◄ Carnets Geol. 21 (19) ►
![]()
Outline:
[1. Introduction]
[2. Geological setting and material]
[3. Systematic palaeontology]
[4. Discussion about evolutionary and biological considerations on
paracomatulids] [5. Conclusions] and ... [Bibliographic references]
Department
of Earth Sciences, "Sapienza" University of Rome, P.le A. Moro 5, I-00185
Roma (Italy)
Via del Teatro Greco 6, I-00019 Tivoli, Roma (Italy)
Published online in final form (pdf) on December 25, 2021
DOI 10.2110/carnets.2021.2119
![]()
[Editor:
Bruno R.C. Granier; language editors: Stephen Carey
and Bruno Ferré]
![]()
A new paracomatulid crinoid, Tiburtocrinus toarcensis gen. et sp. nov., is described from the Lower Jurassic of Tivoli (central Apennines, Italy). This type of stemless crinoids has never previously been recorded in Italy, and this report bridges a significant gap. Morphofunctional analysis of the radial facets suggests that Tiburtocrinus toarcensis gen. et sp. nov. may have been a crawling paracomatulid, very different from other paracomatulids that probably swam.
Echinoderm;
Crinoidea;
Isocrinida;
Paracomatulidae;
Lower Jurassic;
Toarcian;
central Apennines;
morphofunctional analysis
Manni R. & Di Nardo R. (2021).- A possible crawling paracomatulid crinoid from the Lower Jurassic of central Italy.- Carnets Geol., Madrid, vol. 21, no. 19, p. 523-532.
Un crinoïde paracomatulide probablement rampant du Jurassique inférieur d'Italie centrale.- Un nouveau crinoïde paracomatulide, Tiburtocrinus toarcensis gen. et sp. nov., est décrit dans le Jurassique inférieure de Tivoli (Apennins centraux, Italie). Jamais signalée jusqu'à present en Italie, cette découverte comble une lacune au sein des descriptions de ce type de crinoïde sans tige. L'analyse morphofonctionnelle de ses facettes radiales montre que Tiburtocrinus toarcensis gen. et sp. nov. était probablement un paracomatulide rampant, très différent des autres paracomatulides connus qui, eux, nageaient selon toute probabilité.
Échinoderme ;
Crinoidea ;
Isocrinida ;
Paracomatulidae ;
Jurassique inférieur ;
Toarcien ;
Apennins centraux ;
analyse morphofonctionnelle
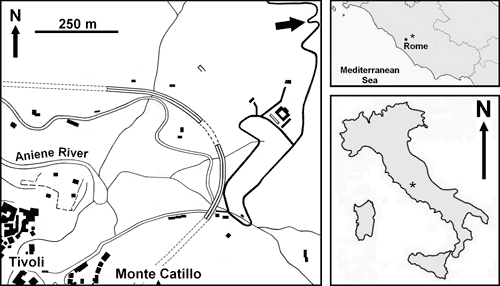
A well-preserved cup of a primitive "comatulid-type" crinoid was retrieved from a lower Toarcian limestone sequence in the Monte Catillo Natural Reserve near Tivoli City. The fossil is a paracomatulid, a group of crinoids of considerable evolutionary importance because they were allegedly the first pentacrinitids to lose the stem in order to move freely. Its major characteristic is a reduced column consisting of few distinct columnals with and without cirri. This crinoid displays columnals with cirri typical of pentacrinitids, organised not as a stem, but as a centrodorsal, such an anatomical architecture being typical of comatulids.
The outcrop that yielded the specimen studied here lies
in the Monte Catillo Natural Reserve, a protected area of the Province of Rome,
near Tivoli City, east of Rome (central Italy). Specifically, the specimen was
found at Colle Piano (Fig. 1 ![]() ). Colle Piano is a hill where Lower Jurassic
limestone layers of the Corniola Formation are exposed (sheet 150 Rome from the
1:100,000 geological map of Italy). These beds consist of white and light grey
micritic limestones with chert lenses and yellowish mudstones. The specimen was found in a
marly-pebbly, slump deposit. In this slump
deposit were found several other crinoids and brachiopods, as well as ammonites,
belemnites, gastropods, and sponges. Colle Piano is well-known
in the palaeontological literature: Manni and Nicosia
(1990, 1994), Vörös
(2002), and Venturi (2006) described specimens of crinoids, brachiopods
and ammonites respectively, several of these as new species. The stratigraphical age of
these layers is Toarcian, though Vörös (2002) also considered a
Pliensbachian age possible. Manni and Nicosia
(1990) regarded
these beds as Toarcian (probably Serpentinus
Zone) based on the associated ammonites. Vörös (2002) considered a
Pliensbachian age more likely based on brachiopod fauna (mainly of the
koninckinid type). However, in the context of the discovery of specimens of Koninckodonta
davidsoni, Linguithyris aspasia and Securithyris adnethensis, Vörös also considered an early Toarcian age plausible. On the other hand, due
to the occurrence of Canavaria retroplicata, Dactylioceras
pseudocommune and Harpophylloceras eximium, Venturi
(2006) correlated the sequence with the upper
part of the first ammonite zone of the lower Toarcian (Polymorphum Zone).
