◄ Carnets Geol. 13 (L04) ►
![]()
[Introduction]
[Geological setting]
[Systematics] [Bibliographic references] and ... [Plates]
Lerchenauerstr. 167, D-80935 München (Germany)
Department of Geology and Center for Integrated Geological
Studies, Babeş-Bolyai University, Kogălniceanu 1,
Ro-400084 Cluj-Napoca (Romania)
Department of Geology, Payame Noor University, PO Box 19395-3697 Tehran (Iran)
Ferdowsi University of Mashhad, International Branch, Mashhad (Iran)
Manuscript online since August 2, 2013
[Scientific and technical
editor: Bruno ;
language editor: Simon F. ]
![]()
The new orbitolinid foraminifer, Praeorbitolina claveli n.sp., is described from the Lower Aptian (Bedoulian) of Central Iran. It is characterized by an eccentric embryonic apparatus displaying both a subdivided subembryonic zone and a deuteroconch. The Barremian-Aptian orbitolinid association of this area has a typical northern Tethyan character (e.g., "association à Valserina" of and , 1973).
Foraminifera; orbitolinids; Praeorbitolina; Mesorbitolina; Lower Aptian; Bedoulian; Iran; palaeobiogeography.
F., I.I., K. & B. (2013).- Praeorbitolina claveli n.sp. (benthic Foraminifera) from the Lower Aptian sensu lato (Bedoulian) of Central Iran.- Carnets de Géologie [Notebooks on Geology], Brest, Letter 2013/04 (CG2013_L04), p. 255-272.
Praeorbitolina claveli n.sp. (foraminifère benthique) de l'Aptien inférieur sensu lato (Bédoulien) du centre de l'Iran.- Un nouveau foraminifère orbitolinidé, Praeorbitolina claveli n. sp., est identifié dans l'Aptien inférieur (Bédoulien) de l'Iran central. Il se caractérise par un appareil embryonnaire constitué d'une zone subembryonnaire subdivisée ainsi que d'une déutéroconque. L'association d'orbitolinidés d'âge Barrémien-Aptien de cette région présente un caractère typiquement nord-téthysien (e.g., "association à Valserina" de et , 1973).
Foraminifère ; Orbitolinidae ; Praeorbitolina ; Mesorbitolina ; Aptien inférieur ; Bédoulien ; Iran ; paléobiogéographie.
Lower Cretaceous orbitolinid foraminifers were reported from different structural regions of Iran, e.g., the Zagros Zone in the south, Central Iran and the northern part (Kopet Dagh, Alborz) (e.g., , 1948; , 1965; , 1965; , 1969; , 1973; et al., 2000; et al., 2010; et al., 2010; & , 2010; & , 2013). Currently, several sections of Lower Cretaceous orbitolinid-bearing, shallow-water carbonates from Central Iran are being investigated with respect to their benthic foraminifers and calcareous algae. In the present contribution, a new species of orbitolinid foraminifer is described.
The mountainous region around Anarak is part of the Yazd Block
of Central Iran ( & ,
1981; 2004, for
overviews). It is bounded northward by the Doruneh fault and towards the west by the Naien Baft
ophiolite belt. Trangressive Lower Cretaceous shallow-water limestones overlie
conglomerates and sandstones which in turn are underlain by the Late Paleozoic
to Triassic Anarak metamorphic complex (e.g., ,
2011; et al.,
2013). The
Lower Cretaceous carbonates of the Yazd Block that are generally assigned to the Barremian-Aptian interval were investigated with respect
to microfacies, sedimentology and micropaleontology by et al.
(2012) and et al.
(2013). Three
sections of Lower Cretaceous, shallow-water carbonates are currently being studied in the area
of Anarak-Naien-Ardakan
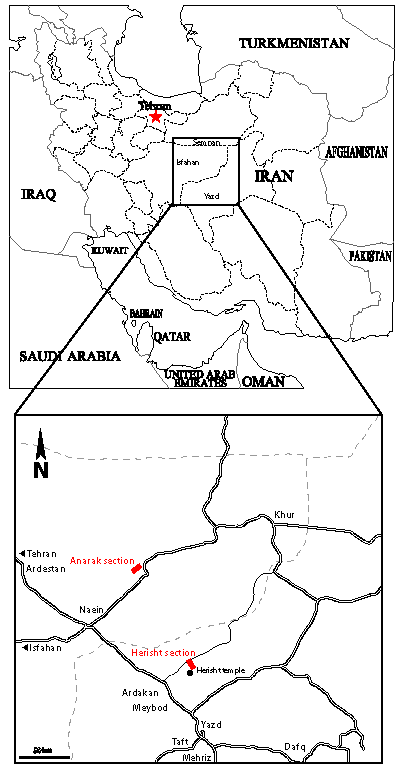
(Fig. 1 ![]() ): the Anarak section near Anarak and two
sections at Herisht (T and H) close to Ardakan. Orbitolinid foraminifers represent frequent
constituents of the carbonates. As the first result of the ongoing studies, a new species is described as Praeorbitolina claveli n.sp. herein.
): the Anarak section near Anarak and two
sections at Herisht (T and H) close to Ardakan. Orbitolinid foraminifers represent frequent
constituents of the carbonates. As the first result of the ongoing studies, a new species is described as Praeorbitolina claveli n.sp. herein.
Click on thumbnail to enlarge the image.
