◄ Carnets Geol. 14 (12) ►
![]()
Contents
[1. Introduction] [2. Study area] [3. Material and methods]
[4. Paleontology]
[5. Distribution of ostracodes and paleoecology] [6. Paleogeography]
[7. Conclusions] and ... [Bibliographic references]
itt Fossil - Instituto Tecnológico de Micropaleontologia,
UNISINOS, Av. Unisinos, 950, Săo Leopoldo, RS (Brazil)
;
Universidade de Lisboa, Faculdade de Cięncias, Departamento de Geologia e Centro de Geologia, Campo Grande, C6, 4º, 1749-016 Lisboa (Portugal)
Universidade de Lisboa, Faculdade de Cięncias, Centro de Geologia, Campo Grande, C6, 4º, 1749-016 Lisboa (Portugal); deceased ( et al., 2013)
itt Fossil - Instituto Tecnológico de Micropaleontologia, UNISINOS, Av. Unisinos, 950, Săo Leopoldo, RS (Brazil)
itt Fossil - Instituto Tecnológico de Micropaleontologia, UNISINOS, Av. Unisinos, 950, Săo Leopoldo, RS (Brazil)
Published online in final form (pdf) on July 3, 2014
[Editor: Bruno ; language editor:
Robert W. ]
![]()
This paper describes thirteen new species and two new genera of marine and brackish water ostracodes from the Turonian deposits of Potiguar Basin, NE Brazil, among a total of 53 taxa. The new species include Cophinia grekoffi, Fossocytheridea tiberti, Haughtonileberis dinglei, Hemicytherura viviersae, Jandairella obesa (new genus and species), Loxocorniculum? narendrai, Ovocytheridea posteroprojecta, O. reymenti, Perissocytheridea caudata, P. mossoroensis, Potiguarella grosdidieri (new genus and species), P. coimbrai and Procytherura ballentae. The diversity and dominance indexes vary according to the paleoenvironment. Three assemblages were identified: a predominantly mixohaline fauna in the basal part of the section, followed by a diversified shallow marine fauna and, in the upper part, a marine and brackish water ostracode fauna. Most of the recorded species are endemic. Eight species are common to Northwest and North Africa, indicating a faunal link during the Turonian. The study of the Turonian faunas of the Potiguar Basin represents a significant improvement to understand the dynamic evolution of the Brazilian basins and the paleobiogeographical relationship with other regions.
Ostracodes; Turonian; Potiguar Basin; Jandaíra Formation; paleoecology; paleobiogeography.
E.K., M.C., J.-P., G. & C. (2014).- Ostracodes from the Upper Cretaceous deposits of the Potiguar Basin, northeastern Brazil: taxonomy, paleoecology and paleobiogeography. Part 1: Turonian.- Carnets de Géologie [Notebooks on Geology], Brest, vol. 14, nº 12, p. 211-252.
Les ostracodes des sédiments du Crétacé supérieur du Bassin de Potiguar, NE Brésil : taxonomie, paléoécologie et paléobiogéographie. Premičre partie : Le Turonien.- Sur un total de 53 taxons d'ostracodes d'eaux marines ou saumâtres recensés dans les sédiments turoniens du Bassin de Potiguar Basin, NE Brésil, treize espčces nouvelles et deux genres nouveaux sont décrits dans cette publication. Ces nouvelles espčces sont Cophinia grekoffi, Fossocytheridea tiberti, Haughtonileberis dinglei, Hemicytherura viviersae, Jandairella obesa (note : il s'agit d'un nouveau genre fondé sur une nouvelle espčce), Loxocorniculum? narendrai, Ovocytheridea posteroprojecta, O. reymenti, Perissocytheridea caudata, P. mossoroensis, Potiguarella grosdidieri (note : il s'agit d'un nouveau genre fondé sur une nouvelle espčce), P. coimbrai et Procytherura ballentae. Les indices de diversité et de prédominance varient en fonction du paléoenvironnement. Trois associations ont été distinguées : une faune d'ostracodes essentiellement mixohaline (d'environnement saumâtre) dans la partie basal de la série, suivie par une faune diversifiée de milieu marin peu profond et, dans la partie supérieure, par une faune mixte d'eaux marines ou saumâtres. La plupart des espčces observées sont endémiques. Huit espčces sont cependant également connues en Afrique du Nord et du NW, suggérant un lien faunistique au cours du Turonien. L'étude des faunes turoniennes du bassin de Potiguar représente une avancée significative dans l'état des connaissances concernant notamment l'évolution dynamique des bassins brésiliens et leur lien paléobiogéographique avec d'autres régions.
Ostracodes ; Turonien ; Bassin de Potiguar ; Formation Jandaíra ; paléoécologie ; paléobiogéographie.
Few studies on Upper Cretaceous ostracodes have been published hitherto from the northeastern Brazilian margin. The first taxonomic study of Cretaceous marine ostracodes from Brazilian basins was carried out by (1964, 1966a, 1966b, 1967) in the Albian-Turonian interval of the Sergipe Basin. Later, (1973a, 1979) studied marine ostracodes from the Upper Cretaceous-Paleogene of the same basin, identifying 49 species, seven of which were new and mostly of them from the Paleogene. Northwards, in the Pernambuco Basin, et al. (2005), identified 30 species belonging to 13 genera and described a new genus and eight new species in the Cretaceous-Paleogene interval. As regards the study of the ostracodes from the marine Upper Cretaceous of the Potiguar Basin, the work of (1995) represents the initial stage of research. Later, et al. (2000) identified a fauna composed of Cytherella austinensis , Cytherella gambiensis , Leguminocythereis reymenti and the new species Bairdia potiguarensis, Soudanella laciniosa paucicostata and Brachycythere ventrocomplanatus. Concomitantly, et al. (2000) illustrated a well diversified fauna from the same basin, focusing their work on biostratigraphic and paleobiogeographic issues.
The sedimentary basins of the East Brazilian and West African margins have significant and almost continuous carbonate sections in the Cretaceous that, despite the different denominations, essentially represent sedimentation associated with the establishment of the Atlantic Ocean. These successions initially deposited in shallow carbonate platforms, evolved into deep waters, due to the expansion of the proto-Atlantic Ocean ( et al., 1993). Due to the proximity of the American and African margins in the Turonian, the ostracode assemblages have high similarities as already demonstrated by (1982) and et al. (2000). More studies of the West African Cretaceous basins have been published than of the Brazilian basins, including several regions in West Africa (, 1961), Morocco (, 1991, 1992, 1995, 1996, 2002; & , 1996; & , 2008), Nigeria (, 1960, 1963; , 1973b; , 1987, 1992; , 1999a, 1999b), Algeria (, 1985; , 1989), Senegal (, 1963), Cameroon (, 1951), Gabon (, 1979) and Tunisia ( et al., 1981, 1995).
The present study contributes to the knowledge of the taxonomy of the Turonian ostracode assemblages from a long and fairly complete borehole of the Jandaíra Formation, Potiguar Basin and gives additional data on the paleoecology and paleobiogeography, based on these microfossils.
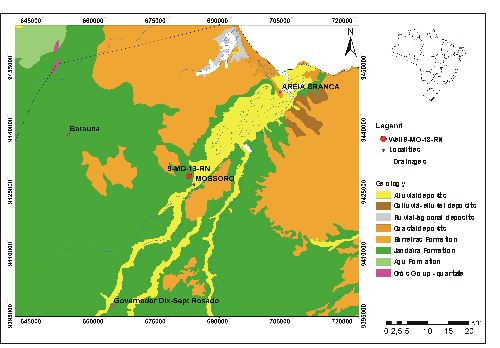
The Potiguar Basin (Fig. 1 ![]() ) is located at the intersection of the Equatorial Margin with the East Continental Margin, covering an area of approximately 48,000 km˛. Geologically, it is limited on the South, East and West by the crystalline basement, extending northwards to the 2,000 m
isobath. This basin is distributed mostly in the state of Rio Grande do Norte and partially in the state of Ceará (,
2003;
et al., 2007).
) is located at the intersection of the Equatorial Margin with the East Continental Margin, covering an area of approximately 48,000 km˛. Geologically, it is limited on the South, East and West by the crystalline basement, extending northwards to the 2,000 m
isobath. This basin is distributed mostly in the state of Rio Grande do Norte and partially in the state of Ceará (,
2003;
et al., 2007).
The Potiguar Basin was formed by extensional processes during the Early Cretaceous associated with rifting that culminated in the separation of the South American and African plates. et al. (2007) divided the stratigraphic record into three supersequences: rift, deposited during the Early Cretaceous (Berriasian-Lower Aptian), represented by fluvial-deltaic and lacustrine deposits of the Pendęncia and Pescada formations; post-rift, deposited during the Late Aptian-Early Albian and characterized by fluvial-deltaic deposits, with the first records of marine ingression (Alagamar Formation); drift, comprising the entire marine sedimentation that occurred after the Early Albian until the Recent.
The Jandaíra Formation was deposited during the first major marine transgression from the north (, 1976). This unit consists predominantly of bioclastic calcarenites and calcilutites deposited in marginal marine environments on a shallow platform ( & , 1988; & , 1994). & (1994) reviewed internal reports of Petrobras and attributed the Turonian-Campanian age to this formation, which was later corroborated by et al. (2007). The base of Turonian is marked by the last occurrence of the palynomorph Gnetaceaepollenites similis and the top is marked by the last occurrence of Tricolpites S427 ( et al., 2000). The ostracode biozone OP-4 (from et al., 2000) is characterized by the presence of Jandairella obesa gen. nov. sp. nov. (=Trachyleberididae sp. P5 of et al., 2000) and Indet. gen 1 POT 1 (= Cypridopsis? sp. P1 et al., 2000).
Click on thumbnail to enlarge the image.
Figure 1: Potiguar Basin, showing the position of the studied well, 9-MO-13-RN, located in Mossoró city, Rio Grande do Norte State, Brazil (from Geological Survey of Brazil).
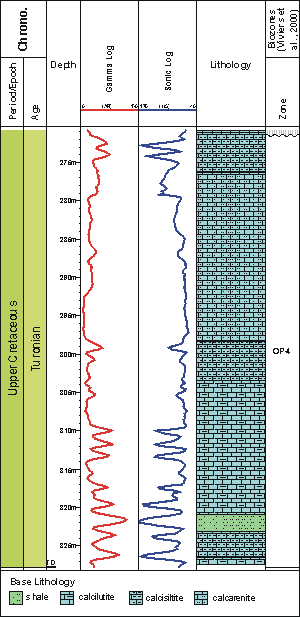
Twenty-eight samples were analyzed, covering 57 m of the Turonian interval in the 9-MO-13-RN well
(Fig. 2 ![]() ). The average weight of the samples was 60 g and were dominantly calcarenites. The samples were processed using standard laboratory techniques for the study of fossil
ostracodes, which consists in disaggregation with hydrogen peroxide (H2O2), washing through sieves with mesh 250, 180 and 63 micrometers and dried at 60° C. More than 4,800 specimens were picked for this study. The selected specimens were imaged using a Zeiss EVO MA15 scanning electron microscope. Some of the material is only illustrated
(Appendix 1) and includes the taxa that remained undetermined due to the small number of the specimens, their poor preservation, limited stratigraphic value, or because it was impossible to include them in any known
taxon.
). The average weight of the samples was 60 g and were dominantly calcarenites. The samples were processed using standard laboratory techniques for the study of fossil
ostracodes, which consists in disaggregation with hydrogen peroxide (H2O2), washing through sieves with mesh 250, 180 and 63 micrometers and dried at 60° C. More than 4,800 specimens were picked for this study. The selected specimens were imaged using a Zeiss EVO MA15 scanning electron microscope. Some of the material is only illustrated
(Appendix 1) and includes the taxa that remained undetermined due to the small number of the specimens, their poor preservation, limited stratigraphic value, or because it was impossible to include them in any known
taxon.
The study area map was created using ArcGIS® software version 9.3 by ESRI (Environmental Systems Research Institute). The complete micropaleontological data were plotted in colour using the StrataBugs® software, with depth in Y-axis and the identified taxa in X-axis in the well. The statistical data were run with the PAST software ( et al., 2001; & , 2006) and Microsoft Excel®.
Click on thumbnail to enlarge the image.
Figure 2: Lithological profile and electrical logs of the studied interval of the well 9-MO-13-RN.
Taxonomy follows the classification of (2005). In the systematic descriptions, the following abbreviations/conventions are employed: L: length, H: height, W: width; very small (<0.400 mm), small (0.400-0.500 mm), medium (0.510-0.700 mm), large (0.710-0.900 mm), very large (>0.900 mm); C: carapace, RV: right valve, LV: left valve, DV: dorsal view, VV: ventral view, EV: external view (valve), IV: internal view; f: female, m: male. All dimensions are in mm. Type and figured specimens are deposited in the collections of Museu de História Geológica do Rio Grande do Sul, Universidade do Vale do Rio dos Sinos, under the prefix ULVG followed by their respective catalogue numbers. All dimensions of the figured specimens are in the plate captions.
Class Ostracoda , 1802
Subclass Podocopa , 1866
Order Platycopida , 1866
Suborder Platycopina , 1866
Superfamily Cytherelloidea , 1866
Family Cytherellidae , 1866
Genus Cytherella , 1849
Cytherella mediatlasica , 1996
(Pl. 1 ![]() ,
figs. A-D)
,
figs. A-D)
1987 Cytherella sp. , p. 25, Pl. 13, figs. 5-6.
1992 Cytherella sp. , p. 328, Pl. 2, fig. 20.
1996 Cytherella mediatlasica , p. 484-485, 488, Pl. 1, figs. 1-10.
2000 Cytherella sp. P6 , , & , p. 415, Figs. 8, 14-15.
Material: 268 specimens.
Age: Turonian.
Stratigraphic and geographic distribution: Turonian-Santonian of Nigeria (, 1987, 1992), Santonian of Morocco (, 1996), Turonian-Campanian of Potiguar Basin, Brazil ( et al., 2000; this work).
Remarks: The presence of different morphotypes in this species was observed by (1996) in the Santonian of Morocco. The specimens recorded in this work are smaller than those recorded in Morocco.
Order Podocopida , 1866
Suborder Bairdiocopina , 1887
Superfamily Bairdioidea , 1887
Family Bairdiidae , 1887
Genus Bairdoppilata , & , 1935
Bairdoppilata potiguarensis (, & , 2000)
(Pl. 1 ![]() ,
figs. E-H)
,
figs. E-H)
2000 Bairdia potiguarensis , & , p. 333-334, Figs. 8.5-8.7.
Material: 267 specimens.
Age: Turonian.
Stratigraphic and geographic distribution: Upper Cretaceous, Potiguar Basin, Brazil ( et al., 2000; this work).
Remarks: This species was reassigned to the genus Bairdoppilatta due to the observation of the auxiliary dentition of the hinge at anterior and posterior ends, which corresponds to Bairdoppilata's diagnosis. The specimens illustrated by et al. (2000) are smaller (average = 0.850 mm) and were considered juveniles. Bairdia sbaensis , 1996, described from the Upper Turonian (?)-Coniacian (?)-Santonian of Morocco is more elongate and the posterior margin is straighter. (1999a, 1999b) recorded Bairdia sp. in the Turonian of Nigeria, which is similar to B. potiguarensis, but more elongate and with the dorsal margin straighter. Bairdia sp. C , 1964, from the Turonian of Egypt, differs by the more rounded dorsal margin and by the pronounced overlap. B. potiguarensis differs from B. cespedesensis (, 1946) from the Maastrichtian of Cuba and the Pará-Maranhăo Basin ( et al., 2009) by the marked difference in the size and by the overlapping of the dorsal margin.
Family Bythocyprididae , 1969
Genus Bythocypris , 1880
"Bythocypris" POT 1
(Pl. 1 ![]() ,
figs. I-K)
,
figs. I-K)
Material: 141 specimens.
Brief description: carapace large, sub-rectangular to suboval in lateral view, narrow in dorsal view, with the greatest height at mid-length. LV overlaps the right one along all margins, except in the dorsal one. Dorsal margin convex, ventral margin almost straight. Anterior and posterior margins rounded. Surface predominantly smooth with punctuations scarcely and irregularly distributed. Sexual dimorphism not observed.
Age: Turonian.
Remarks: Although the Cretaceous species with this shape are usually assigned to the genus Bythocypris, we are not sure about the generic status of this species. Bythocypris , 1880 is more reniform and the posterior margin is much more asymmetrical than in the species recorded here. In addition, the internal features of the studied material are not available. This species differs from Bythocypris gohrbandti , 1968, described from the Upper Cretaceous of Tunisia and recorded in the Santonian of Algeria by (1985), by its symmetry of the anterior and posterior margins.
Suborder Cypridocopina , 1850
Superfamily Cypridoidea , 1845
Family Candonidae , 1900
Subfamily Paracypridinae , 1923
Genus Paracypris , 1866
Paracypris aff. caudata (, 1964)
(Pl. 1 ![]() ,
figs. L-O)
,
figs. L-O)
aff. 1964 Ovocytheridea caudata , p. 119, Pl. 14, figs. 4a-b.
1973a Paracypris caudata (van den ) - , p. 125-126, Pl. 7.3, fig. 4.
1985 Paracypris sp. 2 , p.150-151, Pl. 3, fig. 10.
Material: 52 specimens.
Brief description: Carapace large, robust, sub-triangular in lateral view and suboval in dorsal view. Overlap of the LV along all margins of the RV, more pronounced in the dorsal margin. Maximum height and maximum width at mid-length. Dorsal margin strongly convex, with a small concavity in the anterodorsal part of the RV, ventral margin almost straight in the LV and slightly concave at mid-length of the RV. Anterior margin obliquely rounded; posterior acuminate. Surface smooth.
