné par des replis. Les costules sont un peu infléchies sur les flancs avec convexité tournée vers l'avant, elles se redressent avant de passer le bord ventral radialement ou un peu infléchies vers l'avant. Les costules et les replis sont assez discrets du début jusqu'au milieu du dernier tour, leur relief s'accentue ensuite. La coquille des spécimens des Marnes Bleues rapportées à P. (H.) cypris cytherae est plus comprimée que celle de P. (H.) cypris sensu stricto. Les spécimens étudiés sont très semblables aux illustrations de
(1964,
Pl. 12, figs. 2 & 7 et Pl. 15, fig. 8).
) ont encore leur test bien conservé présentant une belle ornementation. Les costules partent de l'ombilic réunies en faisceaux d'une dizaine, le départ des faisceaux est souligné par des replis. Les costules sont un peu infléchies sur les flancs avec convexité tournée vers l'avant, elles se redressent avant de passer le bord ventral radialement ou un peu infléchies vers l'avant. Les costules et les replis sont assez discrets du début jusqu'au milieu du dernier tour, leur relief s'accentue ensuite. La coquille des spécimens des Marnes Bleues rapportées à P. (H.) cypris cytherae est plus comprimée que celle de P. (H.) cypris sensu stricto. Les spécimens étudiés sont très semblables aux illustrations de
(1964,
Pl. 12, figs. 2 & 7 et Pl. 15, fig. 8).
La ligne cloisonnaire du moule interne du spécimen MD1527 est de type diphylle-triphylle comme celle figurée par pour un petit spécimen de l'Albien inférieur des Baléares (1964, fig. 51.a dans le texte).
Âge : les récoltes vocontiennes de P. (H.) cypris cytherae s'échelonnent du sommet de l'Aptien inférieur (Zone à Furcata) à l'Aptien terminal (sommet de la Zone à Jacobi). Ce taxon n'était jusqu'alors connu qu'au "Gargasien" (, 2000a, p. 130) ; son extension est donc moins importante que celle de P. (H.) cypris sensu stricto.
Phylloceras (Hypophylloceras) delanoyi , 2000a
| V | 1995 | Phylloceras (Hypophylloceras) ponticuli () plicatum & , p. 125, Pl. 1, fig. 5 ; Pl. 2, fig. 1.a-b ; Pl. 3, fig. 1. |
| 2000a | Phylloceras (Hypophylloceras) delanoyi , p. 130, Pl. 33, figs. 1-2. | |
| 2005 | Salfeldiella (Gyrophyllites) paquieri ().- , p. 72, Pl. 6, figs. 1-2 (non Pl. 5, figs. 1-8 = paquieri) | |
| Synonymie plus complète, holotype, localité-type, âge de l'holotype : , 2000a, p. 131. | ||
Matériel étudié : 9 spécimens collection MD1529, 1530, 1531 et 1532.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1529a | 47,7 | 26,8 | 15,4 | 4,6 | 0,56 | 0,32 | 0,10 | 0,57 |
| 1529b | 51,2 | 30,4 | 15,0 | 5,8 | 0,59 | 0,29 | 0,11 | 0,49 |
| 1532 (1) | 39,0 | 23,3 | 15,8 | 3,0 | 0,60 | 0,41 | 0,08 | 0,68 |
| (1) forme plus épaisse | ||||||||
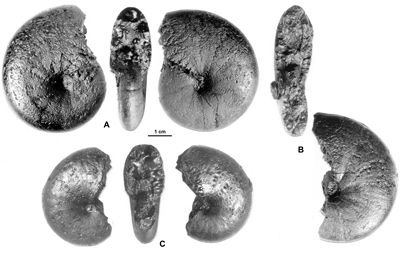
Cliquer sur la vignette pour agrandir l'image.
Figure 16 : Phylloceras (Hypophylloceras) delanoyi ,
2000a.
A : MD1529a U1s/TR, Aptien inférieur (Zone à Furcata) - Ravin du Terrois, Drôme.
B : MD1529b U1s/TR, Aptien inférieur (Zone à Furcata) - Ravin du Terrois, Drôme.
C : MD1532 U1s/TR, Aptien inférieur (Zone à Furcata) - Ravin du Terrois, Drôme.
Description : les spécimens rapportés à cette espèce sont des moules internes comprimés pyritisés. Plusieurs présentent la costulation caractéristique figurée par (2000a, p. 132, Pl. 33, figs. 1-2) : fines costules réunies en faisceaux, replis à la naissance des faisceaux sur les flancs, les replis sont localisés sur les flancs jusqu'au bord ventral mais sans le franchir, les costules franchissent le bord ventral. Les flancs sont à peine convexes et la section du tour est elliptique haute.
a figuré deux spécimens déterminés P. (H.) paquieri (2005, Pl. 6, figs. 1-2). Ces spécimens ont une ornementation de costules en faisceaux caractéristique de l'espèce P. (H.) delanoyi.
La ligne cloisonnaire (Fig. 17 ![]() ) reste de type diphylle-triphylle même chez les spécimens les plus grands, les phyllites internes ne sont jamais assez développés pour atteindre la tétraphyllie.
) reste de type diphylle-triphylle même chez les spécimens les plus grands, les phyllites internes ne sont jamais assez développés pour atteindre la tétraphyllie.
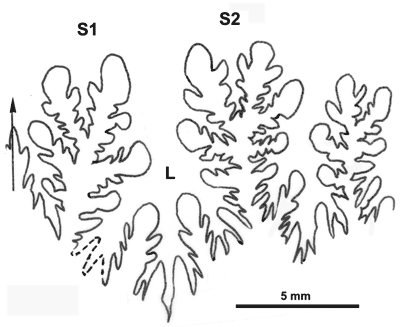
Cliquer sur la vignette pour agrandir l'image.
Figure 17 : Ligne cloisonnaire de Phylloceras (Hypophylloceras) delanoyi.
MD1529b U1s/TR : Aptien inférieur (Zone à Furcata) Ravin du Terrois, Drôme.
Âge : P. (H.) delanoyi était jusqu'alors connue du Barrémien à l'Aptien inférieur (, 2000a, p. 131). Les récoltes vocontiennes étendent sa présence jusqu'au début de l'Aptien moyen (base de la Zone à Martini).
Phylloceras (Hypophylloceras) moriezense , 1920
| V | 1920 | Phylloceras moriezense , p. 194, Pl. 1, figs. 2-2.a & 3. |
| 2000a | Phylloceras (Hypophylloceras) moriezense .- , p. 134, Pl. 32, figs. 10.a-b & 11 ; Pl. 33, figs. 3-4 ; figs. 282-286 dans le texte. | |
| 2005 | Phylloceras (Hypophylloceras) moriezense .- , p. 47, Pl. 1, figs. 12-13. | |
| Synonymie plus complète, holotype, localité-type, âge de l'holotype : , 2000a, p. 134. | ||
Matériel étudié : 9 spécimens collection MD1534, 1535, 1536, 1537, 1538 et 1539.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1534 | 17,2 | 10,3 | 6,3 | 2,0 | 0,60 | 0,37 | 0,12 | 0,61 |
| 1536a | - | 13,8 | 9,0 | - | - | - | - | 0,65 |
| 1536b | - | 20,0 | 14,0 | - | - | - | - | 0,70 |
| 1537a | 29,2 | 17,1 | 11,6 | 2,4 | 0,59 | 0,40 | 0,08 | 0,68 |
Cliquer sur la vignette pour agrandir l'image.
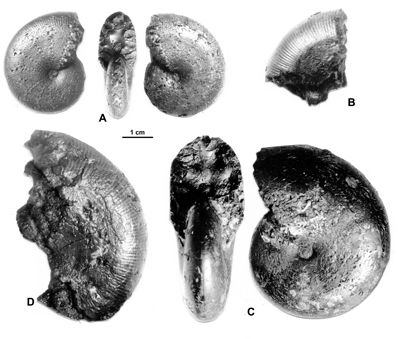
Figure 18 : Phylloceras (Hypophylloceras) moriezense ,
1920.
A : MD1534 U7m/CT, Albien supérieur (Zone à Blancheti) - Ravine de Bert, Drôme.
B : MD1536a U4m/BT, Aptien supérieur (Zone à Nolani) - Bartalasse, Drôme.
C : MD1537a U2i/GB, Aptien moyen (Zone à Martini) - Ravin du Terrois, Drôme.
D : MD1536b - U4m/BT, Aptien supérieur (Zone à Nolani) - Bartalasse, Drôme.
Description : tous les spécimens sont des moules internes pyritisés à flancs plats et ombilic peu ouvert. Lorsque la costulation est visible, elle n'apparaît que sur le bord ventral, les costules n'étant jamais présentes sur la partie plate des flancs au moins chez les spécimens les plus proches de l'holotype.
Les deux spécimens de l'Albien supérieur (MD1534 : Fig. 18.A ![]() et MD1539) sont de petite taille mais leurs flancs sont plats et l'un d'eux laisse apercevoir la costulation ventrale (MD1539), la costulation est très nette chez le spécimen MD1534. Aucun des spécimens n'a un méplat à bords aussi nets que chez l'holotype de l'espèce (,
2000a, Pl. 33, fig. 4), mais le méplat est toujours présent. Le méplat n'est pas très apparent chez les spécimens figurés par
(2005, Pl. 1, figs. 12-13). Donc il semblerait que le méplat des flancs ne soit pas toujours bien apparent au moins chez les petits spécimens.
et MD1539) sont de petite taille mais leurs flancs sont plats et l'un d'eux laisse apercevoir la costulation ventrale (MD1539), la costulation est très nette chez le spécimen MD1534. Aucun des spécimens n'a un méplat à bords aussi nets que chez l'holotype de l'espèce (,
2000a, Pl. 33, fig. 4), mais le méplat est toujours présent. Le méplat n'est pas très apparent chez les spécimens figurés par
(2005, Pl. 1, figs. 12-13). Donc il semblerait que le méplat des flancs ne soit pas toujours bien apparent au moins chez les petits spécimens.
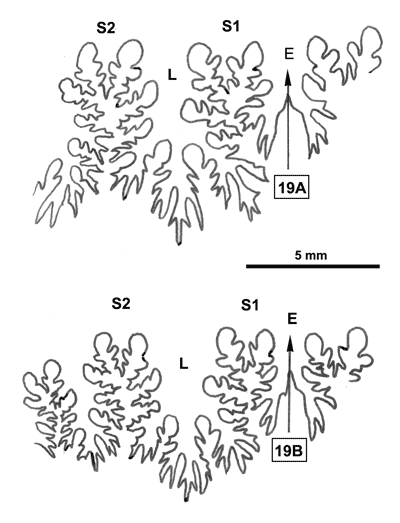
La ligne cloisonnaire (Fig. 19 ![]() ) est de type diphylle-triphylle avec cependant un certain développement des phyllites internes des selles S1 et S2.
) est de type diphylle-triphylle avec cependant un certain développement des phyllites internes des selles S1 et S2.
Cliquer sur la vignette pour agrandir l'image.
Figure 19 : Lignes cloisonnaires de Phylloceras (Hypophylloceras) moriezense.
A : MD1534 U7m/CT : Albien supérieur (Zone à Blancheti) Ravine de Bert, Drôme.
B : MD1537a U2i/GB : Aptien moyen (Zone à Martini) Ravin du Terrois, Drôme.
Âge : P. (H.) moriezense n'était jusqu'alors connue que dans le "Gargasien" (, 2000a, p. 135). L'espèce est désormais connue du sommet de l'Aptien inférieur (Zone à Deshayesi) à l'Albien supérieur (Zone à Blancheti).
Phylloceras (Hypophylloceras) moriezense , 1920 tenuicostulata nov. subsp.
Holotype : collection MD1540.
Localité : Les Vallons, commune de Tartonne, Alpes-de-Haute-Provence.
Âge : début de l'Aptien supérieur : base de la Zone à Nolani.
Origine du nom : en référence à la présence de très fines costules.
Dimensions du spécimen figuré
| N° MD | D | H | E | O | h | e | o | l |
| 1540 | 37,5 | 22,5 | 17,5 | 3,1 | 0,60 | 0,47 | 0,08 | 0,78 |
| 32,0 | 18,6 | 13,8 | 3,5 | 0,58 | 0,43 | 0,11 | 0,74 |
Cliquer sur la vignette pour agrandir l'image.
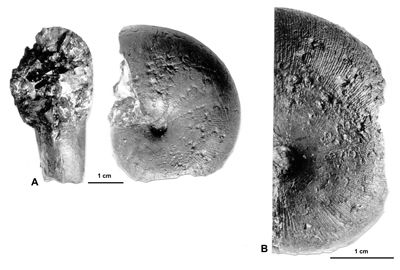
Figure 20 : Phylloceras (Hypophylloceras) moriezense ,
1920 tenuicostulata nov. subsp.
A : MD1540 U4m/BT, Aptien supérieur (Zone à Nolani) - Les Vallons, Alpes-de-Haute-Provence.
B = fig. A agrandie pour le détail de l'ornementation.
Diagnose : variant de P. (H.) moriezense caractérisé par une section du tour subrectangulaire et un méplat moins marqué que chez la forme typique et montrant une fine costulation descendant un peu sur les flancs.
Description : Parmi les spécimens rattachés à P. (H). moriezense, l'un d'entre eux a les flancs plats de l'espèce mais sa costulation commence vers le milieu des flancs. Les costules, mal conservées, sont visibles par endroits sur le moule, elles sont extrêmement fines. Ces caractères sont assez éloignés de ceux du lectotype, la ligne cloisonnaire et la morphologie du spécimen cependant restent proches de celles de P. (H.) moriezense.
Âge : Le nouveau taxon P. (H.) moriezense tenuicostulata est connu à la base de l'Aptien supérieur (Zone à Nolani).
Phylloceras (Hypophylloceras) paquieri , 1896
| 1890 | Phylloceras aff. P. ernesti .- , p. 12, Pl. 1, fig. 2.a-b. | |
| 1896 | Phylloceras paquieri in , p. 765. | |
| V | 1920 | Phylloceras paquieri , p. 198, Pl. 1, figs. 8-8.a, 9-9.a- b, 10-10.a & 11. |
| 2000a | Phylloceras (Hypophylloceras) paquieri .- , p. 135, Pl. 33, figs. 5.a-c, 6.a-c, 7.a-b & 8.a-b ; fig. 287-292 dans le texte. | |
| 2005 | Salfeldiella (Gyrophyllites) paquieri .- , p. 72, Pl. 5, figs. 1-8, non Pl. 6, figs. 1-2 (= delanoyi) | |
| ? | 2005 | Salfeldiella (Gyrophyllites) lateumbilicatum ().- , p. 69, Pl. 5, figs. 9-11 |
| 2006 | Hypophylloceras paquieri (, 1920).- & , p. 96, Pl. 9, figs. 5-9 (figs. 5-7 = Hypophylloceras onoense ?, figs. 8-9 = Hypophylloceras sp. ?) | |
| Synonymie plus complète, lectotype, localité-type, âge du lectotype : , 2000a, p. 135. | ||
Matériel étudié : 14 spécimens collection MD1541, 1542, 1543, 1544, 1545, 1546, 1547, 1548 et 1549.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1544a | 19,3 | 10,1 | 7,0 | 3,0 | 0,52 | 0,36 | 0,16 | 0,69 |
| 1544b (1) | 7,8 | 3,5 | 3,3 | 2,9 | 0,37 | 0,42 | 0,37 | 0,94 |
| 1547 | 38,5 | 20,3 | 11,3 | 6,0 | 0,53 | 0,29 | 0,16 | 0,56 |
| 1548a (1) | 9,9 | 4,7 | 3,6 | 3,0 | 0,47 | 0,36 | 0,30 | 0,76 |
| 1549 | 49,1 | 27,5 | 16,1 | 6,2 | 0,56 | 0,33 | 0,13 | 0,58 |
| (1) nuclei | ||||||||
Cliquer sur la vignette pour agrandir l'image.
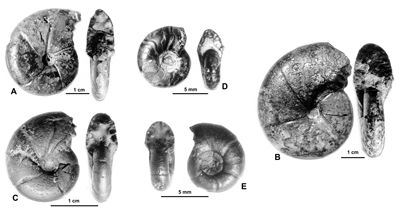
Figure 21 : Phylloceras (Hypophylloceras) paquieri ,
1896.
A : MD1547 U4m/BT : Aptien moyen (Zone à Melchioris) - Les Vallons, Alpes-de-Haute-Provence.
B : MD1549 - U3s/VL : Aptien moyen (Zone à Melchioris) - Les Vallons, Alpes-de-Haute-Provence.
C : MD1544a U1s/?TR : Aptien inférieur (Zone à Furcata) - Montsalier, Vaucluse.
D : MD1548a U1s/TR : Aptien inférieur (Zone à Furcata) - Ravin du Rivet, Alpes-Maritimes.
E : MD1544b U1s/?TR : Aptien inférieur (Zone à Furcata) - Montsalier, Vaucluse.
Description : la plupart des spécimens sont des moules internes pyritisés comprimés à flancs assez aplatis. Les spécimens les plus petits ont des constrictions profondes, d'abord infléchies vers l'avant sur les flancs, elles se redressent avant de passer le bord ventral dirigées vers l'arrière (MD1541 par exemple). Chez les plus grands spécimens (Fig. 21.A-B ![]() ) les constrictions sont encore bien marquées sur les flancs mais presque effacées sur le bord ventral, elles sont presque droites sur les flancs mais proverses au passage du bord ventral. On compte 5 à 6 constrictions sur le dernier tour. Lorsque le test est conservé il montre une fine costulation bien visible à la partie externe des flancs et sur le bord ventral.
) les constrictions sont encore bien marquées sur les flancs mais presque effacées sur le bord ventral, elles sont presque droites sur les flancs mais proverses au passage du bord ventral. On compte 5 à 6 constrictions sur le dernier tour. Lorsque le test est conservé il montre une fine costulation bien visible à la partie externe des flancs et sur le bord ventral.
Un spécimen (Fig. 21.E ![]() à D = 7,8 mm) est très comparable à la sous-espèce S. (G.) lateumbilicatum pygmaeum créée par
(1964, Pl. 17, fig. 6). C'est un petit moule interne, lisse, presque évolute. Le dernier tour porte 5 constrictions proverses mais un peu convexes vers l'avant sur les flancs, elles se redressent pour passer le bord ventral radialement. L'ombilic est très ouvert. Un autre petit spécimen (Fig. 21.D
à D = 7,8 mm) est très comparable à la sous-espèce S. (G.) lateumbilicatum pygmaeum créée par
(1964, Pl. 17, fig. 6). C'est un petit moule interne, lisse, presque évolute. Le dernier tour porte 5 constrictions proverses mais un peu convexes vers l'avant sur les flancs, elles se redressent pour passer le bord ventral radialement. L'ombilic est très ouvert. Un autre petit spécimen (Fig. 21.D ![]() à D = 8,8 mm) a une morphologie assez proche de pygmaeum quoique un peu plus comprimée. De ce fait, on pourrait être tenté de le rattacher au type de Salfeldiella lateumbilicatum. Un examen attentif de la ligne cloisonnaire montre que celle-ci, à la fin du tour, n'a pas les phyllites terminaux développés caractéristiques du genre Salfeldiella, d'autre part les phyllites terminaux portent à leur base une échancrure (petit phyllite) que l'on ne voit jamais chez les Salfeldiella de petite taille. Pourtant au début du tour de ce petit spécimen les selles sont de type Salfeldiella (gros phyllites terminaux sans échancrure à leur base). En fait, ce spécimen est un nucleus de l'espèce P. (H.) paquieri dont il a déjà à la fin du tour la ligne cloisonnaire. C'est sans doute aussi le cas des S.
(G.) lateumbilicatum figurés par
(2005, Pl. 5,
figs. 9-11). Ils ont beaucoup de ressemblance avec les petits P. (H.) paquieri. Rappelons aussi que la ligne cloisonnaire adulte de P. (H.) paquieri est de type Phylloceras-Hypophylloceras et en aucun cas de type Salfeldiella, (,
2000a, p. 135, fig. 292), c'est pourquoi nous rangeons P. (H.) paquieri parmi les espèces du genre Phylloceras, sous-genre Hypophylloceras. Les deux P. (H.) paquieri de la Pl. 6 (,
2005, figs. 1-2) sont des P. (H.) delanoyi par la morphologie de leur coquille et leurs costules réunies en faisceaux, ornementation qui les rend parfaitement reconnaissables.
à D = 8,8 mm) a une morphologie assez proche de pygmaeum quoique un peu plus comprimée. De ce fait, on pourrait être tenté de le rattacher au type de Salfeldiella lateumbilicatum. Un examen attentif de la ligne cloisonnaire montre que celle-ci, à la fin du tour, n'a pas les phyllites terminaux développés caractéristiques du genre Salfeldiella, d'autre part les phyllites terminaux portent à leur base une échancrure (petit phyllite) que l'on ne voit jamais chez les Salfeldiella de petite taille. Pourtant au début du tour de ce petit spécimen les selles sont de type Salfeldiella (gros phyllites terminaux sans échancrure à leur base). En fait, ce spécimen est un nucleus de l'espèce P. (H.) paquieri dont il a déjà à la fin du tour la ligne cloisonnaire. C'est sans doute aussi le cas des S.
(G.) lateumbilicatum figurés par
(2005, Pl. 5,
figs. 9-11). Ils ont beaucoup de ressemblance avec les petits P. (H.) paquieri. Rappelons aussi que la ligne cloisonnaire adulte de P. (H.) paquieri est de type Phylloceras-Hypophylloceras et en aucun cas de type Salfeldiella, (,
2000a, p. 135, fig. 292), c'est pourquoi nous rangeons P. (H.) paquieri parmi les espèces du genre Phylloceras, sous-genre Hypophylloceras. Les deux P. (H.) paquieri de la Pl. 6 (,
2005, figs. 1-2) sont des P. (H.) delanoyi par la morphologie de leur coquille et leurs costules réunies en faisceaux, ornementation qui les rend parfaitement reconnaissables.
Cet exemple illustre la difficulté de déterminer les Salfeldiella qui sont étroitement apparentés aux Hypophylloceras et aux Goretophylloceras. La possession d'une ligne cloisonnaire de type Salfeldiella et la présence de constrictions ne suffisent donc pas pour ranger le spécimen dans le genre Salfeldiella, ces caractères doivent être aussi présents chez l'adulte.
& (2006) considèrent que P. (H.) paquieri est identique à P. (H.) onoense voire à Holcophylloceras sp. ( & , Pl. 9, figs. 8-9). Cette opinion a le mérite de justifier l'intégration de P. (H.) paquieri au sous-genre Hypophylloceras. Il est certain que P. (H.) paquieri et P. (H.) onoense sont des espèces très proches mais il reste cependant à prouver qu'elles sont identiques.
La ligne cloisonnaire a un tracé complexe de type Phylloceras (Hypophylloceras) au moins chez les grands spécimens. Chez les petits spécimens, elle a l'allure moins complexe comme celle qui a été figurée récemment (, 2000a, fig. 291, p. 135). Cette espèce a la morphologie de la coquille des Holcophylloceras du Jurassique mais elle n'en a pas la ligne cloisonnaire. C'est une convergence qui ne doit pas masquer la place réelle de P. (H.) paquieri parmi les P. (Hypophylloceras).