). Colle Piano is a hill where Lower Jurassic
limestone layers of the Corniola Formation are exposed (sheet 150 Rome from the
1:100,000 geological map of Italy). These beds consist of white and light grey
micritic limestones with chert lenses and yellowish mudstones. The specimen was found in a
marly-pebbly, slump deposit. In this slump
deposit were found several other crinoids and brachiopods, as well as ammonites,
belemnites, gastropods, and sponges. Colle Piano is well-known
in the palaeontological literature: Manni and Nicosia
(1990, 1994), Vörös
(2002), and Venturi (2006) described specimens of crinoids, brachiopods
and ammonites respectively, several of these as new species. The stratigraphical age of
these layers is Toarcian, though Vörös (2002) also considered a
Pliensbachian age possible. Manni and Nicosia
(1990) regarded
these beds as Toarcian (probably Serpentinus
Zone) based on the associated ammonites. Vörös (2002) considered a
Pliensbachian age more likely based on brachiopod fauna (mainly of the
koninckinid type). However, in the context of the discovery of specimens of Koninckodonta
davidsoni, Linguithyris aspasia and Securithyris adnethensis, Vörös also considered an early Toarcian age plausible. On the other hand, due
to the occurrence of Canavaria retroplicata, Dactylioceras
pseudocommune and Harpophylloceras eximium, Venturi
(2006) correlated the sequence with the upper
part of the first ammonite zone of the lower Toarcian (Polymorphum Zone).
|
Figure 1: Schematic map of central Italy, and location of fossiliferous outcrop (star). Figure 1 :
Carte schématique d'Italie centrale et localisation géographique du gisement (étoile). |
The Corniola Formation is a Sinemurian-lower Toarcian lithostratigraphic unit of the Umbrian-Marchean succession. It is characterised by centimetric and decimetric beds of white and beige micritic limestone, often with chert lenses. During the Hettangian-early Sinemurian, the area of the central Apennines was covered by a tropical sea where limestones of shallow-water carbonate platform were deposited. From the Sinemurian the Neo-Tethys Ocean opening started. An intense phase of rifting induced the partial drowning of the platform (Centamore et al., 1971; Farinacci et al., 1981; Santantonio, 1994; Fabbi & Santantonio, 2012). Two domains are now recognised: 1) a shallow-water setting, called the Latium-Abruzzi carbonate platform, with tidal limestones, and 2) a deeper-marine setting, the Umbrian-Marchean pelagic domain, with pelagic limestones. Between these two domains lies a transitional domain characterised by a slope down which sediments from the carbonate platform had flowed and often settled. The paracomatulid at hand was found in this transitional domain.
The specimen examined and illustrated here was collected in situ and is now stored in the Museum of the Earth Sciences Department, "Sapienza" University of Rome, Italy.
Hess (1951) erected the new genus Paracomatula and assigned it to the new family Paracomatulidae Hess, 1951.
Rasmussen (1978) assigned Paracomatulidae to the order Comatulida A.H. Clark, 1908.
Simms (1988) assigned Paracomatulidae to the order Isocrinida Sieverts-Doreck, 1952, suborder Comatulidina A.H. Clark, 1908.
Nicosia (1991), Hess (2006), and Hess and Messing (2011), all following the systematics of Rasmussen, assigned Paracomatulidae to the Comatulida.
On the other hand, Hagdorn and Campbell (1993) followed the systematics of Simms and assigned Paracomatulidae to the Isocrinida, suborder Comatulidina.
Finally, Hess (2014, p. 31) assigned Paracomatulidae to the order Isocrinida, suborder Pentacrinitina Gray, 1842, because he believed that the paracomatulids have no real centrodorsal (CD), but a reduced CD-like column.
The varying assignment to Isocrinida or Comatulida, to Comatulidina or Pentacrinitina, is due to the subjective interpretation of the main characters considered by the successive authors. The real issue is whether this CD is a genuine one or not. Secondarily it also raises the issue of whether or not the basal plates have a pentacrinitid-type articulation. The basal plates are obviously articulated with the CD by means of a pattern of narrow petals bordered with short crenulae, like that of pentacrinitids; likewise, the columnals of the CD display cirri as the columnals of the pentacrinitids do. But the CD of true comatulids consists of a single plate, in contrast to the CD of Paracomatula consisting of several distinct columnals. Therefore, the Paracomatula CD is not a true one; it is simply an extremely small stem that served as a CD. In the comatulids, the CD is a single and distinct element that completely lost the functionality of the stem. Hess (2014, p. 26) considered "The segmented centrodorsals of paracomatulids and the centrodorsals of comatulids s. str." to be "homologous structures descended from a pentacrinitid stem group". Thus, the solution is philosophical: Is Paracomatula the first comatulid, or is it one of the intermediates between Pentacrinitina and comatulids? Following Hess (2014), Paracomatula is here considered one of the forms before the real comatulids, and, in this paper, the systematics proposed by Hess (2014) is followed.
Order Isocrinida, Sieverts-Doreck, 1952
Suborder Pentacrinitina Gray, 1842
Family Paracomatulidae Hess, 1951
Diagnosis: The cup of these crinoids is small and characterised by a CD-like stem, five basal plates and five radial plates. The CD-like stem has one to several non-fused columnals with and without cirri. The basal circle, low and star-shaped, has interradial petals bordered by short crenulae. The radial circle, low and of pentagonal outline, is commonly recumbent or overhanging. The radial facets are large and often subvertical. Arms are divided at second primibrachials. First pinnules are on second secundibrachials. The syzygial articulations have only a few coarse culmina.
Composition: Paracomatula Hess, 1951, and Singillatimetra Hess, 2012.
Genus Tiburtocrinus gen. nov.
Type species: T. toarcensis gen. et sp. nov.
Etymology: From Tiburto. According to mythology, Tiburto was one of the three founders of Tivoli City (from the Latin name of Tibur) in 1,215 BC.
Diagnosis: Small sleek cup with CD-like stem, five basal plates and five radial plates. The CD-like stem has five sides, and is moderately high, with three columnals not fused, two of which have cirrus sockets. The basal circle is low. Each basal plate has a lower edge curved downwards, overhanging the top of the first columnal. The radial circle is moderately higher, pentagonal in outline. Each radial facet is separated from the adjacent one by false interradial process. The radial facets are oblong, with a wide aboral ligament surface and smaller interarticular ligament fossae and muscular fossae.
Remarks: The main characters of this crinoid are three columnals, a rather protruding radial circle, oblong radial facets weakly inclined outwards, weakly extended muscular fossae, and basal plates whose lower edges curve downwards. Together, these characters, which are not shared by any other paracomatulid, justify the erection of a new genus, assigned to the family Paracomatulidae. The complete anatomy of this new genus remains unknown because no complete specimen has been found so far. Tiburtocrinus gen. nov. differs from Paracomatula in having non-subvertical radial facets, smaller muscular fossae and basal plates whose lower edges curve downwards. Finally, this new genus differs from Singillatimetra because the CD-like stem of the latter is characterised by a single columnal. Other comparisons are not possible because the basal and radial plates of Singillatimetra are unknown.
