◄ Carnets Geol. 22 (15) ►
![]()
Outline:
[1. Introduction] [2. Geographic and geological setting]
[3. Paleontological study] [4. Discussion] [5. Conclusion] and ...
[Bibliographic references]
Laboratoire de Paléontologie, UFR Sciences & Techniques, Université de Bretagne Occidentale, 6 avenue Le Gorgeu, CS 93837, 29938 Brest (France)
Wezep (The Netherlands)
Harderwijk (The Netherlands)
Published online in final form (pdf) on September 14, 2022
DOI 10.2110/carnets.2022.2215
![]()
[Editor: Brian R. Pratt; technical editor: Bruno R.C. Granier]
![]()
A species of Proporidae (Heliolitida) forming very small, discoid coralla, is fully described. It is mainly preserved in natural moulds in the Öjlemyr chert, an erratic silicified limestone found in Eridanos deposits surfacing in the west and north-east of the Dutch Overijssel-province and adjoining German area, generally accepted to be Uppermost Katian. The material is assigned to the rehabilitated Pinacopora Nicholson & Etheridge, 1878, a genus closely allied to Propora Milne-Edwards & Haime, 1849, and a new species, Pinacopora baltica is named. The main feature of this species, besides the characteristics of the genus and the biometric data, is the presence of wide, short, and convex septal ridges bearing scattered septal spines, alternating with narrow interseptal furrows. For the first time in heliolitids a pleurodictyoid pattern of growth is described. Only two species of Pinacopora are recorded: P. baltica n. sp. (Upper Ordovician) and P. grayi Nicholson & Etheridge, 1878 (Lower Silurian), respectively from Baltica and Laurentia.
• Heliolitida;
• Proporidae;
•
Pinacopora;
• Öjlemyr chert;
• Upper Ordovician;
• Baltica
Plusquellec Y., Eyzenga J. & Keulen P.S.F. van (2022).- A new discoid heliolitid (Proporidae) from the Upper Ordovician of the Baltic area: Morphology and modalities of increase.- Carnets Geol., Madrid, vol. 22, no. 15, p. 685-698.
Un heliolitide discoďde Proporidae nouveau de l'Ordovicien supérieur de la région baltique : Morphologie et modalités de croissance.- Une trčs petite espčce de Proporidae discoďde (Heliolitida), principalement conservée ŕ l'état de moulage naturel dans le chert d'Öjlemyr, un calcaire silicifié de l'Ordovicien supérieur (Katien terminal), connu sous formes de blocs erratiques dans les dépôts du paléofleuve Eridanos, dans l'Ouest et le Nord-Est de la province hollandaise de l'Overijssel et des deux côtés de la frontičre allemande est décrite en détail. Le matériel est attribué au genre Pinacopora Nicholson & Etheridge, 1878, réhabilité ici, proche de Propora Milne-Edwards & Haime, 1849, et une nouvelle espčce, P. baltica, est proposée. L'espčce, en plus des caractčres propres au genre et des données biométriques, se caractérise principalement par des rides septales larges, faiblement proéminantes, portant de petites épines disposées sans ordre et alternant avec des sillons interseptaux étroits et lisses. Pour la premičre fois chez les heliolitides, un mode de développement de la colonie de type pleurodictyoďde est décrit. Seulement deux espčces de Pinacopora sont connues, P. baltica n. sp. (Ordovicien supérieur) et P. grayi Nicholson & Etheridge, 1878 (Silurien inférieur), respectivement dans Baltica et Laurentia.
• Heliolitida ;
• Proporidae ;
•
Pinacopora ;
• chert d'Öjlemyr ;
• Ordovicien supérieur ;
• Baltica
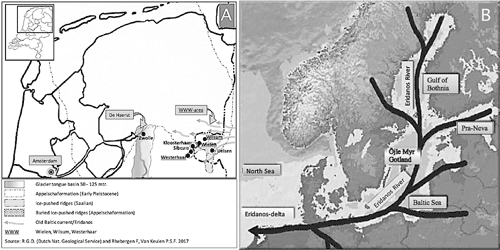
The study of thousands of Upper Ordovician erratic silicified limestone from both sides of the German border (the so-called WWW triangle, named after the villages Wilsum, Wielen and Westerhaar) and De Haerst near Zwolle in the Netherlands, has provided a rich fauna of brachiopods, crinoids, trilobites, ostracods, cornulitids, and other groups (Schallreuter, 1987a, 1987b; Keulen & Rhebergen, 2017; Vinn & Eyzenga, 2021). Among material from this source, a very small discoid heliolitid – not previously described – has drawn our attention and its description, systematic affinities and modalities of increase is the aim of this paper.
The Öjlemyrflint [henceforth: Öjlemyr chert] is a silicified carbonate, only known as erratic boulders, and containing an uppermost Katian fauna. The maximum diameter of the clasts is 17 cm. Many boulders, however, are much smaller. Three types of Öjlemyr chert have been recognized:
Öjlemyr chert of the Wielen type (after the village of Wielen; Schallreuter,
1984, p. 9; Eiserhardt,
1992, p. 3-4). This is only found in the North-Eastern Netherlands and across the German border, mainly in the so-called WWW-area, and in De Haerst (Fig. 1.A
![]() );
);
Öjlemyr chert from the isle of Sylt (Braderuper type);
Öjlemyr chert from Gotland. This is the original Öjlemyrflint, named after the peatswamp Öjle Myr (Wiman, 1901). Unlike the other types, this is a glacial (Weichselian) erratic.
