![]()
Sommaire
[Introduction] [I - Les ammonites du gisement de Saint-Florent ZI ...] [II - De la position stratigraphique ...]
[III - Analyse synécologique ...] [Conclusion] [Références bibliographiques] et ... [Planches]
![]()
Une nouvelle récolte d'ammonites dans les marnes à ammonites ferrugineuses de la région de Niort (France), autrefois étudiées par de (1922), apporte de très nombreuses informations biostratigraphiques, paléoenvironnementales et paléobiogéographiques. La faune est datée de l'Oxfordien inférieur : Zone à Cordatum, Sous-Zone à Cordatum, partie inférieure probable. De plus, elle est unique en Europe occidentale 1) par les espèces trouvées -dont certaines ne semblent connues que dans ce gisement- 2) par la domination très atypique du binôme Taramelliceras - Creniceras qui représente les 2/3 des individus. Enfin, le spectre faunique plaide en faveur d'un milieu de plate-forme distale inconnu dans les autres gisements européens de même âge. Une figuration détaillée des différentes espèces est donnée, parfois accompagnée d'une nouvelle interprétation taxonomique.
Ammonites ; Oxfordien inférieur ; Zone à Cordatum ; spectre faunique ; paléoenvironnement ; paléobiogéographie.
P., D., R., A., D. & P. (2009).- La faune ammonitique des marnes à fossiles ferrugineux de la région de Niort, France (Oxfordien inférieur, Zone à Cordatum, Sous-Zone à Cordatum).- Carnets de Géologie / Notebooks on Geology, Brest, Article 2009/05 (CG2009_A05)
The ammonite fauna of the 'marls with ferruginous fossils' from the Niort region, France (Lower Oxfordian, Cordatum Zone, Cordatum Subzone).- A new collection of ammonites from the 'marls with ferruginous ammonites' in the Niort region (France), previously studied by (1922), provides a large amount of biostratigraphic, paleoenvironmental and paleogeographic information. The fauna is dated Early Oxfordian: Cordatum Zone, Cordatum Subzone, probably the lower part of it. In addition, it is unique in western Europe owing to: 1) the species found of which some appear to be known only in this deposit 2) by the extremely atypical domination of the couple Taramelliceras - Creniceras which represent 2/3 of the specimens. Furthermore, the aspect of the fauna strongly suggests that the habitat was a distal platform unknown in other European strata of the same age. A detailed description of the several species is given, sometimes accompanied by a new taxonomic interpretation.
Ammonites; Lower Oxfordian; Cordatum Zone; faunal spectrum; palaeoenvironment; palaeobiogeography.
Dans la notice de la carte de Niort au 1/50.000ème ( et alii, 1983), au-dessus des marnes grises et calcaires fins, épais d'environ 7m, datés du Callovien supérieur (Zone à Lamberti), les auteurs indiquent la présence d'une épaisse série marneuse (35 à 40 m) qu'ils subdivisent en deux parties : 1) à la base, des marnes gris-bleu à noirâtres, à petites ammonites pyriteuses sur environ 8 m et 2) au dessus, une trentaine de mètres de marnes grises à spongiaires. Ils datent la première formation de l'Oxfordien inférieur (Zone à Cordatum) par son association ammonitique qui renferme en autre l'espèce Cardioceras costicardia , et la seconde de l'Oxfordien moyen (Zone à Plicatilis) à la base de l'Oxfordien supérieur (Zone à Bifurcatus).
Au cours de l'année 2001, des travaux ont été effectués à environ 3 km au sud de Niort afin de créer, à partir de la N 201, la sortie Saint-Florent ZI. Ces travaux ont permis de remettre à jour la formation des marnes gris-bleu et ainsi de récolter une très abondante faune d'ammonites ferrugineuses, rarement pyritisées, soit 4342 individus dans l'échantillon ici analysé.
La faune de ce niveau a été décrite pour la première fois par de (1922) à la suite de travaux effectués au sud de Niort (Deux Sèvres) dans la tranchée des Grosses Terres (voie ferrée reliant Niort à La Rochelle) et dans la première tranchée juste avant la station d'Aiffres (voie ferrée reliant Niort à Saint-Jean-d'Angely). Cet auteur constate que la faune d'ammonites est abondante, toujours de petite taille (rarement au dessus de 30 mm) et qu'elle est diversifiée puisqu'il cite 10 genres pour un total de 17 espèces. Pour lui, cette faune caractérise, à juste raison, la Zone à Cordatum qui, à cette époque, était rangée dans l'Oxfordien moyen et non pas comme aujourd'hui dans la partie sommitale de l'Oxfordien inférieur. En effet, il avait remarqué que le genre Creniceras n'était pas représenté par l'espèce C. renggeri mais par l'espèce C. crenatum et que le genre Cardioceras était surtout représenté par l'espèce C. cordatum .
Les récoltes effectuées par l'un d'entre nous (), à la manière d'A. de , "sur des talus d'éboulement", permettent de préciser : 1) l'âge de la faune, qui est clairement homogène non seulement par les Cardioceratinae mais aussi par les autres taxons, 2) le spectre faunique de la population ammonitique qui donne un 'accès biologique' aux conditions environnementales du dépôt de ces marnes et 3) de compléter les descriptions d'A. de , en renommant de façon moderne ses espèces (en particulier au niveau du genre), de décrire des espèces non signalées et enfin de proposer des couples microconque - macroconque, ce qui ne se faisait pas à cette époque.
(Pl. 1 ![]() )
)
De dit être en possession de sept spécimens de Cardioceras cordatum , d'un spécimen de C. cf. quadratoides , et de deux individus de petite taille qu'il range dans l'espèce C. alternans von . Si la première détermination est correcte, par le nom et l'âge, les deux autres ne le sont pas. En effet, son espèce C. cf. quadratoïdes est clairement une forme de l'Oxfordien moyen par son ornementation mais aussi, à n'en pas douter, par sa fossilisation qui correspond à celle des ammonites des marnes à spongiaires sus-jacentes. Quant à sa troisième espèce, le nom est à l'évidence erroné puisque l'espèce C. alternans von appartient en réalité au genre Amoeboceras, un genre qui ne débute qu'au sommet de l'Oxfordien moyen et persiste durant tout l'Oxfordien supérieur.
Dans notre échantillon, le nombre d'individus appartenant à ce genre est de 30 et les diamètres vont de 6 à 23 mm, la majorité se situant entre 8 et 13 mm. Parmi eux, certains possèdent une partie de la loge d'habitation mais aucun ne présente les sutures rapprochées caractéristiques du stade adulte, ce qui en font des nuclei. Ces Cardioceras ont en commun des côtes primaires bien marquées, avec le plus souvent des tubercules bien développés ; les côtes secondaires sont relativement courtes car les points de bifurcation sont situés au dessus du milieu du flanc. Enfin, on observe quelques côtes tertiaires qui naissent à l'approche du méplat ventral voire un peu au-dessus.
Les deux grands groupes morphologiques autrefois rangés dans les sous-genres Cardioceras et Subvertebriceras sont présents.
Dans le premier groupe, la section est ogivale, les côtes primaires sont peu tuberculées et les secondaires sont souvent très proverses à l'approche du ventre, avec un effilement accusé des côtes qui se terminent chacune par un renflement, ce qui donnera les crénulations de la carène.
