◄ Carnets Geol. 13 (L06) ►
![]()
[Introduction]
[Original description, dimensions ...]
[Age assignment]
[About the subfamilial attribution of Megatyloceras casei]
[About the generic attribution of Megatyloceras casei]
[Conclusions] and ...
[Bibliographic references]
Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad Universitaria, Coyoacán, 04510, México, D.F. (Mexico)
Département des Sciences de la Terre, Université de Nice-Sophia
Antipolis, 28 Avenue Valrose, 06100 Nice (France)
Manuscript online since November 11, 2013
[Scientific editor:
Michel ; technical editor: Bruno ;
language editor: Stephen ]
![]()
In the present work we review the generic attribution of the Mexican ammonoid species Megatyloceras casei , 1949, through a careful examination of the holotype housed in the University of Michigan and with reference to new biostratigraphic data from the type locality. We assign here this species to the subfamily Cheloniceratinae and to the genus Epicheloniceras , 1954.
Ammonites; Cheloniceratinae; Epicheloniceras; Lower Cretaceous; Aptian; Mexico.
J.A. & G. (2013).- About the generic attribution of Megatyloceras casei , 1949 (Ammonoidea, Ancyloceratina), from the Aptian of Mexico.- Carnets de Géologie [Notebooks on Geology], Brest, Letter 2013/06 (CG2013_L06), p. 315-323.
Sur l'attribution générique d'une ammonite de l'Aptien du Mexique: Megatyloceras casei , 1949 (Ammonoidea, Ancyloceratina).- Dans ce travail, nous révisons l'attribution générique de l'espèce mexicaine d'ammonite Megatyloceras casei , 1949. Grâce à un examen minutieux de l'holotype conservé à l'Université du Michigan et grâce aussi aux nouvelles données biostratigraphiques obtenues dans la localité-type, cette révision nous permet d'inclure maintenant cette espèce dans le genre Epicheloniceras , 1954.
Ammonites ; Cheloniceratinae ; Epicheloniceras ; Crétacé inférieur ; Aptien ; Mexique.
In Europe, the genus Megatyloceras ,
1949, is restricted to the Roloboceras hambrovi Subzone ( et al.,
2011) that coincides with the Oceanic Anoxic Event 1a (=OAE 1a) [e.g., et al.,
1998; et al., 2005; et al.,
2010; et al.,
2009, 2010; et al.,
2011]. Currently, this subzone and the OAE 1a are placed in the Deshayesites forbesi Zone (e.g., et al.,
2011; et al.,
2012a; et al.,
2013) or the Deshayesites deshayesi Zone (e.g., et al.,
2005; et al.,
2011). Megatyloceras casei ,
1949, is known from only a single specimen from the La Peña Formation in the Sierra del Rosario, Durango State, Northern Mexico. Based on the most recent research on the ammonoid record of Mexico ( et al.,
2012b, 2013), we propose assigning this species to the interval between the uppermost lower Aptian and the lowermost upper Aptian. The Mexican Megatyloceras is younger than the two possible ages of the Roloboceras hambrovi Subzone proposed in Europe. This biostratigraphic inconsistency is reflected in the stratigraphy. The deposition of the La Peña Formation is related to the Ap 4 sequence ( et al.,
2011, 2012b) and the Roloboceras hambrovi Subzone is associated with the older sequence Ap 3 ( et al.,
2012). In the present work we review the generic attribution of Megatyloceras casei ,
1949, by means of the taxonomic review of its holotype, specimen UMMP 21865
(Fig. 1 ![]() ) housed in the University of Michigan Museum of Paleontology (=UMMP).
) housed in the University of Michigan Museum of Paleontology (=UMMP).
's (1949, p. 149-150) original description reads as follows:
"This species is represented by one large internal cast on which the inner whorls are imperfectly shown.
Form discoidal, depressed, evolute. Whorl section coronatiform, much wider than high, apparently embracing only one-fifth of the preceding whorls. Flanks narrowly convex, venter broadly rounded. Umbilicus deep, rather wide; umbilical wall high, steeply inclined.