Age: Turonian.
Stratigraphic and geographic distribution: probably in the Turonian of Egypt (, 1964), Turonian of Gabon (, 1973a), Middle Turonian-Santonian of Algeria (, 1985) and Turonian of Brazil (this work).
Remarks: This species is higher in the posterior third, when compared with P. caudata (, 1964). In general, P. aff. caudata differs from other species of Paracypris from Potiguar Basin in being higher and wider and in having a more sub-triangular outline.
Paracypris aff. dubertreti & , 1972
(Pl. 1 ![]() ,
figs. P-Q)
,
figs. P-Q)
aff. 1972 Paracypris dubertreti & , p. 276, Pl. 1, fig. 1.
1985 Paracypris dubertreti & - , p.149, Pl. 3, figs. 6-7.
Material: 71 specimens.
Brief description: Carapace of medium size, thin, sub-triangular and elongate in lateral view; narrow and lanceolate in dorsal view. LV slightly larger than the right one along all margins. Greatest height positioned at the anterior third. Dorsal margin almost straight; ventral margin with a pronounced concavity at mid-length. Anterior margin rounded, posterior very acuminate. Surface smooth.
Age: Turonian.
Stratigraphic and geographic distribution: Cenomanian-Turonian of Algeria (, 1985) and Turonian of Potiguar Basin, Brazil (this work).
Remarks: This species is similar to Paracypris dubertreti & , 1972, recorded in the Upper Cenomanian of Lebanon and a common species in the Cenomanian of Africa (e.g., Israel: & , 1974; Algeria: , 1989; Morocco: , 1991; et al., 2013; Tunisia: et al., 1995; Jordan: & , 2010). However, the Brazilian species is significantly smaller than P. dubertreti and has a straighter dorsal margin, the ventral margin is more concave and the cardinal angles less marked.
Paracypris POT 1
(Pl. 1 ![]() ,
figs. R-T)
,
figs. R-T)
Material: 45 specimens.
Brief description: Carapace sub-triangular and elongate in lateral view, lanceolate in dorsal view. LV overlaps the right one along all margins, more pronounced in the anterodorsal margin, where the RV has a small concavity. Maximum height and greatest width just in front of the mid-length. Dorsal margin convex; ventral margin straight. Anterior margin obliquely rounded, posterior margin subacute, with the extremity positioned ventrally. Surface smooth or with faint longitudinal stries near the ventral margin.
Age: Turonian.
Remarks: This species differs from Paracypris sp. P3 , , & , 2000, from the Coniacian-Santonian of Potiguar Basin, Brazil, by its more acute posterior margin and the location of the maximum length, which is more ventral.
Suborder Cytherocopina , 1850
Superfamily Cytheroidea , 1850
Family Cytherideidae , 1925
Genus Cophinia , 1961
Cophinia grekoffi , & sp. nov.
(Pl. 2 ![]() ,
figs. A-E)
,
figs. A-E)
Derivatio nominis: This species is named after N. , in honor of his important contribution to the knowledge of ostracodes.
Material: 140 specimens.
Holotype: C, f, ULVG-9941
(Pl. 2 ![]() , fig. A), sample 297.60 m.
, fig. A), sample 297.60 m.
Paratypes: ULVG-9942, ULVG-9943, ULVG-9944, ULVG-9946.
Dimensions: Holotype: L: 0.900, H: 0.570, W: 0.401. Paratypes: L: 0.817-0.939, H: 0.470-0.521, W: 0.409-0.427.
Type-locality: 9-MO-13-RN, coordinates UTM: 682595E / 9428410N (zone 24S), 297.60 m, Potiguar Basin, Brazil.
Diagnosis: A species of Cophinia characterized by an ovoid outline in lateral and dorsal views, with the anterior and posterior regions strongly depressed.
Description: carapace very large, ovoid and elongate in lateral view, ovoid and inflate in dorsal view. LV larger than the right one, uniformly overlapping it along all margins, except in the anteroventral margin. Maximum height and maximum width at mid-length. Dorsal margin convex on both valves. Ventral margin convex in the LV, almost straight and with a concavity in the anterior half in the RV. Anterior margin obliquely rounded, posterior margin with a projection in both valves below mid-height. Anterior and posterior regions strongly compressed, more easily seen in dorsal view. Merodont hinge. Surface smooth.
Sexual dimorphism: pronounced, with the females higher, less elongated and less inflated than males. The maximum width in the females is located at mid-length, in the males it is located at the posterior third and the LV is more inflated.
Age: Turonian.
Remarks: This species is more elongated and has the anterior and posterior regions more compressed than Cophinia POT 2
(Pl. 9 ![]() ,
figs. N-O). Moreover, in dorsal view, Cophinia POT 2, exhibits more symmetric valves than C. grekoffi sp. nov. "Cophinia" GA B I ,
1979, from the Cenomanian-Turonian of Gabon has the posterior projection more pronounced and the overlap more developed than in C. grekoffi sp. nov.
,
figs. N-O). Moreover, in dorsal view, Cophinia POT 2, exhibits more symmetric valves than C. grekoffi sp. nov. "Cophinia" GA B I ,
1979, from the Cenomanian-Turonian of Gabon has the posterior projection more pronounced and the overlap more developed than in C. grekoffi sp. nov.
Cophinia POT 1
(Pl. 2 ![]() ,
figs. F-H)
,
figs. F-H)
Material: 8 specimens.
Brief description: carapace large, sub-triangular to sub-rectangular in lateral view, ovoid with almost parallel margins, in dorsal view. Strong overlap of the right valve along all margins. Dorsal margin convex in the LV, almost straight in the RV, with well defined anterior cardinal angle. Ventral margin convex in the LV and straight in the RV. Anterior margin asymmetrically rounded, posterior obliquely rounded. In the posterior region, the left valve has a projection, located below the mid-height of the carapace. Surface irregularly punctuated. Sexual dimorphism not observed, probably due to the scarcity of the material.
Age: Turonian.
Remarks: This species has similarities with Cophinia pulvinata , 1963 (Senonian of Gambia), Cophinia apiformis (, 1960) from the Coniacian-lower Santonian of Nigeria and Ovocytheridea ashakaensis , 1987, from the Turonian of Nigeria. However, in dorsal view, this species differs markedly from the others by its parallel margins. The species was not described due to the scarcity of the material and the lack of internal views.
Genus Fossocytheridea & , 1964
Fossocytheridea tiberti , & nov. sp.
(Pl. 2 ![]() ,
figs. I-M)
,
figs. I-M)
Derivatio nominis: In honor of Dr. Neil for his contribution to the knowledge of the genus Fossocytheridea.
Material: 52 specimens.
Holotype: C, f, ULVG-9938 (Pl. 2 ![]() ,
figs. I-J), sample 319.30 m.
,
figs. I-J), sample 319.30 m.
Paratypes: ULVG-10661, ULVG-9939, ULVG-9940.
Dimensions: Holotype: L: 0.880, H: 0.519, W: 0.431. Paratypes: L: 0.775-0.856, H: 0.424-0.500, W: 0.400.
Type-locality: 9-MO-13-RN, coordinates UTM: 682595E / 9428410N (zone 24S), 319.30 m, Potiguar Basin, Brazil.
Diagnosis: A species of the genus Fossocytheridea characterized by a large and sub-rectangular to sub-ovoid carapace in lateral view, evenly swollen in dorsal view; surface smooth.
Description: Carapace large, sub-rectangular to sub-ovoid in lateral view, ovoid in dorsal view. Left valve larger than right one, along all margins, more pronounced in the ventral margin. Greatest height near the mid-length in females, and in males at the anterior third. Dorsal margin convex, ventral margin nearly straight in the RV, slightly convex in LV. Anterior margin rounded, posterior margin rounded to slightly acuminate. Surface smooth. Anterior region slightly compressed. Cardinal angles well developed. The hinge of the RV is the typical of the genus.
Age: Turonian.
Remarks: This species resembles Clithrocytheridea senegali , 1961, from the Senonian of Senegal, in general outline and dimensions of the carapace, but differs in the position of the maximum length which is higher. Moreover, the African species is smaller and has a DV with almost parallel margins in the female.
Genus Ovocytheridea , 1951
Ovocytheridea reymenti , & sp. nov.
(Pl. 2 ![]() ,
figs. N-T)
,
figs. N-T)
1987 Ovocytheridea symmetrica , 1960 - , p. 61-62, Pl. 3, fig. 11; Pl. 4, figs. 1-3, 12-13, 20.
1992 Ovocytheridea symmetrica - , p. 334, Pl. 2, fig. 7.
2000 Ovocytheridea aff. symmetrica - , , & ,, p. 423, Figs. 13, 17-20.
Derivatio nominis: In honor of Dr. Richard for his contribution to the knowledge of the Cretaceous ostracodes from Africa.
Material: 448 specimens.
Holotype: C, f, ULVG-9959 Pl. 2 ![]() ,
figs. N-O), sample 272.40 m.
,
figs. N-O), sample 272.40 m.
Paratypes: ULVG-9960 - ULVG-9963.
Dimensions: Holotype: L: 0.631, H: 0.386, W: 0.297. Paratypes: L: 0.647-0.657, H: 0.369-0.400, W: 0.280-0.292.
Type-locality: 9-MO-13-RN, coordinates UTM: 682595E / 9428410N (zone 24S), 272.40 m, Potiguar Basin, Brazil.
Diagnosis: A species of Ovocytheridea with a medium-size carapace, suboval to sub-rectangular in lateral view, narrow in dorsal view and strongly punctuated.
Description: carapace of medium size, suboval to sub-rectangular in lateral view, ovoid and narrow in dorsal view. Slightly wider at mid-length. Left valve overlapping the right one strongly along all margins, less pronounced in the posteroventral margin. Anterior region very slightly compressed. Anterior margin asymmetrically rounded, posterior margin obliquely rounded, with marked posterior cardinal angle. Dorsal margin convex. Ventral margin straight in the RV and convex in the LV. Greatest height just in front of the mid-length. Surface densely punctuated, except near the margins, where it is smooth. Adductor muscle represented by four scars vertically aligned: the upper and the lower ones are rounded and the median ones more elongated. Frontal muscle scar rounded and mandibular scar more elongated. Antimerodont hinge: in the LV the anterior socket is larger than the posterior one.
Sexual dimorphism: males are proportionally narrower, lower and more elongated than females.
Age: Turonian.
Stratigraphic and geographic distribution: Cenomanian-Maastrichtian of Nigeria (, 1987, 1992) and Turonian, Potiguar Basin, Brazil ( et al., 2000).
Remarks: This species is similar to Ovocytheridea symmetrica , 1960, recorded in the Coniacian of Nigeria. However, the Brazilian species is shorter, less inflated and more inequivalve than O. symmetrica. In the description of (1960), the punctuations in the valves of O. symmetrica are treated as "false ornamentation". In the Potiguar species, the surface is strongly covered with punctuations.
Ovocytheridea posteroprojecta , & sp. nov.
(Pl. 3 ![]() , figs. A-E)
, figs. A-E)
Derivatio nominis: Related to the diagnostic feature in the posteroventral margin of this species.
Material: 40 specimens.
Holotype: C, f, ULVG-9976
(Pl. 3 ![]() , figs. A-C), sample 277.25 m.
, figs. A-C), sample 277.25 m.
Paratypes: ULVG-10486, ULVG-9977.
Dimensions: Holotype: L: 1.040, H: 0.600, W: 0.460. Paratypes: L: 0.760-1.100, H: 0.438-0.580, W: 0.410.
Type-locality: 9-MO-13-RN, coordinates UTM: 682595E / 9428410N (zone 24S), 277.25 m, Potiguar Basin, Brazil.
Diagnosis: An elongate species of the genus Ovocytheridea with a posteroventral projection on the right valve.
Description: carapace very large and elongate, sub-triangular in lateral view, ovoid in dorsal view. Strongly inequivalve, with the LV overlapping the right one along all margins, except in the posteroventral margin where the RV is larger. Anterior region slightly compressed, posterior region projecting downward in the RV. This projection is more pronounced in juvenile specimens. Dorsal margin convex. Ventral margin almost straight in males and slightly convex in females. Anterior margin rounded, posterior margin acuminate and projected downward. Surface smooth. Middle region with a shallow sulcus, more visible in DV in the female carapaces, which are more inflated in the posterior region. Hinge in LV with a crenulated and elongated anterior socket extending anteriorly from dorsal apex, the middle element is a crenulated bar along the posterodorsal slope, and another long crenulated socket is observed in posterodorsal angle.
Sexual dimorphism: females with a more ovoid outline, higher and shorter than males.
Age: Turonian.
Remarks: In the posterior region, this species is similar to Ovocytheridea rostrata , 1963, from the Senonian of Gambia. However, O. rostrata is smaller, higher and has a very pronounced anterodorsal sulcus. Ovocytheridea sp. B 780 , 1973, recorded by (1985) in the Coniacian of Algeria is similar in lateral view, though the left valve overlaps the right one in the posterior region and differs clearly in dorsal view.
Genus Perissocytheridea 1938
Perissocytheridea caudata , & sp. nov.
(Pl. 3 ![]() , figs.
F-K)
, figs.
F-K)
Derivatio nominis: Related to the diagnostic feature, the caudal process in the posterior end of this species.
Material: 515 specimens.
Holotype: C, f,
ULVG-9932
(Pl. 3 ![]() , figs. F-H), sample 272.40 m.
, figs. F-H), sample 272.40 m.
Paratypes: ULVG-9933, ULVG-9934.
Dimensions: Holotype: L: 0.546, H: 0.322, W: 0.330. Paratypes: L: 0.550-0.606, H: 0.310-342, W: 0.303.
Type-locality: 9-MO-13-RN, coordinates UTM: 682595E / 9428410N (zone 24S), 272.40 m, Potiguar Basin, Brazil.
Diagnosis: A species of Perissocytheridea characterized by a sub-pyriform carapace in lateral view, with a short and narrow caudal process; surface of the valves covered with small punctuations.
Description: carapace of medium size, sub-pyriform in lateral view, suboval in dorsal view. LV larger with overlap more pronounced in the anterior margin. Greatest height and width near the mid-length. Anterior and posterior regions compressed. Dorsal margin straight, ventral margin convex, partially covered by the ventrolateral expansion. Anterior margin broadly rounded, posterior slightly triangular, forming a short and narrow caudal process, situated near the upper third of the height and projected upwards. Anterodorsal sulcus moderately deep. The males present a posteroventral depression. Surface completely punctuated and with weak ribs in the ventral and ventrolateral regions. Antimerodont hinge.
Sexual dimorphism: pronounced, with males more elongated and narrower than females. The ventrolateral expansion and the posteroventral depresssion are more prominent in the males.
Age: Turonian.
Remarks: (1991, 1992) recorded a similar species in the Cenomanian of Morocco, which was identified as Pterygocythere? sp. 3. The African species is significantly higher and narrower, with a deeper anterodorsal sulcus and the caudal process and ventrolateral expansion more developed than the Brazilian species.
Perissocytheridea mossoroensis , & sp. nov.
(Pl. 3 ![]() , figs.
L-P)
, figs.
L-P)
Derivatio nominis: From Mossoró, city where the well 9-MO-13-RN was drilled.
Material: 20 specimens.
Holotype: C, f, ULVG-9930
(Pl. 3 ![]() , figs. L-N), sample 319.80 m.
, figs. L-N), sample 319.80 m.
Paratypes: ULVG-9931, ULVG-10555 (not illustrated).
Dimensions: Holotype: L: 0.450, H: 0.259, W: 0.233. Paratypes: L: 0.480-0.539, H: 0.270-0.272, W: 0.247-0.250.
Type-locality: 9-MO-13-RN, coordinates UTM: 682595E / 9428410N (zone 24S), 319.80 m, Potiguar Basin, Brazil.
Diagnosis: A species of Perissocytheridea with a small to medium carapace, with dorsal margin straight to slightly convex, posterior margin sub-triangular and projected downwards. Surface entirely reticulated.
Description: Carapace small (females) to medium (males), tumid, sub-pyriform in lateral view, sub-rhomboid (females) or ovoid (males) in dorsal view. Maximum height in the anterior cardinal angle. LV larger than RV. Dorsal margin slightly convex to almost straight, ventral margin slightly convex, partially covered by the ventrolateral expansion. Anterior margin obliquely rounded; posterior margin sub-triangular with extremity projected downward and located below the mid-height. The ornamentation consists of strong reticulation covering the entire surface of the valves, with small and dense punctuations in the solum of the reticulum; concentric ribs parallel to the periphery of the valves, running from the anteroventral region to dorsomedian region can also be seen. Anterodorsal sulcus deep. Internal features not seen.
Sexual dimorphism: pronounced, males larger, more inflated in the posterior region, with a more developed ventrolateral expansion and marked anteromedian sulcus than females.
Age: Turonian.
Remarks: This species resembles Perissocytheridea cavelieri ( & , 1995), recorded in the Campanian-Maastrichtian of Mali ( et al., 1996). Both species have a sub-triangular posterior margin, although in P. mossoroensis sp. nov it is much more pronounced. Also the dorsal margin of P. mossoroensis sp. nov. is more convex and the ventrolateral expansion is much more developed than P. cavelieri. Perissocytheridea cretacea (, & , 2010), recorded in the Santonian-Campanian of Brazilian continental margin, differs by the posterior margin, which is more rounded and projected upwards, and it is wider than the species recorded herein. Metacytheropteron? sp. , 1973, from the Maastrichtian of Argentina, which is probably a species of Perissocytheridea, is more elongated, with the ventrolateral projection less pronounced and more inflated and with the posterior margin projected upwards.
