![]()
Contents
[Introduction]
[Systematic micropaleontology]
[Miscellaneidae]
[Miscellaneinae]
[Miscellanea]
[Miscellanitinae]
[Miscellanites]
[Ornatononion]
[Miscellanites globularis (, 1978) and Bolkarina aksarayi , 1981]
[Conclusions] [Bibliographic references]
[Figures] [Plates]
[Page with frames] and ... [Plate cross-comparison]
![]()
The Miscellaneidae are divided into two groups of species: forms with a single intercameral foramen and forms with two or a row of multiple foramina. Ten taxa ascribed to this family are revised, amply illustrated and discussed considering both micro- and megalospheric generations. The Family Miscellaneidae is assigned to the Superfamily Nonionacea by reason of their planispiral-involute coiling combined with an interiomarginal position of the foramina. Their combined range covers SBZ zones 2-5 and an area comprising the Central and Western Neotethys including the Pyrenean Gulf. They do not reach the western shores of the Atlantic. Miscellanites meandrinus and Bolkarina aksarayi exhibit extreme morphological features, respectively meandrine alar extensions and expanse chambers. These features are of general interest for the comparative anatomy of the shells of the larger foraminifera in order to understand their biological significance.
Foraminifera; structural analysis of the shells; distribution of intercameral foramina; shallow benthic zones 2-5; meandrine alar extensions; expanse chambers.
L. (2009).- The Paleocene and earliest Eocene foraminiferal Family Miscellaneidae: neither nummulitids nor rotaliids.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2009/06 (CG2009_A06)
La Famille des Miscellaneidae, foraminifères du Paléocène et de l'Éocène inférieur : ni des nummulitidés, ni des rotaliidés.- Les Miscellaneidae, formes lamellaires-perforées à coquille planispiralée involute, sont subdivisées en taxons à un seul foramen en position symétrique par rapport à l'équateur du test (Miscellanea s.str.) et taxons en ayant deux (Ornatononion) ou plus disposés en une rangée intériomarginale (Miscellanites). Dix espèces sont décrites, amplement illustrées et discutées en tenant compte du dimorphisme considérable des générations. La Famille des Miscellaneidae est attribuée comme groupement de grands foraminifères spécialisés à la Superfamille des Nonionacea, caractérisée entre autres par l'enroulement planispiralé-involute du test combiné avec une position intériomarginale des foramens. L'extension biostratigraphique de la famille couvre les biozones SBZ 2-5. La répartition paléogéographique correspond à toute la Néotéthys y compris le Golfe Pyrénéen. L'Atlantique n'est pas franchi ; les Caraïbes restent sans miscellanea. Deux structures extrêmes marquent la famille et sont d'un intérêt particulier pour l'anatomie comparée et fonctionnelle des grands foraminifères: Miscellanites meandrinus présente des extensions alaires méandriformes et Bolkarina aksarayi un test à loges d'expansion en forme de dôme en arrangement bisérié. On retrouve ces deux structures chez d'autres grands foraminifères sans aucune relation systématique ce qui permet d'interpréter ces structures extrêmes comme des analogies. Il doit exister des raisons fonctionnelles pour justifier qu'une telle structure soit répétée plusieurs fois dans des groupes systématiques sans relation mutuelle.
Foraminifères ; analyse structurale des tests ; disposition des foramens ; zones SBZ 2-5 des foraminifères benthiques de plates-formes ; extensions alaires méandriformes ; loges d'expansion.
The Miscellaneidae are a group of index microfossils that are essential for the biozonation of shallow marine deposits of the Paleocene and the earliest Eocene. They have their own discrete mode of architectural design and ornamentation that distinguishes them at the family level from nummulitids and from rotaliids. This paper is intended to be a revision on the species level of the various taxa belonging to the Miscellaneidae in order to lay the foundation for a consistent use of these taxa for zonal biostratigraphy and eventually for biogeography. Significant results in biozonation and in biogeography can be anticipated only when the foraminiferal taxa are examined at the species level. The species can be correctly identified only if parallel evolutionary lineages are recognized by differences in their structural design that remain stable during the time range of the lineage in question. Such differences are usually indicated through the use of different taxonomic names on the generic level.
The identification of larger foraminifera to the species level is difficult and requires experience: most fossil occurrences are in cemented limestones and must be studied in random thin sections. The first, immediate hurdle is to identify the approximate orientation of the shell in the section. Prerequisite is the correct identification of the septa between the chamber lumina in the shell. The shape of the sectioned chamber in relation to the outline of the sectioned shell permits recognition of the mode of chamber arrangement: trochospiral, planispiral involute or planispiral evolute, uni-or multiserial, etc. Random sections of the same taxon and the same generation of a species can not always be recognized. Sections of randomly cut specimens must be selected to obtain all the orientations that are necessary for a correct determination of the architectural design of the shell. Therefore it is important to have enough thin sections of a rock sample to find the sections of the shells in all appropriate orientations.
In bilamellar-perforate larger foraminifera the chamber arrangement, disposition of apertures and the configuration of canal systems are the most important features diagnostic for the identification of genera. The species that constitute these genera are defined by quantitative morphological characters that change with time in one direction. They are interpreted as phylogenetic trends. Their measurement is tricky and often has been attempted with great naivety. The only feature that may be quantified by simple linear measurement is the diameter of the megalospheric proloculus if it is a walled sphere. Other characters like the mode of coiling of successive chambers will not reveal any useful phylogenetic trends based on progressive biological processes, if only simple linear measurements - of the spiral step for instance - are used. However, the human brain is able and used to recognize even individual fellow members of the human species: a photograph of a human face is a means of identifying an individual. The current debate about the biometric data of a passport holder illustrates the complexity of a relevant biometry. In the same way the most accurate method of defining species of larger foraminifera under today's circumstances is the illustration of the sections of a number of specimens at a standard enlargement and orientation. Appropriate numbers of illustrated specimens facilitate the distinction between specific characters and their intraspecific variation through the repeated presentation of the several aspects of the specific characteristics of each individual.
If taxonomic papers are to be kept to a reasonable size the description and discussion of the architecture of larger foraminifera must use many specialized terms to describe their morphology. Here, we use the terms defined, discussed and illustrated by a glossary (, 2006) that is available on the web.
This taxonomic revision of the Miscellaneidae is based on material from the Atlantic shores of western Aquitaine, southern France, from the Subbetic realm in southeastern Spain, from the Adriatic platform and its equivalents in Greece and Turkey, from Egypt, Iran and Oman. Material from the Salt Range and adjoining mountain ranges of Pakistan collected during three expeditions under the guidance of A.A. (Lahore) in 1992, 1993 and 1995 are of particular importance because many samples could be washed. They produced free specimens used for oriented sectioning. Selected series of deposits will be analyzed and published as soon as other groups of larger foraminifera are revised and can be integrated into the zonal system. In particular, the distribution of species belonging to the genera Assilina and Nummulites will be important in the control of correlations based on the Miscellaneidae from the shores of the Atlantic throughout the Mediterranean to the Near and Middle East. Here, we base the biozonal ranges of the revised taxa on the alveolinids and selected associates that occur with the Miscellaneid taxa without discussing the sedimentary or lithostratigraphic details of the their provenance.
The specimens figured in this paper are deposited with supplemental material in the collections of the Museum of Natural History in Basel, Switzerland. Specimens on loan from the British Museum (Natural History) are marked as such in the legends of the plates and are to be sent back to London.
Remarks: and (1987) combine into one family, called the Pellatispiridae, 's Pellatospiridae (1937) and 's Miscellaneidae (in , 1952) of the Superfamily Nummulitacea although the biostratigraphic ranges of the two families are discrete: the Miscellaneidae are restricted to the Paleocene and Earliest Eocene, the Pellatispiridae to the Late Bartonian and the Late Eocene. There are also structural differences between the two families (discussed below) that warrant their separation. Both must be excluded from the Superfamily Nummulitacea de , 1827, and assigned elsewhere in the system.
This subfamily associates all miscellaneids exhibiting a single, interiomarginal, intercameral foramen that is symmetrical with respect to the equatorial plane of the shell.
Miscellanea , 1935, emend. , 1954
Type species: Nummulites miscella d' & , 1853
Generic characters: the shell is lamellar-perforate, the disposition of the chambers planispiral-involute. A single foramen provides direct communication between successive chamber lumina. The foramen consists of a low arch in interiomarginal position and is symmetrical with respect to the equatorial plane as defined by the position of the periphery of the previous whorl.
The chamber bottom is covered by a wall element that produces a kind of carpet extending from the septal flap over all the surface of the previous whorl covered by the new chamber and its alar prolongations. The carpet, called paries proximus, may be attached closely to the previous whorl and therefore be difficult to distinguish from the secondary lamellation of the previous whorl. Often, the paries proximus is attached only to the summits of the pustules that determine the dense ornamentation of the previous whorl and bridges the fissures between them. The widest parts of the interlocular space delimited by the paries proximus and the previous whorl are positioned below the lateral angle of the foramen in the septum. The spiral free part of the paries proximus is called the umbilical plate
(Pl. 5 ![]() ,
figs. 8 & 10;
Pl. 10
,
figs. 8 & 10;
Pl. 10 ![]() , figs.
3-5 & 8). Below the successive septa in a whorl they produce a biumbilical spiral canal that communicates directly with the intraseptal interlocular space. This space is subdivided by radial ribs that produce a simple, radial fan of canals in the septum
(Pl. 8
, figs.
3-5 & 8). Below the successive septa in a whorl they produce a biumbilical spiral canal that communicates directly with the intraseptal interlocular space. This space is subdivided by radial ribs that produce a simple, radial fan of canals in the septum
(Pl. 8 ![]() , fig. 7;
Pl. 10
, fig. 7;
Pl. 10 ![]() , fig. 8).
, fig. 8).
The ornamentation is initiated by a single row of beads aligned over the closed intraseptal interlocular space and flanked by a double row of canal orifices. These early phases of ornamentation can be seen only in penultimate chambers covered by only a few outer lamellas
(Fig. 1 ![]() ). With the addition of more outer lamellas during growth, additional pustules are installed on the peripheral and lateral chamber walls until the initial pattern is completely obscured by a continuous and dense cover of pustules. With the development of the pustular ornamentation the canal systems successively envelops the outer chamber walls until its orifices cover the entire surface of the shell.
). With the addition of more outer lamellas during growth, additional pustules are installed on the peripheral and lateral chamber walls until the initial pattern is completely obscured by a continuous and dense cover of pustules. With the development of the pustular ornamentation the canal systems successively envelops the outer chamber walls until its orifices cover the entire surface of the shell.
The larger-sized members of this family exhibit a considerable dimorphism of generations: the microspheric adults reach at least twice the size of the megalospheric specimens.
Remarks: et alii in 2001 described the architecture of Pellatispira and its consorts in detail. Their architecture is determined by the degree of evoluteness of the planispiral chamber arrangement and by the development of a discrete peripheral structure, the marginal crest, that is as different from that of other groups of larger foraminifera as are the marginal cord of the nummulitid and the retral processes of the elphidiid family.
Axially compressed species of this genus are difficult to separate from the faintly trochospiral Daviesina langhami
(Fig. 2.A-D ![]() ) with a similar, heavily pustular ornamentation. The ornamental pattern of D. langhami, however, begins with a deep feathering of the intraseptal interlocular space. A first generation of pustules develops as a row on each rib that produces the feathering. With the addition of successive outer lamellas the lateral and peripheral chamber walls are covered by a dense mass of pustules like that of Miscellanea. The original equatorial asymmetry of the weakly trochspiral shell in D. langhami is revealed by the asymmetric-triangular foramen with a tooth-like structural element that incompletely subdivides the foramen. The umbilical plates are doubled but also are asymmetric, larger on the ventral side, smaller on the dorsal side of the shell
(Fig. 2.C-D
) with a similar, heavily pustular ornamentation. The ornamental pattern of D. langhami, however, begins with a deep feathering of the intraseptal interlocular space. A first generation of pustules develops as a row on each rib that produces the feathering. With the addition of successive outer lamellas the lateral and peripheral chamber walls are covered by a dense mass of pustules like that of Miscellanea. The original equatorial asymmetry of the weakly trochspiral shell in D. langhami is revealed by the asymmetric-triangular foramen with a tooth-like structural element that incompletely subdivides the foramen. The umbilical plates are doubled but also are asymmetric, larger on the ventral side, smaller on the dorsal side of the shell
(Fig. 2.C-D ![]() ).
).