Dans le second groupe, la section est plus rectangulaire et, corrélativement, les côtes sont moins nombreuses, la tuberculisation plus marquée et les côtes tertiaires plus rares. À noter que les côtes secondaires sont situées sur une sorte de méplat latéral qui caractérise les formes du sommet de la Sous-Zone à Cordatum et de la base de la Sous-Zone à Vertebrale.
Ces individus se distinguent des Cardioceras de la Sous-Zone à Costicardia sous-jacente par un ensemble de caractères. En effet, les spécimens récoltés dans la Sous-Zone à Costicardia [telle qu'on la connaît dans le Jura grâce aux nombreuses récoltes de l'un d'entre nous () ou en Angleterre, dans le Dorset (environs de Weymouth, récoltes de ), ou dans la région de Studley (, 1946-1947)] présentent une ornementation à tubercules moins accusés, une section dans l'ensemble plus ogivale, des côtes tertiaires plus rares et surtout une projection ventrale des côtes moins accusée.
Les individus de Saint-Florent semblent également se distinguer des Cardioceras du sommet de la Sous-Zone à Cordatum [gisements d'Herznach ( & , 1977 ; & , 1982), de Neuvizy et du Jura (, 1986)] ou de l'extrême base de la Sous-Zone à Vertebrale [gisements des Heersumer Schichten en Allemagne du Nord (, 1953) ou de Sy dans les Ardennes ( et alii, 1977 ; , 1986)]. En effet, les individus à côtes secondaires nombreuses et très effilées à l'approche du ventre, décrites sous les noms de C. tenuicostatum et C. tenuistriatum , typiques de ces faunes un peu plus jeunes, sont absents à Saint-Florent.
(Pl. 2 ![]() )
)
Dans son travail de 1922, de rappelle que (1914) considérait comme Taramelliceras les formes à côtes 'falculiformes', présentant fréquemment des tubercules soit sur les flancs soit sur la région externe, mais toujours des tubercules médians sur la région siphonale. Intéressant à noter, rattachait à ce groupe des formes nettement plus petites caractérisées par une rangée de tubercules tranchants sur la région siphonale regroupant des espèces comme A. renggeri , A. crenatus et A. dentatus toutes représentantes actuelles du genre Creniceras. Il rappelle aussi que (1898) considère les Creniceras comme 'probablement les formes mâles des Neumayria' (= Taramelliceras) et considère lui même que 'le genre Creniceras ne peut pas être distingué des Taramelliceras lorsque les deux ont une taille inférieure à 10-12 mm'.
A Saint-Florent, de a distingué 3 espèces de Taramelliceras :
Cette dernière espèce, la plus abondante, se distingue des deux précédentes par sa faible épaisseur et une ornementation faite de côtes primaires arquées, peu saillantes, donnant naissance à un faisceau de trois côtes ; les tubercules médians sont petits et peu saillants.
Nos observations s'accordent parfaitement avec celles d'A. de puisque nous avons récolté 2071 spécimens de T. frequens contre 4 individus que l'on pourrait rattacher morphologiquement à T. ledonicum, espèce que de (1900) signale dans la Zone à Praecordatum (Oxfordien inférieur, Zone à Mariae).
La taille maximale des Taramelliceras de Niort ne dépasse pas 27 mm et la moyenne se situe vers 15 mm. On ne peut donc donner que quelques éléments de description. L'épaisseur est faible, l'ombilic étroit, le ventre arrondi et les tubercules, toujours ténus à ces diamètres, n'apparaissent jamais avant 10 mm. La costulation est souvent très peu marquée mais ce sont toujours des côtes falciformes où les côtes primaires sont plus marquées que les secondaires. On observe aussi que la ligne de suture se complique très vite au cours de l'ontogenèse.
Il est à noter que, d'un strict point de vue morphologique, cette population se distingue difficilement des nombreux spécimens jurassiens rapportés à l'espèce Taramelliceras richei de et récoltés dans les niveaux de la Zone à Mariae (horizon à Praemartini, niveaux Ox6 et Ox7, , à paraître).
(Pl. 3 ![]() , figs. 3-7)
, figs. 3-7)
En suivant les remarques de (1917), de range les Creniceras de Niort dans l'espèce C. crenatum parce que, chez les adultes, le déroulement de l'ombilic se fait de façon progressive et non brutale comme chez C. renggeri , une espèce de la Zone à Mariae. Il signale aussi que cette espèce, dont la taille maximale atteint 17 mm, est très abondante à Niort. Cette observation est en accord avec notre échantillon puisque "C. crenatum" (N = 892) est, en nombre, la seconde espèce après Taramelliceras frequens de . Ceci confirme que le genre Creniceras est bien, comme le proposait dès 1966, la forme microconque du genre macroconque Taramelliceras.
Au texte d'A. de , on peut ajouter que la variabilité de la taille adulte est grande (de 9 mm à 19 mm) et que la force des tubercules ventraux est variable d'un individu à l'autre mais presque toujours plus forte chez les individus de plus grand diamètre.
On peut toutefois s'interroger sur la pertinence du nom d'espèce choisi par de , dans la mesure où les spécimens figurés par de sont variables d'une publication à l'autre et ne répondent pas bien au critère d'ouverture ombilicale mis en avant par de . En outre, C. crenatum est une espèce de l'Oxfordien moyen, Zone à Transversaruim.
(Pl. 3 ![]() , figs. 1-2)
, figs. 1-2)
Dans son article de 1922, de range dans le genre Creniceras une nouvelle espèce qu'il nomme C. gourbinei. Pour lui, les principales caractéristiques morphologiques de cette espèce sont : un ombilic de grandeur assez variable limité par un rebord anguleux, une épaisseur maximale près de l'ombilic, un ventre étroit "où court une succession très régulière de petits tubercules arrondis, saillants, qui existent déjà à une taille assez faible", tubercules dont la taille croît de façon lente et régulière ; de plus, les côtes sont peu marquées. De signale par ailleurs qu'il y a fréquemment "un déplacement latéral de la selle siphonale rejetée à droite ou à gauche ce qui correspond à un déplacement du siphon qui passe toujours par le milieu de la selle". Pour lui, cette espèce est peu commune, sans être absolument rare.
Nous avons bien sûr retrouvé cette espèce (N = 18) dont la description est parfaite. On peut cependant ajouter les remarques suivantes :
Si l'on met l'ammonite dans sa position de vie, la plupart des individus ont le siphon ventral nettement décalé soit sur le flanc droit (N = 9), soit sur le flanc gauche (N = 3). Deux individus ont un siphon très peu décalé vers la gauche et chez les 4 derniers spécimens le siphon n'est pas visible : était-il parfaitement axial ?
On constate qu'il n'y a jamais de déroulement de l'ombilic même chez le plus grand spécimen (un phragmocône de 24 mm). Le seul spécimen (D = 20 mm) qui a conservé sa loge d'habitation (presque complète) a des sutures non rapprochées et son ombilic ne s'ouvre pas : rien n'indique un individu microconque sub-adulte.
Le plus grand spécimen développe une ornementation de type Campylites à partir de 12 mm ; cette ornementation est inconnue chez tous les autres spécimens. Un spécimen présente un net sillon latéral alors que chez d'autres spécimens, quand ce sillon est présent, il est toujours très atténué.