Figure 1: Location of the studied sections at Anarak and Herisht in Central Iran. The Herisht section is the type-locality of the new species Praeorbitolina claveli n.sp. Anarak section: N 33°20'22.66", E 53°41'53.68"; Herisht section: N 32°25'37.61", E 54°6'54.11''.
Family Orbitolinidae , 1890
Subfamily Orbitolininae , 1890
Genus Praeorbitolina , 1964a
Type-species: Praeorbitolina cormyi , 1964a
The new foraminifer is attributed to the genus Praeorbitolina , 1964a, which is defined essentially by its embryonic apparatus composed of a protoconch, a deuteroconch and the subembryonic zone situated in an eccentric position at the beginning of a small spire ( 1964a, 1975; et al., 1998; et al., 2010).
Praeorbitolina claveli n.sp.
Pl. 1 ![]() ,
figs. 1-17
,
figs. 1-17
Synonymy:
1973. Praeorbitolina sp. , Figs. 11-13.
1992. Praeorbitolina wienandsi et al., Pl. 2, fig. 11.
Origin of the name: Dedicated to Bernard for his numerous contributions to the stratigraphy and systematics of the orbitolinids.
Holotype: Pl. 1 ![]() ,
fig. 4. Axial section, cut perpendicular to the median plane and passing
through the embryonic apparatus; from the Herisht section, thin-section T 47 (number 23868),
Lower Aptian (Bedoulian), Praeorbitolina cormyi orbitolinid zone
of et al. (2010; &,
2013).
,
fig. 4. Axial section, cut perpendicular to the median plane and passing
through the embryonic apparatus; from the Herisht section, thin-section T 47 (number 23868),
Lower Aptian (Bedoulian), Praeorbitolina cormyi orbitolinid zone
of et al. (2010; &,
2013).
Paratypes: Specimens in Pl. 1 ![]() ,
figs. 5, 9 (number 23867) and 16 (number 23871), lower Aptian (Bedoulian), Praeorbitolina cormyi
orbitolinid zone
of et al. (2010; &,
2013).
,
figs. 5, 9 (number 23867) and 16 (number 23871), lower Aptian (Bedoulian), Praeorbitolina cormyi
orbitolinid zone
of et al. (2010; &,
2013).
Material studied: About 40 specimens in 15 thin-sections from the Herisht section, deposited at the Babeş-Bolyai University, Cluj-Napoca, Romania, Department of Geology (collection I.I. , under the inventory numbers: 23865 to 23880).
Type locality: The Herisht section (Esfahan area), located about 9 km northeast of Ardakan city, province of Yazd. Coordinates: N 32°25'37.61", E 54°6'54.11''.
Type level: Lower Aptian (Bedoulian), Praeorbitolina cormyi zone ( et al., 2010; &, 2013).
Diagnosis: Representative of Praeorbitolina displaying subdivided subembryonic zone and deuteroconch (two to three, up to 5 short plates).
Description: In the thin-sections that were studied, juvenile specimens
predominate (e.g., Pl. 1 ![]() ,
figs. 12, 14). These consist of the eccentric embryonic apparatus and
some curved chambers (up to 4-5) only, resulting in a biconvex test outline in axial
sections (Pl. 1
,
figs. 12, 14). These consist of the eccentric embryonic apparatus and
some curved chambers (up to 4-5) only, resulting in a biconvex test outline in axial
sections (Pl. 1 ![]() ,
figs. 6, 14). Later chambers successively build up in an annular plane resulting in the slightly
convex to flat test bases (Pl. 1
,
figs. 6, 14). Later chambers successively build up in an annular plane resulting in the slightly
convex to flat test bases (Pl. 1 ![]() ,
figs. 3-5, 7). In adult specimens the chambers may increase
progressively in width (Pl. 1
,
figs. 3-5, 7). In adult specimens the chambers may increase
progressively in width (Pl. 1 ![]() ,
fig. 17). The marginal zone of the chamber layers is subdivided by
one horizontal plate and vertical plates (= one rafter and beams sensu ,
2006). Chamberlets
of the central zone alternate in position between successive chamber layers (Pl. 1
,
fig. 17). The marginal zone of the chamber layers is subdivided by
one horizontal plate and vertical plates (= one rafter and beams sensu ,
2006). Chamberlets
of the central zone alternate in position between successive chamber layers (Pl. 1 ![]() ,
fig. 3).
,
fig. 3).
The megalospheric embryonic apparatus, situated in an eccentric position at the beginning
of the initial spire, consists of three parts: protoconch, deuteroconch and
subembryonic zone. The differentiation of the deuteroconch and the protoconch is
poorly pronounced and they may appear as
a single subspherical chamber. In axial sections parallel to the median plane, the eccentric
position of the embryonic apparatus is distinct and marked at the outer test surface by a small
elevation (Pl. 1 ![]() ,
figs. 5-6, 14). The deuteroconch is subdivided by mostly
two to three, rarely up to five short plates (or septules) (Pl. 1
,
figs. 5-6, 14). The deuteroconch is subdivided by mostly
two to three, rarely up to five short plates (or septules) (Pl. 1 ![]() ,
figs. 3, 9, 16). In the
subembryonic zone, two to three irregularly spaced partitions can be recognized. As the septules
of the subembryonic zone are arranged parallel to the median plane, they are
more easily seen in sections cut perpendicular to this plane (Pl. 1
,
figs. 3, 9, 16). In the
subembryonic zone, two to three irregularly spaced partitions can be recognized. As the septules
of the subembryonic zone are arranged parallel to the median plane, they are
more easily seen in sections cut perpendicular to this plane (Pl. 1 ![]() ,
figs. 3, 13). Axial
sections cut parallel to the median plane often display areas without
partition (Pl. 1
,
figs. 3, 13). Axial
sections cut parallel to the median plane often display areas without
partition (Pl. 1 ![]() ,
figs. 7, 11). In axial sections cut perpendicular to the median plane the
embryonic apparatus may appear centered (Pl. 1
,
figs. 7, 11). In axial sections cut perpendicular to the median plane the
embryonic apparatus may appear centered (Pl. 1 ![]() ,
fig. 8) or inclined to one side (Pl. 1
,
fig. 8) or inclined to one side (Pl. 1 ![]() ,
figs. 4, 15).