Miscellanea miscella (d' & , 1853)
Pl. 1 ![]() , figs. 14-20;
Pl. 2
, figs. 14-20;
Pl. 2 ![]() , figs. 1-7;
Pl. 3
, figs. 1-7;
Pl. 3 ![]() ,
figs. 1-15; Pl. 4
,
figs. 1-15; Pl. 4 ![]() , figs. 1-11;
Pl. 5
, figs. 1-11;
Pl. 5 ![]() , figs. 1-11;
Pl. 6
, figs. 1-11;
Pl. 6 ![]() , figs. 8-9.
, figs. 8-9.
1853 Nummulites miscella d' E. & J., p. 345, Pl. 35, fig. 4.a-c.
1916 Siderolites miscella (d' & ). H., p. 38, Pl. XV, fig. 5.
1926 Siderolites miscella (d' & ). W.L.F., p. 498.
1927 Siderolites stampi L.M., p. 278, Pl. 21, figs. 1-8.
1935 Miscellanea miscella (d' & ). J., p. 230.
1937 Miscellanea miscella (d' & ) pars. L.M. & E.S., p. 43, Pl. VI, figs. 2-3, ?5 & 7-8 (non fig. 1 = Daviesina langhami ).
1937 Miscellanea stampi (). & , p. 42, Pl. VI, figs. 4, 6, 9-10 & 17-18.
1941 Miscellanea miscella (d' & ). & , p. 32, Pl. 5, figs. 1-2.
1952 Miscellanea miscella (d' & ). P.J., p. 145, Pl. 35, fig. 6.a-c.
1953 Miscellanea miscella (d' & ). W.S., p. 30, Pl. 1, figs. 3-4.
1954 Miscellanea miscella (d' & ). A.H., p. 72.
1960 Miscellanea miscella (d' & ). W.S., p. 196, Pl. 26, figs. 46.
1969 Miscellanea meandrina (). M., Pl. 59/2, Pl. 60 upper left.
1970 Miscellanea miscella (d' & ). M., p. 96, Pl. 9, figs. 8-10.
1972 Miscellanea miscella (d' & ). E., p. 284, Pl. 4, figs. 1-7.
1976 Miscellanea miscella (d' & ). A., p. 59, Pl. 3/2, figs. 7-8.
1983 Miscellanea miscella (d' & ). A., p. 63, Pl. 43, figs. 14-18.
1988 Miscellanea miscella (d' & ), pars. U., p. 703, Pl. 1, fig. 1; Pl. 2, fig. 1; Pl. 7, fig. 4; Pl. 8, fig. 1; non text-fig. 5; Pl. 3, fig. 1; Pl. 7, ?figs. 1-3, figs. 5-6; Pl. 8, fig. 2.
1988 Miscellanea sp. S. & S., p. 170 upper left, p. 171.
1989 Miscellanaea rhomboidea J. & U., p. 303, figs. 6.a-g.
1998 Miscellanea miscella (d' & ). E., p. 94, Pl. 57, figs. 1-10.
2008 Miscellanea miscella (d' & ). M.K., p. 328, Pl. 6/15, figs. 3-5.
Megalospheric generation: the lenticular shells have an unkeeled but sharp periphery. The chambers are planispiral-involute, with septa that are inclined backward and curved over the outer two thirds of their path towards the periphery in the equator of the shell. The septa between successive alar prolongations are radial. Towards the poles of the shell a group of heavy piles obscures the original arrangement of the external ornamentation. The ratio of equatorial to axial diameter of the shell is 1.3 3.6. At an equatorial diameter of 1.8 mm there are 18 chambers per whorl; at a diameter of 2.4 mm we counted 22 chambers. The megalosphere is spherical, with a diameter of 0.24 0.40 mm. The deuteroconch has a semilunar outline in the equatorial section and embraces the proloculus in axial sections. (1970: Pl. 9, fig. 8) has found a twinned specimen where two comparatively large nepionts, both consisting of a megalosphere followed by a full first whorl, produced a common adult shell with two more whorls.
Microspheric generation: shell of large size, of flattened lenticular shape, with a rounded periphery, covered by numerous pustules that are transformed in the slightly raised polar area of the shell into slender piles of modest dimensions. At a shell diameter of 1 cm, the adult size, there are around 40 chambers per whorl. In equatorial sections the septa are inclined backward and are curved over the distal two thirds of their path to the periphery. Their spacing and curvature are often irregular in the outer, adult whorls. We have not seen any free specimens showing the exterior of the shell with its pattern of sigmoidal sutures separating the long alar prolongations. The adult microspheric shells are covered by narrowly spaced beads and papillae. The slightly raised polar centers of the shell are occupied by a group of piles that are not much thicker than ordinary beads.
Remarks. Miscellanea miscella is the key species for understanding the genus and the whole family. It was described by d' and (1853) from Sind, currently the southeastern province of Pakistan. The types have been lost and the exact location of its original discovery is not known. Material collected by Peter in the 1959's and labeled "Jamshoro College, Haiderabad, Sind", provided the specimens that are closest to the topotypes. The material contained a single microspheric and some megalospheric specimens figured in , 1988. These occur together with Ranikothalia nuttalli, Alveolina vredenburgi (see et alii, 1998) and Assilina jiwani () (see , 1977, fig. 20 E,F), an association characteristic for SBZ 5 (Lower Ilerdian, see et alii, 1998). Megalospheric specimens of similar characteristics and size are found frequently in the Salt Range, northwestern Pakistan. In Egypt (Wadi Araba) the two generations of this Miscellanea are associated. They were described under the name Miscellanea rhomboidea by U. ( & , 1989). In the Salt Range, in Egypt and in the Aquitaine (southwestern France) they are associated with larger foraminifera from SBZ 4 (Late Thanetian). The number of microspheric specimens is too small to make a decision as to whether there is a sufficient morphological difference between the SBZ 5 Miscellanea miscella and the SBZ 4 M. rhomboidea to differentiate them as species. The megalospheric specimens from both stratigraphic levels exhibit a considerable variability that prevents a distinction between the two that anyway are closely related. For the time being the two species are treated as synonyms with a range of SBZ 4-5.
Miscellanea yvettae , 1988
Pl. 7 ![]() , figs. 9-16; Pl. 8
, figs. 9-16; Pl. 8 ![]() , figs. 1-9;
Pl. 9
, figs. 1-9;
Pl. 9 ![]() , figs. 1-12.
, figs. 1-12.
1962 Miscellanea sp. J., Pl. 22, fig. 1.
1988 Miscellanea sp. 2 (). et alii, p. 160, Pl. 26, figs. 1-4.
1988 Miscellanea yvettae U., p. 702, Pl. 1, fig. 2; Pl. 2, fig. 2; Pl. 3, fig. 2; Pl. 6, figs. 1-9.
1988 Miscellanea juliettae villattae U., p. 702, Pl. 1, fig. 3; Pl. 2, fig. 3; Pl. 5, figs. 1-8.
1998 Miscellanea juliettae . E., p. 93, Pl. 54, figs. 1-10.
Microspheric generation: the thickly lenticular to vertebra-shaped shells are heavily ornamented with a dense cover of piles and pustules. The adult shell has a diameter to thickness ratio of 1.5 or less. The vertebra-shape is produced by a doubling of the periphery in the ultimate and penultimate whorls. Between the two peripheries the chamber walls lace the shell at the equator up to the point at which these walls touch the previous whorl and thus divide the chambers in two lateral parts. Therefore, the equatorial section of the shell is a spiral that gets much narrower or loses the chamber lumen completely in the last, adult whorls
(Pl. 7 ![]() , fig. 9). The lacing of spiral shells at the equator can be observed in other species as well, in particular in many specimens of Daviesina langhami ,
1954, in which the two peripheries are unequal and often oblique in respect to the equatorial plane
(Pl. 1
, fig. 9). The lacing of spiral shells at the equator can be observed in other species as well, in particular in many specimens of Daviesina langhami ,
1954, in which the two peripheries are unequal and often oblique in respect to the equatorial plane
(Pl. 1 ![]() , figs. 3-4). At the adult diameter of about 2.8 mm there are 25 chambers per whorl.
, figs. 3-4). At the adult diameter of about 2.8 mm there are 25 chambers per whorl.
Megalospheric generation: the shells are lenticular with a diameter to thickness ratio of about 1.8. A doubling of the periphery is rare and where it is present at all it is restricted to few last
chambers (Pl. 9 ![]() , fig. 4). At an adult shell diameter of 1.6 mm there are 19-21 chambers per whorl. The ornamentation of the test seen in sections obliquely tangential to the test
(Pl. 9
, fig. 4). At an adult shell diameter of 1.6 mm there are 19-21 chambers per whorl. The ornamentation of the test seen in sections obliquely tangential to the test
(Pl. 9 ![]() , fig. 5) is dominated by a single row of beads aligned over the intraseptal interlocular space. At the poles of the shell, these are transformed into a group of stout piles. The protoconch has a diameter of 0.16 to 0.24 mm. The hemispherical deuteroconch exhibits a similar volume and together with the protoconch constitutes a biconch embryo that is followed by a third chamber with much smaller volume
(Pl. 7
, fig. 5) is dominated by a single row of beads aligned over the intraseptal interlocular space. At the poles of the shell, these are transformed into a group of stout piles. The protoconch has a diameter of 0.16 to 0.24 mm. The hemispherical deuteroconch exhibits a similar volume and together with the protoconch constitutes a biconch embryo that is followed by a third chamber with much smaller volume
(Pl. 7 ![]() , figs. 10-15).
, figs. 10-15).
Remarks. Miscellanea yvettae is represented by numerous free specimens in the Petites Pyrenees (Aquitaine, southwestern France). In the Montes Obarenes (Spanish Western Pyrenees) U. (1988) described them from thin sections of cemented carbonate rock as a particular subspecies under the name M. juliettae villattae. In my view, this is not justified: there are no significant differences in their morphology. There is also a little taxonomic problem: s two subspecies of Miscellanea, M. juliettae pfenderae and M. juliettae villattae were erected without designation of a nominal subspecies ( i.e. M. juliettae juliettae). Therefore, the subspecific names are not available for upgrading to species level.
Miscellanea yvettae was found in the Pyrenees in association with Ranikothalia sindensis, a nummulitid species characteristic for SBZ 3 (Lower Thanetian). In the region of Wadi Araba (Egypt) it is associated with Plumokathina subsphaerica ()
Miscellanea juliettae , 1988
Pl. 1 ![]() , figs. 21-27; Pl. 10
, figs. 21-27; Pl. 10 ![]() , figs. 1-20;
Pl. 11
, figs. 1-20;
Pl. 11 ![]() , figs. 1-11.
, figs. 1-11.
1972 Miscellanea sp. G., J.J. & L.E., p. 379, Pl. 1, fig. 2.
1988 Miscellanea juliettae pfenderae U., p. 700, Pl. 1, fig. 4; Pl. 2, fig. 4; Pl. 3, figs. 4/1 & 4/2; Pl. 4, figs. 1-8, non figs. 9-10 (= ? Daviesina langhami).
1999a Miscellanea miscella (d' & ). M. & A.A., p. 138, Pl. 1, figs. 1-4.
1999b Miscellanea miscella (d' & ). M. & A.A., p. 192, Pl. 1, fig. 1.
2001 Miscellanea yvettae . E. et alii, pars? Pl. 4, figs. 16-17, fig. ?15.
Microspheric generation: small shells in comparison with those of Miscellanea miscella, with a thickly lenticular shape. The lens is flattened at the poles or even slightly depressed (Pl. 10, fig. 19), always with a sharp periphery. Doubling of the periphery is exceptional. At the adult diameter of about 2.8 mm, there are 23 chambers in the last whorl. The ornamentation is delicate and dominated by small beads. At the polar flattening or depression, they are tranformed into a bundle of slender piles that mark the shell axis. The growth spiral is quite loose; the last volution expands rapidly.
Megalospheric generation: lenticular shells with slightly convex to flattened polar areas with a diameter to thickness ratio of 2.3 to 2.8. The equatorial spiral does not expand much in the adult whorls. At a diameter between 1.6 and 1.8 mm, the adult size, we count 18-22 chambers per whorl. The megalospheric embryo is a biconch with an axial diameter of 0.08 to 0.16 mm. The umbilical plate separates a comparatively large spiral canal from the chamber lumen. Some loopholes in the umbilical plate are present below the foramen. The intraseptal interlocular space opens rapidly and houses a broad supplemental skeleton with an enveloping canal system. In slightly oblique equatorial sections the chambers in a whorl produce a series of pointed arches
(Pl. 10 ![]() , fig. 18;
Pl. 11
, fig. 18;
Pl. 11 ![]() , fig. 12).
, fig. 12).