Âge : P. (H.) paquieri n'était connue en France que dans le "Gargasien" (, 2000a, p. 137). Les récoltes vocontiennes étendent son extension à la fois vers le bas, au sommet de l'Aptien inférieur (Zone à Deshayesi), et vers le haut, à la base de l'Albien inférieur (base de la Zone à Tardefurcata).
Phylloceras (Hypophylloceras) ponticuli (, 1842)
| V | 1841 | Ammonites semistriatus d', p. 136, Pl. 41, figs. 3-4. |
| 1842 | Ammonites ponticuli in , p. 783, Pl. 1, fig. 3 & 3.a-b. | |
| 2000a | Phylloceras (Hypophylloceras) ponticuli (, 1842).- , p. 137, Pl. 34, figs. 1-3 & 4.a-b ; figs. 293-299 dans le texte. | |
| 2006 | Euphylloceras ponticuli (, 1842) & , p. 30. | |
| V | 2006 | Phylloceras (Hypophylloceras) ponticuli (, 1842).- in , p. 44, Pl. 17, figs. 1.a-b & 2 ; fig. 22 dans le texte. |
| Synonymie plus complète, lectotype, localité-type, âge du lectotype : , 2000a, p. 137. | ||
Matériel étudié : 9 spécimens collection MD1550, 1551, 1552, 1553, 1554 (non mesuré) et 1555.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1551a | 52,3 | 34,3 | - | 3,2 | 0,66 | - | 0,06 | - |
| 42,2 | 26,0 | 17,0 | 2,9 | 0,62 | 0,40 | 0,07 | 0,65 | |
| 1555 (1) | 48,6 | 30,0 | 20,7 | 3,0 | 0,62 | 0,43 | 0,06 | 0,69 |
| 41,0 | 25,8 | 17,7 | 2,0 | 0,63 | 0,43 | 0,05 | 0,69 | |
| (1) forme épaisse | ||||||||
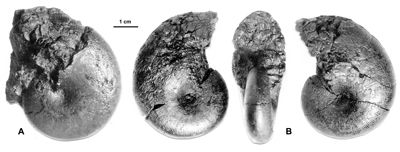
Cliquer sur la vignette pour agrandir l'image.
Figure 22 : Phylloceras (Hypophylloceras) ponticuli (,
1842).
A : MD1551a U1s/TR : Aptien inférieur (Zone à Furcata) - Ravin du Terrois, Drôme.
B : MD1555 U1s/TR : Aptien inférieur (Zone à Furcata) - Ravin du Terrois, Drôme.
Description : les spécimens sont des moules internes lisses sur les flancs mais finement costulés sur le bord externe des flancs et le bord ventral, flancs convexes et section du tour elliptique.
La ligne cloisonnaire (Fig. 23 ![]() ) est du type diphylle-triphylle mais avec un certain développement des phyllites internes des selles S1 et S2, elle est donc complexe, c'est la ligne cloisonnaire d'un Phylloceras (Hypophylloceras).
) est du type diphylle-triphylle mais avec un certain développement des phyllites internes des selles S1 et S2, elle est donc complexe, c'est la ligne cloisonnaire d'un Phylloceras (Hypophylloceras).
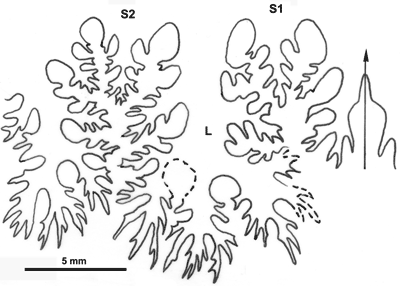
Cliquer sur la vignette pour agrandir l'image.
Figure 23 : Ligne cloisonnaire de Phylloceras (Hypophylloceras) ponticuli.
MD1552 U1s/TR : Aptien inférieur (Zone à Furcata) Ravin du Terrois, Drôme.
Affinités : l'espèce est très proche de P. (H.) moriezense mais s'en distingue surtout par l'absence de méplat sur les flancs.
Âge : P. (H.) ponticuli était connue de l'Hauterivien à l'Aptien inférieur (, 2000a, p. 139). Presque tous les spécimens des Marnes Bleues vocontiennes proviennent de la fin de l'Aptien inférieur (Zone à Furcata); un seul a été récoté dans l'Aptien moyen (MD1554 de la Zone à Melchioris).
Phylloceras (Hypophylloceras) aff. velledae (, 1834)
| 1834 | Ammonites velledae , Pl. 35. | |
| V | 1841 | Ammonites velledae .- d', p. 280, Pl. 82, figs. 1-4. |
| 1989 | Phylloceras (Hypophylloceras) velledae velledae (, 1834).- , p. 114, Pl. 1, figs. 10.a-c & 11. | |
| 2000a | Phylloceras (Hypophylloceras) velledae (, 1834).- , p. 141, Pl. 35, figs. 1-2 ; figs. 314-318 dans le texte. | |
| 2000 | Phylloceras (Hypophylloceras) velledae (, 1834).- et alii, p. 655, fig. 45.i. | |
| 2006 | Euphylloceras velledae (, 1834). & , p. 31, Pl. 2, figs. 1 & 5 ; fig. 13 dans le texte. | |
| V | 2006 | Phylloceras (Hypophylloceras) velledae (, 1834).- in , p. 101, Pl. 39, fig. 1.a-b ; fig. 54 dans le texte. |
| Synonymie plus complète, néotype, localité-type, âge du néotype : , 2000a, p. 141-142. | ||
Matériel étudié : 16 spécimens collection MD1505, 1556, 1557, 1558, 1559, 1560, 1561, 1562, 1563, 1564, 1565, 1566, 1567 et 2444 (de 1556 à 2444 non mesurés).
Dimensions du spécimen de la Fig. 24.E ![]()
| N° MD | D | H | E | O | h | e | o | l |
| 1505b | 19,0 | 11,5 | - | 1,2 | 0,61 | - | 0,06 | - |

Cliquer sur la vignette pour agrandir l'image.
Figure 24 : Phylloceras (Hypophylloceras) aff. velledae (,
1834).
A : MD1563 U8i/BR : Albien supérieur (Zone à Blancheti) - Châtelat, Drôme.
B : MD1558 U7m/CT : Albien supérieur (Zone à Blancheti) - Risou, Hautes-Alpes.
C : MD1562 U8i/BR : Albien supérieur (Zone à Blancheti) - Châtelat, Drôme.
D : MD2444 U3s/?BL : Albien supérieur (Zone à Inflatum) - Ravine de Bert, Drôme.
E : MD1505b U8s/BR : Albien supérieur (Zone à Briacensis) - La Folie, Hautes-Alpes.
Description : tous les spécimens se présentent sous la forme d'empreintes écrasées. Seule leur ornementation constituée de costules partant de l'ombilic permet de les rapprocher de P. (H.) velledae. Chez la plupart des spécimens la costulation est fine. Le spécimen n°
1558 (Fig. 24.B ![]() ) se distingue cependant par ses costules plus épaisses ce qui le rapproche des P. (H.) velledae d'Afrique du Sud figurés par &
(1977).
) se distingue cependant par ses costules plus épaisses ce qui le rapproche des P. (H.) velledae d'Afrique du Sud figurés par &
(1977).
La ligne cloisonnaire (Fig. 25 ![]() ) de l'un des spécimens ressemble à celle de P. (H.) velledae : S1 diphylle à tendance tétraphylle, S2 triphylle.
) de l'un des spécimens ressemble à celle de P. (H.) velledae : S1 diphylle à tendance tétraphylle, S2 triphylle.
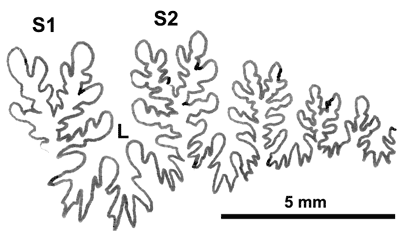
Cliquer sur la vignette pour agrandir l'image.
Figure 25 : Ligne cloisonnaire de Phylloceras (Hypophylloceras) aff. velledae.
MD1505b U8s/BR : Albien supérieur (Zone à Briacensis) Bruis-La Folie, Hautes-Alpes.
Âge : en France, P. (H.) velledae était connue de l'Albien au Cénomanien (, 2000a, p. 142). Les récoltes vocontiennes indiquent la présence de cette espèce dès la fin de l'Aptien supérieur (Zone à Jacobi) jusqu'au début du Cénomanien inférieur (base de la Zone à Mantelli).
Sous-genre Goretophylloceras , 1949
Les ammonites de ce sous-genre se distinguent de celles du sous-genre Hypophylloceras par leurs coquilles épaisses ou assez épaisses et leur ligne cloisonnaire moins découpée à selles S1 et S2 à gros phyllites terminaux spatulés, le lobe E est moins profond que le lobe L (, 2000a, p. 10).
Phylloceras (Goretophylloceras) fortunei (, 1892)
| 1892 | Ammonites fortunei , p. 241, 4 figs. dans le texte non numérotées. | |
| V | 1892 | Phylloceras goreti , p. 5, Pl. 1, fig. 3.a-c. |
| 2000a | Phylloceras (Goretophylloceras) fortunei ().- , p. 145, Pl. 36, figs. 1.a-b, 2.a-b & 3.a-b ; fig. 322-327 dans le texte. | |
| 2005 | Phylloceras (Goretophylloceras) fortunei ().- , p. 54, Pl. 3, figs. 8-12 | |
| Synonymie plus complète, néotype, localité-type, âge du néotype : voir , 2000a, p. 145. | ||
Matériel étudié : 38 spécimens collection MD1465, 1466, 1467, 1468, 1469, 1470, 1471, 1472, 1473, 1474, 1475, 1476, 1477, 1478 (non mesuré), 1479, 1480, 1481, 1498, 1499, 1500, 1501, 1502, 1503 (non mesuré), 2480, 2481, 2482, 2483, 2484 et 2488.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1468 | 19,8 | 11,5 | 9,1 | 2,3 | 0,58 | 0,46 | 0,12 | 0,79 |
| 1477 | 21,3 | 13,2 | 9,6 | 1,7 | 0,62 | 0,45 | 0,08 | 0,73 |
| 1500a | 12,4 | 7,0 | 5,6 | 1,6 | 0,56 | 0,45 | 0,13 | 0,80 |
| 1501 | 50,0 | - | 20,5 | 5,0 | - | 0,41 | 0,10 | - |
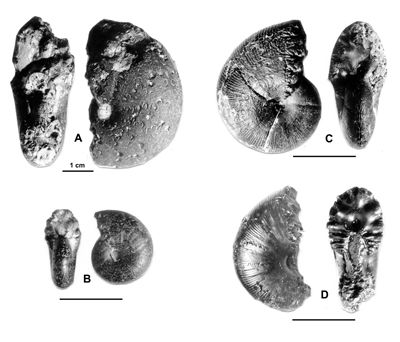
Cliquer sur la vignette pour agrandir l'image.
Figure 26 : Phylloceras (Goretophylloceras) fortunei (,
1892).
A : MD1501 U4m/?BT : Aptien supérieur (Zone à Nolani) - Vallon du Pin, Alpes-de-Haute-Provence.
B : MD1500a U3s/G1-NO : Aptien moyen (Zone à Melchioris) - Gaubert, Hautes-Alpes.
C : MD1477 U3s/Fa-NO : Aptien moyen (Zone à Martini) - Aps Nord, Alpes-de-Haute-Provence.
D : MD1468 U4m/BT : Aptien supérieur (Zone à Nolani) - Bartalasse, Drôme.
Description : les spécimens des Marnes Bleues, même si leur état de conservation est loin d'être parfait, se laissent reconnaître sans difficulté. Ils sont épais, le maximum de l'épaisseur étant vers l'ombilic la section est ovale. L'ornementation est constituée de costules fines, falciformes, groupées en faisceaux. Cette ornementation est particulièrement caractéristique chez le spécimen MD1468.
La ligne cloisonnaire (Fig. 27 ![]() ) est du type Phylloceras (Goretophylloceras) c'est-à-dire diphylle-triphylle, sans véritable tendance vers la tétraphyllie, les phyllites internes des terminaisons des selles restant peu développés.
) est du type Phylloceras (Goretophylloceras) c'est-à-dire diphylle-triphylle, sans véritable tendance vers la tétraphyllie, les phyllites internes des terminaisons des selles restant peu développés.
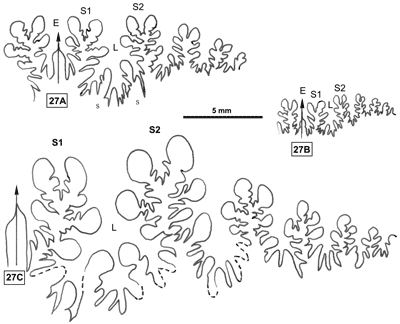
Cliquer sur la vignette pour agrandir l'image.
Figure 27 : Lignes cloisonnaires de Phylloceras (Goretophylloceras) fortunei.
A : MD1501 U4m/?BT : Aptien supérieur (Zone à Nolani) Lioux, Alpes-de-Haute-Provence.
B : MD1500a U3s/G1-NO : Aptien moyen (Zone à Melchioris) Gaubert, Alpes-de-Haute-Provence.
C : MD1479 U3i/FA-G1 : Aptien moyen (Zone à Melchioris) Les Vallons, Alpes-de-Haute-Provence.
Âge : l'espèce P. (G.) fortunei n'était connue jusqu'alors qu'au "Gargasien" (, 2000a, p. 146). Les récoltes vocontiennes étudiées ici s'étendent de l'Aptien moyen (sommet de la Zone à Martini) au début de l'Aptien supérieur (Zone à Nolani).
Phylloceras (Goretophylloceras) subalpinum (d', 1841)
| V | 1841 | Ammonites alpinus d', p. 283, Pl. 83, figs. 1-3. |
| 1850 | Ammonites subalpinus d', p. 124, 19ème étage, Albien, n° 35. | |
| 1989 | Phylloceras (Hypophylloceras) subalpinum subalpinum (d').- , p. 114, Pl. 1, figs. 5.a-b & 6.a-b. | |
| 2000a | Phylloceras (Goretophylloceras) subalpinum (d', 1841).- , p. 146, Pl. 36, fig. 4.a-b, 5, 6.a-b & 7.a-b ; fig. 328-333 dans le texte. | |
| V | 2006 | Phylloceras (Goretophylloceras) subalpinum (d', 1841), in , p. 102, Pl. 38, fig. 3.a-b ; fig. 55 dans le texte. |
| Synonymie plus complète, lectotype, localité-type, âge du lectotype : , 2000a, p. 146. | ||
Matériel étudié : 108 spécimens collection MD1482, 1483, 1484, 1485, 1486, 1487, 1488, 1489 et 2448 (non mesurés).
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1487a | 30,3 | 18,5 | 14,0 | 2,6 | 0,61 | 0,46 | 0,09 | 0,76 |
| 1489a | 31,0 | 18,8 | 14,9 | 2,1 | 0,61 | 0,48 | 0,07 | 0,79 |

Cliquer sur la vignette pour agrandir l'image.
Figure 28 : Phylloceras (Goretophylloceras) subalpinum (d',
1841).
A : MD1489a U7m/CT : Albien supérieur (Zone à Blancheti) - Ravine de Bert, Drôme.
B : MD1487a U7m/CT : Albien supérieur (Zone à Blancheti) - Ravine de Bert, Drôme.
Description : les spécimens vocontiens sont tous des moules internes pyritisés lisses à flancs très convexes et section du tour ovale, le maximum d'épaisseur étant près de l'ombilic.
La ligne cloisonnaire (Fig. 29 ![]() ) est celle des Phylloceras (Goretophylloceras), c'est-à-dire diphylle-triphylle peu découpée. Il n'a jamais été figuré de ligne cloisonnaire plus complexe pour cette espèce.
) est celle des Phylloceras (Goretophylloceras), c'est-à-dire diphylle-triphylle peu découpée. Il n'a jamais été figuré de ligne cloisonnaire plus complexe pour cette espèce.
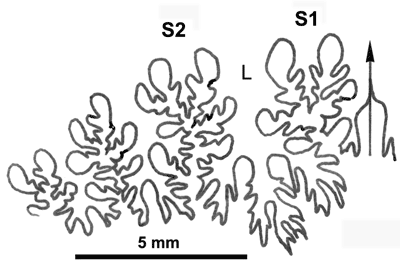
Cliquer sur la vignette pour agrandir l'image.
Figure 29 : Ligne cloisonnaire de Phylloceras (Goretophylloceras) subalpinum.
MD1486 U7i/CT : Albien supérieur (Zone à Blancheti) Le Risou, Hautes-Alpes.
Affinités : (1964) ainsi que & (1977) ont rattaché l'espèce ellipticum à l'espèce subalpinum, point de vue que nous adoptons ici et qui sera discuté avec la description du morphe ellipticum.
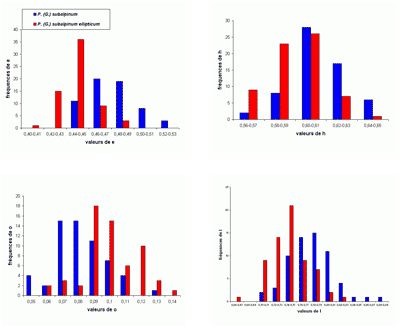
Les histogrammes des fréquences des paramètres relatifs (Fig. 30 ![]() ) pour P. (G.) subalpinum et P. (G.) subalpinum morphe ellipticum sont tous unimodaux, sauf pour les histogrammes de o qui paraissent être bimodaux, il y a peut-être là l'indice d'un processus d'altération de la croissance qui, chez les Goretophylloceras, affecte l'ouverture de l'ombilic, la ligne cloisonnaire et l'ornementation.
) pour P. (G.) subalpinum et P. (G.) subalpinum morphe ellipticum sont tous unimodaux, sauf pour les histogrammes de o qui paraissent être bimodaux, il y a peut-être là l'indice d'un processus d'altération de la croissance qui, chez les Goretophylloceras, affecte l'ouverture de l'ombilic, la ligne cloisonnaire et l'ornementation.
Cliquer sur la vignette pour agrandir l'image.
Figure 30 : Histogrammes des fréquences de e, h, o et l chez Phylloceras (Goretophylloceras) subalpinum et Phylloceras (Goretophylloceras) subalpinum morphe ellipticum.
L'espèce la plus proche est P. (G.) fortunei de l'Aptien. P. (G.) subalpinum en dérive très probablement par accentuation de la pédomorphose, le caractère le plus évident de l'altération de la croissance étant la disparition de la costulation chez P. (G.) subalpinum, ou tout au moins son apparition plus tardive, que chez P. (G.) fortunei.
Âge : P. (G.) subalpinum a été récoltée de l'Aptien terminal (sommet de la Zone à Jacobi) à l'Albien supérieur (Zone à Blancheti). Cet âge correspond parfaitement avec celui qui vient de lui être attribué en France par (2000a, p. 147).
Phylloceras (Goretophylloceras) subalpinum (d', 1841) morphe ellipticum , 1895
| 1865 | Ammonites subalpinus d', , p. 114, Pl. 58, fig. 3 & 3.a-b. | |
| 1895 | Phylloceras ellipticum , p. 11, Pl. 1, fig. 2.a-b; Pl. 6, fig. 1.a-b. | |
| 1989 | Phylloceras (Hypophylloceras) subalpinum (d') ellipticum .- , p. 114, Pl. 1, figs. 7, 8.a-c & 9.a-b. | |
| 2000a | Phylloceras (Hypophylloceras) ellipticum .- , p. 131, Pl. 32, fig. 5.a-b ; fig. 276 dans le texte. | |
| Synonymie plus complète, holotype, localité-type, âge de l'holotype : , 2000a, p. 131. | ||
Matériel étudié : 106 spécimens collection MD1490, 1491, 1492, 1493 et 2445.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1490 | 27,0 | 15,8 | 12,0 | 2,5 | 0,59 | 0,44 | 0,09 | 0,76 |
| 1493a | 19,2 | 11,5 | 8,7 | 1,8 | 0,60 | 0,45 | 0,09 | 0,76 |
| 1493b | 21,9 | 13,6 | 9,7 | 1,6 | 0,62 | 0,44 | 0,07 | 0,71 |
| 2445a | 32,7 | 19,8 | 14,3 | 2,0 | 0,60 | 0,44 | 0,06 | 0,72 |
Cliquer sur la vignette pour agrandir l'image.
Figure 31 : Phylloceras (Goretophylloceras) subalpinum morphe ellipticum ,
1895.
A : MD1490 U7i/FS-CT : Albien supérieur (Zone à Inflatum) - Ravine de Bert, Drôme.
B : MD1493a U7m/CT : Albien supérieur (Zone à Blancheti) - Ravine de Bert, Drôme.
C : MD1493b - U7m/CT : Albien supérieur (Zone à Blancheti) - Ravine de Bert,
Drôme.
D : MD2445a - U7m/CT : Albien supérieur (Zone à Blancheti) - Ravine de Bert, Drôme.
Description : tous les spécimens étudiés sont des moules internes pyritisés lisses, modérément épais, à flancs convexes et section du tour elliptique. Les histogrammes des fréquences des paramètres relatifs h, e et l (voir Fig. 30 ![]() ), rassemblant subalpinum et ellipticum sont ceux d'une population unimodale. Il existe cependant des différences entre les deux taxons, P. (G.) subalpinum est une population globalement plus épaisse que P. (G.) subalpinum morphe ellipticum. Tandis que l'ombilic est globalement un peu plus ouvert chez ellipticum que chez subalpinum. La valeur de h est sensiblement la même chez l'espèce et son morphe ellipticum. Une différence importante est dans la section du tour, ovale chez subalpinum, elliptique chez ellipticum, c'est le caractère qui les distingue le plus facilement.
), rassemblant subalpinum et ellipticum sont ceux d'une population unimodale. Il existe cependant des différences entre les deux taxons, P. (G.) subalpinum est une population globalement plus épaisse que P. (G.) subalpinum morphe ellipticum. Tandis que l'ombilic est globalement un peu plus ouvert chez ellipticum que chez subalpinum. La valeur de h est sensiblement la même chez l'espèce et son morphe ellipticum. Une différence importante est dans la section du tour, ovale chez subalpinum, elliptique chez ellipticum, c'est le caractère qui les distingue le plus facilement.
