◄ Carnets Geol. 17 (7) ►
![]()
Contents
[1. Introduction]
[2. Geological background]
[3. Material and methods]
[4. Results]
[5. Discussion]
[6. conclusions] and ...
[Bibliographic references]
Department of Geology, University of Tartu, Ravila 14A, 50411 Tartu (Estonia)
Institut für Geologie, Universität Hamburg, Bundesstr. 55, 20146 Hamburg (Germany)
Institute of Geology, Tallinn University of Technology, Ehitajate tee 5, Tallinn, 19086 (Estonia)
Published online in final form (pdf) on October 16,
2017
DOI 10.4267/2042/62664
[Editor: Bruno Granier; language editor: Robert W. Scott]
![]()
Two relatively large specimens of the rugosan Lambelasma sp. are fully intergrown with the bryozoan Stigmatella massalis colony. The intergrown specimen occurs in the Oandu Regional Stage (lower Katian) of Estonia and constitutes the earliest record of bryozoan-rugosan intergrowth from Baltica. Most likely this symbiotic association was accidental. Rugosans presumably benefitted from the bryozoan, which served as an anchor to stabilize them in hydrodynamically active waters. The lack of malformations and no decrease in the size of bryozoan zooids near the rugosans indicate a lack of negative effect of the rugosans on the bryozoan. Bryozoan-rugosan symbiosis is only known from the Ordovician of Baltica and Laurentia.
symbiosis;
bioclaustrations;
bryozoans;
corals;
Katian;
Baltica.
Vinn O., Ernst A. & Toom U. (2017).- Rare rugosan-bryozoan intergrowth from the Upper Ordovician of Estonia.- Carnets Geol., Madrid, vol. 17, no. 7, p. 145-151.
Rare intercroissance tétracoralliaire-bryozoaire dans l'Ordovicien supérieur d'Estonie.- Deux spécimens relativement grands du tétracoralliaire Lambelasma sp. montrent une intercroissance étroite avec une colonie du bryozoaire Stigmatella massalis. Le matériel a été récolté dans l'Étage Régional Oandu (Katien inférieur) d'Estonie et constitue la plus ancienne mention de intercroissance tétracoralliaire-bryozoaire dans l'Ordovicien de Baltica. Très vraisemblablement cette symbiose est accidentelle. Le tétracoralliaire tire probablement bénéfice du bryozoaire dont il se sert comme point d'ancrage pour se stabiliser dans un milieu marin agité. L'absence de malformations et de diminution de la taille des zoécies du bryozoaire au contact du tétracoralliaire indiquent que ce dernier ne perturbe pas le développement du bryozoaire. La symbiose tétracoralliaire-bryozoaire n'est connue que dans l'Ordovicien de Baltica et de Laurentia.
symbiose ;
bioclaustrations ;
bryozoaires ;
madréporaires ;
Katien ;
Baltica.
Among the best examples of symbiosis are endobionts embedded (i.e., bioimmured) in the living tissues of host organisms (Taylor, 1990).There are cases of Paleozoic rugose corals being embedded in skeletons of tabulate corals, bryozoans, crinoids, and stromatoporoids (Elias, 1982; McAuley & Elias, 1990; Sorauf & Kissling, 2012; Lee et al., 2016; Vinn et al., 2016). Bioimmured rugosans differ from organisms which formed bioclaustrations (Palmer & Wilson, 1988) by having their own skeleton, rather than being soft bodied. Skeletonized invertebrates that live in close proximity will often grow into one another to form a fused pair of skeletons, which differs from simple encrustation where one skeleton overlaps another (Tapanila, 2008). The full intergrowth of two invertebrates provides the most continuous evidence of symbiosis (Tapanila, 2008). Intergrown fossils are especially common in muddy settings or reef environments where competition for hard substrate settling is high (Tapanila, 2008).
The faunas of bryozoans and rugose corals in the Ordovician of Estonia are relatively well described (Bassler, 1911; Modzalevskaya, 1953; Reiman, 1958; Kaljo, 1958, 1961; Männil, 1959; Neuman, 1969, 1986; Lavrentjeva, 1990; Pushkin & Gataulina, 1992; Gorjunova, 1992, 1996; Gorjunova & Lavrentjeva, 1993), but only a single example of intergrowth between rugosans and bryozoans has been previously reported (Vinn et al., 2016) and this topic needs further study. Modern bryozoans also encrust living substrates, for example, live crabs. In this association, the negative impact on the crab is minimal, the relationship is more beneficial to the bryozoans, and there is no special symbiotic relationship between the crab and the bryozoans (Key et al., 1999).