Remarks. Miscellanea juliettae is a good representative of the genus with all its characters but it is significantly smaller than, M. miscella, about half its size, and with more delicate walls. Compared to M. yvettae at similar diameters the number of chambers per whorl is also similar. Both smaller species are associated with larger foraminifera from strata of SBZ 3 (Thanetian). They may represent two ecophenotypes of the same epoch found in the Pyrenees, in the area of Wadi Araba (Egypt) and near Dandot village in the Salt Range, Pakistan.
Miscellanea dukhani , 1954
Pl. 6 ![]() ,
figs. 1-7; Pl. 7
,
figs. 1-7; Pl. 7 ![]() , figs. 1-8.
, figs. 1-8.
1954 Miscellanea miscella var. dukhani A.H., p. 73, ül. 14, figs. 2-6.
2008 Miscellanea miscella var. dukhani , 1954. M.K., p. 128, Pl. 6/15, fig. 9.
Microspheric generation: the shells are much compressed to form a disc (diameter to thickness ratio about 6.5) with approximately plane-parallel lateral surfaces. These are covered by numerous pustules that maintain their small diameter even at the poles of the shell. In equatorial sections the chambers are curved backward over most of their radial extension. and are closely spaced. There are about 40 of them per whorl at an adult shell diameter of 5 mm. The periphery is unkeeled, narrowly rounded in axial sections.
Megalospheric generation: shells flatly lenticular, with lateral surfaces covered by numerous pustules. The septa are much inclined and closely spaced, like those of the microspheric generation. The diameter of the proloculus is 0.32 mm.
Remarks. The material at our disposal is is but 7 specimens from Qatar
(Pl. 7 ![]() , figs. 1-8) on loan from the British Museum of Natural History under the
number P 40151. They are labeled as 's 1954
topotypes, from the "highest zone of the Paleocene". Their preservation is rather poor and does not permit a verification of all the structural features that characterize the genus. In addition, a few specimens of both generations were found in samples from the Salt Range (Pakistan) associated to Daviesina langhami and Ranikothalia nuttalli of SBZ 4-5 zones.
, figs. 1-8) on loan from the British Museum of Natural History under the
number P 40151. They are labeled as 's 1954
topotypes, from the "highest zone of the Paleocene". Their preservation is rather poor and does not permit a verification of all the structural features that characterize the genus. In addition, a few specimens of both generations were found in samples from the Salt Range (Pakistan) associated to Daviesina langhami and Ranikothalia nuttalli of SBZ 4-5 zones.
Miscellanitinae, new subfamily
In this subfamily, all miscellaneid genera with multiple intercameral foramina are united. Their apertures are a single interiomarginal row in a comparatively low chamber. In an analogy with the elphidiids, forms with two symmetric foramina are grouped with the forms with multiple foramina rather than with those having a single intercameral foramen: Porosononion and Cribroelphidium versus Haynesina ( et alii, 2001).
Miscellanites, new genus
Type species: Miscellanea iranica , 1983.
Diagnosis: globular or subglobular shells formed by a planispiral-involute chamber arrangement. The globular shape of the shell is linked with a single row of multiple interiomarginal foramina
(Pl. 13 ![]() , fig. 12) that tend to facilitate the formation of low but elongate chambers reaching from pole to pole
(Pl. 13
, fig. 12) that tend to facilitate the formation of low but elongate chambers reaching from pole to pole
(Pl. 13 ![]() , fig. 7), as in the alveolinids. The free, outer surface of the chamber walls are covered by a simple and shallow enveloping canal system that may develop a single generation of slender pustules between the orifices of the enveloping canals. The megalosphere is more or less spherical followed by a deuteroconch of similar size and shape.
, fig. 7), as in the alveolinids. The free, outer surface of the chamber walls are covered by a simple and shallow enveloping canal system that may develop a single generation of slender pustules between the orifices of the enveloping canals. The megalosphere is more or less spherical followed by a deuteroconch of similar size and shape.
The bottom of the chamber is covered by a paries proximus that extends from the septal flap into the alar prolongations. The paries proximus is loosely attached to the previous whorl on the summits of the relief that furnishes the ornament of the previous whorl. As in spherical alveolinids, the umbilicus is restricted to the polar region of the shell and consequently the spiral plexus of the canal system is very narrow and difficult to see in ordinary thin-sections. The septum is pierced by a single row of foramina with a small diameter that may be even smaller than the free space between the paries proximus and the previous whorl
(Pl. 12 ![]() , fig. 6;
Pl. 13
, fig. 6;
Pl. 13 ![]() , fig. 12;
Pl. 16
, fig. 12;
Pl. 16 ![]() , figs.
6-7 & 11). The elphidiid architecture ( et alii,
2001) illustrates the narrow biumbilical spiral structure in planispiral involute shells with a single row of apertures. The strong proximal inclination of the septa in Miscellanites makes it difficult to demonstrate the multiple apertures.
, figs.
6-7 & 11). The elphidiid architecture ( et alii,
2001) illustrates the narrow biumbilical spiral structure in planispiral involute shells with a single row of apertures. The strong proximal inclination of the septa in Miscellanites makes it difficult to demonstrate the multiple apertures.
Miscellanites iranicus (, 1983)
Pl. 12 ![]() , fig. 1-6;
Pl. 13
, fig. 1-6;
Pl. 13 ![]() , figs. 1-12.
, figs. 1-12.
1972 Pellatispirella cf. antillea . G., J.J. & L.E., p. 379, Pl. 1, fig. 4.
1983 Miscellanea iranica A., p. 63, Pl. 44, figs. 1-21; Pl. 45, figs. 1-2 & 14-15, non Pl. 44, fig. 22.
1997 Miscellanea agriensis E., p. 81, Pl. 6, figs. 1-11.
1998 Miscellanea agriensis . E., p. 95, Pl. 55, figs. 1-8.
2004 Miscellanea agriensis . E., p. 70, Pl. 64, figs. 1-8.
Microspheric generation: spherical to ovoid tests with a diameter up to 3.5 mm. The diameter to thickness ratio is only 1.1. The chambers are arranged in a simple involute planispiral. They are low and short, almost isometric in an equatorial section. At a diameter of 2.8 mm there are 24 chambers in the last whorl. The path of the septa from pole to pole follows closely the corresponding axial plane. The spiral of the chambers in the equatorial plane grows in both generations at an even step shortening slightly in the last whorls. The polar region is marked by few, stout piles that start their growth in the narrow, tiny first whorls. The supplemental skeleton is quite simple and produces an enveloping canal system with uniform, short radial canals feeding uniformly spaced canal orifices distributed over the whole surface of the test.
Megalospheric generation: ovoid tests with a planispiral-involute chamber arrangement. The diameter to thickness ratio varies fom 1.2 to 1.3. The periphery of the test is rounded, the polar area marked by a large group of stout piles that may occupy about a third of the lateral surface of the shell. There are 4 to 6 such piles in an axial section. They originate at the surface of the megalosphere
(Pl. 13 ![]() , figs.
1-2 & 6) and restrict the alar prolongations. The spherical megalosphere present a diameter varying from 0.20 0.28 mm. At an equatorial diameter of 1.8 mm there are 21 chambers in the last whorl.
, figs.
1-2 & 6) and restrict the alar prolongations. The spherical megalosphere present a diameter varying from 0.20 0.28 mm. At an equatorial diameter of 1.8 mm there are 21 chambers in the last whorl.
Remarks. The material of this species originates from a section of the Kuh-e-Kargan near Kermanshah in the Iranian Zagros ranges collected by J. around 1968. The rich sequence of Paleocene larger foraminifera represents SBZ zones 3 to 5 or 6 (Thanetian to Basal Ilerdian) and will be described elsewhere. At Kuh-e-Kargan, Miscellanites iranicus is associated to Glomalveolina primaeva, Fallotella alavensis, Dictyoconus turriculus and "Taberina" daviesi, index fossils for the SBZ 3 (Lower Thanetian).
Miscellanites meandrinus (, 1861)
Pl. 14 ![]() , figs. 1-14;
Pl. 15
, figs. 1-14;
Pl. 15 ![]() , figs. 1-14.
, figs. 1-14.
1861 Alveolina meandrina H.J., p. 251, Pl. 17, figs. 4.a-f.
1951 Miscellanea meandrina (). Y., p. 43.
1954 Miscellanea meandrina (). A.H., p. 74, Pl. X, fig. 1-14, Pl. XI, fig. 12.
1975 Miscellanea meandrina (). E. & Ö., p. 193, Pl. 1, figs. 1-6; Pl. 2, figs. 1-4.
1983 Miscellanea iranica (pars). A., p. 63, Pl. 44, fig. 22.
1989 Miscellanea meandrina (). J. & U., p. 307, fig. 7.a-c.
1998 Miscellanea ? meandrina (). E., p. 99, Pl. 59, figs. 1-6; Pl. 60, figs. 1-10.
2008 Miscellanea aff. juliettae . et alii, Pl. 6, fig. 7; p. 134, Pl.
2008 Miscellanea meandrina (). M.K., p. 328, Pl. 6/15, figs. 6-8.
There can be little doubt that Miscellanites meandrinus from SBZ 4 is to be interpreted as direct phylogenetic successor of M. iranicus in SBZ 3. As in M. iranicus, this species is characterized by large ovoid shells with a planispiral-involute arrangement of numerous, very low and short chambers. The chambers of the later whorls of the microspheric generation lengthen their extension from pole to pole by replacing their linear shape with a meandrine pattern. In the megalosheric generation, the last chambers may be deformed to a simple chevron shape. The megalosphere is particularly large, varying from 0.24 to 0.40 mm in diameter. The deuteroconch is semilunar in axial section and has a volume similar to that of the next succeeding spiral chambers. Both generations are also characterized by their supplemental skeleton that houses an enveloping canal system and provides the chambers with outer walls as thick or even thicker than the corresponding chamber lumen. The microspheric generation is easy to identify by its size and the meandrine pattern of the chambers in the last whorls. The megalospheric generation is distinguished from Miscellanea miscella A by its rounded periphery and by the much tighter outer whorls.
Miscellanites meandrinus occurs in Aquitaine (southwestern France) in association with Assilina yvettae, Nummulites katari, Dictyokathina simplex and Miscellanea miscella. In Wadi Araba, Egypt, and in Turkey it is associated with a similar fauna of larger foraminifera all in the SBZ 4 (Upper Thanetian) faunal zone.
Miscellanites primitivus (, 1983)
Pl. 16 ![]() , figs. 1-24;
Pl. 17
, figs. 1-24;
Pl. 17 ![]() , figs.
1-16.
, figs.
1-16.
1983 Miscellanea primitiva A., p. 61, Pl. 42, figs. 8-16.
1998 Miscellanea sp. 2. J. et alii, Pl. 4, figs. 6-7.
2008 Miscellanea aff. iranica . J. et alii, p. 134, Pl. 6, fig. 6.
Exceptionally well preserved, free specimens of a small Miscellanites were discovered in Narp, a locality of SBZ 3 age in western Aquitaine, southwestern France). I identified them as Miscellanites because of their multiple foramina and as M. primitivus () on the basis of their relatively small size and their ovoid shape. Their heavy piles in the polar area restrict the length of the alar prolongations almost to a point at which the shell whorls become evolute. Their exoskeleton provides the free chamber walls with a comparatively deep enveloping canal system. Many axial sections show a conspicuous change from an angular to a rounded periphery in the last whorl.
The microspheric generation attains an equatorial diameter of 1.5 mm, the megalospheric generation 1mm. At these diameters, there are 16 and 14 chambers, respectively, in the last whorl. The septa are radial and straight at the equator, moderately curved and inclined backward in lateral areas. Megalospheric forms have a single row of six interiomarginal foramina
(Pl. 16 ![]() , figs. 6-7). There are heavy umbilical plates separating the chamber lumen from a comparatively spacious spiral canal
(Pl. 16
, figs. 6-7). There are heavy umbilical plates separating the chamber lumen from a comparatively spacious spiral canal
(Pl. 16 ![]() , figs. 8-10,
16 & 24). The megalospheres have a diameter of only 0.06 to 0.1 mm.
, figs. 8-10,
16 & 24). The megalospheres have a diameter of only 0.06 to 0.1 mm.
Miscellanea primitiva has been recognized sofar in Iran, in the Apulian Domain and on the shores of the Paleocene Atlantic in southwestern France. Its range seems to be limited to the zone SBZ 3. However, the unconspicuous shells of this taxon have probably been overlooked in many places and may have a larger range in time and space.
Miscellanites minutus (, 1983)
Pl. 18 ![]() , figs. 1-21;
Pl. 19
, figs. 1-21;
Pl. 19 ![]() , figs. 1-11.
, figs. 1-11.