Position systématique : Cette espèce, qui a une taille adulte très supérieure à celle des Creniceras s.s., des tubercules ventraux très différents et des sutures simplifiées, ne peut pas appartenir au genre Creniceras. Et comme la position déjetée du siphon ventral ne nous est connue que dans la Sous-Famille des Hecticoceratinae, l'espèce gourbinei peut en toute logique appartenir à cette sous-famille. Les éléments d'ornementation qui peuvent être relevés sur une partie des individus (côtes, sillon latéral), classiques chez les Campylites de l'Oxfordien inférieur, confirment pour nous cette attribution.
Dans son travail de 1898, de ne figure aucun spécimen de ce type. Par contre, dans celui de 1900, deux spécimens appelés Oppelia baylei (Pl. III, figs. 27-28) présentent des similitudes : ombilic assez ouvert, ligne de petits tubercules axiaux, flancs lisses, sutures peu complexes. Mais des différences existent chez ces O. baylei : 1) l'épaisseur est un peu plus forte, 2) on note l'apparition de deux carènes ventro-latérales lisses, très écartées de l'axe ventral vers 15 mm et 3) des tubercules ventraux plus allongés. Il nous paraît donc raisonnable de considérer ces deux formes comme deux espèces diachrones faisant partie d'un même genre, que nous désignerons provisoirement par 'Hecticoceras'. Mais il nous semble, étant donné la présence d'une rangée de petits tubercules ventraux, qu'un nouveau nom de genre serait plus approprié. Mais comme nous ne disposons pas d'un matériel suffisant (adultes, grands spécimens, dimorphisme ?) pour établir une diagnose fiable, il nous semble raisonnable d'attendre.
(Pl. 4 ![]() , figs. 1-2)
, figs. 1-2)
Ce genre a été créé par en 1909 pour l'espèce calcarata (1853), une forme caractérisée par "un ventre aplati, tricaréné et bisulqué" d'après ses propres termes. En 2002, à partir de formes récoltées dans la Zone à Mariae du Jura, et alii ont décrit Sphaerodomites comme un taxon micromorphe. Au sein de la faune de Niort, à la différence d'A. de , nous avons récolté 4 individus qui présentent à l'évidence des analogies avec cette espèce micromorphe. On peut cependant noter que les formes de Niort ont un ventre moins aplati et une expression variable de la carène selon les individus. Pour ces raisons, nous proposons de l'appeler Sphaerodomites cf. calcarata .
(Pl. 6 ![]() )
)
Pour de , l'espèce la plus fréquente est P. bernensis de . Dans notre échantillon, tous les individus appartenant à ce genre sont étonnement similaires, en particulier par l'épaisseur qui est relativement forte, ce qui les distingue très aisément des formes figurées par de (1898, 1900). Étant donné que les diamètres ne dépassent que très rarement 25 mm, nous n'avons pas pu mettre en évidence des adultes de petite taille à sutures rapprochées qui seraient alors des microconques. Mais il est intéressant de noter que les morphologies observées sont proches de celles récemment figurées par & (2004) en provenance d'un gisement polonais daté de la Sous-Zone à Praecordatum (horizon à Alphacordatum probable) et nommées Perisphinctes (Properisphinctes) cf. bernensis de .
(Pl. 8 ![]() , figs. 1-16)
, figs. 1-16)
Dans toutes les populations d'âge Callovien supérieur (au moins depuis la Zone à Lamberti) et Oxfordien inférieur à moyen, les macroconques Euaspidoceras coexistent toujours avec les microconques Mirosphinctes (, 1995 ; et alii, 2002).
(Pl. 8 ![]() , figs. 1-11)
, figs. 1-11)
De a rangé les Euaspidoceras de Niort dans l'espèce Perisphinctes perisphinctoides et var. armata de . Il précise que les individus de cette espèce ont une ornementation qui va de côtes régulières sans renflements à des côtes avec tubercules très saillants. De plus, il signale à juste titre que les cloisons des individus de Niort rapportés à P. perisphinctoides sont très simples et que ces formes, contrairement à Perisphinctes bernensis de , ont des sutures sans lobe suspensif. Ce dernier caractère sutural est de fait très important puisqu'il permet de distinguer sans risque d'erreur les Perisphinctidae des Aspidoceratidae.
À Saint-Florent, le genre Euaspidoceras est représenté de façon très inégale par deux espèces : tous les spécimens peuvent être attribués à E. freii excepté un individu qui doit être rattaché à E. ovale .
E. freii
La plupart des individus sont totalement cloisonnés. Quelques rares juvéniles possèdent une partie de leur chambre d'habitation. L'enroulement est subserpenticône et la section entre les tubercules est subcirculaire.
Après un stade lisse très court (quelques millimètres de diamètre), l'ornementation débute avec des côtes assez fortes, proverses, dont certaines possèdent des formations paraboliques : d'abord de simples paraboles puis des nuds paraboliques qui se redressent et donnent naissance aux tubercules épineux externes qui sont fossilisés sur le moule interne : ces tubercules ne sont donc pas cloisonnés ; ils apparaissent précocement, entre 11 et 15 mm de diamètre. Les paraboles internes sont peu développées. Après l'apparition des tubercules externes, les côtes s'espacent et on observe entre elles une fine striation qui s'efface assez rapidement. L'apparition des tubercules internes n'a pas pu être observée avec précision, mais un fragment présente déjà, à un diamètre d'environ 28 mm, de petits tubercules internes sur le rebord ombilical.
E. freii représente la morphologie "moyenne" des Euaspidoceras, une morphologie qui débute au Callovien supérieur et perdure jusqu'à l'Oxfordien supérieur ; les espèces successives ne se distinguent que par des détails d'ornementation. Ainsi, si E. freii est très proche par son enroulement d'E. douvillei de la Sous-Zone à Bukowskii, il s'en distingue aisément par la précocité de son stade tuberculé puisque, chez E. douvillei , les tubercules externes apparaissent entre 10 et 25 mm alors que les tubercules internes, nettement plus tardifs, n'apparaissent qu'entre 70 et 140 mm.
E. ovale ()
Il s'agit d'un très petit nucleus (D = 12mm) totalement cloisonné, qui montre la morphologie classique de l'espèce à ce diamètre : enroulement subserpenticône, section ovale, ornementation très réduite, irrégulière, avec seulement quelques côtes un peu plus fortes (visibles à la loupe), mais non encore paraboliques. Il n'est pas possible de savoir si ce spécimen est un macroconque ou un microconque.
(Pl. 8 ![]() , figs. 12-16)
, figs. 12-16)
Tous les microconques peuvent être attribués à Mirosphinctes niedzwiedzkii . La plupart des spécimens sont totalement cloisonnés, mais les 2 ou 3 dernières cloisons ne sont jamais rapprochées ; quelques individus ont une partie de la chambre d'habitation. L'enroulement est subserpenticône et la section subcirculaire. Il n'y a pas de constrictions. Après un stade lisse très court, débute le stade à ornementation parabolique : côtes assez fortes, proverses, à paraboles internes et paraboles externes évoluant en nuds paraboliques, et côtes intermédiaires rapidement convexes, simples ou bifurquées à mi-flanc, passant sur le ventre. Les nuds paraboliques sont encore présents sur la chambre d'habitation juvénile (alors qu'ils sont absents sur la chambre adulte, au moins chez les spécimens d'âge Callovien supérieur à Oxfordien inférieur). La variabilité touche essentiellement la fréquence des formations paraboliques et la force de la costulation non parabolique sur la chambre d'habitation.
