![]()
Contents
[1. Introduction] [2. Studied material]
[3. Quantitative method]
[4. Size variations of vestibula on Krithe gnoma] [5. Remarks]
[Bibliographic references] and ... [Figures]
* Corresponding author
UnB - Universidade de Brasília, Instituto de Geocięncias, Brasília - DF, CEP 70919-970 ( Brazil)
UnB - Universidade de Brasília, Instituto de Geocięncias, Brasília - DF, CEP 70919-970 (Brazil)
UnB - Universidade de Brasília, Instituto de Química, Brasília - DF, CEP 70910-900 (Brazil)
UnB - Universidade de Brasília, Instituto de Química, Brasília - DF, CEP 70910-900 (Brazil)
Manuscript online since December 31, 2009
![]()
The species Krithe gnoma was described from Holocene sediments on the continental margin of Brazil, where it occurs on both shelf and slope. The shelf distribution of this species is restricted to those areas influenced by the Malvinas current between 32°11' and 22°31'S. K. gnoma was selected due to the excellent resolution of images of the anterior vestibule. So the range in size of the vestibula of this species was used to determine whether or not the dimensions of the anterior vestibule were to some degree related to amount of dissolved oxygen in the water at the sampling site. Female valves of the 7th and 8th instars were measured and their sizes and those of the anterior vestibule were compared. These valves and the reference carapace were collected at six localities in the southern shelf area. The anterior vestibule of right and left valves of the 7th instar and one of the 8th instar from discrete localities were measured. In the reference carapace, the larger left valve also has the larger vestibule. A gerontic 8th instar has valves larger than those of the previous instars, but its vestibula are smaller than those of the 7th instar. The vestibula of the left valves of the 7th instar collected at discrete localities showed consistent differences in size related to the oxygen content of the sampling site. The relationship between vestibule size and oxygen content is inversely proportional. However, the poor correlation factor (R2<<1) of these results are not precise enough to support 's hypothesis. But the size of the vestibula of K. gnoma does show some degree of relationship to oxygen content. So it is important to measure the vestibula of species of Krithe using the procedure described here. Rigorous application of this procedure will eliminate the possibility of discrimination based on variations in size due to ontogeny and dimorphism leaving only those related to actual valve size. In any case, variations in the size of vestibula may have a phenotypic origin.
Ostracoda; Krithe gnoma; size variation of vestibula; 's hypothesis; Malvinas current.
D.A., R.P., P.A.Z. & V.M. (2009).- Size variations of the vestibula of Krithe gnoma & , 1999 (Ostracoda): a new procedure for their analysis.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2009/09 (CG2009_A09)
Variaçőes de tamanho do vestíbulo de Krithe gnoma & , 1999 (Ostracoda): novo procedimento para análise.- A espécie Krithe gnoma foi descrita a partir de sedimentos do Holoceno da margem continental do Brasil onde a mesma ocorre da plataforma ao talude. Nesta margem continental, a distribuiçăo desta espécie é restrita a áreas de influęncia da corrente das Malvinas entre latitudes de 32°11'/22°31'S. K. gnoma foi selecionada para ser utilizada no presente estudo devido a excelente resoluçăo alcançada no imageamento do vestíbulo anterior. Desta forma, K. gnoma foi utilizada para medir as diferenças de tamanho do vestíbulo e para avaliar a relaçăo entre o tamanho do vestíbulo e a quantidade de oxigęnio dissolvido na água do mar nas áreas de coleta. Valvas de fęmeas adultas foram comparadas e analisadas através das mediçőes dos tamanhos do vestíbulo anterior. Os resultados mostram que o vestíbulo das valvas direita e esquerda de uma mesma carapaça săo distintos. Neste caso, a valva maior apresenta também o maior vestíbulo. Um estágio gerôntico do 8°. Instar e, por isso com tamanho de valva maior do que aqueles das outras valvas do 7°. Instar, apresenta o menor tamanho do vestíbulo. Com a análise das valvas esquerdas do 7°. Instar, coletadas em distintas localidades, tornou-se aparente que existiam diferenças consistente no tamanho do vestíbulo. Neste caso, foi possível observar uma relaçăo inversamente proporcional entre o tamanho do vestíbulo o conteúdo de oxigęnio. No entanto, considerando a pobreza do fator de correlaçăo (R2<<1), estes resultados năo săo válidos para se avaliar a hipótese de . Mesmo assim, está claramente mostrado que há variaçőes do vestíbulo de K. gnoma. Neste sentido, vale salitentar que é importante avaliar as variaçőes do vestíbulo em espécies de Krithe seguindo o procedimento aqui sugerido. Pois, desta forma pode-se evitar variaçőes de tamanho devido a ontogenia, dimorfismo e aqueles relacionados a tamanhos de valvas distintos. Estes resultados mostram que variaçőes do vestíbulo săo potencialmente de natureza fenotípica.
