◄ Carnets Geol. 17 (11) ►
![]()
Contents
[Introduction]
[Geological setting]
[Material and methods]
[Mecaster from the Sergipe-Alagoas Basin]
[Systematic palaeontology]
[Discussion and conclusions] and ...
[Bibliographic references]
Universidade Federal de Sergipe, Departamento de Biociências, Campus Prof. Alberto
Carvalho, Av. Olimpio Campos s/nº, Itabaiana, 49500-000, Sergipe
(Brazil)
Fundação Paleontológica Phoenix, Rua Geraldo Menezes de Carvalho, 218, Suissa,
49050-360, Aracaju, Sergipe (Brazil)
Published online in final form (pdf) on December 24,
2017
DOI 10.4267/2042/64287
[Editor: Bruno Granier; language editor:
Stephen Carey]
![]()
The spatangoid echinoid Mecaster texanus (Roemer) was first described from the Austin Chalk in Texas (USA). In northeast Brazil, this species was recorded from the Upper Cretaceous (Turonian-Coniacian) Jandaíra Formation in the Potiguar Basin. Here we extend the palaeogeographical distribution of M. texanus southwards to the Sergipe-Alagoas Basin, where this species occurs in the Turonian-Coniacian Cotinguiba Formation. A key to identification of spatangoids from the Cretaceous-Cenozoic sequence at Sergipe-Alagoas is added and considerations about other spatangoid species from that basin are presented.
Echinodermata;
Spatangoida;
Cretaceous;
South America;
comparison.
Manso C.L. de C. & Souza-Lima W. (2017).- Mecaster texanus (Echinoidea) from Turonian-Coniacian strata in the Sergipe-Alagoas Basin, Brazil.- Carnets Geol., Madrid, vol. 17, no. 11, p. 221-234.
Mecaster texanus (Echinoidea) du Turonien-Coniacien du Bassin de Sergipe-Alagoas, Brésil.- Mecaster texanus (Roemer) a été décrit à l'origine dans les calcaires d'Austin au Texas (États-Unis). Au Brésil, cette espèce n'était jusqu'à présent connue que dans la Formation Jandaíra du Crétacé supérieur (Turonien-Coniacien) dans le bassin Potiguar (Nord-Est du Brésil). Cette étude étend paléogéographiquement sa distribution plus au Sud jusqu'au bassin de Sergipe-Alagoas, où l'espèce a été identifiée dans la Formation Cotinguiba du Crétacé supérieur (Turonien-Coniacien). Une clé pour aider à l'identification des spatangoïdes des séries crétacées et tertiaires de ce bassin est proposée. Une dernière partie présente quelques considérations sur d'autres espèces de spatangoïdes rencontrées dans ce bassin.
Echinodermata ;
spatangoïdes ;
Crétacé ;
Amérique du Sud ;
comparaisons.
Echinoids from the Sergipe-Alagoas Basin have been studied for over one hundred years since White (1887) described the first specimens from Sergipe. Maury (1925) identified numerous new species for the Albian-Turonian sequence, while Brito and Ramirez (1974) and Brito (1980, 1981a, 1981b, 1981c) reviewed and reassigned many species from this basin. Smith (in Smith & Bengtson, 1991) published a detailed taxonomic review of Albian-Coniacian echinoids from northeast Brazil, including those from Sergipe. Manso (2003, 2011, 2014), Manso and Souza-Lima (2003a, 2003b, 2005, 2011, 2012), Manso and Andrade (2008), and Manso and Lemos (2008) studied echinoids mainly from the Aptian-Albian sequence, and established new stratigraphic distributions, recorded new occurrences and erected new species.
A recent study of echinoids of the genus Mecaster from the Sergipe-Alagoas Basin housed in the Fundação Paleontológica Phoenix collections (Aracaju, Sergipe, Brazil) has resulted in the recognition of some species whose apical system supports their confident assignment to Mecaster texanus (Roemer, 1849), a species previously unknown from this part of Brazil. This new record for the Turonian-Coniacian sequence of the Sergipe-Alagoas Basin is discussed in the present paper, which also focuses on its ontogeny and palaeoenviromental distribution.
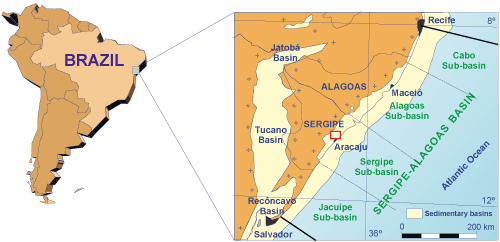
The
Sergipe-Alagoas Basin, located in the coastal region of the states of Sergipe
and Alagoas in northeast Brazil (Fig. 1 ![]() ), features one of the most complete outcrops of Cretaceous
sedimentary sequences on either side of the South Atlantic. Its Aptian-Coniacian
sequence records a broad carbonate platform that developed in response to a
marine ingression that culminated with the establishment of the South Atlantic
Ocean during the breakup of Gondwana from the Early Cretaceous onwards. The
Aptian-Albian Riachuelo Formation represents deposition on a shallow, mixed
siliciclastic-carbonate platform. The Cenomanian-Coniacian Cotinguiba Formation
documents the drowning of the previous platform, currently recorded by deeper
facies, as subsequent tilt of the basin led to erosion of the shallower portions
of this sequence (Souza-Lima, 2006).
), features one of the most complete outcrops of Cretaceous
sedimentary sequences on either side of the South Atlantic. Its Aptian-Coniacian
sequence records a broad carbonate platform that developed in response to a
marine ingression that culminated with the establishment of the South Atlantic
Ocean during the breakup of Gondwana from the Early Cretaceous onwards. The
Aptian-Albian Riachuelo Formation represents deposition on a shallow, mixed
siliciclastic-carbonate platform. The Cenomanian-Coniacian Cotinguiba Formation
documents the drowning of the previous platform, currently recorded by deeper
facies, as subsequent tilt of the basin led to erosion of the shallower portions
of this sequence (Souza-Lima, 2006).
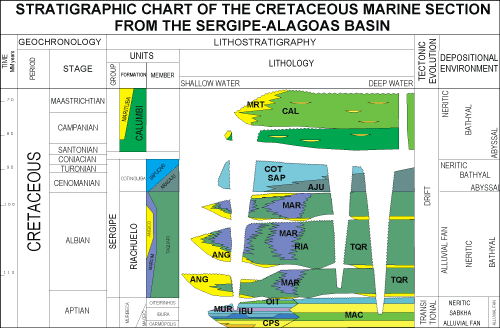
The material
studied comes from the uppermost portion of the Cotinguiba Formation, which is
dated as Turonian-Coniacian on the basis of abundant ammonites from throughout
the section (Bengtson, 1983) (Fig. 2 ![]() ).