,
figs. 4, 15).
Dimensions:
| Measurements: | Minimum | Maximum |
| Height of test: | 0.14 mm | 0.75 mm |
| Diameter of test: | 0.2 mm | 1.2 mm |
| Diameter embryonic apparatus: | 0.09 mm | 0.19 mm |
| Number of chambers: | 4 | 25 |
| Height of chamber: | 0.02 mm | 0.04 mm |
Comparisons: The new species was described as Praeorbitolina sp. by h
(1973)
from the Lower Aptian of the Esfahan region, Central Iran, who noted the
subdivision of the deuteroconch by short plates. Praeorbitolina claveli n.sp. differs
from the type-species Praeorbitolina cormyi ,
1964a, and Praeorbitolina wienandsi
, 1964a, by the complexity of the embryonic apparatus. While the former
shows an undivided subembryonic zone and undivided deuteroconch, the latter exhibits a
partioned subembryonic zone (Pl. 2 ![]() ). Praeorbitolina claveli n.sp. is characterized by the presence
of short plates (2 to 5) in the deuteroconch and plates in the subembryonic
zone. The latter may be scattered and irregularly spaced. Furthermore, the size
of the embryonic apparatus is larger. According to
(1975, p. 121), the average diameter
of the embryonic apparatus is 0.07 mm (0.08 mm in & ,
2009) in P.
cormyi, and 0.1 mm in P. wienandsi (0.13 mm in & ,
2009). In P. claveli, the
embryonic apparatus ranges from 0.09 mm (, 1973: 0.08 mm) to 0.19 mm
(most values between 0.13 and 0.15 mm).
). Praeorbitolina claveli n.sp. is characterized by the presence
of short plates (2 to 5) in the deuteroconch and plates in the subembryonic
zone. The latter may be scattered and irregularly spaced. Furthermore, the size
of the embryonic apparatus is larger. According to
(1975, p. 121), the average diameter
of the embryonic apparatus is 0.07 mm (0.08 mm in & ,
2009) in P.
cormyi, and 0.1 mm in P. wienandsi (0.13 mm in & ,
2009). In P. claveli, the
embryonic apparatus ranges from 0.09 mm (, 1973: 0.08 mm) to 0.19 mm
(most values between 0.13 and 0.15 mm).
In the original description, the deuteroconch of Praeorbitolina wienandsi was defined as completely lacking any plates (see also Fig. B in , 1964a). Later, a "first weak indication" of subdivision of the deuteroconch (= alveolar layer) was mentioned (, 1975: p. 122). et al. (1998: p. 548) defined P. wienandsi as having "a deuteroconch sometimes showing one or two rudimentary septula in its uppermost part". The schematic drawing of the embryonic apparatus again was shown to lack septula in the deuteroconch ( et al., 1998: Fig. 3B). In fact, there are only few a illustrations in the literature for such a feature. For example, et al. (1999: Fig. 5g) illustrated an axial section (perpendicular to the median plane) of P. wienandsi showing a single short plate (or septula) in the deuteroconch. We interpret these specimens as transitional forms between P. wienandsi and P. claveli n.sp. It is worth mentioning that transitional forms between P. cormyi and P. wienandsi also exist ( et al., 1998: Pl. 4, figs. 2, 8). (2005) described a new orbitolinid, Praeorbitolinoides japonica from the Japanese Lower Aptian, assigning it a new genus and species. This taxon has also been interpreted by & (2009) as representing a transitional form between Praeorbitolina cormyi and P. wienandsi, a view later accepted by et al. (2011).
Remarks: Orbitolinids display distinct phylogenetic evolutionary lineages within different species within a single genus or connecting one genus with another. Such lineages are known from both the Dictyoconinae (with simple embryonic apparatus) and the Orbitolininae (with complex embryonic apparatus). In the case of the Dicytoconinae, for instance, such lineages show an increasing complexity of the marginal zone (from forms without plates, to forms with horizontal plates, to forms with both horizontal and vertical plates) (e.g., et al., 2002). Within the Orbitolininae such trends are exhibited by a continuous complexity of the embryonic zone along with an increasing size of the latter (e.g., 1975; et al., 2010).