Ostracoda; Krithe gnoma; variaçőes do tamanho de vestíbulo; hipótese de ; corrente das Malvinas.
Nouvelle procédure pour analyser les variations des tailles de vestibules chez Krithe gnoma & , 1999 (Ostracoda).- L'espčce Krithe gnoma a été décrite dans des sédiments de l'Holocčne de la marge continentale du Brésil, oů elle est présente de la plate-forme au talus. Le long de cette marge continentale, la distribution de cette espčce sur la plate-forme est restreinte aux régions influencées par le courant des Malouines ŕ des latitudes comprises entre 32°11' et 22°31'S. K. gnoma a été choisie pour cette étude en raison des excellents résulats apportés par la forme du vestibule antérieur. On a mesuré les différentes tailles de vestibule et évalué la relation entre la taille de celui-ci et la concentration en oxygčne des eaux océaniques sur les sites d'échantillonnage. Les valves des femelles adultes ont été analysées en mesurant et comparant la taille de leur vestibule antérieur. Les résultats sont basés sur l'observation du vestibule des 2 valves de carapaces d'individus appartenant tous au stade ontogénétique 7 dans des sites différents. Le vestible le plus grand se trouvant sur la valve gauche, la plus grande. Un stade ontogénétique 8, de taille supérieure, a été observé montrant un vestibule plus petit que chez le stade 7. Aprčs avoir analysé les vestibules des valves gauches des stades 7 recoltés dans des sites différents, il est apparu que des différences de tailles notables existaient. Dans ce cas, il a été possible d'observer une relation inversement proportionelle entre la taille du vestibule et la concentration en oxygčne. Compte tenu du faible facteur de corrélation (R2<<1), ces résulats ne permettent pas d'évaluer l'hypothčse de . Néanmoins, des variations de taille du vestibules de K. gnoma ont été clairement observées. Il est important d'analyser les vestibules des espčces de Krithe en utilisant la procédure présentée dans ce travail. En suivant ce protocole, on évite l'effet des variations de taille due ŕ l'ontogénie, le dimorphisme ou d'autres facteurs. Les résultats de cette étude montrent que les variations de la taille du vestibule sont potentiellement d'origine phénotypique.
Ostracodes ; Krithe gnoma ; variation de la taille des vestibules ; hypothčse de ; courant des Malouines.
The continental shelf of Brazil includes three associations of ostracodes: Southern, Transitional and Northern/Eastern ( & , 1989). On this huge shelf, Krithe gnoma & , 1999, is restricted to that portion of the Southern association under the influence of the Malvinas current ( & , 1999).