It could also be hypothesised that this CD-like stem is a case of stem regrowth, after a traumatic event that would have involved the rupture and detachment of proximal columnals (Nakano et al., 2004). However, there is no evidence to support this. Furthermore, the characters of this specimen are typical of paracomatulids. It should also be remarked that the cases described by Nakano et al. (2004) were all experimentally induced and that no similar cases of regeneration of columnals in living crinoids have ever been observed.
Other species: Unknown.
Occurrence: Lower Toarcian of central Italy.
Tiburtocrinus toarcensis sp. nov.
Holotype: Specimen NS 6/950.
Material: Holotype specimen NS 6/950 only.
Etymology: Referring to the Toarcian age of the specimen.
Type horizon and type locality: Lower Toarcian; Colle Piano, near Tivoli (Rome, Italy).
Diagnosis: Characters as for genus.
Description: The specimen at hand
consists of a tall CD-like stem, articulated with a low basal circle and
moderately higher radial circle (Figs. 2.A ![]() ,
3
,
3 ![]() ,
4.A
,
4.A ![]() ).
Sutures between radial plates, between basal and radial plates and between
columnals are not prominent. The CD-like stem is relatively high and has five
sides. The five sides are not flat but slightly irregular due to the presence of
cirrus sockets. The CD-like stem consists of three columnals (Figs.
2.A
).
Sutures between radial plates, between basal and radial plates and between
columnals are not prominent. The CD-like stem is relatively high and has five
sides. The five sides are not flat but slightly irregular due to the presence of
cirrus sockets. The CD-like stem consists of three columnals (Figs.
2.A ![]() ,
3
,
3 ![]() - 4
- 4 ![]() ). The two proximal columnals are cirriferous (cirrinodals) whereas
the distal one is without any cirrus (nudinodal). The first proximal cirrinodal
is the widest columnal whereas the distal one is the smallest and lowest. The
distal columnal is the underside of the CD-like stem which is thin and
pentagonal in outline. Its distal surface is irregular, probably a little
weathered, and without any central canal, the latter meaning there was no other
articulated columnal (Fig. 4.B
). The two proximal columnals are cirriferous (cirrinodals) whereas
the distal one is without any cirrus (nudinodal). The first proximal cirrinodal
is the widest columnal whereas the distal one is the smallest and lowest. The
distal columnal is the underside of the CD-like stem which is thin and
pentagonal in outline. Its distal surface is irregular, probably a little
weathered, and without any central canal, the latter meaning there was no other
articulated columnal (Fig. 4.B ![]() ). Each cirrinodal is characterised by five
wide cirrus sockets that are arranged in 10 columns. Each socket is sub-circular,
slightly concave and with the edge slightly raised. They are lacking any
distinct articular feature. The largest cirri sockets lie in the proximal
cirrinodal. Between the radial circle and the CD-like stem lies the basal circle
consisting of five basal plates. The basal plates are only visible interradially.
The basal circle is probably star-shaped; the basal plates are hidden by the
radial circle and CD-like stem. Similarly, the upper and lower articular
surfaces of basals cannot be observed. The basal plates are relatively small,
their lower edges curved downwards and overhanging the top of the first columnal
(Fig. 4.C
). Each cirrinodal is characterised by five
wide cirrus sockets that are arranged in 10 columns. Each socket is sub-circular,
slightly concave and with the edge slightly raised. They are lacking any
distinct articular feature. The largest cirri sockets lie in the proximal
cirrinodal. Between the radial circle and the CD-like stem lies the basal circle
consisting of five basal plates. The basal plates are only visible interradially.
The basal circle is probably star-shaped; the basal plates are hidden by the
radial circle and CD-like stem. Similarly, the upper and lower articular
surfaces of basals cannot be observed. The basal plates are relatively small,
their lower edges curved downwards and overhanging the top of the first columnal
(Fig. 4.C ![]() ). The lower edges are not flat but moderately convex and sub-triangular
in cross-section. The radial circle, pentagonal in outline, is strongly
protruding, being significantly wider than the CD-like stem. It consists of five
relatively high radial plates, each with exposed free surface slightly lobed and
clearly turned downwards (Figs. 2.A
). The lower edges are not flat but moderately convex and sub-triangular
in cross-section. The radial circle, pentagonal in outline, is strongly
protruding, being significantly wider than the CD-like stem. It consists of five
relatively high radial plates, each with exposed free surface slightly lobed and
clearly turned downwards (Figs. 2.A ![]() ,
4.A
,
4.A ![]() ). The upper side of each radial is
occupied by an oblong radial facet (Fig.
5
). The upper side of each radial is
occupied by an oblong radial facet (Fig.
5 ![]() ). This facet is relatively wide,
neither steep, nor vertical, but gently tilted inwards. The aboral side is
smaller than the adoral one. The aboral ligament fossa is wide and strongly
inclined outwards (almost subvertical). The fossa is slightly concave and
quarter-moon-shaped, and is characterised by a wide and deep ligament pit. The
adoral side is moderately inclined inwards. The fulcral ridge is clearly evident
and long. Near the fulcral ridge is a wide and raised, elliptical axial canal.
The two interarticular ligament surfaces are wide, almost subvertical, slightly
concave and sub-triangular. The two muscle surfaces are somewhat smaller and not
subvertical. These fossae are sub-triangular in shape and ventrally bound by a
high and thick ridge. Each ridge is sub-triangular with the interradial side
significantly higher than the radial side. The ridges of two adjacent muscle
fossae compose a false interradial "process". The upper side of this false "process" is characterised by a distinct groove just where the suture,
between two adjacent radial plates, is presumed to be (Fig.