1983 Miscellanea minuta A., p. 62, Pl. 43, figs. 1-13.
2008 Miscellanea aff. juliettae . et alii, p. 134, Pl. 6, figs. 8-9.
2008 Miscellanea cf. juliettae . et alii, p. 134, Pl. 4, figs. 4-5.
The rare microspheric forms are lenticular, with a sharp periphery and attain a maximum diameter of 2.2mm. The diameter to thickness ratio is around 2.0. The shell is planispiral and clearly involute, with sigmoidal septal sutures
(Pl. 18 ![]() , fig. 13). In a restricted area around the poles of the shell there is a group of slender piles that bound a considerable number of funnels of the umbilical canal system
(Pl. 18
, fig. 13). In a restricted area around the poles of the shell there is a group of slender piles that bound a considerable number of funnels of the umbilical canal system
(Pl. 18 ![]() , figs.
7 & 10).
, figs.
7 & 10).
The megalospheric forms are lenticular, about 1.4 to 1.5 mm in maximum diameter. At this diameter we count about 25 rather isometric chambers in the last whorl. There are about 3 piles visible in axial sections. Four low slits in an interiomarginal position constitute the intercameral connection. The umbilical plate coats the entire floor of the chambers. The only open space between the chamber floor and the previous whorl is a narrow spiral canal. The megalosphere is small; its diameter is never more than 0.1 mm.
Miscellanites minutus is distinguished from M. primitivus by a greater number of chambers per whorl, a sharper periphery, a more compressed megalospheric shell and by a lesser number of slitlike foramina.
M. minutus is known only from several localities in Iran where it is associated with Glomalveolina primaeva, Dictyoconus turriculus and other index fossils of SBZ 3.
Type species: Nonion? ornatum van in , 1982 = Ornatononion moorkensii n.sp.
Derivatio nominis: in allusion to the ornament on the nonionid shell.
Diagnosis: bilamellar-perforate, planispiral-involute, bi-umbilicate test covered by an ornamentation consisting of closely spaced, delicate, imperforate or sparsely perforate pustules thickening in an adaxial direction. Both umbilici are crambed with piles.
Apertures were not seen. The intercameral foramina are two, symmetrical, interiomarginal, low arches flanking the periphery of the previous whorl. During ontogeny, the two foramina appear in early whorls of the shell but we do not know if the proloculus already has two apertures.
The two intercameral foramina are the most important diagnostic feature. They can be recognized in axial or transverse sections if the section runs tangentially through the basal part of a septum. In more or less centered oblique sections, the presence of two foramina is shown by interruptions of the septum in the adaxial, but not in the equatorial sectors, where interruptions would be present if the aperture were a single equatorial opening. There are no lips.
A strong umbilical plate extends from the umbilical end of the foramen towards the axis of the shell and produces a cover for the alar chamber floor. The umbilical plate is attached to the previous whorl below the foramen and below the tips of the chamber wings
(Pl. 20 ![]() , figs. 8-9). The free-standing part of the umbilical plate delimits a low, spiral umbilical space communicating with narrow intraseptal and vertical umbilical spaces both opening to the ambient environment in and around the ubilical area on either side of the shell. In the equatorial zone there are no interlocular spaces, the septal flap reaching the base of the apertural face and touching the previous whorl.
, figs. 8-9). The free-standing part of the umbilical plate delimits a low, spiral umbilical space communicating with narrow intraseptal and vertical umbilical spaces both opening to the ambient environment in and around the ubilical area on either side of the shell. In the equatorial zone there are no interlocular spaces, the septal flap reaching the base of the apertural face and touching the previous whorl.
Differential diagnosis: Ornatononion is distinguished from Miscellanea mainly by the two foramina in each septum and its much more delicate ornamentation. In elphidiids, we observe a similar phenomenon: two apertures characterize Porosononion ( et alii, 2001), a planispiral-involute smaller benthic form.
Ornatononion is assigned here to the Subfamily Miscellanitinae because of its truly planispiral architecture comprising a symmetrical pair of umbilical plates in each chamber, its spiral umbilical interlocular space communicating with vertical umbilical canals opening between umbilical piles to the ambient environment and the lack of a horizontal subdivision of the umbilical cavities. The absence of a marginal cord separates this group from all true nummulitids.
Ornatononion moorkensii, n.gen. n.sp.
Pl. 20 ![]() , figs. 1-13;
Pl. 21
, figs. 1-13;
Pl. 21 ![]() , figs. 1-16.
, figs. 1-16.
1946 Nonion ornatum van R.C., p. 44, Pl. 4, figs. 7-10.
1962 "petites" Miscellanea. J., Pl. 22, fig. 2.
1964 Nonion ornatum van . P., Pl. 2, figs. 3.a-b.
1982 Nonion ornatum van . T., p. 98, Pl. 9, fig. 2.
Derivatio nominis: in honor of Thierry (Latin: Moorkensius) for his interest and engagement in foraminiferal micropaleontology.
Diagnosis: specimens with the architecture of Ornatononion, as defined above, presenting a thickly lenticular test with flattened poles, covered by pustules and pile-heads in the bi-umbilical, axial zone. The ratio of equatorial to axial diameter ranges from 2.6 to 1.8 in the adult that attains an equatorial diameter of 1.5 to 1.7 mm. Since the last whorl of the test is comparatively fragile, it may often have been broken away prior to its final deposition in the encasing sediment. Therefore many small specimens have significantly lower diameter-thickness ratios. The periphery of the shell is rounded to bluntly angular, never keeled.
In axial section, 3 to 5 piles fill the umbilical cavity. Adult specimens have 18 to 23 chambers. The septa are radial, sharply bent backward near their periphery as they are in the operculinids, but this occurs only in a comparatively narrow equatorial zone of the shell while in the lateral zones the septum is straight and only slightly inclined backward. Some septa may be oblique in respect to the axial plane. Therefore, axial or transverse sections parallel to the axial plane may show only one half of the septum and only one of the two laterally-positioned foramina that characterize the
genus (Pl. 20 ![]() , figs. 9 & 13). The proloculus is small, only 0.06 mm in diameter. Therefore, a dimorphism of generations can not be demonstrated. This small size might present a case similar to that of the Eocene alveolinas with a threshold value of proloculus diameter around 0.11 mm, below which no dimorphism can be detected (,
1963).
, figs. 9 & 13). The proloculus is small, only 0.06 mm in diameter. Therefore, a dimorphism of generations can not be demonstrated. This small size might present a case similar to that of the Eocene alveolinas with a threshold value of proloculus diameter around 0.11 mm, below which no dimorphism can be detected (,
1963).
Differential diagnosis: O. minutus must be distinguished from Miscellanites primitivus , 1983, of similar size by its two symmetrical intercameral foramina, by its much more delicate ornamentation and by its distinctly more involute chambers. M. primitiva, in our view, is not a Miscellanea because of its multiple foramina in a basal row.
Material and distribution: about 30 free specimens sectioned in various defined directions and many more in random thin sections of cemented limestones. This material comes from the Vaquemorte, Buholoup, Ruisseau de Gosse and Sabarat localities (all in the Petites Pyrénées, southwestern France), Nouts near Pau (southwestern France) and Mons (Belgium, , 1982). All ocurrences are dated by the accompanying fauna and/or by their stratigraphic position in relation to typical SBZ 3-faunas for they are significantly older, i.e. SBZ 2.
Remarks: this species was already in use in 1962 under the informal name "petites Miscellanea", as an index fossil for the so-called Montian in the Petites Pyrénées by J. , the foremost expert on the stratigraphy of the Paleocene of this region. A few isolated specimens from a borehole on Montaimé near Mons available from Yvette 's collection permitted the establishment of the identity of these specimens with ' (1982) Nonion ornatum van , 1946. The species name is not available because it is a homonym of Nonionina ornata , 1856. By creating the new generic name Ornatononion, we acknowledge the previous work done with these difficult small benthics.
Miscellanites globularis (, 1978) and Bolkarina aksarayi , 1981
These two taxa are treated here under a common heading but under different names. That they are but two generations of a single species is strongly supported by the fact that in the field they are always found together. However, the structural details of Bolkarina are known only in part (see below). Two separate names must be used to designate the A- and B-forms until the riddle is solved through the study of appropriately preserved specimens of Bolkarina, yet to be found.
Miscellanites globularis (, 1978)
Pl. 22 ![]() , figs. 1-24;
Pl. 23
, figs. 1-24;
Pl. 23 ![]() , figs. 7-13.
, figs. 7-13.
1978 Miscellanea globularis A., p. 61, Pl. 12, figs. 10-20.
1983 Miscellanea globularis . A., p. 61, Pl. 42, figs. 1-5, 7, non fig. 6.
1998 Miscellanea? globularis . E., p. 97, Pl. 58, figs. 1-8, 10 & 12-14.
Megalospheric shells spherical, produced by chambers arranged in an involute planispiral 1mm or less in diameter. At a maximum diameter there are 12 to 15 chambers in the last whorl. The chambers are isometric in the equatorial plane, with almost radial septa. Towards the poles the septa are progressively inclined backward. In some specimens we see a chevron-shaped deformation of the chambers that is symmetrical in respect to the equatorial plane
(Pl. 20 ![]() , fig. 11). The proloculus is spherical with a flattened surface of the wall separating a large hemispherical deuteroconch from the protoconch. There is a thin umbilical plate that adheres to the previous whorl. A spiral canal was not seen but a few funnels exist between some of the piles in the narrow polar region. The supplemental skeleton covering the free chamber walls has a shallow enveloping canal system, which provides the surface of the shell with a cover of canal orifices in addition to the pores.
, fig. 11). The proloculus is spherical with a flattened surface of the wall separating a large hemispherical deuteroconch from the protoconch. There is a thin umbilical plate that adheres to the previous whorl. A spiral canal was not seen but a few funnels exist between some of the piles in the narrow polar region. The supplemental skeleton covering the free chamber walls has a shallow enveloping canal system, which provides the surface of the shell with a cover of canal orifices in addition to the pores.
Bolkarina aksarayi , 1981
Fig. 3 ![]() ;
Pl. 21
;
Pl. 21 ![]() , figs. 1-6;
Pl. 22
, figs. 1-6;
Pl. 22 ![]() , figs.
1-9;
Pl. 23
, figs.
1-9;
Pl. 23 ![]() , figs.
1-5; Pl. 24
, figs.
1-5; Pl. 24 ![]() , figs.
1-9;
Pl. 25
, figs.
1-9;
Pl. 25 ![]() , figs.
1-5.
, figs.
1-5.
1981 Bolkarina aksarayi E., p. 76, Pls. 1-3.
1987 Bolkarina aksarayi . A. & H., p. 681, Pl. 801, figs. 1-5.
1988 Bolkarina sp. K. et alii, p. 157, Pl. 25, figs. 1-2.
1998 Bolkarina aksarayienis . E., p. 91, Pl. 53, figs. 1-5.
2004 Bolkarina aksarayensis . E., p. 55, Pl. 49, figs. 1-3; Pl. 50, figs. 1-4; Pl. 51, figs. 1-6.
Possibly microspheric large discoidal shells with a sharp to rounded unkeeled periphery. The center of the disc exhibits a nepiont of 19 to 20 planispiral-involute chambers. They provide the disc with a central thickening that covers the one and a half involute whorls of the spiral nepiont. The proloculus is small attaining a diameter of 0.08 mm, a rather large size for a microsphere. The walls of the nepiont are thick, heavily ornamented by a group of piles in the polar area.
The ornate planispiral-involute nepiont is the most important argument supporting the attribution of Bolkarina to the
Family Miscellaneidae. The structure of the adult growth stage is not yet fully understood: to distinguish the lamellar-perforate chamber wall from the shell cavities is difficult without the aid of shells with an infill of opaque sediment that are yet to be found.
Apparently the multiple layers of the disc hamper the filling of the chambers
with sediment before the shell is finally interred. However, there is a main layer of annular chambers
(Fig. 3 ![]() ). These alternate in position in respect to the equatorial plane as in Dicyclina
(Pl. 24
). These alternate in position in respect to the equatorial plane as in Dicyclina
(Pl. 24 ![]() , fig. 8). The annular chambers maintain a wide free space in preseptal position
(Pl. 24
, fig. 8). The annular chambers maintain a wide free space in preseptal position
(Pl. 24 ![]() , fig. 9). The septa between successive annuli are oblique with respect to the equatorial plane and bear multiple foramina
(Pl. 25
, fig. 9). The septa between successive annuli are oblique with respect to the equatorial plane and bear multiple foramina
(Pl. 25 ![]() , fig. 4). Their number and pattern of distribution is unclear.
, fig. 4). Their number and pattern of distribution is unclear.
There is a one-sided lateral subdivision of the annular chambers. Their radial alignment is continuous in successive annuli on one or the other side of the equator, alternating on either side of the equatorial plane. These radial subdivisions appear to be transformed into lateral walls that produce a pile of dome-shaped expanse chambers
(Pl. 23 ![]() , figs.