In order to evaluate size differences in the size of the
anterior vestibule specimens of K. gnoma were
sampled from localities where information was available about bathymetry and the
level of dissolved oxygen in ambient water. These
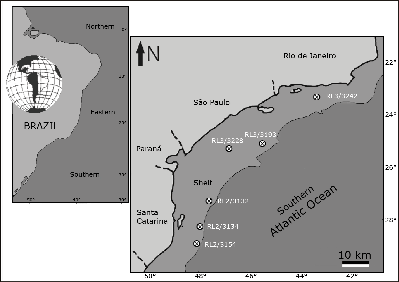
factors restrict the area studied to the shelf and slope of a tract bounded by 28°23'S and 23°28'S,
and 47°34'W and 43°17'W
(Fig. 1 ![]() ). The physical-chemical parameters of the areas are based on oceanographic cruises performed by
the Brazilian Navy (Anonymous, 1969, 1970,
1972; & ,
1977).
Here we present a procedure for the analysis of size variation of vestibula presumed
to have been caused by phenotypic plasticity. We also discuss the relationship between
the quantity of oxygen dissolved in sea water and the size of the vestibula of K. gnoma.
). The physical-chemical parameters of the areas are based on oceanographic cruises performed by
the Brazilian Navy (Anonymous, 1969, 1970,
1972; & ,
1977).
Here we present a procedure for the analysis of size variation of vestibula presumed
to have been caused by phenotypic plasticity. We also discuss the relationship between
the quantity of oxygen dissolved in sea water and the size of the vestibula of K. gnoma.
K. gnoma was selected for analysis of size variations in vestibula after a detailed taxonomic study of this species' structure. Female and male instars are easily recognized based on valve and carapace morphology. K. gnoma occurs in the southern shelf areas of Brazil in different latitudes and bathymetries, dependent to a large degree on the influence of the Malvinas current on temperature and salinity ( & , 1995, 1999). K. gnoma ranges from Late Eocene to the Miocene - Holocene ( et alii, 2002). Understanding the relationship between the ecology and morphology of this species and basic environmental variables, like dissolved oxygen, is important for these features are fundamental to palaeoenvironmental reconstructions.
(1975) recognized variations in the size of the vestibula of Krithe and Parakrithe, and was the first to note an inverse proportional relationship between the size of the vestibule and the oxygen content of ambient sea water. This relationship was later named 's hypothesis and was tested by et alii (1989), (1990) and et alii (1993).
et alii (1989), using quantitative methods based on multivariate analysis, tested this hypothesis for the first time. Based on digital images, valves of Krithe praetexta praetexta (, 1866) were measured in order to estimate the areas of the vestibula. Through canonical analyses, a total of four groups of valves were identified, two for each sex. The vestibular area of each group remained unchanged, although oxygen content differed significantly with locality. (1990), after analysing soft tissues of other genera, failed to find a cellular or histological mechanism for the storage of oxygen in vestibula. et alii (1993) discussed 's hypothesis, suggesting that it should be rejected based on studies of the ecology, morphology and distribution of many species of Krithe. Supporting their position, no respiratory function for the vestibule has been identified and no difference in morphology has been observed between cells that comprise the vestibule and those making up other portions of the carapace.
Taxonomic data on the genus Krithe and morphology of their vestibula were considered useful for distinguish species ( et alii, 1994). Considering the influence of temperature on relative numbers, et alii (1996) present the ratios of Mg/Ca and Sr/Ca in Krithe from upper Quaternary strata in the Arctic Ocean. Their development had been markedly increased by a lower water temperature. For bottom water shallower than 900m, low Mg/Ca ratios in their carapaces correlate with periods of relative cold, when the genus Krithe is dominant among the ostracodes. et alii (1999) studied Mg/Ca ratios and the dominance of Krithe in strata of the upper Quaternary in the deep sea portion of the North Atlantic Chain 82-24. They recognized that the dominance of the genus Krithe is clearly related to the glacial maximum and the corresponding low Mg/Ca ratios.