À ces considérations s'ajoutent les résultats statistiques obtenus sur les deux taxons et résumés dans le tableau suivant :
| P. (G.) subalpinum ellipticum | P. (G.) subalpinum subalpinum | ||||||
| paramètres | moyennes | écarts-types | coefficients de variation | paramètres | moyennes | écarts-types | coefficients de variation |
| D | 19,95 | 3,85 | 19,30% | D | 25,12 | 3,81 | 15,17% |
| H | 11,67 | 2,65 | 22,71% | H | 15,31 | 2,54 | 16,59% |
| E | 8,83 | 1,81 | 20,50% | E | 11,89 | 1,94 | 22,66% |
| O | 1,92 | 0,32 | 16,67% | O | 2,03 | 0,46 | 22,66% |
| Moyennes | 19,80% | moyennes | 17,69% | ||||
| h | 0,60 | 0,02 | 3,33% | h | 0,61 | 0,02 | 3,28% |
| e | 0,44 | 0,02 | 4,55% | e | 0,48 | 0,02 | 4,17% |
| o | 0,10 | 0,02 | 20,00% | o | 0,08 | 0,02 | 25,00% |
| l | 0,74 | 0,03 | 4,05% | l | 0,78 | 0,03 | 3,85% |
| Moyennes | 7,98% | moyennes | 9,08% | ||||
On remarque surtout la grande similitude des valeurs des écarts-types des deux taxons. Jusqu'alors les auteurs admettaient généralement qu'ellipticum et subalpinum étaient deux espèces différentes. Le fait que les taxons sont, dans les Marnes Bleues, souvent présents dans les mêmes gisements, donne un argument pour faire de ellipticum un morphe de préférence à une sous-espèce de subalpinum (, 1964 ; & , 1977). L'étude des spécimens des Marnes Bleues nous a amenés à modifier l'opinion exprimée récemment ( in , 2006, p. 102) et à admettre que ellipticum est un morphe de subalpinum, les histogrammes des deux taxons montrant suffisamment de points communs et de différences (voir ci-dessus) pour justifier cette interprétation. Il n'est pas possible actuellement d'affirmer ou d'infirmer que subalpinum et ellipticum pourraient constituer un couple dimorphe. En ne considérant que l'épaisseur maximale de la coquille proche de l'ombilic (section ovale) ou au milieu des flancs (section elliptique) l'hypothèse du dimorphisme de section a déjà été envisagée (, 1977, p. 28 ; 2000a, p. 15). Le dimorphisme de type microconque-macroconque paraît plus probable mais le matériel des Marnes Bleues ne permet pas de trancher cette question.
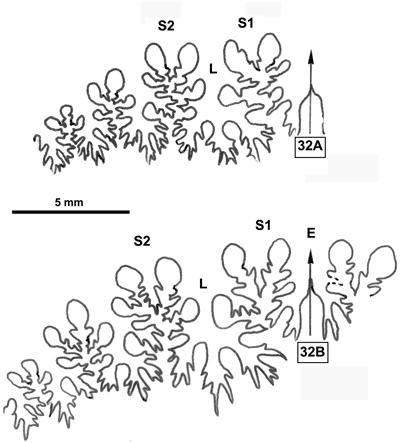
La ligne cloisonnaire (Fig. 32 ![]() ) est de type Phylloceras (Goretophylloceras) chez les spécimens des Marnes Bleues. La différence la plus importante avec l'holotype réside surtout dans les terminaisons des selles apparemment peu développées. Mais il s'agit peut-être d'une conséquence de l'état de conservation de la ligne cloisonnaire (l'usure du moule interne pouvant simplifier le tracé de la ligne cloisonnaire).
) est de type Phylloceras (Goretophylloceras) chez les spécimens des Marnes Bleues. La différence la plus importante avec l'holotype réside surtout dans les terminaisons des selles apparemment peu développées. Mais il s'agit peut-être d'une conséquence de l'état de conservation de la ligne cloisonnaire (l'usure du moule interne pouvant simplifier le tracé de la ligne cloisonnaire).
Cliquer sur la vignette pour agrandir l'image.
Figure 32 : Lignes cloisonnaires de Phylloceras (Goretophylloceras) subalpinum morphe ellipticum.
A : MD1493a U7m/CT : Albien supérieur (Zone à Blancheti) Ravine de Bert, Drôme.
B : MD1490 U7i/FS-CT : Albien supérieur (Zone à Inflatum) - Ravine de Bert, Drôme.
Les lignes cloisonnaires figurées (par exemple & , 1977, fig. 22, p. 356 ou celle de l'holotype de , 1865) justifiaient jusqu'alors l'opinion selon laquelle ellipticum pouvait appartenir à un autre sous-genre que celui de subalpinum. Mais il n'est pas certain que les lignes cloisonnaires figurées par différents auteurs aient été relevées sur des spécimens bien conservés. Alors que les lignes cloisonnaires des Marnes Bleues l'ont été sur des spécimens bien conservés. Ces lignes cloisonnaires sont de manière certaine celles du sous-genre Goretophylloceras et non celle d'Hypophylloceras comme le croyaient (1964) et & (1977).
Âge : P. (G.) morphe ellipticum n'avait été identifiée avec certitude que dans l'Albien moyen en France (, 2000a, p. 131). Les spécimens des Marnes Bleues vocontiennes proviennent tous de l'Albien supérieur (Zone à Blancheti).
Phylloceras (Goretophylloceras) vocontium nov. sp.
Holotype : collection MD1495.
Localité : Les Vallons, commune de Tartonne, Alpes-de-Haute-Provence.
Âge : base de l'Aptien supérieur : Zone à Nolani.
Origine du nom : du nom de la peuplade des Voconces établie dans la région de Vaison-la-Romaine.
Matériel étudié : 4 spécimens collection MD1494, 1495, 1504 et 2515.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1494 | 17,6 | 9,5 | 7,1 | 3,9 | 0,54 | 0,40 | 0,22 | 0,75 |
|
1495 holotype |
19,4 | 10,6 | 9,1 | 4,7 | 0,55 | 0,47 | 0,24 | 0,86 |
| 1504 | 11,7 | 6,0 | 4,3 | 2,4 | 0,51 | 0,37 | 0,21 | 0,72 |
Cliquer sur la vignette pour agrandir l'image.
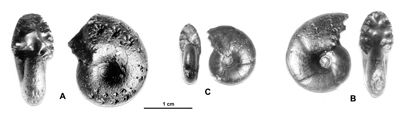
Figure 33 : Phylloceras (Goretophylloceras) vocontium nov. sp.
A : MD1495, holotype - U4m/BT : Aptien supérieur (Zone à Nolani) - Les
Vallons, Alpes-de-Haute-Provence.
B : MD1494 U3s/?VL : Aptien moyen (Zone à Melchioris) - Les Briers, Alpes-de-Haute-Provence.
C : MD1504 U5i/?SZ : Albien inférieur (Zone à Tardefurcata) - Chênebuis, Alpes-de-Haute-Provence.
Diagnose : coquille épaisse, flancs convexes, bord externe arrondi et aminci, maximum d'épaisseur du tour vers l'ombilic, ombilic ouvert, constrictions au moins au début du dernier tour, section du tour ovale épaisse, ligne cloisonnaire de type Goretophylloceras.
Description : trois moules internes pyritisés, lisses, assez épais, flancs convexes, bord externe un peu aminci et arrondi, maximum d'épaisseur du tour vers l'ombilic, section du tour ovale épaisse, ombilic ouvert (plus de 20 %) laissant voir les tours internes, forte pente ombilicale. Le spécimen le plus petit (Fig. 33.C ![]() ) a 3 constrictions droites et un peu proverses sur le dernier tour, le second (Fig. 33.B
) a 3 constrictions droites et un peu proverses sur le dernier tour, le second (Fig. 33.B ![]() ) a 4 constrictions sur le dernier tour mais la dernière près de l'ouverture est presque effacée. L'holotype (Fig. 33.A
) a 4 constrictions sur le dernier tour mais la dernière près de l'ouverture est presque effacée. L'holotype (Fig. 33.A ![]() ), le spécimen le plus grand, ne montre qu'une seule constriction au début du dernier tour.
), le spécimen le plus grand, ne montre qu'une seule constriction au début du dernier tour.
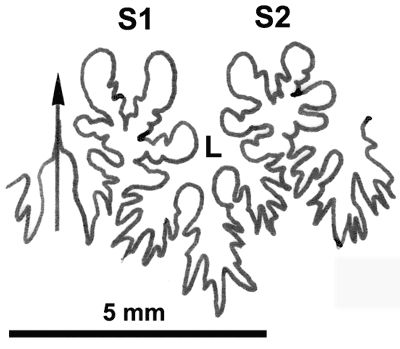
La ligne cloisonnaire (Fig. 34 ![]() ) est plus complexe que celle des Salfeldiella avec S1 diphylle et S2 triphylle à terminaisons assez développées et petits phyllites internes.
) est plus complexe que celle des Salfeldiella avec S1 diphylle et S2 triphylle à terminaisons assez développées et petits phyllites internes.
Cliquer sur la vignette pour agrandir l'image.
Figure 34 : Ligne cloisonnaire de Phylloceras (Goretophylloceras) vocontium, holotype.
MD1495 U4m/BT : Aptien supérieur (Zone à Nolani) - Les Vallons, Alpes-de-Haute-Provence.
Affinités : P. (G.) vocontium a une morphologie qui rappelle le port de P. (G.) fortunei. Elle en est probablement issue par altération de la croissance se manifestant surtout par l'ouverture de l'ombilic et l'absence d'ornementation. Il n'a pas été observé, dans la population étudiée, de formes de passage entre les deux espèces, qui se distinguent donc sans difficulté.
Âge : P. (G.) vocontium nov. sp. a été récoltée de la fin de l'Aptien moyen (sommet de la Zone à Melchioris) au début de l'Albien inférieur (base de la Zone à Tardefurcata).
Phylloceras (Goretophylloceras) zanebonii , 2000a
| 2000a | Phylloceras (Goretophylloceras) zanebonii , p. 147, Pl. 36, figs. 8.a-b, 9, 10 & 11.a-b ; figs. 334-338 dans le texte. | |
| Synonymie plus complète, holotype, localité-type, âge de l'holotype : , 2000a, p. 147. | ||
Matériel étudié : 2 spécimens collection MD1496 et 1497.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1496 | 14,2 | 7,6 | 5,0 | 2,9 | 0,54 | 0,35 | 0,20 | 0,66 |
| 1497 | 17,9 | 10,5 | 7,8 | 2,6 | 0,59 | 0,43 | 0,14 | 0,74 |

Cliquer sur la vignette pour agrandir l'image.
Figure 35 : Phylloceras (Goretophylloceras) zanebonii ,
2000a.
A : MD1497 U2s/FA : Aptien moyen (Zone à Martini) - Crouas, Alpes-de-Haute-Provence.
B : MD1496 U3s/VL : Aptien moyen (Zone à Melchioris) Serre-Chaitieu, Drôme.
Description : moules internes pyritisés, modérément épais, lisses, à flancs plats, bord externe arrondi, ombilic profond un peu ouvert à pente assez prononcée, section du tour subrectangulaire. Le plus petit des spécimens porte une discrète costulation du moule interne sur le bord ventral. Les deux spécimens de cette nouvelle récolte sont parmi les plus petits actuellement connus, ce qui explique que leur ombilic soit un peu ouvert (chez les Phyllocerataceae, l'involution s'accroît au cours du développement).
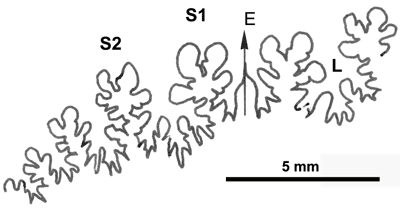
La ligne cloisonnaire diphylle-triphylle est de type Goretophylloceras (Fig. 36 ![]() ).
).
Cliquer sur la vignette pour agrandir l'image.
Figure 36 : Ligne cloisonnaire de Phylloceras (Goretophylloceras) zanebonii.
MD1497 U2s/FA : Aptien moyen (Zone à Martini) Crouas, Alpes-de-Haute-Provence.
Âge : P. (G.) zanebonii était déjà connue dans le "Gargasien" (, 2000a, p. 147). Les deux spécimens des Marnes Bleues vocontiennes proviennent de l'Aptien moyen (sommet de la Zone à Martini et Zone à Melchioris).
Sous-famille des Neocalliphylloceratinae , 2000a
Les ammonites de cette sous-famille ont une coquille lisse ou finement costulée portant des constrictions proverses même aux plus grands diamètres et parfois des bourrelets ventraux, les constrictions du moule internes ont même allure que les costules du test, la ligne cloisonnaire est peu complexe (, 2000a, p. 10).
Genre Salfeldiella , 1927
Coquille assez comprimée, ornée de costules ainsi que le moule interne, constrictions falciformes sur le test et le moule interne, selles diphylles ou triphylles, E moins profond que L, ligne cloisonnaire modérément découpée (, 2000a, p. 11).
La plupart des taxons décrits dans les Marnes Bleues du bassin vocontien étaient auparavant considérés comme des "variétés" de Salfeldiella guettardi. Les récoltes de cette étude ont permis de constituer des populations suffisantes pour comparer les "variétés" entre elles et les comparer à la population des Salfeldiella guettardi (proches du type + les sous-espèces gracile et victoriana). On verra plus loin les enseignements que l'on peut tirer de cette étude statistique.
Salfeldiella belgradensis (, 1913)
| 1899 | Phylloceras guettardi (), , p. 97, Pl. 5, fig. 5.a-c. | |
| ? | 1907 | Phylloceras guettardi ().- , p. 60. |
| 1913 | Phylloceras belgradensis , p. 66, Pl. 3, fig. 5.a- b. | |
| 1969 | Salfeldiella pura , p. 131, Pl. 1, figs. 8.a-c & 9.a-c ; Pl. 21, fig. 7. | |
| 2000a | Salfeldiella belgradensis (, 1913).- , p. 149, Pl. 36, fig. 13.a-c ; figs. 339-340 dans le texte. | |
| Synonymie plus complète, holotype, localité-type, âge de l'holotype : , 2000a, p. 149. | ||
Matériel étudié : 10 spécimens collection MD1605, 1606, 1607, 1608, 1609, 2485 et 2486.
Dimensions des spécimens figurés et de S. pura , 1969
| N° MD | D | H | E | O | h | e | o | l |
| MD1605 | 18,8 | 10,0 | 8,4 | 3,1 | 0,53 | 0,45 | 0,16 | 0,84 |
| MD1606 | 28,7 | 16,0 | 12,5 | 4,0 | 0,56 | 0,44 | 0,14 | 0,78 |
| MD1607 | 15,2 | 8,6 | 6,8 | 2,6 | 0,57 | 0,45 | 0,17 | 0,79 |
|
S. pura n° 237 |
35,5 | 18 | 14,2 | 5,3 | 0,51 | 0,40 | 0,15 | 0,79 |
|
S. pura n° 238 |
24 | 12 | 8,7 | 4 | 0,50 | 0,36 | 0,17 | 0,72 |
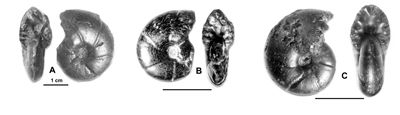
Cliquer sur la vignette pour agrandir l'image.
Figure 37 : Salfeldiella belgradensis (,
1913).
A : MD1606 U3s/VL : Aptien moyen (Zone à Melchioris) - Les Vallons, Alpes-de-Haute-Provence.
B : MD1607 U3i/FA-G1 : Aptien moyen (?Zone à Melchioris) - Gaubert, Alpes-de-Haute-Provence.
C : MD1605 U2i/GB : Aptien moyen (Zone à Martini) - Ravin du Terrois, Drôme.
Description : les spécimens rapportés au type de l'epèce belgradensis sont des moules internes pyritisés, épais, lisses, à ombilic un peu ouvert, flancs convexes, maximum d'épaisseur du tour vers l'ombilic et section du tour ovale. Le moule interne porte 5 constrictions proverses chez le spécimen le plus complet (MD1606), un autre (MD1605) a aussi 5 constrictions ainsi que le spécimen MD1607 dont la mauvaise conservation du début du tour masque la première constriction. Le spécimen n° 1609 est mal conservé mais ses dimensions, sa section et ses constrictions sont celles de l'espèce belgradensis. Au passage du bord siphonal la constriction s'infléchit un peu plus fortement vers l'avant en formant un chevron assez prononcé.
(2005, p. 65) pense que l'on ne peut réunir belgradensis et caucasica étant donné leur différence de taille et leur âge différent. En réalité la différence des tailles n'est pas considérable et à part parfois une ornementation plus discrète chez les petits spécimens et au-delà de 20 mm, on voit peu de différences dans la morphologie et l'ornementation des spécimens de cette taille. Bien sûr il y a des exceptions quand l'écart des tailles est nettement plus important mais les Salfeldiella ne sont jamais très grands, leurs coquilles et leurs lignes cloisonnaires conservent les caractères des tours internes de taxons plus grands, ce qui explique en particulier une certaine discrétion de l'ornementation, un tracé simplifié de la ligne cloisonnaire et un ombilic souvent assez ouvert. Un autre argument de serait que belgradensis est une espèce albienne alors que caucasica serait aptienne. En réalité, grâce à la présente étude, nous savons que les deux taxons sont connus dans l'Aptien et l'Albien. D'autre part on ne voit pas pourquoi une forme aptienne ne pourrait pas être réunie à une forme albienne si celles-ci se révèlent identiques. Cela n'a rien d'exceptionnel chez les Phylloceratidae dont les taxons sont connus parfois sur quatre étages. Il est difficile de mettre en doute l'espèce belgradensis, qui, rappelons-le a la priorité sur caucasica (voir la sous-espèce suivante et , 2000a, p. 150).
La ligne cloisonnaire (Fig. 38 ![]() ) est du type Salfeldiella : diphylle-triphylle, à phyllites terminaux développés.
) est du type Salfeldiella : diphylle-triphylle, à phyllites terminaux développés.
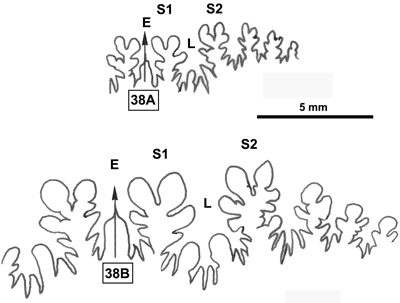
Cliquer sur la vignette pour agrandir l'image.
Figure 38 : Lignes cloisonnaires de Salfeldiella belgradensis.
A : MD1607 U3i/FA-G1 : Aptien moyen (?Zone à Melchioris) Gaubert, Alpes-de-Haute-Provence.
B : MD1609 U3s/FA-NO : Aptien moyen (Zone à Melchioris) Aps nord, Alpes-de-Haute-Provence.
Affinités : il n'y a pas de différences évidentes entre les spécimens des Marnes Bleues et le S. pura , 1969 (= P. guettardi , 1899, non A. guettardi , 1831), comme le montrent les dimensions données par pour S. pura (tableau ci-dessus). On doit admettre que S. pura est synonyme plus récent de S. belgradensis.
Les spécimens vocontiens sont parfaitement identiques à celui de l'Albien de l'Aube figuré récemment (, 2000a, Pl. 36, fig. 13).
Âge : Salfeldiella belgradensis était connue jusqu'alors de l'Albien inférieur (, 2000a, p. 150). Les spécimens vocontiens proviennent de l'Aptien moyen (Zone à Martini et Zone à Melchioris).
Salfeldiella belgradensis (, 1913) sayni , 2000a
| 1920 | Phylloceras guettardi () var. caucasica , p. 197, fig. 7 & 7.a. | |
| 2000a | Salfeldiella belgradensis (, 1913) sayni, , p. 150, Pl. 37, figs. 1-2 ; figs. 341-343 dans le texte. | |
| 2005 | Salfeldiella caucasica ().- , p. 65, Pl. 1, figs. 3-7. | |
| Synonymie plus complète, holotype, localité-type, âge de l'holotype de la sous-espèce : , 2000a, p. 150. | ||
Matériel étudié : 22 spécimens collection MD1610, 1611, 1612, 1613, 1614, 1616*, 1617, 1618, 1619, 1620, 1621, 2450 et 2489.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1610 | 14,7 | 8,5 | 5,6 | 1,9 | 0,58 | 0,38 | 0,13 | 0,66 |
| 1611 | 21,7 | 11,1 | 6,6 | 5,4 | 0,51 | 0,30 | 0,25 | 0,59 |
| 1616 (1) | 13,4 | 7,0 | 5,5 | 2,2 | 0,52 | 0,41 | 0,16 | 0,78 |
| (1) microconque | ||||||||
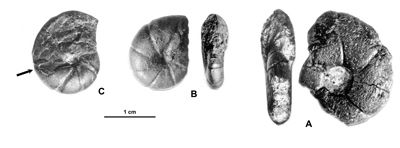
Cliquer sur la vignette pour agrandir l'image.
Figure 39 : Salfeldiella belgradensis sayni ,
2000a.
A : MD1611 U4m/BT : Aptien moyen (Zone à Melchioris) - Les Vallons, Alpes-de-Haute-Provence.
B : MD1610 U1s/TR : Aptien inférieur (Zone à Furcata) - Serre Chaïtieu, Drôme.
C : MD1616 - microconque (flèche au début de la loge) U2i/GB : Aptien moyen (Zone à Martini) - Gaubert, Alpes-de-Haute-Provence.
Description : S. belgradensis sayni se distingue essentiellement de S. belgradensis sensu stricto par son ombilic plus ouvert (à D élevé). Les spécimens sont des moules internes portant 5 à 6 constrictions presque droites et nettement proverses, elles passent le bord siphonal nettement infléchies vers l'avant, cette inflexion est très prononcée chez les plus grands spécimens. La ligne cloisonnaire (, 2000a, p. 150, fig. 343) a le tracé simplifié caractéristique du genre Salfeldiella.
Affinités : les histogrammes des fréquences des paramètres relatifs (Fig. 40 ![]() ) montrent qu'il existe bien deux populations distinctes S.
guettardi et S. belgradensis sayni pour h, e et surtout o et l.
) montrent qu'il existe bien deux populations distinctes S.
guettardi et S. belgradensis sayni pour h, e et surtout o et l.
Les individus appartenant à l'espèce S. belgradensis sayni forment donc une population dont les moyennes de h, e et l sont plus petites que celles de S. guettardi alors que la moyenne de o est plus élevée chez sayni.
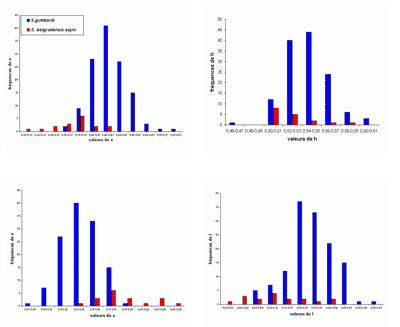
Cliquer sur la vignette pour agrandir l'image.
Figure 40 : Histogrammes des fréquences de e, h, o et l chez Salfeldiella belgradensis sayni et Salfeldiella guettardi.
Âge : S. belgradensis sayni était jusqu'alors connue dans l'Albien (, 2000a, p. 150). Les récoltes des Marnes Bleues sont datées de la fin de l'Aptien inférieur (Zone à Furcata) à l'Aptien terminal (Zone à Jacobi).