4.D
). This facet is relatively wide,
neither steep, nor vertical, but gently tilted inwards. The aboral side is
smaller than the adoral one. The aboral ligament fossa is wide and strongly
inclined outwards (almost subvertical). The fossa is slightly concave and
quarter-moon-shaped, and is characterised by a wide and deep ligament pit. The
adoral side is moderately inclined inwards. The fulcral ridge is clearly evident
and long. Near the fulcral ridge is a wide and raised, elliptical axial canal.
The two interarticular ligament surfaces are wide, almost subvertical, slightly
concave and sub-triangular. The two muscle surfaces are somewhat smaller and not
subvertical. These fossae are sub-triangular in shape and ventrally bound by a
high and thick ridge. Each ridge is sub-triangular with the interradial side
significantly higher than the radial side. The ridges of two adjacent muscle
fossae compose a false interradial "process". The upper side of this false "process" is characterised by a distinct groove just where the suture,
between two adjacent radial plates, is presumed to be (Fig.
4.D ![]() ). The
ventral cavity, sub-pentagonal in outline, is wide and sub-conical. The walls of
the ventral cavity, at least at the top, are smooth and steep. The interradial sutures
are not evident. Unfortunately, the lower part of the cavity is smeared with sediment. The
space just below the aboral ligament surface and between two basal plates
probably belongs to the corresponding radial plate.
). The
ventral cavity, sub-pentagonal in outline, is wide and sub-conical. The walls of
the ventral cavity, at least at the top, are smooth and steep. The interradial sutures
are not evident. Unfortunately, the lower part of the cavity is smeared with sediment. The
space just below the aboral ligament surface and between two basal plates
probably belongs to the corresponding radial plate.
|
Figure 2: Cup of Tiburtocrinus toarcensis gen. et sp. nov. (NS 6/950). A: Lateral view; B: Ventral view. Scale bar: 2 mm. Figure 2 :
Thèque de Tiburtocrinus
toarcensis gen. et sp. nov. (NS 6/950). A : Vue latérale ; B : Vue ventrale. Barre
d'échelle : 2 mm. |
|
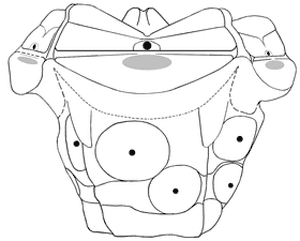
Figure 3: Reconstruction of cup of Tiburtocrinus toarcensis gen. et sp. nov. Figure 3 : Reconstitution de la thèque de Tiburtocrinus
toarcensis gen. et sp. nov. |
|
Figure 4: Tiburtocrinus toarcensis gen. et sp. nov. (NS 6/950). A: Slightly tilted lateral view of the cup; B: Detail of distal columnal (arrow). Note absence of central canal; C: Detail of basal plate (arrow) and cirrals (thin arrows); D: Detail of false interradial "process" and interradial groove (arrow). Scale bars: 2 mm. Figure 4 : Tiburtocrinus toarcensis
gen. et sp. nov. (NS 6/950). A : Vue latérale légèrement
basculée de la thèque ; B : Détail de columnale distale (flèche) ;
C : Détail de la plaque basale (flèche) et des cirrales (flèches
minces). Notez l'absence de canal central ; D : Détails du faux
processus interradial. Détail du sillon interradial (flèche). Barres d'échelle :
2 mm. |
Remarks: This specimen still bears
plates articulated between them among which CD-like stem, basal and radial
plates. In regard to the CD-like stem, the characters of cirrus sockets are
unclear and, in particular, the transverse ridge and the central pore are not
evident. In the case of the ventral cavity, partial occlusion by sediment hamper
its detailed description. Certainly, because the radial circle is not very high,
the ventral cavity is not very deep. Another element that cannot be described in
detail is the basal circle. Situated between the CD-like stem and the radial
circle, it is completely hidden. However, by analogy with those of other
paracomatulid species, it can be inferred that the basal circle is more or less
star-shaped. With respect to the radial facets, the interarticular ligament
surface and the muscular surface are not very evident. Nevertheless, they can be
deduced with some confidence as being planes of differing slope and because the
lateral edges of each facet displays a notch due to the differing lengths of the
interarticular ligament and the muscular fossae (Fig.
5 ![]() ).
).
At first sight the CD-like stem of Tiburtocrinus toarcensis gen. et sp. nov. is similar to that of Paracomatula liasica Hess, 2006. However, the CD-like stem of T. toarcensis gen. et sp. nov. has three columnals whereas the CD-like stem of P. liasica has only two. If the radial facets are also analysed, the differences become more obvious, due to the radial facets of T. toarcensis gen. et sp. nov. being rather smaller and more inclined (they are subvertical in P. liasica).
The specimen at hand bears sufficiently distinctive features to support the erection of a new species. T. toarcensis gen. et sp. nov. differs from all other species of paracomatulids in the presence of basal plates characterised by a lower edge curved downwards and because the articular radial facets are oblong and not subvertical. Furthermore, the CD-like stem is distinctive in consisting of three columnals. An important diagnostic element to classify paracomatulids is the number of columnals of the CD-like stem. Each species displays a well-defined number of columnals: Paracomatula triadica Hagdorn & Campbell, 1993, is characterised by six columnals; P. helvetica Hess, 1951, by five columnals; P. liasica and P. morator Hess, 2014, by only two columnals; and Paracomatula sp., found in Turkey, is characterised by seven columnals. The CD-like stem of the specimen here is characterised by only three columnals. There is no evidence of any axial canal on the most distal columnal and, consequently, it is reasonable to conclude that the CD-like stem of T. toarcensis gen. et sp. nov. consists only of three columnals.