& (1999) present remarkable data on the diversity of species of the genus Krithe recovered from the Florida-Hatteras slope of the North Atlantic. A brief history concerning the vestibula of Krithe is included. Note that only & (1992) give data corroborating 's hypothesis, along with an evaluation of the vestibula of the species of Krithe. Vestibule shape and size in a range of bathymetries were analyzed by & (op.cit.). Their study does not support 's hypothesis. Three major points militate against it. First, species with both large and small vestibula were recovered from all sites when the hypothesis postulates a zonation in size depending on the depth of the sea-floor. Second, occurrences of Krithe pernoides with a relatively small vestibule are found when the opposite would be expected if 's hypothesis were correct in the OMZ (Oxygen Minimum Zone). Finally, large vestibula were found in specimens from the glacial interval, when if 's hypothesis were well-founded they should be smaller.
et alii (2000) present intriguing results based on thermocultures evaluating the effect of temperature on the growth rate of Krithe praetexta praetexta. They show that at lower temperatures (5° C), the average size of a specimen is greater than that of those reared at higher temperatures (14° C).
After studying recent adult specimens of Leptocythere lacertosa, & (2009) gave an oral presentation during the 16th International Symposium on Ostracoda - ISO - held in Brasília (Brazil). They reported that vestibula are not formed in the first 30 hours after ecydisis but were completely formed 60 hours after ecydisis. In conclusion, the vestibular origin and stages of development took place in steps, even in adults. On the other hand, the shapes of vestibula and their pore canals were found to be valid criteria for taxonomic purposes.
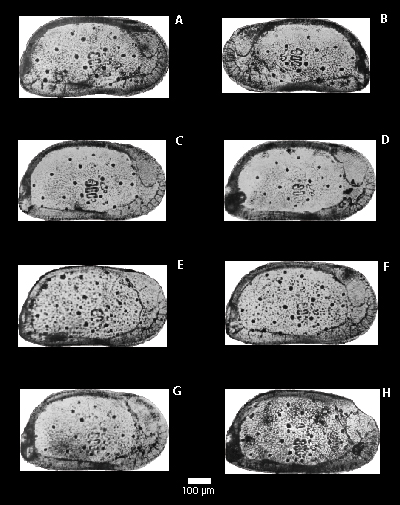
In K. gnoma, the left valve is the larger of the two, as is common in other members of this genus. To analyze and quantify the variability of vestibula in K. gnoma, the reference selected was an adult carapace, 7th instar
(sample 3184). It is important to note that both valves were considered
in the analysis. The remaining five left valves of 7th instar (samples 3193, 3132, 3228, 3242,
and 3279) and one from the 8th instar (sample 3154) were used for comparisons of
the size of vestibula. This one left valve of 8th
instar, probably gerontic, was used to determine whether or not an older and larger instar would
have a comparatively larger vestibule
(Fig. 2 ![]() ).
).
Vestibule areas of left and right valves were measured using digital images. Image distortions were avoided by imaging each valve following exactly the same procedure, internal view, magnification, illumination,
etc.
(Fig. 2 ![]() ). The anterior vestibule was chosen for study due its excellent resolution in imaging. To
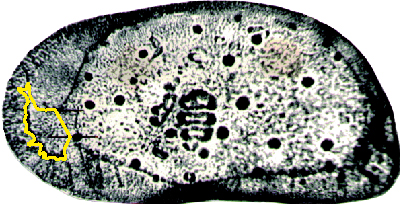
delimit and standardize the area of the vestibule of each sample, two tangent lines in the neck
of the vestibular area were used (Fig. 3
). The anterior vestibule was chosen for study due its excellent resolution in imaging. To
delimit and standardize the area of the vestibule of each sample, two tangent lines in the neck
of the vestibular area were used (Fig. 3 ![]() ). Before measuring the
area of a vestibule, it was necessary to select an adult female's instars and valve (right or left) to be used to avoid size variations due to ontogeny, differences of right and left valves, or dimorphism.
). Before measuring the
area of a vestibule, it was necessary to select an adult female's instars and valve (right or left) to be used to avoid size variations due to ontogeny, differences of right and left valves, or dimorphism.