The so-called Praeorbitolina cormyi Mesorbitolina lineage is the
longest of these trends ranging from the Lower Aptian to the Lower Cenomanian (e.g., et al.,
2010). (1964a) established the Praeorbitolina lineage with P.
cormyi as the ancestor of P. wienandsi, the latter with a more complex
embryonic apparatus that had also shifted to a more central position. This
evolution leads to the most primitive representative of the genus Mesorbitolina
with Mesorbitolina lotzei appearing in the uppermost
Bedoulian. This species is characterized by a centrically positioned embryonic apparatus
showing an undivided deuteroconch (or with rare rudimentary plates) and a well
partitioned subembryonic zone (, 1964b; et al.,
1998; et al.,
2010). Up
to now, P. wienandsi has been considered the direct precursor of Mesorbitolina
lotzei, that does not bear plates in its deuteroconch. Praeorbitolina claveli
only partly fits into the scheme of increasing complexity of the embryonic apparatus, but the position
of the embryonic apparatus (shifting from a clearly eccentric
to a more central position) does not (Fig. 2 ![]() ). This leads to a conclusion that either the details
of the transition from Praeorbitolina to Mesorbitolina must be
reconsidered or that we are dealing with a separate parallel evolution.
). This leads to a conclusion that either the details
of the transition from Praeorbitolina to Mesorbitolina must be
reconsidered or that we are dealing with a separate parallel evolution.

Click on thumbnail to enlarge the image.
Figure 2: Comparison of the embryonic apparatus of representatives of Praeorbitolina , 1964a (further details in the text).
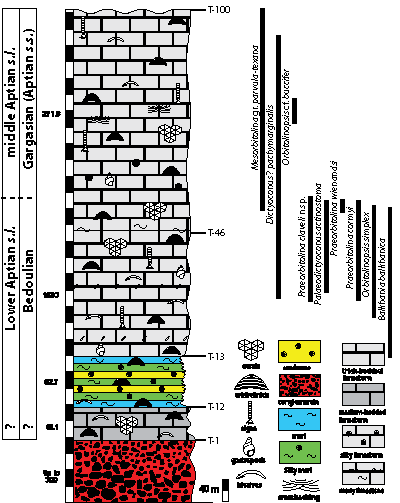
Stratigraphy: The biostratigraphy of the Lower Cretaceous sections in Central Iran is based
on orbitolinid foraminifers (Fig. 3 ![]() ). Some of the taxa observed in different
sections under study are illustrated in plates 1 to 5. They include species that
co-occur with Praeorbitolina claveli n.sp. as well as species from older strata, e.g., Eopalorbitolina
charollaisi (Pl. 4
). Some of the taxa observed in different
sections under study are illustrated in plates 1 to 5. They include species that
co-occur with Praeorbitolina claveli n.sp. as well as species from older strata, e.g., Eopalorbitolina
charollaisi (Pl. 4 ![]() ,
figs. 5, 8), Eopalorbitolina cf. transiens (&) (Pl. 4
,
figs. 5, 8), Eopalorbitolina cf. transiens (&) (Pl. 4 ![]() ,
fig. 12), Eygalierina? turbinata (Pl. 4
,
fig. 12), Eygalierina? turbinata (Pl. 4 ![]() ,
fig. 9)
or Valserina broennimanni (Pl. 5
,
fig. 9)
or Valserina broennimanni (Pl. 5 ![]() ,
figs. 1-6) from the Lower Barremian
of the Anarak section. These taxa and others not illustrated here will be the subject
of a separate contribution.
,
figs. 1-6) from the Lower Barremian
of the Anarak section. These taxa and others not illustrated here will be the subject
of a separate contribution.
Click on thumbnail to enlarge the image.
Figure 3: Lithostratigraphic column of the Herisht section (T numbers) with distribution (vertical range) of orbitolinids and Balkhania balkhanica (for the palaeobiogeographic importance of this taxon, see et al., 2013).
et al. (2010; &,
2013)
developed an orbitolinid biozonation for the Arabian plate shallow-water
carbonates comprising the late Early Barremian Middle Albian interval. For
southeastern France and the Swiss Jura, a detailed biostratigraphic distribution
of 39 orbitolinid taxa (based on ammonites) was recently presented by et al.
(2013)
comprising the Late Hauterivian Bedoulian interval. Both zonations can be taken as a base
for the stratigraphic setting of the sections studied. However, we note that et al.
(2010), ascribed the Furcata
Zone to the Early Gargasian, whereas et al.
(2013) ascribe it
to the late Bedoulian. Praeorbitolina claveli spans the entire Praeorbitolina
cormyi zone sensu et al.
(2010) and stratigraphically
occurs also a little bit higher in the Herisht T section (see Fig. 3 ![]() ). It is
not definitely proven whether this observation indeed represents a longer range than P.
cormyi or just reflects a lack of observation of the latter. To summarize, P. claveli is
recorded from the Praeorbitolina cormyi zone (= Deshayesites deshayesi and D. furcata
ammonite zones, et al., 2010, p. 67) that is late Bedoulian (Early Aptian) early Gargasian (middle Aptian,
or Aptian sensu stricto of et al.,
2011) in age. For a different
concept of the Aptian stage (contrasting et al.,
2011), see
also & (2012).
). It is
not definitely proven whether this observation indeed represents a longer range than P.
cormyi or just reflects a lack of observation of the latter. To summarize, P. claveli is
recorded from the Praeorbitolina cormyi zone (= Deshayesites deshayesi and D. furcata
ammonite zones, et al., 2010, p. 67) that is late Bedoulian (Early Aptian) early Gargasian (middle Aptian,
or Aptian sensu stricto of et al.,
2011) in age. For a different
concept of the Aptian stage (contrasting et al.,
2011), see
also & (2012).