Occurrence: Lower Toarcian of central Apennines (Italy).
|
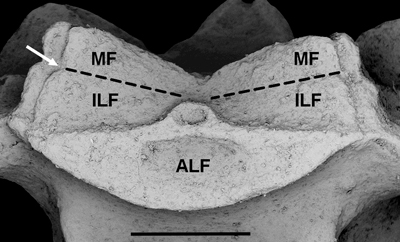
Figure 5: Tiburtocrinus toarcensis gen. et sp. nov. (NS 6/950). Detail of radial facet. MF: muscular fossa; ILF: interarticular ligament fossa; ALF: aboral ligament fossa; dotted line delimits muscular fossa; arrow shows notch between interarticular ligament fossae and muscle fossae. Scale bar: 1 mm. Figure 5 : Tiburtocrinus toarcensis
gen. et sp. nov.
(NS 6/950). Détails de la facette radiale et du faux processus interradial. MF : fosse musculaire ; ILF : fosse ligamentaire
interarticulaire ; ALF : fosse ligamentaire aborale ; la ligne
pointillée délimite la fosse musculaire ; la flèche montre l'encoche
entre la fosse du ligament interarticulaire et la fosse du muscle. Barre d'échelle : 1 mm. |
Origin: Hess (2014) thought that the paracomatulids derived from the pentacrinitids due to similarity in their anatomical characteristics. On the other hand, Hagdorn and Campbell (1993, p. 13) suggested that Paracomatula derived from the holocrinids because the paracomatulids retain "long cylindrical cirrals and the tall brachials" and "symplectial lower nodal articula". The Holocrinida Jaekel, 1918, are the earliest Articulata crinoids because they are reported from the Lower Triassic. The Holocrinida Jaekel feature a thick-walled cup, consisting of low infrabasal plates, basal plates generally higher than the radial plates, a narrow ventral cavity, radial facets with small muscular fossae, uniserial arms, columns more or less pentagonal or circular, and low columnals with symplectial articular surfaces with petaloid pattern, and cirrinodals with two or five cirrus sockets.
The hypothesis of Hess is to be preferred. The downward curvature of the lower edges of the basal plates in T. toarcensis
gen. et sp. nov. (Fig. 4.C ![]() ) supports the hypothesis that paracomatulids
derived from pentacrinitids. Indeed, pentacrinitids have the lower edge of their
basal plates directed downward and covering the proximal columnals. In contrast,
holocrinids have basal plates lacking downward curvature of the lower edges.
Additionally, the basal and radial plates of holocrinids are much taller than
those of paracomatulids.
) supports the hypothesis that paracomatulids
derived from pentacrinitids. Indeed, pentacrinitids have the lower edge of their
basal plates directed downward and covering the proximal columnals. In contrast,
holocrinids have basal plates lacking downward curvature of the lower edges.
Additionally, the basal and radial plates of holocrinids are much taller than
those of paracomatulids.
Ancestors: This new species has basal plates very similar to those of pentacrinitids, and radial facets not exactly of comatulid type (the comatulids have different types of radial facets, one of the most common types being subvertical, with a very wide aboral ligament fossa and wide muscular fossae). The basal plates, as already mentioned, have downward curvature of the lower edges, a feature not present in Paracomatula species but particularly evident in Pentacrinites Blumenbach, 1804, and Seirocrinus Gislén, 1924. All the Paracomatula species have their basal plates with tips well-exposed interradially. The radial facets of T. toarcensis gen. et sp. nov. are outward-sloping and with proportionally smaller muscular fossae; they also differ from those of the pentacrinitids in being narrower and having articular elements markedly different in shape and size. P. triadica, the stratigraphically oldest species of paracomatulids, is an ambiguous species: On the one hand, it shows primitive characters (a CD-like stem with several columnals), and on the other, it has more derived characters (the large, subvertical facets of the comatulids here are considered an apomorphic character, as it appears for the first time in P. triadica). Unfortunately, the basal plates are not well-exposed (Hagdorn & Campbell, 1993) and therefore nothing can be said about their shape and characters.
P. liasica and P. helvetica have comatulid-type radial facets, and basal plates with tips clearly evident interradially. So, Paracomatula triadica, P. liasica and P. helvetica have more or less similar cup characters. All this suggests that T. toarcensis gen. et sp. nov. has more primitive characters (type of radial facets, type of basal plates) than any species of Paracomatula. It is therefore reasonable to infer that T. toarcensis gen. et sp. nov. may belong to an archaic lineage that likely links the pentacrinitids to Paracomatula.
Evolutionary trends: Unfortunately, apart from P. triadica and P. helvetica (whole specimens of which are known), P. liasica, P. morator and T. toarcensis gen. et sp. nov. are still poorly known. In fact, only their CD-like stem is known, and in some cases their basal circle and radial circle. Nevertheless, the two documented evolutionary trends of paracomatulids are the marked reduction in the number of columnals from six (P. triadica) to two (P. morator), and the significant increase in the length of the arms (Hagdorn & Campbell, 1993, p. 13). Finally, another trend could be the disappearance of downward curvature of the lower edge of the basal plates in the paracomatulid species. Such a disappearance, as well as the reduction in number of the columnals of the CD-like stem, would have had a functional advantage: The weight of the cup would have been reduced with a consequent energy advantage, i.e., less weight meaning less energy consumption during movement. As far as arm length is concerned, it must be admitted that arm extension also has two significant functional advantages: Enhancement of displacement capacity [swimming comatulids have a longer arm length than crawling ones (Janevski & Baumiller, 2010)] and increase in filtering capacity.
Paracomatulids, swimming and crawling crinoids: One of the most prominent differences between T. toarcensis gen. et sp. nov. and the known species of Paracomatula concerns the size and orientation/positioning of the radial articular facets. Muscular facets are important because they permit the movement of the arms. Large muscular fossae indicate large muscles and therefore a better capability to move; on the other hand, smaller fossae indicate more limited mobility. Janevski and Baumiller (2010: p. 595), comparing the respective radial facets of the antedonid Florometra asperrima (A.H. Clark, 1907) and the comasterid Davidaster discoideus (Carpenter, 1888), noted that the swimming antedonid has "an enlargement of the area of the muscular fossae relative to the overall radial articular facet, but the crawling comasterid possesses comparatively smaller muscular fossae". F. asperrima has the typical subvertical articular facet of the comatulids, with a wide aboral ligament fossa, and wide muscular fossae; D. discoideus, meanwhile, has a narrower, oblong articular facet, is not subvertical and has smaller muscular fossae.