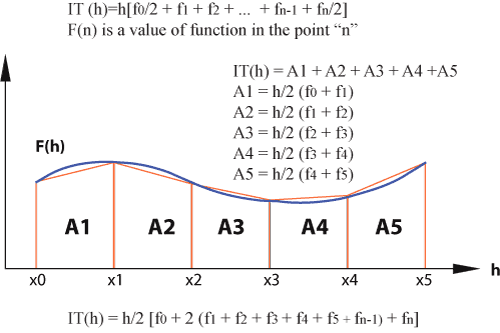
The method was used to measure
the areas of
vestibula. Usually, this method is used to calculate irregular areas. It consists of iterating the results calculated for an integral through the trapezium method for different "h". These "h" must belong to a geometric progression divided by two. For the trapezium method, the inferior area is divided in a series of trapeziums, whose base is "h" and height is the value of a function in a given point,
f(h) (Fig. 4 ![]() ). Today, there is sophisticated software to measure areas of carapace as well as to calculate
the areas of vestibula using digital images.
). Today, there is sophisticated software to measure areas of carapace as well as to calculate
the areas of vestibula using digital images.
The size of the vestibule is different in the right and left valves of the carapace used as a reference. However, the left (and larger) valve had the bigger vestibule (Table 1). The area of the vestibula measured in several left valves averaged 87.37 µm, with a standard deviation of 10.64 µm, thus representing a variation of 12.77 %.
The left valve of gerontic 8th instar (sample 3154), the largest carapace studied, had the smallest vestibule; it is 38.35 % smaller than the average of the left valves of 7th instars (Table 1). The two left valves of the 7th instar (sample 3228) differ in size; one is 9.58 % larger than the other. The left and right valves of the reference carapace (sample 3134) have a proportional relationship between the area of the vestibula and the size of the valves. Consequently, the larger left valve has the bigger vestibule.
Table 1: Krithe gnoma & ,
1999. Oceanographic data [Depth (m), Temperature (°C), Salinity (‰), Oxygen
(ml), Size of Vestibule (μm2) and Instar] (Anonymous, 1972).
Krithe gnoma & ,
1999. Dados oceanográficos [Profundidade
(m), Temperatura (°C), Salinidade (‰), Oxigęnio (ml), tamanho do vestíbulo
(µm2) e estágio] (Anônimo,
1972).
Krithe gnoma & ,
1999: Données
océanographiques [Profondeur (m), Température (°C), Salinité (‰), Oxygčne
(ml), Taille du vestibule (μm2) et stade ontogénétique] (Anonyme,
1972).
To evaluate the vestibule area in relation to oxygen content, salinity and depth, several curves and lines were examined to determine the procedure that presented the data best (straight line, exponential and logarithm curves). Using the least square method, a straight line was found to be the most appropriate to portray the relationship between vestibular size and oxygen content, temperature, salinity and other factors.
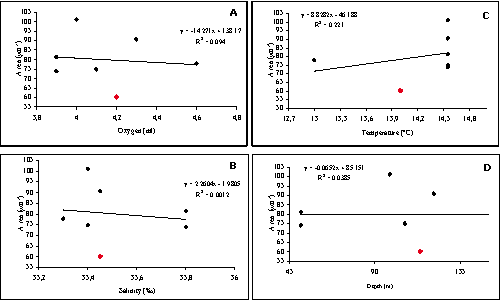
The results presented here show that variations in size of the vestibula have no clear relationship to oxygen content (R2=0.06047), temperature (R2=0.02814), depth (R2=0.04932) and salinity (R2=0.01735). On the other hand the data show that the vestibula range considerably in size even when several intrinsic biologic factors are taken into consideration: dimorphism, ontogeny and dfferences in the dimensions of left and right valves.