|
|
Figure 1: A, northern part of the Netherlands with locations of De Haerst and WWW-area. Arrows indicate the main directions of the current. 2M years ago the coastline ran almost parallel to the Dutch-German border. B, drainage system of the Eridanos. |
These types share many faunal characteristics, and the distinction between them is mainly based on petrological grounds. Nevertheless, they do show significant differences in faunal composition. For instance the heliolitid described herein seems to have never been reported from Öjlemyr chert from Sylt and Gotland and, as yet, it is known only from Öjlemyr chert of the Wielen type. Lindström (1899, p. 99) did, however, indicate that Wiman had sent him small complete "nummuloid disks of a thickness of 3 mm" with "calicles deeper than usual" collected in the morainic accumulations of Öjle myr, Gotland. This material - not illustrated - clearly shares some features with our specimens. In addition, Lindström (1899, Pl. XI, figs. 16-17) illustrated a specimen of Proheliolites from Öjlemyr (Gotland), but this form is quite different from our material.
The fine-grained Öjlemyr chert of the Wielen type is often partially, or even completely leached out. Various pieces are concretion-like, carrying a light-colored, leached-out crust. The fossil assemblage is generally accepted to be uppermost Katian, and stratigraphically it should be situated close below the boundary between the Estonian Pirgu (F1c) and Porkuni (F2) regional stages (Schmidt, 1881; Eiserhardt, 1992; see also Rhebergen, 2009, Fig. 3).
The fossil fauna of Öjlemyr chert of the Wielen type is rich and varied. It is dominated by bryozoans (e.g., Pachydichtya borkholmensis, Sceptropora facula, Ptilodichtya sp.), ostracods, and brachiopods (Resserella pirguensis, Dalmanella sp., strophomenids). Trilobites include Erratencrinurus kiaeri, Pharostoma sp., Ascetopeltis bockeli, Harpidella sp., Sphaerexochus sp., Valdariops sp.). A new cornulitid (Cornulites spinosus) has been recently described (Vinn & Eyzenga, 2021). Quite remarkable fossils include rostroconchs (Bitrigonocardia lindstroemi) and algae (Palaeoporella variabilis, Apidium cf. sororis), which often have preserved organic carbon.
Since the assemblage shows a clear affinity with that of the Estonian Upper Ordovician, without being identical, it is believed that Öjlemyr chert originates from Ordovician formations close to Estonia (probably west of Estonia, in an area now covered by the Baltic Sea). These formations have never been found, and they may have disappeared altogether due to glacial erosion.
In the Netherlands, Öjlemyr cherts of the Wielen type are mainly found in gravel deposits of the Eridanos river-system (see Fig. 1.B
![]() ) (Appelscha Formation), which date from the Early-Middle Pleistocene (Menapian, ca. 1 MA). These deposits are now exposed in the Weichselian ice-pushed ridges of the WWW-area and in De Haerst. Little is known about the course the Öjlemyr cherts took from their place of origin in the Baltic all the way to the WWW-area. Since many of the boulders are big and angular, it is generally assumed that they have been transported fluvially, embedded in ice floes of the Eridanos. Upriver, these boulders were frozen in anchor ice which formed at the river bottom. In period of thaw, the anchor ice floated downstream in the shape of ice floes (Hucke,
1928). In this way, boulders may have been transported repeatedly during the cold periods which marked the Late Pliocene and Early Pleistocene. One million years ago, the coastline of the North sea lay along the current Dutch-German border. Apparently, where one arm of the Eridanos discharged into sea (near the WWW-area; see Fig. 1.B
) (Appelscha Formation), which date from the Early-Middle Pleistocene (Menapian, ca. 1 MA). These deposits are now exposed in the Weichselian ice-pushed ridges of the WWW-area and in De Haerst. Little is known about the course the Öjlemyr cherts took from their place of origin in the Baltic all the way to the WWW-area. Since many of the boulders are big and angular, it is generally assumed that they have been transported fluvially, embedded in ice floes of the Eridanos. Upriver, these boulders were frozen in anchor ice which formed at the river bottom. In period of thaw, the anchor ice floated downstream in the shape of ice floes (Hucke,
1928). In this way, boulders may have been transported repeatedly during the cold periods which marked the Late Pliocene and Early Pleistocene. One million years ago, the coastline of the North sea lay along the current Dutch-German border. Apparently, where one arm of the Eridanos discharged into sea (near the WWW-area; see Fig. 1.B
![]() , the ice floes melted and the boulders fell out.
, the ice floes melted and the boulders fell out.
In the first stages of this study, the available material was preserved only in natural moulds. The structure of the skeleton thus remained unknown and could have been a thick wall, a coenenchyme, or another common skeletal tissue. However, one incomplete silicified colony discovered while this work was in progress has provided the missing data. Although the calices are rather large (see Fig. 9
![]() ), the specimen undoubtedly belongs to the same species as the material preserved as natural moulds, evidenced by its discoid morphology, the presence of deep calices (some of them with complete tabulae bearing numerous small spines) and, overall, 12 rather flat and broad septal ridges with scattered small spines alternating with narrow interseptal furrows. In addition, a vertical split shows the presence of a narrow coenenchyme of diaphragms and dissepiments and allows the assignment to the Heliolitida.
), the specimen undoubtedly belongs to the same species as the material preserved as natural moulds, evidenced by its discoid morphology, the presence of deep calices (some of them with complete tabulae bearing numerous small spines) and, overall, 12 rather flat and broad septal ridges with scattered small spines alternating with narrow interseptal furrows. In addition, a vertical split shows the presence of a narrow coenenchyme of diaphragms and dissepiments and allows the assignment to the Heliolitida.