The aims of this paper are to: 1) describe the earliest known symbiotic association of rugosans and bryozoans from Baltica, here reported from the lower Katian of Estonia, and 2) discuss the paleoecology of this rugosan-bryozoan association.
The Ordovician limestones of Estonia are well exposed and form a broad belt from the Narva River in the northeast to Hiiumaa Island in the west (Nestor & Einasto, 1997). The total thickness of the Ordovician System in Estonia is 70 to 180 m (Nestor & Einasto, 1997). In the early Late Ordovician, the western part of the East-European Platform was covered by a shallow, epicontinental sea. This basin had little bathymetric differentiation and was characterized by an extremely low sedimentation rate (Nestor & Einasto, 1997). Along the entire extent of the basin ramp a series of grey calcareous - argillaceous sediments accumulated. The sedimentation trend was to decrease bioclasts and increase clay in the offshore direction (Nestor & Einasto, 1997). During the Ordovician the Baltica paleocontinent moved from the temperate climatic zone to the subtropics (Nestor & Einasto, 1997; Cocks & Torsvik, 2005; Torsvik et al., 2013). The Late Ordovician climatic change resulted in an increase in carbonate production and sedimentation rate on the carbonate ramp. During the Late Ordovician the first carbonate buildups appeared, marking a striking change in the overall character of the paleobasin (Nestor & Einasto, 1997).
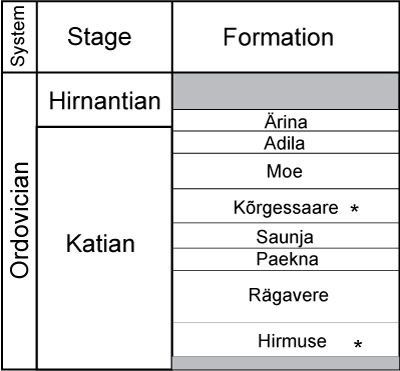
The Hirmuse Formation of the Oandu Regional Stage (lower Katian) consists of a succession of argillaceous limestones and marls of normal marine origin. Sedimentation took place on a relatively shallow water shelf with a strong influx of clay material. The marls of the Hirmuse Formation are very rich in fossils; especially common are brachiopods (most abundant is Sowerbyella tenera) and bryozoans (Stigmatella massalis, Dittopora colliculata, Homotrypa similis). The distribution of the Hirmuse Formation is relatively restricted to northern and central Estonia. The thickness of the formation varies from 0.6 to 10.8 m (Rõõmusoks, 1983).
The collections
of the Institute of Geology, Tallinn University of Technology (GIT), contain
about 500 hemispherical bryozoans from the Hirmuse Formation of the Oandu
Regional Stage (Katian) (Figs. 1 ![]() - 2
- 2 ![]() ). Most hemispherical bryozoans from the
Hirmuse Formation belong to Stigmatella
massalis Bassler, 1911. The bryozoans in the collection were searched
for the presence of intergrowth with the other invertebrates. A single specimen
of Stigmatella massalis contained rugosans. It was photographed using a Nikon D7000 digital camera. The dimensions
of both rugosans and the bryozoan zoarium were obtained from calibrated
photographs. Part of the bryozoan colony was polished perpendicular to the
growth surface in order to study the internal structures.
). Most hemispherical bryozoans from the
Hirmuse Formation belong to Stigmatella
massalis Bassler, 1911. The bryozoans in the collection were searched
for the presence of intergrowth with the other invertebrates. A single specimen
of Stigmatella massalis contained rugosans. It was photographed using a Nikon D7000 digital camera. The dimensions
of both rugosans and the bryozoan zoarium were obtained from calibrated
photographs. Part of the bryozoan colony was polished perpendicular to the
growth surface in order to study the internal structures.
|
Figure 1:
Locality map. Kullaaru ditch marked with square. |
|
Figure 2:
Stratigraphy of the Katian of Estonia. Occurrences of bryozoan-rugosan
intergrowth marked with asterisks. |
A single
specimen from among of hundreds of Stigmatella
massalis specimens shows intergrowth with rugosans (Fig.