).
|
Figure 1:
Location map of
the Sergipe-Alagoas Basin in northeast Brazil (Souza-Lima
et al., 2002). The red rectangle indicates the map of Figure 4 |
|
Figure 2:
Stratigraphic chart of the marine Cretaceous sequences in
the Sergipe-Alagoas Basin (adapted from Feijó, 1995). |
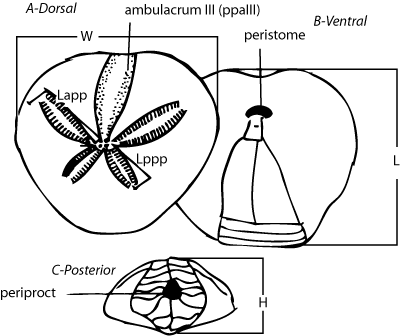
The specimens studied are housed in the fossil echinoderm collections of
the Fundação Paleontológica Phoenix (FPH), Aracaju (Sergipe, Brazil). For
identification of the taxon studied, a series of biometric analyses of tests
were performed: L = test length; W = test width; H = test height;
ppaIII = number of differentiated
pore-pairs in the ambulacrum III, Lapp = length of anterior paired petals; Lppp
= length of posterior paired petals (Fig.
3 ![]() ).
).
|
Figure 3:
Dimensions of tests of Mecaster texanus (Roemer,
1849). |
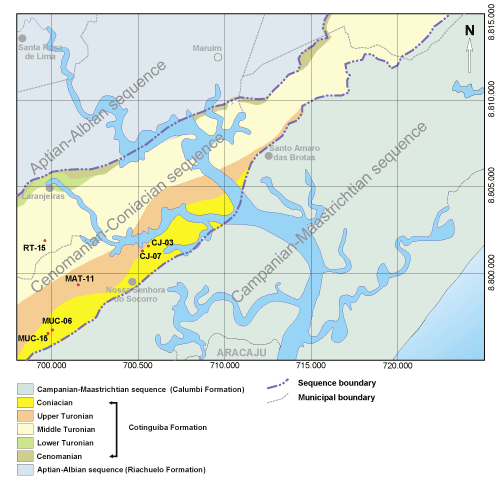
The locality description follows the pattern adopted by Bengtson
(1983, pp. 30-31) for the Sergipe-Alagoas Basin, Brazil. The co-ordinates were
obtained from the Córrego Alegre datum and rounded off to the nearest 50
metres. UTM co-ordinates are referenced to the central 39° meridian. The code
after the location identifies the outcrop on the map (Fig. 4
![]() ).
).
Kcsp = Cotinguiba Formation, Sapucari Member
Cajaíba 3 (CJ-03): UTM 8 801 550N/705 700E. Topographical map sheet: SC.24-Z-B-IV Aracaju. Geological map sheet: SC.24-Z-B-IV-4 Aracaju.
Quarry facing east. Altitude c. 5-20 m.
Kcsp: "Massive, yellowish and brown-grey Laranjeiras limestones with abundant echinoids" (Bengtson, 1983).
References: Probably "Jiquy" of Wanderley (1938: 49) and "Pedreira do Jiqui" of Brito (1980: 572, 575; 1981c: 409).
Cajaíba 7 (CJ-07): UTM 8 801 300N/705 250E. Topographical map sheet: SC.24-Z-B-IV Aracaju. Geological map sheet: SC.24-Z-B-IV-4 Aracaju.
Quarry facing east. Altitude c. 10-20 m.
Kcsp: "Massive, cream and blue-grey Laranjeiras limestones. Large ammonites locally abundant" (Bengtson, 1983).
Mata 11 (MAT-11): UTM 8 799 350N/701 500E. Topographical map sheet: SC.24-Z-B-IV Aracaju. Geological map sheet: SC.24-Z-B-IV-4 Aracaju.
Quarry facing west. Altitude c. 20-30 m.
Kcsp: Hard, cream mudstones, locally with abundant echinoids and inoceramids.
References: "Seção exposta em pedreira ativa com cerca de 110m no Município de Nossa Senhora do Socorro" (Andrade, 2005; Manso & Andrade, 2008).
Mucuri 6 (MUC-06): UTM 8 796 750N/700 050E. Topographical map sheet: SC.24-Z-B-IV Aracaju. Geological map sheet: SC.24-Z-B-IV-4 Aracaju.
Quarry facing southwest. Altitude c. 50-60 m.
Kcsp: "Cream Laranjeiras limestones, locally with abundant ammonites" (Bengtson, 1983).
Mucuri 15 (MUC-15): UTM 8 796 550N/699 800E. Topographical map sheet: SC.24-Z-B-IV Aracaju. Geological map sheet: SC.24-Z-B-IV-4 Aracaju.
Quarry on hillside facing east. Altitude c. 30-35 m.
Kcsp: "Coquinoid, yellowish Laranjeiras limestones" (Bengtson, 1983).
Retiro 15 (RT-15): UTM 8 800 400N/699 450E. Topographical map sheet: SC.24-Z-B-IV Aracaju. Geological map sheet: SC.24-Z-B-IV-4 Aracaju.
Quarry facing northwest. Altitude c. 20-40 m.
Kcsp: "Dominantly massive, cream and blue-grey Laranjeiras limestones with several discontinuity surfaces, some of which are underlain by breccia banks. Also nodular and coquinoid Laranjeiras limestones. Inoceramid bivalves and small ammonites locally abundant" (Bengtson, 1983).
|
Figure
4:
Chronostratigraphic map of the
Aptian-Maastrichtian sequence in the Sergipe-Alagoas Basin, showing outcrops of
the Turonian-Coniacian interval from where Mecaster texanus was collected. |
Three species of the genus Mecaster have been recorded from the Sergipe-Alagoas Basin (Smith in Smith & Bengtson, 1991), namely Mecaster batnensis (Cenomanian-Turonian), M. fourneli (Turonian-Coniacian) and M. africanus (Turonian). According to Smith, M. batnensis usually occurs as a small, widely distributed form in the Cenomanian deposits of the Sergipe-Alagoas Basin. Forms in the mid-Turonian, here called "transitional Mecaster batnensis", are characterised by a greater number of pore pairs in ambulacrum III (Smith in Smith & Bengtson, 1991). The apical system plates' arrangement of these so-called transitional forms is nevertheless similar to that observed in Cenomanian M. batnensis, in which the madreporite separates the posterior genitals I and IV, but not the posterior oculars 1 and 5. Mecaster fourneli differs in having a larger number and narrower plates in ambulacrum III where the two pores of each pair are more closely spaced, and a madreporite separates the two posterior ocular plates. As Smith noted, they probably form an evolutionary lineage (see also Hook & Cobban, 2017). Finally, Mecaster africanus stands out for its greater height than in the other species, with more rectilinear distally open petals, and with madreporite not inserted between the posterior ocular plates.