However, some of the orbitolinids from Central Iran, for example Dictyoconus? pachymarginalis
(Pl. 3 ![]() ,
figs. 6-7, 10-11), Praeorbitolina div. sp. (Pls. 1
,
figs. 6-7, 10-11), Praeorbitolina div. sp. (Pls. 1 ![]() - 2
- 2 ![]() ), Dictyoconus? sp. 1 and 2
(Pl. 3
), Dictyoconus? sp. 1 and 2
(Pl. 3 ![]() ,
figs. 5, 8), Orbitolinopsis simplex () (Pl. 5
,
figs. 5, 8), Orbitolinopsis simplex () (Pl. 5 ![]() ,
figs. 7-10), and Mesorbitolina div. sp. (Pl. 4
,
figs. 7-10), and Mesorbitolina div. sp. (Pl. 4 ![]() ,
figs. 7, 10, 13) have
not been recorded from the SW-French-Suisse Jura area (see
et al., 2013).
,
figs. 7, 10, 13) have
not been recorded from the SW-French-Suisse Jura area (see
et al., 2013).
On the contrary, the stratigraphic ranges of the species Palaeodictyoconus
actinostoma &
(Pl. 4 ![]() ,
figs. 3-4, 6) and Orbitolinopsis cf. buccifer & (Pl. 3
,
figs. 3-4, 6) and Orbitolinopsis cf. buccifer & (Pl. 3 ![]() ,
figs. 1-4) as indicated by et al. (2013) can be slightly increased
to include younger strata where orbitolinids have not been found from southern France. This
conclusion stems from the total ranges of both Praeorbitolina cormyi and P. wienandsi extending
to the top of the Furcata Zone ( et al.,
2010, Fig. 10) and the
distribution of the orbitolinids in the Herisht section (T section). However, in the absence
of ammonite data, it is not possible to provide further specifications.
,
figs. 1-4) as indicated by et al. (2013) can be slightly increased
to include younger strata where orbitolinids have not been found from southern France. This
conclusion stems from the total ranges of both Praeorbitolina cormyi and P. wienandsi extending
to the top of the Furcata Zone ( et al.,
2010, Fig. 10) and the
distribution of the orbitolinids in the Herisht section (T section). However, in the absence
of ammonite data, it is not possible to provide further specifications.
The stratigraphy of the still poorly-known species Dictyoconus? pachymarginalis needs further clarification. (1965) described it from the upper Bedoulian-Gargasian strata of the Alborz Mountains, northern Iran. In south-east Spain, it is described as characteristic of the lower Gargasian, i.e., the biozone of "Dictyoconus pachymarginalis, with the initial appearance of Mesorbitolina minuta" ( et al., 1992). Data from the Herisht T section show that this species co-occurs with Mesorbitolina gr. texana () at its upper range limit and is recorded throughout the whole Praeorbitolina cormyi zone.
The above also holds true for the stratigraphic distribution of Orbitolinopsis simplex described by
(1948) as Iraqia simplex
from the "Lower to middle Cretaceous" of Iraq and Iran (for generic
affiliation of the species see , 1963, 1964b). In the Urgonian
of Spain (Teruel region), (1964b) established the Orbitolinopsis simplex
zone between the Mesorbitolina lotzei zone (below) and the Mesorbitolina texana parva
zone (above). According to recent Lower Cretaceous orbitolinid zonation
of the Arabian plate and adjacent regions ( et al.,
2010), this
short term interval can be placed around the Bedoulian-Gargasian boundary. et al.
(1992) place their "Iraqia simplex
zone" below the Praeorbitolina zone. In the Herisht section, O. simplex spans the
whole range of Praeorbitolina cormyi
(Fig. 3 ![]() ).
).
Microfacies and palaeoenvironment: Praeorbitolina claveli n.sp. was observed in wackestones with orbitolinids and other benthic foraminifers such as Maycina bulgarica et al., Sabaudia minuta (), Vercorsella div. sp., Everticyclammina hedbergi () and others. Calcareous algae are represented by Morelletpora turgida (), Terquemella sp., Marinella lugeoni and others. The microfacies and associated microfossils indicate a quiet water inner platform-ramp depositional setting.
Palaeobiogeography: The taxa (or assemblage) reported from the Barremian-Aptian of Central Iran show the "European" character of the orbitolinid fauna of Central Iran ( et al., 1981), namely the "Valserina association" sensu & (1973). For example, Orbitolinopsis simplex is known from non-European areas NE Iraq (type-locality; , 1948), the Zagros Zone of Iran (, 1948), as well as from Spain ( & , 1962; , 1963, 1964b; et al., 1992). Its occurrence in Japan ( et al., 2007) has so far not been verified ( et al., 2011). Other forms like Dictyoconus? pachymarginalis are not palaeobiogeographically diagnostic and were reported from both northern (e.g., et al., 1992) and southern Tethyan carbonate platforms ( & , 2006). The northern Tethyan margin affinity is also further substantiated by the larger benthic foraminifers Balkhania balkhanica ( et al., 2013), Torremiroella hispanica & and the dasycladalean algal assemblages ( et al., 2012; et al., 2013). For the differences between the orbitolinid fauna of Central Iran and that from southern Iran (Zagros zone, eastern part of the Arabian Plate) see the species compilation in et al. (2010).
Helpful remarks by reviewers Esmeralda (Barcelona), Jean (Genève), Bernard (Messery), and Danielle (Geneva) are kindly acknowledged. Simon F. (Kingston) and Antun (St. Lawrence) helped to improve the English. Finally we would also like to thank Bruno (Brest) for his careful editing.