Shell ornamented by irregularly alternating primary and secondary ribs. Primaries begin on umbilical wall and form exaggerated lateral bullae from which, on posterior part of outer whorl, two or three rounded ribs may branch. One secondary may rise independently on broad venter between branching primaries. All ribs cross venter transversely without alteration, being somewhat irregularly spaced and subequal in size. On anterior portion of outer whorl, bifurcation from large bullae is rare, and two secondaries may be present between two simple primaries. On inner whorls there is suggestion that the bullae are represented by small, double nodes and that whorl height may be relatively greater. Suture lines not shown.
Holotype U.M 21865."
The dimensions (in mm), given by , are: diameter =176 mm, width of umbilicus = 85, whorl height = 68, whorl thickness = 150.
Remarks: "The new species is named in honor of Professor Emeritus Ermine Cowles of the University of Michigan, Ann Arbor, Michigan."

Click on thumbnail to enlarge the image.
Figure 1: Megatyloceras casei , 1949, lateral (1a), ventral (1b) and frontal (1c) views of the holotype, specimen UMMP 21865. Scale bar is 10 mm.
The ammonoid record of the La Peña Formation contains three zones: the Dufrenoyia justinae Zone of the uppermost lower Aptian, the Gargasiceras ? adkinsi Zone that contains the boundary between the lower and upper Aptian and the Caseyella aguilerae Zone of the lowermost upper Aptian ( et al.,
2013). The oldest zone to which Megatyloceras casei ,
1949, can be assigned, i.e., Dufrenoyia justinae Zone, is younger than the record of Megatyloceras in the Roloboceras hambrovi Subzone of Europe (Fig. 2 ![]() ). The older Aptian taxa, which come from the basal strata of the La Peña Formation, are referable to the genera Dufrenoyia and Burckhardtites, Dufrenoyia justinae Zone.
(1949) and and
(1956) reported some Dufrenoyia and Burckhardtites in the lower part of the La Peña Formation or laterally equivalent units including the Cuchillo Formation of Chihuahua State, the Sierra de los Muertos, Sierra de Parras, Cuesta del Cura, Puerto de las Palomas, Cañón de San Antonio and other localities of Coahuila State; some localities in Nuevo León State such as Cerro de la Silla and Arroyo de San Roque; and the El Mulato Ranch and several other localities in the Nazas River area of Durango State. Other authors have correlated the base of the La Peña Formation with the Dufrenoyia justinae Zone. Recently, et al.
(2011, 2012b) concluded that the base of La Peña Formation is isochronous and assignable to the Dufrenoyia justinae Zone.
(1949) does not specify the position where Megatyloceras casei ,
1949, was found within the La Peña Formation. Thus, M. casei could come from anywhere within the interval, uppermost lower Aptian to lowermost upper Aptian, that contains the ammonoid record of the La Peña Formation (Fig. 2
). The older Aptian taxa, which come from the basal strata of the La Peña Formation, are referable to the genera Dufrenoyia and Burckhardtites, Dufrenoyia justinae Zone.
(1949) and and
(1956) reported some Dufrenoyia and Burckhardtites in the lower part of the La Peña Formation or laterally equivalent units including the Cuchillo Formation of Chihuahua State, the Sierra de los Muertos, Sierra de Parras, Cuesta del Cura, Puerto de las Palomas, Cañón de San Antonio and other localities of Coahuila State; some localities in Nuevo León State such as Cerro de la Silla and Arroyo de San Roque; and the El Mulato Ranch and several other localities in the Nazas River area of Durango State. Other authors have correlated the base of the La Peña Formation with the Dufrenoyia justinae Zone. Recently, et al.
(2011, 2012b) concluded that the base of La Peña Formation is isochronous and assignable to the Dufrenoyia justinae Zone.
(1949) does not specify the position where Megatyloceras casei ,
1949, was found within the La Peña Formation. Thus, M. casei could come from anywhere within the interval, uppermost lower Aptian to lowermost upper Aptian, that contains the ammonoid record of the La Peña Formation (Fig. 2 ![]() ). However, in the section of the La Peña Formation studied by et al.