2 & 5-6). Thus, the main annulus communicates with the expanse chamber by a restriction of the annular chamber lumen to a radial passage
(Pl. 24
, figs.
2 & 5-6). Thus, the main annulus communicates with the expanse chamber by a restriction of the annular chamber lumen to a radial passage
(Pl. 24 ![]() , fig. 3). The transition from the nepionic planispiral-involute, uniserial chambers to the neanic stage with biserial annuli is not clear. In particular we do not understand what foraminal pattern is required to produce this structural development.
, fig. 3). The transition from the nepionic planispiral-involute, uniserial chambers to the neanic stage with biserial annuli is not clear. In particular we do not understand what foraminal pattern is required to produce this structural development.
Successive dome-shaped expanse chambers produce the lateral skeleton of the disc that has irregular, vermicular intersections where the section cuts successive layers of the dome. Successive dome chambers communicate by series of small foramina that appear to be aligned in an interiomarginal position all along the vermicular sutures. In addition there seems to be a canal system
(Pl. 23 ![]() , fig. 3;
Pl. 24
, fig. 3;
Pl. 24 ![]() , fig. 2) arranged as polygonal network between piles or pustules that ornate the lateral surface of the shell.
, fig. 2) arranged as polygonal network between piles or pustules that ornate the lateral surface of the shell.
The question arises as to whether or not the genus Lakadongia & , 2003, from Thanetian beds of northeastern India, is identical to Bolkarina or has a fundamentally different architecture. The photographic documentation said to illustrate the architecture of the shell is difficult to interpret because the authors do not identify any structural element in the pictures. Their figs. 3-4 and 7 on Pl. 1 seem to depict orbitoidiforms, not Lakadongia. The same must be said about the drawing at the bottom of text fig. 4a. On the other hand, the drawing at the top of the same figure seems to me to reflect quite clearly the basic architecture of Bolkarina with its biserially arranged expanse chambers. The sections stated to be transverse, perpendicular to the equatorial plane, seem to me rather oblique with an inclination of about 45°. Keeping in mind the difficulties of structural analysis in this group I recommend the name Lakadongia not be used.
Taxonomical remark: in 1998 changed the specific name aksarayi to aksarayiensis to show that the name refers to a location, not a person. Changes of this kind are not permitted, as long as any combination of letters that sound like Latin is recognized as a name by the international commission. If, however, Bolkarina aksarayi and Miscellanites globularis are confirmed to be two generations of one species, a name in common, M. globularis, would then have priority.
The Family Miscellaneidae is treated here as a separate group of planispiral larger foraminifera that has neither rotaliid nor nummultid diagnostic features. Moreover, it is unrelated to the Pellatispiridae for it lacks a marginal crest in the supplemental skeleton. The placement of the Miscellaneidae among the bilamellar benthic foraminifera depends on the hierarchical weight for classification given the mode of coiling, planispiral versus trochospiral, and the placement and type of the aperture: interiomarginal over its whole length or areal. There must be a link between planispiral coiling with the position of the foramen that is symmetrical to the periphery of the test or consists of a single row of multiple foramina in interiomarginal position. A similar trend can be observed in agglutinated and porcelaneous shells (fusulinids, Barkerinidae, Alveolinidae). Consequently, the planispiral mode of coiling may be used as guide to fix the position of the Miscellaneidae within the system. The Nonionidae are defined as a planispiral shell with a single interiomarginal aperture. Nonion faba & , 1798 (see & , 1980) exhibits in addition an umbilical structure in which interlocular spaces are fringed with feathering or fine beads. A systematic progression toward the periphery of such a structure by the addition of outer lamellae ( & , 1980, p. 117, fig. 1) produced by a much greater number of chambers per whorl, would form a kind of enveloping canal system like the one of Miscellanea. Consequently, I advocate the attribution of the Miscellaneidae to the Superfamily Nonionacea , 1854 (see & , 1964).
The combined range of the ten miscellaneid taxa treated in this paper is confined to zones SBZ 2-5
(Fig. 4 ![]() ). The geographic distribution of the members of the family extends from Tibet to Somalia, covers all the Near and Middle East including the Arabian Peninsula, reaches Egypt and crosses the Mediterranean to the Adriatic platform and into the Pyrenean Gulf in Spain and southwestern France. Apparently, like the alveolinids, the Paleocene Mid-Atlantic was not crossed by any member of the Miscellaneidae but the Paleocene nummulitid Ranikothalia did.
). The geographic distribution of the members of the family extends from Tibet to Somalia, covers all the Near and Middle East including the Arabian Peninsula, reaches Egypt and crosses the Mediterranean to the Adriatic platform and into the Pyrenean Gulf in Spain and southwestern France. Apparently, like the alveolinids, the Paleocene Mid-Atlantic was not crossed by any member of the Miscellaneidae but the Paleocene nummulitid Ranikothalia did.
The family is divided into the Miscellaneinae with a single foramen and the Miscellanitinae with two symmetrical (Ornatononion or a single row of numerous foramina (Miscellanites).
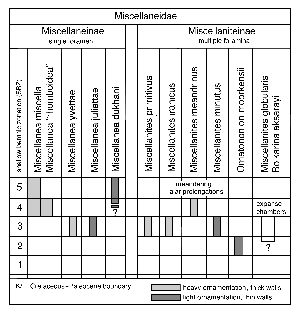
Fig. 4 ![]() sketches a corresponding hypothesis concerning the phylogenetic relationships of the taxa taking into account their age relationships, their apertural features and their heavy or light ornamentation which may reflect details of discrete patterns in the enveloping canal system. The separation of species with a single foramen from species with multiple foramina is new and must have taxonomic consequences on the generic level.
sketches a corresponding hypothesis concerning the phylogenetic relationships of the taxa taking into account their age relationships, their apertural features and their heavy or light ornamentation which may reflect details of discrete patterns in the enveloping canal system. The separation of species with a single foramen from species with multiple foramina is new and must have taxonomic consequences on the generic level.
The meandrine alar prolongations in Miscellanites meandrinus and the expanse chambers on either side of a main chamber layer in the discoidal Bolkarina are of particular interest for the comparative and functional anatomy of larger foraminifera. The two extreme morphological variants repeat features known elsewhere in unrelated forms. Meandering alar prolongations are present in porcelaneous families (Meandropsinidae, Archaiasinidae; , 2005) and in Nummulites (, 1988) with a lamellar-perforate test. In all cases, the shells are planispiral-involute, that is, with long and narrow alar prolongations. All porcelaneous forms have multiple apertures along the interiomargin of the alae. In nummulites, such foramina are unknown but an intraseptal interlocular canal system feeds the numerous orifices of the sutural or trabecular canals (, 2000, fig. 20). In miscellaneids, the meandrine septal sutures are marked by the orifices of a double row of sutural canals that begin to envelop the free chamber walls only after the deposition of several supplemental lamellae. In all cases, the number of orifices of the chambers or of the interlocular space is increased by the progressive elongation of the meandrine sutures. Therefore, the independent rise of meandrine sutural patterns reflects the implementation of common functions that support the success in the life of the benthic foraminifer.
Expanse chambers as lateral skeletons of lenticular shells as in Bolkarina are known also in unrelated forms, for instance in Vaughanina cubensis (, 2005). The expanse chamber developed in permanently sessile forms fixed to a solid substrate, as for instance Planogypsina squamiformis (). The chessboard pattern of the successive expanse chambers obviously helps the occupation on the substrate of as much surface as possible with minimal costs, that is, with the minimum amount of new chamber lumen that must be filled with new protoplasm during the successive steps of growth. In the orbitoidiform Vaughanina a cyclical marginal crest is overgrown from both sides by the lateral skeleton in the form of expanse chambers. In Bolkarina aksarayi biserial expanse chambers form an orbitoidiform structure without a differentiation into a true main layer of chamberlet cycles and lateral chamberlets. In orbitoidiforms such an architecture is generally interpreted as being greenhouses for symbiontic algae. However, Planogypsina has no symbionts. Here the expanse chamber increases the number of orifices on the surface of the shell in order to collect food, i.e. pellets from the plankton, that fall from the water column. The large number of coccoliths in the feeding cysts confirms the validity of this interpretation.
Both reviewers Antonino (Vienna) and Ercan (Istambul) have contributed by their remarks to substantial ameliorations of the manuscript and Nestor has improved the readability of the English text. Many thanks to all.
C.G. (1988).- Septa, septal traces and septal filaments in the foraminiferal genus Nummulites .- Journal of Micropalaeontology, London, vol. 7, n° 1, p. 89102.
M. & A.A. (1999a).- Lower Tertiary biostratigraphy of the Kala Chitta Range, Northern Pakistan.- Revue de Paléobiologie, Genève, vol. 18, n° 1, p. 123-146.
M. & A.A. (1999b).- Microfacies and foraminiferal assemblages from the early Tertiary rocks of the Kala Chitta Range (Northern Pakistan).- Géologie méditerranéenne, Marseille, t. 26, n° 3-4, p. 185201, 4 Pls.
A. d' & J. (1853).- Description des animaux fossiles du groupe nummulitique des Indes, précédée d'un résumé géologique et d'une monographie des Nummulites.- Gide & Baudry, Paris, 373 p., 36 Pls.
R.C. van (1946).- Foraminifera from the Middle Eocene in the southern part of the Netherlands Province of Limburg.- Mededelingen Geologische Stichting, Leiden, (series C), vol. 5, 144 p., 13 Pls.
H.M. de (1827).- Manuel de malacologie et de conchyologie.- Levrault, Paris, 664 p., 97 Pls.
P.J. (1952).- Estudio sistematico de los foraminiferos rotaliformes.- Boletin de Geologia, Caracas, vol. II, n° 4, 230 p., 35 Pls.
G., J.J. & L.E. (1972).- Étude stratigraphique et paléogéographique des formations tertiaires de la région de Neyriz (Fars interne, Zagros iranien).- Revue de l'Institut Français du Pétrole, Rueil-Malmaison, vol. 27, n° 3, p. 369-405.
M.K. (2008).- Evolution and geological significance of larger benthic foraminifera.- Developments in Palaeontology & Stratigraphy, Amsterdam, vol. 21, 540 p.
H.J. (1861).- Further observations on the structure of Foraminifera and on the larger fossilized forms of Sind, etc., including a new genus and species.- Journal of the Bombay Branch of the Royal Asiatic Society, vol. VI, p. 31-96.
W.S. (1953).- Criteria for the recognition of certain assumed camerinid genera.- Bulletins of American Paleontology, Ithaca, vol. 35, n° 147, p. 1-12, 2 Pls.
W.S. (1960).- The genus Camerina.- Bulletins of American Paleontology, Ithaca, vol. 41, n° 190, p. 189-205, 4 Pls.
O.G. (1856).- Paleontologia del regno di Napoli, parte II.- Atti della Accademia Pontaniana, Napoli, vol. 7, n° 2, p. 113-378.
L.M. (1927).- The Ranikot beds of Thal.- Quaterly Journal of the Geological Society of London, vol. 83, p. 260-290.
L.M. & E.S. (1937).- The Eocene Beds of the Punjab Salt Range.- Palaeontologia Indica, Calcutta, vol. 24, Memoir 1, 79 p., 7 Pls.
. H. (1916).- Le Crétacé et l'Éocène du Tibet central.- Paleontologia Indica, Calcutta, (new series), vol. 5, part 3, p. 1-84.
K., B., M., M-L. & D. (1988).- Maastrichtian, Danian and Thanetian beds in Dolenja Vas (NW Dinarides, Yugoslavia): microfacies, foraminifers, rudists and corals.- Razprave, Slovenska Akademija Znanosti in Umenosti, Ljubljana, (classis IV Historia naturalis), vol. 29, n° 6, p. 147-224, 35 Pls.
H.J. & F. (1980).- What is Nonion? Problems involving foraminiferal genera described by 1808 and the type species of & 1798.- Journal of Foraminiferal Research, Lawrence, vol. 10, p. 173-179.
A.F.M.M. (1958).- Cincoriola, a new generic name for Punjabia , 1956.- Contributions from the Cushman Foundation for Foraminiferal Research, Washington, vol. 9, p. 103.
L. von & J.P.C. von (1798).- Testacea microscopica, aliaque minuta ex generibus Argonauta et Nautilus, ad naturam picta et descripta.- Camesina, Vienna, 123 p.
S. (1937).- Notes on some interesting Creaceous and Tertiary Foraminifera from the West Indies.- Journal of Paleontology, Tulsa, vol. 11, p. 110-117.
L. (1963).- Les Alvéolines paléogènes, Exemple d'un genre polyphylétique.- In: G. von, J.D., W.L. & C.W. (eds.), Evolutionary trends in Foraminifera.- Elsevier, Amsterdam, p. 298-314.