Salfeldiella guettardi (, 1831)
| 1779 | sans nom spécifique, , p. 841, Pl. 10, figs. 1?, 2, 3? & 4. | |
| 1830 | Ammonites guettardi , Pl. 11, fig. 2 ; Pl. 12, fig. 8, description en 1831. | |
| V | 1841 | Ammonites guettardi .- d', p. 169, Pl. 53, figs. 1-3. |
| ? | 1989 | Sowerbyceras (Holcophylloceras) guettardi ().- , p. 114, Pl. 1, fig. 15.a-b. |
| 2000a | Salfeldiella guettardi ().- , p. 151, Pl. 37, figs. 4.a-b, 5.a-b, 6 & 7.a-b ; figs. 345-350 dans le texte. | |
| 2005 | Salfeldiella guettardi ().- , p. 62, Pl. 1, figs. 1-2 ; Pl. 4, figs. 6-9 (non fig. 10 = victoriana). | |
| V | 2006 | Salfeldiella guettardi ().- in , p. 61, Pl. 31, fig. 7.a-b ; fig. 38 dans le texte. |
| Synonymie plus complète, néotype, localité-type, âge du néotype : , 2000a, p. 152. | ||
Matériel étudié : 48 spécimens collection MD 1533, 1622, 1623, 1624, 1625, 1626, 1627, 1628 (dont une forme à fortes constrictions), 1629 (non mesurés), 1630, 1631 (dont une forme à fortes constrictions), 1632, 1633, 1634, 1635, 1636, 1637, 1638, 1639, 1640, 1641, 1642, 1643, 1644, 1645, 1646, 2451 et 2452.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1628c (1) | 25,8 | 13,5 | - | 3,6 | 0,52 | - | 0,14 | - |
| 23,7 | 12,5 | 10,6 | 3,4 | 0,53 | 0,45 | 0,14 | 0,85 | |
| 1642 | 42,0 | 22,3 | 18,9 | 5,3 | 0,53 | 0,45 | 0,13 | 0,85 |
| (1) forme à fortes constrictions | ||||||||
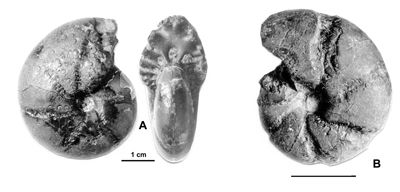
Cliquer sur la vignette pour agrandir l'image.
Figure 41 : Salfeldiella guettardi (,
1831).
A : MD1642 U2i/GB : Aptien moyen (Zone à Martini) - Gaubert, Alpes-de-Haute-Provence.
B : MD1628c - spécimen à fortes constrictions U2i/GB : Aptien moyen (Zone à Martini) - Gaubert, Alpes-de-Haute-Provence.
Description : moules internes pyritisés, épais, à ombilic légèrement ouvert, flancs convexes. Le maximum d'épaisseur du tour est vers l'ombilic, la section du tour est ovale. Le dernier tour porte 5 à 8 constrictions falciformes sur les flancs légèrement proverses sur le bord siphonal.
La ligne cloisonnaire (Fig. 42 ![]() ) est de type Salfeldiella, c'est-à-dire diphylle-triphylle et les selles S1 et S2 ont des terminaisons développées.
) est de type Salfeldiella, c'est-à-dire diphylle-triphylle et les selles S1 et S2 ont des terminaisons développées.
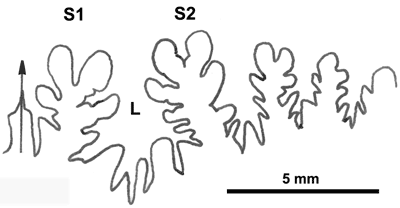
Cliquer sur la vignette pour agrandir l'image.
Figure 42 : Ligne cloisonnaire de Salfeldiella guettardi.
MD1634a U1sz/TR : Aptien inférieur (Zone à Furcata) Ravin du Pissourier, Drôme.
Affinités : la comparaison avec les autres taxons a été déjà évoquée avec S. belgradensis sayni. Ces relations seront approfondies plus loin.
Âge : les précédentes récoltes (pour celles qui étaient vérifiables) provenaient du "Gargasien" (, 2000a, p. 152). Les Salfeldiella guettardi les plus typiques ont été récoltés de la fin de l'Aptien inférieur (Zone à Furcata) à la base de l'Aptien supérieur (Zone à Nolani) L'espèce est surtout présente dans l'Aptien moyen. Ces précisions nouvelles ne modifient guère les conclusions auxquelles l'étude des Phylloceratoidea de France avait conduit, puisque S. guettardi n'a vécu que durant une partie de l'Aptien. L'espèce reste une exception parmi ce groupe pour sa durée de vie modérée.
Salfeldiella guettardi (, 1831) gracile , 1957
| 1957 | Salfeldiella gracile , p. 88, Pl. 1, figs. 7.a-b & 8.a-b ; fig. 7 dans le texte. | |
| ? | 1964 | Phylloceras (Hypophylloceras) grothi .- , p. 182, Pl. 12, fig. 6.a-c ; fig. 39 dans le texte. |
| ? | 1989 | Phylloceras (Hypophylloceras) grothi .- , p. 113, Pl. 1, fig. 3.a-c. |
Holotype : désigné par R. (1957), c'est la fig. 7 de la Pl. 1.
Localité : Medjez Sfa, Algérie.
Âge : Barrémien basal.
Matériel étudié : 103 spécimens collection MD1647, 1649, 1650, 1651, 1652, 1653, 1654, 1655, 1656, 1657, 1658, 1659, 1660, 1661, 1662, 1663, 1664, 1665, 1666, 1667, 1668 (forme à constrictions serrées), 1669, 1670, 1671, 1672, 1673, 1674, 1675, 1676, 1677, 1678, 1679, 1680, 1681, 1682, 1683, 1684, 1685, 1715, 2453, 2454, 2490, 2491, 2492, 2493, 2494, 2495, 2496 et 2497.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1650 (1) | 28,7 | 15,1 | 12,0 | 4,2 | 0,53 | 0,42 | 0,15 | 0,79 |
| 1663 | 29,0 | 16,7 | 11,6 | 3,6 | 0,58 | 0,40 | 0,12 | 0,69 |
| 1667 | 29,5 | 15,7 | 12,3 | 3,9 | 0,53 | 0,42 | 0,13 | 0,78 |
| 1668 (2) | - | 22,0 | 17,1 | - | - | - | - | 0,78 |
| 1670 | 32,7 | 18,4 | 14,5 | 4,6 | 0,56 | 0,44 | 0,14 | 0,79 |
| 1679 (3) | 32,3 | 17,0 | - | 4,0 | 0,53 | - | 0,12 | - |
| - | 15,5 | 13,0 | 3,3 | - | - | - | 0,84 | |
| (1) forme à constrictions profondes | ||||||||
| (2) forme à constrictions serrées | ||||||||
| (3) forme épaisse | ||||||||
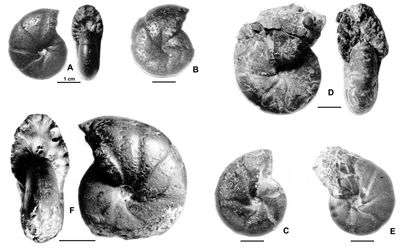
Cliquer sur la vignette pour agrandir l'image.
Figure 43 : Salfeldiella guettardi gracile ,
1957.
A : MD1667a U2i/GB : Aptien moyen (Zone à Martini) - Gaubert, Alpes-de-Haute-Provence.
B : MD1650a - U2i/GB : Aptien moyen (Zone à Martini) - Ravin du Terrois, Drôme.
C : MD1670 - U2i/GB : Aptien moyen (Zone à Martini) - Gaubert, Alpes-de-Haute-Provence.
D : MD1679a forme épaisse - U2i/GB : Aptien moyen (Zone à Martini) -
Gaubert, Alpes-de-Haute-Provence.
E : MD1663 U3s/VL : Aptien moyen (Zone à Melchioris) - Serre Chaïtieu, Drôme.
F : MD1668 forme à constrictions serrées - U2i/GB : Aptien moyen (Zone à Martini) - Gaubert, Alpes-de-Haute-Provence.
Description : moules internes pyritisés lisses à l'exception d'indice d'une costulation ventrale chez quelques spécimens, comprimés, à flancs presque plats ou légèrement convexes et ombilic un peu ouvert. Le maximum de l'épaisseur du tour est vers le milieu du tour, plus près de l'ombilic chez les spécimens les plus épais, section du tour elliptique. Le dernier tour porte 5 à 6 constrictions falciformes légèrement proverses sur le bord siphonal.
La ligne cloisonnaire (Fig. 44 ![]() ) est celle de S. guettardi.
) est celle de S. guettardi.
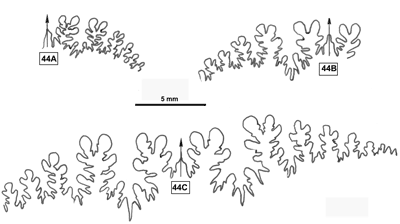
Cliquer sur la vignette pour agrandir l'image.
Figure 44 : Lignes cloisonnaires de Salfeldiella guettardi gracile.
A : MD1664a U3s/VL : Aptien moyen (Zone à Melchioris) Serre Chaïtieu, Drôme.
B: MD1667a U2i/GB : Aptien moyen (Zone à Martini) Gaubert, Alpes-de-Haute-Provence.
Affinités : les spécimens vocontiens étudiés ici ne diffèrent de S. guettardi que par leurs flancs aplatis et la forme de la section. Au sein de la population globale S. guettardi + S. guettardi gracile et S. guettardi victoriana, S. guettardi gracile rassemble les individus les moins épais. C'est, au total la principale différence entre les guettardi typiques et gracile. Il semble intéressant de conserver la distinction entre les deux formes. D'abord parce que les spécimens vocontiens, que nous rapprochons de l'espèce créée par
(1957), ont un aspect très proche des illustrations que ce dernier a donné ; ils sont seulement un peu plus épais. En quelque sorte ils sont intermédiaires entre les guettardi typiques et les gracile d'Algérie. D'autre part, on peut être frappé par le fait que R. n'a pas cité la présence de guettardi en Algérie. Quoi qu'il en soit l'allure unimodale des histogrammes de h, e, o et l donne bien l'image d'une population homogène (Fig. 45 ![]() ).
).
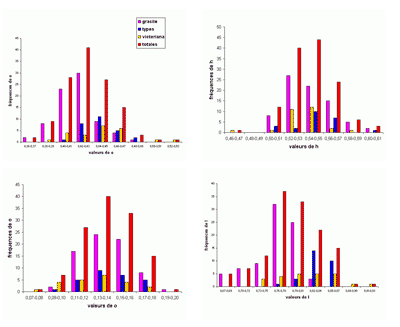
Cliquer sur la vignette pour agrandir l'image.
Figure 45 : Histogrammes des fréquences de e, h, o et l chez Salfeldiella guettardi et ses sous-espèces.
Le taxon gracile est donc interprété comme étant une sous-espèce du taxon guettardi apparue avant les formes typiques. L'absence du taxon gracile dans la partie moyenne de l'Aptien inférieur des Marnes Bleues (voir S. aff. guettardi) est difficilement explicable si les formes les plus anciennes de gracile sont barrémiennes, alors qu'une petite population de formes rapprochées du type et du taxon victoriana existe déjà. Remarquons que (1957) a figuré des spécimens de taille modeste qui peuvent avoir la morphologie de certains Phylloceras (Hypophylloceras) comme P. (H.) paquieri. La connaissance des lignes cloisonnaires de ces spécimens pourrait éclairer la question de leurs affinités. Rappelons que la ligne cloisonnaire de P. (H.) paquieri est clairement celle d'un Phylloceras (Hypophylloceras). Les lignes cloisonnaires figurées par (1957) sont celles d'un spécimen de l'Aptien du Djebel Ouach, elles appartiennent incontestablement à un spécimen du genre Salfeldiella. Ces remarques amènent à penser que les S. gracile d'Algérie sont plus probablement ceux de l'Aptien que ceux du Barrémien. De plus, il ne fait aucun doute, que les récoltes vocontiennes établissent clairement que gracile est une sous-espèce de guettardi. La sous-espèce est plus comprimée que les individus les plus typiques et sa section est elliptique au lieu d'être ovale.
Le spécimen que (1964, Pl. 12, fig. 6) rattache à l'espèce P. (H.) grothi est en réalité très proche de S. guettardi gracile par ses dimensions et l'allure de ses constrictions. En revanche si la ligne cloisonnaire de la fig. 4a dans le texte de (1920b) appartient bien à un spécimen de l'espèce grothi relevée à un petit diamètre (D = 6 mm), elle est sans aucun doute celle d'un Phylloceras (Hypophylloceras), avec S1 tétraphylle. L'espèce grothi de (non ) est un incontestable Phylloceras (Hypophylloceras) et ne peut se confondre avec gracile. Le grothi figuré par (1989) a bien la morphologie de la sous-espèce gracile, malheureusement ses dimensions sont inconnues.
Âge : S. guettardi gracile a été récoltée en Algérie dans le Barrémien basal à l'Aptien (, 1957). Elle est peut-être apparue avant les guettardi typiques. Le spécimen des Baléares provient de l'Aptien supérieur ; celui figuré par (1989) provient de l'Aptien inférieur-moyen du Vorarlberg.
Les diverses récoltes des Marnes Bleues vocontiennes s'échelonnent de la fin de l'Aptien inférieur (Zone à Furcata) à la fin de l'Aptien moyen (Zone à Melchioris) mais l'espèce atteint son acmé à la base de l'Aptien moyen (Zone à Martini).
Salfeldiella guettardi (, 1831) victoriana (, 1913)
| 1883 | Phylloceras cf. guettardi (), , p. 182, Pl. 4, figs. 9.a-b (= var. Victoriana ), fig. 10. | |
| 1913 | Phylloceras guettardi (), , p. 332. | |
| 2005 | Salfeldiella guettardi ().- , p. 62, Pl. 4, fig. 10 (non figs. 6-9) |
Lectotype de victoriana : fig. 9.a-b, Pl. 4, (1883).
Localité : Malenovice, Carpathes, République Tchèque.
Âge : Barrémien (couches de Wernsdorf = couches de Verovice ?).
Matériel étudié : 37 spécimens collection MD1686, 1688, 1689, 1690, 1691, 1692, 1693, 1694, 1695, 1696, 1697, 1698, 1699, 1700, 1701, 1702, 2446 (dont un microconque), 2449, 2455, 2456, 2457, 2458, 2460, 2498, 2499, 2500 et 2501.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1691 | 36,6 | 20,7 | 16,0 | 4,0 | 0,57 | 0,44 | 0,11 | 0,77 |
| 1692 | 29,8 | 16,8 | 15,6 | 3,0 | 0,56 | 0,52 | 0,10 | 0,93 |
| 1693 | 29,4 | 15,6 | 11,6 | 3,9 | 0,53 | 0,39 | 0,13 | 0,74 |
| 1694 | 35,5 | 19,7 | 14,5 | 4,4 | 0,55 | 0,41 | 0,12 | 0,74 |
| 1695 | 38,0 | 20,5 | 17,2 | 5,4 | 0,54 | 0,45 | 0,14 | 0,84 |
| 2446 (1) | 15,5 | 8,3 | 6,5 | 2,2 | 0,54 | 0,42 | 0,14 | 0,78 |
| (1) microconque | ||||||||
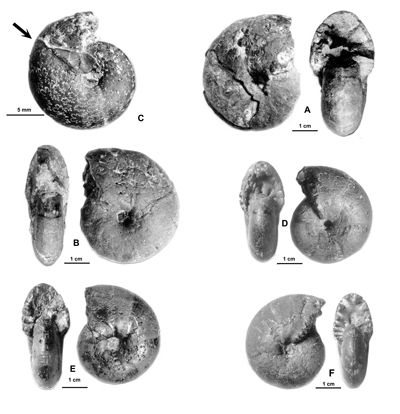
Cliquer sur la vignette pour agrandir l'image.
Figure 46 : Salfeldiella guettardi victoriana (,
1913).
A: MD1692 spécimen épais U2i/GB : Aptien moyen (Zone à Martini) - Gaubert, Hautes-Alpes.
B : MD1693a spécimen aplati U2i/GB : Aptien moyen (Zone à Martini) - Gaubert, Hautes-Alpes.
C : MD2446 microconque (flèche au début de la loge) - U2i/GB : Aptien moyen (Zone à Martini) -
Gaubert, Hautes-Alpes.
D : MD1691 - U2i/GB : Aptien moyen (Zone à Martini) - Gaubert, Hautes-Alpes.
E : MD1695 - U2i/GB : Aptien moyen (Zone à Martini) - Gaubert, Hautes-Alpes.
F : MD1694 - U2i/GB : Aptien moyen (Zone à Martini) - Gaubert, Hautes-Alpes.
Description : moules internes pyritisés, épais (mais l'épaisseur est tout de même variable, voir par exemple Fig. 46.A-B ![]() ), lisses le plus souvent mais pouvant montrer une fine costulation ventrale débutant de façon ténue sur la partie
externe des flancs. Le moule interne est toujours lisse autour de l'ombilic, l'ombilic est légèrement ouvert, la section du tour est ovale.
), lisses le plus souvent mais pouvant montrer une fine costulation ventrale débutant de façon ténue sur la partie
externe des flancs. Le moule interne est toujours lisse autour de l'ombilic, l'ombilic est légèrement ouvert, la section du tour est ovale.
Les constrictions falciformes sur les flancs, et fortement proverses sur le bord ventral, sont assez fortes au début du tour, elles s'effacent ensuite plus ou moins complètement. Le spécimen microconque (Fig. 46.C ![]() ) montre deux constrictions obsolètes au début du dernier tour, ensuite il est complètement lisse, il ne montre qu'une partie de la loge.
) montre deux constrictions obsolètes au début du dernier tour, ensuite il est complètement lisse, il ne montre qu'une partie de la loge.
Le S. guettardi figuré par (2005, Pl. 4, fig. 10) a bien la morphologie de l'espèce guettardi mais ses constrictions sont effacées, ce spécimen appartient donc à la sous-espèce victoriana.
La ligne cloisonnaire (Fig. 47 ![]() ) des grands spécimens atteint la complexité que l'on observe chez les S. guettardi typiques, elle est toujours diphylle-triphylle avec des phyllites terminaux spatulés, mais à la différence des petits Salfeldiella on observe le développement de petits phyllites internes à la base des gros phyllites. Le lobe siphonal est sensiblement moins profond que le lobe L.
) des grands spécimens atteint la complexité que l'on observe chez les S. guettardi typiques, elle est toujours diphylle-triphylle avec des phyllites terminaux spatulés, mais à la différence des petits Salfeldiella on observe le développement de petits phyllites internes à la base des gros phyllites. Le lobe siphonal est sensiblement moins profond que le lobe L.
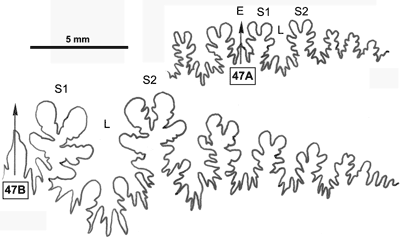
Cliquer sur la vignette pour agrandir l'image.
Figure 47 : Lignes cloisonnaires de Salfeldiella guettardi victoriana.
A : MD2446 U2i/GB : Aptien moyen (Zone à Martini) Gaubert, Alpes-de-Haute-Provence.
B : MD2449 U2i/GB : Aptien moyen (Zone à Martini) Ravin du Terrois, Drôme.
Affinités : S. guettardi victoriana est considérée comme une sous-espèce de S. guettardi dont elle se distingue surtout par l'effacement de ses constrictions (, 2000a, p. 152).
Les nombreux spécimens récoltés dans les Marnes Bleues vocontiennes ont permis de constater que ce taxon constituait une population bien proche des S. guettardi sensu stricto par ses paramètres relatifs (voir Fig. 52 ![]() ).
).
Âge : S. guettardi victoriana a été récoltée de la fin de l'Aptien inférieur (sommet de la Zone à Furcata) à l'Aptien moyen (base de la Zone à Melchioris) avec son acmé à la base de l'Aptien moyen (Zone à Martini).
Salfeldiella aff. guettardi (, 1831)
Matériel étudié : 2 spécimens collection MD1648.
Dimensions du spécimen figuré
| N° MD | D | H | E | O | h | e | o | l |
| 1648b | 11,2 | 6,2 | 5,1 | 2,0 | 0,55 | 0,45 | 0,18 | 0,82 |

Cliquer sur la vignette pour agrandir l'image.
Figure 48 : Salfeldiella aff. guettardi (,
1831).
MD1648b U1s/?TR : Aptien inférieur (Zone à Furcata) - Gaubert, Hautes-Alpes.
Description : les trois spécimens étudiés sont des moules internes pyritisés, lisses, à maximum d'épaisseur vers l'ombilic, à section du tour ovale épaisse, ombilic un peu ouvert. Le dernier tour porte 6 constrictions droites ou à peine incurvées au milieu des flancs et proverses chez l'un des spécimens (D = 11,2 mm). Un autre spécimen porte 5 constrictions (D = 7,6 mm) mais moins régulièrement réparties sur le dernier tour. Le spécimen incomplet a des constrictions effacées comme chez la sous-espèce S. guettardi victoriana.
La ligne cloisonnaire du plus grand spécimen est de type Salfeldiella, c'est-à-dire à gros phyllites terminaux sans échancrure à leur base.
Les trois spécimens diffèrent des spécimens du sommet de l'Aptien inférieur et de l'Aptien moyen par les flancs plus bombés, les constrictions plus droites, l'ombilic plus ouvert et leur petite taille. La forme de la section est celle des formes les plus typiques de l'espèce. Ces modestes différences sont probablement dues à la petite taille des spécimens. Ils sont actuellement les plus anciens, parmi les spécimens connus rattachés à l'espèce S. guettardi, ils sont certainement les représentants d'une population ancestrale de l'espèce.
Âge : les spécimens vocontiens étudiés ici ont été récoltés à la fin de l'Aptien inférieur (sommet Zone à Furcata). Au même niveau existe un spécimen incomplet rattaché à la sous-espèce victoriana.
Conclusion d'ensemble sur l'espèce Salfeldiella guettardi : prise dans son ensemble, l'espèce apparaît extrêmement polymorphe. Ce constat va à l'encontre de l'opinion assez générale selon laquelle les Phyllocératidés auraient été peu variables. Il faut nuancer cette opinion. En fait, l'impression de faible variabilité est liée à la grande durée de vie de beaucoup d'espèces (, 2000a). L'espèce étant stable dans le temps cela donne l'impression d'une faible variabilité du groupe. Quand les espèces eurent une plus faible durée de vie, et c'est le cas de l'espèce S. guettardi dans son ensemble, la variabilité est plus importante.
On peut admettre que l'espèce comprend :
Ces coupures peuvent apparaître comme artificielles. Toutefois, elles paraissent se justifier du fait qu'elles se reconnaissent assez facilement sur les histogrammes des fréquences (voir Fig. 45 ![]() ).
).