(2013), three large fragments of a poorly preserved Cheloniceratinae were collected in beds 136 and 138 of the Caseyella aguilerae Zone (lowermost upper Aptian). In this particular section it seems that this is the only part of the La Peña Formation that provides large Cheloniceratinae. The dimensions and preservation of these large fragments of Cheloniceratinae resemble Megatyloceras casei. Accordingly, it is likely that Megatyloceras casei was collected from a similar position within the La Peña Formation and its age is probably earliest late Aptian. In order to test this probable age we reviewed all of the Cheloniceratinae of a similar size to Megatyloceras casei housed in the University of Michigan, Museum of Paleontology (UMMP), that come from the same or nearby areas to where Megatyloceras casei was collected. In this collection we recognize specimens of Cheloniceras
(Fig. 3a-b
). However, in the section of the La Peña Formation studied by et al.
(2013), three large fragments of a poorly preserved Cheloniceratinae were collected in beds 136 and 138 of the Caseyella aguilerae Zone (lowermost upper Aptian). In this particular section it seems that this is the only part of the La Peña Formation that provides large Cheloniceratinae. The dimensions and preservation of these large fragments of Cheloniceratinae resemble Megatyloceras casei. Accordingly, it is likely that Megatyloceras casei was collected from a similar position within the La Peña Formation and its age is probably earliest late Aptian. In order to test this probable age we reviewed all of the Cheloniceratinae of a similar size to Megatyloceras casei housed in the University of Michigan, Museum of Paleontology (UMMP), that come from the same or nearby areas to where Megatyloceras casei was collected. In this collection we recognize specimens of Cheloniceras
(Fig. 3a-b ![]() ;
Fig. 4a-d
;
Fig. 4a-d ![]() ), all of which must be assigned to the lower Aptian, as this genus is restricted to this interval. One of these specimens
(Fig. 4c
), all of which must be assigned to the lower Aptian, as this genus is restricted to this interval. One of these specimens
(Fig. 4c ![]() ) with a high density of ribbing resembles Cheloniceras meyendorffi
(d', 1845). In Europe this species is assigned to the uppermost lower Aptian (e.g., ,
1961a; et al.,
2008; et al.,
2012a). We also found one specimen that belongs to the genus Epicheloniceras
(Fig. 3c-d
) with a high density of ribbing resembles Cheloniceras meyendorffi
(d', 1845). In Europe this species is assigned to the uppermost lower Aptian (e.g., ,
1961a; et al.,
2008; et al.,
2012a). We also found one specimen that belongs to the genus Epicheloniceras
(Fig. 3c-d ![]() ). Given the particular ammonoid record of the PFZ section, the presence of Cheloniceras (uppermost lower Aptian) in the UMMP collection precludes a lowermost upper Aptian position for Megatyloceras casei. Thus, the range of M. casei is uppermost lower Aptian to lowermost upper Aptian
(Fig. 2
). Given the particular ammonoid record of the PFZ section, the presence of Cheloniceras (uppermost lower Aptian) in the UMMP collection precludes a lowermost upper Aptian position for Megatyloceras casei. Thus, the range of M. casei is uppermost lower Aptian to lowermost upper Aptian
(Fig. 2 ![]() ).
).
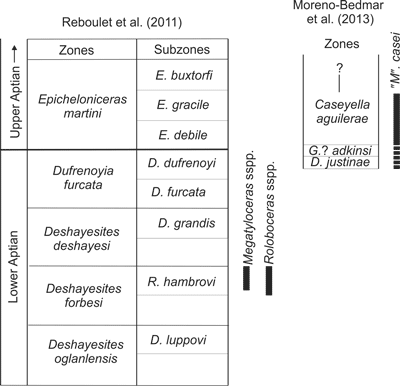
Click on thumbnail to enlarge the image.
Figure 2: Tethyan lower Aptian and lowermost upper Aptian standard ammonite zonation of et al. (2011) with the ranges of the species of Megatyloceras and Roloboceras, and Mexican uppermost lower Aptian and lowermost upper Aptian ammonite zonation of et al. (2013) with the range of Megatyloceras casei.

Click on thumbnail to enlarge the image.