Subclass Tabulata Milne-Edwards & Haime, 1850
Order Heliolitida Frech, 1897
Family Proporidae Sokolov, 1949
Genus Pinacopora Nicholson & Etheridge, 1878
Pinacopora baltica n. sp.
(Figs. 2
![]() - 3
- 3
![]() - 4
- 4
![]() - 5
- 5
![]() - 6
- 6
![]() - 7
- 7
![]() - 8
- 8
![]() - 9
- 9
![]() )
)
Holotypus: specimen RGM 1 333 571
Locus typicus: Erratics at the WWW-area, Wilsum, Germany.
Stratum typicum/age: Öjlemyr flint of the Wielen type, Upper Ordovician, uppermost Katian.
Diagnosis. Very small discoid "epithecate" corallum, proximal side flat to slightly convex, more rarely concave, outline of corallum weakly polygonal in early stages, distal side convex, calicinal aperture lined by a narrow rim, calices of corallites (tabularia) deep, slightly conical, more rarely truncated cone-shaped and with flat calicinal spiny bases, corallite wall furnished with 12, wide and rather flat, very short septal ridges bearing scattered septal spines more or less in two or three rows, interseptal furrows narrow and smooth, tabulae absent in younger specimens, scarce in older one, and covered by numerous small spines. Narrow coenenchyme of scarce diaphragms and dissepiments. Increase of corallites epithecal, arrangement of corallites belonging to the pleurodictyoid category. Corallum diameter mainly between 6.0 and 10 mm, height about a quarter of the diameter, usually 8 to 20 corallites, diameter of calices mainly 1.3-1.8 mm, width of coenenchymal platform generally not exceeding 0.5 mm.
Material. The studied material consists of one specimen with a silicified skeleton, and a set of 10 specimens preserved in natural moulds, for most of them the two sides of the moulds (i.e., proximal [lower] and distal [upper] surfaces of the coral) have been collected and thus, nearly complete morphological features of the coralla are available. Latex casts of both sides have been taken - giving rise to restored specimens - and the material otherwise stated is described as such.
The specimens collected by two of us, J. Eyzenga and P.S.F. van Keulen, are deposited in the collections of the Naturalis Biodiversity Center Leiden (Netherlands), catalog number RGM 1 333 568-578. These are from sand pits at De Hearst (specimen RGM 1 333 568, RGM 1 333 577 and RGM 1 333 578, coll. J. Eyzenga) and from the WWW-triangle (specimens RGM 1 333 569-RGM 1 333 576, coll. P.S.F. van Keulen).
Proximal side. The proximal side could be more or less irregular, roughly flat, but is generally slightly convex (concave in natural mould) or more rarely concave such as in the largest known specimen. The central area is occupied by a small piece of bioclast which is surrounded by the so-called epitheca showing tiny concentric growth lines and some coarser ones (Figs. 2
![]() , 3.C, .G
, 3.C, .G
![]() ).
).
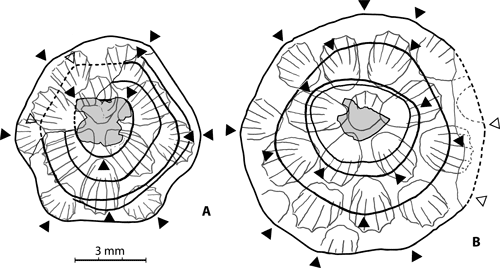
Outline of the corallum. It shows an unusual feature for discoid coralla. Although roughly circular, it exhibits - especially in the smallest colonies - a clear trend to be polygonal (rounded polygonal) instead of being lobate (see for example
Paracleistopora granulosa [Plusquellec, 2007, Pl. 5, fig. 8], and
Vaughaniopsis lafusti [Plusquellec, 2007, Pl. 6, fig. 8]). This is particularly obvious in the specimen illustrated Figure 3.A
![]() and in the earliest stage of another one (Fig. 2.B
and in the earliest stage of another one (Fig. 2.B
![]() ).
).
On the proximal side of a corallum, the main growth lines indicate successive
"stages" corresponding to different outlines of the corallum. The specimen illustrated in Figure 2.B
![]() shows an early 3-sided stage followed by a 5 and then 6-sided one. Another specimen (Fig. 2.B
shows an early 3-sided stage followed by a 5 and then 6-sided one. Another specimen (Fig. 2.B
![]() ) exhibits successively a 4 (trapezoidal), 5 (pentagonal) and more than 6 (weakly polygonal) sequence. In the central areas of coralla the spacing between two main growth ridges could be highly variable (Fig. 2.B
) exhibits successively a 4 (trapezoidal), 5 (pentagonal) and more than 6 (weakly polygonal) sequence. In the central areas of coralla the spacing between two main growth ridges could be highly variable (Fig. 2.B
![]() ) because their maximum curvatures are not situated on the same radial lines.
) because their maximum curvatures are not situated on the same radial lines.
|
|
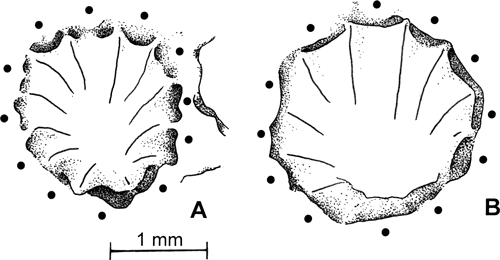
Figure 2:
Pinacopora baltica n. sp. Proximal side showing the main growth lines. The rounded angles
(i.e., maximum curvature of the growth lines) giving rise to the successive polygonal outlines of the corallum are indicated by black triangles. Bioclast to which the larva was attached in grey. The upper surface (drawing of the natural mould) is seen in "transparency". A, specimen RGM 1 333 568 (see also Fig. 3.A
|
Distal side. This side is slightly convex and the corallum consists generally of a few closely spaced corallites (tabularia) surrounded by a narrow coenenchyme.