With regard to the inclination of the radial facets, it
is difficult to assess how much the functionality of the arms varies with the
inclination of the radial facets. In any case, if the radial facets are very
steep, i.e., they are positioned vertically, the proximal part of the
arms is positioned sub-horizontally; conversely, if the radial facets are
slightly inclined, the proximal part of the arms is slightly inclined upwards.
This should lead to a greater upward thrust in the first case, thus resulting in
higher performance than in the second case. This would be supported by the fact
that the swimming Florometra serratissima (A.H. Clark,
1907) can
swim using only the proximal part of the arms (Janevski & Baumiller,
2010: p. 590). Paracomatula triadica, P. liasica
and P. helvetica have radial facets similar to those of the
majority of comatulids. In fact, they have large, steep (almost subvertical)
radial facets, with a very wide aboral ligament fossa and wide muscular fossae. T. toarcensis
gen. et sp. nov., on the other hand, has smaller radial facets moderately
sloping outward with narrower aboral ligament fossa and muscular fossae;
otherwise, its articular facet does not differ much from that of D. discoideus
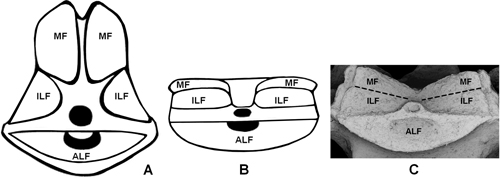
(Fig. 6 ![]() ). So, by analogy with D. discoideus, it can
be concluded that, due to a reduced musculature and the differing slope of the
radial facets, T. toarcensis gen. et sp. nov. may have
had reduced mobility of the arms and consequently it was perhaps able to crawl.
In contrast, the species of Paracomatula, displaying the opposite
condition, may have been more mobile and probably able to swim. It is important
to emphasise that this hypothesis is based only on the morphological similarity
of the radial facets of T. toarcensis gen. et sp. nov. to those
of D. discoideus. In fact, because the length of the arms, the shape
of the brachials, and the number of muscles in each arm of T. toarcensis
gen. et sp. nov. are unknown, nothing can be said about the real locomotion
capacity of this new species.
). So, by analogy with D. discoideus, it can
be concluded that, due to a reduced musculature and the differing slope of the
radial facets, T. toarcensis gen. et sp. nov. may have
had reduced mobility of the arms and consequently it was perhaps able to crawl.
In contrast, the species of Paracomatula, displaying the opposite
condition, may have been more mobile and probably able to swim. It is important
to emphasise that this hypothesis is based only on the morphological similarity
of the radial facets of T. toarcensis gen. et sp. nov. to those
of D. discoideus. In fact, because the length of the arms, the shape
of the brachials, and the number of muscles in each arm of T. toarcensis
gen. et sp. nov. are unknown, nothing can be said about the real locomotion
capacity of this new species.
|
Figure 6: Radial facets of comatulid crinoids and Tiburtocrinus toarcensis gen. et sp. nov. A: Florometra asperrima, swimming crinoid; B: Davidaster discoides, crawling crinoid; C: Tiburtocrinus toarcensis gen. et sp. nov. (NS 6/950), possible crawling crinoid. Images A and B modified from Janevski and Baumiller (2010). Figure 6 : Facettes radiales de crinoïdes comatulides et de
Tiburtocrinus toarcensis gen. et sp. nov. A : Florometra
asperrima, crinoïde natatoire; B : Davidaster discoides, crinoïde
rampant; C : Tiburtocrinus toarcensis gen. et sp. nov. (NS 6/950), un
crinoïde rampant possible. Images A et B modifiées d'après Janevski
and Baumiller (2010). |
Ecology: Understanding how these crinoids lived requires comparing them with living comatulids. The comatulids are free crinoids because they are stemless. Not being fixed, they can move over short distances; this way, they hide in rocky crevices during the day and crawl on coral reefs and/or climb on corals for nourishment at night. Living comatulids feed mainly at night so as not to encounter their predators (Rutman & Fishelson, 1969; Meyer et al., 1984). They are also able to swim very slowly over short distances. This freedom of movement allows comatulids both to choose the best habitat and to escape from predators (Meyer & Macurda, 1977; Baumiller, 2008; Baumiller et al., 2008, 2010; Gorzelak & Salamon, 2009).
Like comatulids, the paracomatulids lack an encrusting disc or a root. They attached themselves either to the rocky surface or to some obstacle of biological origin, such as shells, corals, etc., with the cirri of their short, CD-like stem filtering water. Note that the cirri of P. helvetica were equipped with a terminal claw, which was very useful in anchoring the individual on a hard surface (Hess, 1999). By detaching the cirri from the substrate, they could move either to hide from predators or to look for new places favourable for nutrient supply.
Palaeogeography: Until now, no paracomatulid has ever been found in Italy. This new finding is of great importance because it widens the geographical distribution of this type of crinoid. Nevertheless paracomatulids are very rare crinoids. Four species of Paracomatula are currently known: P. triadica from the Otapirian [the Otapirian belongs to the Norian, though today it is considered Rhaetian (Raine et al., 2015)] (Late Triassic) of Ile Ducos (New Caledonia), P. liasica from the Pliensbachian of Arzo (Switzerland), P. helvetica from the Bajocian of Hottwil (Switzerland), and P. morator Hess, 2014, from the Oxfordian of Andelot-Morval (France); and two species of Singillatimetra are known: S. truncata Hess & Thuy, 2016, from the Toarcian of Feuguerolles (France) and S. inordinata Hess, 2012, from the lower Bathonian of La Pouza (France) (only isolated CDs are known). In addition, Paracomatula sp. from the upper Sinemurian-lower Pliensbachian of Günören and Iznik (Turkey) was described by Nicosia (1991) (only isolated CDs are known). Finally, Simms (1989) dubiously assigned some brachials from the Toarcian of Ilminster (Somerset, England) to the paracomatulids. Apart from P. triadica found in the Triassic sediments of New Caledonia, that is, in a very different and distant palaeogeographic context, paracomatulids are mainly found in strata of the western Neo-Tethys Ocean. The findings are all situated between N18° and N40° palaeolatitude. The southernmost species is T. toarcensis gen. et sp. nov., the northern- and easternmost species is Paracomatula sp. from Turkey; and the westernmost species is P. morator. Certainly, new discoveries are necessary to understand, on the one hand, whether New Caledonia was in fact the area of radiation of these crinoids and, on the other, whether the western sector of the Neo-Tethys Ocean provided the paracomatulids with the optimal conditions to develop.