Figure 3: a-b) Cheloniceras sp. of 210 mm of maximum diameter lateral (3a) and ventral (3b) views of the specimen UMMP 16414. Black triangle indicates the end of the phragmocone. El Mulato Ranch, Durango State; c-d) Epicheloniceras sp., ventral (3c) and lateral (3d) views of the specimen UMMP 23215. White and black triangles indicate the equivalent ventral tubercle, characteristic of this genus. North of Rio Nazas Valley, Durango State. Scale bar is 10 mm.

Click on thumbnail to enlarge the image.
Figure 4: a) Cheloniceras sp. of 175 mm of maximum diameter, lateral view of specimen UMMP 23458. South of Sierra del Rosario, Durango State; b) Cheloniceras sp. of 320 mm of maximum diameter, lateral view of the specimen UMMP 23479. Black triangle indicates the end of the phragmocone. Rio Nazas Valley, Durango State; c) Cheloniceras cf. meyendorffi of 160 mm of maximum diameter, lateral view of specimen UMMP 23485. West side of Sierra del Rosario, Durango State; d) Cheloniceras sp. of 225 mm of maximum diameter, lateral view of specimen UMMP 23480. North of Rio Nazas Valley, Durango State. Scale bar is 10 mm.
The genus Megatyloceras , 1949, possesses "but one row of prominent lateral bullae in the adult stages" (, 1949, p. 149). The generic assignment of Megatyloceras casei is placed in doubt by 's (1949, p. 150) observation, "On inner whorls there is a suggestion that the bullae are represented by small, double nodes". (1961b) comments that at 176 mm diameter this taxon is similar to the genus Roloboceras , 1954, (1961b) also remarks on the "double nodes" mentioned by , which are problematic as Roloboceras has one tubercle, similar to a bulge, in the peri-umbilical position. (1961b) concludes that the Mexican taxon is more similar to the subfamily Cheloniceratinae , 1923, than the subfamily Roloboceratinae , 1961b, which includes the genera Megatyloceras , 1949, and Roloboceras , 1954. Later, and (1970) make a similar argument in coming to the same conclusion as .
Megatyloceras casei , 1949, cannot be assigned to the genus Megatyloceras because of some clear morphological differences. At the same diameter, the large mid-lateral tubercles that coronate the whorl section appear to be absent and the ribs do not bifurcate or trifurcate regularly as is common in species of Megatyloceras. In the Mexican taxon, ribs are clearly visible in the umbilical wall whereas in Megatyloceras and Roloboceras, ribs are more discreet or absent in this position. Megatyloceras casei , 1949, cannot be included in the genus Roloboceras , 1954, because the majority of Roloboceras species of a similar size to the Mexican specimen possess the bulges characteristic of the genus. The only exception is Roloboceras saxbyi , 1961b, in which bulges disappear very early during ontogeny, but this species differs from the Mexican taxon because the ribs are stronger and have a more regular costulation pattern. In addition, the whorl section of Megatyloceras casei , 1949, is of maximum width at the middle of the flank while in all species of Roloboceras the maximum width is located at the lower flank position.
The inner whorls of the Mexican species are very badly preserved (Fig. 5 ![]() ).
).

Click on thumbnail to enlarge the image.
Figure 5: Lateral view of the holotype of Megatyloceras casei , 1949, UMMP 21865 with two enlargements. The two white triangles show two very rudimentary tubercles. Scale bar is 10 mm.
However, in the last whorl it seems that two very rudimentary tubercles occur in one rib, as indicated by the two white triangles in
Fig. 5 ![]() . This observation accords with the "double nodes" of
(1949). The presence of two such tubercles is a characteristic of Cheloniceratinae ,
1923. The presence of two tubercles therefore enables us to eliminate the possibility that this ammonoid belongs to the genus Megatyloceras ,
1949, or the genus Roloboceras ,
1954, from the Roloboceratinae, in accordance with the opinions of
(1961b) and and
(1970). Meanwhile, the fact that the whorl section of Megatyloceras casei ,
1949, has the maximum width at midflank is also characteristic of the genera Cheloniceras ,
1903, and Epicheloniceras ,
1954.