3 ![]() ). Two
relatively large rugosans are partially embedded within the bryozoan colony. The
diameter of the bryozoan colony is 23 mm. Maximal diameters of the rugosans are
15 mm and 18 mm. The proximal parts of the rugosans are deeply embedded within
the bryozoan while the apertural parts are partially covered by thin bryozoan
encrustation. The calyxes of the rugosans are devoid of bryozoan encrustation.
One rugosan is oriented vertically in the center of the bryozoan colony, while
the other is strongly tilted. The bryozoan colony shows no malformations in
contact with the rugosans. Bryozoan zooids around the rugosans are the same size
as those in other regions of the colonies.
). Two
relatively large rugosans are partially embedded within the bryozoan colony. The
diameter of the bryozoan colony is 23 mm. Maximal diameters of the rugosans are
15 mm and 18 mm. The proximal parts of the rugosans are deeply embedded within
the bryozoan while the apertural parts are partially covered by thin bryozoan
encrustation. The calyxes of the rugosans are devoid of bryozoan encrustation.
One rugosan is oriented vertically in the center of the bryozoan colony, while
the other is strongly tilted. The bryozoan colony shows no malformations in
contact with the rugosans. Bryozoan zooids around the rugosans are the same size
as those in other regions of the colonies.
|
Figure 3:
1-2,
Bryozoan Stigmatella massalis Bassler,
1911, intergrowth with rugose corals Lambelasma sp. from Kullaaru ditch northern
Estonia, Hirmuse
Formation, Oandu Regional Stage (lower Katian) (GIT 406-1272). 3,
Longitudinal section of Stigmatella
massalis Bassler, 1911, from Kullaaru ditch northern Estonia, Hirmuse
Formation, Oandu Regional Stage (lower Katian) (GIT 406-1272). |
Among
the known species from the Oandu Regional Stage (lower Katian), the studied
bryozoan matches best with Stigmatella massalis Bassler,
1911. However there are a few
reservations: the colony is encrusting not hemispheric as in Bassler's
type material, but the latter actually consist of multilayered crusts. The
longitudinal section shows abundant diaphragms, which are less abundant on the
images by Bassler. However, the number of diaphragms can be variable. In
other characters the studied bryozoan matches very well with S.
massalis. The available material has
been studied externally and on a polished surface (Fig. 3.3 ![]() ), whereas the
preparation of thin sections is a necessary method for study of Palaeostomata.
The latter was not possible because the material is a part of a museum
collection.
), whereas the
preparation of thin sections is a necessary method for study of Palaeostomata.
The latter was not possible because the material is a part of a museum
collection.
Among the known species from the Oandu Regional Stage, the studied rugosans match best with species of Lambelasma.
Symbiotic association
This association between two rugosans and a bryozoan colony most likely occurred during the life of these organisms. The syn vivo nature of this association is supported by the perpendicular to sub-perpendicular orientation of the rugosans relative to the growth surface of bryozoan. The embedding of rugosans in the bryozoan is also restricted to areas outside of the coral calyxes. This association is rare. In fact, the lack of rugose coral-bryozoan intergrowth among the rest of the investigated material might have been caused by anti-biofouling agents secreted by the bryozoans.