Salfeldiella opegana (, 1920)
| V | 1920 | Phylloceras guettardi () var. opegana , p. 197, Pl. 1, fig. 6 & 6.a-b. |
| 2000a | Salfeldiella opegana ().- , p. 152, Pl. 37, figs. 8.a-b, 9.a-b & 10.a-b ; figs. 351-355 dans le texte. | |
| 2005 | Salfeldiella opegana ().- , p. 60, Pl. 4, figs. 1-5. | |
| Synonymie plus complète, holotype, localité-type, âge de l'holotype de la sous-espèce : , 2000a, p. 15. | ||
Matériel étudié : 30 spécimens collection MD1703, 1704, 1705, 1706, 1707, 1708, 1709, 1710, 1711, 1712, 1713 et 1714.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1704a | 35,5 | 19,1 | 15,6 | 4,3 | 0,54 | 0,44 | 0,12 | 0,82 |
| 1704b | 42,0 | 24,3 | 20,4 | 5,0 | 0,58 | 0,49 | 0,12 | 0,84 |
| 1705 | 48,1 | 27,4 | 21,6 | 5,3 | 0,57 | 0,45 | 0,11 | 0,79 |
| 1711 | 19,4 | 10,7 | 8,4 | 3,2 | 0,55 | 0,43 | 0,16 | 0,79 |
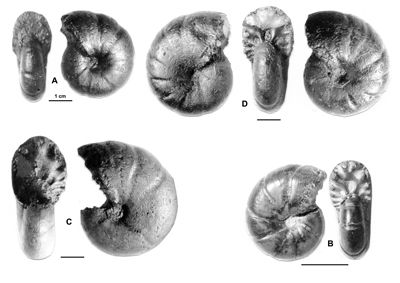
Cliquer sur la vignette pour agrandir l'image.
Figure 49 : Salfeldiella opegana (,
1920).
A : MD1704a U1s/TR : Aptien inférieur (Zone à Furcata) - Ravin du Terrois, Drôme.
B : MD1711 - U1s/TR : Aptien inférieur (Zone à Furcata) - Ravin du Rivet,
Alpes-Maritimes.
C : MD1705a - U1s/TR : Aptien inférieur (Zone à Furcata) - Ravin du Terrois,
Drôme.
D : MD1704b - U1s/TR : Aptien inférieur (Zone à Furcata) - Ravin du Terrois, Drôme.
Description : la plupart des spécimens sont des moules internes pyritisés, lisses, flancs aplatis, au bord externe arrondi et section du tour subrectangulaire. Quand le test est conservé, on observe des costules développées surtout sur le bord ventral (MD1708 et MD1711 en Fig. 49.B ![]() par exemple). Le test et le moule sont parcourus de nombreuses constrictions (on en compte environ 6 chez les petits spécimens et une dizaine chez les plus grands). Elles sont droites ou légèrement falciformes, proverses sur les flancs, très proverses sur le bord ventral où elles forment un chevron très accentué chez les plus grands spécimens.
par exemple). Le test et le moule sont parcourus de nombreuses constrictions (on en compte environ 6 chez les petits spécimens et une dizaine chez les plus grands). Elles sont droites ou légèrement falciformes, proverses sur les flancs, très proverses sur le bord ventral où elles forment un chevron très accentué chez les plus grands spécimens.
La ligne cloisonnaire (Fig. 50 ![]() ) est assez découpée pour une espèce du genre Salfeldiella, mais le style en reste diphylle-triphylle avec des phyllites terminaux développés et spatulés. Le lobe L est sensiblement moins profond que le lobe siphonal.
) est assez découpée pour une espèce du genre Salfeldiella, mais le style en reste diphylle-triphylle avec des phyllites terminaux développés et spatulés. Le lobe L est sensiblement moins profond que le lobe siphonal.
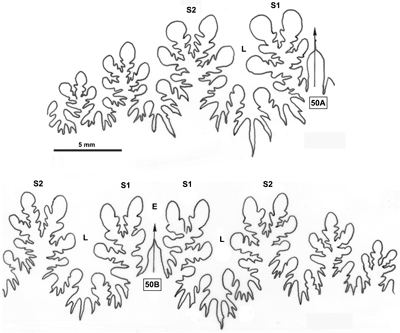
Cliquer sur la vignette pour agrandir l'image.
Figure 50 : Lignes cloisonnaires de Salfeldiella opegana.
A : MD1703a U1s/TR : Aptien inférieur (Zone à Furcata) Ravin du Terrois, Drôme.
B : MD1703b U1s/TR : Aptien inférieur (Zone à Furcata) Ravin du Terrois, Drôme.
Affinités : S. opegana avait été décrite comme variété de S. guettardi par (1920, p. 197). Cependant, elle a trop de différences avec cette dernière et les formes intermédiaires n'existent pas vraiment. La variété a donc récemment été élevée au rang d'espèce (, 2000a, p. 152).
Les histogrammes des fréquences des paramètres relatifs des espèces S. guettardi et S. opegana (Fig. 51 ![]() ) montrent d'ailleurs que les deux taxons forment des populations bien distinctes, surtout pour les paramètres relatifs e, o et l. S. opegana est plus épaisse que S. guettardi mais son ombilic est moins ouvert.
) montrent d'ailleurs que les deux taxons forment des populations bien distinctes, surtout pour les paramètres relatifs e, o et l. S. opegana est plus épaisse que S. guettardi mais son ombilic est moins ouvert.
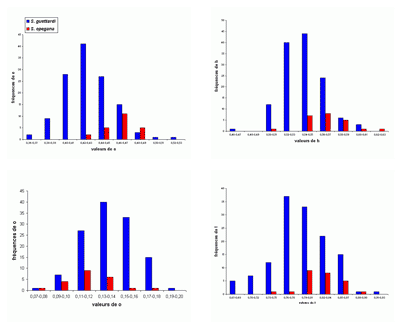
Cliquer sur la vignette pour agrandir l'image.
Figure 51 : Histogrammes des fréquences de e, h, o et l chez Salfeldiella guettardi et Salfeldiella opegana.
Âge : S. opegana avait été signalée dans le "Gargasien" (, 2000a, p. 153). Dans les Marnes Bleues vocontiennes, elle est surtout abondante dans l'Aptien inférieur (Zone à Furcata et Zone à Deshayesi). De rares spécimens ont été récoltés dans l'Aptien moyen (Zone à Martini : MD1714) et la base de l'Aptien supérieur (Zone à Nolani : MD1713).
Sous-genre Gyrophyllites , 1964
Coquille lisse à section du tour subcirculaire, constrictions radiales, E aussi profond que L (, 2000a, p. 11).
Salfeldiella (Gyrophyllites) falloti nov. sp.
| Non ? | 1920b | Phylloceras grothi , p. 20, Pl. 1, figs. 3.a-b, 4.a-b, 5.a-b & 6.a-b ; non figs. 4.a-b & 5 dans le texte. |
| 1920b | Phylloceras cf. lytoceroide , p. 30, Pl. 1, fig. 12.a-c. | |
| 1964 | Sowerbyceras (Gyrophyllites) lytoceroides ().- , p. 261, Pl. 17, fig. 7a-c ; fig. 63 dans le texte. |
Holotype : collection MD1580a.
Localité : Les Vallons, commune de Tartonne, Alpes-de-Haute-Provence.
Âge : Aptien moyen (Zone à Melchioris).
Origine du nom : en hommage à Paul à qui l'on doit une importante étude sur les ammonites de l'Aptien et l'Albien des Baléares ainsi qu'une autre étude en collaboration avec Henri également sur sur les faunes des Baléares.
Matériel étudié : 174 spécimens collection MD1568, 1569, 1570 (non mesuré), 1571, 1572, 1573, 1575, 1576, 1578, 1579, 1580, 1581, 1582, 1583, 1584, 1585, 1586 (non mesuré), 1587, 1588, 1589, 1590 (non mesuré), 1591, 1592, 1593, 1594, 1595, 1596, 1597, 1598, 1599, 1600, 1601, 1602, 1603, 2447, 2461, 2462, 2463, 2502, 2503, 2504, 2505, 2506, 2507 et 2508.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1569 | 8,4 | 5,2 | 3,6 | 2,1 | 0,62 | 0,43 | 0,25 | 0,69 |
| 1571 | 14,0 | 7,3 | 5,6 | 3,0 | 0,52 | 0,40 | 0,21 | 0,77 |
| 1578 | 10,0 | 5,5 | 4,2 | 2,1 | 0,55 | 0,42 | 0,21 | 0,76 |
| 1580a | 22,8 | 11,7 | 10,1 | 4,8 | 0,51 | 0,44 | 0,21 | 0,86 |
| 1580b | 21,5 | 10,6 | 9,5 | 5,0 | 0,49 | 0,44 | 0,23 | 0,90 |
| 1594a | 12,6 | 6,6 | 5,3 | 3,0 | 0,52 | 0,42 | 0,24 | 0,80 |
| 1594b | 17,0 | 8,3 | 6,8 | 3,1 | 0,49 | 0,40 | 0,18 | 0,82 |
| 1596 | 12,0 | 5,9 | 5,6 | 3,4 | 0,49 | 0,47 | 0,28 | 0,95 |
| 1598 | 16,2 | 9,0 | - | - | 0,56 | - | - | - |
| 15,2 | 8,9 | 6,9 | 2,0 | 0,59 | 0,45 | 0,13 | 0,78 | |
| 1599 | 9,0 | 4,8 | 4,4 | 2,5 | 0,53 | 0,50 | 0,28 | 0,92 |
| 2447b | 16,7 | 8,3 | 7,3 | 3,5 | 0,50 | 0,44 | 0,21 | 0,88 |
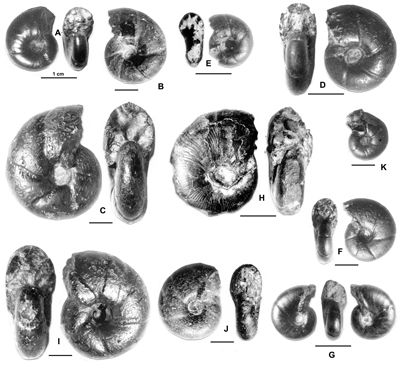
Cliquer sur la vignette pour agrandir l'image.
Figure 52 : Salfeldiella (Gyrophyllites) falloti nov. sp.
A : MD1599 U4m/BT : Aptien moyen (Zone à Melchioris) - Les Vallons, Alpes-de-Haute-Provence.
B : MD2447b U3s : Aptien moyen (Zone à Melchioris) - Ravin des Prés, Alpes-de-Haute-Provence.
C : MD1580a holotype - U4m/BT : Aptien moyen (Zone à Melchioris) - Les
Vallons, Alpes-de-Haute-Provence.
D : MD1594a U3s/G1-NO : Aptien moyen (Zone à Melchioris) - Gaubert, Hautes-Alpes.
E : MD1596 U5i/SZ : Aptien supérieur (Zone à Jacobi) - Les Sauzeries, Alpes-de-Haute-Provence.
F : MD1571 U3s/VL : Aptien moyen (Zone à Melchioris) - Les Vallons, Alpes-de-Haute-Provence.
G : MD1569 U4m/BT : Aptien moyen (Zone à Melchioris) - Les Vallons, Alpes-de-Haute-Provence.
H : MD1598 spécimen à test conservé U2i/GB : Aptien moyen (Zone à Martini) - Gaubert, Alpes-de-Haute-Provence.
I : MD1580b U4m/BT : Aptien moyen (Zone à Melchioris) - Les Vallons, Alpes-de-Haute-Provence.
J : MD1594b U3s/G1-NO : Aptien moyen (Zone à Melchioris) - Gaubert, Alpes-de-Haute-Provence.
K : MD1578 U2i/GB : Aptien moyen (Zone à Martini) - Ravin du Terrois, Drôme.
Diagnose : coquille à test finement costulé, moule interne lisse, ombilic ouvert, flancs presque plats, bord externe arrondi, section du tour subquadratique, constrictions du moule interne proverses pouvant être radiales chez les plus petits spécimens, à sinus ventral chez les plus grands spécimens, ligne cloisonnaire de type Gyrophyllites.
Description : moules internes pyritisés, lisses, épais, ombilic assez ouvert laissant voir les tours internes, flancs presque plats, bord externe arrondi, section du tour subquadratique. Le dernier tour porte 3 à 7 constrictions droites plus ou moins profondes, proverses sur les flancs et le bord ventral. Chez les plus grands spécimens, la constriction sur le bord ventral forme un véritable sinus (Fig. 52.C et I ![]() par exemple). Chez des spécimens plus petits comme MD2461 (D = 14,0 mm), les constrictions sont proverses sauf l'avant dernière (près de l'ouverture) qui se redresse avant de passer le bord ventral qu'elle passe en direction pratiquement radiale. Un autre spécimen (Fig. 52.A
par exemple). Chez des spécimens plus petits comme MD2461 (D = 14,0 mm), les constrictions sont proverses sauf l'avant dernière (près de l'ouverture) qui se redresse avant de passer le bord ventral qu'elle passe en direction pratiquement radiale. Un autre spécimen (Fig. 52.A ![]() à D = 9,0 mm), muni de son test, porte 3 constrictions, le test est finement costulé entre les constrictions, la costulation est nette surtout sur le bord ventral. Sur le côté gauche, ce spécimen porte les traces d'une blessure qui a perturbé le tracé des costules, disposition assez comparable à celle qui a été signalée chez un Calliphylloceras du Lias (,
1972, Pl. 1, fig. 8). Le plus petit des spécimens étudiés (Fig. 52.G
à D = 9,0 mm), muni de son test, porte 3 constrictions, le test est finement costulé entre les constrictions, la costulation est nette surtout sur le bord ventral. Sur le côté gauche, ce spécimen porte les traces d'une blessure qui a perturbé le tracé des costules, disposition assez comparable à celle qui a été signalée chez un Calliphylloceras du Lias (,
1972, Pl. 1, fig. 8). Le plus petit des spécimens étudiés (Fig. 52.G ![]() ) porte une fine costulation surtout visible sur le bord siphonal. Les costules partent de l'ombilic en faisceaux soulignés par des replis, caractère de Phylloceras.
) porte une fine costulation surtout visible sur le bord siphonal. Les costules partent de l'ombilic en faisceaux soulignés par des replis, caractère de Phylloceras.
La ligne cloisonnaire (Fig. 53 ![]() ) est très simplifiée, les selles S1 et S2 sont du type diphylle-diphylle (tendance triphylle ?), E est un peu moins profond que L. Elle est du même type que
les figures 8 et 9 de (1920b, p. 29), c'est-à-dire de type Gyrophyllites.
) est très simplifiée, les selles S1 et S2 sont du type diphylle-diphylle (tendance triphylle ?), E est un peu moins profond que L. Elle est du même type que
les figures 8 et 9 de (1920b, p. 29), c'est-à-dire de type Gyrophyllites.
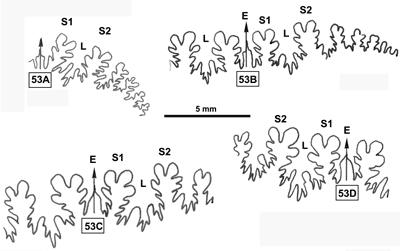
Cliquer sur la vignette pour agrandir l'image.
Figure 53 : Lignes cloisonnaires de Salfeldiella (Gyrophyllites) falloti.
A : MD2462a U3s/VL : Aptien moyen (Zone à Melchioris) Les Vallons, Alpes-de-Haute-Provence.
B : MD1594a U3s/G1-NO : Aptien moyen (Zone à Melchioris) Gaubert, Alpes-de-Haute-Provence.
C : MD1587a U3s/G1-NO : Aptien moyen (Zone à Melchioris) Gaubert, Alpes-de-Haute-Provence.
D : MD1588a U3s/G1-NO : Aptien moyen (Zone à Melchioris) Gaubert, Alpes-de-Haute-Provence.
Affinités : la morphologie des spécimens vocontiens est parfaitement identique aux illustrations données par pour ses "Phylloceras cf. lytoceroides" (1920b, fig. 12). La ligne cloisonnaire est du même type avec toutefois un lobe E moins profond que L chez les spécimens des Marnes Bleues. Cela s'explique peut-être par une plus grande taille des spécimens que celui figuré par . L'ombilic est moins ouvert que chez les S. lytoceroides des Baléares, ce qui justifie de faire de la population des Marnes Bleues une nouvelle espèce.
L'espèce la plus proche morphologiquement de S. (G.) falloti étant S. opegana, il était intéressant de comparer les histogrammes des fréquences de leurs paramètres relatifs (Fig. 54 ![]() ). On constate qu'il y a deux populations bien distinctes pour les paramètres relatifs e, h et l, pour o la différence est encore plus nette. S. (G.) falloti a une hauteur du tour et une épaisseur moindre que chez opegana. En revanche, elle a un ombilic sensiblement plus ouvert.
). On constate qu'il y a deux populations bien distinctes pour les paramètres relatifs e, h et l, pour o la différence est encore plus nette. S. (G.) falloti a une hauteur du tour et une épaisseur moindre que chez opegana. En revanche, elle a un ombilic sensiblement plus ouvert.
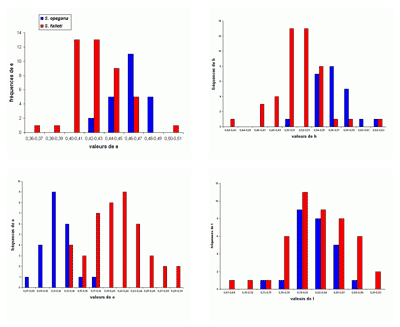
Cliquer sur la vignette pour agrandir l'image.
Figure 54 : Histogrammes des fréquences de e, h, o et l chez Salfeldiella opegana et Salfeldiella (Gyrophyllites) falloti.
Âge : S. (G.) falloti a été récoltée de la fin de l'Aptien inférieur (Zone à Furcata) au début de l'Aptien supérieur (Zone à Nolani). Le spécimen figuré par provient de l'Hauterivien de Noyers-sur-Jabron (Alpes-de-Haute-Provence). Aux Baléares, l'espèce S. lytoceroides serait d'âge Aptien supérieur (, 1964, p. 262) ; en Espagne du Sud (Province de Murcie), elle proviendrait de l'Albien (, 1964, p. 262).
Salfeldiella (Gyrophyllites) micipsa (, 1880)
| 1880 | Ammonites micipsa , p. 24. | |
| 1886 | Ammonites micipsa , la figure non numérotée se trouve dans l'Atlas de , fossiles décrits par , Pl. 1. | |
| 1920b | Phylloceras micipsa ().- , p. 26, Pl. 1, figs. 7.a-c & 8.a-c ; figs. 6-7 dans le texte. |
Néotype : le type de lespèce micipsa nayant pas été retrouvé par Ottlia (Muséum dHistoire Naturelle de Budapest), nous proposons la fig. 7.a-c de la Pl. 1 de (1920b) comme néotype.
Localité : Oued Cheniour, Algérie. Collection Université de Grenoble, n° UJF-ID.12 pour le spécimen de la fig. 7.a-c (, 1920b, Pl. 1) désigné comme néotype.
Âge : Aptien.
Matériel étudié : 2 spécimens collection MD1604.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1604a | 13,7 | 7,3 | 5,5 | 2,3 | 0,53 | 0,40 | 0,17 | 0,75 |
| 1604b | 10,4 | 5,4 | 4,3 | 2,3 | 0,52 | 0,41 | 0,22 | 0,80 |
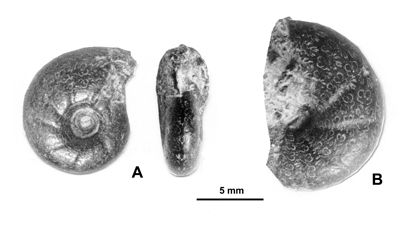
Cliquer sur la vignette pour agrandir l'image.
Figure 55 : Salfeldiella (Gyrophyllites) micipsa (,
1880).
A : MD1604a U3s/VL : Aptien moyen (Zone à Melchioris) - Les Vallons, Alpes-de-Haute-Provence.
B : MD1604b - U3s/VL : Aptien moyen (Zone à Melchioris) - Les Vallons, Alpes-de-Haute-Provence.
Description : deux petits moules internes sont rapportés à l'espèce créée par telle que
(1920b, p. 26) l'a interprétée. Ils sont lisses, assez épais, portant des constrictions serrées sur le dernier tour, 6 pour le dernier demi-tour du plus grand spécimen (Fig. 55.B ![]() ), 7 pour le dernier tour du second spécimen (Fig. 55.A
), 7 pour le dernier tour du second spécimen (Fig. 55.A ![]() ), ce qui est un nombre élevé pour des Salfeldiella de petite taille. Ces constrictions sont droites et fortement proverses sur les flancs et le bord ventral. Les flancs sont à peine convexes et la section est ovale.
), ce qui est un nombre élevé pour des Salfeldiella de petite taille. Ces constrictions sont droites et fortement proverses sur les flancs et le bord ventral. Les flancs sont à peine convexes et la section est ovale.
La ligne cloisonnaire (Fig. 56 ![]() ) est de type Salfeldiella, elle est très peu découpée et très semblable
aux figures 6 et 7 de (1920b, p. 27), les selles S1 et S2 sont diphylles, et non diphylles-triphylles cas le plus habituel.
) est de type Salfeldiella, elle est très peu découpée et très semblable
aux figures 6 et 7 de (1920b, p. 27), les selles S1 et S2 sont diphylles, et non diphylles-triphylles cas le plus habituel.
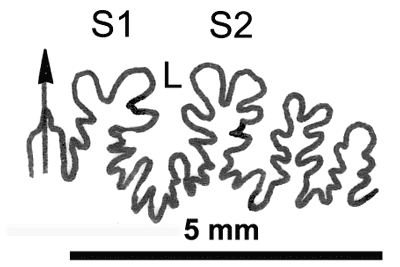
Cliquer sur la vignette pour agrandir l'image.
Figure 56 : Ligne cloisonnaire de Salfeldiella (Gyrophyllites) micipsa.
MD1604a U3s/VL : Aptien moyen (Zone à Melchioris) Les Vallons, Alpes-de-Haute-Provence.
Affinités : les dimensions des deux spécimens vocontiens correspondent bien à celles données par pour les ammonites de l'Oued Cheniour en Algérie. Il n'y a pas de différences notables entre les spécimens des Marnes Bleues vocontiennes et ceux d'Algérie. Ce sont bien des micipsa au moins au sens où avait interprété l'espèce décrite par (1880). pensait que l'espèce créée par n'avait jamais été figurée avant lui, il se trompait puisque l'espèce micipsa est figurée dans l'Atlas de (1886). Mais la figure publiée dans l'Atlas est médiocre et pratiquement inexploitable. L'interprétation de l'espèce donnée par permet de conserver le nom micipsa avec une diagnose et des illustrations de qualité. Nous pensons que l'espèce micipsa doit être conservée en acceptant l'interprétation de .
Âge : les deux spécimens vocontiens de Salfeldiella (Gyrophyllites) micipsa sont datés du sommet de l'Aptien moyen (Zone à Melchioris). Les spécimens d'Algérie sont datés de l'Aptien probablement moyen.
Sous-famille des Phyllopachyceratinae , 1937
Genre Phyllopachyceras , 1925
Les Phyllopachyceras pyriteux du Crétacé ont la réputation d'être difficiles à distinguer les uns des autres. Pourtant on verra plus loin que lorsque l'on dispose de nombreux spécimens bien datés la distinction des taxons est moins malaisée que ce que l'on croit habituellement.