Phylum Echinodermata Bruguière, 1791
Class Echinoidea Leske, 1778
Cohort Irregularia Latreille, 1825
Superorder Atelostomata von Zittel, 1879
Order Spatangoida Claus, 1876
Suborder Hemiasterina Fischer in Moore, 1966
Family Hemiasteridae H.L. Clark, 1917
Genus Mecaster Pomel, 1883
Mecaster texanus (Roemer, 1849)
Synonymy:
1849 Hemiaster texanus Roemer, p. 77, 393.
1852 Hemiaster texanus Roemer - Roemer, p. 78, 85, Pl. 10, fig. 4 a-c.
1893 Hemiaster texanus Roemer - Clark, p. 86, Pl. 47, fig. 1a-i.
1915 Hemiaster texanus Roemer - Clark & Twitchell, p. 94, Pl. 49, fig. la-j.
1925 Hemiaster sancti-sebastiani Maury, p. 569, Pl. 24, fig. 13.
1928 Hemiaster texanus Roemer - Adkins, p. 299, Pl. 34, figs. 4-5.
1953 Hemiaster texanus Roemer - Cooke, p. 33, Pl. 13, figs. 1-4.
1955 Hemiaster texanus Roemer - Cooke, p. 109, Pl. 29, figs. 5-10.
1964 Hemiaster catandubensis Maury - Beurlen, p. 157, Pl. 20, figs. 118 a-b, 120 a-b.
1966 Hemiaster sancti-sebastiani Maury - Beurlen, p. 463.
1991 Mecaster texanum (Roemer) - Smith in Smith & Bengtson, p. 13.
2003 Hemiaster sanctisebastiani Maury, 1925 - Cassab, p. 75.
2003 Mecaster texanum (Roemer, 1852) - Cassab, p. 76.
2013 Mecaster texanum (Roemer, 1852) - Oliveira et al., p. 11, Figs. 7 A-C, 9 A-I.
Type: Hemiaster texanus Roemer, 1849, from the Austin Chalk at the New Braunfels waterfall of the Guadalupe River (Adkins, 1928, p. 299).
Material: FPH-1457-I; FPH-1500-I; FPH-1887-I; FPH-1888-I; FPH-1889-I; FPH-1890-I; FPH-1891-I; FPH-1892-I; FPH-1955-I.
Localities: Cajaíba 3, Cajaíba 7, Mata 11, Mucuri 6, Mucuri 15 and Retiro 15.
Diagnosis: The peristome is small and surrounded by a well-marked lip. Naked pouches in the interambulacral regions adjacent to the peristome. The apical disc is very elongate, and the madreporite is large and strongly separates the posterior ocular plates (modified from Smith in Smith & Bengtson, 1991).
Description (based on
FPH-1890-I): test longer than wide, measuring 26.7 mm in length, 25,0 mm in
width and 17.5 mm in height, with rounded anterior. The posterior side is
inclined so that the periproct is visible from above (Fig. 6.C ![]() ). The tallest point of
the test is just behind the apical plates. A narrow peripetalous
orthofasciole can be observed around the petals.
). The tallest point of
the test is just behind the apical plates. A narrow peripetalous
orthofasciole can be observed around the petals.
The apical disc, tetrabasal and
ethmolytic, is off-centre and transversely wide (Figs. 7.D![]() ,
8
,
8 ![]() ;
Table 1). Genital plate 4 is just in contact with
the madreporite and well separated from genital plate 1 (e.g., specimen
FPH-1955-I; see Fig. 5.E
;
Table 1). Genital plate 4 is just in contact with
the madreporite and well separated from genital plate 1 (e.g., specimen
FPH-1955-I; see Fig. 5.E ![]() ). The posterior ocular plates are widely separated by
the madreporite so that occasionally they go beyond the ocular plates and touch
the other plates (Fig. 7.C, E
). The posterior ocular plates are widely separated by
the madreporite so that occasionally they go beyond the ocular plates and touch
the other plates (Fig. 7.C, E ![]() ).
).
The frontal ambulacrum is relatively broad and
sunken. There are 35 rounded pore pairs with elevation raised surface between
them (Fig. 6.C ![]() ).
).
Petals are sunken, and almost closed at the end. There are 39 pore pairs in the anterior pair, 11 mm in length, and 30 pore pairs in the posterior petals, 8 mm in length. The petal pores are elongated and slit-like in both series.
The
periproct is small and almost oval in outline and located longitudinally in an
elevated position at the posterior end (Fig. 6.C ![]() ). This feature is well preserved in specimen FPH-1888-I (Fig.
5.D
). This feature is well preserved in specimen FPH-1888-I (Fig.
5.D ![]() ).
).
The peristome is not preserved in
specimen FPH -1890-I, but in FPH-1457-I a well-marked lip is visible around the
peristome (Fig. 5.B ![]() ).
).
Stratigraphical and geographical distribution: Upper Cretaceous Austin Chalk, Anacacho Limestone and Taylor Marl in Texas (United States; Cooke, 1953, 1955); upper part of the Napo Series of Late Cretaceous age in Ecuador (Cooke, 1953, 1955); Jandaíra Formation, Potiguar Basin (Turonian-Coniacian) (Oliveira et al., 2013); and Cotinguiba Formation, Sergipe-Alagoas Basin (Turonian-Coniacian) in the present paper.
|
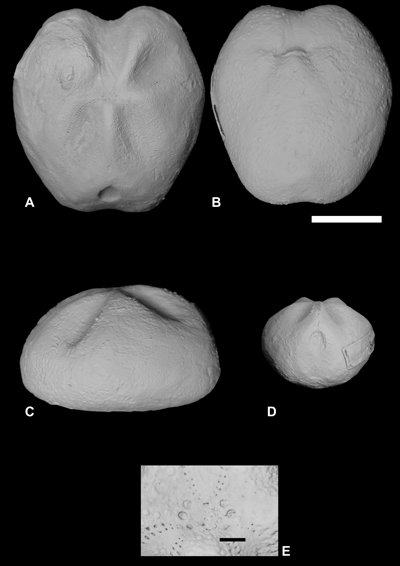
Figure
5:
Mecaster
texanus (Roemer) from the Sergipe-Alagoas Basin : A. Dorsal view
(FPH-1955-I) ; B. Ventral view (FPH-1457-I); C. Lateral view (FPH-1457-I) ; D.