Presumably, the described symbiotic association does not represent any adaptive strategy of the involved rugosan or bryozoan species. Most likely this association was accidental. This is supported by the partial embedment of rugosans in the bryozoan. In well-established symbiotic associations one would expect rugosans to be more completely embedded in bryozoans, so that only their calyxes were left free. The lack of similarly intergrown specimens among the studied material indicates that this association was rare, which also indicates the accidental nature of this association. It is likely that the rugosan behavior may have been somewhat opportunistic. The corals presumably used bryozoans as a hard substrate that they needed for initial growth. They may also have used the bryozoan as an anchor to stabilize them in hydrodynamically active waters. The bryozoan did not provide the rugosans with a higher tier for feeding. The studied bryozoan colony is not highly domical in shape, so even in its central region the rugosans did not achieve a significantly greater elevation. It is likely that the rugosans initially cemented to dead parts of the young bryozoan colony and then they were later partially overgrown by bryozoan. The walls of the symbiotic rugosans are not thinner or peculiar in any way as compared to non-symbiotic specimens of Lambelasma, indicating that the rugosan symbionts were not anatomically adapted to a life within bryozoans. The lack of malformations and decrease in the size of bryozoan zooids near the rugosans indicate a lack of negative effect of the rugosans on the bryozoan. The two relatively large rugosans occupied some part of the bryozoan colony's feeding surface and may have slightly decreased the feeding efficiency of the whole colony. Feeding competition between Lambelasma sp. and Stigmatella massalis seems unlikely as bryozoans and corals feed on different sources. Corals usually feed on zooplankton, whereas bryozoans take much smaller phytoplankton. Their feeding methods are also different; corals hunt food with nematocysts, and bryozoans are filter feeders.
Other examples of bryozoan coral symbiosis
Several cases of bryozoan-rugosan symbiosis are known from the Ordovician. Endobiotic rugose coral symbionts occur in Upper Ordovician (Katian) bryozoans of Estonia (Vinn et al., 2016). Multiple rugosans (Bodophyllum sp. and Lambelasma sp.) were partially embedded in colonies of the cystoporate bryozoan Ceramopora intercellata Bassler, 1911, leaving only their apertures free on the bryozoan growth surface (Vinn et al., 2016). Thus, Lambelasma sp. also reported in this study may have been prone to symbiosis with bryozoans. Rugosans had a presumably mutualistic relationship with their bryozoan hosts in the Upper Ordovician of Estonia. Symbiosis between rugosans and the bryozoans was most likely facultative (Vinn et al., 2016). Streptelasma divaricans (Nicholson) intergrowths have been reported from Upper Ordovician (Richmondian) bryozoans of the Cincinnati Arch region in the USA. The coral larvae frequently settled on living bryozoan colonies, as evidenced by upward growth of the host around these epizoans (Elias, 1982, p. 23, Pl. 3, fig. 5s). A somewhat similar association also occurs in the Upper Ordovician (Gamachian) of Missouri in the USA where Streptelasma sp. A of McAuley and Elias (1990) had a symbiotic relationship with bryozoans. In the latter association the host bryozoan colony eventually grew around the sides of the epizoic corals (McAuley & Elias, 1990, p. 47, Pl. 9, figs. 8-9; Pl. 10, figs. 15), which is very similar to the situation in the rugosan-bryozoan association from the Oandu Regional Stage of Estonia. As with rugosans, tabulates formed symbiotic associations with bryozoans. Encrusting species of Aulopora and the bryozoan Leioclema formed a symbiotic association in the Lower Devonian of western Tennessee (McKinney et al., 1990) so that Aulopora corallites, except for their calyxes, were entirely embedded by a thin encrustation of Leioclema sp. This association was likely mutualistic with benefits including escape from limited space on the substratum into a higher tier of suspension feeders (McKinney et al., 1990).
Other cases of symbiosis in Ordovician bryozoans
Ordovician bryozoans were common hosts for a variety of symbiotic endobionts. The earliest bryozoan endobionts appeared in the Middle Ordovician of Estonia where worm-like Anoigmaichnus bioclaustrations occur in Mesotrypa bystrowi (Vinn et al., 2014). Bioclaustrations made by colonial hydroids or ascidian tunicates have been reported from bryozoans of the Upper Ordovician of North America (Palmer & Wilson, 1988). In addition to bioclaustrations, skeletonized endobiotic invertebrates such as conulariids are also known in Ordovician bryozoans. Conulariids occur in several bryozoan species in the Sandbian and Katian of Estonia (Vinn & Wilson, 2015). They usually were completely embedded in their bryozoan hosts, leaving only apertures free on the growth surface of host. Each bryozoan may host several conulariid specimens (Männil, 1959). It is likely that conulariids sought additional protection against predators by their embedment within the bryozoan skeleton and a relatively stable growth substrate (Vinn & Wilson, 2015).