As a result, the discoidal colonies are mainly asymmetric flat biconvex lenses.
i. Corallites.
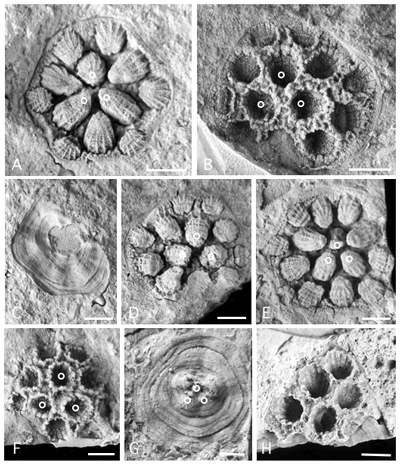
Calices. At the calicinal level, the apertures of the calices appear rounded or scalloped, circular or slightly elliptical, and each is lined by a narrow raised rim (Fig. 3.B, .F
). The calices of the corallites are conical in shape, but in some colonies they are truncated cone-shaped and thus exhibit indistinct to clear flat calicinal bases (Figs. 4.B
The calicinal base of each corallite is part of the basal plate and thus these corallites belong to the eucorallite category defined for Pleurodictyum-like discoid corals by Plusquellec (2007, p. 11). The interstitial corallites - those whose apex does not reach the basal plate - are not recorded in our material.
Septal ridges. The corallite wall bears 12 contiguous septal ridges whose number is not easy to establish at first sight. Nevertheless, the best preserved calicinal moulds show the print of 12 indisputable ridges alternating with 12 interseptal furrows (Fig. 3.A
Tabulae. The conical calices of the natural moulds (predominating or exclusive in younger specimens) do not show distinct features corresponding to the tabulae and it is inferred that they are missing. However, they are very likely present - horizontal, complete and covered with numerous small spines - in truncated cone-shaped calices (older specimens) since their flat base is interpreted as the surface of a tabula. However, indisputable spiny tabulae, only one in each corallite and situated close to their base, are exposed by the vertical split of the silicified specimen (Fig. 6
Pores. This kind of structure is usually very well preserved by natural moulds. Here, mural pores as well as basal pore (or "basal mural pore" of Swann, 1947, Pl. 2, fig. 8, = "apical pore" of Beecher, 1891, p. 211) are undoubtedly absent.
|
|
Figure 3:
Pinacopora baltica n. sp. Photographs of corallum in natural moulds (A, C, D, E), in latex cast (B, F, G) and in silicified preservation (H). Distal side (A, B, D, E, F, H), proximal side (C, G). Small open circles identify the prototriad (cf. Fig. 7
|
|
|
Figure 4:
Pinacopora baltica n. sp. Photographs of distal side of the corallum. A, latex cast of specimen RGM 1 333 571 (see also Fig. 3.E
|
|
|
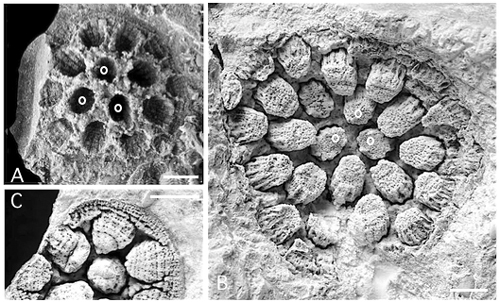
Figure 5:
Pinacopora baltica n. sp. Drawings of the calicinal moulds, view along their axis showing the 12 septal ridges. Black circles in front of the septal ridges, radial solid lines as prints of the interseptal furrows. Note the slight bilateral symmetry emphasized by the peripheral ridge (top of figure) broader than the others. Prints of the septal spines omitted. A, specimen RGM 1 333 573, corallite 21 right; B, specimen RGM 1 333 572, corallite 4x right. |
ii. Coenenchyme.
In the natural moulds, the empty space between the fillings of the corallites corresponds to the coenenchyme. Its surface morphology is well exposed in the restored specimen RGM 1 333 569 (Fig. 3.B
The accurate observation of coenenchyme is problematic in mouldic preservation, but its structure is shown by the vertical split of the silicified specimen (Fig. 6
|
|
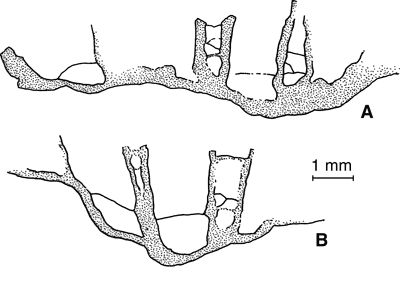
Figure 6:
Pinacopora baltica n. sp. Drawings of vertical split in a silicified specimen showing the presence of tabulae in some corallites and of diaphragms - complete or cystose - in the
coenenchyme; respectively part A and B of the specimen RGM 1 333 577. |
Development of the colony (astogeny). According to Oliver (1968), astogeny can be described in terms of form, increase (process whereby individuals multiply) and pattern (special arrangement of individuals).