Posterior view (FPH-1888-I) ; E. Detail of apical plating (FPH-1955-I). Scale
bar 10 mm (a-d) and 1 mm (e). |
|
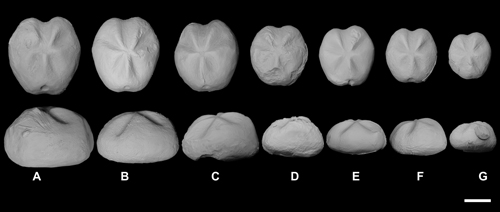
Figure
6:
Ontogenetic series of tests of
Mecaster texanus (Roemer) from the
Sergipe-Alagoas Basin (from larger to smaller, A-G): A (FPH-1955-I), C
(FPH-1890-I), D (FPH-1889-I), F (FPH-1888-I), G (FPH-1891-I) (Coniacian); B
(FPH-1457-I), E (FPH-1500-I) (upper Turonian). Scale bar 10 mm. |
Ontogenetic considerations
In the
small specimen FPH-1891-I (16,2 mm), anterior and posterior petals are almost
the same in size (Fig. 8 ![]() ). The madreporite is located between genital plates 1
and 4, and is smaller than in other, larger specimens (Fig.
6.G
). The madreporite is located between genital plates 1
and 4, and is smaller than in other, larger specimens (Fig.
6.G ![]() ). In this
small individual, ocular plates 1 and 5 are widely separated and the madreporite
between them has only few pores (Fig. 7.A
). In this
small individual, ocular plates 1 and 5 are widely separated and the madreporite
between them has only few pores (Fig. 7.A ![]() ). We believe that this condition was temporary and, with
growth, the madreporite pores could have occupied this place. In madreporites of
other specimens larger than 16 mm in test length, the pores occupy almost the
entire plate. In specimen (FPH-1888-I; 21 mm) the anterior petals are slightly
longer than the posterior ones (Table 1; Fig. 8
). We believe that this condition was temporary and, with
growth, the madreporite pores could have occupied this place. In madreporites of
other specimens larger than 16 mm in test length, the pores occupy almost the
entire plate. In specimen (FPH-1888-I; 21 mm) the anterior petals are slightly
longer than the posterior ones (Table 1; Fig. 8 ![]() ), the apical system is laterally
very wide and the madreporite is located between posterior genitals and ocular
plates but does not go beyond these (Fig. 7.B
), the apical system is laterally
very wide and the madreporite is located between posterior genitals and ocular
plates but does not go beyond these (Fig. 7.B ![]() ). In individuals larger than 21 mm in test length, the
madreporite moves further away from the posterior ocular plates 1 and 5 (Fig.
7.C-E
). In individuals larger than 21 mm in test length, the
madreporite moves further away from the posterior ocular plates 1 and 5 (Fig.
7.C-E ![]() ).
).
Ontogenetic differences observed in the specimens studied suggest a modification of the development of the apical system with growth. It is noted that changes occur in the madreporite, passing from an initial stage with more concentrated pores that became more evenly distributed in adult specimens. Additionally, it is noted that the position of the madreporite shifts progressively more posteriorly with growth, increasing the relative distance between ocular plates 1 and 5.
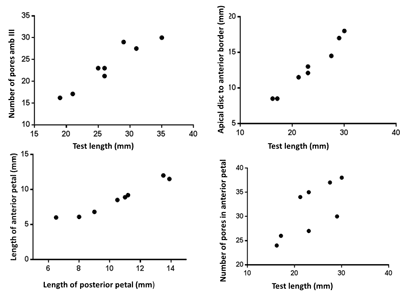
The number of pore
pairs in ambulacrum III between
the apical system and the fasciole was plotted against the test length (Fig. 8 ![]() ). There is a notable increase of pore
pair number with test growth. In comparing these results with the observations
of Smith (in
Smith & Bengtson, 1991, p. 12) for the genus Mecaster
from the Cenomanian-Coniacian
interval in the Sergipe-Alagoas Basin, M.
texanus is very similar to M. fourneli.
Different results were observed by Smith (in
Smith & Bengtson, 1991) for other Mecaster
species from Cenomanian-lower Turonian which showed fewer pore pairs on
ambulacrum III.
). There is a notable increase of pore
pair number with test growth. In comparing these results with the observations
of Smith (in
Smith & Bengtson, 1991, p. 12) for the genus Mecaster
from the Cenomanian-Coniacian
interval in the Sergipe-Alagoas Basin, M.
texanus is very similar to M. fourneli.
Different results were observed by Smith (in
Smith & Bengtson, 1991) for other Mecaster
species from Cenomanian-lower Turonian which showed fewer pore pairs on
ambulacrum III.
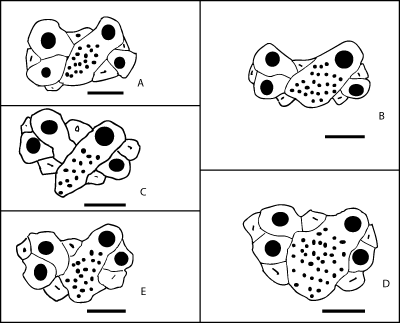
|
Figure
7:
Line
drawings of the apical disc of Mecaster texanus (Roemer) from the Sergipe-Alagoas Basin,
arranged according to increasing test length: A. FPH-1891-I (test length 16.2
mm), B. FPH-1888-I (21.0 mm); C. FPH-1889-I (23.0 mm), D. FPH-1890-I (26.7 mm);
E. FPH-1955-I (30.0 mm). Scale bars 1 mm. |
|
Figure
8:
Biometric
data of selected specimens (n = 8) of Mecaster
texanus (Roemer) from Turonian-Coniacian the Sergipe-Alagoas
Basin. |
Table 1 - Biometric data of Mecaster texanus (Roemer). W - Width; H - Height; L - Length; PAIII - Number of differentiated pore-pairs in the ambulacrum III; PAIII/L - Number of differentiated pair pores on ambulacrumIII/Length; Adab - Apical disc to anterior border; Npap - Number of pores in anterior petals; Nppp - Number of pores in posterior petals; Lppa - length of anterior pair petals; Lppp - Length of posterior pair petals.