Genus Rostrocytheridea , 1969
Rostrocytheridea? POT 1
(Pl. 3 ![]() , figs.
Q-S)
, figs.
Q-S)
Material: 10 specimens.
Brief description: carapace of medium size, suboval to sub-triangular in lateral view, ovate in dorsal view. Maximum height near the mid-length in females, at anterior third in males; maximum width at mid-length. LV larger than the right one, with a strong overlap in the dorsal region. Dorsal margin convex, ventral margin almost straight to slightly convex. Anterior margin rounded, posterior margin obliquely rounded, with the extremity under mid-height and slightly projected downwards. Surface with large and irregularly distributed pits, which decrease in size towards the margins. Anterior margin with stout and short spines, posteroventral region with four robust spines. Very strong sexual dimorphism, with males lower and more elongate than females.
Age: Turonian.
Remarks: The general outline of this species has similarities to Rostrocytheridea , 1969, but its generic attribution is doubtful because the diagnostic internal features are poorly preserved and the ventral margin is straight, while in all the species of the genus it is clearly convex. et al. (2009) recorded a possible species of Rostrocytheridea in the Brazilian equatorial margin, in the Maastrichtian, but it is larger, not punctuated and has no spines in the anterior and posteroventral margins.
Family Cytheruridae , 1894
Genus Eucytherura , 1894
Eucytherura aff. speluncosus (, 1996)
(Pl. 4 ![]() , figs. A-E)
, figs. A-E)
aff. 1996 Schizocythere speluncosus , p. 494-495, Pl. 5, figs. 10-18.
Material: 102 specimens.
Brief description: carapace very small, sub-rectangular in lateral view, posteromedian region inflated in dorsal view. LV slightly larger than the RV. Greatest height at anterior cardinal angle. Dorsal margin straight, ventral margin convex. Anterior margin obliquely rounded, posterior sub-rounded and projected upwards with the extremity situated in the upper third of the height. Anterior and posterior regions compressed. The entire surface is reticulated and with the presence of three tubercles in posteroventral, posterodorsal and anterodorsal regions. Anteromedian sulcus very deep. Eye tubercle present, followed by post-ocular sulcus. Sexual dimorphism pronounced, with females less elongated and wider.
Age: Turonian.
Remarks: The species recorded in the Potiguar Basin is very similar to the males of Eucytherura speluncosus (, 1996), from the Santonian of Morocco, but some features of the females from Potiguar Basin are different: they are more inflated, the posterior region is more compressed, more projected upwards and with the ribs more robust. The species was reassigned to the genus Eucytherura because Schizocythere , 1950, has the caudal process at mid-height and has no sulcus or tubercles.
Genus Hemicytherura , 1941
Hemicytherura viviersae , & sp. nov.
(Pl. 4 ![]() , figs.
F-I)
, figs.
F-I)
Derivatio nominis: This species is named after Marta Claudia , in honor of her important contribution to the study of Brazilian marine ostracodes.
Material: 53 specimens.
Holotype: C,
ULVG-9902
(Pl. 4 ![]() , figs. F-H), sample 275.80 m.
, figs. F-H), sample 275.80 m.
Paratypes: ULVG-10669, ULVG-9901 (not figured).
Dimensions: Holotype: L: 0.390, H: 0.251, W: 0.165. Paratypes: L: 0.340-0.380, H: 0.200-0.210, W: 0.149-0.150.
Type-locality: 9-MO-13-RN, coordinates UTM: 682595E / 9428410N (zone 24S), 275.80 m, Potiguar Basin, Brazil.
Diagnosis: A species of the genus Hemicytherura, very small, sub-rhomboidal to suboval in lateral view, with a regular reticulation and a triangular caudal process at mid-height, straightly pointing backwards.
Description: Carapace very small, sub-rhomboidal to suboval in lateral view, suboval elongated in dorsal view. The LV overlaps slightly the right one along all margins, except in the dorsal margin, where the RV overlaps the LV strongly. Greatest height at mid-length. Dorsal margin strongly convex, ventral margin convex, especially in the posterior third. Anterior margin rounded, posterior acuminate. Anterior region slightly compressed, posteroventral region compressed. Caudal process at mid-height, weakly reticulated. Presence of a small ventrolateral expansion. Surface ornamented by asymmetrically rounded reticula, regularly arranged. Dorsal region densely punctuated near the margin. Ribs in the ventral region, with intercostal region punctuated. Internal features not observed.
Sexual dimorphism: evident, with males lower and more elongated than females.
Age: Turonian.
Remarks: Few records of the genus Hemicytherura in the "mid" Cretaceous are known, possibly due to its difficult identification, which is strongly dependent on internal features. (1996) described, in the Santonian of Boulmane region, Middle Atlas, Morocco, the species Hemicytherura sexangula. The outline of both species is similar, but the African species is much more robust, with a different reticulation, ventral margin straight to slightly convex and a caudal process projected downwards.
Genus Pelecocythere , 1979
Pelecocythere POT 1
(Pl. 4 ![]() , figs.
J-L)
, figs.
J-L)
Material: 6 specimens.
Brief description: carapace of medium size, sub-trapezoidal in lateral view. Dorsal margin straight, ventral margin flattened and not visible due to a large ventrolateral carina. Anterior and posterior margins very similar, asymmetric and projected downwards. Maximum height and maximum width at mid-length. Anterodorsal region compressed. Surface with fine ribs, more visible in the posterior and posterodorsal regions.
Age: Turonian.
Remarks: (2009) suggested that, during the Cretaceous, Pelecocythere lived in shallow water and during the Cenozoic it migrated into deeper or colder environments, characterizing a retrothermal adaptation. et al. (2010) proposed the reassignment of some Cretaceous species into Pelecocythere, in addition to describing a new species from the Brazilian margin (Pelecocythere dinglei) found in the Santonian-Campanian interval of the Santos Basin.
Genus Procytherura , 1970
Procytherura ballentae , & sp. nov.
(Pl. 4 ![]() , figs.
M-P)
, figs.
M-P)
Derivatio nominis: In honor of the late Dr. Sara for her great legacy in the study of ostracodes.
Material: 52 specimens.
Holotype: C, f, ULVG-9913
(Pl. 4 ![]() , figs. M-N), sample 275.80 m.
, figs. M-N), sample 275.80 m.
Paratypes: ULVG-9914, ULVG-10675.
Dimensions: Holotype: L: 0.389, H: 0.196, W: 0.123. Paratypes: L: 0.394-0.430, H: 0.166-0.190, W: 0.120-0.140.
Type-locality: 9-MO-13-RN, coordinates UTM: 682595E / 9428410N (zone 24S), 275.80 m, Potiguar Basin, Brazil.
Diagnosis: A species of the genus Procytherura with a very small, elongate and fragile carapace, very narrow in dorsal view and with faintly developed ribs on the surface of the valves.
Description: Carapace very small and fragile, sub-ovoid elongate to sub-rectangular in lateral view, acuminate and posteroventraly compressed, very narrow in dorsal view. "Caudal process" at mid-height. RV overlapping the LV dorsally. Greatest height at anterior cardinal angle. Maximum width at mid-length. Dorsal and ventral margins almost straight, both with a slight concavity at mid-length. Anterior margin rounded, posterior margin acuminate. Presence of an eye swelling and a very weak dorso-median sulcus. The ornamentation consists of weak ribs, extending along the entire length of the valves. Internal features not observed.
Sexual dimorphism: females higher and less elongated than males.
Age: Turonian.
Remarks: This is the first record of this genus in the Brazilian Cretaceous. (1991) recorded a similar species in the Vraconian-Cenomanian of Morocco, Procytherura? sp. 1. The species recorded by differs by the smooth surface and the caudal process projected downwards. Procytherura? sp. 2 recorded in the Cenomanian of Algeria (, 1985) is smooth and has the caudal process in the upper third of the height, differing from the species recorded herein.
Genus Semicytherura , 1957
Semicytherura aff. adversainflata , 1996
(Pl. 4 ![]() , figs.
Q-U)
, figs.
Q-U)
aff. 1996 Semicytherura adversainflata , p. 493-494, Pl. 4, figs. 18-26.
Material: 8 specimens.
Description: carapace very small, sub-rectangular and elongate in lateral view, inflated in dorsal view. Valves nearly equal, with the LV slightly larger than the right one. Greatest width in the posterior third. Dorsal margin straight, parallel to the ventral margin which is almost straight with a slight median concavity just behind the mid-length. Anterior margin rounded, posterior sub-triangular, with a caudal process in the upper third of the height. The ventrolateral region has a small alar expansion. Ventral region flattened and striate, particularly in the anterior part. Surface with large and weak reticula. Intramural region with a dense secondary reticulation. Sexual dimorphism not observed.
Age: Turonian.
Remarks: Semicytherura adversainflata , 1996, is very similar to this species, but has a more regular reticulated ornamentation and the caudal process projected slightly downward.
Family Loxoconchidae , 1925
Genus Loxocorniculum & , 1963
Loxocorniculum? narendrai , & sp. nov.
(Pl. 5 ![]() , figs.
A-G)
, figs.
A-G)
Derivatio nominis: In honor of Dr. Narendra Kumar for his significant contribution to the knowledge of the Potiguar Basin.
Material: 1215 specimens.
Holotype: C, f, ULVG-9924
(Pl. 5 ![]() , figs. A-B), sample 272.40 m.
, figs. A-B), sample 272.40 m.
Paratypes: ULVG-9925, ULVG-10681, ULVG-9926.
Dimensions: Holotype: L: 0.390, H: 0.252, W: 0.202. Paratypes: L: 0.377-0.410, H: H: 0.224-0.249, W: 0.198-0.202.
Type-locality: 9-MO-13-RN, coordinates UTM: 682595E / 9428410N (zone 24S), 272.40 m, Potiguar Basin, Brazil.
Diagnosis: A species assigned with doubt to the genus Loxocorniculum, because it lacks the diagnostic "hornlike protuberance on posterodorsum", characterized by very compressed anterior and posterior regions, straight dorsal margin, eye tubercle developed, surface with a distinct reticulation and a weak posterodorsal rib.
Description: Carapace small, sub-rhomboidal in lateral view, inflated in dorsal view. LV larger than the right one, with overlap not pronounced. Maximum height and width at mid-length. Dorsal margin straight, ventral margin slightly convex. Anterior margin obliquely rounded, with an extremity more developed in males; posterior margin almost truncated and projected upwards. Anterior, posterior and posteroventral regions strongly compressed. Surface covered by reticulation of different sizes. The reticulum is larger in the middle part of the valves and shorter towards the periphery. In the anterior and posterior regions, where the carapace is compressed, the reticulum is large and with thin muri. A small rib, starting below and after the eye tubercle, is present in the dorsal region. Presence of a well marked tubercle in the posteroventral region. Prominent eye tubercle. Hinge gongylodont with the anterior element of the LV composed of a tooth and a very small socket separated from the posterior one (elongated horseshoe-shaped socket with a prominent tooth in the middle) by a crenulated bar. Central muscle scars composed of only a vertical row of four elongate adductor scars and a rounded frontal scar.
Sexual dimorphism: males longer and narrower than females.
Age: Turonian.
Remarks: Loxocorniculum? narendrai sp. nov. resembles Loxocorniculum miocenicum et al., 1992, from the Miocene of Tunisia. The Brazilian species, however, differs by the less robust ornamentation and the absence of the typical posterodorsal tubercle.
Genus Phlyctocythere , 1958
"Phlyctocythere"? POT 1
(Pl. 5 ![]() , figs.
H-J)
, figs.
H-J)
Material: 31 specimens.
Brief description: carapace very small, fragile, sub-rhomboidal in lateral view, inflated in the anteromedian region in dorsal view. Valves nearly equal, the LV being slightly larger than the right one. Dorsal and ventral margins nearly straight. Anterior margin sub-rounded, posterior margin projected upwards. Anterior, posterior and posteroventral regions strongly compressed. Surface smooth. Sexual dimorphism with males longer and narrower than females.
Age: Turonian.
Remarks: The Cretaceous records of this genus are few and uncertain. (2002), in a review of the biochronology of ostracodes from Morocco, cites the occurrence of Phlyctocythere? in the upper Cenomanian of Agadir Basin. (1985) described Phlyctocythere citreum in the Cenomanian-Turonian of Algeria, also recorded by et al. (1995) in the Cenomanian of Tunisia. P. citreum is shorter and has the dorsal margin more convex and the anterior part less compressed than the Brazilian species. The carapace of P. citreum is poorly calcified, which usually results in poor preservation, commonly with a broken posterior end. More recently, et al. (2013) recorded a very similar species in the Cenomanian of Morocco, identified as Pseudocythere sp. 21 which presents a more straight dorsal margin. Probably, this is the first record of the genus in the Brazilian Cretaceous.
Family Trachyleberididae , 1948
Genus Bicornicythereis , 2009
Bicornicythereis? POT 1
(Pl. 5 ![]() , figs.
K-L)
, figs.
K-L)
Material: 1 specimen.
Brief description: carapace of medium size, robust, sub-rectangular in lateral view, very compressed anteriorly and posteriorly in dorsal view. LV overlaps the RV, less pronounced in the dorsal margin. Maximum height at anterior cardinal angle. Greatest width at posterior third. Dorsal margin almost straight, ventral margin convex. Anterior margin broadly rounded, posterior margin sub-triangular and projected downwards. Anterior and posterior regions compressed. Ornamentation represented by three strong and large ribs: ventral, median and dorsal, the two last ones linked in the posterodorsal part; both dorsal and ventral ribs almost linked in the posterior region, forming a kind of vertical rib. Intercostal area reticulated. Subcentral and eye tubercles present.
Age: Turonian.
Remarks: This species is similar to Bicornicythereis bicornis (, 1929), from the Santonian to Campanian of North America (see revision of the genus in , 2009) being, however, significantly smaller. On the other hand, the sinuosity of the ventral margin and the presence of more robust ribs indicate that they might even belong to different genera.
Genus Haughtonileberis , 1969
Haughtonileberis dinglei , & sp. nov.
(Pl. 5 ![]() , figs.
M-S)
, figs.
M-S)
1992 Dumontina sp. , p. 330, Pl. 1, figs. 21, 25.
Derivatio nominis: After Dr. Richard , in honor of his important contribution to the knowledge of ostracodes.
Material: 104 specimens.
Holotype: C, f,
morphotype 1, ULVG-10002
(Pl. 5 ![]() , fig. N), sample 277.25 m.
, fig. N), sample 277.25 m.
Paratypes: ULVG-10676 (morphotype 1), ULVG-10001 (morphotype 1), ULVG-10009 (morphotype 2), ULVG-10603 (morphotype 2), ULVG-10008 (morphotype 2).
Dimensions: Holotype: L: 0.654, H: 0.324, W: 0.280. Paratypes: L: 0.624-0.760, H: 0.260-0.350, W: 0.220-0.265.
Type-locality: 9-MO-13-RN, coordinates UTM: 682595E / 9428410N (zone 24S), 277.25 m, Potiguar Basin, Brazil.
Diagnosis: Species of Haughtonileberis sub-rectangular and elongate in lateral view; finely to strongly reticulated, with the median and dorsal ribs linked, in the posterior zone, by a small vertical rib.
Description: Carapace of medium (females) to large (males) size, sub-rectangular in lateral view. In dorsal view, outline sub-rectangular, with anterior and posterior zones compressed. LV larger than RV. Greatest height at anterior cardinal angle. Dorsal and ventral margins straight, the latter with a concavity before the mid-length. Anterior margin broadly rounded, posterior sub-triangular. Anteromarginal rib prominent. Dorsal rib convex and protruding slightly beyond the margin; ventral rib weakly developed; median rib parallel to the dorsal one and bifurcated in the anterior region. The dorsal and median ribs are linked by a small vertical rib in the posterior end. Subcentral tubercle poorly developed. Spines in the anterior, posterior and posteroventral margins. Eye tubercle moderately developed. Internal features not seen. The morphotypes differ in the ornamentation:
Morphotype 1: intercostal region strongly reticulated and ribs less developed.
Morphotype 2: intercostal region finely reticulated and ribs well developed.
Sexual dimorphism: Males more elongated and narrower than females.
Age: Turonian.
Stratigraphic and geographic distribution: Turonian-Santonian of Nigeria (, 1992); Turonian (this work).
Remarks: Haughtonileberis propeplanus (1995) from the Late Santonian of Morocco is similar in the outline, but differs by the much more developed marginal rib, the stronger spines in the anterior margin and the dorsal and the median rib, which are not linked. H. mdaouerensis ( & , 1969), from the Turonian of Algeria, also recorded by (1985) in the same age and country has not the bifurcated median rib and the dorsal and median ribs are not linked. Cythereis aff. mdaouerensis ( & , 1969) recorded by (1973a) in the Cenomanian of Gabon is much more inflated and compressed in the anterior region. "Dumontina" GA E 20 , 1979 (Turonian of Gabon), is probably a representative of Haughtonileberis, but has a straighter ventral margin and it is less spinose in the anterior and posteroventral margins. According to the original description (, 1966), Dumontina has no frontal lobe (though it has a small eye tubercle) and has a strong anteromarginal rib.
Haughtonileberis POT 1
(Pl. 6 ![]() , figs.
A-C)
, figs.
A-C)
Material: 10 specimens.