. This observation accords with the "double nodes" of
(1949). The presence of two such tubercles is a characteristic of Cheloniceratinae ,
1923. The presence of two tubercles therefore enables us to eliminate the possibility that this ammonoid belongs to the genus Megatyloceras ,
1949, or the genus Roloboceras ,
1954, from the Roloboceratinae, in accordance with the opinions of
(1961b) and and
(1970). Meanwhile, the fact that the whorl section of Megatyloceras casei ,
1949, has the maximum width at midflank is also characteristic of the genera Cheloniceras ,
1903, and Epicheloniceras ,
1954.
The Mexican specimen is characterized by the irregularity of its ribbing pattern, especially of the secondary ribs. This important feature was also noted by (1949): "Shell ornamented by irregularly alternating primary and secondary ribs". Species of the genus Cheloniceras , 1903, have a more regular rib pattern. Species of the genus Epicheloniceras have a less regular rib pattern, especially in the secondary ribs. In juvenile specimens of the genus Epicheloniceras we can clearly see the characteristics of the genus Epicheloniceras: siphonal depression in the primary ribs and ventrolateral tubercles. In the current specimen, which is a subadult or adult, inner whorls are not visible and these characteristics cannot be observed. Therefore we compare our specimen with other subadult or adult specimens that belong to the genus Epicheloniceras , 1954. (2005) uses an interesting conception of the macroconch and microconch for the genus Epicheloniceras. According to (2005) Epicheloniceras microconchs are small forms with low costulation density, very regular rib pattern, less pronounced tuberculation and little difference between the primary and secondary ribs. In contrast, Epicheloniceras macroconchs are larger specimens with higher costulation density, more irregular rib pattern especially in secondary ribs, well developed tuberculation during the first ontogenetic stages and a pronounced difference between the primary and secondary ribs. According to 's conception, our specimen would be a subadult-adult macroconch. (1906, Pl. 3, figs. 1-3) figured very well preserved specimens of macroconchs of Epicheloniceras tschernyschewi (, 1906), the type species of the genus. In his Pl. 3, figs. 2 (diameter,D=100 mm) and 3, the high costulation density and the irregularity of the secondary ribs are clearly evident. Further, they show how, during the ontogeny, primary ribs become less tuberculated and more similar to the secondary ribs. On 's larger specimen (op. cit., Pl. 3, fig. 1) the tuberculation virtually disappears; in its inner whorls it is difficult to see the initial tuberculated stages. (1915, Pl. 3, fig. 2) shows a larger specimen (D=174 mm) of the same species with identical features, but in this case the inner whorls are more distinct and it is possible to see the initial tuberculated stages. On the bigger specimen (D=215 mm) (, 1915, Pls. 4-5), the tuberculation seems to disappear completely. This absence of clear tuberculation, and the rib pattern of the large Mexican specimen (D=176 mm), resemble features of Epicheloniceras specimens of similar size figured by (1915, Pls. 4-5). More recent authors show examples of large specimens of the genus Epicheloniceras with the same, previously noted features (e.g., , 1962, text-fig. 85c; et al., 2008, Pls. 17, 19-20; Pl. 21, fig. 1; Pl. 22 & Pl. 24, fig. 3).
In addition, the Mexican specimen seems to have a big tubercle placed on a robust primary rib
(Fig. 5 ![]() ). Robust primary ribs with a well developed tubercle can be seen on some specimens of
(2005) e.g., Pl. 21, fig. 1b and Pl. 22, 2a
& 2c. The comparison of the Mexican taxon with other Epicheloniceras specimens illustrated in the literature shows clear similarities.
). Robust primary ribs with a well developed tubercle can be seen on some specimens of
(2005) e.g., Pl. 21, fig. 1b and Pl. 22, 2a
& 2c. The comparison of the Mexican taxon with other Epicheloniceras specimens illustrated in the literature shows clear similarities.