| Chronostratigraphy | Fossils N=8 | Length (mm) | Width (mm) | Hight (mm) | W/L | H/L | PAIII | PAIII/L | Adab | Npap | Nppp | Lppa | Lppp |
| Turonian | FPH-1457-I | 29.0 | 26.5 | 18.0 | 0.1 | 0.62 | 29 | 1.0 | 17.0 | 30 | 27 | 13.5 | 12.0 |
| Turonian | FPH-1500-I | 23.0 | 22.5 | 14.0 | 0.97 | 0.60 | 25 | 1.08 | 12.1 | 35 | 25 | 10.5 | 8.5 |
| Turonian | FPH-1955-I | 30.0 | 28.1 | 20.0 | 0.93 | 0.66 | 35 | 1.16 | 18.0 | 38 | 31 | 13.9 | 115 |
| Coniacian | FPH-1889-I | 23.0 | 21.9 | 15.0 | 0.95 | 0.65 | 26 | 1.13 | 13.0 | 27 | 23 | 11.0 | 8.9 |
| Coniacian | FPH-1890-I | 26.7 | 25.0 | 17,5 | 0.93 | 0.65 | 31 | 1.10 | 14.5 | 39 | 30 | 11.0 | 8.0 |
| Coniacian | FPH-1891-I | 16.2 | 15.5 | 11.0 | 0.95 | 0.66 | 19 | 1.12 | 8.5 | 24 | 22 | 8.0 | 6.1 |
| Coniacian | FPH-1887-I | 17.1 | 15.5 | 11.5 | 0.94 | 0.67 | 21 | 1.22 | 8.5 | 26 | 20 | 6.5 | 6.0 |
| Coniacian | FPH-1888-I | 21.0 | 20.1 | 14.3 | 0.87 | 0.67 | 26 | 1.22 | 11.5 | 34 | 26 | 9.0 | 6.8 |
Spatangoid occurrences in the Sergipe-Alagoas Basin
The Order Spatangoida accounts for a substantial portion of the Cretaceous echinoid fauna from the Sergipe-Alagoas Basin. Until now, representatives of this order were recorded from the Aptian-Albian (Riachuelo Formation) and Cenomanian-Coniacian interval (Cotinguiba Formation), although unidentifiable isolated fragments are commonly recovered from the Cenozoic section of this basin (Calumbi Formation). One extant species, Moira atropos, was recorded from Sergipe by Manso et al. (2002).
Echinoid distribution appears to be strongly linked to substrate type, because the larvae of these organisms can only develop when substrate conditions are favourable (Smith, 1984). Therefore, irregular echinoids, such as spatangoids, are restricted to certain environments, as they are morphologically adapted to burying themselves and feeding in very specific sediments (Smith, 1984).
The oldest spatangoid from the Sergipe-Alagoas Basin, Douvillaster benguelensis de Loriol, 1888 (Manso & Souza-Lima, 2003a), now Macraster benguelensis (Kroh, 2014), was identified in the upper Aptian. Their tests exhibit evidence to suggest that they lived epifaunally in an anoxic-dysoxic environment (Neumann, 1996; Manso & Souza-Lima, 2003a; Villier et al., 2004; Manso, 2011; Kamyabi Shadan et al., 2014). Micraster (Epiaster) dartoni (Cooke, 1955), now Epiaster dartoni, was recorded from the lower Albian. Their tests suggest they were semi-infaunal (Manso, 2011; Manso & Souza-Lima, 2012; Kroh, 2015). Recorded from the Albian also were Hemiaster proclivus Cotteau et al., 1878, with low depressed petals, and H. zululandensis Besairie & Lambert, 1930, with more deeply sunken petals and rounded tests, suggesting they were capable of burying themselves in arenaceous sediments close to the water/sediment interface (Manso, 2011). From the Cenomanian to Turonian, a new marine transgression occurred in the Sergipe-Alagoas Basin, with deposition of fine-grained sediments (marls and mudstones of the Cotinguiba Formation), and many species of the genus Mecaster appeared. The tests of this genus are adapted for living buried more deeply than the spatangoids from the Riachuelo Formation. Burial capacity involves modification in the disposition of fascioles, the number and disposition of the frontal ambulacral pores, the depth of petals and even the test layout (Smith, 1984; Kanazawa, 1992; Smith & Stockley, 2005). The establishment of more specialised species for living in sediments can be related to the presence of predators in these palaeocommunities, and burial could have been a defence strategy. According to Vermeij (1977), during the Mesozoic modification of the structure of marine benthic communities occurred, caused by the diversification of the gastropods. Previously, gastropods were mainly grazers but, with morphological change, they acquired new feeding habits, becoming predators of other species. These molluscs rapidly diversified towards the end of the Cretaceous. In the sedimentary basins of northeastern Brazil the presence of carnivorous gastropods has previously been recorded, and has been considered in a global Cretaceous evolutionary context (Andrade & Félix, 2012).
On the basis of the diverse record of the Order Spatangoida of the Sergipe-Alagoas Basin, a key is proposed for the identification of such echinoids from this basin; this may prove useful for group identification in other areas as well.
Identification key for species of the Order Spatangoida currently known from the Sergipe-Alagoas Basin and their stratigraphical occurrence.