1. Form: as previously stated in this paper, the colonies are discoidal (mainly flat biconvex lenses) and very likely fall in the tabular field of the growth-form variation defined by Young and Scrutton (1991).
2. Increase: owing to the lack of "basal mural pore", offsets could arise from the coenenchyme that unites individual corallites and thus, the mode of increase of the corallites belongs to the coenenchymal category (this is consistent with that previously described in Heliolitida as the most common feature [Lindström, 1899; Bondarenko, 1971]) or to the so-called epithecal category sensu Lindström (1899, p. 23) "in which calicles go out directly from the epitheca". Lindström (1899) illustrated this second mode of increase, giving very convincing figures for Heliolites interstinctus (Pl. I, fig. 28), Plasmopora scita (Pl. VII, fig. 11) and Propora tubulata (Pl. VIII, fig. 17) in which the coenenchyme appear to be developed prior to offsetting and for a specimen he called Propora (Pinacopora) girvanensis (Pl. IX, figs. 38-39) in which the coenenchyme appear to be developed after offsetting.
Remarks: the epithecal increase is not - in our opinion - a case of lateral increase as suggested by Oliver (1968), but more likely a case of peripheral increase (cf. Oliver, 1968, Fig. 1.B). Indeed, the presence of a basal mural pore between parent and offset is diagnostic of the lateral increase (Oliver, 1968, p. 21, Fig. 1.C; Stel, 1978, p. 167-168, Figs. 5-6); this kind of pore is missing in the heliolitids and halysitids. In addition, note that Webby (1975, Fig. 1.C-D) used "lateral increase" for Halysites sp. and Cystihalysites cavernosa, and that Hill (1981) on her Figure 289 4e, used "peripheral budding" for the above Cystihalysites cavernosa and Webby and Semeniuk (1969, Fig. 8) also peripheral budding for Halysites praecedens.
As a result, given the confusion in the nomenclature, it seems that the designation "epithecal increase" is particularly suitable, on one hand for some "epithecate" discoid corals, and in other hand for corals devoid of pores such as Heliolitina and Halysitina. Note that in some discoid Favositida, Pleurodictyum, Petridictyum etc, whose morphology is similar with that of Pinacopora, the increase is clearly lateral owing to the presence of a basal pore (see for example Pleurodictyum sp. in Plusquellec, 2007, Fig. 10).
The structure exposed by the specimen with silicified skeleton (Fig. 6
![]() ) makes clear that the increase of the Baltic material can be assigned to the epithecal category and it seems that the coenenchyme appear more likely to be developed prior offsetting.
) makes clear that the increase of the Baltic material can be assigned to the epithecal category and it seems that the coenenchyme appear more likely to be developed prior offsetting.
3. Pattern: in its first stages of development, the pattern seems to have never been clearly studied in heliolitids. The preservation of our material in natural moulds provides new data that help to fill this gap, taking into account some similarities with the Pleurodictyum-like corals (Plusquellec, 2007). Nevertheless, note that Lindström (1899, p. 45, Pl. 1, figs. 25-27) described and illustrated colonies "consisting of a few polypierites, which have sprung out from a single primary one" and, later, Noble and Lee (1990, Fig. 5) identified the three first corallites - but not the protocorallite - at the earliest portion of colony growth. In addition, Young and Elias (1995, p. 48) recognize the earliest growth stages in some specimens of Propora thebetensis but they stated that "the protocorallite cannot be distinguished" and the "small group of up to 10 similar closely spaced corallites [occurring] at the absolute base of the corallum (Text-fig. 12A)" does not allow to identify the prototriad.
In some colonies depicted herein, the protocorallite could be difficult to identify, especially because the basal pore is missing. Nevertheless, based on the best bilateral symmetry of the corallum the modalities of increase can be established. The numbering of the corallites and their nomenclature follow that was used by Fuchs and Plusquellec (1982) and Plusquellec (2007). Note that in these papers the founding corallite is called protocorallite as usual and metacorallite is used instead of hysterocorallite.
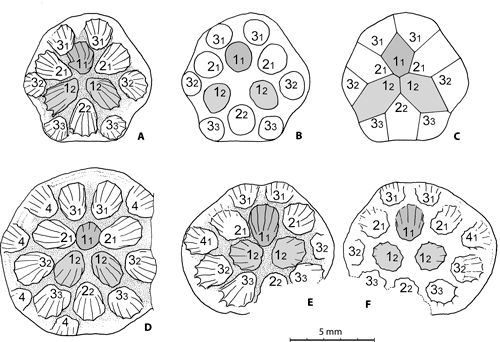
In our material the prototriad (protocorallite 11 + first pair of metacorallites 12) is of the contiguous type and the centrifugal development of the colony is marked by the addition of corallites of order 2 (21 22 21) followed by those of order 3 and 4 (Fig. 7
![]() ). In the colonies with more than 14-15 corallites (specimen RGM 1 333 572 and RGM 1 333 575) the numbering of the peripheral corallites becomes difficult to establish.
). In the colonies with more than 14-15 corallites (specimen RGM 1 333 572 and RGM 1 333 575) the numbering of the peripheral corallites becomes difficult to establish.
|
|
Figure 7:
Pinacopora baltica n. sp. Pleurodictyoid pattern of the colonies, drawings of the distal side, prototriad in light grey, protocorallite darker (explanations in text). A-C specimen RGM 1 333 568; A, natural mould of the calices of the corallites at the calicinal base level (see also Fig. 3.A
|
The development of the colonies belongs to the pleurodictyoid category (Plusquellec, 2007, Fig. 14A-D) owing to the fact that the prototriad occupy a central position within the corallum. In contrast, in the petrioid category the protocorallite - often larger than the metacorallites - is surrounded by a complete circle of 6 metacorallites (Plusquellec, 2007, Fig. H-I).