Brief description: Carapace medium (females) to large (males) in size, sub-rectangular in lateral view, narrow in dorsal view. Maximum height at anterior region. Dorsal and ventral margins almost straight, converging slightly posteriorly. Anterior margin broadly rounded, posterior margin sub-triangular. Anterior and posterior regions strongly compressed. Thin anteromarginal rib. Surface ornamented with three subparallel, sharp and unconnected ribs: ventral, dorsal and median. A small rib below the eye tubercle is present. Intercostal surface smooth. Anterior margin denticulate. Subcentral and eye tubercles weakly developed. Sexual dimorphism: females higher, shorter and slightly wider than males.
Age: Turonian.
Remarks: This species is similar to "Imhotepia" GA C 8 , 1979 (Cenomanian of Gabon), but it is higher, narrower and the subcentral tubercle is less developed.
Genus Jandairella , & gen. nov.
Derivatio nominis: From the Jandaíra Formation, Potiguar Basin, Brazil.
Type-species: Jandairella obesa , & sp. nov.
Diagnosis: Carapace sub-rectangular in lateral view, with anterior margin rounded, posterior margin sub-triangular, subparallel and nearly straight dorsal and ventral margins. Overlap of the RV pronounced at anterodorsal and posterodorsal regions. Surface with three subparallel weak ribs; intercostal region with small and dense punctuation. The median ridge is connected with the subcentral tubercle. Eye tubercle prominent, subcentral tubercle present. Spines in the anterior and posterior margins. Anteromarginal rib absent. Anterior and posterior regions compressed. Robust holamphidont hinge: in the RV it consists of an anterior tooth and a posterior lobate tooth separated by a smooth median groove, anteriorly enlarged forming a deep socket. Central muscle scars: frontal V-shaped and four adductors scars arranged in semi-circle; mandibular scar elliptical. Very strong sexual dimorphism, with females higher and more inflated than males.
Stratigraphic and geographic distribution: Turonian, Potiguar Basin, Brazil.
Remarks: Both external and internal features of Jandairella gen. nov. are consistent with the characteristics of the Trachyleberididae and according to its morphology it cannot be included in any known genus of this family. Sergipella , 1967, emend. et al., 2012, is possibly the closest genus, but Jandairella has no anteromarginal rib, a less prominent "hinge ear", ribs less developed, with the median one connected to the subcentral tubercle and posterior margin more symmetrical, not upturned. Until now it can be considered as a monospecific genus.
Jandairella obesa , & sp. nov.
(Pl. 6 ![]() , figs.
D-M)
, figs.
D-M)
2000 Trachyleberididae sp. P5 , , & , p. 435, figs. 21, 5-7.
Derivatio nominis: After the general aspect of the female carapace.
Material: 79 specimens.
Holotype: C, f, ULVG-10012
(Pl. 6 ![]() , figs. D-E), sample 272.40 m.
, figs. D-E), sample 272.40 m.
Paratypes: ULVG-10634, ULVG-10013, ULVG-10014, ULVG-10015, ULVG-10016, ULVG-10017.
Dimensions: Holotype: L: 0.684, H: 0.420, W: 0.334. Paratypes: L: 0.676-0.773, H: 0.375-0.443, W: 0.323-0.390.
Type-locality: 9-MO-13-RN, coordinates UTM: 682595E / 9428410N (zone 24S), 272.40 m, Potiguar Basin, Brazil.
Diagnosis: A species of Jandairella with the posterior margin sub-triangular to slightly rounded; inflated in dorsal view; surface with three weak ribs, almost straight and parallel; intercostal area with dense and small punctuation; eye tubercle large, sub-central tubercle moderately developed.
Description: Carapace of medium (females) to large (males) size, sub-rectangular in lateral view. LV larger than RV, with more pronounced overlap in the anterodorsal and posterodorsal margins. Greatest height before the anterior third, at anterior cardinal angle, greatest width near the mid-length. Dorsal margin straight, ventral margin with a slight concavity in front of the mid-length. Anterior margin obliquely rounded, posterior margin sub-triangular to sub-rounded. Ornamentation: three fine ribs, median, ventral and dorsal. The median rib is fused to the subcentral tubercle; the ventral rib is curved and it is bifurcated in the posterior region. The dorsal rib, strongly convex, starts in the anterodorsal region, under the eye tubercle, and ends in the posterodorsal region. Intercostal region densely covered by punctuation. Posterior and anterior margins spinoses. Eye tubercle developed and rounded. In dorsal view, carapace very inflated in females, thinner in males with well marked dorsal and median ribs. Hinge and muscle scars as in the genus.
Sexual dimorphism: pronounced, with females higher and much more inflated than males.
Age: Turonian.
Stratigraphic and geographic distribution: Turonian, Potiguar Basin, Brazil ( et al., 2000; this work).
Remarks: There are no other species of the same genus to be compared.
Genus Nigeria , 1963
Nigeria cf. rotunda , sensu , 1991
(Pl. 6 ![]() , figs.
N-R)
, figs.
N-R)
1991 Veenia (Nigeria) cf. rotunda , 1978 - , p. 687-689, Pl. 70, figs. 1-5.
Material: 23 specimens.
Brief description: Carapace large, sub-rectangular in lateral view, inflate in dorsal view. LV larger than the right one, with more pronounced overlap in the anterodorsal and posterodorsal margins. Greatest height at anterior cardinal angle, greatest width at posterior third. Dorsal margin straight, ventral margin slightly concave. Anterior margin broadly rounded, posterior margin sub-triangular. Anterior and posterior regions compressed. Surface reticulate. The reticulation is larger near the sub-central tubercle and decreases in size towards the periphery of the valves. Dorsal rib poorly developed. Four spines present in the posteroventral region. Eye tubercle well developed. Sexual dimorphism present, with females proportionally higher and wider.
Age: Turonian.
Stratigraphic and geographic distribution: Cenomanian of Morocco (, 1991) and Turonian of Potiguar Basin, Brazil (this work).
Remarks: The specimens recorded in Potiguar Basin are smaller than Veenia (Nigeria) cf. rotunda recorded by (1991), in the Cenomanian of Morocco. Despite this difference, we prefer to consider them as the same species due to the strong similarity in shape and ornamentation.
Genus Potiguarella , & gen. nov.
Derivatio nominis: From Potiguar Basin.
Type-species: Potiguarella grosdidieri , & sp. nov.
Other species: P. coimbrai sp. nov and P. POT 1.
Diagnosis: Medium to large size carapace, sub-rectangular in lateral view; tricostate with the dorsal rib strongly convex, starting below and behind the eye tubercle; median rib irregular, thin to relatively thick and acute, with sub-central tubercle weak and linked to it; ventral rib short and not connected with the anteromarginal rib; intercostal area reticulate. Internal features not seen.
Stratigraphic and geographic distribution: Turonian, Potiguar Basin, Brazil.
Remarks: Although the internal features were not observed, all the external characteristics of the carapaces correspond to those of the family Trachyleberididae. The most similar genera are Rehacythereis , 1973, and Cythereis , 1849. The new genus differs from Rehacythereis by the weaker sub-central tubercle and the ventral rib not connected with the anteromarginal rib; the dorsal rib of Cythereis is discontinuous and represented by a row of spines and the ventral rib is connected with the anteromarginal rib too.
Potiguarella grosdidieri , & sp. nov.
(Pl. 7 ![]() , figs.
A-H)
, figs.
A-H)
1979 "Rehacythereis" GA E 10 , Pl. 2, fig. 5.
1979 "Rehacythereis" GA F 18 , Pl. 2, fig. 6.
2000 Cythereis? sp. P11 , , & , p. 434, Fig. 20, 1-2.
Derivatio nominis: In honor of the late Dr. Emmanuel for his important contribution to the knowledge of African ostracodes.
Material: 62 specimens.
Holotype: C, f, morphotype 1,
ULVG-9998
(Pl. 7 ![]() , figs. A-B), sample: 272.40 m.
, figs. A-B), sample: 272.40 m.
Paratypes: ULVG-9999 (morphotype 1), ULVG-10689 (morphotype 1), ULVG-10647 (morphotype 1), ULVG-10649 (morphotype 1), ULVG-9993 (morphotype 2), ULVG-10663 (morphotype 2).
Dimensions: Holotype: L: 0.910, H: 0.483, W: 0.433. Paratypes: L: 0.665-1.100, H: 0.373- 0.480, W: 0.272-0.410.
Type-locality: 9-MO-13-RN, coordinates UTM: 682595E / 9428410N (zone 24S), 272.40 m, Potiguar Basin, Brazil.
Diagnosis: A species of the genus Potiguarella characterized by a very robust carapace, strongly reticulated, with sharp and pronounced ribs and post-ocular sulcus well marked.
Description: Carapace very large, sub-rectangular in lateral view, very compressed anteriorly and posteriorly in dorsal view. LV larger than RV, with more pronounced overlapping in the posterodorsal margin. Greatest height at anterior cardinal angle, greatest width in the posterior region. Dorsal margin straight, partially covered by the dorsal rib. Ventral margin almost straight, with a concavity in front of the mid-length. Anterior margin broadly rounded, posterior margin sub-triangular. Anterior region strongly compressed, with a very prominent anteromarginal rib. Dorsal rib convex starting in the anterodorsal region, after the deep ocular sulcus, extending until the posterodorsal region. Median rib very irregular extending from the subcentral tubercle to the posteromedian region. Ventral rib straight in the middle, convex in the anterior extremity. Intercostal region heavily reticulated. In the posterior compressed region, the reticulation is more tenuous. Posteroventral region with four stout spines. Anterior margin with small spines. The morphotype 2 is smaller and with weak ornamentation pattern. Internal features not seen.
Sexual dimorphism: Males more elongated than females.
Age: Turonian.
Stratigraphic and geographic distribution: Turonian, Gabon (, 1979) and Turonian, Potiguar Basin, Brazil ( et al. 2000; this work).
Remarks: This species resembles Cythereis gabonensis , 1973a (Cenomanian-Turonian of Gabon), but differs in the rib pattern. The median rib of C. gabonensis is divided in two parts; the ventral rib is bifurcated and connected with the anteromarginal rib.
Potiguarella coimbrai , & sp. nov.
(Pl. 7 ![]() , figs.
I-N)
, figs.
I-N)
aff. 1979 Rehacythereis GA F 10 , p. 16, Pl. 3, figs. 15a-c.
Derivatio nominis: In honor of Dr. Joăo Carlos for his important contribution to the knowledge of Brazilian ostracodes.
Material: 24 specimens.
Holotype: C, m,
ULVG-9990
(Pl. 7 ![]() , figs. I-J), sample 319.80 m.
, figs. I-J), sample 319.80 m.
Paratypes: ULVG-10554, ULVG-10660.
Dimensions: Holotype: L: 0.880, H: 0.419, W: 0.431. Paratypes: L: 0.780-0.860, H: 0.420-0.460, W: 0.380-0.520.
Diagnosis: A species of the genus Potiguarella characterized by a broadly rounded anterior margin, very convex dorsal rib and rounded reticulum ornamentation.
Description: Carapace large, elongated and sub-rectangular in lateral view, with parallel margins in dorsal view, anteriorly and posteriorly compressed. LV larger than RV, with pronounced overlapping in the posterior margin. Greatest height at anterior cardinal angle. Maximum length below the mid-height and maximum width in front of the mid-length. Dorsal margin almost straight and partially hidden by the dorsal rib. Ventral margin very slightly sinuous. Anterior margin broadly rounded, posterior margin strongly triangular and acuminate. Anterior and posterior regions very compressed. Ornamentation: reticulated, with the reticulum rounded and larger in the central region of the carapace and punctuated in the posterior region; posteroventral margin with few spines. Anterior margin denticulate. Anteromarginal rib weakly developed. Presence of three sub-parallel and horizontal ribs not connected: the dorsal one very pronounced, sharp and strongly convex, the median one thin and fused to subcentral tubercle and the ventral one less prominent and slightly convex. Eye tubercle present. Internal features not seen.
Sexual dimorphism: Females less elongated and wider than males.
Age: Turonian.
Remarks: (1979) recorded a very similar species, Rehacythereis GA F 10, from the Turonian of Gabon, but it differs in the ornamentation, which consists mainly of polygonal reticulations and in the anterior region, which is less compressed in the African species. P. i sp. nov. has a less strongly convex dorsal rib, a polygonal reticulation and a much more marked anteromarginal rib.
Potiguarella POT 1
(Pl. 8 ![]() , figs.
A-C)
, figs.
A-C)
Material: 17 specimens.
Brief description: Carapace large, sub-rectangular in lateral view, inflated in dorsal view, with the anterior and posterior regions very compressed. LV larger than the right one, with most marked overlap in the ventral margin. Greatest height at anterior cardinal angle. Dorsal margin straight, partially covered by the dorsal rib, ventral margin slightly concave. Anterior margin rounded, posterior margin sub-triangular to sub-rounded. Anterior region very compressed. Surface reticulate with three prominent ribs: dorsal rib strongly convex, median rib slightly convex and ventral rib slightly sinuous. Anteromarginal rib moderately marked. Eye tubercle present, sub-central tubercle poorly developed.
Age: Turonian.
Remarks: The posterior margin of this species is less elongated and much more rounded than P. grosdidieri sp. nov. and P. coimbrai sp. nov.
Subfamily Brachycytherinae , 1954
Genus Brachycythere , 1933
"Brachycythere" aff. jodhpurensis , 1997
(Pl. 8 ![]() , figs.
D-I)
, figs.
D-I)
aff. 1997 Brachycythere jodhpurensis , p. 9-10, Pl. 2, figs. 9-12; Pl. 3, fig. 1.
aff. 2007 Brachycythere jodhpurensis - et al., Pl. 3, figs. 11-13.
Material: 283 specimens.
Brief description: Carapace of medium (females) to large (males) size, sub-triangular in lateral view, inflated in dorsal view. LV larger than RV, with overlap more pronounced in the dorsal margin. Maximum height at anterior cardinal angle; maximum width behind the mid-length. Dorsal and ventral margins almost straight. Anterior margin asymmetrically rounded, posterior margin sub-triangular and projected downward. Anterior and posterior regions compressed. Surface densely punctuated, except in the anterior and posterior regions. Ventral region flat and with parallel rows of punctuations. Anterior denticles present. Ventrolateral rib well developed, more clearly observed in ventral view. Eye tubercle small, post-ocular sulcus deep. Sexual dimorphism present with females higher and more inflated than males.
Age: Turonian.
Remarks: This species is similar to B. jodhpurensis , 1997, from theTuronian-Coniacian of India, but differs by its much more elongated general outline and by the position of the maximum width. B. ventrocomplanatus , & , 2000, also from the Upper Cretaceous Jandaíra Formation, Potiguar Basin, is similar but it is less elongated and with the ventrolateral rib less pronounced. This species is also similar to B. multidifferentis & , 2013, from the Cenomanian-Turonian of the Santos Basin, Brazil, but differs in the position of maximum width and in the more compressed anterior region. According to (2002) Brachycythere is restricted to North America and the brachycytherines of South America and Africa should belong to a different, but undescribed genus (& , in prep.).
"Brachycythere" POT 1
(Pl. 8 ![]() , figs.
J-N)
, figs.
J-N)
Material: 25 specimens.
Brief description: Carapace large, sub-triangular in lateral view, inflated in dorsal view. LV larger than RV, with overlap more pronounced in dorsal, ventral and posterior margins. Maximum height at anterior cardinal angle; maximum width at mid-length. Dorsal margin almost straight; ventral margin of the RV almost straight, slightly convex in the LV. Anterior margin broadly and obliquely rounded; posterior margin sub-triangular, strongly acute and projected downwards. Carapace very compressed in the anterior and posterior regions. The middle part of the carapace is punctuated; the anterior and posterior regions are almost smooth. Eye tubercle poorly developed. Sexual dimorphism present: females shorter, higher and wider than males.
Age: Turonian
Remarks: This species differs from Brachycythere aff. jodhpurensis (see above) by the much more acuminate posterior margin and by the position of maximum width. B. reymenti , 1989, recorded in the Campanian of Iran resembles "B." POT 1 due to its acuminate posterior end, but B. reymenti is more elongated and the posterior projection is located near the mid-height.
Family Xestoleberididae , 1928
Genus Xestoleberis , 1866
Xestoleberis POT 1
(Pl. 8 ![]() , figs.
O-P)
, figs.
O-P)
Material: 60 specimens.
Brief description: Carapace small, suboval in lateral and dorsal views. LV larger than RV, with overlap less pronounced in the ventral margin. Maximun height near the mid-length; maximum width at the posterior third. Dorsal margin convex, ventral margin almost straight. Anterior margin obliquely rounded, posterior margin rounded. Surface smooth.
Age: Turonian.
Remarks: Xestoleberis POT 1 differs from Xestoleberis dissimilissummis , 1996 from the Santonian of Morocco, mainly by the straighter ventral margin, the position of the maximum width and by the maximum height.

Click on thumbnail to enlarge the image.
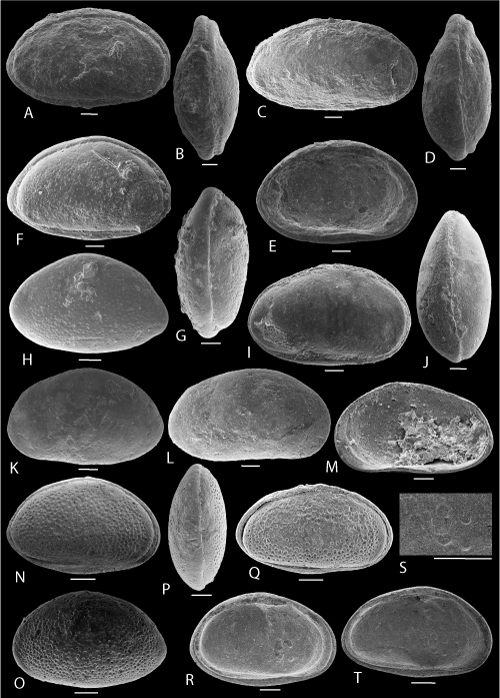
Plate 1: Scale bar: 100 µm
A-D: Cytherella mediatlasica , 1996. A: C, left view, f, ULVG-10615, sample 276.65 m; L: 0.700, H: 0.356, W: 0.280; B-C: C, f, ULVG-9845, sample 276.25 m, L: 0.735, H: 0.365, W: 0.275, B: right view, C: DV; D: C, left view, m, ULVG-10686, sample 276.25 m, L: 0.673, H: 0.356, W: 0.239.