A. (2004).- The geology of Iran.- Geological Survey of Iran, 389 p. [in Farsi].
S.A., S.N. & F. (2010).- Biostratigraphy and evolutionary trend of the Cretaceous orbitolinids in the sedimentary succession of east Lut Block, south west of Qayen.- Sedimentary Facies, Mashhad, vol. 3, n° 1, p. 1-10.
J.P. & M. (1962).- Les Orbitolinidae du Crétacé inférieur de la Sierra du Montsech, Province de Lerida (Espagne).- Revue de Micropaléontologie, Paris, vol. 5, n° 2, p. 104-114.
M. & G.C.P. (1981).- Towards a palaeogeography and tectonic evolution of Iran.- Canadian Journal of Earth Sciences, Montréal, vol. 18, n° 2, p. 210-265.
A., A. & R. (1999).- Stratigraphic evidence for an Early Aptian sea-level fluctuation: the Graua Limestone of south-eastern Ethiopia.- Cretaceous Research, London, vol. 20, n° 6, p. 783-791.
D., S., L., J. & R. (2013).- Paleozoic to Triassic ocean opening and closure preserved in Central Iran: Constraints from the geochemistry of meta-igneous rocks of the Anarak area.- Lithos, Amsterdam, vol. 172, n° 3, p. 267-287.
I.I., K. & B. (2012).- Early Cretaceous calcareous algae from central Iran (Taft Formation, south of Alibad, near Yazd).- Facies, Berlin, vol. 58, n° 4, p. 605-636.
A., M. & R. (1981).- Aperçu paléogéographique sur les Provinces à grands Foraminifères du Crétacé inférieur dans le cadre structural méditerranéen.- Commission internationale pour l'Exploration scientifique de la Mer Méditerranée, Rapport, Marseille, vol. 27, n° 8, p. 115-119.
A. & R. (1973).- Sur la biogéographie de l'association à Valserina du Barremian et la rotation de la Sardaigne.- Comptes Rendus de l'Académie des Sciences, Paris, (Série D), t. 277, p. 829-832.
A. & R. (1999).- Late Barremian orbitolinid Foraminifera from northern Somalia.- Bollettino della Società Paleontologica Italiana, Modena, vol. 38, n° 1, p. 3-13.
A. & R. (2009).- Revision of the orbitolinid foraminiferal genus Praeorbitolinoides 2005 from the Aptian of Hokkaido, Japan.- Micropaleontology, New York, vol. 55, n° 4, p. 421-424.
A. & R. (2013).- The Praeorbitolina/Palorbitolinoides Association: an Aptian biostratigraphic key-interval at the southern margin of the Neo-Tethys. In: P., B. & M. (eds.), Special issue: Spatial patterns of change in Aptian carbonate platforms and related events.- Cretaceous Research, London, vol. 39, p. 70-77.
A., R. & M. (1998).- Early Aptian orbitolinid foraminifera from the Qishn Formation of Al Mukalla (Hadramawt, Southern Yemen). Comparisons with adjacent regions.- Zeitschrift für geologische Wissenschaften, Berlin, vol. 26, n° 5/6, p. 543-561.
B., M., R., J. & B. (2013).- Mapping the rise and demise of Urgonian platforms (Late Hauterivian - Early Aptian) in southeastern France and the Swiss Jura. In: P., B. & M. (eds.), Special issue: Spatial patterns of change in Aptian carbonate platforms and related events.- Cretaceous Research, London, vol. 39, p. 29-46.
A. (1965, unpublished).- A micropaleontological study of the Upper Jurassic and Lower Cretaceous of southern Iran.- Ph.D. Thesis, London, 629 p., 240 Pls.
FRS. (1948).- Larger imperforate Foraminifera of south-western Asia. Families Lituolidae, orbitolinidae and Meandropsinidae.- Monograph British Museum (Natural History), London, 127 p.
S., M.A. & P. (2013).- Iranella inopinata 1965, a puzzling dasycladalean alga from the Lower Cretaceous shallow carbonate shelf deposits of the Zagros fold-thrust belt, SW Iran.- Facies, Berlin, vol. 59, n° 1, p. 231-245.
L. (2006).- Illustrated glossary of terms used in foraminiferal research.- Carnets de Géologie [Notebooks on Geology], Brest, Memoir 2006/02 (CG2006_M02), 126 p. DOI: 10.4267/2042/5832
Y., S.I. & T. (2011).- Orbitolinid foraminifers in the Northwest Pacific: their taxonomy and stratigraphy.- Micropaleontology, New York, vol. 57, n° 2, p. 163-171.
A. & M. (2006).- Cretaceous benthic foraminifers and calcareous algae from Monte Cairo (southern Latium, Italy).- Bolletino della Società Paleontologica Italiana, Modena, vol. 45, n° 1, p. 91-113.
J.P., C. & L. (1992).- Stratigraphy and biozonation of a reference Aptian - Albian p. p. Tethyan carbonate platform succession: The Sierra del Carche series (oriental Prebetic zone Murcia, Spain).- Schriftenreihe der Erdwissenschaftlichen Kommission, Wien, vol. 9, p. 201-221.
K. (2005).- Praeorbitolinoides, a new orbitolinid foraminiferal genus from the Lower Aptian (Cretaceous) of Hokkaido, Japan.- Micropaleontology, New York, vol. 51, n° 1, p. 93-99.
K., M., K. & A. (2007).- Note of orbitolinid foraminifera from the Lower Aptian (Cretaceous) Shimanoshita Mudstoner, Lower Yezo Group, Hokkaido, Japan.- Journal of Saitama University Faculty of Education, Saitama, vol. 56, n° 1, p. 367-372.