| 1 - Apical disc ethmophract with four gonopores; | madreporite not separating genital plates 1 and 4 | jump to 2 |
| Apical disc ethmophract with four gonopores; | madreporite separating genital plates 1 and 4 | jump to 5 |
| Apical disc ethmolytic with four gonopores; | madreporite separating genital plates 1 and 4 and posterior ocular plates 1 and 5 | jump to 6 |
| Apical disc ethmolytic with two gonopores | jump to 7 | |
| 2 - Petals slightly ridged and distally open; | posterior petals slightly shorter than anterior ones | jump to 3 |
| Petals almost closed; | posterior petals less than approximately half length of anterior pair | jump to 4 |
| 3 - Pore pairs in ambulacrum III slit-shaped | Macraster benguelensis (upper Aptian-lower Albian; e.g., Manso & Souza-Lima, 2003a) | Fig. 9AI |
| Pore pairs in ambulacrum III rounded | Epiaster dartoni (lower Albian; e.g., Manso & Souza-Lima, 2012) | Fig. 9.B |
| 4 - Petals shallow; test taller in posterior region | Hemiaster proclivus (Albian; e.g., Smith & Bengtson, 1991) | Fig. 9.C |
| Petals more deeply sunken; test profile much more depressed | Hemiaster zululandensis (upper Albian; e.g., Manso & Souza-Lima, 2003b) | Fig. 9.D |
| 5 - Petals deeply sunken and broad; anterior and posterior pairs of almost same size | Mecaster batnensis (Cenomanian-lower Turonian; e.g., Smith & Bengtson, 1991) | Fig. 9.E |
| Petals sunken, straight, distally opened and subequal in length | Mecaster africanus (upper Turonian; e.g., Smith & Bengtson, 1991) | Fig. 9.F |
| 6 - Apical disc not very elongate, with madreporite narrowly separating ocular plates | Mecaster fourneli (Turonian-Coniacian; e.g., Smith & Bengtson, 1991) | Fig. 9.G |
| Apical disc very elongate, with madreporite broadly separating ocular plates, occasionally extending beyond these plates | Mecaster texanus (Coniacian; the present paper) | Fig. 9.H |
| 7 - Posterior genital plates widely separated | Moira atropos (Pleistocene to Recent; e.g., Manso et al., 2002) | Fig. 9.I |
|
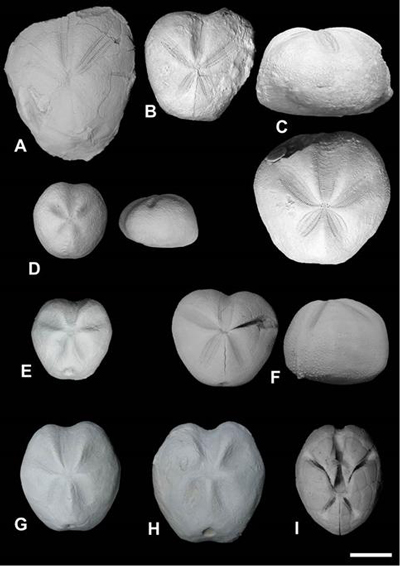
Figure
9:
Spatangoid echinoids from
the Sergipe-Alagoas Basin: A. Macraster
benguelensis (FPH-1961-I); B. Epiaster
dartoni (FPH-423-I); C. Hemiaster
zululandensis (FPH-341-I); D. Hemiaster
proclivus (FPH-645-I); E. Mecaster
batnensis (FPH-1963-I); F. Mecaster
africanus (FPH-1964-I); G. Mecaster
fourneli (FPH-1948-I); H. Mecaster
texanus (FPH-1955-I); I. Moira atropos
(FPH-1957-I). Scale bar 10 mm. |
Mecaster texanus co-occurs with M. fourneli (Deshayes in Agassiz & Desor, 1847) at all localities studied. These species are very close, showing the same number of pore pairs in ambulacrum III. The main difference between them is the larger size of the madreporite plate in M. texanus in individuals of the same test size. Another difference concerns the peristome which is wider in M. fourneli than in M. texanus.
Smith (in Smith & Bengston, 1991), who assessed other species of Mecaster of Cretaceous (Cenomanian-Coniacian) age in the Sergipe-Alagoas Basin, concluded that the density of pore pairs between the apical system and the fasciole in ambulacrum III exhibits a general increase through time. Thus, Cenomanian Mecaster batnensis has fewer pore pairs than Turonian M. africanus and Coniacian M. fourneli. This also appears to be true for M. texanus which has a similar number of pore pairs to M. fourneli from the Turonian-Coniacian of this basin.
Mecaster also exhibits a decrease in test roundness from the Cenomanian to the Turonian. In the Coniacian, the tests again display greater width (Smith in Smith & Bengtson, 1991).
Cretaceous spatangoids can be subdivided into two groups on the basis of the length of their even petals (Zoeke, 1951): those with long petals with many pore pairs and those with short petals (especially the posterior pair) with few pore pairs. As echinoid metabolism is influenced by water temperature and since their distribution may be limited by this variable (Smith, 1984), some inferences can be made. Those species with longer petals would have higher metabolic rates and would occur in warmer waters. In contrast, those species with shorter petals, with lower metabolism, would be found in colder waters. According to Smith (1984), hemiasterids that lived around the palaeoequator during the Cretaceous had long petals and species with shorter petals existed in more temperate regions in both hemispheres. In the Sergipe-Alagoas Basin, spatangoids with long petals appeared in the late Aptian and lived on a shallow platform in warm waters in anoxic-dysoxic palaeoenvironments (Manso & Souza-Lima, 2003a). Organisms with long petals and similar pores and slits in all ambulacra would have more efficient gas exchange (Neumann, 1996; Manso & Souza-Lima, 2003a; Kamyabi Shadan et al., 2014). During the Albian, species of Hemiaster with posterior petals shorter than the frontal ones appeared, and were better adapted to colder, oxygenated and probably deeper palaeoenvironments (Smith & Bengtson, 1991; Néraudeau & Floquet, 1991; Manso & Souza-Lima, 2003a). It is assumed that at the start of the Albian the Sergipe-Alagoas Basin was further away from the palaeoequator. From the Cenomanian to Coniacian the position of this basin gradually moved to higher latitudes probably with warmer waters. During this interval there was greater communication between oceanic basins and greater oxygenation of bottom waters globally (Smith & Stockley, 2005). At higher water temperatures than during Albian, the species of Mecaster with longer posterior petals were widely distributed on the Sergipe-Alagoas Basin platform.
Regarding the size of even petals, and the number and arrangement of pores in the anterior ambulacrum in species of Mecaster from the Cretaceous of the Sergipe-Alagoas Basin, Guimarães (2016) observed that in the Turonian two distinct morphological groups co-occurred. The first, comprising M. batnensis and M. africanus, has narrow and open petals and fewer pores in ambulacrum III than their counterparts. The second group, formed by M. fourneli and M. texanus, exhibits closed petaloids, less rounded tests and a larger number of pores in ambulacrum III. In comparing these two groups with results obtained by Néraudeau and Floquet (1991) for the Upper Cretaceous of the Castilla and Navarra-Cantabria platforms of Spain, Guimarães (2016) concluded that the species from the Sergipe-Alagoas Basin could have been distributed in a similar way during the Turonian. Mecaster fourneli and M. texanus would have occupied shallower environments on the inner platform where they would have buried themselves more deeply into the sediment. Mecaster batnensis and M. africanus would have preferred deeper areas but buried themselves more closely to the sediment/water interface. Néraudeau and Floquet (1991) also observed that the spatial distribution of echinoids reflected a palaeoecological control and that differences in depths in distinct geographic outcrops could be related to transgressive and regressive events.
Mecaster texanus and M. fourneli were recorded from the Sergipe-Alagoas Basin up to the Coniacian. From the Santonian onwards, a new trangressive event, possibly related to a climatic turnover, laid down siliciclastic strata (Calumbi Formation) over the previous carbonate platform in which these species of Mecaster probably could not survive.
The authors wish to thank Dr Andreas Kroh (Naturhistorisches Museum Wien, Vienna) for suggestions on an earlier typescript, Dr Loïc Villier and Dr John Jagt for their valuable suggestions that helped to improve the final manuscript, and Dr Stephen Carey by the English corrections.