Phyllopachyceras baborense (, 1880)
| 1880 | Ammonites baborensis , p. 26. | |
| 1886 | Ammonites baborensis in , Pl. 1, 2 figures non numérotées pour 1 spécimen. | |
| ? | 1989 | Partschiceras baborense ().- , p. 114, Pl. 1, fig. 4. |
| 2000a | Phyllopachyceras baborense ().- , p. 153, Pl. 37, fig. 11.a-b ; fig. 356-360 dans le texte. | |
| (pars) | 2000 | Phyllopachyceras baborense ().- et alii, p. 656, fig. 40.a-c, fig. 39.j-k ?, non fig. 40.d-f (= P. rouyanum), non fig. 39.f-h, l-n (= P. rouyanum). |
| 2005 | Phyllopachyceras baborense ().- , p. 77, Pl. 3, figs. 13-15. | |
| Synonymie plus complète, néotype, localité-type, âge du néotype : , 2000a, p. 153. | ||
Matériel étudié : 193 spécimens collection MD1718, 1721, 1722, 1723, 1724, 1726, 1727, 1730, 1733, 1735, 1736, 1738, 1739, 1741, 1742, 1743, 1747, 1756, 1761, 1764 (non mesuré), 1765, 1768, 1769, 1774, 1779, 1789, 1790, 1791, 1793, 1794, 1796, 1797, 1801, 2464, 2465, 2466, 2467, 2509, 2510, 2511, 2512, 2513 et 2514.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1735 | 21,0 | 12,4 | 11,4 | 1,3 | 0,59 | 0,54 | 0,06 | 0,92 |
| 1742a | 24,4 | 16,0 | 13,0 | 1,5 | 0,66 | 0,53 | 0,06 | 0,81 |
| 1742b | 13,2 | 8,6 | 7,3 | 0,9 | 0,65 | 0,55 | 0,07 | 0,85 |
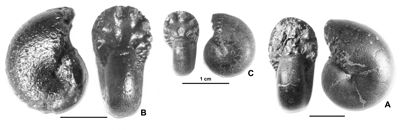
Cliquer sur la vignette pour agrandir l'image.
Figure 57 : Phyllopachyceras baborense (,
1880).
A : MD1742a U2i/GB : Aptien moyen (Zone à Martini) - Gaubert, Alpes-de-Haute-Provence.
B : MD1735 U2i/GB : Aptien moyen (Zone à Martini) - Ravin du Terrois, Drôme.
C : MD1742b U2i/GB : Aptien moyen (Zone à Martini) - Gaubert, Drôme.
Description : tous les spécimens vocontiens sont des moules internes pyritisés, lisses, épais, à flancs convexes. Le bord ventral est arrondi, la section du tour subcirculaire et rarement circulaire (un seul spécimen a ce type de section, c'est l'un des MD1736).
La ligne cloisonnaire (Fig. 58 ![]() ) est du type Phyllopachyceras : S1 et S2 tétraphylles, E aussi profond que L.
) est du type Phyllopachyceras : S1 et S2 tétraphylles, E aussi profond que L.
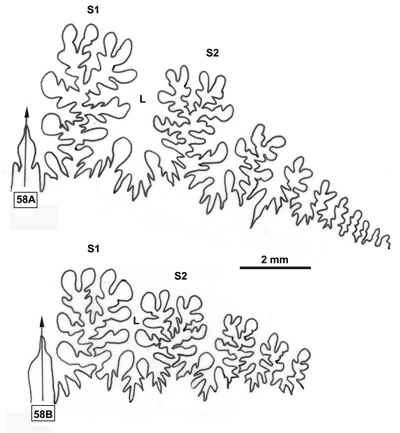
Cliquer sur la vignette pour agrandir l'image.
Figure 58 : Lignes cloisonnaires de Phyllopachyceras baborense.
A : MD1756a U3s/VL : Aptien moyen (Zone à Melchioris) Les Vallons, Alpes-de-Haute-Provence.
B : MD1794a U2i/GB : Aptien moyen (Zone à Martini) Gaubert, Alpes-de-Haute-Provence.
Affinités : il n'est pas nécessaire de revenir en détail sur cette espèce bien connue. Toutefois, il ne faut pas la confondre avec d'autres Phyllopachyceras comme P. brehereti n. sp. ou P. rouyanum. Les rapports avec ces espèces seront discutés plus loin. Notons seulement que, dans les Marnes Bleues, P. baborense est fréquemment récoltée en compagnie d'un autre Phyllopachyceras aux flancs aplatis et à section quadratique qui est décrit plus loin sous le nom de P. brehereti.
Âge : P. baborense est connue du Barrémien supérieur à l'Albien basal (, 2000a, p. 155). Les récoltes des Marnes Bleues vocontiennes s'échelonnent de l'Aptien inférieur (Zone à Deshayesi) au début de l'Albien inférieur (Zone à Tardefurcata).
Phyllopachyceras brehereti nov. sp.
| ? | 1977 | Partschiceras sp. juv. ? cf. baborense ().- & , p. 371, Pl. 6, figs. 2.a-b & 3.a-b. |
Holotype : collection MD1732a.
Localité : Gaubert, commune de Bayons, Alpes-de-Haute-Provence.
Âge : Aptien moyen (Zone à Melchioris).
Origine du nom : en hommage à Jean dont le travail fondamental sur les Marnes Bleues vocontiennes est à l'origine du renouveau de l'intérêt stratigraphique et paléontologique porté à cette formation.
Matériel étudié : 62 spécimens collection MD1731, 1732, 1737, 1740 (non mesuré), 1750 (non mesurés), 1754, 1755, 1759, 1763, 1767 (non mesuré), 1771, 1772, 1773 (non mesurés), 1782, 1792, 1799, 1800a, 1803, 2468, 2470, 2471, 2472 et 2473.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
|
1732a holotype |
13,9 | 8,1 | 6,6 | 1,1 | 0,58 | 0,47 | 0,08 | 0,81 |
| 1771b | 11,6 | 7,2 | 5,8 | 0,9 | 0,62 | 0,50 | 0,08 | 0,81 |
| 1803 | 14,7 | 8,7 | 6,8 | 1,4 | 0,59 | 0,46 | 0,09 | 0,78 |
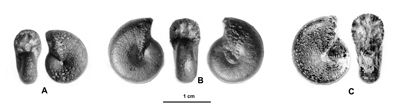
Cliquer sur la vignette pour agrandir l'image.
Figure 59 : Phyllopachyceras brehereti nov. sp.
A : MD1771b U3s/VL : Aptien moyen (Zone à Melchioris) - Les Vallons, Alpes-de-Haute-Provence.
B : MD1732a holotype - U3s/G1-NO : Aptien moyen (Zone à Melchioris) -
Gaubert, Alpes-de-Haute-Provence.
C : MD1803 U3s/VL : Aptien moyen (Zone à Melchioris) - Les Vallons, Alpes-de-Haute-Provence.
Diagnose : coquille épaisse, flancs plats, bord ventral aplati ou légèrement convexe, entonnoir ombilical évasé, section du tour quadratique, ligne cloisonnaire de type Phyllopachyceras à selles tétraphylles spatulées allongées.
Description : moules internes pyritisés, lisses, épais, à flancs plats et bord ventral aplati ou très légèrement convexes. L'ombilic est peu ouvert au fond d'une pente ombilicale en entonnoir évasé, la section du tour est quadratique. Chez un petit spécimen (MD1800 à D = 11,5 mm) la loge écrasée est partiellement conservée. L'espèce n'est donc pas le nucleus d'une espèce plus grande, il s'agit d'un taxon de petite taille, sans exclure la possibilité que ce spécimen soit un microconque.
La ligne cloisonnaire découpée a des selles S1 et S2 tétraphylles, E est aussi profond que L, c'est bien la ligne cloisonnaire d'un Phyllopachyceras (Fig. 60 ![]() ).
).
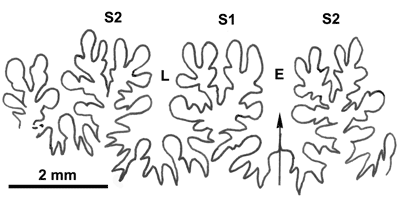
Cliquer sur la vignette pour agrandir l'image.
Figure 60 : Ligne cloisonnaire de Phyllopachyceras brehereti.
MD1799a U3s/G1-NO : Aptien moyen (Zone à Melchioris) Gaubert, Alpes-de-Haute-Provence.
Affinités : la forme de la section, les flancs aplatis permettent de distinguer la nouvelle espèce de tous les autres Phyllopachyceras. Ce caractère permet de l'identifier facilement parmi les P. baborense avec lesquels elle est souvent récoltée.
Les histogrammes des fréquences des paramètres dimensionnels (Fig. 61 ![]() ) montrent que, sur une population importante, la moyenne de l'épaisseur est moins grande chez P. brehereti que chez P. baborense. Les deux populations, P. baborense et P. brehereti, déjà bien reconnaissables par leurs caractères morphologiques, constituent des ensembles distincts sur les histogrammes des fréquences des paramètres relatifs.
) montrent que, sur une population importante, la moyenne de l'épaisseur est moins grande chez P. brehereti que chez P. baborense. Les deux populations, P. baborense et P. brehereti, déjà bien reconnaissables par leurs caractères morphologiques, constituent des ensembles distincts sur les histogrammes des fréquences des paramètres relatifs.
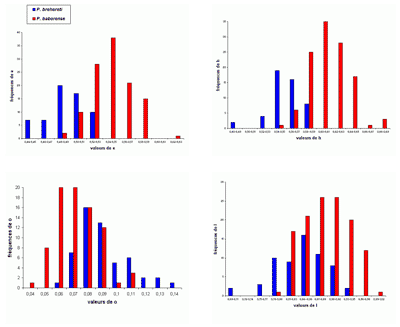
Cliquer sur la vignette pour agrandir l'image.
Figure 61 : Histogrammes des fréquences de e, h, o et l chez Phyllopachyceras baborense et Phyllopachyceras brehereti.
Âge : les récoltes vocontiennes sont datées du début de l'Aptien moyen (base de la Zone à Martini) au début de l'Aptien supérieur (Zone à Nolani).
Phyllopachyceras eichwaldi (, 1907)
| 1895 | Phylloceras eichwaldi , p. 13. | |
| 1907 | Phylloceras eichwaldi .- , p. 41, Pl. 2, fig. 10.a-b ; Pl. 3, figs. 4, 6 & 13 ; Pl. 24, figs. 3 & 5. | |
| Synonymie plus complète, lectotype, localité-type, âge du lectotype : , 2000a, p. 155. | ||
L'espèce P. eichwaldi est représentée dans les Marnes Bleues par sa sous-espèce P. eichwaldi occidentale.
Phyllopachyceras eichwaldi (, 1907) occidentale (, 1964)
| 1964 | Partschiceras eichwaldi () occidentale , p. 237, Pl. 16, fig. 5 ; figs. 55 & 56.a-c dans le texte. | |
| V | 1995 | Phyllopachyceras eichwaldi () occidentale ().- & , p. 130, Pl. 1, fig. 2. |
| 2000a | Phyllopachyceras eichwald (, 1907) occidentale (, 1964).- , p. 155, Pl. 37, fig. 12.a-b & 13, figs. 361-362. | |
| Synonymie plus complète, holotype, localité-type, âge de l'holotype de la sous-espèce : , 2000a, p. 155. | ||
Matériel étudié : 3 spécimens collection MD1725 et 1734.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1725a | 13,9 | 10,0 | 8,4 | - | 0,72 | 0,60 | - | 0,84 |
| 1734 | 14,5 | 9,2 | 8,2 | 1,2 | 0,63 | 0,57 | 0,08 | 0,89 |

Cliquer sur la vignette pour agrandir l'image.
Figure 62 : Phyllopachyceras eichwaldi occidentale (,
1964).
A : MD1734 U1s/TR : Aptien inférieur (Zone à Furcata) - Ravin du Terrois, Drôme.
B : MD1725a U1s/TR : Aptien inférieur (Zone à Furcata) - Ravin du Rivet, Alpes-Maritimes.
Description : Phyllopachyceras eichwaldi occidentale est représentée par trois moules internes lisses sur le bord ventral, portant une douzaine de replis à peine perceptibles sur les flancs, les flancs sont convexes. Le bord ventral est arrondi, la section du tour est subcirculaire, plus haute que large.
La ligne cloisonnaire montre des selles S1 et S2 tétraphylles caractéristiques du genre Phyllopachyceras.
La présence des replis sur les flancs et la valeur de e inférieure à 1 permettent de rattacher en toute certitude ces spécimens à la sous-espèce créée par (1964).
Âge : Phyllopachyceras eichwaldi occidentale était déjà connue dans le Barrémien du Sud-Est de la France ( & , 1995, p. 130 ; , 2000a, p. 156) et dans le Barrémien des Baléares (, 1964, p. 237). Dans les Marnes Bleues vocontiennes, ce taxon a été récolté uniquement dans des niveaux datés de la fin de l'Aptien inférieur (Zone à Furcata).
Phyllopachyceras picturatum (d', 1841)
| V | 1841 | Ammonites picturatus d', p. 178, Pl. 54, figs. 4-6. |
| V | 2000a | Phyllopachyceras picturatum (d').- , p. 156, Pl. 38, fig. 3.a-b, 4.a-b & 5.a-b ; figs. 367-370 dans le texte. |
| V | 2006 | Phyllopachyceras picturatum (d', 1841).- in , p. 66, Pl. 31, fig. 11.a-b ; fig. 42 dans le texte. |
| Synonymie plus complète, néotype, localité-type, âge du néotype : , 2000a, p. 156-157. | ||
Matériel étudié : 2 spécimens collection MD1785 et 1795.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1785 | 26,2 | 17,3 | 12,1 | - | 0,66 | 0,46 | - | 0,70 |
| 1795 | 10,0 | 6,6 | 5,0 | - | 0,66 | 0,50 | - | 0,76 |
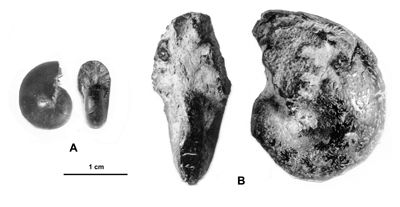
Cliquer sur la vignette pour agrandir l'image.
Figure 63 : Phyllopachyceras picturatum (d',
1841).
A : MD1795 U2i/GB : Aptien moyen (Zone à Martini) - Gaubert, Hautes-Alpes.
B : MD1785 spécimen écrasé U5i/CB : Albien inférieur (Zone à Tardefurcata) - Pillart, Alpes-de-Haute-Provence.
Description : les deux spécimens étudiés sont incomplets mais reconnaissables. Ce sont des moules internes lisses à section du tour ovale, le maximum d'épaisseur du tour étant proche de l'ombilic. Ce type de section est peu courant chez les Phyllopachyceras.
La ligne cloisonnaire est typiquement celle des Phyllopachyceras : S1 et S2 sont tétraphylles, E est aussi profond que L. Comme chez les spécimens récemment étudiés les phyllites terminaux des selles sont allongés (Fig. 64 ![]() ).
).
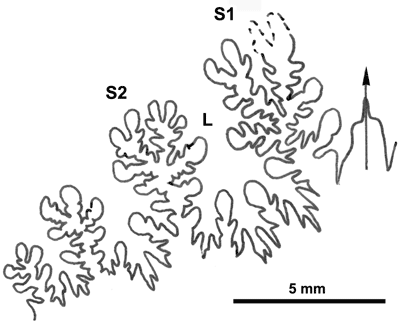
Cliquer sur la vignette pour agrandir l'image.
Figure 64 : Ligne cloisonnaire de Phyllopachyceras picturatum.
MD1785 U5i/CB : Albien inférieur (Zone à Tardefurcata) Pillart, Alpes-de-Haute-Provence.
La forme de la section du tour permet une identification facile de l'espèce, mais elle est plus rare que les autres Phyllopachyceras.
Âge : les deux spécimens proviennent de gisements différents : MD1795 est daté de l'Aptien moyen (Zone à Martini), MD1785 provient de la limite Aptien-Albien. C'est probablement le plus jeune P. picturatum connu. Les précédentes récoltes provenaient du "Gargasien" (, 2000a, p.157).
Phyllopachyceras rouyanum (d', 1841)
| V | 1841 | Ammonites rouyanus d', p. 362, Pl. 110, figs. 3-5. |
| 2000a | Phyllopachyceras rouyanum (d').- , p. 159. | |
| (pars) | 2000 | Phyllopachyceras baborense ().- et alii, p. 656, fig. 39.f-h, l-n, non fig. 39.j-k (= P. baborense ?) ; fig. 40.d-f, non fig. 40.a-c (= P. baborense). |
| V | 2006 | Phyllopachyceras rouyanum (d').- & in , p. 132, Pl. 17, figs. 3.a-c, 4.a-b, 5.a-b & 6.a-b ; fig. 62 dans le texte. |
| Synonymie plus complète, lectotype, localité-type, âge du lectotype : , 2000a, p. 159. | ||
Matériel étudié 131 spécimens collection MD1728, 1729, 1745, 1746, 1775, 1778, 1784, 1786, 1787, 1788, 1804, 1805, 1806 et 1807.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1728b | 13,9 | 8,7 | 9,4 | 0,9 | 0,63 | 0,68 | 0,06 | 1,08 |
| 1728c | 12,9 | 7,8 | 7,7 | 0,8 | 0,60 | 0,60 | 0,06 | 0,99 |
| 1729a | 24,6 | 16,0 | 14,2 | - | 0,65 | 0,58 | - | 0,89 |
| 23,5 | 14,5 | 13,7 | 1,2 | 0,62 | 0,58 | 0,05 | 0,94 | |
| 1729b | 15,4 | 9,7 | 9,2 | 0,8 | 0,63 | 0,60 | 0,05 | 0,95 |
| 1786a | 18,9 | 11,8 | 12,4 | - | 0,62 | 0,66 | - | 1,05 |
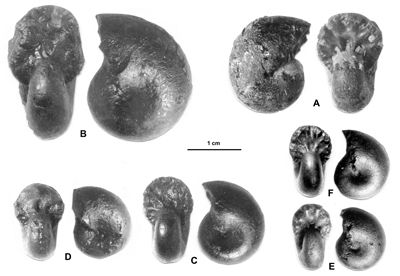
Cliquer sur la vignette pour agrandir l'image.
Figure 65 : Phyllopachyceras rouyanum (d',
1841).
A : MD1786a U5i/CB : Albien inférieur (Zone à Tardefurcata) - Pillart, Alpes-de-Haute-Provence.
B : MD1729a U5i/CB : Albien inférieur (Zone à Tardefurcata) - Chênebuis, Alpes-de-Haute-Provence.
C : MD1729b - U5i/CB : Albien inférieur (Zone à Tardefurcata) - Chênebuis,
Alpes-de-Haute-Provence.
D : MD1728b - U5i/CB : Albien inférieur (Zone à Tardefurcata) - Chênebuis,
Alpes-de-Haute-Provence.
E : MD1728c - U5i/CB : Albien inférieur (Zone à Tardefurcata) - Chênebuis,
Alpes-de-Haute-Provence.
F : MD1728a - U5i/CB : Albien inférieur (Zone à Tardefurcata) - Chênebuis, Alpes-de-Haute-Provence.
Description : les spécimens étudiés ici sont tous des moules internes pyritisés, lisses, très épais (la valeur de e est proche, égale ou supérieure à 1). L'ombilic est très petit, les flancs sont très convexes, le bord ventral est arrondi, la section du tour est circulaire.
La ligne cloisonnaire (Fig. 66 ![]() ) a des selles S1 et S2 tétraphylles, E aussi profond que L, elle est très semblable à la fig. 5 de d'
(1841).
) a des selles S1 et S2 tétraphylles, E aussi profond que L, elle est très semblable à la fig. 5 de d'
(1841).
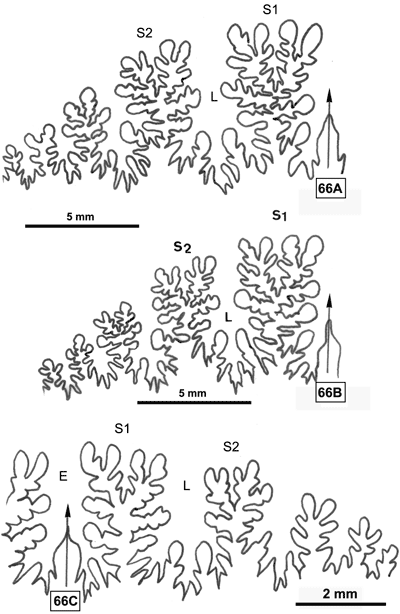
Cliquer sur la vignette pour agrandir l'image.
Figure 66 : Lignes cloisonnaires de Phyllopachyceras rouyanum.
A-B : MD1786a U5i/CB : Albien inférieur (Zone à Tardefurcata) Pillart, Alpes-de-Haute-Provence.
C : MD1784a U5i/?CB : Albien inférieur (Zone à Tardefurcata) Pillart, Alpes-de-Haute-Provence.
Affinités : les spécimens vocontiens sont, sans aucun doute, à rattacher à l'espèce créée par d'. En fait, contrairement à ce que l'on croit parfois, ce taxon ne se confond pas avec P. baborense, comme le prouvent la comparaison des histogrammes des fréquences des paramètres relatifs des deux taxons (Fig. 67 ![]() ). Pour tous les paramètres, les deux populations sont parfaitement distinctes mais c'est surtout par son épaisseur plus grande que rouyanum se distingue de baborense, les histogrammes de e et l ont nettement une allure bimodale.
). Pour tous les paramètres, les deux populations sont parfaitement distinctes mais c'est surtout par son épaisseur plus grande que rouyanum se distingue de baborense, les histogrammes de e et l ont nettement une allure bimodale.
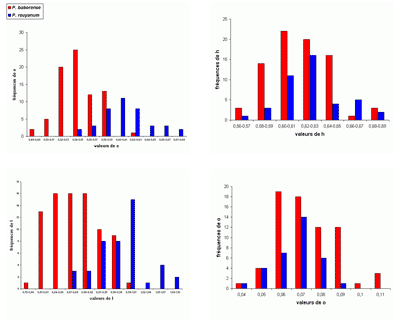
Cliquer sur la vignette pour agrandir l'image.
Figure 67 : Histogrammes des fréquences de e, h, o et l chez Phyllopachyceras baborense et Phyllopachyceras rouyanum.
Pourquoi l'espèce P. rouyanum est-elle si difficile à identifier dans les récoltes ? En partie, parce que sa morphologie est proche de P. baborense ; d'autre part, parce que dans une population de P. baborense on trouve toujours des individus à section circulaire (l = 1,00), mais jamais de spécimens chez lesquels l a une valeur sensiblement supérieure à 1,00. On est alors tenté de déterminer comme P. rouyanum ces P. baborense. Il est évidemment possible aussi de considérer les P. baborense les plus épais comme les premiers représentants de l'espèce P. rouyanum.
Cet exemple montre la nécessité de dépasser, pour le paléontologue, la conception purement typologique des espèces pour s'intéresser à l'étude des populations.