E-H: Bairdoppilata potiguarensis (, & , 2000). E-F: C, ULVG-9856, sample 275.80 m, L: 1.024, H: 0.644, W: 0.344, E: right view, F: DV; G: LV, IV; ULVG-9857, sample 272.40 m, L: 0.999, H: 0.558; H: RV, IV; ULVG-9858, sample 272.40 m; L: 1.018, H: 0.641.
I- K: "Bythocypris" POT 1. I-J: C, ULVG-9872, sample 276.65 m; L: 0.767, H: 0.384, W: 0.277, I: left view, J: DV; K: C, right view, ULVG-9873, sample 277.25 m, L: 0.735, H: 0.375, W: 0260.
L-O: Paracypris aff. caudata (, 1964). L-N: C, ULVG-9883, sample 275.80 m; L: 0.820, H: 0.434, W: 0.348, L: right view, M: DV; N: left view; O: LV, IV, ULVG-9884, sample 276.25 m; L: 0.698, H: 0.374.
P-Q: Paracypris aff. dubertreti & , 1972. P-Q: C, ULVG-9877, sample 277.25 m, L: 0.627, H: 0.640, W: 0.173, P: right view, Q: DV.
R-T: Paracypris POT 1. R: C, right view, ULVG-9878, sample 276.25 m, L: 0.808, H: 0.344, W: 0.208; S-T: C, ULVG-9879, sample 276.25 m, L: 0.790, H: 0.334, W: 0.240, S: left view, T: DV.
Click on thumbnail to enlarge the image.
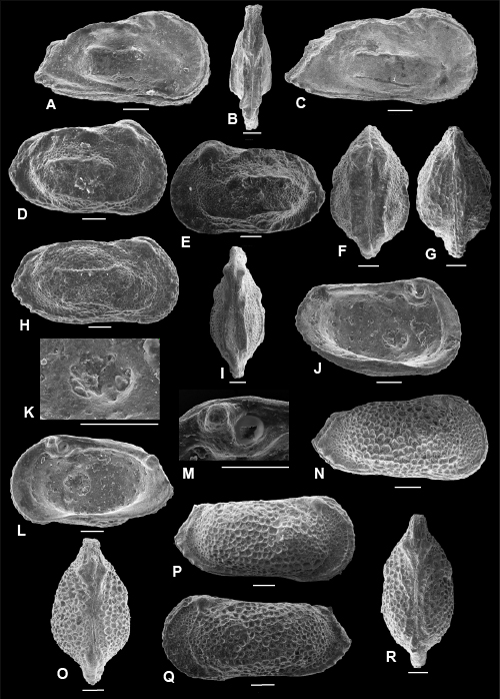
Plate 2: Scale bar: 100 µm
A-E: Cophinia grekoffi , & sp. nov. A: holotype, C, right view, f, ULVG-9941, sample 297.60 m, L: 0.900, H: 0.570, W: 0.401; B: paratype 1, C, DV, f, ULVG-9942, sample 297.60 m, L: 0.875, H: 0.515, W: 0.409; C: paratype 2, C, right view, m, ULVG-9943, sample 297.60 m, L: 0.939, H: 0.470, W: 0.409; D: paratype 3, C, DV, m, ULVG-9944, sample 297.60 m. L: 0.899, H: 0.498, W: 0.427; E: paratype 4, LV, IV, f, ULVG-9946, sample 297.60 m. L: 0.817, H: 0.521.
F-H: Cophinia POT 1. F-G: C, ULVG-9957, sample 276.65 m, L: 0.735, H: 0.465, W: 0.371, F: right view, G: DV; H: C, left view, ULVG-9958, sample 275.80 m, L: 0.737, H: 0.450, W: 0.349.
I-M: Fossocytheridea tiberti , & sp. nov. I-J: holotype: C, f, ULVG-9938, sample 319.30 m, L: 0.880, H: 0.519, W: 0.431, I: right view, J: DV; K: paratype 1, C, left view, f, ULVG-10661, sample 311.90 m; L: 0.840, H: 0.500, W: 0.400; L: RV, EV, m, ULVG-9939, sample 319.80 m, L: 0.856, H: 0.461; M: RV, IV, ULVG-9940, sample 319.80 m, L: 0.775, H: 0.424.
N-T: Ovocytheridea reymenti , & sp. nov. N-O: holotype, C, f, ULVG-9959, sample 272.40 m, L: 0.631, H: 0.386, W: 0.297, N: right view, O: left view; P: paratype 1, C, DV, f, ULVG-9960, sample 272.40 m, L: 0.647, H: 0.369, W: 0.292, Q: paratype 2, C, right view, m, ULVG-9961, sample 272.40 m, L: 0.657, H: 0.375, W: 0.280, R: paratype 3, LV, IV, f, ULVG-9962, sample 272.40 m, L: 0.653, H: 0.400, S-T: paratype 4, m, ULVG-9963, sample 272.40 m, L: 0.654, H: 0.396, S: detail of muscle scars, T: LV, IV.

Click on thumbnail to enlarge the image.
Plate 3: Scale bar: 100 µm
A-E: Ovocytheridea posteroprojecta , & sp. nov. A-C: holotype, C, f, ULVG-9976, sample 277.25 m; L: 1.040, H: 0.600, W: 0.460, A: right view, B: DV, C: left view; D: paratype 1, C, right view, m, ULVG-10486, sample 275.80 m, L: 1.100, H: 0.580, W: 0.410; E: paratype 2, LV, IV, f, ULVG-9977, sample 277.25 m, L: 0.760, H: 0.438.
F-K: Perissocytheridea caudata , & sp. nov. F-H: holotype, C, f, ULVG-9932, sample 272.40 m, L: 0.546, H: 0.322, W: 0.330, F: right view, G: DV, H: left view; I-J: paratype 1, C, m, ULVG-9933, sample 297.60 m, L: 0.606, H: 0.310, W: 0.303, I: right view, J: dorsal view; K: paratype 2, LV, IV, f, ULVG-9934, sample 297.60 m; L: 0.550, H: 0.342.
L-P: Perissocytheridea mossoroensis , & sp. nov. L-N: holotype, C, f, ULVG-9930, sample 319.80 m; L: 0.450, H: 0.259, W: 0.233, L: right view, M: DV, N: left view; O-P: paratype 1, C, m, ULVG-9931, sample: 319.80 m; L: 0.539, H: 0.272, W: 0.247, O: right view, P: DV.
Q-S: Rostrocytheridea? POT 1. Q-R: C, f, ULVG-9954, sample 276.25 m. L: 0.510, H: 0.338 mm, W: 0.283 mm, Q: right view, R: DV; S: EV, m, ULVG-9955, sample 276.25 m. L: 0.539, H: 0.289.

Click on thumbnail to enlarge the image.
Plate 4: Scale bar: 20 µm
A-E: Eucytherura aff. speluncosus (, 1996). A-B: C, f, ULVG-9908, sample 272.40 m, L: 0.357, H: 0.204, W: 0.181, A: right view, B: DV; C-D: C, m, ULVG-9909, sample 277.25 m, L: 0.399, H: 0.189, W: 0.175, C: right view, D: DV; E: C, left view, f, ULVG-10668, sample 277.25 m, L: 0.370, H: 0.210, W: 0.190.
F-I: Hemicytherura viviersae , & sp. nov. F-H: holotype, C, ULVG-9902, sample 275.80 m, L: 0.390, H: 0.251, W: 0.165, F: right view, G: DV, H: VV, I: paratype 1, C, left view, ULVG-10669, sample 277.25 m; L: 0.380, H: 0.210, W: 0.150.
J-L: Pelecocythere POT 1. J: C, right view, ULVG-9911, sample: 275.80 m, L: 0.563, H: 0.191, W: 0.252; K-L: C, ULVG-9912, sample 275.80 m, L: 0.560, H: 0.220, W: 0.293, K: DV, L: left view.
M-P: Procytherura ballentae , & sp. nov. M-N: holotype, C, ULVG-9913, sample 275.80 m, L: 0.389, H: 0.196, W: 0.123, M: right view, N: DV; O: paratype 1, C, right view, ULVG-9914, sample 276.65 m, L: 0.394, H: 0.166, W: 0.120, P: paratype 2, C, left view, ULVG-10675, sample 276.65 m, L: 0.430, H: 0.190, W: 0.140.
Q-U: Semicytherura aff. adversainflata , 1996. Q-S: C, ULVG-9915, sample 276.25 m, L: 0.352, H: 0.146, W: 0.174, Q: right view, R: DV; S: VV; T-U: C, ULVG-10667, sample 277.25 m, L: 0.370, H: 0.160, W: 0.190, T: left view, U: detail of ornamentation of the central part of the carapace.
Click on thumbnail to enlarge the image.
Plate 5: Scale bar: 20 µm (A-J); 100 µm (K-S)
A-G: Loxocorniculum? narendrai , & sp. nov. A-B: holotype, C, f, ULVG-9924, sample 272.40 m, L: 0.390, H: 0.252, W: 0.202, A: right view, B: DV; C-D: paratype 1, C, m, ULVG-9925, sample 272.40 m, L: 0.410, H: 0.249, W: 0.198, C: right view, D: DV; E: paratype 2, C, left view, m, ULVG-10681, sample 275.80 m, L: 0.377, H: 0.220, W: 0.202; F-G: paratype 3, LV, m, ULVG-9926, sample 272.40 m, L: 0.407, H: 0.224, F: detail of central muscle scar, G: LV, IV.
H-J: "Phlyctocythere"? POT 1. H-I: C, f, ULVG-9929, sample 275.80 m, L: 0.379, H: 0.222, W: 0.160, H: right view, I: DV; J: C, left view, m, ULVG-10480, sample 275.80 m, L: 0.420, H: 0.210, W: 0.140.
K-L: Bicornicythereis? POT 1. K-L: C, ULVG-10023, sample 285.30 m, L: 0.522, H: 0.269, W: 0.241, K: DV, L: right view.
M-S: Haughtonileberis dinglei , & sp. nov. M: holotype, morphotype 1, C, right view, f, ULVG-10002, sample 277.25 m, L: 0.654, H: 0.324, W: 0.280; N: paratype 1, morphotype 1, C, left view, m, ULVG-10676, sample 276.65 m; L: 0.760, H: 0.350, W: 0.260; O-P: paratype 2, morphotype 1, C, m, ULVG-10001, sample 276.65 m, L: 0.710, H: 0.328, W: 0.265, O: right view, P: DV; Q: paratype 3, morphotype 2, C, right view, m, ULVG-10009, sample 276.25 m, L: 0.624, H: 0.266, W: 0.228; R: paratype 4, morphotype 2, C, left view, m, ULVG-10603, sample 276.25 m; L: 0.640, H: 0.260, W: 0.220; S: paratype 5, morphotype 2, C, DV, m, ULVG-10008, sample 277.25 m, L: 0.639, H: 0.276, W: 0.223.
Click on thumbnail to enlarge the image.
Plate 6: Scale bar: 100 µm
A-C: Haughtonileberis POT 1. A-B: C, f, ULVG-10005, sample 272.40 m, L: 0.685, H: 0.365, W: 0.236, A: right view, B: DV; C: C, right view, m, ULVG-10006, sample 272.40 m, L: 0.718, H: 0.362, W: 0.229.
D-M: Jandairella obesa , & sp. nov. D-E: holotype, C, f, ULVG-10012, sample 272.40 m, L: 0.684, H: 0.420, W: 0.334, D: right view, E: left view; F: paratype 1, C, f, ULVG-10634, sample 272.40 m, L: 0.763, H: 0.443, W: 0.390, G: paratype 2, C, VV, f, ULVG-10013, sample 272.40 m, L: 0.770, H: 0.417, W: 0.323, F: DV, G: VV; H: paratype 3, C, right view, m, ULVG-10014, sample 272.40 m, L: 0.742, H: 0.396, W: 0.360; I: paratype 4, C, DV, m, ULVG-10015, sample 272.40 m; L: 0.773, H: 0.406, W: 0.325; J-K: paratype 5, RV, ULVG-10016, sample 272.40 m, L: 0.678, H: 0.390, J: LV, IV, K: detail of muscle scars; L-M: paratype 6, LV, ULVG-10017, sample 272.40 m; L: 0.676, H: 0.375, L: RV, IV, M: detail of the hinge anterior element.
N-R: Nigeria cf. rotunda , sensu , 1991. N-O: C, f, ULVG-9980, sample 272.40 m, L: 0.694, H: 0.331, W: 0.365, N: right view, O: DV; P-R: C, m, ULVG-9981, sample 272.40 m, L: 0.756, H: 0.345, W: 0.338, P: right view, Q: left view, R: DV.
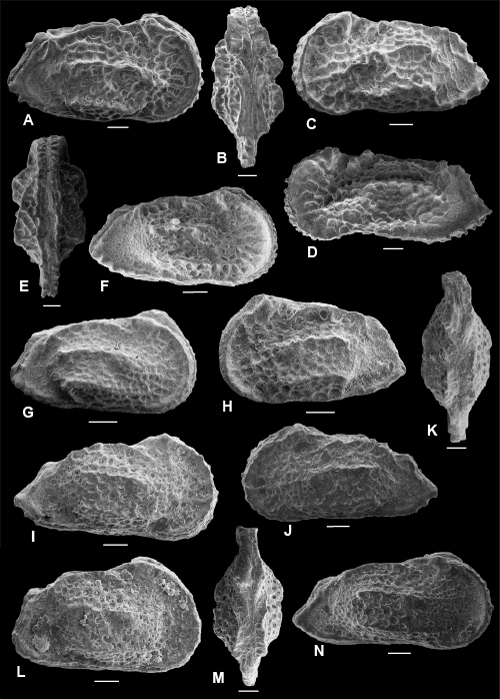
Click on thumbnail to enlarge the image.
Plate 7: Scale bar: 100 µm
A-H: Potiguarella grosdidieri , & sp. nov. A-B: holotype, morphotype 1, C, f, ULVG-9998, sample: 272.40 m, L: 0.910, H: 0.483, W: 0.433, A: right view, B: DV; C: paratype 1, morphotype 1, C, left view, f, ULVG-9999, sample: 272.40 m, L: 0.890, H: 0.458, W: 0.408; D: paratype 2, morphotype 1, C, left view, m, ULVG-10689, sample: 272.40 m, L: 1.100, H: 0.450, W: 0.400; E: paratype 3, morphotype 1, C, VV, f, ULVG-10647, sample 272.40 m, L: 0.900, H: 0.480, W: 0.410; F: paratype 4, morphotype 1, C, right view, juvenile, ULVG-10649, sample 272.40 m; L: 0.800, H: 0.400, W: 0.350; G: paratype 5, morphotype 2, C, right view, f, ULVG-9993, sample 272.40 m. L: 0.678, H: 0.396, W: 0.272; H: paratype 6, morphotype 2, C, left view, f, ULVG-10663, sample 304.10, L: 0.665, H: 0.373, L: 0.250.
I-N: Potiguarella coimbrai , & sp. nov. I-K: holotype, C, m, ULVG-9990, sample 319.80 m; L: 0.880, H: 0.419, W: 0.431, I: right view, J: left view, K: DV; L-M: paratype 1, C, f, ULVG-10554, sample 319.30 m, L: 0.780, H: 0.460, W: 0.520, broken, L: right view, M: DV; N: paratype 2, C, right view, m, ULVG-10660, sample 319.30 m; L: 0.860, H: 0.420, W: 0.380.
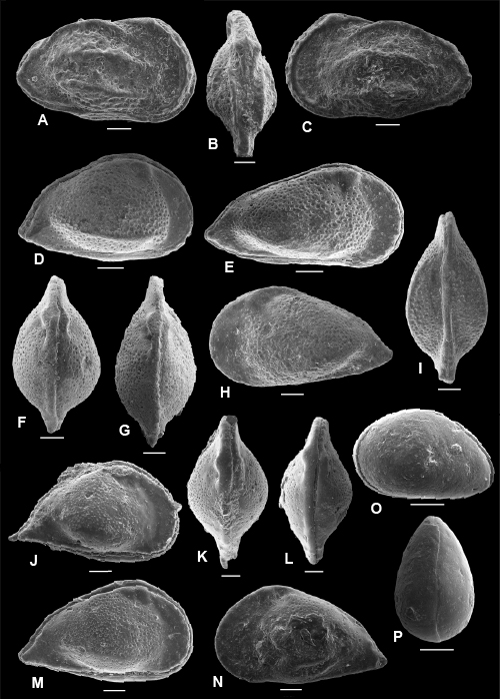
Click on thumbnail to enlarge the image.
Plate 8: Scale bar: 100 µm
A-C: Potiguarella POT 1. A: C., right view, ULVG-10019, sample 319.30 m, L: 0.755, H: 0.426, W: 0.325; B-C: C, ULVG-10020, sample 319.80 m, L: 0.747, H: 0.419, W: 0.350, B: DV, C: left view.