Adkins W.S. (1928).- Handbook of Texas Cretaceous fossils.- University of Texas Bulletin, Austin, vol. 2838, p. 1-381.
Andrade E.J. (2005).- Turonian inoceramids and biostratigraphy of the Sergipe Basin, northeastern Brazil: An integrated study of the Votorantim and Nassau quarries.- PhD thesis, Universität Heidelberg, 155 p.
Andrade E.J. & Félix I.L.C. (2012).- Gastrópodos marinhos do Turoniano (Cretáceo Superior) da Bacia de Sergipe.- Cuadernos de Geociências, Bahia, vol. 9, no. 2, p. 103-111.
Bengtson P. (1983).- The Cenomanian-Coniacian of the Sergipe Basin, Brazil.- Fossils and Strata, Oslo, no. 12, 78 p.
Beurlen K. (1964).- A fauna do calcário Jandaíra da região de Mossoró (Rio Grande do Norte).- Coleção Mossoroense, Editora Pongetti, Rio de Janeiro, 215 p.
Beurlen K. (1966).- Novos equinóides do Cretáceo do Nordeste do Brasil.- Anais da Academia brasileira de Ciências, Rio de Janeiro, vol. 38, nos. 3-4, p. 455-464.
Brito I.M. (1980).- Os equinóides fósseis do Brasil: I. Os endocíclicos.- Anais da Academia Brasileira de Ciências, Rio de Janeiro, vol. 52, no. 3, p. 569-590.
Brito I.M. (1981a).- Os equinóides fósseis do Brasil: II. Holectypoides e cassiduloides.- Anais da Academia Brasileira de Ciências, Rio de Janeiro, vol. 53, no. 3, p. 513-527.
Brito I.M. (1981b).- Os equinóides fósseis do Brasil: III. Espatangóides, exceto Hemiasteridae.- Anais da Academia Brasileira de Ciências, Rio de Janeiro, vol. 53, no. 3, p. 569-578.
Brito I.M. (1981c).- O gênero Hemiaster no Cretáceo do Brasil (Echinoidea Spatangoida). In: Sanguinetti Y.T. (ed.), Anais do II Congresso Latino-Americano de Paleontologia.- Editora Pallotti, Porto Alegre, vol. 1, p. 399-414.
Brito I.M. & Ramirez L.V.O. (1974).- Contribuição ao conhecimento dos equinóides albianos (Cretáceo inferior) do Brasil.- Anais da Academia Brasileira de Ciências, Rio de Janeiro, vol. 46, no. 2, p. 275-282.
Cassab R. de C.T. (2003).- Paleontologia da Formação Jandaíra, Cretáceo Superior da Bacia Potiguar, com ênfase na paleobiologia dos gastrópodos.- PhD thesis, Universidade Federal do Rio de Janeiro, Instituto de Geociências, Rio de Janeiro, 184 p. URL: http://www.cprm.gov.br/publique/media/rita.pdf
Clark W.B. (1893).- The Mesozoic Echinodermata of the United States.- United States Geological Survey, Bulletin, vol. 97, p. 1-207.
Clark W.B. & Twitchell M.W. (1915).- The Mesozoic and Cenozoic Echinodermata of the United States.- United States Geological Survey, Monographs, vol. 14, 468 p.
Cooke C.W. (1953).- American Upper Cretaceous Echinoidea.- United States Geological Survey, Professional Paper, vol. 254-A, p. 1-44.
Cooke C.W. (1955).- Some Cretaceous echinoids from the Americas.- United States Geological Survey, Professional Paper, vol. 264-E, p. 87-112.
Feijó F.J. (1995).- Bacias de Sergipe e Alagoas.- Boletim de Geociências da Petrobras, Rio de Janeiro, vol. 8, no. 11 (for 1994), p. 149-161.
Fisher A.G. (1966).- Spatangoids. In: Moore R.C. (ed.), Treatise on Invertebrate Paleontology, Part U, Echinodermata 3.- Geological Society of America, Boulder, and University of Kansas, Lawrence, vol. 2, U543-U628.
Guimarães C.D.M. (2016).- Identificação e distribuição das espécies de Mecaster (Echinodermata: Echinoidea) do Cretáceo da Bacia de Sergipe-Alagoas, nordeste do Brasil.- Dissertação, Departamento de Geologia, Universidade Federal de Sergipe, São Cristóvão, 42 p. URL: https://ri.ufs.br/handle/riufs/5402
Hook S.C. & Cobban W.A. (2017).- Mecaster batnensis (Coquand, 1862), a late Cenomanian echinoid from New Mexico, with a compilation of Late Cretaceous echinoid records in the Western Interior of the United States and Canada.- Acta Geologica Polonica, vol. 67, no. 1, p. 1-30.
Kamyabi Shadan H., Villier L., Sadeghi A. & Adabi M.H. (2014).- A revision of the Macraster species (Echinodermata, Echinoidea) occurring in the Albian deposits of the Zagros Basin, southwest Iran.- Annales de Paléontologie, Paris, vol. 100, no. 1, p. 51-62.
Kanazawa K. (1992).- Adaptation of test shape for burrowing and locomotion in spatangoid echinoids.- Palaeontology, vol. 35, part 4, p. 733-750.
Kroh A. (2014).- Macraster benguellensis (de Loriol, 1888) . In: Kroh A. & Mooi R. (2017).- World Echinoidea Database. Accessed through: World Register of Marine Species at http://www.marinespecies.org/traits/aphia.php?p=taxdetails&id=746347 on 2017-11-08
Kroh A. (2015).- Epiaster dartoni Cooke, 1955 . In: Kroh A. & Mooi R. (2015).- World Echinoidea Database. Accessed through: World Register of Marine Species at http://www.marinespecies.org/echinoidea/aphia.php?p=taxdetails&id=755656 on 2016-08-22
Manso C.L. de C. (2003).- Paleoecologia, paleobiogeografia e sistemática dos equinóides do Aptiano-Albiano (Cretáceo) do Brasil.- PhD thesis, Universidade Federal da Bahia, Instituto de Geociências, Salvador, 211 p.
Manso C.L. de C. (2011).- Adaptação e distribuição dos espatangóides (Echinodermata: Echinoidea) no Cretáceo da Sub-bacia de Sergipe, Brasil. In: Carvalho I.S., Srivastava N.K., Strohschoen O. Jr & Lana C.C. (eds.).- Paleontologia: Cenários da Vida, Rio de Janeiro, vol. 4, p. 123-132.