Additionally, if we compare the species studied here with other Mexican Cheloniceratinae of a comparable size that come from the same or nearby areas to where Megatyloceras casei was collected, it is clear that the specimens assigned to the genus Cheloniceras
(Fig. 3a-b ![]() ;
Fig. 4a-d
;
Fig. 4a-d ![]() ) are very different in their regular rib pattern. The only specimen with an irregular rib pattern is assigned to the genus Epicheloniceras
(Fig. 1c-d
) are very different in their regular rib pattern. The only specimen with an irregular rib pattern is assigned to the genus Epicheloniceras
(Fig. 1c-d ![]() ).
).
Despite poor preservation of the inner whorls, the morphological and ornamental features of Megatyloceras casei lead us to conclude that this taxon should be assigned to the subfamily Cheloniceratinae. Further, we consider it appropriate to place M. casei within the genus Epicheloniceras as Epicheloniceras casei (, 1949). E. casei (, 1949) is assigned to the uppermost lower Aptian.
We appreciate the assistance and access to facilities to review the Mexican Aptian ammonoids housed in the University of Michigan, Museum of Paleontology, provided by Dr. Dan , Coordinator of the Invertebrate Collection of the Museum of Paleontology, and Dr. William , Preparator of the Vertebrate Fossil Preparation Laboratory, both of the University of Michigan, Ann Arbor, Michigan, USA. We are very grateful for the helpful corrections and suggestions made by Dr. Ottilia and Dr. Yves . We are very grateful to the Language Editor, Stephen , for his corrections which allow significant improvements to the manuscript.
T., J.A., R., P.W., K. & E. (2010).- Sedimentary evolution of an Aptian syn-rift carbonate system (Maestrat Basin, E Spain): effects of accommodation and environmental change.- Geologica Acta, Barcelona, vol. 8, n° 3, p. 249-280.
R. (1954).- New genera and subgenera of Lower Cretaceous ammonites.- Journal of the Washington Academy of Sciences, vol. 44, n° 4, p. 106-115.
R. (1961a).- The stratigraphical palaeontology of the Lower Greensand.- Palaeontology, London, vol. 3, p. 487-621.
R. (1961b).- A monograph of the Ammonoidea of the Lower Greensand, part III.- Monograph of the Paleontographical Society, London, vol. 115, p. 119-216.
R. (1962).- A monograph of the Ammonoidea of the Lower Greensand, part IV.- Monograph of the Paleontographical Society, London, vol. 116, p. 217-288.
Y. (2005, unpublished).- Biostratigraphie, évolution et renouvellement des ammonites de l'Aptien supérieur (Gargasien) du bassin vocontien (Sud-Est de la France).- Doctoral thesis, Université Claude Bernard Lyon I, 1-302 p.
T., F.J-M.R. & J.A. (2013).- Stable carbon-isotope stratigraphy and ammonite biochronology at Madotz, Navarra, northern Spain: implications for the timing and duration of oxygen depletion during OAE-1a.- Cretaceous Research, London, vol. 40, p. 143-157.
F.M., J.G., M. & G. (eds., 2012).- A geologic time scale.- Elsevier, Amsterdam, 1176 p.
W.E. (1949).- Geology of Sierra de Los Muertos area, Mexico (with descriptions of Aptian cephalopods from the La Peña Formation).- Geological Society of America, Bulletin, Tulsa, vol. 60, p. 89-176.
W.E. & T. (1956, unpublished).- Jurassic and Lower Cretaceous stratigraphy and tectonics of northeast Mexico.- Petróleos Mexicanos, internal report, Mexico, 390 p.
A. (1903).- Pseudoceratites of the Cretaceous.- Monographs of the United States Geological Survey, Washington, vol. 54, 351 p.
J.A., M., T., R., G., R. & A. (2009).- Biostratigraphic characterization by means of ammonoids of the lower Aptian Oceanic Anoxic Event (OAE 1a) in the eastern Iberian Chain (Maestrat Basin, eastern Spain).- Cretaceous Research, London, vol. 30, p. 864-872.
J.A., M., T., R., G., F.J.-M.R., A. & R. (2010).- Lower Aptian ammonite biostratigraphy in the Maestrat Basin (Eastern Iberian Chain, Eastern Spain). A Tethyan transgressive record enhanced by synrift subsidence.- Geologica Acta, Barcelona, vol. 8, n° 3, p. 281-299.