Mirosphinctes niedzwiedzkii est proche de M. mirus (= Euaspidoceras douvillei , microconque), mais ce dernier est nettement plus grand et ses formations paraboliques sont nettement plus prononcées.
La ligne de suture est construite de façon identique chez les dimorphes, même si, à diamètre identique, celle du microconque est toujours moins découpée que celle du macroconque. Ils partagent une même ornementation dans les tours initiaux, mais les formations paraboliques sont plus fréquentes chez le macroconque : pour un diamètre compris entre 15 et 20 mm, on dénombre de 9 à 13 paraboles chez le macroconque contre 5 à 10 chez le microconque. Au cours de son ontogenèse, le macroconque présente une puis deux rangées de tubercules alors que le microconque reste toujours simplement costulé.
(Pl. 8 ![]() , figs. 17-19)
, figs. 17-19)
De ne signale pas sa présence à Niort. Pour notre part, nous sommes en possession de trois nuclei qui peuvent être attribués à ce genre. La section est sub-trapézoïdale à rebords anguleux et il existe un mur ombilical individualisé, oblique puis vertical.
Après un court stade lisse, les côtes prennent naissance sur le rebord ombilical. Elles sont d'abord très proverses, puis s'infléchissent sous le milieu des flancs à partir duquel elles sont franchement rétroverses. Elles passent sur la région ventrale en s'atténuant légèrement. Certaines de ces côtes sont plus fortes et montrent des paraboles ombilicales. Vers 11-12 mm, des nuds paraboliques apparaissent sur le rebord latéro-ventral, rapidement suivis chez le spécimen le plus grand par des tubercules spatulés (caractéristiques des macroconques).
Ces 3 individus sont morphologiquement proches de l'individu figuré par et alii en 1997.
Dans son travail, de indique qu'il est en possession d'un spécimen de petite taille qu'il rapporte à Peltoceras aff. eugenii d' (= Peltoceratoides cf. eugenii). Ce genre, abondant dans certains niveaux de l'Oxfordien inférieur, devient très rare à son sommet et n'est plus connu à la base de l'Oxfordien moyen. Il est absent de notre échantillon. L'absence, ou l'extrême rareté de ce genre dans un gisement de cet âge, constitue la seule exception connue en Europe.
(Pl. 5 ![]() )
)
De rappelle que ce genre, créé par en 1911, se caractérise par un ventre tricaréné et des côtes falciformes peu nombreuses et peu marquées vers l'ombilic. Puis il ajoute, avec justesse : 1) que les côtes sont moins effacées chez les spécimens dont l'ombilic est un peu plus large et 2) que les individus de petite taille ont encore un ventre arrondi qui deviendra plus anguleux avant l'apparition des deux carènes latérales. Après une discussion assez longue sur les espèces d'âge Oxfordien, de propose de regrouper les individus de Niort au sein d'une nouvelle espèce, Trimarginites sauvageti caractérisée par des côtes 'falculiformes' serrées qui se bifurquent au point d'inflexion. Il ajoute que le rapport O/D varie de 0,13 à 0,25 et le rapport E/H de 0,37 à 0,50.
Dans notre population on constate que le morphe 'trimarginites', tel que de l'a décrit, est très largement majoritaire, surtout aux diamètres les plus grands qui peuvent atteindre 30 mm. Cependant, à côté de ces individus 'typés', on trouve aussi des formes à ombilic plus ouvert, à costulation falciforme plus visible, voire accusée : dans ce dernier cas, les côtes sont moins nombreuses. Mais chez ces formes, on constate que la tricarénation est moins nette puisque le ventre est un peu plus arrondi. En ce qui concerne les lignes de sutures, on constate que les formes à ombilic étroit et costulation peu marquée -voire absente- ont des sutures plus complexes que les formes plus costulées à ombilic plus ouvert.
Entre ces deux pôles extrêmes, il existe en réalité un continuum morphologique. Ce phénomène a été signalé par (2003) pour les formes récoltées par en Suisse dans la région d'Herznach et le canton de Schaffhausen. Dans ces faunes, a montré qu'il existe de nombreuses formes de passage entre un pôle trimarginites mince, involute, peu costulé, à sutures complexes et un pôle campylites un peu plus épais, plus évolute, plus costulé, à sutures moins complexes. Il a montré aussi que le pôle trimarginites était clairement plus abondant dans les milieux plus ouverts. Dans le cas des faunes suisses, il y aurait donc une continuité morphologique entre le genre Trimarginites et le genre Campylites . Dans le gisement de Niort, le pôle trimarginites est clairement le plus abondant.
Nous proposons ici d'abandonner pour nos formes l'appellation trimarginites qui renvoie dans la littérature à toutes sortes de formes, situées à des niveaux stratigraphiques très différents (Callovien supérieur, Oxfordien inférieur : zones à Mariae et Cordatum, Oxfordien moyen : zones à Plicatilis et Transversarium). Rappelons que l'espèce type du genre proposée par provient du sommet de l'Oxfordien moyen (Trimarginites arolicus, Zone à Transversarium).
De nombreux spécimens de la population de Niort présentent toutes les caractéristiques du genre Campylites tel qu'il a été défini par à partir de l'espèce delmontanus . Cette morphologie, qui est déjà connue dès le milieu de la Zone à Mariae, se poursuit donc dans la Zone à Cordatum ; de ce fait, nous préférons ranger tous les individus de Niort dans le genre Campylites.
L'espèce sauvageti, telle qu'elle a été proposée par de , tient compte par définition de la variabilité intra-spécifique : en effet, tous les individus récoltés sont rangés par lui dans la même espèce. Citons de : "De cette espèce il y a des variétés à petit ombilic, à flancs lisses ou avec ornementation très atténuée, et d'autres avec ombilic plus large et ornementation plus ou moins accentuée."
Cette espèce sauvageti est donc une meilleure candidate au statut de bio-espèce que l'espèce delmontanus , utilisée par (1951) pour décrire les seuls morphes costulés provenant de la Sous-Zone à Cordatum d'Herznach. Et de plus, la dénomination d' correspond à un individu provenant de l'Oxfordien moyen.
(Pl. 4 ![]() , figs. 4-6)
, figs. 4-6)
Ce genre a été créé par (1892) pour des formes du Bajocien supérieur. Et dans le 'Treatise on Invertebrate Palaeontology' ( et alii, 1957), le genre Cadomoceras n'est cité que dans le Dogger. Mais pour de , ce genre cryptogène persisterait plus longtemps puisqu'il propose d'y ranger les formes "scaphitoïdes" trouvées à Niort. Ces formes sont regroupées au sein de sa nouvelle espèce Cadomoceras cossmanni et pour lui, si son espèce est proche d'Ammonites scaphitoides et d'A. paucirugatus , elle s'en distingue : 1) par un ombilic moins fermé dans les tours internes, 2) par un déroulement plus progressif et 3) par un dernier tour beaucoup moins géniculé. De plus, les flancs de sa nouvelle espèce sont presque plats à la différence des flancs bombés d'A. scaphitoides appelé aujourd'hui Scaphitodites scaphitoides ( et alii, 1997 ; et alii, 2002). Et de note aussi très justement que, au moins vers le début du dernier tour (en réalité sur le début de la loge d'habitation), il y a une arête ventrale peu accusée où se trouvent de petits tubercules irréguliers, par la taille et l'espacement. Il signale même une petite dépression spirale située un peu au dessus du milieu des flancs qu'il pense être le canal de la languette buccale si classique dans la famille des Oppeliidae lorsqu'il existe une apophyse jugale.