Other cases of symbiosis in rugosans
In contrast to the middle Palaeozoic, endobiotic rugose corals are rare in the Ordovician. Tryplasmastromatoporoid intergrowths are common in the Upper Ordovician of China (Lee et al., 2016). Several organisms have been found within coenostea of the stromatoporoid Clathrodictyon, including the branching tetradiid Rhabdotetradium, the solitary rugose corals Streptelasma and Tryplasma, the tabulate coral Heliolites and a phaceloid or branching tubular organism identified as the tabulate Bajgolia (Lee et al., 2016). Similarly, the stromatoporoid Ecclimadictyon hosted the rugosans Tryplasma and Streptelasma in the Upper Ordovician of China (Lee et al., 2016). Ordovician rugosans were not only symbionts, but they were also hosts for other invertebrates. The rugosan Grewingkia from the Upper Ordovician of Manitoba hosted worm-like invertebrates, which made Chaetosalpinx ferganensis Sokolov bioclaustrations (Elias, 1986). Pickett (2016) described Carboniferous rugose corals embedded in syringoporid tabulates, where the rugose coral symbionts probably used sweeper tentacles to clear areas of host polyps. The situation in this study of bryozoan-rugosan intergrowth is different, demonstrating that Palaeozoic rugosans used different strategies as symbionts.
Financial support to O.V. was provided by Estonian Research Council project IUT20-34. We are grateful to Dimitri Kaljo, Institute of Geology, Tallinn University of Technology, for identifications of the rugosans. We are also grateful to Gennadi Baranov, Institute of Geology, Tallinn University of Technology, for digital photography of the specimens. We are also grateful to Carlton E. Brett and Owen A. Dixon for the constructive reviews. This paper is a contribution to project IGCP 653 "The onset of the Great Ordovician Biodiversity Event".
Bassler R.S. (1911).- The early Paleozoic Bryozoa of the Baltic provinces.- Bulletin of the United States National Museum, Washington D.C., vol. 77, p. 1382.
Cocks L.R.M. & Torsvik T.H. (2005).- Baltica from the late Precambrian to mid-Palaeozoic times: The gain and loss of a terrane's identity.- Earth-Science Reviews, vol. 72, p. 3966.
Elias R.J. (1982).- Latest Ordovician solitary rugose corals of eastern North America.- Bulletins of American Paleontology, New York, vol. 81, p. 1116.
Elias R.J. (1986).- Symbiotic relationships between worms and solitary rugose corals in the Late Ordovician.- Paleobiology, New York, vol. 12, p. 3245.
Gorjunova R.V. (1992).- Morphology and system of Palaezoic bryozoans.- Trudy Paleontologicheskogo Instituta, Moscow, vol. 251, p. 1152 [in Russian].
Gorjunova R.V. (1996).- Phylogeny of the Paleozoic bryozoans.- Trudy Paleontologicheskogo Instituta, Moscow, vol. 267, p. 1163 [in Russian].
Gorjunova R.V. & Lavrentjeva V.D. (1993).- Morphology and system of the cryptostome bryozoans.- Trudy Paleontologicheskogo Instituta, Moscow, vol. 257, p. 1151 [in Russian].
Kaljo D.L. (1958).- Some new and little-known Baltic tetracorals.- Trudy Institut Geologii AN ESSR, Tallinn, vol. 3, p. 101123 [in Russian].
Kaljo D.L. (1961).- Some additional data on the study of Ordovician streptelasmids in Estonia.- Trudy Institut Geologii AN ESSR, Tallinn, vol. 6, p. 5167 [in Russian].
Key M.M., Winston J.E., Volpe J.W., Jeffries W.B. & Voris H.K. (1999).- Bryozoan fouling of the blue crab Callinectes sapidus at Beaufort, North Carolina.- Bulletin of Marine Science, vol. 64, p. 513-533.
Lavrentjeva V.D. (1990).- New genus of cryptostomate bryozoans.- Paleontologicheski Zhurnal, Moscow, vol. 2, p. 127130 [in Russian].
Lee M., Elias R.J., Choh S.-J. & Lee D.-J. (2016).- Insight from early coralstromatoporoid intergrowth, Late Ordovician of China.- Palæogeography, Palæoclimatology, Palæoecology, vol. 463, p. 192204.
Männil R. (1959).- Problems of Stratigraphy and Bryozoans from the Ordovician of Estonia (Candidate of Sciences Thesis) Institute of Geology.- Academy of Sciences ESSR, Tallinn, p. 542 [in Russian].