Manso C.L. de C. (2014).- Os equinodermos como indicadores paleoclimáticos do Cretáceo do Brasil. In: Carvalho I.S., Srivastava N.K., Strohschoen O. Jr & Lana C.C. (eds.), Paleoclimas.- Paleontologia: Cenários da Vida, Rio de Janeiro, vol. 5, p. 63-70.
Manso C.L. de C. & Andrade E.J.A. (2008).- Equinóides do Turoniano (Cretáceo Superior) de Sergipe, Brasil.- Geociências, Rio Claro, vol. 27, no. 3, p. 319-327.
Manso C.L. de C. & Lemos A.C.C. (2008).- Os Echinoidea (Echinodermata) da localidade Catete Velho I (Cretáceo) em Sergipe, Nordeste do Brasil.- Revista Brasileira de Paleontologia, Porto Alegre, vol. 11, no. 2, p. 129-138.
Manso C.L. de C. & Souza-Lima W. (2003a).- O equinóide Douvillaster Lambert, 1917, na Formação Riachuelo, Bacia de Sergipe, Brasil.- Revista Brasileira de Paleontologia, Porto Alegre, no. 5, p. 29-37.
Manso C.L. de C. & Souza-Lima W. (2003b).- O registro do equinóide Hemiaster zululandensis Besairie & Lambert, 1930, no Cretáceo (Albiano superior) de Sergipe.- Revista Brasileira de Paleontologia, Porto Alegre, no. 6, p. 61-67.
Manso C.L. de C. & Souza-Lima W. (2005).- Os equinóides Pygorhynchus colombianus (Cooke, 1955) e Pseudholaster altiusculus (White, 1887) no Neoalbiano da Bacia de Sergipe, Brasil.- Revista Brasileira de Paleontologia, Porto Alegre, vol. 8, p. 229-238.
Manso C.L. de C. & Souza-Lima W. (2011).- New genus and new species of echinoid (Echinodermata, Orthopsidae) from the Albian (Lower Cretaceous) of Sergipe-Alagoas Basin, Brazil.- Arquivos do Museu Nacional, Rio de Janeiro, vol. 68, no. 1-2, p. 41-46.
Manso C.L. de C. & Souza-Lima W. (2012).- Novos equinóides para o Aptiano-Albiano (Cretáceo inferior) da sub-bacia de Sergipe, Nordeste do Brasil.- Geociências, Rio Claro, vol. 31, no. 4, p. 584-605.
Manso C.L. de C., Souza-Lima W. & Souza-Lima R. (2002).- Ocorrência de Moira atropos Lamarck, 1816 (Echinodermata: Echinoidea) no litoral de Sergipe. In: Congresso Brasileiro de Zoologia, 24, Itajaí, Santa Catarina, 2002.- Boletim de Resumos, Sociedade Brasileira de Zoologia, vol. 1, p. 92.
Maury C.J. (1925).- Fósseis terciarios do Brasil com descripção de novas formas cretáceas.- Monografia, Serviço Geologico e Mineralógico, Rio de Janeiro, no. 4, p. 1-305.
Néraudeau D. & Floquet M. (1991).- Les échinides Hemiasteridae: marqueurs écologiques de la plate-forme castillane et navarro-cantabre (Espagne) au Crétacé supérieur.- Palæogeography, Palæoclimatology, Palæoecology, no. 88, p. 265-281.
Neumann C. (1996).- The mode of life and paleobiogeography of the genus Douvillaster Lambert (Echinoidea: Spatangoida) as first recorded in the Lower Cretaceous (Albian) of Spain.- Berliner geowissenschaftliche Abhandlungen, E18, p. 257-265.
Oliveira J., Manso C.L. de C., Andrade E.J. & Souza-Lima W. (2013).- O gênero Mecaster (Echinodermata: Spatangoida) do Cretáceo superior da Formação Jandaíra, Bacia Potiguar, Nordeste do Brasil.- Scientia Plena, São Cristóvão, vol. 9, no. 8, p. 1-17.
Pomel A. (1883).- Classification méthodique et genera des Échinides vivants & fossiles.- 1ère thèse, Docteur ès Sciences Naturelles, Faculté des Sciences de Paris, no. 503, 131 p. URL: https://www.biodiversitylibrary.org/item/43038#page/5/mode/1up
Roemer F. (1849).- Texas, mit besonderer Rucksicht auf Deutsche Auswanderung und die physischen Verhältnisse des Landes nach eigener Beobachtung geschildert; mit einem naturwissenschaftlichen Anhange und einer topographisch-geognostischen Karte.- Bonn, Adolph Marcus, 626 p.
Roemer F. (1852).- Die Kreidebildungen von Texas und ihre organischen Einschlusse.- Bonn, Adolph Marcus, 100 p.
Smith A.B. (1984).- Echinoid Palaeobiology.- George Allen & Unwin, London, 190 p.
Smith A.B. & Bengtson P. (1991).- Cretaceous echinoids from north-eastern Brazil.- Fossils and Strata, Oslo, no. 31, 88 p.
Smith A.B. & Stockley B. (2005).- Fasciole pathways in spatangoid echinoids: A new source of phylogenetically informative characters.- Zoological Journal of the Linnean Society, no. 144, p. 15-35.
Souza-Lima W. (2006).- Litoestratigrafia e evolução tectono-sedimentar da bacia de Sergipe-Alagoas: Introdução.- Phoenix, Aracaju, no. 89, p. 1-10.
Souza-Lima W., Andrade E.J., Bengtson P. & Galm P.C. (2002).- A bacia de Sergipe-Alagoas. Evolução geológica, estratigrafia e conteúdo fóssil [The Sergipe-Alagoas Basin. Geological evolution, stratigraphy and fossil content].- Phoenix, Aracaju, Edição Especial, no. 1, p. 1-34.
Vermeij G.J. (1977).- The Mesozoic marine revolution: evidence from snails, predators and grazers.- Paleobiology, no. 3, p. 245-258.
Villier L., Néraudeau D., Clavel B. & Neumann C. (2004).- Phylogeny of early Cretaceous spatangoids (Echinodermata: Echinoidea) and taxonomic implications.- Palaeontology, no. 47, p. 265-292.
Wanderley A. (1938).- Estado de Sergipe. Estudos geológicos e levantamentos topográficos. In: Oliveira E.P. de (ed.).- Relatório Annual do Director, anno de 1935, p. 46-57.
White C.A. (1887).- Contribuições à paleontologia do Brazil.- Archivos do Museu Nacional do Rio de Janeiro, no. 7, p. 1-273.
Zoeke E. (1951).- Étude des plaques des Hemiaster (Échinides).- Bulletin du Muséum national d'Histoire naturelle de Paris, (2), no. 23, p. 696-705.