La population d'A. de se distingue surtout du genre Scaphitodites défini par en 1924 à partir de son espèce navicula par un déroulement progressif et non clairement 'scaphitoïde'. D'un point de vue strictement morphologique, les formes de Niort sont proches des Glochiceras de l'Oxfordien moyen bien que les tubercules ventraux soient beaucoup moins marqués. C'est donc avec réserve que nous acceptons ce nom de genre pour les individus de Niort.
L'espèce cossmanni d'A. de se distingue de toutes les espèces signalées dans la littérature (C. scaphitoides , 1853, C. navicula , 1924, C. paucirugatus , 1887), espèces qui sont classiquement rangées dans le genre Scaphitodites. Par contre, l'espèce C. ogerieni , 1928, est très proche de celle récoltée à Niort et pourrait être un synonyme. Mais comme le nom d'espèce cosmanni a été créé par de en 1922, il a priorité de date et doit donc être conservé. Par contre, nous pensons que le nom de genre Cadomoceras doit être abandonné pour cette espèce que nous nommons, avec doute, Glochiceras ? cossmanni de .
(Pl. 7 ![]() )
)
De regroupe les rares individus appartenant à cette famille au sein du genre Sphaeroceras sans procéder à une tentative de description précise. En effet, il considère que les spécimens sont trop peu nombreux et trop petits pour valider la création d'une espèce nouvelle.
En réalité, les 9 spécimens en notre possession, dont le diamètre varie de 8 à 16 mm, montrent des caractéristiques morphologiques bien précises. La coquille, de type sphaerocone, est plus ou moins épaisse et possède une section arrondie à légèrement ogivale. La plus grande épaisseur se situe sur le rebord ombilical et la forte croissance en épaisseur entraîne un ombilic profond. Les côtes sont radiales à faiblement proverses, simples ou divisées en deux ou trois secondaires, qui traversent la région ventrale sans se modifier. La costulation est très discrète sauf sur un seul spécimen où elle est plus accusée. De rares côtes sont paraboliques, avec une parabole interne (ombilicale) allongée et une parabole externe (ventrale) arrondie. Chez un autre spécimen, les paraboles sont nettement plus fortes, associées à une augmentation brutale de la hauteur. À signaler enfin que les trois spécimens qui possèdent une partie de leur chambre d'habitation n'ont jamais de sutures rapprochées.
Ces 9 spécimens sont rangés dans le genre Tornquistes , mais leur très petite taille ne permet pas de leur attribuer un nom spécifique.
On peut rappeler que dans le Jura, les populations du sommet de la Zone à Mariae présentent un dimorphisme net, avec des paraboles présentes essentiellement dans les tours internes des individus microconques, tandis que les macroconques semblent en être dépourvus. On peut être tenté de transposer ces observations à la population plus récente de Niort, malgré l'absence de formes indiscutablement adultes. Dans le Jura, les Tornquistes de petite taille récoltés au niveau de la Sous-Zone à Cordatum se distinguent nettement des spécimens plus anciens par une section plus déprimée et sont proches de ceux de la faune de Niort.
De rapporte à cette famille un fragment de Phylloceras qu'il range avec doute dans l'espèce P. plicatum, une espèce dont l'holotype serait une forme de . Nous n'avons pas retrouvé cette morphologie.
En revanche nous avons récolté quatre individus appartenant à cette famille. Un spécimen de grande taille (25 mm) est attribuable sans ambiguïté à l'espèce Sowerbyceras tortisulcatum d'. Trois autres spécimens de très petite taille (inférieure à 10 mm) ne peuvent être déterminés précisément : tours internes de Sowerbyceras ou de Calliphylloceras ?
De dit n'avoir récolté qu'un seul exemplaire de Lytoceras (dont le diamètre atteint 14 mm) qui présente des analogies avec Lytoceras adelae d' mais s'en distingue par une costulation plus flexueuse et la présence de côtes intercalaires. Pour ces deux raisons, il a préféré créer une nouvelle espèce qu'il nomme L. parvulum. Il note aussi que les sutures à ce stade sont simples et que les cloisons se regroupent deux par deux.
Nous avons récolté un individu de 11,5 mm, d'allure très différente par son ombilic très ouvert et ses tours à section circulaire. Nous n'avons trouvé aucune espèce décrite s'accordant à ce spécimen.
Comme dit plus haut, la faune de Saint-Florent indique sans ambiguïté la Zone et la Sous-Zone à Cordatum. Par contre, il est délicat d'être plus précis (partie inférieure ou partie supérieure?), car les informations biostratigraphiques données par les Euaspidoceratinae () et les Cardioceratinae () de Saint-Florent ne sont pas strictement concordantes avec celles provenant des gisements européens connus de même âge.
La faune du "niveau vert" de Zalas (, 1981), constituée de spécimens carbonatés d'assez grande taille, a été datée de l'extrême base de la Sous-Zone à Cordatum ( & , 1992). Chez les Euaspidoceratinae ( et alii, 1994) on constate que : 1) les microconques polonais (g. Mirosphinctes) sont nettement plus grands et 2) le stade bituberculé des macroconques (g. Euaspidoceras) apparaît plus tardivement. Ces deux caractères rapprochent la population polonaise d'Euaspidoceras douvillei (macroconque et microconque), une espèce dont l'acmé se situe dans la Sous-Zone à Bukowskii. Bien qu'il soit assez délicat de comparer les formes pyriteuses de Saint-Florent avec celles carbonatées de Zalas, la population décrite ici semble clairement plus récente.
La faune d'Herznach ( & , 1998) est celle qui se rapproche le plus de la population de Saint-Florent, bien que la variabilité de cette dernière soit moindre. En effet, on note chez les macroconques l'absence du pôle épais correspondant à Euaspidoceras kobyi () ainsi que l'absence chez Euaspidoceras freii des morphotypes spathi et elegans; chez les microconques, le dimorphe de E. kobyi n'a pas été reconnu.
Dans le Jura, région de Crotenay ( et alii, 2000), les spécimens récoltés à Saint-Florent se rapprochent incontestablement de ceux de l'unité 135 datée de la Sous-Zone à Cordatum s.l..
Les rares formes de Neuvizy (Ardennes) datées du sommet de la Sous-Zone à Cordatum (, 1986) sont attribuées uniquement à Euaspidoceras freii et E. ovale.
Par rapport aux spécimens de Zalas ( & , 1992), les spécimens de Niort, tous beaucoup plus petits, présentent un aspect un peu plus évolué dans la mesure où les côtes secondaires sont mieux individualisées et la tuberculisation plus nette. La population de Niort semble donc un peu plus récente.