Note that in our material the colonies show a strong component of lateral growth, that the interstitial corallites are lacking and therefore the new offsets appear exclusively on the margin of the corallum (cf. peripheral growth strategy of Scrutton, 1997).
Remark: The pleurodictyoid development of the corallum probably occurs in each form of heliolitid with a flat or weakly convex "epithecate" proximal side such as Protarea illustrated by Sokolov (1962, Pl. I, fig. 2), but in large specimens - especially those with preserved skeleton - it is hardly recognizable. In this particular case, the mouldic preservation provides irreplaceable data. In addition, in large epithecate domed or bulbous colonies, together with the presumed pleurodictyoid development of their proximal part, new offsets (interstitial corallites) could arise at various levels within the corallum from the coenenchymal dissepiments or tubuli (see examples in Lindström, 1899: Heliolites interstinctus, Pl. 1, fig. 31, and Propora speciosa, Pl. 9, figs. 42, 46); in Scrutton, 1993: Propora tubulata, Fig. 4.4 left).
Measurements. The measurements of this very small species are mainly taken from camera lucida drawings of the natural moulds, some of them (relative to the distal surface) from latex replica.
1. Diameter of corallum: two series of measurements were taken, one along the plane of bilateral symmetry (which is also that of the
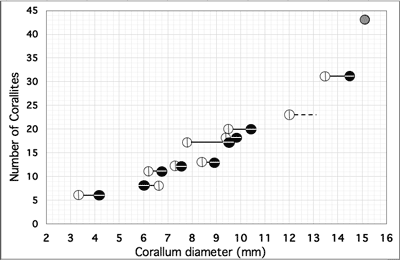
protocorallite), and the other normal to this plane. As a result, 1) the diameter of corallum along the plane of bilateral symmetry mainly varies between 6.2-9.4 mm, and normal to this plane between 6.6-10.4 mm; the largest corallum reaches about 15.1 mm, and 2) in almost cases (8 on 9) the later diameter is slightly greater (Fig. 8
![]() , Table 1).
, Table 1).
Remark: a brief review of the corallum diameter of some colonies of Devonian discoid tabulate corals illustrated by Plusquellec (2007) and belonging to Pleurodictyum, Petridictyum, Pterodictyum, Ligulodictyum, Cleistopora, Paracleistopora and Vaughanopsis, shows that the diameter normal to the plane of bilateral symmetry is usually greater than the other. On 25 measured colonies: 1) 72 % belong to the first category, and 2) the two categories could coexist within the same species.
|
|
Figure 8:
Pinacopora baltica n. sp. Relationship between corallum diameter and number of corallites. Open circle with vertical bar as diameter in the plane of bilateral symmetry, black circle with horizontal bar as diameter normal to this plane and grey circle as average diameter of the largest specimen. This specimen
(RGM 1 333 578) is somewhat deformed nearly the plane of bilateral symmetry of the colony and thus the two above diameter are not available with sufficient confidence, it is why only an apparent average (15.1) between maximum (16.0) and minimum (14.2) diameter is used. Some data, diameter or number of corallites, are estimated (see Table 1). Specimen with silicified skeleton
(RGM 1 333 577) too incomplete, not taken into account. |
Table 1:
Pinacopora baltica n. sp. Biometric data. Those with an* are estimated (corallum diameter as well as number of corallites). First column, diameter of corallum in the plane of bilateral symmetry, second column, normal to this plane, third column, average. The measurements of "specimen" RGM 1 333 5711 correspond to a potential colony inferred from its innermost growth line (see Fig. 2.B
![]() ). The specimen RGM 1 333 572 is incomplete and its diameter normal to the plane of bilateral symmetry is presumed to be probably greater than the one in this plane. About specimen RGM 1 333 578 see caption of Figure 8
). The specimen RGM 1 333 572 is incomplete and its diameter normal to the plane of bilateral symmetry is presumed to be probably greater than the one in this plane. About specimen RGM 1 333 578 see caption of Figure 8
![]() . Specimens RGM 1 333 572 and RGM 1 333 578 are not taken into account in the results got with 9 specimens.
. Specimens RGM 1 333 572 and RGM 1 333 578 are not taken into account in the results got with 9 specimens.
| Specimen |
Corallum |
Corallum diameter = |
Corallum diameter average |
Nbr corallites preserved |
Nbr corallites or Nbr estimated |
| RGM 1 333 568 | 7.3 | 7.4 | 7.35 | 12 | 12 |
| RGM 1 333 569 | 8.4 | 8.8 | 8.60 | 13 | 13 |
| RGM 1 333 570 | 7.8 | 9.5* | 8.65 | 14 | 17* |
| RGM 1 333 5711 | 3.3 | 4.2 | 3.75 | 6 | 6 |
| RGM 1 333 5712 | 9.4 | 9.7* | 9.55 | 17 | 18* |
| RGM 1 333 572 | 12.0 | ≥12.0 | ≥12.0 | 16 | 23* |
| RGM 1 333 573 | 6.2 | 6.6* | 6.40 | 11 | 11 |
| RGM 1 333 574 | 6.5 | 6.0* | 6.25 | 8 | 8 |
| RGM 1 333 575 | 13.5 | 14.5 | 14.0 | 28 | 31* |
| RGM 1 333 576 | 9.5* | 10.4 | 9.95 | 13 | 20* |
| RGM 1 333 578 | ≤15.1* | ≥15.1* | 15.1* | 42 | 43 |
| N 9 Average | 7.99 | 8.57 | 8.28 | 15.1 | |
| # 8.0 | # 8.6 | # 8.3 | # 15 | ||
| N 11 Average | 9.0 | 9.47 | 9.24 | 18.3 | |
| 9.0 | # 9.5 | # 9.25 | # 18 |
2. Thickness (height) of corallum: estimated on vertical/axial "sections" is difficult to establish; the colonies are rather flat and height generally does not exceed 2.5 mm.