D-I: "Brachycythere" aff. jodhpurensis , 1997. D: C, right view, f, ULVG-10028, sample 272.40 m, L: 0.684, H: 0.407, W: 0.373; E: C, right view, m, ULVG-10031, sample 272.40 m, L: 0.739, H: 0.390, W: 0.352; F: C, DV, f, ULVG-10586, sample 272.40 m, L: 0.690, H: 0.390, W: 0.390; G: C, DV, m, ULVG-10030, sample 272.40 m, L: 0.760, H: 0.380, W: 0.370; H: C, left view, m, ULVG-10587, sample 272.40 m, L: 0.740, H: 0.385, W: 0.380; I: C, VV, f, ULVG-10588, sample 272.40 m, L: 0.710, H: 0.430 mm, W: 0.385.
J-N: "Brachycythere" POT 1. J-K: C, f, ULVG-10025, sample 276.65 m, L: 0.801, H: 0.445, W: 0.402, J: right view, K: DV; L-M: C, m, ULVG-10026, sample 275.80 m; L: 0.799, H: 0.441, W: 0.380, L: DV, M: right view; N: C, left view, m, ULVG-10027, sample 277.25 m; L: 0.800, H: 0.430, W: 0.390.
O-P: Xestoleberis POT 1. O: C, right view, ULVG-10047, sample: 275.80 m, L: 0.426, H: 0.291, W: 0.283; P: C, DV, ULVG-10683, sample: 275.80 m; L: 0.410, H: 0.280, W: 0.280.
Appendix 1: Illustrated species.
This appendix (Pls. 9-11) presents the taxa which remained in open nomenclature or undetermined due to their scarcity and poor preservation, or that were not included in the taxonomy chapter due to their reduced stratigraphic value.
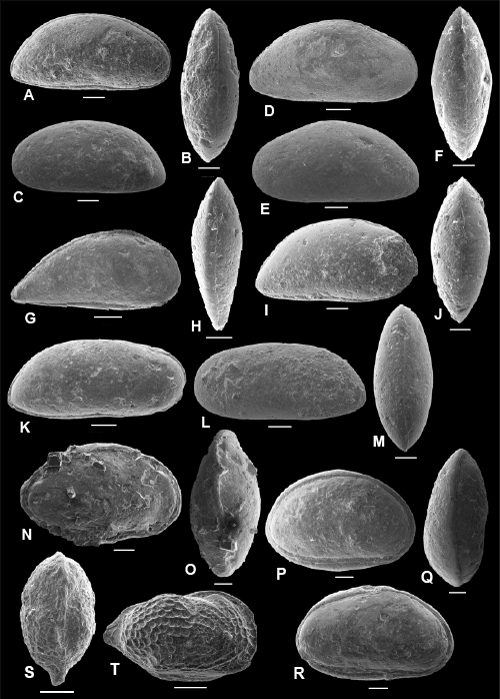
Click on thumbnail to enlarge the image.
Plate 9: Scale bar: 100 µm
A-C: Paracypris POT 2. A-B: C, ULVG-9885, sample 277.25 m, L: 0.754, H: 0.373, W: 0.311, A: right view, B: DV; C: C, left view, ULVG 9986, 277.25 m, L: 0.720, H: 0.320, W: 0.294.
D-F: Paracypris POT 3. D-F: C, ULVG-9887, sample 275.80 m, L: 0.730, H: 0.370, W: 0.270, D: right view, E: left view, F: DV.
G-H: Paracypris POT 4. G-H: C, ULVG-9889, sample 275.80 m, L: 0.718, H: 0.342, W: 0.254, G: right view, H: DV.
I-J: Paracypris POT 5. I-J: C, ULVG-9890, sample 275.80 m, L: 0.749, H: 0.407, W: 0.365, I: right view, J: DV.
K-M: Pontocyprella POT 1. K; M: C, ULVG-9895, sample 275.80 m, L: 0.700, H: 0.314, W: 0.283, K: right view, M: DV; L: C, left view, ULVG-9897, sample 276.65 m, L: 0.710, H: 0.330, W: 0.260.
N-O: Cophinia POT 2. N-O: C, ULVG-9974, sample 327.80 m, L: 0.782, H: 0.473, W: 0.350, N: right view, O: DV.
P-R: Ovocytheridea? aff. palapiformis , 1973a. P-Q: C, f, ULVG-9969, sample 297.60 m, L: 0.810, H: 0.520, W: 0.380, P: right view, Q: DV; R: C, right view, m, ULVG-9970; sample 272.40 m, L: 0.791, H: 0.470, W: 0.358.
S-T: Perissocytheridea? POT 1. S-T: C, ULVG-9919, sample 297.60 m, L: 0.420, H: 0.240, W: 0.240, S: DV, T: right view.
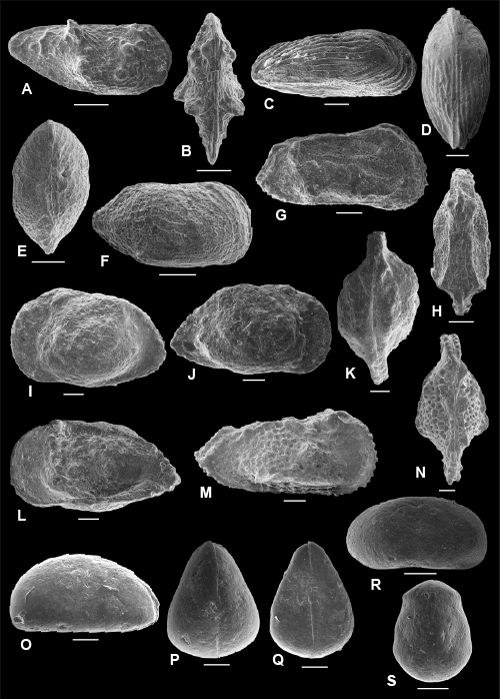
Click on thumbnail to enlarge the image.
Plate 10: Scale bar: 100 µm
A-B: Acuminacythere? POT 1 A-B: C, ULVG-9910, sample 276.65 m, L: 0.308, H: 0.138, W: 0.145, A: right view, B: DV.
C-D: Metacytheropteron? POT 1. C-D: C, ULVG-9903, sample 272.60 m, L: 0.525, H: 0.262, W: 0.210, C: right view, D: DV.
E-F: Semicytherura POT 1. E-F: C, ULVG-9916, sample 319.80 m, L: 0.424, H: 0.220, W: 0.257, E: DV, F: right view.
G-H: Curfsina? POT 1. G-H: C, ULVG-10003, sample 276.65 m, L: 0.619, H: 0.300, W: 0.209, G: right view, H: DV.
I-L: "Brachycythere"? POT 2. I; K: C, ULVG-10034, sample 271.40 m, L: 0.800, H: 0.440, W: 0.420, I: left view, K: DV; J; L: RV, ULVG-10035; 271.40 m, L: 0.800, H: 0.400, J: EV, L: RV, IV.
M-N: Trachyleberididae indet. gen. 1. M-N: C, ULVG-10024, sample 272.40 m, L: 0.889, H: 0.429, W: 0.410, M: right view, N: DV.
O-Q: Platyleberis? POT 1. O-P: C, ULVG-10048, sample 275.80 m, L: 0.480, H: 0.222, W: 0.340, O: right view, P: DV; Q: C, VV, ULVG-10670, sample 277.25 m, L: 0.450, H: 0.220, W: 0.320.
R-S: Xestoleberididae indet. gen. 1. R-S: C, ULVG-10049, sample 276.65 m, L: 0.485, H: 0.251, W: 0.380, R: right view, S: DV.

Click on thumbnail to enlarge the image.
Plate 11: Scale bar: 100 µm
A-D: Indet. gen. 1 POT 1. A-B: C, f, ULVG-9892, sample 276.65 m, L: 0.645, H: 0.444, W: 0.416, A: right view, B: DV; C-D: C, m, ULVG-9893; 276.25 m, L: 0.614, H: 0.419, W: 0.349, C: right view, D: DV.
E-G: Indet. gen. 2 POT 1. E-F: C, f, ULVG-9952, sample 272.40 m, L: 0.675, H: 0.413, W: 0.278, E: right view, F: DV; G: C, left view, m, ULVG-9953, sample 272.40 m, L: 0.730, H: 0.370, W: 0.300.
H-J: Indet. gen. 3 POT 1. H-J: C, ULVG-9904, sample 271.40 m, L: 0.525, H: 0.262, W: 0.210, H: right view, I: DV, J: left view.
K-M: Indet. gen. 4 POT 1. K-L: C, ULVG-9917, sample 319.80 m, L: 0.537, H: 0.254, W: 0.306, K: right view, L: DV; M: C, left view, ULVG-10659, sample 319.80 m, L: 0.460, H: 0.233, W: 0.270.
N-P: Indet. gen. 5 POT 1. N-O: C, f, ULVG-9874, sample 317.90 m, L: 0.705, H: 0.441, W: 0.354, N: right view, O: DV; P: C, left view, m, ULVG-9875, sample 319.80 m, L: 0.760, H: 0.420, W: 0.302.
Diversity and dominance indexes were used to identify ecologically induced variation in Cretaceous ostracode assemblages. Studies by (1971), (1975), (1980), et al. (1982), & (1983), (1985), et al. (2012), using Shannon or Simpson indexes, have demonstrated how the diversity varies according to the environmental fluctuations. The advantage of using diversity indexes is the measurement of two attributes of the community together (richness and evenness), despite the sampling effort ( & , 1983; , 2008). The Shannon index is calculated on not only the number of species but how their abundance is distributed within the community.
The studied Turonian assemblages characterize shallow shelf environments. A total of 4,841 specimens were recorded, distributed among 53 taxa (Appendix 2). Based on the vertical distribution of the species in the well, their abundance, diversity and dominance, three ostracode assemblages have been identified, with variable ecological preferences.
This assemblage (Fig. 3 ![]() ) is situated in the interval 327.80 m - 287.10 m and it is characterized by the presence of several species mainly belonging to brackish genera, such as Perissocytheridea and Fossocytheridea. It may be divided in two sub-assemblages: Assemblage 1A (327.80m - 310.90 m) and Assemblage 1B (304.10 m - 287.10 m). The most abundant species in the first sub-assemblage are Fossocytheridea tiberti sp. nov., Perissocytheridea
mossoroensis sp. nov., Indet. Gen. 4 POT 1, and Indet. Gen. 5 POT 1. The two Indet. Gen. seem to be brackish because both possess a median sub-vertical sulcus, which is very common in brackish genera (e.g., Fossocytheridea and Perissocytheridea). In the second sub-assemblage (1B) Perissocytheridea caudata sp. nov. (~57%) is the dominant species, followed by Cophinia grekoffi sp. nov. (~18%) and Perissocytheridea? POT 1 (~10%).
) is situated in the interval 327.80 m - 287.10 m and it is characterized by the presence of several species mainly belonging to brackish genera, such as Perissocytheridea and Fossocytheridea. It may be divided in two sub-assemblages: Assemblage 1A (327.80m - 310.90 m) and Assemblage 1B (304.10 m - 287.10 m). The most abundant species in the first sub-assemblage are Fossocytheridea tiberti sp. nov., Perissocytheridea
mossoroensis sp. nov., Indet. Gen. 4 POT 1, and Indet. Gen. 5 POT 1. The two Indet. Gen. seem to be brackish because both possess a median sub-vertical sulcus, which is very common in brackish genera (e.g., Fossocytheridea and Perissocytheridea). In the second sub-assemblage (1B) Perissocytheridea caudata sp. nov. (~57%) is the dominant species, followed by Cophinia grekoffi sp. nov. (~18%) and Perissocytheridea? POT 1 (~10%).
Although some authors state that the genus Cophinia is commonly associated with typical marine species (e.g., , 1963; et al., 1998), C. grekoffi sp. nov. seems to be euryhaline, associated with brackish species and with a few marine ones (Bairdoppilata potiguarensis, Brachycythere POT 1, Cytherella mediatlasica) too. The levels 297.60 m and 287.10 m, the two highest ones of Assemblage 1, have the strongest marine signal of all this assemblage.
The Shannon values are low and the dominance is high (Fig. 4 ![]() ), which is characteristic of brackish water environments ( & ,
2006).
), which is characteristic of brackish water environments ( & ,
2006).
The mixohaline character (probably lagoon) of this first interval, especially at the base (Assemblage 1A) is reinforced by the presence of the Fossocytheridea-Perissocytheridea association. This association is considered the fossil equivalent of the Cyprideis-Perissocytheridea association found from the Neogene to the Recent. Previous studies show that these associations can occur both with marine ostracodes ( & , 1979; , 1988; , 1996; et al., 2007; & , 2010) and non-marine ostracodes ( & , 1978; et al., 1996; & , 2008; et al., 2013). The genera Cyprideis and Perissocytheridea, associated with marine ostracodes, have also been found in hyperhaline environments (, 1988).
In the upper interval of Assemblage 1B, the fauna records an increase in salinity as evidenced by the presence of the marine genera Cophinia, Bairdopillata, Brachycythere, Xestoleberis and Jandairella associated with the brackish genus Perissocytheridea.
In the interval 285.30 m - 275.80 m a highly diversified shallow marine association is recorded
(Fig. 4 ![]() ). The dominant species are Loxocorniculum? narendrai sp. nov. (~21%), Cytherella mediatlasica (~15%), "Bythocypris" POT 1 (~8%) and Bairdoppilata potiguarensis (~7%), though many species of trachyleberidids, cypridids and cytherurids were also recorded.
). The dominant species are Loxocorniculum? narendrai sp. nov. (~21%), Cytherella mediatlasica (~15%), "Bythocypris" POT 1 (~8%) and Bairdoppilata potiguarensis (~7%), though many species of trachyleberidids, cypridids and cytherurids were also recorded.
In contrast to the first assemblage, the diversity is high and the dominance is low
(Fig. 4 ![]() ), except in the sample 285.30, where only one specimen was recorded. et al.
(2012) found similar values in the Upper Cretaceous of Caribbean that was considered by the authors as indicative of normal marine and stable environments.
), except in the sample 285.30, where only one specimen was recorded. et al.
(2012) found similar values in the Upper Cretaceous of Caribbean that was considered by the authors as indicative of normal marine and stable environments.
The increase in diversity could be related to sea-level highstands that contribute to an expansion of available biotopes and promote evolutionary radiations of shallow-marine organisms.
The interval 272.60 m - 270.80 m reflects a mixture of faunas from marine and brackish water environments
(Fig. 3 ![]() ). Diversity decreases and dominance increases
(Fig. 4
). Diversity decreases and dominance increases
(Fig. 4 ![]() ), characterizing more unstable environmental conditions.
), characterizing more unstable environmental conditions.
The species Loxocorniculum? narendrai sp. nov. (40%), Ovocytheridea reymenti sp. nov. (21%) and "Brachycythere" aff. jodhpurensis (13%) are dominant. Species of the brackish genus Perissocytheridea are also representative of this assemblage and, to a lesser degree, Fossocytheridea spp. According to a study on Recent sediments from Florida, Perissocytheridea is an euryhaline genus, with a species inhabiting mesohaline waters and another preferring polyhaline waters (, 1977). This paleoecological plasticity was also observed in the Upper Cretaceous species of Morocco, where (1996) recorded Perissocytheridea konatei , 1985, associated with several marine ostracode species. Another example is P. cretacea (, & , 2010), which occurs predominantly with Fossocytheridea spp. in Upper Cretaceous deposits from Santos Basin, Brazil.
Based on these data, we can infer that the depositional environment of the Jandaíra Formation was marginal marine. Initially, a more restricted lagoonal environment was dominated by almost only brackish genera (assemblage 1A); subsequently the environment became more open in the upper part (Assemblage 1B), with the presence of typical brackish genera (dominant) mixed with some more marine ones such as Cophinia grekoffi. This is corroborated by the interpretation of (2001) who suggests a tidal flat to lagoonal facies for this interval. Superimposing these deposits is a fauna representative of higher salinity (assemblage 2), and probably deeper water, dominated by the species Loxocorniculum? narendrai, Cytherella mediatlasica and Bairdoppilata potiguarensis. Above this a reduction in salinity was probably related to the more restricted environments, though not so restricted as in assemblage 1, which is represented by tidal flat and open lagoonal facies dominated by Loxorniculum? narendrai and "Brachycythere" aff. jodhpurensis associated with the cytherideid Ovocytheridea reymenti and the brackish Perissocytheridea and Fossocytheridea.
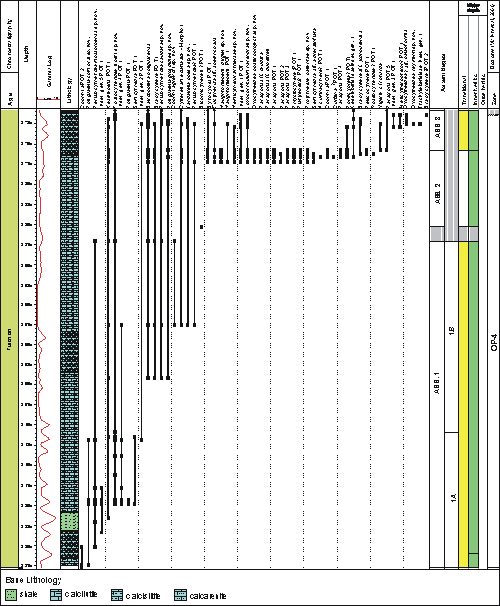
Click on thumbnail to enlarge the image.
Figure 3: Stratigraphic ranges and paleoecological assemblages of the Turonian species from the well 9-MO-13-RN.