J.A., T., R. & R. (2011).- La transgresión tetisiana del Aptiense inferior terminal: comparación entre su registro en México y España y relación con el ciclo global de tercer orden Ap4.- Paleontologia i Evolució Memòria especial, Sabadell, vol. 5, p. 259-262.
J.A., M., J., J.M., T., R., G., F.J.-M.R. & R. (2012a).- Lower Aptian ammonite and carbon isotope stratigraphy in the eastern Prebetic Domain (Betic Cordillera, southeastern Spain).- Geologica Acta, Barcelona, vol. 10, n° 4, p. 333-350.
J.A., T., R. & R. (2012b).- Uppermost Lower Aptian transgressive records in Mexico and Spain: chronostratigraphic implications for the Tethyan sequences.- Terra Nova, Oxford, vol. 24, p. 333-338.
J.A., R., M. & L.G. (2013).- Aptian (Lower Cretaceous) ammonite biostratigraphy of the Francisco Zarco Dam stratigraphic section (Durango State, northeast Mexico).- Journal of South American Earth Sciences, Amsterdam, vol. 42, p. 150-158.
M., W., J.A., J.-P. & G. (1998).- Correlation of biostratigraphic and stable isotope events in the Aptian historical stratotype of La Bédoule (southeast France).- Comptes-Rendus de l'Académie des Sciences, Paris, (Série II - Sciences de la Terre et des Planètes), vol. 327, p. 693-698.
M., B. & G. (2011).- The Aptian Stage: Back to fundamentals.- Episodes, Bangalore, vol. 34, n° 3, p. 148-156.
M., I., J.A., G.A., E., M. & G. (2011).- High-resolution chemo- and biostratigraphic records of the Early Aptian oceanic anoxic event in Cantabria (N Spain): Palaeoceanographic and palaeoclimatic implications.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 299, p. 137-158.
J. (1915).- Représentants du genre Douvilleiceras de l'Aptien du versant septentrional du Caucase.- Mémoire du Comité géologique, Paris, (Nouvelle Série), Livre 121, 53 p. [in Russian]
A. d' (1845).- In: R.I., E. de & A., Géologie de la Russie d'Europe et des montagnes de l'Oural.- London and Paris, vol. 2, p. 419-498.
S. & E. (1970).- Sur la présence de l'Aptien inférieur dans le secteur du sud-ouest des Monts Perşani Megatyloceras persaniense nov. sp.- Analele universitatii Bucuresti, Geologie, Bucarest, vol. 19, p. 1-41.
S., P.F., J.A., M.B., R., Y., M., C., V., A., B., V., H., Z., E.J., D., S., T.N., L.G., J.-L., I.A., P. & O. (2011).- Report on the 4th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the " Group" (Dijon, France, 30th August 2010).- Cretaceous Research, London, vol. 32, p. 786-793.
M., M. de, L., M., J.-P., W., J.A. & G. (2005).- Early Aptian δ13C and manganese anomalies from the historical Cassis-La Bédoule stratotype sections (S.E. France): relationship with a methane hydrate dissociation event and stratigraphic implications.- Carnets de Géologie [Notebooks on Geology], Brest, Article CG2005/04 (CG2005_A04), 18 p., DOI: 10.4267/2042/3229
P., G., M., G. & R. (2008).- The Douvilleiceratidae (Ammonoidea) of the Lower Aptian historical stratotype area at Cassis-La Bédoule (SE France).- Carnets de Géologie [Notebooks on Geology], Brest, Memoir 2008/03 (CG2008_M03), 60 p., DOI: 10.4267/2042/18125
J. (1906).- Die Beschreibung einiger Douvilleiceras-Arten aus dem oberen Neocom Russlands.- Verhandlungen der Russisch-Kaiserlichen Mineralogischen Gesellschaft zu St. Petersburg, (series 2), vol. 44, n° 1, p. 157-198.
L.F. (1923).- A monograph of the Ammonoidea of the Gault, part 1.- Monograph of the Paleontographical Society, London, (1921), vol. 75, 72 p.