Âge : si la détermination des spécimens ne laisse aucun doute, en revanche leur niveau stratigraphique élevé pose problème. En effet, les gisements sont datés du sommet de l'Aptien supérieur (Zone à Jacobi) au début de l'Albien inférieur (base de la Zone à Tardefurcata).
Or, & , dans le cadre de la Révision critique de la Paléontologie française (in , 2006, p. 132), ont retrouvé le lectotype de l'espèce de d' qui proviendrait bien de l'Hauterivien supérieur.
Il est probable qu'un certain nombre de rouyanum n'ont pas été reconnus en tant que tels, et, qu'à l'inverse, certains spécimens déterminés "rouyanum" sont en réalité des P. baborense. Les spécimens des Marnes Bleues vocontiennes correspondent parfaitement à la définition que d' a donné de l'espèce rouyanum en y ajoutant cependant la notion de variabilité qui ne peut s'appréhender que par l'étude des populations. Il est essentiel pour comprendre les taxons chez les Phyllopachyceras de tenir compte de la grande variabilité de leur épaisseur.
Famille des Neophylloceratidae , 1993
Sous-famille des Hyporbulitinae , 1993
Genre Hyporbulites , 1947
Hyporbulites seresitensis (, 1907)
| 1907 | Phylloceras velledae () var. seresitensis , p. 52. | |
| 1910 | Phylloceras velledae () var. seresitensis .- , p. 9, Pl. 1, fig. 1.a-b, 2.a-b & 3.a-c ; fig. 2 dans le texte. | |
| 1989 | Phylloceras (Hypophylloceras) seresitense .- , p. 114, Pl. 1, fig. 2. | |
| 1989 | Phylloceras (Hypophylloceras) tanit .- , p. 114, Pl. 1, fig. 13. | |
| 2000a | Hyporbulites seresitensis ().- , p. 164, Pl. 39, fig. 13.a-b, 14 & 15.a-b, figs. 389-394 dans le texte. | |
| 2006 | Neophylloceras seresitensis (). & , p. 41, Pl. 5, figs. 5-7 & 10-14. | |
| Synonymie plus complète, lectotype, localité-type, âge du lectotype : , 2000a, p. 164. | ||
Matériel étudié : 71 spécimens collection MD1454, 1455, 1456, 1457, 1458, 1459, 1460, 1461, 1462 et 1463.
Dimensions des spécimens figurés
| N° MD | D | H | E | O | h | e | o | l |
| 1454a | 29,8 | 17,7 | 10,5 | 1,8 | 0,59 | 0,35 | 0,06 | 0,59 |
| 1455a | 31,0 | 18,9 | 11,2 | 2,0 | 0,61 | 0,36 | 0,06 | 0,59 |

Cliquer sur la vignette pour agrandir l'image.
Figure 68 : Hyporbulites seresitensis (,
1907).
A : MD1455a U7m/CT : Albien supérieur (Zone à Blancheti) - Ravine de Bert, Drôme.
B : MD1454a U7m/CT : Albien supérieur (Zone à Blancheti) - La Folie, Hautes-Alpes.
Description : moules internes pyritisés, lisses, comprimés, ombilic fermé, flancs presque plats, bord externe arrondi, section du tour ogivale haute.
La ligne cloisonnaire est découpée, avec S1 et S2 tétraphylles mais les terminaisons des selles tendent à perdre l'aspect spatulé qui jusque là avait caractérisé le genre Phylloceras. Ce sont les caractères du genre Hyporbulites appartenant à la sous-famille des Hyporbulitinés (Fig. 69 ![]() ).
).
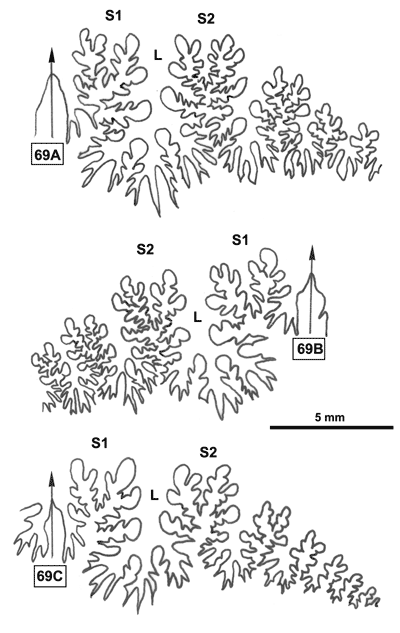
Cliquer sur la vignette pour agrandir l'image.
Figure 69 : Lignes cloisonnaires de Hyporbulites seresitensis.
A : MD1455a U7m/CT : Albien supérieur (Zone à Blancheti) Ravine de Bert, Drôme.
B-C : MD1454a U7m/CT : Albien supérieur (Zone à Blancheti) La Folie, Hautes-Alpes.
Les histogrammes des fréquences des paramètres relatifs sont unimodaux (Fig. 70 ![]() ) ce qui permet d'affirmer que l'on est bien en présence d'une seule espèce malgré la variabilité de l'épaisseur.
) ce qui permet d'affirmer que l'on est bien en présence d'une seule espèce malgré la variabilité de l'épaisseur.
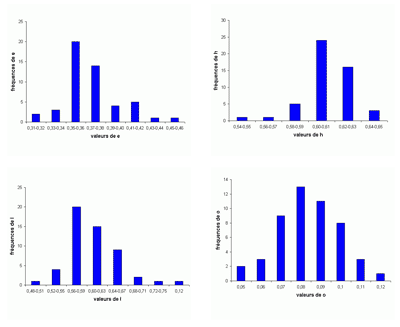
Cliquer sur la vignette pour agrandir l'image.
Figure 70 : Histogrammes des fréquences de e, h, o et l chez Hyporbulites seresitensis.
Âge : Hyporbulites seresitensis était déjà connue en France dans l'Albien et au moins jusqu'au Cénomanien moyen (, 2000a, p. 165). Les nouvelles récoltes sont datées de l'Albien supérieur (Zone à Blancheti) et du Cénomanien inférieur (Zone à Mantelli).
Deux espèces problématiques non présentes dans les récoltes récentes du bassin vocontien : Phylloceras (Hypophylloceras) morelianum (d', 1841) et Salfeldiella belus (d', 1841).
Il est étonnant que ces deux espèces introduites par d' en 1841 n'aient pas été retrouvées dans les faunes récemment récoltées dans le bassin vocontien.
La "Révision critique" de la Paléontologie française de d' (, 2006, p. 65) a montré que P. (H.) morelianum a été créée sans doute à partir d'un matériel hétérogène dont malheureusement les spécimens les plus intéressants n'ont pu être retrouvés dans la collection d' du Muséum national d'Histoire naturelle de Paris. En outre, la fig. 1 de d' (1841, Pl. 54) est erronée car elle exagère manifestement l'ouverture de l'ombilic. Avant d'être égaré, le lectotype de l'espèce a été figuré par (1964, Pl. 13, fig. 2.a-b) et son ombilic est nettement plus fermé que celui de la fig. 1 de d'. Le lectotype de l'espèce étant perdu, un néotype a été désigné ( in , 2006, p. 65). Ce néotype provient de l'Aptien de Gargas (Vaucluse) donc d'une localité proche de celle du lectotype (Carpentras, Vaucluse). Le néotype est conservé au Musée d'Avignon (collection n° MR 2001-015).
Les spécimens de l'espèce morelianum sont rares. L'espèce a été certainement souvent confondue avec P. (H.) aptiense , 1920 beaucoup plus fréquente. Outre, ceux de Carpentras et de Gargas des collections anciennes, (2005, p. 49, Pl. 3, figs. 5-6) a figuré deux spécimens de P. (H.) morelianum provenant de La Tuilière près d'Apt (zones à Deshayesites et Furcata) ainsi qu'un spécimen plus jeune (Zone à Martini) récolté à Teyssières au nord de Nyons (Drôme). En outre, (1920, p. 192) a décrit un spécimen déterminé P. aff. Moreli provenant de Saint André Les Alpes (Alpes de Haute-Provence). La ligne cloisonnaire de ce spécimen étant assez différente de celle du néotype, (2000a, p. 134) a rattaché avec doute ce spécimen à l'espèce morelianum. À ce jour, P. (H.) morelianum semble donc être une espèce restreinte à la plate-forme nord-provençale et sur les bordures du bassin vocontien, distribution un peu paradoxale pour un représentant des Phylloceratoidea!
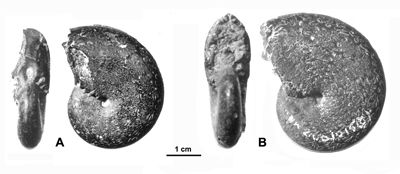
Cliquer sur la vignette pour agrandir l'image.
Figure 71 : Phylloceras (Hypophyloceras) morelianum (d',
1841).
A : néotype, collection , Musée , Avignon, n° MR-2001-015(1),
Aptien (?Zone à Furcata) - Gargas, Vaucluse.
B : para-néotype, collection , Musée , Avignon, n° MR-2001-015(4),
Aptien (?Zone à Furcata) - Gargas, Vaucluse.
Quant à Salfeldiella belus, espèce jusqu'alors difficile à identifier et très rarement
citée, la "Révision critique" a mis en évidence une erreur originelle sur l'un des dessins de d'. Les figures 4 et 5
(1841, Pl. 52) sont bien celles d'un Salfeldiella mais le tracé de la ligne cloisonnaire est celui d'un Desmoceratidae (Fig. 6 ![]() ). Dans ces conditions, la seule connaissance des figures de d' rendait quasiment impossible la mise en évidence de
l'espèce. Dans la collection d' du Muséum national d'Histoire naturelle de Paris, le lectotype et le paralectotype de l'espèce sont de petits spécimens
(respectivement D = 14,8 mm et 11,2 mm). Ils proviennent de "l'Aptien" de la région de Gap
(Hautes-Alpes). S. belus a des affinités avec S. lateumbilicatum (,
1907) et peut-être avec les tours les plus internes de P. (H.) paquieri ,
1920 ( in ,
2006, p. 59).
). Dans ces conditions, la seule connaissance des figures de d' rendait quasiment impossible la mise en évidence de
l'espèce. Dans la collection d' du Muséum national d'Histoire naturelle de Paris, le lectotype et le paralectotype de l'espèce sont de petits spécimens
(respectivement D = 14,8 mm et 11,2 mm). Ils proviennent de "l'Aptien" de la région de Gap
(Hautes-Alpes). S. belus a des affinités avec S. lateumbilicatum (,
1907) et peut-être avec les tours les plus internes de P. (H.) paquieri ,
1920 ( in ,
2006, p. 59).
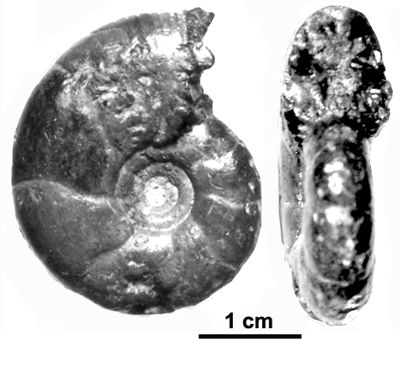
Cliquer sur la vignette pour agrandir l'image.
Figure 72 : Salfeldiella belus (d',
1841).
lectotype, Collection d' (n° 5579 bis-1), MNHN de Paris n° LPMP-B46094 -
"Aptien" des environs de Gap, Hautes-Alpes.
Grâce à un abondant matériel repéré stratigraphiquement à l'échelle de la zone
d'ammonite, nous pouvons dresser un tableau satisfaisant de la distribution lithostratigraphique et biostratigraphique des Phylloceratoidea connus à ce jour dans le bassin vocontien entre la fin de l'Aptien inférieur et la fin de l'Albien (Figs. 73 ![]() - 74
- 74 ![]() ).
).
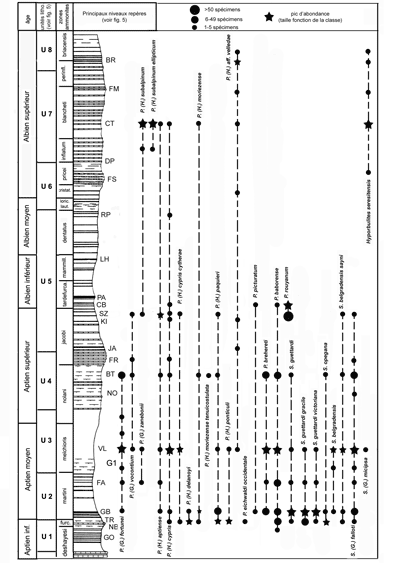
Cliquer sur la vignette pour agrandir l'image.
Figure 73 : Répartition lithostratigraphique semiquantitative des Phylloceratoidea des Marnes Bleues du bassin vocontien (à l'exception de P. morelianum et S. belus non récoltées dans ce travail).
Avec une trentaine de taxons (dont 4 identifiés comme nouveaux), les Marnes Bleues du bassin vocontien sont remarquables par leur richesse et leur variété en Phylloceratoidea. Cette composition présente cependant d'importantes fluctuations dans le temps tant sur le plan qualitatif (diversité taxonomique) que sur le plan quantitatif (proportion relative d'individus dans l'ammonitifaune).
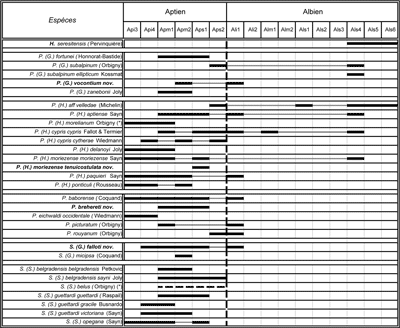
Cliquer sur la vignette pour agrandir l'image.
Figure 74 : Répartition biostratigraphique des 30 espèces de Phylloceratoidea connues dans les Marnes Bleues du bassin vocontien. (*) : espèces non récoltées dans ce travail.
Lorsqu'on analyse la distribution stratigraphique des Phylloceratoidea dans les Marnes Bleues, on remarque trois pics d'abondance au sein desquels ils constituent jusqu'à plus des deux tiers de l'ammonitifaune :
Notons que les fluctuations des populations de Tetragonitoidea suivent assez fidèlement celles des Phylloceratoidea. Si l'on replace l'évolution de ces deux groupes dans l'évolution des séquences de dépôt (voir Fig. 9 ![]() ), on note une bonne corrélation avec les variations du niveau marin puisque les pics 1 à 3 se situent à proximité des surfaces d'inondation maximale. Cependant, les épisodes de haut niveau marin ne favorisent pas à eux seuls le développement des formes du biome pélagique téthysien. Ainsi, le niveau (flèche noire sur Fig. 77
), on note une bonne corrélation avec les variations du niveau marin puisque les pics 1 à 3 se situent à proximité des surfaces d'inondation maximale. Cependant, les épisodes de haut niveau marin ne favorisent pas à eux seuls le développement des formes du biome pélagique téthysien. Ainsi, le niveau (flèche noire sur Fig. 77 ![]() ), en position de haut niveau, est remarquable par sa très grande pauvreté en Phylloceratoidea et en Lytoceratoidea (les Tetragonitoidea semblent même totalement absents) tandis que pullulent Leymeriellidae (Fig. 75
), en position de haut niveau, est remarquable par sa très grande pauvreté en Phylloceratoidea et en Lytoceratoidea (les Tetragonitoidea semblent même totalement absents) tandis que pullulent Leymeriellidae (Fig. 75 ![]() ), Brancoceratidae, Desmoceratidae et Ancyloceratidae.
), Brancoceratidae, Desmoceratidae et Ancyloceratidae.

Cliquer sur la vignette pour agrandir l'image.
Figure 75 : Biofaciès du Niveau à Leymeriella. MD1156 - base U5 : Albien inférieur (Zone à Tardefurcata) - Pré-Guittard, Drôme.
La meilleure interprétation de cet épisode défavorable aux représentants du biome pélagique téthysien et, par ailleurs, riche en matière
organique semble être celui d'un déficit en oxygène des eaux profondes ( et alii, 1986). On note également une baisse sensible de la diversité des Phylloceratoidea dans un autre épisode de couches riches en matière organique, les niveaux (FA sur Fig. 77 ![]() ).
).
Les études récentes qui montrent une absence d'augmentation notable de la productivité des eaux de surface ( et alii, 2003) confirment cette interprétation et le fait que le mode de vie des Phylloceratoidea, Lytoceratoidea et Tetragonitoidea dépendent des eaux profondes. Ces observations rejoignent les interprétations des Phylloceratoidea comme des migrateurs planctoniques verticaux ayant vécu à assez grande profondeur ( & , 2006, p. 58) comparables aux modes de vie suggérés pour certaines ammonites du Trias ( & , 2000, p. 112).
Par contre, le rôle de la température est plus ambigu car l'épisode est considéré comme un épisode particulièrement chaud par et alii (2003) alors que l'on note par ailleurs un afflux d'ammonites nord-tempérés (Leymeriellidae). Mais il est possible que le gradient latitudinal de la température des eaux ait été moins important que le gradient vertical. En ce sens, les formes d'eau profonde pourraient avoir été sténothermes.
D'autres facteurs que le déficit en oxygène des couches profondes du bassin vocontien doivent être recherchés pour expliquer la quasi absence en Phylloceratoidea constatée dans la partie supérieure de l'unité U5 et dans l'unité U6, soit durant l'intervalle allant du sommet de l'Albien inférieur à la base de l'Albien supérieur. Dans le premier cas (unité U5s), peu d'ammonites sont fossilisées mais cela ne signifie pas obligatoirement qu'elles aient été rares pour autant (Fig. 7.L-M ![]() ). Il semblerait plutôt qu'il s'agisse d'une mauvaise fossilisation affectant l'ensemble des ammonites et plus particulièrement les formes à test mince. Un fait un peu semblable a été signalé chez les Juraphyllitidae et les Phylloceratidae du Carixien et du Domérien de l'Auxois (,
1977, p. 43).
). Il semblerait plutôt qu'il s'agisse d'une mauvaise fossilisation affectant l'ensemble des ammonites et plus particulièrement les formes à test mince. Un fait un peu semblable a été signalé chez les Juraphyllitidae et les Phylloceratidae du Carixien et du Domérien de l'Auxois (,
1977, p. 43).
Le cas de l'unité U6 est tout autre car elle renferme souvent une profusion d'ammonites appartenant au genre Puzosia et à la famille des Brancoceratidae mais aucun Phylloceratoidea. Cette composition particulière de l'ammonitifaune jointe à la présence de nombreux bivalves (Birostrina et Aucellina) formant des horizons lumachelliques (Fig. 76 ![]() ) coincide avec une importante augmentation du détritisme terrigène.
) coincide avec une importante augmentation du détritisme terrigène.

Cliquer sur la vignette pour agrandir l'image.
Figure 76 : Biofaciès du Faisceau silteux à Puzosia et Birostrina sulcata. MD1982a - U6s : Albien supérieur (Zone à Pricei) - Col de Palluel, Hautes-Alpes.
Cet ensemble de faits suggère une notable diminution de la profondeur du Bassin vocontien vers le milieu de l'Albien supérieur. Cet épisode culmine au sommet de l'unité U6 dans l'horizon à Pinna (base de la Zone à Inflatum : Fig. 5 ![]() ) où l'étude des bivalves réalisée par
(1995, p. 463) a conduit cet auteur à envisager une paléoprofondeur inférieure à 200 m. Ceci confirme que les Phylloceratoidea et les Tetragonitoidea sont des indicateurs de vie relativement profonde à bathymétrie sensiblement supérieure à 200 m.
) où l'étude des bivalves réalisée par
(1995, p. 463) a conduit cet auteur à envisager une paléoprofondeur inférieure à 200 m. Ceci confirme que les Phylloceratoidea et les Tetragonitoidea sont des indicateurs de vie relativement profonde à bathymétrie sensiblement supérieure à 200 m.
Sur le plan de la diversité taxonomique, les 3 pics à Phylloceratoidea ne sont pas comparables ainsi que le montre le graphique de la figure 77. La période la plus propice aux Phylloceratoidea se situe clairement au début de l'Aptien moyen où ils constituent à la fois une forte proportion de l'ammonitifaune (45%) et présentent une grande diversité spécifique (13 taxons). La situation est bien différente pour les pics 2 et 3 qui présentent certes une forte proportion de Phylloceratoidea (respectivement 55% et 38%) mais une faible diversité spécifique (respectivement 4 et 6 taxons).
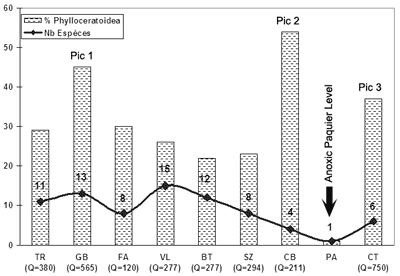
Cliquer sur la vignette pour agrandir l'image.
Figure 77 : Évolution stratigraphique des l'importance quantitative et de la diversité spécifique des Phylloceratoidea des Marnes Bleues du bassin
vocontien
(abréviations des horizons fossilifères en Fig. 5 ![]() ).
).
Après un dernier épisode situé au début du Cénomanien (, 1987a) où les Phylloceratoidea représentent encore une part notable de l'ammonitifaune vocontienne, ce groupe montre un fort appauvrissement qui est à corréler à un remblaiement du bassin d'ouest en est. Cette évolution va réduire la profondeur en cantonnant les formes d'eau profonde dans une étroite aire orientale (aire à Leiostraca de , 1965, 1980). L'évolution des Phylloceratoidea au début du Crétacé supérieur en domaine vocontien traduit ainsi le changement de la subsidence qui conduira à faire du bassin une zone positive dès la fin du Crétacé.
La présente étude a apporté d'importantes précisions sur l'extension stratigraphique des Phylloceratoidea téthysiens. Elle confirme que la période cruciale pour l'évolution de la famille des Phylloceratidae a bien été l'Aptien-Albien. Au sein de la Téthys occidentale, le Bassin vocontien semble ainsi avoir été l'un des derniers lieux de peuplement important et d'évolution des représentants de cette famille remplacés progressivement par les Neophylloceratidae.
Si les faunes aptiennes sont encore très riches en Phylloceratidae à constrictions de l'Aptien à l'Albien, le nombre de taxons se réduit considérablement au début de l'Albien, de moitié environ, le nombre de genres et de sous-genres restant identique dans les deux étages (Fig. 75 ![]() ). Cette évolution est l'amorce du déclin des Phylloceratidae, les formes à constrictions comme les Salfeldiella et les Goretophylloceras ne subsistant dans l'Albien que par de rares formes. Les Salfeldiella sensu stricto semblent même avoir disparu du Bassin vocontien alors que, curieusement, ils sont encore connus dans l'Albien de l'Aube tout en étant cependant très rares (,
1998). Les Phylloceratidae, quoique en décroissance, sont cependant encore abondants dans les Marnes Bleues où ils peuvent constituer une part essentielle de l'ammonitifaune de certains horizons pyriteux de la fin de l'Albien supérieur (horizon du Châtelat : Fig. 9
). Cette évolution est l'amorce du déclin des Phylloceratidae, les formes à constrictions comme les Salfeldiella et les Goretophylloceras ne subsistant dans l'Albien que par de rares formes. Les Salfeldiella sensu stricto semblent même avoir disparu du Bassin vocontien alors que, curieusement, ils sont encore connus dans l'Albien de l'Aube tout en étant cependant très rares (,
1998). Les Phylloceratidae, quoique en décroissance, sont cependant encore abondants dans les Marnes Bleues où ils peuvent constituer une part essentielle de l'ammonitifaune de certains horizons pyriteux de la fin de l'Albien supérieur (horizon du Châtelat : Fig. 9 ![]() ).