L. (1977).- Foraminifères operculiniformes.- Mémoires du Muséum National d'Histoire Naturelle, Paris, (Nouvelle série C), t. XL, 159 p., 66 Pls.
L. (2000).- Functional morphology of benthic foraminiferal shells, envelopes of cells beyond measure.- Micropaleontology, New York, vol. 46, supplement 1, p. 57-86, 21 figs.
L. (2001).- The shell cavity systems in elphidiid and pellatispirine bilamellar foraminifera, Editor's preface.- Micropaleontology, New York, vol. 47, supplement 2, p. 1-4.
L. (2005).- Geometrical constraints in foraminiferal architecture: consequences of change from planispiral to annular growth. In: Methods and applications in micropalaentology.- Studia Geologica Polonica, Kraków, vol. 124, p. 99-115.
L. (2006).- Illustrated glossary of terms used in foraminiferal research.- Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2006/02 (CG2006_M02), 126 p.
URL: https://carnetsgeol.net/cg/06/M02/
L. & S. (1980).- The structure of calcarinid foraminifera.- Schweizerische Paläontologische Abhandlungen, Basel, Vol. 101, p. 115-154.
L., Z. & M. (2001).- Spiral canals of some Elphiidae.- Micropaleontology, New York, vol. 47, supplement 2, p. 5-34.
L., J. & A.A. (1998).- Emendation of Alveolina vredenburgi & 1937 from the Surghar Range, Pakistan.- Dela SAZU, Ljubljana, vol. 34, n° 2, p. 155-163.
M. (1970).- Die alttertiären Grossforaminiferen Südost-Afghanistans unter besonderer Berücksichtigung der Nummulitiden-Morphologie, Taxonomie und Biostratigraphie.- Münsterische Forschungen zur Geologie und Paläontologie, Münster, Heft 16/17, 400 p., 19 Pls.
J. & U. (1989).- The early Tertiary (middle-late Paleocene) limestones from the western Gulf of Suez, Egypt.- Neues Jahrbuch Geologie und Paläontologie, Abhandlungen, Stuttgart, Band 177, Heft 3, p. 289-332.
U. (1988).- Structural analysis and taxonomic revision of Miscellanea, Paleocene larger Foraminifera.- Eclogae Geologicae Helvetiae, Basel, vol. 81, n° 3, p. 689-721.
A.R. & H. (1964).- Sarcodina, chiefly "Thecamoebians" and Foraminiferida.- In: R.C. (ed.), Treatise on Invertebrate Paleontology, Part C, Protista 2.- The University of Kansas Press, Lawrence, and The Geological Society of America, Boulder, vol. 1, 510 p.; vol. 2, p. 511-900.
A.R. & H. (1987).- Foraminiferal genera and their classification.- Van Nostrand Reinhold, New York, vol. 1, 970 p.; vol. 2, 212 p., 847 Pls.
P. (1964).- Les facies du Montien (France, Belgique, Hollande). In: Colloque sur le Paléogène, Bordeaux 1962.- Mémoires du Bureau de Recherches Géologiques et Minières, Orléans, vol. 28 (2), p. 1077-1101, 2 Pls.
K. & A.K. (2003).- Lakadongia, a new Orbitoidal foraminiferal genus from the Thanetian (Paleocene) of Meghalaya, NE India.- Micropaleontology, New York, vol. 49, n° 3, p. 277-291, 4 Pls.
T.L. (1982).- Foraminifera of the Montian stratotype and of subjacent strata in the "Mons Well 1969" with a review of Belgian Paleocene stratigraphy.- Mémoires pour servir à l'Explication des Cartes Géologiques et Minières de la Belgique, Bruxelles, n° 17.2, 186 p., 18 Pls.
Y. (1951).- The stratigraphic value of Miscellanea and Pellatispira in India, Pakistan and Burma.- Proceedings of the Indian Academy of Sciences, Bangalore, vol. 33, p. 41-48.
W.L.F. (1926).- The larger Foraminifera of the Upper Ranikot (Lower Eocene) of Sind, India.- Geological Magazine, London, vol. 63, n° 3, p. 112-121, 2 Pls.
E., E., S. & S. (2001).- Late Paleocene orthophragminae (Foraminifera) from the Haymana- Polatli Basin (central Turkey) and description of a new taxon, Orbitoclypeus haymanaensis.- Micropaleontology, New York, vol. 47, n° 4, p. 339-357, 4 Pls.
J. (1935).- À propos de Siderolites vidali et de quelques autres.- Bulletin de la Société géologique de France, Paris, (série 5), vol. 4, p. 225-236.
J., M., A., C., A., M., R., G. & M. (2008).- SBZ 2-6 larger foraminiferal assemblages from the Apulian and Pre-Apulian domains.- Atti del Museo Civico di Storia Naturale di Trieste, vol. 53, supplement, p. 131146.
J., R., T. & L. (1998).- Larger foraminiferal biostratigraphy of the Maastrichtian-Ypresian Wadi Mashib succession (South Hadramawt Arch, SE Yemen).- Zeitschrift geologische Wissenschaften (Berlin), vol. 26 (5-6), p. 609-635.
A. (1976).- Contribution à l'étude de quelques grands foraminifères de l'Iran.- Société Nationale Iranienne des Pétroles, Laboratoire de Micropaléontologie, Teheran, Publication n° 6, 79 p., 10 Pls.
A. (1978).- Paleogene biostratigraphy of some parts of Iran.- National Iranian Oil Company, Geological Laboratories, Teheran, Publication n° 7, 82 p., 41 Pls.
A. (1983).- Stratigraphy and faunal assemblage of Paleocene-Lower Eocene in Iran.- National Iranian Oil Company, Geological Laboratories, Teheran, Publication n° 10, 73 p., 49 Pls.
D.M. & A.V. (eds., 1959).- Principles of paleontology. Part 1. General part and Protozoa.- Academiya Nauk SSSR, Moskow, 482 p. [in Russian]; English translation: Israel Program for Scientific Translations (1962), S. Monson, Jerusalem, 728 p.
M. (1969).- Microfacies and microfossils of the Zagros Area, southwestern Iran (from Pre-Permian to Miocene).- International Sedimentary Petrographical Series, Brill, Leiden, vol. XII, 102 p., 105 Pls.
D. & S. (1988).- Southern Tethys biofacies.- Agip Mineraria, San Donato Milanese, 235 p.
M.S. von (1854).- Über den Organismus der Polythalamien (Foraminiferen), nebst Bermerkungen über die Rhizopoden in Allgemeinen.- Wilhem Engelmann, Leipzig, xviii + 68 p., 7 Pls.
J., L., E., K., C., A.K., G., R., J., J.M., H., E., A., Y., J. & E. (1998).- Larger foraminiferal biostratigraphy of the Tethyan Paleocene and Eocene.- Bulletin de la Société géologique de France, Paris, vol. 169, n° 2, p. 281-299.
J. (1952).- Ordre des Foraminifera. In: J. (ed.).- Traité de paléontologie. Tome I.- Masson, Paris, p. 133-301.
E. (1972).- Systematic study of new species of the genera Fabularia and Kathina from Paleocene.- Türkiye Jeoloji Kurumu Bülteni, Ankara, c. 15, p. 277-249. 8 Pls.
E. (1981).- Bolkarina, new genus (Foraminiferida) and some associated species from the Thanetian limestone (Central Turkey).- Eclogae Geologicae Helvetiae, Basel, vol. 74, n° 1, p. 75-95, 5 Pls.
E. (1997).- The species of Miscellanea , 1935 (Foraminiferida) in the Thanetian-Ilerdian sediments of Turkey.- Revue de Paléobiologie, Genève, vol. 16, n° 1, p. 77-99.
E. (1998).- Foraminiferal description and biostratigraphy of the Paleocene-Lower Eocene shallow-water limestones and discussion on the Cretaceous-Tertiary boundary in Turkey.- Mineral Research and Exploration (MTA), General Directorate, Ankara, (Monography Series), n° 2, 117 p., 68 Pls.
E. (2004).- Türkije'nin Mesozoyik ve Senozoyik Yeni Bentik Foraminiferleri.- Jeoloji Mühendisleri Odasi Yayinlari 84 (Chamber of Geological Engineers of Turkey: publication 84), Ankara, Emegin Bilimsel Sentezi [Scientific Synthesis of Life Time], özel Sayi [Special Volume 1] 1, 219 p., 66 Pls.
E. & Ö. (1975).- Miscellanea meandrina () 'in Turkiye'de zuhuru ve yasi. (The occurrence and the age of Miscellanea meandrina () in Turkey.- Türkiye Jeoloji Kurumu Bülteni, Ankara, c. 18, p. 193-194, 2 Pls.
A.H. (1954).- Lower Tertiary foraminifera of the Qatar peninsula.- British Museum (Natural History), London, 96 p., 44 figs., 15 Pls.
T.W. & W.S. (1941).- Preliminary report on the Cretaceous and Tertiary larger Foraminifera of Trinidad British West Indies.- Geological Society of America, Special Papers, Boulder, vol. 30, 137 p.
J. (1962).- Étude stratigraphique et paléontologique du Montien des Petites Pyrénées et du Plantaurel.- Privat, Toulouse, 331 p., 22 Pls.
Click on thumbnail to enlarge the image.
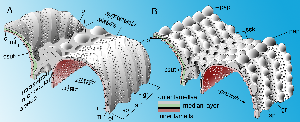
Figure 1: Patterns of ornaments in ultimate and penultimate chambers of Miscellanea (A) and Daviesina langhami (B). Stereographs showing the bilamellar primary wall and the independent geometry of the first supplementary outer lamella that is folded over itself. The folds cover the interlocular space except the radial canals. These connections between the interlocular space and the ambient environment represent the impending enveloping canal system that determines the nature of the supplemental skeleton. Note the distribution of pores that can be generated only in the presence of a median layer that separates the two lamellas in the primary chamber wall. Schematic, not to scale.
Abbreviations: af: apertural face; b: beads; gr: grooves of the feathering along the shoulders of the intraseptal interlocular space; il: inner lamella; ml: median layer; ol: outer lamella; p: pore; pap: papilla; ssk: supplemental skeleton; sp: spurs of the feathering.
Click on thumbnail to enlarge the image.
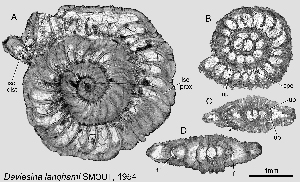
Figure 2: Daviesina langhami , 1954. A: Microspheric specimen, equatorial section. B: Megalospheric specimen, equatorial section. C, D: megalospheric specimens, axial sections. The horizontal orientation of the axial sections indicates a dorsal side (upwards) and a ventral side (downwards). Note the asymmetry of the foramen.
Abbreviations: f: foramen; isc dist: distal end of intraseptal canal system determining the basic ornamental pattern; isc prox: proximal part of intraseptal canal system; spc: spiral canal; up: umbilical plate. Specimens A & D from sample 92010a, specimens B & C from sample 93504; Dhak pass, Salt Range, Pakistan, top of Lockhard Lst - SBZ 4.
Click on thumbnail to enlarge the image.
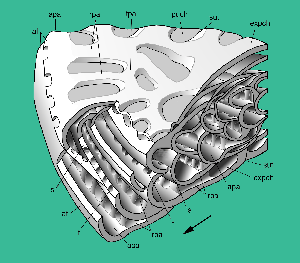
Figure 3: Bolkarina aksarayi , 1981. Stereograph to explain the structure, schematic, not to scale. Peripheral part of sector; radial and oblique sections. Supplemental apertures, possibly along the sutures of the expand chambers, are omitted, main equatorial foramina tentatively placed in interiomarginal position. Note the biserial arrangement of the annular passages. Compare with , 2005, fig. 4 on p. 107.
Abbreviations: af: apertural face; apa: annular passage (in preseptal position); expch: expanse chamber; f: foramen; puch: penultimate (expanse) chamber; rpa: radial passage; s: septum; sut: (chamber) suture; tpa: transverse tubular passage. Arrow: direction of growth.
Click on thumbnail to enlarge the image.
Figure 4: Stratigraphic distribution of the taxa belonging to the Miscellaneidae and an hypothesis as to their phylogenetic relationships.

Click on thumbnail to enlarge the image.