La faune d'Herznach ( & , 1982) a été datée du sommet de la Sous-Zone à Cordatum. Les formes de Niort présentent de nombreuses similitudes mais on constate que les morphologies 'tenuistriatum' et 'tenuicostatum' sont absentes alors qu'elles sont présentes à Herznach. Les Cardioceratinae de Niort seraient donc légèrement plus anciens. Les mêmes remarques doivent être faites lorsque l'on compare les formes de Niort avec la faune de Neuvizy (équivalente en âge de celle d'Herznach).
Les représentants de ces deux sous-familles indiquent donc soit la base de la Sous-Zone à Cordatum (Cardioceratinae), soit plutôt son sommet (Euaspidoceratinae). Ceci n'est pas illogique. En effet, il est facile d'envisager que des changements morphologiques significatifs, utilisables en biostratigraphie fine, ne soient pas synchrones dans deux groupes d'ammonites phylétiquement très séparés.
Les travaux de (à paraître), effectués dans la partie du Jura où se sont déposées les 'Marnes à Creniceras renggeri', montre que des espèces comme Creniceras crenatum , 'Glochiceras cossmanni' de , Campylites sauvageti de apparaissent dès la Sous-Zone à Costicardia alors que 'Hecticoceras' gourbinei de n'apparaîtrait que dans la Sous-Zone à Cordatum.
Tous ces résultats plaident donc plutôt en faveur de l'hypothèse : 'moitié inférieure de la Sous-Zone à Cordatum'.
Les ammonites récoltées dans ce gisement par l'un d'entre nous () l'ont été de façon exhaustive comme en témoigne la variation des diamètres qui varient de 5mm à 30 mm. Chacun des 4342 individus a été regardé à la loupe afin de détecter la présence de cloisons rapprochées associées ou non à une loge d'habitation partielle ou complète.
Le but de cette observation systématique était de deux ordres : 1) détecter les formes adultes de petite taille correspondant soit à des formes microconques, soit à des macroconques micromorphes ( et alii, 2002) et 2) distinguer les Taramelliceras des Creniceras aux plus petites tailles afin d'avoir un comptage le plus proche possible de la réalité puisqu'en dessous de 5-6 mm ces deux groupes sont vraiment très délicats à séparer.
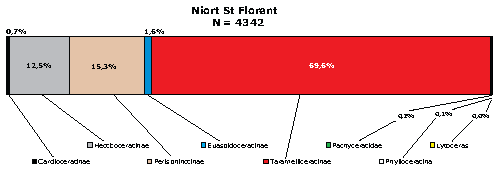
Les résultats obtenus, résumés dans le cartouche de la Fig. 1 ![]() ,
sont les suivants :
,
sont les suivants :
Cardioceratinae : 0.69%
g. Cardioceras [M et m] : N = 30 soit 0.69%
Hecticoceratinae : 12.50%
g. Campylites sauvageti : N = 525 soit 12.09%
g. 'Hecticoceras' gourbinei : N = 18 soit 0.41%
Perisphinctinae : 15.29%
g. Properisphinctes [M et m] : N = 664 soit 15.29%
Euaspidoceratinae : 1.61%
g. Euaspidoceras [M] : N = 25 soit 0.58%
g. Mirosphinctes [m] : N = 24 soit 0.55%
M ou m indéterminés : N = 18 soit 0.41%
soit pour ces deux groupes : N = 67 soit 1.54%
g. Paraspidoceras [M et m] : N = 3 soit 0.07%
Taramelliceratinae : 69.59%
g. Taramelliceras [M] : N = 2103 soit 48.44%
g. Creniceras [m] : N = 880 soit 20.27%
Ta + C : N = 2983 soit 68.70%
g. 'Glochiceras' cossmanni [m] : N = 34 soit 0.79%
g. Sphaerodomites cf. calcarata : N = 4 soit 0.09%
Pachyceratinae : 0.21%
g. Tornquistes sp. [M et m] : N = 9 soit 0.21%
Phylloceratina : 0.09%
g. Sowerbyceras sp. : N = 4 soit 0.09%
Lytoceratina : 0.02%
g. Lytoceras sp. : N = 1 soit 0.02%
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Ce spectre faunique est intéressant dans la mesure où il est très différent de tous les autres spectres connus en Europe occidentale dans la Sous-Zone à Cordatum (, 1986).
L'originalité de la faune de Niort (Fig. 1 ![]() ) est sans conteste liée à la rareté des Cardioceratinae, à l'absence du genre Peltoceratoïdes et à l'extrême abondance du couple Taramelliceras
- Creniceras qui représente, avec 68,71%, un peu plus des 2/3 de la faune ammonitique. En Europe, cette abondance ne nous est connue que dans ce gisement.
) est sans conteste liée à la rareté des Cardioceratinae, à l'absence du genre Peltoceratoïdes et à l'extrême abondance du couple Taramelliceras
- Creniceras qui représente, avec 68,71%, un peu plus des 2/3 de la faune ammonitique. En Europe, cette abondance ne nous est connue que dans ce gisement.
D'après les travaux de l'un d'entre nous ( & , 1979 ; , 1984, 1992), les Taramelliceras sont considérés comme des formes qui nécessitent une 'tranche d'eau' relativement importante puisque leur abondance sur les plates-formes proximales est toujours faible. Mais lorsque les Phylloceratidae (g. Sowerbyceras très dominant) abondent, comme dans le bassin du Sud-Est de la France ( et alii, 1997), on constate durant tout l'Oxfordien inférieur la rareté ou la fréquence toujours faible du genre Taramelliceras et l'absence absolue du genre Creniceras.
Par contre, dans le Jura, les niveaux à ammonites pyriteuses datés de la Sous-Zone à Cordatum (récoltes ) mais situés sous les premiers bancs de calcaires argileux datés de l'extrême sommet de cette sous-zone (, 1986), on constate : 1) que le couple Taramelliceras - Creniceras est fréquent et 2) que les Phylloceratina sont, comme à Niort, très rares. Mais surtout, à la différence de Niort, le genre Peltoceratoides est nettement plus fréquent dans le Jura.
Enfin on constate que dans les gisements riches en Cardioceratinae (Herznach en Suisse, Ardèche, Ardennes) il y a une nette diminution de la diversité spécifique et une diminution drastique du couple Taramelliceras - Creniceras.
L'ensemble de ces constatations permet de proposer que les Taramelliceratinae préfèrent les milieux ouverts mais pas trop profonds (absence des Phylloceratina) alors que les Cardioceratinae colonisent de préférence des milieux moins profonds. Avec l'ensemble de ces données, on peut penser que le gisement de Niort correspondrait à un environnement de plate-forme distale.
Mais il est possible aussi, comme certains le suggèrent, que les masses d'eau situées dans la partie ouest de la France présentent des caractéristiques "physico-chimiques et trophiques" différentes de celles que l'on trouve dans l'Est de la France et le Bassin subalpin. "Profondeur et ouverture du milieu" ne seraient donc pas forcément responsables des différences fauniques observées entre des peuplements datés de la même sous-zone.