McAuley R.J. & Elias R.J. (1990).- Latest Ordovician to earliest Silurian solitary rugose corals of the east-Central United States.- Bulletins of American Paleontology, Washington D.C., vol. 98, p. 182.
McKinney F.K., Broadhead T.W. & Gibson M.A. (1990).- Coral-bryozoan mutualism: Structural innovation and greater resource exploitation.- Science, Washington D.C., vol. 248, p. 466468.
Modzalevskaya E.A. (1953).- Ordovician Treptostomata from Baltics and their significance in stratigraphy.- Trudy VNIGRI, Stratigrafiya i Fauna Ordovika i Silura Zapada Russkoy Platformy, St. Petersburg, vol. 78, p. 91167 [in Russian].
Nestor H. & Einasto R. (1997).- Ordovician and Silurian carbonate sedimentation basin. In: Raukas A. & Teedumäe A. (eds.), Geology and mineral resources of Estonia.- Estonian Academy Publishers, Tallinn, p. 192204.
Neuman B. (1969).- Upper Ordovician streptelasmatid corals from Scandinavia.- University of Uppsala, Geological Institutions Bulletin (new series), vol. 1, p. 173.
Neuman B. (1986).- Rugose corals from the Upper Ordovician erratic boulders of Öland.- GFF, Stockholm, vol. 108, p. 349365.
Palmer T.J. & Wilson M.A. (1988).- Parasitism of Ordovician bryozoans and the origin of pseudoborings.- Palaeontology, London, vol. 31, p. 939949.
Pickett J.W. (2016).- Settlement strategy in Symplectophyllum (Cnidaria, Rugosa).- Geologica Belgica, Brussels, vol. 19, p. 4356.
Pushkin V.I. & Gataulina, G.M. (1992).- Revision of the early Palaeozoic Bryozoa (Cystoporida) from the collection of E.I. Eichwald.- Voprosy Paleontologii, St. Petersburg, vol. 10, p. 8897.
Reiman V.M. (1958).- Neue Tetrakorallen des baltischen Oberordoviziums und des Llandovery.- Trudy Institut Geologii AN ESSR, Tallinn, vol. 2, p. 3347 [in Russian].
Rõõmusoks A. (1983).- Eesti aluspõhja geoloogia.- Valgus, Tallinn, 223 p.
Sorauf J.E. & Kissling D.L. (2012).- Rugosans immured in Silurian Paleofavosites; Brassfield Formation (Llandovery) of Ohio.- Geologica Belgica, Brussels, vol. 15, p. 220225.
Tapanila L. (2008).- Direct evidence of ancient symbiosis using trace fossils. In: Kelley P.H. & Bambach R.K. (eds.), From evolution to geobiology: Research questions driving paleontology at the start of a new century.- Paleontological Society Short Course, October 4; Paleontological Society Papers, Lawrence, vol. 14, p. 271287.
Taylor P.D. (1990).- Preservation of soft-bodied and other organisms by bioimmuration - a review.- Palaeontology, vol. 33, p. 117.
Torsvik T.H., Cocks L.R.M. & Harper D.A.T. (2013).- New global palaeogeographical reconstructions for the Early Palaeozoic and their generation. In: Servais T. (ed.), Early Palaeozoic biogeography and palaeogeography.- Geological Society Memoirs, London, vol. 38, p. 524.
Vinn O., Ernst A. & Toom U. (2016).- Earliest symbiotic rugosans in cystoporate bryozoan Ceramopora intercellata Bassler, 1911 from Late Ordovician of Estonia (Baltica).- Palæogeography, Palæoclimatology, Palæoecology, vol. 461, p. 140144.
Vinn O. & Wilson M.A. (2015).- Symbiotic interactions in the Ordovician of Baltica.- Palæogeography, Palæoclimatology, Palæoecology, vol. 436, p. 5863.
Vinn O., Wilson M.A., Mõtus M.-A. & Toom U. (2014).- The earliest bryozoan parasite: Middle Ordovician (Darriwilian) of Osmussaar Island, Estonia.- Palæogeography, Palæoclimatology, Palæoecology, vol. 414, p. 129-132.