Plate 1. External lateral views of some heavily ornate involute shells; figs. 1-13. Daviesina langhami , 1954. Note the whorl sutures indicating the imperfect involution of the test and the doubling of the periphery in 3 and 4. Specimens 1-4 from sample 92010a, Dhak pass, Salt Range, Pakistan, top Lockhart Lst., SBZ 4. Specimens 5-13 from sample 92014, Nammal Gorge, Salt Range, Lowest part of Nammal shales, SBZ 5; figs. 14-20. Miscellanea miscella (d' & , 1853). All specimens from samples 95108 and 95109, Dhak pass, Salt Range, lowest part of Patala shales, SBZ 4. Note specimen 20 that is figured from both sides in order to show the degree of reflection in the ornament; figs. 21-27. Miscellanea juliettae , 1988. All specimens from sample 93563, Dandot village, Salt Range, Pattala shales below coal seam, SBZ 3.
Abbreviations: dp: double periphery; per: simple periphery; wsut: whorl suture.

Click on thumbnail to enlarge the image.
Plate 2. Miscellanea miscella (d' & , 1853). Megalospheric specimens, equatorial sections in transparent light. All specimens from samples 95108 and 95109, Dhak pass, Salt Range, lower Patala shales, SBZ 4.
Abbreviations: bic: straight wall between protoconch and deuteroconch that indicates the biconch quality of the nepiont; f: foramen; isc: intraseptal canal system; spc: spiral canal, px: paries proximus; up: umbilical plate.

Click on thumbnail to enlarge the image.
Plate 3. Miscellanea miscella (d' & , 1853). Megalospheric specimens, axial sections in transparent light. All specimens except 5 and 15 from samples 95108 and 95109, Dhak pass, Salt Range, Pakistan, lower Patala shales, SBZ 4; 5 & 15 from sample 95601, Le Quillet, western Aquitaine (southwestern France), SBZ 4.
Abbreviations: alp: lumen of alar prolongation; bic: biconch wall; f: foramen; isc: intraseptal canal system; pct: peripheral chamber tip; px: paries proximus; s: septum; spc: spiral canal, up: umbilical plate.
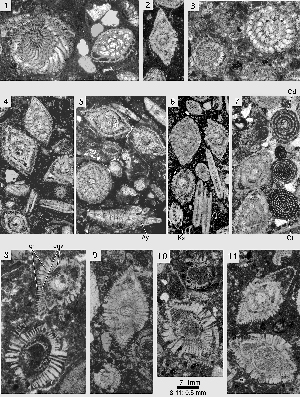
Click on thumbnail to enlarge the image.
Plate 4. Miscellanea miscella (d' & , 1853). Megalospheric topotypes of M. "rhomboidea" , 1988. Specimens collected by & from Wadi Araba area, Egypt, SBZ 4.; fig. 1: tangential and centered-oblique sections; fig. 2: axial section; fig. 3: tangential and equatorial sections; fig. 4: axial sections; fig. 5: axial and oblique-centered sections. Ay: Assilina yvettae ; fig. 6: axial sections. Ks: Kathina selveri , 1954; A: Assilina sp; fig. 7: oblique sections with glomalveolinids. Gd: Glomalveolina dachelensis (); Gt: Glomalveolina telemetensis ; fig. 8: axial sections, details showing the enveloping canal system (env) and its orifices (or); fig. 10: axial section; fig. 11: axial sections. Note on the upper right a section of the penultimate chamber with incipient enveloping canals inspite of the extremely low number of secondary lamellae.
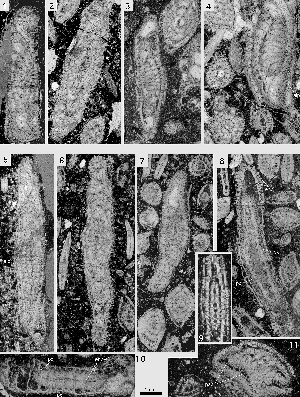
Click on thumbnail to enlarge the image.
Plate 5. Miscellanea miscella (d' & , 1853). Microspheric topotypes of M. "rhomboidea" , 1988. Specimens collected by & from Wadi Araba area, Egypt, SBZ 4. fig. 1: oblique section inclined to about 30° in respect to the axial plane; fig. 2: axial section; fig. 3: transverse section parallel to the axis of the shell; fig. 4: oblique sections of both gnerations; fig. 5: transverse section close to the axial one; fig. 6: axial section; fig. 7: transverse section parallel to but quite far from the shell axis, axial section of megalospheric specimen and of Plumokathina aff. subsphaerica () on lower left; fig. 8: transverse section far from center. Note the enveloping canal system in the last whorl; figs. 9-10: tranverse sections; fig. 11: tangential section showing ornamentation.
Abbreviations: alp: alar prolongations; env: enveloping canal system; isc: intraseptal canal system; pap: papilla; px: paries proximus.

Click on thumbnail to enlarge the image.
Plate 6. Miscellanea dukhani , 1954: figs. 1-7, and Miscellanea miscella (d' & , 1853): figs. 8-9; figs. 1-2: micro- and megalospheric specimens, equatorial sections. from sample Gill 123. Salt Range, Pakistan; fig. 3: megalospheric specimen, equatorial section. From sample 96108, Dhak pass, Salt Range. Patala shales, SBZ 4; figs. 4-5: microspheric specimens, equatorial and axial sections. From sample Gill-"Dhak pass, Salt Range. Patala shales", SBZ 4. Note the megalospheric Ranikothalia nuttalli () cemented with specimen; figs. 5-7: axial sections, not quite centered, of megalosheric specimens. Note in 6 the delicate last whorl that is compressed and broken by the compaction of the sediment. From sample Gill 123, Salt Range, Patala shales.
Miscellanea miscella (d' & , 1853); figs. 8-9: axial sections of a microspheric and a megalospheric specimen. From sample 95109, Dhak pass, Salt Range, lower Patala shales, SBZ 4.
Abbreviations: alp: alar prolongations; f: foramen; lh: loophole; mc: marginal cord of Ranikothalia nuttalli; pct: peripheral chamber tip; up: umbilical plate.

Click on thumbnail to enlarge the image.
Plate 7. Miscellanea dukhani , 1954; figs. 1-8: paratypes from Qatar, "highest zone of the Paleocene" according to , 1954. British Museum (Natural History) P 40151; figs. 1-2 & 6: megalospheric generation, equatorial and axial sections. 1 and 6 are the same specimen at different standart enlargements to facilitate comparisons; figs. 3-8: microspheric generation, in part poorly preserved, equatorial and axial sections. Note in 3 and 7 the peculiar ornamentation with double rows of beads (b) along the sutures and with a single row of papillae (pap) in between the septa.
Miscellanea yvettae , 1988; figs. 9-16: all specimens from sample 73932 H. , Tapiau-Cazères, Petites Pyrénées, Aquitaine, southwestern France, SBZ 3; fig. 9: equatorial section of microspheric specimen. Note the narrowing of the spire in the last whorls that corresponds to the rise of two peripheries in the adult stages of growth; figs. 10-15: megalospheric specimens in equatorial sections; fig. 16: axial section of megalospheric specimen.

Click on thumbnail to enlarge the image.
Plate 8. Miscellanea yvettae , 1988. Tapiau-Cazères, Petites Pyrénées, Aquitaine (southern France), SBZ 3; figs. 1-2: transverse sections perpendicular to the shell axis of microspheric specimens. Note the ornamentation on the inner whorls as seen in tangential section; figs. 3-4 & 6: detail (3) and complete axial sections (4 & 6) of microspheric specimens showing a double periphery due to equatorial lacing of the shell (arrow); fig. 5: axial section of microspheric specimen without equatorial lacing; fig. 7: axial section of megalospheric specimen. Details showing the apertural features and the fan-shaped intraseptal canal system (isc); figs. 8-10: oblique random sections of microspheric specimens; fig. 11: oblique random section of megalospheric specimen.
Abbreviations: f: foramen; isc: intraseptal canals; s: septum; spc: spiral canal; up: umbilical plate.
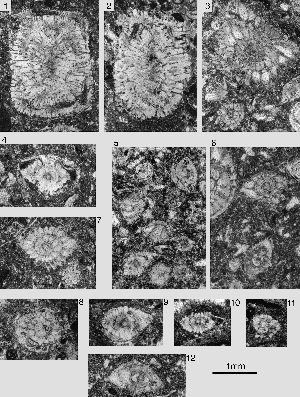
Click on thumbnail to enlarge the image.
Plate 9. Miscellanea yvettae , 1988. Wadi Araba, Egypt, from samples collected by & , SBZ 3; figs. 1-3: microspheric specimens, oblique sections; fig. 4: megalospheric specimen with an equatorial lacing in the ultimate half whorl ; axial section; figs. 7-12: megalospheric specimens, random sections near to the equatorial plane (7-8) and near the axis of the shell (6 & 9-12). Note in 5 the tangential section of M. yvettae in the middle left of the picture, showing a single row of beads, and on the upper right an oblique section of Plumokathina subsphaerica ().

Click on thumbnail to enlarge the image.
Plate 10. Miscellanea juliettae , 1988; figs. 1-15 from sample 93563, Dandot village, Salt Range (Pakistan). SBZ 3; figs. 17-21 from samples 93501 93503, Nammal Gorge, Salt Range, Lockhart Limestone. SBZ 3; figs. 1-3: megalospheric specimens, equatorial sections; figs. 4-5: microspheric specimen, not quite centered equatorial section with structural details; figs. 6-12: megalospheric specimens, equatorial (6) and axial sections showing structural details; figs. 13-16: megalospheric specimen of small size, equatorial sections; figs. 17-18: equatorial sections of megalospheric (17) and microspheric (18) specimens; figs. 19-20: axial sections of microspheric (19) and megalospheric (20) specimens; fig. 21: equatorial section of microspheric specimen with narrowly spaced septa figured at half standard enlargement.
Abbreviations: co: canal orifices; f: foramen; isc: intraseptal canals; pct: peripheral chamber tip; spc: spiral canal; up: umbilical plate.
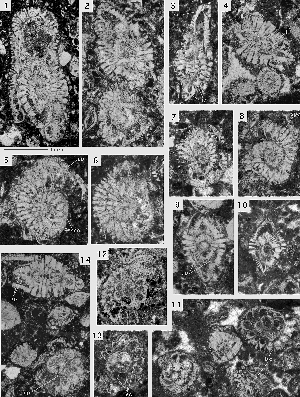
Click on thumbnail to enlarge the image.
Plate 11. Miscellanea juliettae , 1988. All from Wadi Araba area, Egypt, collected by & . SBZ 3; figs. 1-8: random sections of microspheric specimens. Note the openig of the spiral in the last whorl; figs. 9-11: random sections of megalospheric specimens. Note the petal-like aspect of the chambers in sections with no or little inclination in respect to the equatorial plane of the shell. The petal-like intersection of the chambers is due to a particularly large interlocular space that straddles the proximal end of the septum. In 4 and 11 there are also sections of Plumokathina subsphaerica ().
Abbreviations: co: canal orifices; f: foramen; isc: intraseptal canal system; pct: peripheral chamber tip; spc: spiral canal; up: umbilical plate.
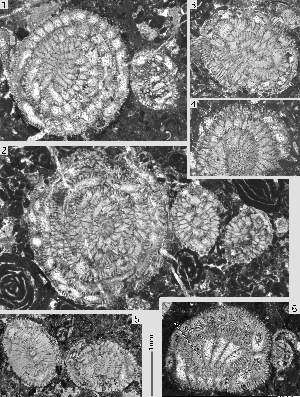
Click on thumbnail to enlarge the image.
Plate 12. Miscellanites iranicus (, 1983). All from sample Kar 13 collected by A. , Kuh-e-Kargan near Kermanshah, Taurus (Iran). SBZ 3; figs. 1-4 & 6: microspheric specimens, randomly oblique sections; figs. 1-2 & 5: additional megalospheric specimens, oblique sections, except specimen 5 with a good axial section.Note the multiple foramina (f) in the tangential section 6. On the right an oblique section of Redmondina henningtoni .
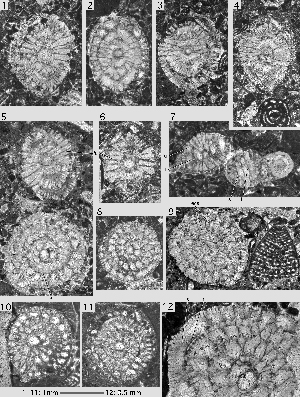
Click on thumbnail to enlarge the image.
Plate 13. Miscellanites iranicus (, 1983). Megalospheric specimens. All from sample Kar 13, Kuh-e-Kagan near Kermanshah. SBZ 3; figs. 1-4: axial sections; fig. 5: equatorial and axial sections; fig. 6: axial section; fig. 7: oblique and tangential sections. In addition there is a section perpendicular to the shell axis of Cincoriola ovoidea , 1958; figs. 8-11: more or less perfect equatorial sections; fig. 9: in addition: Fallotella alavensis ; fig. 12: oblique-centered section showing detail of foraminal features.