L'étude d'une nouvelle population d'ammonites dans la région de Niort - Saint-Florent a permis de retrouver les espèces décrites par de en 1922, d'apporter de nombreuses précisions sur la systématique de ces espèces et de préciser l'âge de cette faune (Zone à Cordatum, Sous-Zone à Cordatum, partie inférieure par les Cardioceratinae, partie sommitale par les Euaspidoceratinae). Enfin, l'abondance des ammonites a permis d'analyser la composition faunique qui indique un milieu de plate-forme distale, contrairement aux gisements de même âge déjà connus en Europe occidentale et qui indiquent tous un environnement plus proximal et moins anoxique. Le 'bassin' du Sud-Est constitue une exception dans la mesure où les Phylloceratidae, et en particulier le genre Sowerbyceras, sont toujours fréquents voire dominants et les Taramelliceratinae toujours rares, voire très rares.
Nous adressons nos plus vifs remerciements aux deux correcteurs, R. et J. , dont les remarques étaient toujours justifiées et les propositions toujours constructives.
D. (2003, inédit).- Diversité et disparité chez les Oppelidés (Ammonites) du sommet de l'Oxfordien inférieur des cantons d'Argovie et de Schaffhausen (Suisse).- Mémoire de DEA, Géosystème, Évolution, Environnement (option Paléontologie évolutive), Université de Bourgogne, Dijon, 50 p.
R., B. & D. (1977).- L'Oxfordien moyen de la région de Sy (Ardennes). Précisions stratigraphiques, paléontologiques et paléogéographiques.- Comptes Rendus des Séances de l'Académie des Sciences, Paris, (série D), t. 285, p. 299-302.
W. (1946-1947).- A monograph on the ammonites of the English Corallian Beds.- Paleontolographical Society, London, 509 p., Pls. A-E + 1-78.
W.J., W.M., B., A.K., R.C., O.H., P.C. & C.W. (1957).- Part L, Mollusca, Cephalopoda, Ammonoidea. In : R.C. (ed.), Treatise on Invertebrate Palaeontology.- Geological Society of America, Boulder, and University of Kansas, Lawrence, vol. 4, 509 p.
A. (1995, inédit).- Les Aspidoceratidae d'Europe occidentale au Callovien supérieur et à l'Oxfordien inférieur.- Thèse Université de Dijon, Centre des Sciences de la Terre, 487 p., 15 Pls.
A., P. & D. (2002).- Parallel Biozonation in the Upper Callovian and the Lower Oxfordian Based on the Peltoceratinae subfamily (Ammonitina, Aspidoceratidae). In : H., K. & A. (eds.), Cephalopods - Present and Past.- Abhandlungen der Geologischen Bundesanstalt, Wien, Band 57, p. 501-507.
A. & R.A. (1998).- Les Euaspidoceratinae (Ammonitina, Aspidoceratidae) d'Herznach (Suisse septentrionale) à la fin de la zone à Cordatum (Oxfordien inférieur).- Eclogae Geologicae Helvetiae, Basel, vol. 91, n° 3, p. 493-512, 3 Pls.
A., P., R. & D. (1994).- Mirosphinctes et Euaspidoceras du niveau vert de Zalas (Pologne) (Oxfordien inférieur, zone à Cordatum) : dimorphes sexuels?.- Bulletin of the Polish Academy of Sciences, Earth Sciences, Varsaw, vol. 42, n° 3, p. 181-205, 1 Pl.
S.S. (1909-1930).- Yorkshire types ammonites.- Published by the author and Wheldon & Wesley, London, vol. 17, 790 Pls.
G. (1887).- Uber die Jurabildungen von Czentochau in Polen.- Beitrage zur Paläontologie Österreich-Hungarns, Wien, Band V, Heft IV, p. 75-171, Pls. 25-30.
E., B., J. & J. (1983).- Notice explicative de la feuille de Niort à 1/50 000.- Notice explicative de la Carte géologique, B.R.G.M., Orléans, n° 142, 35 p.
H. (1853).- Description d'espèces nouvelles de coquilles fossiles du Musée de Besançon.- Journal de Conchyliologie, Paris, t. IV, p. 439-443, 1 Pl.
P., A., P.Y., R. & D. (2000).- Une série oxfordienne de référence, à mi-chemin des régions boréales et téthysiennes (Jura Central, France).- Revue de Paléobiologie, Genève, vol. 19, n° 2, p. 381-397, 3 Pls.
S. & D. (1979).- Réflexions sur la répartition des Ammonoïdés dans l'est et le sud du Bassin parisien au Callovien supérieur (zone à Lamberti) et à l'Oxfordien inférieur (zone à Mariae).- Bulletin du Bureau de Recherches Géologiques et Minières, Orléans, (2ème série), section IV, Géologie générale, n° 2, p. 103-112.
R. (1914).- Études sur les Oppeliidées de Dives et Villers-sur-Mer.- Mémoire de Paléontologie, Mémoires de la Société Géologique de France, Paris, n° 48, 26 p., Pls. I-II.
D., D. & A. (1997).- Les coupes de Thuoux et de Savournon (SE de la France) et la limite CallovienOxfordien.- Geobios, Villeurbanne, n° 30, p. 519540.
A. de (1922).- L'Oxfordien moyen des environs de Niort.- Bulletin de la Société Géologique de France, (4ème série), t. XXI, fasc. 7-9, p. 297-316, Pl. XV.
R.A. & D. (1982).- Les faunes de Cardioceratinae (Ammonoidea) du Callovien terminal et de l'Oxfordien inférieur et moyen (Jurassique) de la Suisse septentrionale : stratigraphie, paléoécologie, taxonomie préliminaire.- Geobios, Villeurbanne, n° 15, fasc. 4, p. 517-571, 13 Pls.
A. (1951).- Stratigraphie und Paläeontologie des oolithischen Eisenerzlagers von Herznach und seiner Umgebung.- Beiträge zur Geologie der Schweiz. Geotechnische, Zürich, Lief. 13, 5, 240 p., 107 Pls.
P. de (1898).- Étude sur les mollusques et brachiopodes de l'Oxfordien inférieur ou zone à Ammonites renggeri du Jura bernois.- Mémoires de la Société Paléontologique Suisse, Genève, vol. 25, 115 p., 6 Pls.
P. de (1900).- Étude sur les mollusques et brachiopodes de l'Oxfordien inférieur ou zone à Ammonites renggeri du Jura Lédonien.- Mémoires de la Société Paléontologique Suisse, Genève, vol. 27, 143 p., 6 Pls.
V. (1928).- Contribution à la connaissance de la faune des marnes à Creniceras renggeri dans la Franche-Comté septentrionale. Étude sur les Oppélidés.- Travaux du Laboratoire de Géologie de la Faculté de Lyon, Mémoire 12, Fasc. 10, 60 p., 3 Pls.
D. (1984).- Ammonites et paléoenvironnements : une nouvelle approche. In : L. & , Paléoécologie. 1er Congrès international de Paléoécologie (Lyon, 1983).- Geobios, Mémoir spécial, Villeurbanne, n° 8, p. 101-107.
D. (1986, inédit).- L'évolution des Cardioceratinae d'Europe occidentale dans leur contexte paléobiogéographique (Callovien supérieur Oxfordien moyen).- Thèse de Doctorat d'État, Université de Dijon, 601 p., 22 Pls.