M. (1973).- Eine orbitoliniden-Fauna aus der Unterkreide von Esfahan (Zentral Iran).- Neues Jahrbuch für Geologie und Paläontologie Monatshefte, Stuttgart, vol. 6, p. 374-382.
M., B. & G. (2011).- The Aptian Stage: Back to fundamentals.- Episodes, Bangalore, vol. 34, n° 3, p. 148-156.
J.G. & L.A. (2012).- Cretaceous. In: F., J., M. & G. (eds.), The geologic time scale 2012.- Elsevier, Amsterdam, vol. 2, p. 793-854.
M. (1969).- Microfacies and microfossils of the Zagros Area, Southwestern Iran (from Pre-Permian to Miocene).- International Sedimentary Petrographical Series, Leiden, vol. 12, 102 p.
R. (1963).- Grundlagen einer orbitoliniden-Biostratigraphie des tieferen Urgons im pyrenäisch-kantabrischen Grenzgebiet (Nordspanien).- Neues Jahrbuch für Geologie und Paläontologie Monatshefte, Stuttgart, n° 6, p. 320-326.
R. (1964a).- Communication préalable sur l'origine des orbitolines.- Comptes Rendus sommaire de la Société géologique de France, Paris, vol. 10, p. 411-413.
R. (1964b).- Orbitoliniden-Biostratigraphie des Urgons nordöstlich von Teruel (Spanien).- Neues Jahrbuch für Geologie und Paläontologie Monatshefte, Stuttgart, 1964, p. 462-473.
R. (1965).- Dictyoconus pachymarginalis n.sp. aus dem Apt des Elburz-Gebirges (Nord-Iran) (Studien über primitive Orbitolinidae III).- Eclogae Geologiace Helvetiae, Basel, vol. 58, n° 2, p. 976-979.
R. (1975).- General evolutionary trends in Orbitolinas.- Revista Española de Micropaleontologia, Madrid, Num. esp., p. 117-128.
R. & A. (1979).- Upper Barremian - lowermost Aptian orbitolinid foraminifers from the Grand Banks continental rise, northwestern Atlantic (DSDP Leg 43, Site 384).- Initial Reports DSDP, Washington D.C., vol. 43, p. 575-583.
R., F.S.P. van, A., D., B., A. & B. (2010).- Revised orbitolinid biostratigraphic zonation for the Barremian-Aptian of the eastern Arabian plate and implications for regional stratigraphic correlations.- GeoArabia, Manama, Special Publication 4, vol. 1, p. 49-96.
R., B., A., R., J. & D. (2002).- Lignées phylétiques d'orbitolinidés de l'intervalle Hauterivien supérieur-Aptien inférieur : leur importance stratigraphique.- Revue de Paléobiologie, Genève, vol. 21, n° 2, p. 853-863.
M.P.N. & F. (2013).- Lower Cretaceous orbitolinid (Foraminiferida) record from the southwest of Iran (Zagros, Shiraz).- Open Journal of Geology, Irvine, vol. 3, n° 1, p. 1-6.
M.D., J.E. & R.W. (2000).- Orbitolinids from Cretaceous sediments of the Middle East a revision of the F.R.S. and Associates Collection. In: M.B., M.A. & C.W. (eds.), Proceedings of the 5th International Workshop on Agglutinated Foraminifera.- Grzybowski Foundation Special Publication, Krakow, vol. 7, p. 411-437.
M., M.A., A.A. & A.R. (2010).- Barremian-Aptian Dasycladalean algae, new and revisited from the Tirgan Formation in the Kopet Dagh, NE Iran.- Carnets de Géologie [Notebooks on Geology], Brest, Article 2010/05 (CG2010_A05), 13 p. DOI: 10.4267/2042/33368
M., F., S.H., A.A. & A.R. (2013).- Balkhania balkhanica , 1966 (benthic foraminifer) and Kopetdagaria sphaerica , 1960 (dasycladale alga) from the Lower Cretaceous Tirgan Formation of the Kopet Dagh mountain range of NE-Iran and their palaeobiogeographic significance.- Facies, Berlin, vol. 59, n° 1, p. 267-285.
G. (2011).- Late Permian blueschist from Anarak ophiolite (Central Iran, Isfahan province), a mark of multi-suture closure of the Paleo-Tethys ocean.- Revista Mexicana de Ciencas Geológicas, Mexico City, vol. 28, n° 3, p. 544-555.
M., F.T. & M.R. (2013).- The Shah Kuh Formation, a latest Barremian Early Aptian carbonate platform of Central Iran (Khur area, Yazd Block). In: P., B. & M. (eds.), Special issue: Spatial patterns of change in Aptian carbonate platforms and related events.- Cretaceous Research, London, vol. 39, p. 183-194.
M. & F. (2010).- Lower Cretaceous agglutinated larger benthic foraminifera from the Sarvestan section, south of Esfahan, Iran.- The 1st International Applied Geological Congress, Department of Geology, Islamic Azad University-Mashad Branch, Iran, 26-28 April 2010, p. 976-980.

Click on thumbnail to enlarge the image.
Praeorbitolina claveli n.sp.
Herisht section, Bedoulian p.p. Gargasian of Central Iran.
Fig. 1: Axial section. Thin-section T 36.
Fig. 2: Oblique transverse section through the embryonic apparatus. Thin-section T 47.