So, as regards the influence of oxygen content on vestibule size these findings do not corroborate this theory, but do not refute it either. Therefore, additional studies should be made to determine whether or not the size of the vestibula in K. gnoma and other species of Krithe is controlled to any degree by oxygen content, temperature, salinity or any other factor.
Variation in vestibular size merits further discussion. The 7th instar reference carapace has the largest vestibule in the largest valve. This suggests a proportional relationship between carapace and vestibule size. On the other hand, the left valve of the 8th instar is even larger but has the smallest vestibule of all the reference specimens. Nevertheless, one may conclude that in any one carapace the vestibula are always larger in the left valve.
When considering a vestibular analysis of Krithe, several steps are required: 1. Selection of the species to analysed, 2. Choice of an instar and the valve (left or right), and the gender to be examined in order to avoid size variations caused by ontogeny and dimorphism and 3. Selection and standardization of the portion of the vestibule to be analyzed.
In summary, the vestibule-oxygen relationship proposed by is not clearly
verified
here (R2 < 0.1). The result providing the most support was
obtained using a cross-plot of vestibular surface versus water temperature. Because
K. gnoma has
a broad distribution stratigraphically (Late
Eocene/Miocene in the Pelotas and Santos basins), these
supportive results suggest that evaluating such relationships may be found
useful in paleoceanographic studies as indicators - "proxies" - of temperate or cold
water inferences regarding
temperature (temperate or cold, Fig. 5 ![]() ). However,
in view of the polemic nature of the subject, future studies
should involve a larger number of specimens and include other species of Krithe
while carrying out the procedures presented here.
). However,
in view of the polemic nature of the subject, future studies
should involve a larger number of specimens and include other species of Krithe
while carrying out the procedures presented here.
If it is true that the development size of species of Krithe is affected by the temperature of oceanic waters ( et alii, 1996, 1999; et alii, 1999), it would be interesting to conduct experiments to evaluate how and why the size of the vestibule varies in the several species attributed to this genus.
Thanks to & (2009), a methodology to study the origin and development of vestibula has been introduced. It would be interesting to combine the experiment conducted by et alii (2000) with the methodology introduced by & (op.cit.). This experimental might make it possible to determine whether or not the pattern recognized for L. lacertosa holds for Krithe praetexta praetexta. Also in thermocultures of Krithe praetexta praetexta, study of vestibular sizes and isotope ratios would be very useful.
The first author is grateful to CNPq (Brazilian Research Council) and to FINATEC (Fundaçăo de Empreendimentos Científicos e Tecnológicos) for financial support with respect to his participation on the 15th International Symposium on Ostracoda. Special thanks to Luiz Filipe , a chemical engineer and an old friend for support in programming the calculator to optimize measurements. P.A.Z. and the first author thank CNPq for their grants. Special thanks to Nestor who kindly corrected our English in the final version. Thanks are also due Pierre , Thomas and Bruno for reviewing the manuscript.
This paper was first presented at the 22nd ROLF (Réunion des Ostracodologistes de Langue Française) held in Brussels (June 2-4, 2008) and organized by J.-G. (Institut royal des Ssciences naturelles de Belgique, Brussels) and J.-P. (Cestas, France, and Faculdade de Cięncias, Universidade de Lisboa, Portugal).
Anonymous (1969).- Comissăo Oceanográfica, 2, NE Almirante Saldanha (15 a 28/02/1957).- Marinha do Brasil, Diretoria de Hidrográfia e Navegaçăo, Niterói (RJ), 80 p.
Anonymous (1970).- Comissăo Oceanográfica, 30, N.Oc. Almirante Saldanha (06/06 a 18/07/1966).- Marinha do Brasil, Diretoria de Hidrográfia e Navegaçăo, Niterói (RJ), 80 p.