D. (1992).- Ammonites et paléoprofondeur : les faits, les interprétations.- Paleovox, Lyon, vol. 1, p. 49-68.
D., P., A., J. & Q. (2002).- Very small ammonites (micromorphs) from Lower Oxfordian marls (Mariaezone). In : H., K. & A. (eds.), Cephalopods - Present and Past.- Abhandlungen der Geologischen Bundesanstalt, Wien, Band 57, p. 467-478, 1 Pls.
D. & R.A. (1977).- L'Oxfordien inférieur d'Herznach (canton d'Argovie, Suisse). Précisions paléontologiques et stratigraphiques.- Comptes Rendus des Séances de l'Académie des Sciences, Paris, (série D), t. 285, p. 853-856.
D. & R.A. (1992).- Les ammonites du niveau vert de Zalas (Oxfordien inférieur, Pologne du Sud), condensation ou concentration de faunes.- Bulletin of the Polish Academy of Sciences, Earth Sciences, Varsaw, vol. 40, n° 1, p. 55-65.
C. (1892).- Sur la possibilité d'admettre un dimorphisme sexuel chez les Ammonitidés.- Comptes-Rendus Sommaires des Séances de la Société Géologique de France, Paris, (3ème série), t. XX, n° 14, p. CLXX-CLXXIV.
P., D., J. & J. (1997).- Apparition d'une morphologie scaphitomorphe par miniaturisation chez une ammonite oxfordienne : Scaphitodites scaphitoides , 1853.- Comptes Rendus de l'Académie des Sciences, Paris, (série II), fasc. a, t. 325, N° 4, p. 281-284.
D.F.B. (1966).- Variation and ontogeny of some Oxfordian ammonites: Taramelliceras richei (de ) and Creniceras renggeri (), from Woodham, Buckinghamshire.- Palaeontology, London, vol. 9, part 2, p. 290-311, Pls. 48-52.
P. (1917).- Note sur les fossiles nouveaux rares ou peu connus de l'Est de la France.- Vesoul, 52 p.
L. (1909).- Phylogénie des principaux genres d'ammonoïdés de l'Oolithe (Dogger) et de l'Oxfordien.- Archives des Sciences Physiques et Naturelles, Genève, (série 4), t. 28, p. 611-623.
L. (1911).- Les faciès du Dogger ou Oolithique dans le Jura et les régions voisines.- Georg et Cie, Zürich, 352 p.
Q., D., A., P., M. & V. (1997).- Le contact Callovo-Oxfordien dans les environs de Chaignay : nouvelles données stratigraphiques et paléontologiques.- Bulletin Scientifique de Bourgogne, Dijon, t. 49, p. 45-63.
(1953).- Die Hersumer Schichten im Hildesheimen Jura-Zug.- Geologisches Jahrbuch, Hanover, Band 67, p. 273-360.
R.A. (1981).- Amonity i biostratygrafia dolnego i srodkowego oksfordu okolic Krakowa [Ammonites et biostratigraphie de l'Oxfordien inférieur et moyen des environs de Cracovie].- Thèse [Praca doktorska], AGH Akademia Górniczo-Hutnicza [University of Science and Technology], Wydzial Geologiczno-Poszukiwawczy, Kraków, 123 p., XXV Pls. [en Polonais].
R.A. & D. (2004).- Le dimorphisme chez Perisphinctes (Properisphinctes) cf. bernensis de (Oxfordien inférieur, région de Cracovie, Pologne).- Compte Rendu Paléovol, Paris, vol. 3, n° 3, p. 191-198.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Cardioceras cordatum
figs. 1-8 : figs. 1, 3 & 6 : microconque ? ; figs. 4 & 7 : microconque ; figs. 2 & 5 : macroconque ? ; fig. 8 : macroconque ; fig. 6 : morphe persecans in ; fig. 7 : morphe vertebrale in ; fig. 8 : morphe costicordatum .- Point noir : dernière suture visible ; échelle graphique = 1 cm.
Plate 1
Cardioceras cordatum
figs. 1-8: figs. 1, 3 & 6: microconch ?; figs. 4 & 7: microconch; figs. 2 & 5: macroconch ?; fig. 8: macroconch; fig. 6: morph persecans in ; fig. 7: morph vertebrale in ; fig. 8: morph costicordatum .- Black dot: last visible suture; scale bar = 1 cm.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
figs. 1-7 : Taramelliceras frequens de ; fig. 8 : Taramelliceras frequens de proches de ledonicum de .- Toutes les formes figurées sont des phragmocônes ; échelle graphique = 1 cm.
Plate 2
figs. 1-7: Taramelliceras frequens de ; fig. 8: Taramelliceras frequens de close to ledonicum de .- All the forms figured are phragmocones; scale bar = 1 cm.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
figs. 1-2 : "Hecticoceras" gourbinei de ; figs. 3-7 : Creniceras crenatum .- Point noir : dernière suture visible ; échelle graphique = 1 cm.
Plate 3
figs. 1-2: "Hecticoceras" gourbinei de ; figs. 3-7: Creniceras crenatum .- Black dot: last visible suture; scale bar = 1 cm.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
figs. 1.a-d & 2 : Sphaerodomites cf. calcarata ; fig. 3 : "Hecticoceras" aff. gourbinei de ; figs. 4-6 : Glochiceras ? (ex Cadomoceras) cossmanni de .- Point noir : dernière suture visible ; échelle graphique = 1 cm.
Plate 4
figs. 1.a-d & 2: Sphaerodomites cf. calcarata ; fig. 3: "Hecticoceras" aff. gourbinei de ; figs. 4-6: Glochiceras ? (ex Cadomoceras) cossmanni de .- Black dot: last visible suture; scale bar = 1 cm.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Campylites sauvageti de
figs. 1-9 : figs. 2, 4 et& 8 : pôle Campylites ; figs. 1, 3, 5-7 & 9 : pôle Trimarginites.- Toutes les formes figurées sont des phragmocônes ; échelle graphique = 1 cm.
Plate 5
Campylites sauvageti de
figs. 1-9: figs. 2, 4 & 8: Campylites group; figs. 1, 3, 5-7 & 9: Trimarginites group.- All the forms figured are phragmocones; scale bar = 1 cm.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Properisphinctes cf. bernensis de
figs. 1-8 : fig. 5 : morphe très mince (rare) ; fig. 8 : morphe à rares constrictions (rare).- Toutes les formes figurées sont des phragmocônes ; échelle graphique = 1 cm.
Plate 6
Properisphinctes cf. bernensis de
figs. 1-8: fig. 5: very thin morph (rare); fig. 8: morph with a few constrictions (rare).- All the forms figured are phragmocones; scale bar = 1 cm.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Tornquistes sp.
figs. 1-5.- Point noir : dernière suture visible ; échelle graphique = 1 cm.
Plate 7
Tornquistes sp.
figs. 1-5.- Black dot: last visible suture; scale bar = 1 cm.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
figs. 1-11 : Euaspidoceras freii morphe freii macroconque ; figs. 12-16 : Euaspidoceras freii morphe niedzwiedskii () microconque ; figs. 17-19 : Paraspidoceras sp.- Point noir : dernière suture visible ; échelle graphique = 1 cm.
Plate 8
figs. 1-11: Euaspidoceras freii morph freii macroconch; figs. 12-16: Euaspidoceras freii morph niedzwiedskii () microconch; figs. 17-19: Paraspidoceras sp.- Black dot: last visible suture; scale bar = 1 cm.