3. Number of corallites: This varies between 8 and 20 for colonies of maximum diameter 6.5-10.4 mm and is estimated at 30-31 for a specimen of diameter 14.5 mm and reaches 43 for the largest known specimen (about 15.1 mm). The diagram
"Number of corallites" plotted against "Corallum
diameter" (Fig. 8
![]() ) clearly shows an allometric mode of growth of the colonies. It recalls that of the emblematic discoid
Pleurodictyum
problematicum (Fuchs & Plusquellec,
1982, Fig. 14).
) clearly shows an allometric mode of growth of the colonies. It recalls that of the emblematic discoid
Pleurodictyum
problematicum (Fuchs & Plusquellec,
1982, Fig. 14).
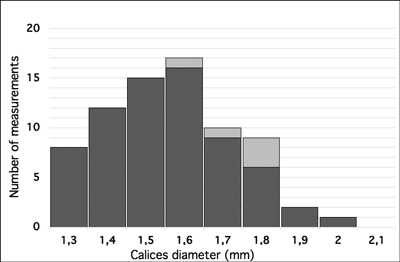
4. Diameter of calices: the measurements are taken normal to the radial line of the corallum (= smaller diameter of elliptical calices) and show small variations; diameter mainly between 1.3-1.8 mm (Fig. 9
![]() ).
).
|
|
Figure 9:
Pinacopora baltica n. sp. Histogram of calices diameter. Measurements of specimens in natural moulds in black, specimen with silicified skeleton
(RGM 1 333 577) in light gray. |
5. Distance between the centers of corallites: mainly between 1.8-2.3 mm.
6. Width of the coenenchymal surface: this measurement does not include the thickness of the peripheral rim of the calices and is taken along a line connecting their center; usually between 0.3-0.5 mm.
Part conclusion: Similar form, size and thickness of the corallum of our material suggests that the coral may have had a determinate growth, each colony lived for a definite time interval, reproduced, and died (G. Young, pers. comm., 2021). Therefore it is not a juvenile stage of a large species.
The lack of mural pores and basal pore together with the presence of 12 septal ridges clearly shown by the natural moulds, as well as the coenenchyme of diaphragms and dissepiments exhibited by a specimen with preserved skeleton, allow to assign the studied material to the Heliolitida.
Propora vs Pinacopora. The structure of tabularia and coenenchyme of the studied material is consistent with that of the Proporidae and is roughly similar to that of some specimens of Propora conferta illustrated by Dixon (1974, Pl. 2, figs. 2, 6) or to Propora tubulata illustrated by Young and Scrutton (1991, Fig. B1. N, P). Thus, at first sight, our material could be assigned to the genus Propora Milne-Edwards & Haime, 1849, which is mainly bulbous and domal but may be (rarely) discoidal as shown by Scrutton (1993) dealing with P. tubulata and P. n. sp.
Nicholson and Etheridge (1878, p. 52) introduced the genus Pinacopora (which can be considered a case of discoid Propora) for a "corallum composite, coin shaped, of extremely short corallites supported upon the convex surface of a free, discoidal, concavo-convex, concentrically-striated epitheca [ ] with large sized corallites [ ] surrounded by a circle of very much smaller corallites ("coenenchymal tubes"). Large tubes furnished with twelve short septum [ ] small tubes without septa. [ ] Large tubes furnished with one or two tabulae [ ] situated close to their base [ ]. Small tubes furnished with from two to four strong and complete tabulae wich extend to close their summit. [ ] No mural pores. No columella".
Laub (1979, p. 332) commented this diagnosis and stated: "Though their description of the genus and its type species P. grayi, is replete with reference to tabulate tubules between the corallites, the figures (e.g., Pl. 3, fig. 3d) clearly show a vesiculose stenosteum, often so narrow due to close approximation of the corallites, as to be about one dissepiment broad. In this, and in all other discernable features, Pinacopora is identical to, and a junior synonym of Propora".
Lindström (1899, Pl. 9, figs. 36-39), used Pinacopora as a subgenus of Propora in the legend of his plate for a specimen he called Propora (Pinacopora) girvanensis Nicholson & Etheridge, but called Pinacopora grayi Nicholson & Etheridge in the text (ibid., p. 99). Moreover, Lindström (ibid., p. 87) added Pinacopora in the synonymy list of Propora and stated (ibid., p. 99) "there would no reason for maintaining the genus Pinacopora as there is so very little to distinguish from Propora. I would prefer at present to let it stand as a synonym of Propora".
Later, Hill (1981, p. F612) indicated that Pinacopora could be a synonym of Propora, and Young (pers. comm., 2021) claimed that he "would prefer to keep the synonymization of Propora and Pinacopora".
Nevertheless, some discoid (= tabular) specimens that could be assigned to Propora, such as our material and P. grayi, are mainly characterized 1) by a strong, persistent component of lateral growth, 2) by scarcity of tabulae which are sometimes limited to only one in a corallite and situated close to its base and 3) by a coenenchyme of diaphragms and dissepiments until close to near the summit. In our opinion these features are sufficiently diagnostic for the resurrection of Pinacopora as a genus closely allied to Propora and our material can be assigned to Pinacopora Nicholson & Etheridge, 1878.