Anonymous (1972, Ediçăo Provisória).- Brasil, Marinha do Brasil, Diretoria de Hidrográfia e Navegaçăo. 1972. Atlântico Sul. Atlas Oceanográfico: Costa Sul do Brasil - condiçőes de verăo de 1972. N.Oc. "Almirante Saldanha". Operaçăo Sul 2, janeiro-fevereiro/1972.- Marinha do Brasil, Diretoria de Hidrográfia e Navegaçăo, Niterói (RJ), 80 p.
J.-P. & O.O. Jr (1977).- Sobre as condiçőes oceanográficas do Rio Grande do Sul.- Atlântica, Rio Grande, vol. 2, n° 2, p. 25-110.
H.A.F. (1983).- Introduçăo Geral. In: PROJETO REMAC: Processo e métodos. Relatório final.- PETROBRAS/SINTEC, Rio De Janeiro, (Série Projeto REMAC), n° 6, p. 1-32.
J.C. & L.P. (1989).- Distribution and ecology of subrecent Orionininae (Ostracoda) in the Brazilian continental shelf.- Revista Brasileira de Geocięncias, Săo Paulo, vol. 19, n° 2, p. 177-186.
G., R. & A. (1994).- The ostracode genus Krithe from the Tertiary and Quaternary of the North Atlantic.- Palaeontology, London, vol. 37, part 1, p. 71-120.
T.M., D.M., G.S., J. (1999).- Deep-sea ostracode species diversity response to late Quaternary climate change.- Marine Micropaleontology, Amsterdam, vol. 37, n° 3, p. 231-249.
T.M., G.S., P.A., J. & W.M. Jr (1996).- Deep-sea ostracode shell chemistry (Mg:Ca ratios) and late Quaternary Artic Ocean history.- Geological Society, London, Special Publications, vol. 111, p. 117-134.
D.A. & Y.T. (1995).- Krithe occurrence on the Brazilian continental margin, an ecological approach. In: J. (ed.), Ostracoda and biostratigraphy.- Balkema Publishers, Rotterdam, p. 407-412.
D.A. & Y.T. (1999).- Taxonomy and palaeoceanographical significance of the Krithe genus in the Brazilian continental margin.- Journal of Micropaleontology, London, vol. 18, n° 2, p. 111-123.
I.A. (1978).- Numerical analysis for computer science.- Elsevier, Amsterdam, p. 431-473.
J.P. (1982).- Marine geology.- Englewood Cliff, Prentice-Hall, New Jersey, 813 p.
D. (1990).- Morphological changes and function of the inner lamella layer of podocopid Ostracoda. In: R.C. & C. (eds.), Ostracoda and global events.- Chapman and Hall, London, p. 401-410.
S., S. & M. (2000).- The effect of temperature on shell and growth rate in Krithe praetexta praetexta ().- Hydrobiologia, Amsterdam, vol. 419, n° 1, p. 141-148.
K.G., S., V. & R.A. (1989).- The Krithe problem - first test of 's hypothesis, with redescription of Krithe praetexta praetexta (Crustacea, Ostracoda).- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 74, n° 3-4, p. 343-354.
C.G., M. & D.A. (2002).- Fish and ostracods remains from the Santos basin (Cretaceous to Recent), Brazil.- Geological Journal, Liverpool, vol. 37, n° 4, p. 297-316.
J.-P. (1975).- Les variations des caractčres morphologiques internes chez les Ostracodes des genres Krithe et Parakrithe : relation possible avec la teneur en oxygčne dissous dans l'eau.- Bulletin de l'Institut de Géologie du Bassin d'Aquitaine, Talence, n° 17, p. 81-88.
J.-P. (1977).- Les ostracodes et la connaissance des paléomilieux profonds. Applications au Cénozoďque de l'Atlantique nord-oriental.- Thčse d'État, Université de Bordeaux 1, 433 p.
J. & T.M. (1999).- Quaternary glacial and deglacial Ostracoda in the thermocline of the Little Bahama bank (NW Atlantic): Palaeoceanographic implications.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 152, n° 3-4, p. 339-364.