The study by et al. (2000) of the Jandaíra Formation in more distal offshore wells records an ostracode fauna characteristic of outer neritic environment. The only species in common with the present work is Bairdoppilata potiguarensis ( et al.), which ranges from the Cenomanian to the Paleocene, according to those authors. About the ecology of Bairdoppilata, (1996) concludes that the temperature and depth are apparently less restrictive than the salinity gradient. In the case of B. potiguarensis the tolerance seems to be also relative to the salinity, because it is associated with brackish species. This difference in faunal composition is probably due to the more distal position of the wells studied by et al. (2000). Another hypothesis is that et al. (2000) did not sample the Turonian interval. However, a more accurate discussion on this subject is not possible because the stratigraphic control is not precise in et al. work.
The top of the studied interval seems to correspond to the K88 Sequence ( et al., 2007), which base is marked by abrupt modifications in seismic profiles, also reflected in both environmental and stratigraphical changes.
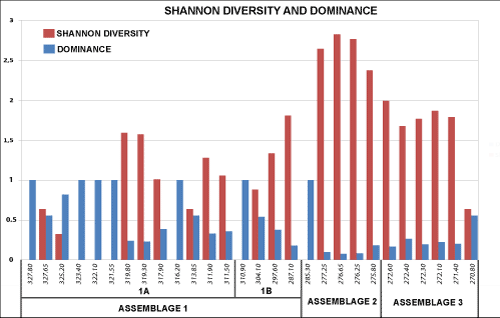
Click on thumbnail to enlarge the image.
Figure 4: Shannon and dominance indexes in the proposed assemblages.
Click on thumbnail to enlarge the image.
Appendix 2: Distribution of the ostracode species, with their abundances.
Paleobiogeographical reconstructions using fossil groups depend on the accuracy of the available taxonomic data from the involved regions. This type of study is still in an early stage in Brazilian basins. This work represents an additional contribution for paleobiogeographic correlations using ostracode faunas. Pioneering works demonstrating the potential of ostracodes in correlation studies were published in the 1960 and 1970 decades (e.g., , 1966a, 1966b, 1976) dealing with faunas from Gabon and NE Brazil. Paleobiogeographic syntheses of the Upper Cretaceous ostracodes were developed by several authors, including (1982), & (1988, 1992), (1988, 1999), (1999c), (2002) and (2003).
et al. (2000) demonstrated that the ostracode fauna of the Potiguar Basin was endemic except for Cytherella austinensis, previously recorded in North America (, 1929; & , 1964) and Africa ( & , 1986), for Cytherella gambiensis, recorded in Senegal (, 1963), and for Veenia glabella recorded in Senegal (, 1963), Gabon (, 1973a) and Algeria (, 1985). At the generic level, et al. (2000) demonstrated a strong affinity between eastern Brazilian basins and West African basins in the Turonian, especially by the presence of the genera Brachycythere and Haughtonileberis.
According to the Turonian ostracode data, the Potiguar Basin and West/North Africa have strong similarity as demonstrated by sharing eight common species
(Fig. 5 ![]() ). Cytherella mediatlasica occurs in the Turonian-Santonian of Nigeria (,
1987, 1992) and Santonian of Morocco (,
1996); both Potiguarella grosdidieri sp. nov.
and Potiguarella coimbrai sp. nov.
occur in the Turonian of Gabon (, 1979); Nigeria cf. rotunda
, sensu , occurs in the Cenomanian of Morocco (,
1991); Paracypris aff. dubertreti was also recorded in the Cenomanian-Turonian of Algeria (,
1985); Paracypris aff. caudata occurs
probably in the Turonian of Egypt (, 1964), Turonian of Gabon (,
1973a) and Middle Turonian-Santonian of Algeria (,
1985); Ovocytheridea reymenti is present in the Cenomanian-Maastrichtian of Nigeria (,
1987, 1992) and Haughtonileberis dinglei sp. nov. was recorded in the Turonian-Santonian of Nigeria (,
1992).
). Cytherella mediatlasica occurs in the Turonian-Santonian of Nigeria (,
1987, 1992) and Santonian of Morocco (,
1996); both Potiguarella grosdidieri sp. nov.
and Potiguarella coimbrai sp. nov.
occur in the Turonian of Gabon (, 1979); Nigeria cf. rotunda
, sensu , occurs in the Cenomanian of Morocco (,
1991); Paracypris aff. dubertreti was also recorded in the Cenomanian-Turonian of Algeria (,
1985); Paracypris aff. caudata occurs
probably in the Turonian of Egypt (, 1964), Turonian of Gabon (,
1973a) and Middle Turonian-Santonian of Algeria (,
1985); Ovocytheridea reymenti is present in the Cenomanian-Maastrichtian of Nigeria (,
1987, 1992) and Haughtonileberis dinglei sp. nov. was recorded in the Turonian-Santonian of Nigeria (,
1992).
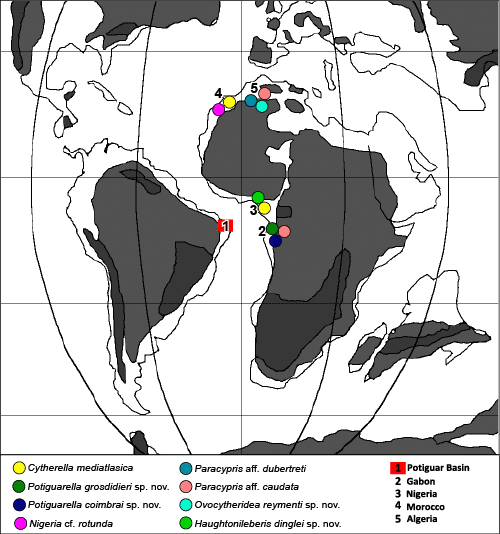
Click on thumbnail to enlarge the image.
Figure 5: Paleogeographic distribution of Turonian species in Potiguar Basin (Turonian map after et al., 1994).
(1993) and et al. (2013) noted the existence of an almost continuous Cenomanian-Turonian carbonate platform in Northwest and North Africa, which acted probably as the migratory route for the species shared with the Potiguar Basin.
The presence of the genera Ovocytheridea, Cophinia, "Brachycythere" and Haughtonileberis is evidence that the Potiguar Basin is in the Afro-Arabian Bioprovince (that includes north-northeastern Africa and the Middle East) proposed by & (1988, 1992). This also reinforces the initial proposal of (1982) of a "Brazil-Central/West Africa Province".
The occurrence, in the Potiguar Basin, of some taxa from South Africa, South and North America and North Africa is related to the bidirectional opening of the Atlantic, which began in the south during the Berrisian-Barremian, followed by Aptian-Albian rifting in the north (, 2000), with the last connection in Northwest Gabon and Northeast Brazil (, 2004). Some authors (, 1982; et al., 1996; , 2000; et al., 2005) suggest that the first marine influences appeared in Equatorial area during Late Aptian, but the open marine circulation between South Atlantic and Central Atlantic would only have been established in Albian times (-, 1987, and references therein; , 2001). The Turonian ostracode data presented in this paper indicate that the migration occurred both directions.
The possible presence of the genus Bicornicythereis , 2009, is particularly interesting. Species of this genus occur in Santonian-Maastrichtian deposits of the United States according to (2009) and they are not present in the Caribbean region ( et al., 2012). (1980) infers that the migration of shallow water marine benthic organisms by passive transport from North Atlantic to South Atlantic occurred until the Cenomanian or even Early Turonian. On the other hand, biogeographic distribution patterns of benthic foraminifers indicate the existence of a "Brazilian-Caribbean-Gulf Coast Province" in the Coniacian-Maastrichtian ( & , 2000).
Some elements from South Gondwana (the Pan-Gondwana Fauna proposed by , 1988), such as the widely distributed genera Brachycythere, Haughtonileberis,and Nigeria, are recorded in this study. Rostrocytheridea, a typical Gondwanine genus (, 1999; & , 2007), probably occurs both in Potiguar and Pará-Maranhăo Basins, Brazilian equatorial margin ( et al., 2009). Similarities with Indian fauna are related to "Brachycythere" aff. jodhpurensis. The species Brachycythere jodhpurensis , 1997, was recorded in the Turonian-Coniacian of India (, 1997; et al., 2007). All these taxa possibly had the same migration pattern as Majungaella and Aracajuia, indicating that the northward migration across the Walvis Ridge-Rio Grande Rise barrier to Northeast Brazilian margin, which began in Late Aptian-Albian, was still effective during the Late Cretaceous (, 1988; et al., 2012; et al., 2013).
This detailed taxonomic study of Turonian ostracodes from the Jandaíra Formation, Potiguar Basin has significantly improved the micropaleontological knowledge of this area. More precisely, 53 taxa were recognized with two new genera and 13 new species; several of the 53 taxa probably are new species, but were only compared with species already described or left in open nomenclature.
We propose three faunal assemblages indicative of different paleoenvironmental contexts, within the marginal marine environment, varying from lagoonal to more open marine (probably subtidal), which supports the potential of ostracodes as paleoecological indicators.
We reinforce the inclusion of this region in the "Brazil-Central/West Africa Province" based on the strong similarity verified by the occurrence of eight common species. The Turonian ostracode data presented in this paper indicate that migration occurred from the South Africa and South America and North America and North Africa, according to the bidirectional opening of the Atlantic Ocean.
We are grateful to Narendra Kumar (UFRN) for his help in the field activities, Ana Cristina (Universidade de Lisboa) for the help in revising the text, Rogério (Petrobras-Cenpes/DIVEX) and Michele (itt Fossil, Unisinos) for the SEM work. The first author thanks Coordenaçăo de Aperfeiçoamento de Pessoal de Nível Superior for the research grant received during the development of this study (CAPES/PDSE, Proc. nº 18919/12-0). Finally, we are greatly indebted to Marta Claudia (Petrobras-Cenpes/DIVEX), without whom this study would not have been possible. Gerson thanks CNPq (The Brazilian Scientific and Technology Developing Council) for the grant (proc. 308544/2012-9). T.M. and J. are thanked for their detailed critical reviews of the manuscript.
C.I. (1929).- Ostracoda of the Cretaceous of North Texas.- University of Texas Bulletin, Austin, nº 2907, 144 p.
C.I. (1933).- Shell structure of the ostracode genus Cytheropteron, and fossil species from the Cretaceous of Texas.- Journal of Paleontology, Ithaca, vol. 7, nº 2, p. 181-214.
B. (1991).- Les ostracodes du Crétacé moyen (Barrémien ŕ Turonien), le long d'une transversale Agadir-Nador (Maroc).- Strata, Toulouse, (Série 2), vol. 14, 765 p.
B. (1992).- Associations d'ostracodes et paléoécologie du Crétacé (Barrémien ŕ Turonien) le long d'une transversale Agadir-Nador (Maroc).- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 99, nº 3-4, p. 291-319.
B. (1993).- Associations d'Ostracodes des marges téthysiennes et atlantiques marocaines de l'Albien au Turonien.- Geobios, Villeurbanne, vol. 26, nº1, p. 69-84.
B. (1995).- Trachyleberididae (Ostracodes) du Turonien supérieur (?)-Santonien de la région de Boulmane, Moyen Atlas (Maroc) : Systématique et biostratigraphie.- Revista Espańola de Micropaleontologia, Madrid, vol. 27, nº 1, p. 85-142.
B. (1996).- Nouvelles espčces d'ostracodes du Turonien supérieur (?)-Coniacien (?)-Santonien de la région de Boulmane, Moyen Atlas, Maroc. Systématique, biostratigraphie et paléoécologie, paléobiogéographie des associations. In: S., I. & J.-P. (eds.), Géologie de l'Afrique et de l'Atlantique Sud.- Actes des Colloques de Géologie d'Angers, 1994, Pau, p. 483-509.
B. (2002).- Cretaceous ostracode biochronology of Morocco.- Eclogae Geologicae Helvetiae, Basel, vol. 95, p. 133-152.
B., J.-P. & J. (2007).- Cretaceous (Albian to Coniacian) Ostracodes from the subsurface of the Jaisalmer Basin, Rajasthan, India.- Micropaleontology, New York, vol. 53, nº 7/8, p. 345-370.
B., V., M.-J., C. & M. (2013).- The upper Cenomanian-lower Turonian carbonate platform of the Preafrican Trough, Morocco: Biostratigraphic, paleoecological and paleobiogeographical distribution of ostracods.- Cretaceous Research, London, vol. 45, p. 216-246.
B. & G. (1996).- Ostracodes et foraminifęres du Crétacé supérieur du synclinal d'El Koubbat, Moyen Atlas, Maroc.- Geobios, Villeurbanne, vol. 29, nº 1, p. 45-71.
L.S., A., D.A. & R.P. (2013).- Taxonomy, biostratigraphy and paleozoogeography of Amphicytherura and , Aracajuia and Dinglecythere et al., n.gen. (Crustacea, Ostracoda).- Marine Micropalentology, Amsterdam, vol. 105, p. 1-17.
V. (1961).- Contribution ŕ l'étude paléontologique (Ostracodes) et stratigraphique des bassins crétacés et tertiaires de l'Afrique Occidental.- Revue de l'Institut Français du Pétrole, Rueil-Malmaison, vol. 16, nº 7-8, p. 779-867.
V. (1963).- Essai de zonation par les Ostracodes dans le Crétacé du bassin du Sénégal.- Revue de l'Institut Français du Pétrole, Rueil-Malmaison, vol. 18, nº 12, p. 1675-1694.
P.T. & F.J. (1994).- Bacia Potiguar.- Boletim de Geocięncias da Petrobras, Rio de Janeiro, vol. 8, nº 1, p. 127-141.
J.-F. & J.-P. (1988).- Palaeobiogeography of Tethyan Cretaceous marine ostracods. In: T., N. & K. (eds.), Evolutionary Biology of Ostracodes. Its Fundaments and Applications.- Elsevier-Kodansha Ltda, Tokyo- Amsterdam, p. 823-839.
J.-F. & J.-P. (1992).- Marine ostracode provincialism in the late Cretaceous of the Tethyan realm and the austral province.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 233, p. 63-95.
S. & R. (2007).- The distribution of the gondwanine ostracod Rostrocytheridea : palaeozoogeographical implications.- Geological Journal, Sussex, vol. 42, p. 113-125.
C., J. & R. (2005).- Phanerozoic geological evolution of the Equatorial Atlantic domain.- Journal of African Earth Sciences, Oxford, vol. 43, p. 275-282.
J.-P. & R. (1969).- Quelques ostracodes nouveaux du Cénomano-Turonien de l'Atlas Saharien Occidentale (Algérie).- Revue de Micropaléontologie, Paris, vol. 12, nº 3, p. 130-144.
H., C., P., J. & P. (1981).- Le Crétacé moyen et supérieur du Djebel Semmama (Tunisie du Centre-Nord).- Bulletin des Centres de Recherches Exploration-Production elf Aquitaine, Pau, vol. 5, p. 193-267.
H., R., F. & R. (1995).- Tunisian Cenomanian ostracodes in a distal environment. In: J. (ed.), Ostracoda and biostratigraphy.- Balkema, Rotterdam, p. 129-139.
W.A. (1964).- Ostracoden aus der Oberkreide von Abu Rawash, Agypten.- Palaeontographica Abt. A, Stuttgart, vol. 123, p. 111-136.
W.A. (1946).- Contribution to the study of Ostracoda with special reference to the Tertiary and Cretaceous microfauna of the Caribbean region.- PhD Thesis, University of Utrecht, J.H. de Bussy, Amsterdam, 167 p.
G., G., A. & H. (1992).- Late Miocene ostracoda from the Ashtart 1 well (Gulf of Gabčs, Tunisia).- Bollettino della Societŕ Paleontologica Italiana, Modena vol, 31, nº 1, p. 3-93.
I.M. (1976).- História Geológica das bacias sedimentares do Nordeste do Brasil.- Anais da Academia Brasileira de Cięncias, Rio de Janeiro, vol. 48, nº 3, p. 463-474.
G.V. (2004).- Diacronismo de eventos no rifte Sul-Atlântico.- Boletim de Geocięncias da Petrobras, Rio de Janeiro, vol. 12, nº 2, p. 203-229.
P., M.C. & D.L. (2013).- Jean-Paul .- Carnets de Géologie [Notebooks on Geology], Brest, Obituary Notice (Jean-Paul_Colin), p. 325-345.
G. & C. (1995).- Ostracodes paléogčnes au débouché de la mer transsaharienne (Mali-Niger).- Documents du Laboratoire de Géologie de Lyon, Lyon, nº 135, p. 1-123.
J.C., A.L., E.A. & B.B. (2007).- Ostracodes (Crustacea) from Cananéia-Iguape estuarine/lagoon system and geographical distribution of the mixohaline assemblages in southern and southeastern Brazil.- Iheringia, Série Zoologia, Porto Alegre, 97 nº 3, p. 273-279.
J.-P., M.A. & J.M. (1982).- Los ostracodos del Cenomaniense Superior y Turoniense de la Cuenca Vasco-Cantábrica.- Revista Espańola de Micropaleontología, Madrid, vol. 14, nº 1-2-3, p. 187-220.
J.-P., Y. & V.A. (1996).- Ostracodes limniques et lagunaires dans le Crétacé supérieur du Mali (Afrique de l'Ouest) : systématique, paléoécologie et affinités paléobiogéographiques.- Revue de Micropaléontologie, Paris, vol. 39, nº 3, p. 211-222.
V.C. (2001).- A evoluçăo da plataforma carbonática Jandaíra durante o Neocretáceo na Bacia Potiguar: análise paleoambiental, diagenética e estratigráfica.- Tese de Doutorado, Instituto de Geocięncias e Cięncias Exatas, UNESP, Rio Claro, 239 p.