).
À la fin de l'Albien supérieur prédominent les populations de Hyporbulites seresitensis, sans cependant atteindre la diversité spécifique connue à Madagascar (, 1993), ce genre étant le premier représentant de la famille des Neophylloceratidae apparu à lHauterivien (, 2000a, p. 53). Cette dernière famille atteindra le Crétacé terminal en plein développement. LAlbien voit donc samorcer le déclin des Phylloceratidae apparus au Lias, seuls les Phyllopachyceras atteindront le sommet du Maastrichtien en compagnie des derniers Neophylloceratidae.
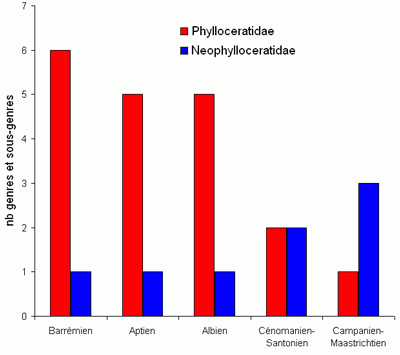
Cliquer sur la vignette pour agrandir l'image.
Figure 78 : Remplacement progressif des Phylloceratidae par les Neophylloceratidae du Barrémien au Maastrichtien.
Le manuscrit a bénéficié des remarques des deux rapporteurs, François (Université de Lyon) et Luc Georges (Université de Marseille). Quant à l'extended English abstract il a été révisé par Nestor Les auteurs les remercient tous trois ici.
F. (2002). Plaidoyer pour un étage Vraconnien entre l'Albien sensu stricto et le Cénomanien (système Crétacé).- Académie royale de Belgique, Publication de la Classe des Sciences, Liège, Collection in-4°, (3e série), t. IV, 128 p.
F. (2008).- Support for a Vraconnian Stage between the Albian sensu stricto and the Cenomanian (Cretaceous System).- Carnets de Géologie / Notebooks on Geology, Brest, Mémoire 2008/02 (CG2008_M02), 83 p.
D.J. (1899).- Über die Kreideformation des Kaukasus.- Beiträge zur Paläontologie und Geologie Österreich- Ungars und des Orients, Wien, vol. 12, n° 13, p. 15-159.
F. & Y. (2002). Nouvelles données biostratigraphiques sur l'Aptien moyen et supérieur du Sud-Est de la France, à la lumière de la succession des ammonites du Domaine vocontien.- Documents des Laboratoires de Géologie de la Faculté des Sciences de Lyon, n° 156, p. 23-24.
F. & Y. (2005). Biostratigraphie et faunes d'ammonites de l'Aptien supérieur (Gargasien) de la région stratotypique de Gargas, près d'Apt (Sud-Est de la France).- Réunions thématiques du Groupe français du Crétacé, Paris 5-6 décembre 2005, "Impact environnemental sur la dynamique des populations au Crétacé", volume des résumés, p. 5-8.
A.F. & P. (1977). Un bassin mésogéen du domaine péri-alpin : le Sud-Est de la France.- Bulletin des Centres de Recherche et Exploration-Production Elf-Aquitaine, Pau, t. 1, n° 1, p. 261-308.
J.G. (1983). Sur des niveaux de black shales dans l'Albien inférieur et moyen du domaine vocontien (sud-est de la France) : étude de nannofaciès et signification des paléoenvironnements.- Bulletin du Muséum National d'Histoire Naturelle, Paris, (4e série), 5, section C, n° 1, p. 113-159.
J.G. (1985). Indice d'un évènement anoxique étendu à la Téthys alpine, à l'Albien inférieur (évènement ).- Comptes Rendus de l'Académie des Sciences, Paris, t. 300, (série II), n° 8, p. 355-358.
J.G. (1988). Épisodes de sédimentation riche en matière organique dans les Marnes Bleues d'âge aptien et albien de la partie pélagique du bassin vocontien.- Bulletin de la Société Géologique de France, Paris, (série 8), t. IV, n° 2, p. 349-356.
J.G. (1995). L'Aptien et l'Albien de la Fosse vocontienne (des bordures au bassin). Évolution de la sédimentation et enseignements sur les événements anoxiques.- Thèse Doctorat de l'Université François Rabelais, Tours, 564 p. et Publication de la Société Géologique du Nord, Lille, t. 25 (1997), 614 p.
J.G., M. & M. (1986). Niveaux riches en matière organique dans l'Albien vocontien, quelques caractères du paléoenvironnement, essai d'interprétation génétique.- Documents du Bureau de Recherche Géologique et Minière, Orléans, n° 110, p. 141-191.
M. (1937). Sur les niveaux fossilifères de l'Albien dans la fosse vocontienne (Drôme, Hautes-Alpes et Basses-Alpes).).- Comptes Rendus de l'Académie des Sciences, Paris, t. 204, p. 1492-1493.
M. (1939). Sur le Cénomanien inférieur de la fosse vocontienne (Hautes-Alpes et Drôme).- Comptes Rendus de l'Académie des Sciences, Paris, t. 208, p. 1514-1516.
M. (1947).- Sur les zones d'ammonites dans l'Albien de France et d'Angleterre.- Travaux du Laboratoire de Géologie de la Faculté des Sciences de Grenoble, t. 26, p. 17-104.
L.G., B. & L. (2005). Modification de la structure des populations d'ammonites dans l'Aptien inférieur sommital et l'Aptien supérieur du domaine vocontien (SE France).- Réunions thématiques du Groupe français du Crétacé, Paris 5-6 décembre 2005, "Impact environnemental sur la dynamique des populations au Crétacé", volume des résumés, p. 15-17.
R. & L. (1957).- Contribution à l'étude des faunes d'ammonoïdes de Medjez-Sfa, Étude paléontologique (par ).- Publication du Service de la carte géologique de l'Algérie, Alger, n° 13 (nouvelle série), p. 67-123.
M. (1937).- Paléontologie de Madagascar, XXII - Les Ammonites pyriteuses de l'Aptien d'Antanatanamirafy.- Annales de Paléontologie, Masson éditeur, Paris, t. 26, p. 3-28.
M. (1949).- Faunes néocomiennes des couches à Criocères de Belohasifaka (Cercle de Sitampiky) (Madagascar).- Annales géologiques du Service des Mines, Gouvernement général de Madagascar et Dépendances, Paris, fasc. 15, p. 53-85, 6 Pls.
H. (1880).- Études supplémentaires sur la paléontologie algérienne.- Bulletin de l'Académie d'Hippone, Bône, n° 15, p. 1-449.
H. (1886).- Fossiles décrits par . in: , Bulletin de l'Académie d'Hippone, Bône, n° 28, 1 Pl.
L. (2002).- Litho-, bio-, et chronostratigraphie comparées dans le bassin Vocontien à l'Aptien.- Thèse de l'Université, Université des Sciences et Technologies de Lille-I, 451 p.
G. & B. (1995).- Sur quelques Phylloceratidae (Phylloceratina, Ammonoidea) du Barrémien supérieur et de l'Aptien inférieur du Sud-Est de la France.- Mésogée, Bulletin du Muséum d'Histoire Naturelle de Marseille, vol. 54, p. 121-138.
L. & G. (1986). Tentative d'élaboration d'une chronologie hémérale de l'Aptien moyen (Système Crétacé) fondée sur les Ammonites.- Comptes Rendus de l'Académie des Sciences, Paris, t. 302, (série II), p. 29-34.
A. de (1840-42).- Voyage dans la Russie méridionale et la Crimée. Description des principaux fossiles de la Crimée par L. . Céphalopodes du Néocomien, du grès vert.- Bourdin édit., Paris, vol. 2 : p. 781-819 et p. 821-823 (explications des planches).
V.V. (1953).- Dannye ob ontogeneze novogo roda ammonitov Euphylloceras [Data on the ontogeny of the new ammonite genus Euphylloceras].- Vestnik Moskovskogo Universitera, n° 9, p. 141-147.
Y. (2005).- Biostratigraphie, évolution et renouvellement des ammonites de l'Aptien supérieur (Gargasien) du bassin vocontien (SE de la France).- Thèse, Université de Lyon 1, 280 p.
N. & S.J. (1972). Punctated equilibria : an alternative to phyletic gradualism. In : T. J. M. (ed.), Model in Paleobiology.- Freeman Cooper Publisher, San Francisco, p. 82-115.
V.L. (1969). [Ammonites of the Clansayes horizon from Northern Caucasus].- [Proceedings of the Krasnodar Filial of the All-Union Petroleum-Gas Research Institut], Moscou, vol. 19, p. 126-189 [en russe].
E. (1885). Étude géologique sur les étages moyens et supérieurs du terrain crétacé dans le Sud-Est de la France.- Annales des Sciences Géologiques, Paris, vol. 18, 268 p.
P. (1920a). Observations sur diverses espèces du Gargasien bathyal alpin et en particulier sur la faune de Blieux.- Mémoires pour servir à l'Explication de la Carte géologique détaillée de la France, Paris, p. 229-266.
P. (1920b).- La faune des marnes aptiennes et albiennes de la région d'Andraitx (Majorque).- Trabajos del Museo Nacional de Ciencias Naturales, serie Geológica, Madrid, n° 26, p. 5-68.
P. & H. (1923).- Ammonites nouvelles des îles Baléares.- Trabajos del Museo Nacional de Ciencias Naturales, serie Geológica, Madrid, n° 32, p. 5-83.
J.-C. (2006). Révision Critique de la Paléontologie Française d'Alcide d'. Volume IV Céphalopodes crétacés.- Backhuys Publishers, Leiden, 292 p.
J. (1963). Remarques stratigraphiques, paléontologiques et structurales sur la région de Séderon.- Bulletin du Service de la Carte géologique de France, Paris, n° 272, p. 815-845.
K.B. (1989). Beschreibung neugefundener Ammonoidea aus der Vorarlberger Garschella-Formation (Aptian-Albian).- Jahrbuch der Geologischen Bundesanstalt, Wien, vol. 132, n° 1, p. 105-189.
G. (1987). Dynamique du bassin subalpin méridional de l'Aptien au Cénomanien.- École des Mines de Paris, Mémoire des Sciences de la Terre, vol. 4, 370 p.
G. & O. (2003). Anatomy of ancient passive margin slope systems : Aptian gravity-driven deposition on the Vocontian paleomargin, western Alps, south-east France.- Sedimentology, Oxford, vol. 50, p. 1231-1270.
J. & P. (1954). Nouvelle contribution à la connaissance du Crétacé moyen dans la Fosse vocontienne.- Comptes Rendus de l'Académie des Sciences, Paris, t. 238, p. 1048-1050.
A.S., W.J., J.A., M. & B.E. (1996). The Late Albian to Early Cenomanian succession at the Mont Risou, near Rosans (Drôme, SE France): an integrated study (ammonites, inoceramids, planktonic foraminifera, nannofossils, oxygen and carbon isotopes).- Cretaceous Research, London, vol. 17, p. 515-606.
G.E. & R.H. (2000). The relationships of color patterns and habitat for Lower Triassic Ammonoids from Crittenden Springs, Elko County, Nevada.- Revue de Paléobiologie, Genève, volume spécial n° 8, p. 109-122.
J.E. (1779).- Mémoire sur la minéralogie du Dauphiné.- Imprimerie de Clousier rue Saint-Jacques, Paris, t. 1, 298 p. ; t. 2, 554 p.
R. (1886).- Fossiles décrits par .- Bulletin de l'Académie d'Hippone, Bône, n° 28, 1 Pl.
J.O., J., O., P. & C. (2003).- Forcing mechanisms for mid-Cretaceous black shale formation: evidence from the Upper Aptian and Lower Albian of the Vocontian Basin (SE France).- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, n° 190, p. 399-426.
E.F. (1892).- Sur une forme nouvelle de céphalopodes du Crétacé inférieur des Basses-Alpes.- Feuille des Jeunes Naturalistes, Paris, 22ème année, n° 264, p. 241-242.
C. (1908). Étude sur quelques ammonites du Crétacé moyen.- Mémoire de la Société Géologique de France, Série Paléontologie, Paris, n° 15, 64 p.
B. (1972). La structure du test et son altération chez les Phylloceratidae du genre Calliphylloceras.- Haliotis, Société Française de Malacologie, Paris, vol. 2, n° 2, p. 179-185.
B. (1977). - Les Phylloceratidae malgaches au Jurassique. Généralités sur quelques Phylloceratidae et quelques Juraphyllitidae.- Documents des Laboratoires de Géologie de la Faculté des Sciences de Lyon, n° 67, 471 p.
B. (1993).- Les Phyllocerataceae malgaches au Crétacé (Phylloceratina, Ammonoidea).- Documents des Laboratoires de Géologie de la Faculté des Sciences de Lyon, n° 127, 171 p.
B. (1998). Les Phyllocerataceae albiens du département de l'Aube.- Bulletin Annuel de l'Association Géologique Auboise, Sainte-Savine, n° 19, p. 33-59.
B. (2000a). Les Juraphyllitidae, Phylloceratidae, Neophylloceratidae (Phyllocerataceae, Phylloceratina, Ammonoidea) de France au Jurassique et au Crétacé.- Géobios, Lyon, mémoire spécial n° 23 & Mémoire de la Société Géologique de France, Paris, n° 174, 204 p.
B. (2000b). Altérations de la croissance chez les Phyllocerataceae , 1884, conséquence sur la classification. Apports de l'étude des Phyllocerataceae aux modalités de l'évolution.- Comptes Rendus de l'Académie des Sciences, Paris, t. 330, p. 709-716.
B. & C. (2006). Découverte récente de deux Phylloceratidae colorés appartenant au genre Adabofoloceras , 1977. Hypothèses sur le lien entre style de coloration de la coquille et mode de vie.- Bulletin de la Société Géologique de France, Paris, t. 177, n° 1, p. 51-59.
N.I. (1895). [Zur Frage ueber die Synonymie von Phylloceras infundibulum und Ph. Rouyi].- Geologische und mineralogische Mittheilungen der St Petersburger Gesellshaft der Naturforschung, Saint-Petersbourg, n° 4, 14 p. [en russe, résumé en allemand].
N.I. (1907). Le Crétacé inférieur de la Crimée et sa faune.- Travaux de la Société Impériale des Naturalistes de Saint Petersbourg, Géologie et Minéralogie, vol. 32, livr. 5, 482 p.
W.J., A.S., P.R., M., R. J., D., & D.S. (2000). Integrated stratigraphy across the Aptian-Albian boundary in the Marnes Bleues, at the Col de Pré-Guittard, Arnayon (Drôme), and at Tartonne (Alpes-de-Haute-Provence), France : a candidate Global Boundary Stratotype Section and Boundary Point for the base of the Albian Stage.- Cretaceous Research, London, vol. 21, p. 591-720,
W.J. & H.C. (1977).- Cretaceous faunas from Zululand and Natal, South Africa. The ammonite family Phylloceratidae.- Bulletin of the British Museum (Natural History), Geology, London, vol. 27, n° 5, p. 347-380.
W. (1892).- Contribution à l'étude des Céphalopodes Crétacés du Sud-Est de la France. Sur quelques Ammonitidés appartenant au Muséum d'Histoire Naturelle de Lyon.- Archives du Muséum d'Histoire Naturelle de Lyon, t. 5, 9 p.
W. (1896).- Note stratigraphique sur les environs de Sisteron et contributions à la connaissance des terrains secondaires du Sud-Est de la France.- Bulletin de la Société Géologique de France, Paris, (3ème série), t. 23, p. 659-803.
W. (1907-1913).- Das Mesozoicum Unterkreide (Palaeocretacicum).- Lethea Geognostica, Stuttgart, teil 2, t. 3, p. 1-168 (1907) et p. 289-398 (1913).
F. (1895).- Untersuchungen über die Südindische Kreideformation.- Beiträge zur Paläontologie und Geologie Österreich-Ungars und des Orients, Wien, t. 9, p. 97-203.
H. (1834).- Coquilles fossiles de Gérodot (Aube).- Magazine de Zoologie, Paris, n° 3, Classe V, Pl. 35.
M. (1966). Étude stratigraphique et micropaléontologique du Crétacé inférieur de la "Fosse vocontienne".- Documents du Laboratoire de Géologie de la Faculté des Sciences de Lyon, vol. 15, 369 p.
M.A. & P.U. (2006). California Early Cretaceous Phylloceratidae (Ammonoidea).- University of California, Riverside, Campus Museum Contribution, n° 7, 98 p.
A. d' (1840-42).- Paléontologie française, terrains crétacés, I - Céphalopodes.- Masson édit., Paris, 662 p. + 148 Pls.
A. d' (1850). Notes sur quelques nouvelles espèces remarquables d'ammonites des étages Néocomien et Aptien de la France.- Journal de Conchyliologie, Paris, t. 1, p. 196-201.
A. d' (1850-1852).- Prodrome de paléontologie stratigraphique universelle des animaux mollusques et rayonnés.- Masson, Paris, t. 1, 394 p. ; t. 2, 427 p. ; t. 3, 196 p.
V. (1900). Recherches géologiques dans le Diois et les Baronnies orientales.- Travaux du Laboratoire de Géologie de la Faculté des Sciences de l'Université de Grenoble, t. 5, n° 2-3, p. 373-806.
L. (1907).- Étude de paléontologie tunisienne - 1. Céphalopodes des terrains secondaires.- Mémoire de la carte géologique de Tunisie, Paris, 438 p.
L. (1910).- Sur quelques ammonites du Crétacé algérien.- Mémoire de la Société Géologique de France, série Paléontologie, Paris, t. 17, fascicules 2-3, mémoire n° 42, 86 p.
V.K. (1913).- Étude sur le Gault en Serbie.- Bulletin de l'Académie royale de Serbie, Belgrade, n° 89, 139 p.
F.V. (1831).- Histoires des ammonites, suivie de la description des ammonites des Basses-Alpes et des Cévennes.- Le Lycée, Paris, n° 27, Librairie Hachette, p. 115 (les Pls. 11-12 sont parues en 1830, in Annales des Sciences d'Observation, Paris, t. 3).
S., F. & O. (2005). Ammonoid abundance variations related to changes in trophic conditions across the oceanic anoxic event 1d (latest Albian, SE France).- Palaios, Research Report, Lawrence, vol. 20, p. 121-141.
L. (1842).- Description des principaux fossiles de la Crimée. In : A. (coord.), "Voyage dans la Russie méridionale et la Crimée".- Bourdin édit., Paris, vol. 2, p. 781-819 et p. 821-823.
J.-L. (1982). Les grès albo-aptiens dans le bassin vocontien et sur sa bordure occidentale.- IX Réunion Annuelle des Sciences de la Terre, Paris, p. 540.
J.-L. (1988). Organisation des séquences de dépôt de la plate-forme au bassin dans l'Aptien et l'Albien du bassin vocontien (S-E de la France).- Géotrope, Lyon, n° 1, p. 56-82.
H. (1924).- Die Bedeutung der Konservativstämme für die Stammesentwicklung der Ammonoitiden.- Max Weg ed., Leipzig, 16 p.
G. (1890). Description des ammonitidés du Barrémien du Djebel Ouach, près Constantine.- Bulletin de la Société agricole et d'Histoire Naturelle de Lyon, (6ème série), t. 3, 78 p.
G. (1920).- Les Phylloceras gargasiens du S.E. de la France (espèces nouvelles ou peu connues).- Mémoire pour servir à l'explication de la carte géologique détaillée de la France, Paris, p. 191- 203.
L.F. (1925). On upper Albian Ammonoidea from Portuguese East Africa with an appendix on Upper Cretaceous ammonites from Maputoland.- Annals of the Transvaal Museum, Pretoria, vol. 11, p. 179-200.
L.F. (1927-33).- Revision of the Jurassic Cephalopod Fauna of Kachh (Cutch).- Palaeontologica Indica, Calcutta, (new series), t. 9, n° 2, 945 p.
F. (1865).- The fossil Cephalopoda of the Cretaceous rocks of Southern India.- Palaeontologica Indica, Calcutta, t. 3, n° 6-9, p. 107-154.
E. (1865).- Über Ammoniten.- Sitzberichte der Akademie der Wissenshaften, mathematisch-naturwissenshaften Classe, Wien, vol. 52, p. 71-89.
G. (1960). Observations sur l'Aptien et sur l'Albien des environs de Puget-Théniers (Alpes-Maritimes).- Bulletin de la Société Géologique de France, Paris, t. 7, n° 2, p. 87-93.
G. (1963). Considérations sur la faune gargasienne du Sud-est de la fosse vocontienne.- Travaux du Laboratoire de Géologie de la Faculté des Sciences de Marseille, t. 7, p. 47-56.
G. (1964). Les zones d'ammonites de l'Aptien des Basses Alpes.- Comptes Rendus de l'Académie des Sciences, Paris, groupe 9, t. 258, p. 4308-4310.
G. (1965). L'évolution post-aptienne des Leiostraca dans le Sud-Est de la France.- Comptes Rendus de l'Académie des Sciences, Paris, groupe 9, t. 260, p. 5606-5608.
G. (1968). À propos de l'Ammoniceratites (Argonauticeras) depereti () du Gargasien des Basses Alpes orientales.- Bulletin de la Société Géologique de France, Paris, t. 7, n° 10, p. 684-687.
G. (1980). Ammonites.- Éditions Serre, Nice, 227 p.
G. (1987a). La famille des Tetragonitidae (Ammonoidea) dans le Cénomanien du Sud-Est de la France.- Annales de Paléontologie (Vertébrés-Invertébrés), Paris, t. 73, n° 4, p. 241-272.
G. (1987b). La faune des marnes gargasiennes des Alpes de Haute-Provence - Phylloceratidae.- Lithos, Paris, vol. 3, p. 42-50.
V. (1883).- Die Cephalopodenfauna der Wernsdorfer Schichten.- Denkschriften der Österreichischen Akademie der Wissenshaften mathematisch-naturwissenschaflichen Classe, Wien, vol. 46, t. 2, p. 1-165.
J. (1964).- Unterkreide-Ammoniten von Mallorca - 2 Lieferung: Phylloceratina.- Abhandlungen der mathematisch-Naturwissenschaftlichen Klasse, Akademie der Wissenschaften und der Literatur in Mainz, Wiesbaden, n° 4, p. 151-256.
J. (1973). The Albian and Cenomanian Tetragonitidae (Cretaceous Ammonoidea) with Special Reference to the Circum-Indic Species.- Eclogae geologicae Helvetiae, Bâle, vol. 66, p. 585-616.
K.A. von (1884).- Handbuch der Paleontologie, Cephalopoden.- R. Oldenbourg, München et Leipzig, t. 2, p. 329-522.