M.I. (1984).- Aspectos da oceanografia física do Atlântico Sul.- Pesquisas em Geocięncias, Porto Alegre, vol. 16, p. 76-90.
R.C. & Z. (1993).- The Krithe problem - a case history of the distribution of Krithe and Parakrithe (Crustacea, Ostracoda) in the South China Sea.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 103, n° 3-4, p. 282-297.
S. & D. (2009).- Calcification of the marginal infold in podocopid ostracods. In: Biostratigraphy and applied ecology. 16th International Symposium on Ostracoda, Brasília, (July 26-30, 2009 - Brazil).- Abstract Volume, p. 8.
B. & N. (1992).- Three species of Krithe (Crustacea : Ostracoda) from Suruga Bay, Central Japan.- Transactions and Proceedings of the Palaeontological Society of Japan, Tokyo, (new series), n° 166, p. 1097–1115.
Click on thumbnail to enlarge the image.
Figure 1:
Map of sampling locations (adapted from , 1983; & ,
1989).
Mapa de localizaçăo das amostras (adaptado de ,
1983; & ,
1989).
Carte des sites d'échantillonage (adaptée de ,
1983 ; & ,
1989).
Click on thumbnail to enlarge the image.
Figure 2:
Specimens of Krithe gnoma. Valves used for measurement of the area of the vestibule.
A-B: carapace base (R.T.2/3134), 7th instar; A: Left valve, B: Right valve; C-G: Left valve, 7th instar;
C: (R.T.3/3193), D: (R.T.2/3132), E-F: (R.T.3/3228), G: (R.T.3/3242), H: (R.T.2/3154) 8th instar,
male.
Espécimes de Krithe gnoma. Valvas utilizadas para medir a área do
vestibulo. A-B: carapace de referęncia (R.T.2/3134), 7° estágio
ontogenético; A: valva esquerda, B: valva direita; C-G: valvas esquerdas, 7° estágio
ontogenético; C: (R.T.3/3193), D: (R.T.2/3132), E-F: (R.T.3/3228), G: (R.T.3/3242),
H: (R.T.2/3154) macho
adulto.
Spécimens de Krithe gnoma. Valves utilisées pour la
mesure de la superficie du vestibule. A-B : Carapace de référence (R.T.2/3134),
7čme stade ; A : Valve gauche, B : Valve droite; C-G : Valve gauche, 7čme stade ;
C :
(R.T.3/3193), D : (R.T.2/3132), E-F : (R.T.3/3228), G : (R.T.3/3242),
H :
(R.T.2/3154) 8čme stade, mâle adulte.
Click on thumbnail to enlarge the image.
Figure 3:
Bounds used to define and delimit the area of the anterior vestibule (sample R.L.2 /3134).
Área utilizada como o limite do vestíbulo (amostra R.L.2/3134).
Surface utilisée pour délimiter le vestibule (échantillon
R.L.2/3134).
Click on thumbnail to enlarge the image.
Figure 4:
Trapezium method for calculating area.
Método para o cálculo do trapézio.
Méthode
du trapčze.
Click on thumbnail to enlarge the image.
Figure 5:
Areas of the vestibule in Krithe gnoma & ,
1999. In black
diamonds the measurements of six left valves of 7th instar [A: Oxygenation (ml) B: Salinity (‰) C: Temperature (°C) D: Depth (m)]
(Anonymous, 1969, 1970,
1972).
Tamanho do vestibulo de Krithe gnoma & ,
1999. Os pontos pretos indicam as seis valvas esquerdas adultas [A: Oxigęnio (ml) B: Salinidade (‰) C: Temperatura (°C) D: Profundidade (m)]
(Anônimo, 1969, 1970,
1972).
Taille du vestibule de Krithe gnoma & ,
1999. Points noirs : six valves du 7čme stade [A: Oxygčne (ml) B: Salinité (‰)
C: Température(°C) D: Profondeur (m)] (Anonyme, 1969,
1970, 1972).