Abbreviations: ecs: enveloping canal system; f: foramen; fu: funnel; s: septum.
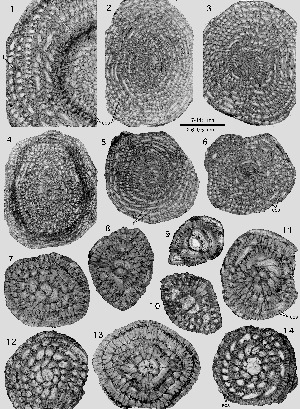
Click on thumbnail to enlarge the image.
Plate 14. Miscellanites meandrinus (, 1861). All from a donation of material by E. , from Agri, northeastern Turkey, except 9 from sample H. 72008, Le Quillet. Aquitaine, (southwestern France). SBZ 4; figs. 1-6: microspheric specimens with structural details in 1; figs. 7-14: megalospheric specimens, subequatorial and centered subaxial sections.
Abbreviations: ecs: enveloping canal system; f: foramen.
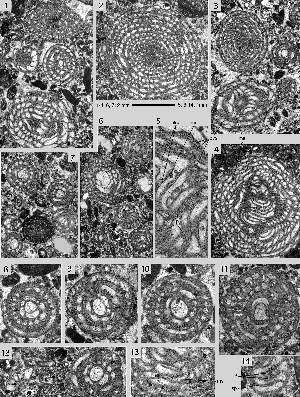
Click on thumbnail to enlarge the image.
Plate 15. Miscellanites meandrinus (, 1861), from Wadi Araba, Egypt, collected by & ; figs. 1-5: microspheric specimens, random oblique sections. The meandrine septa appear in the tangential parts of the sections; figs. 6-14: megalospheric specimens, oblique-centered sections (6-12), tangential sections (13, 14).
Abbreviations: ecs: enveloping canal system; f: foramen; ilsp: interlocular intraseptal space; ms: meandrine septum; p: pores.

Click on thumbnail to enlarge the image.
Plate 16. Miscellanites primitivus (, 1983). All specimens from Narp, near Pau, Aquitaine, (southwestern France). SBZ 3; figs. 1-2: external lateral view; figs. 3-4: external apertural view with apertural face; fig. 5: oblique-centered section; figs. 6-8, 11-18 & 21-24: axial sections of megalospheric specimens; figs. 9 & 19: megalospheric specimens, transverse sections parallel to shell axis; fig. 10: microspheric specimen, axial section. Note the multiple foramina.
Abbreviations: af: apertural face; f: foramen; spc: spiral canal; up: umbilical plate.
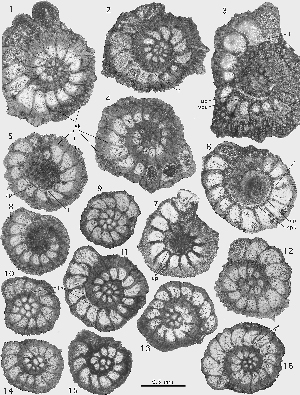
Click on thumbnail to enlarge the image.
Plate 17. Miscellanites primitivus (, 1983). All specimens from Narp, near Pau, Aquitaine, (southwestern France). SBZ 3. Equatorial and subequatorial sections; figs. 1-5: microspheric generation; figs. 6-16: megalospheric generation. Note the heavy umbilical plate separating a wide spiral canal.
Abbreviations: f: foramen; sk: supplemental skeleton as revealed by an enveloping canal system; spc: spiral canal; up: umbilical plate.
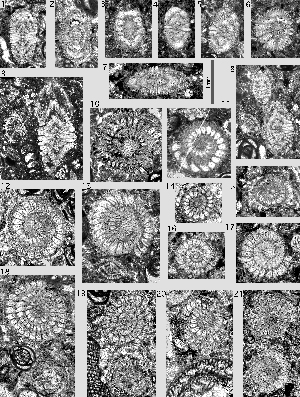
Click on thumbnail to enlarge the image.
Plate 18. Miscellanites minutus (, 1983). All from samples Kar 11 and 12 collected by A. , Kuh-e-Kargan near Kermanshah, Iran. SBZ 3; figs. 1-8: megalospheric specimens, sections parallel to or in the shell axis; fig. 9: microspheric specimen, axial section; figs. 10-13: microspheric specimens, transverse sections perpendicular to shell axis; figs. 11-21: megalospheric specimens, sections perpendicular to shell axis or in the equatorial plane. In addition, there is a fragment of "Taberina" daviesi in 19, of "Globoreticulina" paleocaenica in 20.
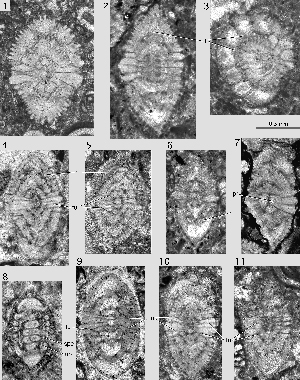
Click on thumbnail to enlarge the image.
Plate 19. Miscellanites minutus (, 1983). Megalospheric specimens from samples Kar 11 and 12, Kuh-e-Kargan near Kermanshah, Iran. SBZ 3; figs. 1 & 9-10: axial sections; figs. 2, 8 & 11: transverse sections parallel to the shell axis; figs. 3, 5 & 7: oblique sections.
Abbreviations: f: foramen; fu: funnel; pr: proloculus; spc: spiral canal; up: umbilical plate.

Click on thumbnail to enlarge the image.
Plate 20. Ornatononion moorkensii n. gen. n. sp. All from Mons, Belgium. SBZ 2; figs. 1-7: equatorial sections; 5 is a little oblique to show the strong umbilical plate and the wide spiral canal; figs. 9-10: transverse sections parallel to coiling axis; fig. 11: oblique section showing spiral canal; figs. 8 & 12-13: axial sections. Note the absence of a noticable dimorphisme in this small-sized, early species.
Abbreviations: f: foramen; s: septum; spc: spiral canal; up: umbilical plate.
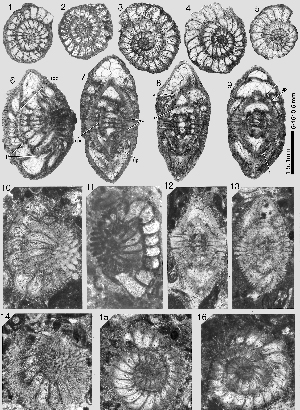
Click on thumbnail to enlarge the image.
Plate 21.
Ornatononion moorkensii n. gen. n.sp.; figs. 1-9: from Mons, Belgium. SBZ 2. Equatorial sections at low enlargment in order to compare with
Pl. 18 ![]() , figs. 10-22;
figs. 6-9: oblique sections progressively more parallel to the coiling axis; figs. 10-16: Specimens from different places in the Petites Pyrénées, Aquitaine, collected by J. & Y. . SBZ 2; fig. 10: oblique section, from S'ours; figs.
11-12: oblique and axial sections, from Bouholoup; fig. 13: oblique-centered section, from
S'ours; fig. 14: transverse section perpendicular to coiling axis, from Bouholoup;
figs. 15-16: subequatorial sections, from Vaquemorte south of Montbrun.
, figs. 10-22;
figs. 6-9: oblique sections progressively more parallel to the coiling axis; figs. 10-16: Specimens from different places in the Petites Pyrénées, Aquitaine, collected by J. & Y. . SBZ 2; fig. 10: oblique section, from S'ours; figs.
11-12: oblique and axial sections, from Bouholoup; fig. 13: oblique-centered section, from
S'ours; fig. 14: transverse section perpendicular to coiling axis, from Bouholoup;
figs. 15-16: subequatorial sections, from Vaquemorte south of Montbrun.
Abbreviations: f: foramen; s: septum; spc: spiral canal; u: umbilicus with some funnel-canals; up: umbilical plate.
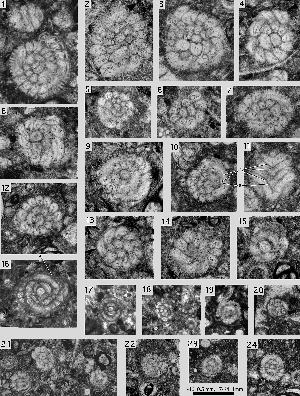
Click on thumbnail to enlarge the image.
Plate 22. Miscellanites globularis (, 1978). All from Haimana, Central Anatolia, Turkey. SBZ 2 or 3; figs. 1, 4, 6 & 12-13: equatorial sections; figs. 11 & 14: tangential sections; fig. 17: axial section; all other sections are more or less oblique, many centered.
Abbreviations: f: foramina; s: septum.
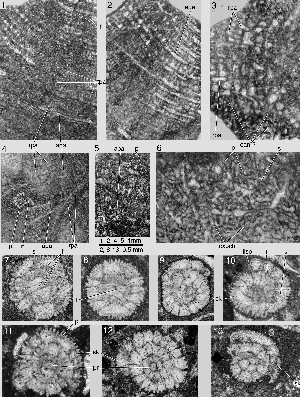
Click on thumbnail to enlarge the image.
Plate 23. Bolkarina aksarayi , 1981; figs. 1-6: all from Haymana, central Anatolia, Turkey. SBZ 2 or 3; figs. 1-3: sections in or near to the equatorial plane; fig. 4: equatorial section with spiral nepiont; figs. 5-6: tangential section of lateral skeleton parallel to the lateral surface of the discoidal shell showing several successive expanse chambers.
Miscellanites globularis (, 1978); figs. 7-13: Haymana, central Anatolia, SBZ 2 or 3; figs. 7-8: tangential sections; figs. 9-10 & 12-13: more or less perfect equatorial sections; fig. 11: subaxial section.
Abbreviations: apa: annular passage (in preseptal position); can ?: canal system orifices ?; expch: expanse chamber; f: foramen; ilsp: interlocular intraseptal space; n: nepiont (end of nepionic spiral); p: pores; pr: proloculus; rpa: radial passage; s: septum; sk: supplemental skeleton (as revealed by a simple canal system); tpa: transverse tubular passage.
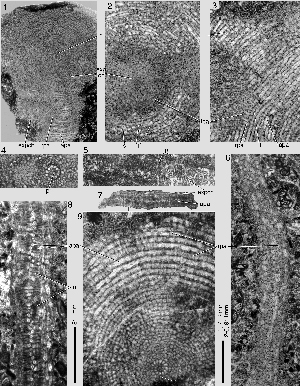
Click on thumbnail to enlarge the image.
Plate 24. Bolkarina aksarayi , 1981. From Haymana, central Anatolia, Turkey. SBZ 2 or 3; fig. 1: equatorial section, overview with spiral nepiont; fig. 2: section perpendicular to the axis of the shell tangentially cutting the early neanic expanse chambers and the annuli in the equatorial plane in later neanic stages of growth; fig. 3: detail of equatorial section showing the intercameral foramina between successive annuli and the lateral connections from the annular to the dome-shaped, expanded parts of the chambers; fig. 4: equatorial section of a nepiont; fig. 5: oblique section with a few annular chambers; fig. 6: oblique section near to involute nepiont; fig. 7: almost centered axial section showing involute spiral nepiont; overview; fig. 8: axial section; details of nepionic involute chamber arrangement and crossections of the annuli; fig. 9: equatorial section with nepiont showing the linear-radial pattern of the passages from the annular to the expanded parts of the chamber.
Abbreviations: apa: annular passage (in preseptal position); expch: expanse chamber; f: foramen; n: nepiont (end of nepionic spiral); p: pores; pr: proloculus; rpa: radial passage; s: septum; tpa: transverse tubular passage.
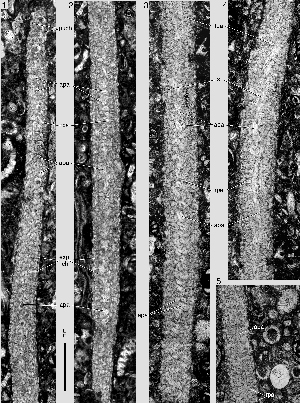
Click on thumbnail to enlarge the image.
Plate 25. Bolkarina aksarayi , 1981. From Haymana, central Anatolia, Turkey. SBZ 2 or 3; figs. 1-2 & 5: Sections parallel to the axis of the shell in more or less distance from the center; figs. 3-4: Sections progressively inclined in respect to the axis of the shell, far from the center. Note in 5 top right the tangential section of Miscellanites globularis (, 1978), possibly the megalospheric generation of Bolkarina aksarayi.
Abbreviations: apa: annular passage (in preseptal position); expch: expanse chamber; puch: penultimate (expanse) chamber; rpa: radial passage; s: septum; tpa: transverse tubular passage.