T.M. (1988).- Ostracods and sea-level changes; case studies from the Quaternary of North and South Carolina, U.S. Atlantic Coast Plain. In: P., J.-P. & J.-P. (eds.), Ostracoda in the Earth Sciences.- Elsevier Science Publishers, Amsterdam, p. 77-88.
R. & P. (1972).- Contribution ŕ la connaissance des Ostracodes crétacés du Liban.- Revue de Micropaléontologie, Paris, vol. 4, nº 3, p. 273-296.
M.P., J.C. & A.L. (2000).- Cretaceous marine Ostracoda from the Potiguar Basin, northeastern Brazil.- Neues Jahrbuch für Geologie und Palaöntologie, Abhandlungen, Sttutgart, vol. 215, nº 3, p. 321-345.
G. (1966).- Cytheracea (Ostracodes) du Maastrichtien de Maastricht (Pays-Bas) et des régions voisines; résultats stratigraphiques et paléontologiques de leur étude.- Mededelingen van de Geologische Stichting, (Serie C), vol. 2, nº 2, 197 p.
D. (1987).- A Bacia de Campos no Mesocretáceo: uma contribuiçăo ŕ paleoceanografia do Atântico Sul primitivo.- Revista Brasileira de Geocięncias, Săo Paulo, vol. 17, nº 2, p. 162-167.
R.V. (1969).- Marine Neocomian Ostracoda from South Africa.- Transactions of the Royal Society of South Africa, Cape Town, vol. 38, nº 2, p. 139-163.
R.V. (1980).- Marine Santonian and Campanian ostracods from a borehole at Richards Bay, Zululand.- Annals of the South Africa Museum, Cape Town, vol. 82, nº 1, p. 1-70.
R.V. (1988).- Marine ostracod distributions during the early breakup of Southern Gondwanaland. In: T., N. & K. (eds.), Evolutionary Biology of Ostracodes. Its Fundaments and Applications.- Elsevier-Kodansha Ltda, Tokyo- Amsterdam, p. 841-854.
R.V. (1999).- Walvis Ridge barrier: its influence on palaeoenvironments and source rock generation deduced from ostracod distributions in the early South Atlantic Ocean. In: N.R., R.H. & V.S. (eds.), The oil and gas habitats of the South Atlantic.- Geological Society, London, Special Publication, nº 153, p. 479-492.
R.V. (2009).- Implications for high latitude gondwanide palaeozoogeographical studies of some New Zealand and Antarctic Peninsula.- Revista Espańola de Micropaleontología, Madrid, vol. 41, nº 1/2, p. 145-196.
D.A., J.P., P.H.P., R.P., M.L.C. & C.M. (2012).- Reassessment of the genus Sergipella , 1967 (Ostracoda, Trachyleberididae), uppermost Aptian-Albian of Brazil and West Africa: Taxonomy and paleogeographic distribution.- Revue de Micropaléontologie, Paris, vol. 55, nº 1, p. 3-15.
V. (1989).- Upper Cretaceous Brachycythere (Ostr., Crust.) from Iran.- Journal of African Earth Sciences, Oxford, vol. 9, nº 3/4, p. 609-616.
G., J.-P., E.A. & P. (2005).- Cretaceous/Tertiary boundary ostracodes from the Poty Quarry, Pernambuco, Northeastern Brazil.- Journal of South American Earth Sciences, Oxford, vol. 19, p. 285-305.
G. (2002).- Paleobiogeography of the Upper Cretaceous to Lower Tertiary marine ostracods from the Atlantic Ocean.- Revista Brasileira de Paleontologia, Rio de Janeiro, vol. 4, p. 65-71.
E. & R.F. (1979).- Zoogeography of Holocene Cytheracean ostracodes in the bays of Texas.- Journal of Paleontology, Ithaca, vol. 53, p. 841-919.
H. (1999a).- Cenomanian to Coniacian ostracodes from the Nkalagu area (SE Nigeria): biostratigraphy and palaeoecology.- Paläontologische Zeitschrift, Sttutgart, vol. 73, nº 1/2, p. 77-98.
H. (1999b).- Occurrence and palaeoecology of Cenomanian to Turonian ostracods from Ashaka (NE Nigeria).- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Sttutgart, vol. 211, nº 1/2, p. 133-149.
H. (1999c).- Cenomanian to Coniacian biogeography and migration of North and West African ostracods.- Cretaceous Research, London, vol. 20, p. 215-229.
H. & I. (2008).- Cenomanian ostracods of the Tarfaya upwelling region (Morocco) as palaeoenvironmental indicators.- Revue de Micropaléontologie, Paris, vol. 51, nº 4, p. 273-286.
N. (1951).- Quelques ostracodes nouveaux du Sénonien supérieur du Cameroun.- Revue de l'Institut Français du Petrole et Annales des Combustibles liquides, Paris, vol. 6, p. 53-59.
E. (1979).- Principaux Ostracodes marins de l'intervalle Aptien-Turonien du Gabon (Afrique Occidentale).- Bulletin des Centres de Recherches Exploration-Production elf Aquitaine, Pau, vol. 3, nº 1, p. 1-35.
M., M.I., M. & W. (2013).- Ostracoda (Crustacea) and their palaeoenvironmental implications for the Solimőes Formation (Late Miocene, Western Amazonia, Brazil).- Journal of South American Earth Science, Oxford, vol. 42, p. 216-241.
Ř. & D.A.T. (2006).- Paleontological data analysis.- Blackwell Publishing, Oxford, 351 p.
Ø., D.A.T. & P.D. (2001).- PAST: Paleontological Statistics Software Package for Education and Data Analysis.- Palaeontologia Electronica, Columbia, vol. 4, nº 1, p. 1-9.
J.E. (1975).- Pattern of marine ostracode Diversity in the Cape Hatteras, North Carolina Area.- Journal of Paleontology, Ithaca, vol. 49, nº 4, p. 731-744.
D.J. (2005).- Ostracoda. In: R.C., R.M. & I.R. (eds.), Encyclopedia of Geology.- Elsevier, Oxford, p. 453-463.
M.C. (1929).- Upper Cretaceous Ostracoda of Arkansas.- Arkansas Geological Survey Bulletin, Little Rock, nº 2, p. 1-29.
D. (1977).- Ecology and zoogeography of recent brackish-water ostracoda (Crustacea) from south-west Florida. In: H. & D. (eds.), Aspects of Ecology and Zoogeography of Recent and Fossil Ostracoda.- Dr W. Junk b.v. Publishers, The Hague, p. 207-221.
C.A. & K. (1986).- Micropaleontology and biostratigraphy of the Coastal basins of West Africa.- Journal of African Earth Sciences, Oxford, vol. 5, nº 1. p. 1-100.
E.A.M. (1982).- Geohistória e paleoecologia das bacias marginais de Florianópolis e Santos.- Anais do XXXI Congresso Brasileiro de Geologia, Salvador, nº 5, p. 2369-2382.
E.A.M., N., N.C.A. & A.R. (1993).- Upper Aptian-lower Coniacian carbonate sequences in the Sergipe Basin, northeastern Brazil. In: J.A, R.W. & J.-P. (eds.), Cretaceous Carbonate Platforms.- The American Association of Petroleum Geologists (AAPG), Tulsa, Oklahoma, vol. 56, p. 127-144.
E.A.M. & I. (2000).- Late Cretaceous foraminiferal biogeography (Families Bolivinidae, Buliminellidae, Gavelinellidae, Siphogenerinoididae, Turrilinidae) in northeastern Brazilian shelf and central West African basins.- Cretaceous Research, London, vol. 21, p. 381-405.
K. (1964).- Brachycythere (Brachycythere) sapucariensis n.sp. aus dem Turonium.- Senckenbergiana lethaea, Frankfurt, vol. 45, p. 489-495.
K. (1966a).- On ''Gondwana Wealden'' Ostracoda from NE Brazil and West Africa. In: J.E. (ed.), Proceedings of the 2nd West African Micropaleontological Colloquium.- E.J. Brill, Leiden, p. 113-118.
K. (1966b).- Preliminary remarks on some marine Cretaceous ostracodes from Northeastern Brazil and West Africa. Proceedings of the 2nd West African Micropalaeontological Colloquium.- E.J. Brill, Leiden, p. 119-123.
K. (1967).- Ostracoden aus der marinen ''Küsten-Kreide Brasiliens'': 2. Sergipella transatlantica n.gen. n.sp. und Aracajuia benderi n.gen. n.sp., aus dem Ober-Aptium/Albium.- Senckenbergiana Lethaea, Frankfurt, vol. 48, p. 525-533.
K. (1976).- Remarks on marine Cretaceous ostracods of gondwanic distribution.- Proceedings of the 5nd West African Micropaleontological Colloquium, Addis Abada, p. 539-551.
P. (2003).- Paleobiogeography of late Early Cretaceous to Early Paleocene marine Ostracoda in Arabia and North to Equatorial Africa.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 196, nº 3-4, p. 319-342.
J.G. (2000).- Continental break up and the distribution of fishes of Western Gondwana during the Early Cretaceous.- Cretaceous Research, London, vol. 21, p. 281-314.
S. (1989).- Mid-Cretaceous Ostracoda of northeastern Algeria.- Fossils and Strata, Oslo, vol. 27, p. 1-67.
J., G.P. & P.D. (coords., 1996).- Proceedings of the Ocean Drilling Program, Initial Reports, College Station, vol. 159, 616 p.
A.S. (2008).- O que ganhamos "confundindo" riqueza de espécies e equitabilidade em um índice de diversidade?- Biota Neotropica, Campinas, vol. 8, nº 3, p. 21-27.
W.U. (2003).- Bacias Sedimentares da Margem Continental Brasileira. In: L.A., C., R.M. & J.H. (eds.), Geologia, Tectônica e Recursos Minerais do Brasil.- CPRM, Brasília, p. 87-94.
M.C. & R.T. (1988).- Planície de maré no poço 9-MO-13-RN, Formaçăo Jandaíra - um exemplo do passado.- Anais do XXXV Congresso Brasileiro de Geologia, Belém, nº 2, p. 809-823.
A.M. & J.E. (2010).- Biostratigraphy, palaeoecology and palaeogeography of the Middle Cenomanian-Early Turonian Levant Platform in Central Jordan based on ostracods. In: C. & M. (eds.), Evolution of the Levant Margin and Western Arabia Platform since the Mesozoic.- Special Publication, Geological Society, London, vol. 341, p. 187-210.
E.A. (1973).- Charophytas y ostrácodos no marinos del Grupo Neuquén (Cretácico Superior) en algunos afloramientos de las Provincias de Río Negro y Neuquén en la República Argentina.- Revista del Museo de La Plata, (Paleontología), vol. 8, p. 1-33.
J.W. (1988).- Ostracods and paleosalinity reconstruction. In: P., J.-P. & J.-P. (eds.), Ostracoda in the Earth Sciences.- Elsevier Science Publishers, Amsterdam, p. 125-155.
E.M.H. (1973a).- Upper Cretaceous-Palaeogene Ostracoda from the South Atlantic.- Publications from the Palaeontological Institution of the University of Uppsala, 205 p.
E.M.H. (1973b).- Ostracoda from the Eze-Aku shale (Turonian, Cretaceous) Nkalagu, Nigeria.- Bulletin of the Geological Institutions of the University of Uppsala, vol. 4, p. 43-52.
E.M.H. (1979).- Upper Cretaceous-Paleogene marine ostracods from the Sergipe-Alagoas Basin, northeastern Brazil.- Bulletin of the Geological Institutions of the University of Uppsala, vol. 8, p. 135-172.
D.D. & J.C. (2008).- Perissocytheridea carrenoae sp. nov. (Crustacea, Ostracoda) and associated calcareous microfauna from Yecua Formation (Miocene), Bolivia.- Revista Brasileira de Paleontologia, Rio de Janeiro, vol. 1, nº 3, p. 179-186.
E.A. (1987).- Ostracod biostratigraphy of the Eastern Dahomey Basin, Niger Delta and the Benue Trough of Nigeria.- Geological Survey of Nigeria, Lagos, vol. 41, 151 p.
E.A. (1992).- Cretaceous ostracod biostratigraphy from Chad Basin in Nigeria.- Journal of African Earth Sciences, Oxford, vol. 14, p. 327-339.
O.C., U.M., J.G.F., E.H., C.P. & C.A.V. (2007).- Bacia Potiguar.- Boletim de Geocięncias da Petrobras, Rio de Janeiro, vol. 15, nº 2, p. 357-387.
E.K, S. & G. (2012).- Cretaceous palaeogeography of southern Gondwana from the distribution of the marine ostracod Majungaella : New data and review.- Cretaceous Research, London, vol. 37, p. 127-147.
E.K., C.T. & G. (2009).- Cretaceous ostracodes from Pará-Maranhăo Basin, Brazil: taxonomy and preliminary paleoecological and paleobiogeographical inferences.- Revue de Paléobiologie, Genčve, vol. 28, nº 2, p. 437-456.
E.K., C.T. & G. (2010).- New ostracode species from the Upper Cretaceous of the Santos Basin, Brazil.- Revista Brasileira de Paleontologia, Rio de Janeiro, vol. 13, nº 3, p. 175-180.
T., J., A.E.L., W., M., F.E., E. & T. (2001).- Cretaceous separation of Africa and South America: the view from the West African margin (ODP Leg 159).- Journal of South American Earth Sciences, Oxford, vol. 14, p. 147-174.
V. (1971).- The diversity of fossil ostracode communities as an indicator of paleogeopraphic conditions.- Bulletin des Centres de Recherches Exploration-Production elf Aquitaine, Pau, vol. 5 (suppl.), p. 45-61.
T.M. (1996).- Ecologic Atlas of Upper Cretaceous Ostracoda.- Geological Survey of Alabama Monograph,, Tuscaloosa, nº 14, 176 p.
T.M. (2002).- Systematics and paleobiogeography of brachycytherine Ostracoda.- Micropaleontology, New York, vol. 48, (suppl. 2), 87 p.
T.M. (2009).- On the global distribution of Late Cretaceous Ostracodes: the Genus Bicornicythereis (n.gen.), with notes on Curfsina.- Micropaleontology, New York, vol. 55, nº. 4, p. 345-364.
T.M., J.-P. & S. (2012).- New species and genera of Ostracoda from the Maastrichtian (Late Cretaceous) of Jamaica.- Micropaleontology, New York, vol. 58, nº 5, p. 397-455.
R.A. (1960).- Studies on Nigerian Upper Cretaceous and Lower Tertiary Ostracoda: part 1. Senonian and Maastrichtian Ostracoda.- Stockholm Contributions in Geology, Stockholm, vol. 7, p. 1-238.
R.A. (1963).- Studies on Nigerian Upper Cretaceous and Lower Tertiary Ostracoda, part 2: Danian, Paleocene and Eocene Ostracoda.- Stockholm Contributions in Geology, Stockholm, vol. 10, p. 1-286.
R.A. (1978).- Quantitative biostratigraphical analysis exemplified by Moroccan Cretaceous ostracods.- Micropaleontology, New York, vol. 24, nº 1, p. 24-43.
R.A. (1980).- Paleo-oceanology and paleobiogeography of the Cretaceous South Atlantic Ocean.- Oceanologica Acta, Paris, vol. 3, nº 1, p. 127-134.
A. & M. (1974).- Cenomanian-Turonian ostracods from the Judea Group in Israel.- Bulletin of the Geological Survey of Israel, Jerusalem, vol. 62, 64 p.
J.E. & R.F. (1983).- Recurrent species associations and species diversity of cytheracean ostracodes in the upper Austin and lower Taylor groups (Campanian, Upper Cretaceous) of Travis County, Texas.- Transactions of the Gulf Coast Association of Geological Societies, Houston, vol. 33, p. 397-406.
P. (1997).- Ostracoda from the subsurface Cretaceous strata of Manhera Tibba well-I and Shahgarh well-B, Jaisalmer Basin, Rajasthan, India with special remarks on foraminiferids.- Geoscience Journal, Lucknow, vol. 18, nº 1, p. 1-57.
A.G., D.G. & B.M. (1994).- Atlas of Mesozoic and Cenozoic coastlines.- Cambrige University Press, Cambridge, 99 p.
F.M. & P.M. (1964).- Cretaceous ostracoda from wells in the southeastern United States.- Bulletin of North Carolina Department of Conservation and Development, North Carolina, vol. 78, p. 1-55.
Y. (1982).- Les Ostracodes et l'histoire géologique de l'Atlantique Sud au Crétacé.- Bulletin des Centres de Recherches Exploration-Production elf Aquitaine, Pau, vol. 6, p. 1-37.
M.A. & E.A. (1978).- Microfósiles calcáreos no marinos del Cretácico superior en Zampal, provincia de Mendoza, Argentina.- Ameghiniana, Buenos Aires, vol. 15, p. 111-135.
J.L. (1985).- Les Ostracodes du Crétacé Supérieur (Vraconien ŕ Campanien basal) de la région de Tébessa (Algérie du Nord-Est): Stratigraphie, Paléoécologie, Systématique.- Ph.D thesis, Université Pierre et Marie Curie, Paris, 261 p.
M.C. (1995).- Ostracodes marinhos na Bacia Potiguar: aplicaçăo nos estudos bioestratigráficos do Cretáceo.- Anais da Academia Brasileira de Cięncias, Rio de Janeiro, vol. 67, p. 392-393.
M.C., E.A.M., Jr A.C. & P. (2000).- Stratigraphy and biogeographic affinities of the late Aptian-Campanian ostracods of the Potiguar and Sergipe Basins in northeastern Brazil.- Cretaceous Research, London, vol 21, p. 407-455.