The pentacrinitids tested different morphologies to identify which one was most adaptive to new environments originating from the opening of the Neo-Tethys Ocean. In this context appeared Paracomatula, Singillatimetra, and Tiburtocrinus gen. nov., all genera with a CD-like stem not attached to the substrate and therefore free to move. Locomotion was a winning adaptation because it allowed these crinoids not only to move into a better environment to inhabit, but also to hide from predators.
The lineage of the paracomatulids also developed two different displacement strategies. Some paracomatulids may have been able to crawl (Tiburtocrinus gen. nov.) and others (Paracomatula) were able to swim (nothing can be said about Singillatimetra because only columnals are known). These two strategies are markedly different: One is linked to the bottom and the other not. These different strategies are also found today in the comatulids. From a logical point of view, and therefore also from an evolutionary one, it is probable that the strategy of crawling appeared first. If this is true, the paracomatulids must have differentiated from the pentacrinitids before the Late Triassic (occurrence of P. triadica). The ability to move was so much the key to the success of the paracomatulids that they successfully ranged from the Rhaetian to the Oxfordian, surviving several biological crises, including the Triassic/Jurassic extinction. This freedom of movement allowed these crinoids to quickly occupy new niches resulting from the environmental instability of the Late Triassic and Early Jurassic. On the other hand, many crinoids with a stem, including holocrinids and several isocrinids, became extinct during that interval, perhaps because, not being able to move, they could not colonise the new habitats that were forming. Certainly, comatulids and paracomatulids entered into competition, even living together in the same environments (Hess & Thuy, 2016). But the paracomatulids probably became extinct in the Oxfordian, whereas the comatulids are still present and now represent well over 80% of modern species of crinoids. This means that the comatulids have been able to adapt better and perhaps faster to environmental stimuli from the Jurassic to the present.
We wish to thank Maria Vinci of "Monte Catillo Natural Reserve"; Bruno Granier and the reviewers, Mariusz A. Salamon and Andrew S. Gale, for their constructive comments on a preliminary version of the manuscript. In addition, thanks are due to Umberto Nicosia for useful discussions and to Marco Albano (IGAG) for the SEM photographs.
Baumiller T.K. (2008).- Crinoid ecological morphology.- Annual Review of Earth and Planetary Sciences, Palo Alto - CA, vol. 36, p. 221-249.
Baumiller T.K., Mooi R. & Messing C.G. (2008).- Urchins in the meadow: Paleobiological and evolutionary implications of cidaroid predation on crinoids.- Paleobiology, New York, vol. 34, p. 35-47.
Baumiller T.K., Salamon M.A., Gorzelak P., Mooi R., Messing C.G. & Gahn F. (2010).- Post-Paleozoic crinoid radiation in response to benthic predation preceded the Mesozoic marine revolution.- Proceedings of the National Academy of Sciences of the United States of America, Los Angeles, vol. 107, no. 3), p. 5893-5896.
Blumenbach J.F. (1802-1804).- Abbildungen naturhistorischer Gegenstände, pt. 7, no. 70.- Göttingen, 4 p.
Carpenter P.H. (1888).- Report on the Crinoidea collected during the voyage of H.M.S. Challenger, during the years 1873-76. In: Part II. The Comatulae.- Reports of the Scientific Results of the Voyage of H.M.S. Challenger, Zoology, vol. 26 (part 60), 402 p.
Centamore E., Chiocchini M., Deiana G., Micarelli A. & Pieruccini U. (1971).- Contributo alla conoscenza del Giurassico dell'Appennino Umbro-marchigiano.- Studi Geologici Camerti, Camerino, vol. 1, p. 7-89.
Clark A.H. (1907).- Description of new species of recent unstalked crinoids from the North Pacific Ocean.- Proceedings of the United States National Museum, Washington - D.C., vol. 33 (1559), p. 69-84.
Clark A.H. (1908).- New genera of unstalked crinoids.- Biological Society of Washington, Proceedings, Washington - D.C., vol. XXI, p. 125-136.
Fabbi S. & Santantonio M. (2012).- Footwall progradation in syn-rift carbonate platform-slope systems (Early Jurassic, northern Apennines, Italy).- Sedimentary Geology, vol. 281, p. 21-34.
Farinacci A., Mariotti N., Nicosia U., Pallini G. & Schiavinotto F. (1981).- Jurassic sediments in the umbro-marchean Apennines: An alternative model. In: Farinacci A. & Elmi S. (eds.), Rosso Ammonitico Symposium Proceedings.- Tecnoscienza, Rome, p. 335-398.
Gislén T. (1924).- Echinoderm studies.- Zoologisk Bidrag från Uppsala, Uppsala, vol. 9, 330 p.
Gray J.E. (1842).- Synopsis of the contents of the British Museum.- British Museum, London, 308 p.
Gorzelak P. & Salamon M.A. (2009).- Signs of benthic predation on Late Jurassic stalked crinoids, preliminary data.- Palaios, Tulsa, vol. 24, p. 70-73.
Hagdorn H. & Campbell H.J. (1993).- Paracomatula triadica sp. nov.: An early comatulid crinoid from the Otapirian (Late Triassic) of New Caledonia.- Alcheringa, Abingdon, vol. 17, p. 1-17.