Fig. 3: Axial section. Thin-section T 77.
Fig. 4: Axial section, perpendicular to the median plane. Holotype specimen. Thin-section T 47
Fig. 5: Axial section. Thin-section T 36.
Fig. 6: Oblique axial section. Thin-section T 36.
Fig. 7: Axial section slightly oblique. Thin-section T 66.
Figs. 8, 15: Axial section, perpendicular to the median plane. Thin-section T 47.
Fig. 9: Axial section. Thin-section T 36.
Figs. 10, 13: Oblique section passing through the embryonic apparatus perpendicular to the median plane. Thin-sections T 47, T 36.
Figs. 11, 14: Axial section of a juvenile specimen. Thin-section T 36, T 47.
Fig. 12: Axial section. Thin-section T 36.
Fig. 16: Oblique axial section. Thin-section M 3.
Fig. 17: Axial section of an adult specimen. Thin-section T 50.
Abbreviations for all plates: AQ = Anarak section, north of
Anarak, T = Herisht
section, M = single sample taken NE of Anarak, G = single sample (see Fig. 1 ![]() ).
).

Click on thumbnail to enlarge the image.
Praeorbitolina cormyi (1-8, 10-11), Praeorbitolina wienandsi (9), Bedoulian of Central Iran
Fig. 1: Axial section of a large adult specimen. Thin-section T 47.
Fig. 2: Axial section. Thin-section T 47.
Fig. 3: Detail from fig. 2 showing embryonic apparatus. Note drop-like subembryonic zone.
Fig. 4: Oblique section. Thin-section T 73.
Fig. 5: Subaxial section, slightly oblique. Thin-section AQ 48B.
Fig. 6: Axial section. Thin-section T 67.
Fig. 7: Axial section, perpendicular to the median plane. Thin-section T 22.
Fig. 8: Axial section of a specimen exhibiting large embryonic apparatus. The specimen compares to Fig. 5c in et al. (1999). Thin-section AQ 49.
Fig. 9: Axial section, perpendicular to median plane. Thin-section T 22.
Fig. 10: Oblique section. Thin-section T 49.
Fig. 11: Axial section, perpendicular to the median plane. Thin-section AQ 48.

Click on thumbnail to enlarge the image.
Orbitolinids from the Anarak (figs. 5, 12) and Herisht sections (all others), Central Iran
Figs. 1-4: Orbitolinopsis cf. buccifer &. Thin-sections T 72, T 86 (fig. 3).
Fig. 5: Dictyoconus? sp. 1. Thin-section AQ 24.
Figs. 6-7, 10-11: Dictyoconus? pachymarginalis . Note the detail of the central zone of a specimen agglutinating nannoconids (see also & , 1979; & , 1999). Thin-sections T 58, T 86, T 74, T 94.
Fig. 8: Dictyoconus? sp. 2. Thin-section T 55.
Figs. 9, 12-13 Orbitolinopsis sp. Thin-section H 25, AQR 1big, H 43n.

Click on thumbnail to enlarge the image.
Orbitolinids from the Anarak (figs. 1-3, 5-7, 9, 10, 12-13) and Herisht sections (figs. 4, 7, 10, 13), Central Iran
Figs. 1-2: Paleodictyoconus cuvillieri (). Thin-sections AQ 7, AQ 18.
Figs. 3-4, 6: Paleodictyoconus actinostoma & . Thin-sections AQ 30, T 67, AQ 1.
Figs. 5, 8: Eopalorbitolina charollaisi . Thin-sections AQ 42, AQn 18.
Fig. 9: Eygalierina? turbinata . Thin-section AQ 49.
Figs. 7, 10, 13: Mesorbitolina gr. texana (). Thin-sections T 48, T 78, T 65.
Fig. 11: Palorbitolina cf. ultima & . Thin-section AQn 42-1.
Fig. 12: Eopalorbitolina cf. transiens (& ). Thin-section AQ 5.
Click on thumbnail to enlarge the image.
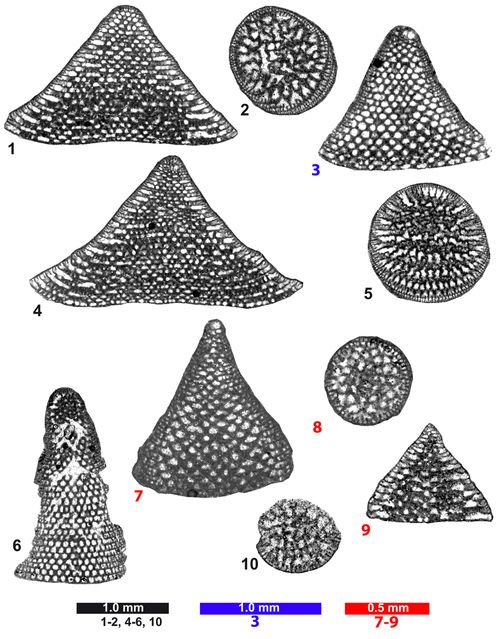
Orbitolinids from the Anarak and Herisht sections (fig. 9), Central Iran
Figs. 1-6: Valserina broennimanni . Thin-sections AQ 8 (figs. 1, 4), AQ 2 (fig. 2), AQ 5 (figs. 3, 5), PA 24 (fig. 6).
Figs. 7-10: Orbitolinopsis simplex (). Thin-sections AQn 18, AQ 20, T 52, AQ 18.