Hess H. (1951).- Ein neuer Crinoide aus dem mittleren Dogger der Nordschweiz (Paracomatula helvetica n. gen. n. sp.).- Eclogae geologicae Helvetiae, Basel, vol. 43, p. 208-216.
Hess H. (1999).- Middle Jurassic of northern Switzerland. In: Hess H., Ausich W.I., Brett C.E. & Simms M.J. (eds.), Fossil Crinoids.- Cambridge University Press, Cambridge, p. 203-215.
Hess H. (2006).- Crinoids from the Lower Jurassic (Pliensbachian) of Arzo, southern Switzerland.- Schweizerische paläontologische Abhandlungen, Basel, vol. 126, 143 p.
Hess H. (2012).- Crinoids from the Middle Jurassic (Bajocian-Lower Callovian) of Ardèche, France.- Swiss Journal of Palaeontology, vol. 131, p. 211-253.
Hess H. (2014).- Origin and radiation of the comatulids (Crinoidea) in the Jurassic.- Swiss Journal of Palaeontology, vol. 133, p. 23-34.
Hess H. & Messing C.G. (2011).- Comatulida. In: Selden P.A. (ed.), Part T, Echinodermata 2, Revised, Crinoidea, vol. 3.- Treatise on Invertebrate Paleontology, The Geological Society of America, Boulder - CO, and The University of Kansas, Lawrence - KS, p. 70-159.
Hess H. & Thuy B. (2016).- Extraordinary diversity of feather stars (Echinodermata: Crinoidea: Comatulida) from a Lower Jurassic (Pliensbachian-Toarcian) rock reef of Feuguerolles (Normandy, France).- Swiss Journal of Palaeontology, vol. 136, p. 301-321.
Jaekel O. (1918).- Phylogenie und System der Pelmatozoen.- Paläontologische Zeitschrift, Berlin, vol. 3, p. 1-128.
Janevski G.A. & Baumiller T.K. (2010).- Could a stalked crinoid swim? A biomechanical model and characteristics of swimming crinoids.- Palaios, Lawrence - KS, vol. 25, no. 9/10, p. 588-596.
Manni R. & Nicosia U. (1990).- New crinoids from Liassic sediments nearby Tivoli (central Italy); rearrangement in the systematics of eudesicrinids and cotyledermatids.- Geologica Romana, vol. 26, p. 81-92.
Manni R. & Nicosia U. (1994).- Crinoidi giurassici dell'Italia centrale.- Studi Geologici Camerti, Camerino, vol. spec. 1994, p. 299-323.
Meyer D.L., La Haye C.A., Holland N.D., Arenson A.C. & Strickler J.R. (1984).- Time-lapse cinematography of feather stars (Echinodermata: Crinoidea) on the Great Barrier Reef, Australia: Demonstrations of posture changes, locomotion, spawning and possible predation by fish.- Marine Biology, vol. 78, p. 179-84.
Meyer D.L. & Macurda D.B. (1977).- Adaptive radiation of comatulid crinoids.- Paleobiology, New York, vol. 3, p. 74-82.
Nakano H., Hibino T, Hara,Y., Oji T. & Amemiya S. (2004).- Regrowth of the stalk of the sea lily, Metacrinus rotundus (Echinodermata: Crinoidea).- Journal of Experimental Zoology, vol. 301A, p. 464-471.
Nicosia U. (1991).- Mesozoic crinoids from the north-western Turkey.- Geologica Romana, vol. 27, p. 389-436.
Raine J.I., Beu A.G., Boyes A.F., Campbell H.J., Cooper R.A., Crampton J.S., Crundwell M.P., Hollis C.J. & Morgans H.E.G. (2015).- Revised calibration of the New Zealand geological timescale: NZGT2015/1.- GNS Science Report, vol. 2012/39. 53 p.
Rasmussen H.W. (1978).- Comatulida. In: Moore R.C. & Teichert C. (eds.), Part T, Echinodermata 2, vol. 1.- Treatise on Invertebrate Paleontology, The Geological Society of America, Boulder - CO, and The University of Kansas, Lawrence - KS, p. T867-T916.
Rutman J. & Fishelson L. (1969).- Food composition and feeding behaviour of shallow-water crinoids at Eilat (Red Sea).- Marine Biology, vol. 3, p. 981-994.
Santantonio M. (1994).- Pelagic carbonate platforms in the geological record: Their classification, and sedimentary and paleotectonic evolution.- American Association Petroleum Geologists Bulletin, Tulsa, vol. 78, p. 122-141.
Sieverts-Doreck H. (1952).- Classification. In: Moore R.C., Lalicker C.G. & Fischer A.G., Invertebrate fossils, Crinoids, chapter 18.- Mc Graw-Hill Book Company, New York, Toronto, London, p. 614.
Simms M.J. (1988).- The phylogeny of post-Paleozoic crinoids. In: Paul C.R.C. & Smith A.B. (eds.), Echinoderm phylogeny and evolutionary biology.- Oxford University Press, vol. 21, p. 269-284.
Simms M.J. (1989).- British Lower Jurassic crinoids.- Monograph of the Palaeontographical Society, London, Publication 581, part of vol. 142, p. 1-103.
Venturi F. (2006).- Brachiopodi, ammoniti ed altri fossili nel Toarciano basale dei Monti Lucretili (Lazio orientale). Bio-cronostratigrafia e paleoambienti.- La FESPEM informa, Bollettino di Mineralogia e Paleontologia, Aqui Terme, vol. 2006, p. 10-16.
Vörös A. (2002).- Early Jurassic koninckinids (Athyridida, Brachiopoda) from Tivoli near Rome, Italy.- Geologica Romana, vol. 36, p. 169-197.
| Nomenclatural note: Life Sciences Identifier (LSID) http://zoobank.org/References/A114F0DE-03BE-424B-AFBB-C7DBEE4986CA Genus Group Species Group |