The three above features are necessary to assign a given form to Pinacopora and - for example - the discoidal Propora n. sp. Scrutton, 1993, although all colonies but one fall in the tabular field of Young and Scrutton (1991), cannot be assigned to Pinacopora owing to the development of very numerous tabulae and dissepiments to near the summit of respectively the corallites and coenenchyme.
Specific assignment. Only three species were previously assigned to Pinacopora:
Pinacopora andersoni. First remark, the so-called "small corallites" described by the authors are clearly a dissepimented or vesicular coenenchyme as in P. grayi. Secondly, the numerous tabulae in the corallites and the considerable number of dissepiments in the coenenchyme - that "seem to be constant" according to Nicholson & Etheridge (1880, p. 273) - do not allow to assign andersoni to Pinacopora although the corallum clearly belongs to the same tabular growth form. Thus, andersoni should be more reasonably regarded as Propora.
According to Dixon (1974, p. 583), Propora (Pinacopora) girvanensis Nicholson & Etheridge, 1880, sensu Lindström, 1899, is "almost certainly a small colony of Propora speciosa, as the trabecular coenenchyme, the trabecular corallite walls, and the tabularium diameter (about 2.0 mm) correspond". We can add that the tabulae in the corallites are obviously more numerous than in Pinacopora (see Lindström, 1899, especially Pl. 9, fig. 39) and moreover that the corallites are tabulate to a point near the surface. As a result the assignment to Propora is the better choice.
Remarks: contrary to what was stated by Dixon (1974) the reference to Pinacopora grayi by Lindström (1899, p. 99) is not an error; Pinacopora grayi Nicholson & Etheridge, 1878, is the type species of Pinacopora whereas Heliolites grayi Milne-Edwards & Haime, 1851, is the type species of Diploepora Quenstedt, 1879. Moreover, it seems that the species girvanensis used by Lindström has never been erected for a Propora or Pinacopora but for Plasmopora (Plasmopora girvanensis Nicholson & Etheridge, 1880, p. 266) and Favosites (Favosites girvanensis Nicholson & Etheridge, 1878, p. 34).
Part conclusion: The only species of Pinacopora with which the material depicted in this paper can be compared is P. grayi.
The material from the Upper Ordovician of the Öjlemyr chert shares the main characteristics of the Silurian
Pinacopora grayi (in fact features diagnostic of
Pinacopora) as shown by the comparison of Nicholson and Etheridge, 1878, Pl. 3, fig. 3E, and this paper Figure 5
![]() . It differs in having a much smaller corallum (6.2 mm-15.2 mm
vs 10 mm-25 mm according to the description of Nicholson & Etheridge, 1878, p. 55, "corallum [ ] varying from six to sixteen lines in diameter") and clearly larger corallites (1.3 mm-1.8 mm
vs about 1 mm ["about two third of a line"]). The number of corallites in
P. grayi reaches up to 350 in the large specimen illustrated by Nicholson and Etheridge (1878, Pl. 3, fig. 3f) and about 200 (estimated from Nicholson and Etheridge, 1878, Pl. 3, fig. 3b) for a corallum of 27 mm according to the stated enlargement. As a result, in colonies of similar diameter, the corallites are clearly more numerous in
P. grayi than in our material.
. It differs in having a much smaller corallum (6.2 mm-15.2 mm
vs 10 mm-25 mm according to the description of Nicholson & Etheridge, 1878, p. 55, "corallum [ ] varying from six to sixteen lines in diameter") and clearly larger corallites (1.3 mm-1.8 mm
vs about 1 mm ["about two third of a line"]). The number of corallites in
P. grayi reaches up to 350 in the large specimen illustrated by Nicholson and Etheridge (1878, Pl. 3, fig. 3f) and about 200 (estimated from Nicholson and Etheridge, 1878, Pl. 3, fig. 3b) for a corallum of 27 mm according to the stated enlargement. As a result, in colonies of similar diameter, the corallites are clearly more numerous in
P. grayi than in our material.
The pleurodictyoid pattern of development of the colonies is not recorded in P. grayi where the corallites are said to be arranged in obliquely intersecting lines. Nevertheless the pattern could be difficult to identify in colonies with very numerous corallites.
The morphology of the septal ridges seems to differ. In the described specimens they are systematically broad, short, slightly convex and alternating with narrow interseptal furrows whereas in P. grayi they seems to be narrow, acute and alternating with concave interseptal furrows (see Nicholson & Etheridge (1878, Pl. 3, fig. 3h, j, g). However, ibid., Pl. 3, fig. 3d, enlarged about twenty times could be more reliable than the former ones and shows a rather wide range of septal ridge morphology (especially some convex ridges with narrow interseptal furrow are recorded).
Another feature of the depicted material such as the presence of scattered small spines on the septal ridges cannot be compared; Nicholson & Etheridge did not provide comments on this topic. Note that this morphology (indicating that each ridge consists of irregular rows of more than one trabecula) has - as far as we know - never been described in the Proporidae in which only a vertical row of septal spines is identified or illustrated, among other in Propora speciosa (Dixon, 1974, p. 581, Fig. 7B) and in Propora tubulata (Young & Scrutton, 1991, Figs. B1, P). In addition, Hill (1981, p. F614) reports "septal spines sparse to ?absent in Rotalites".
The online catalog of Estonian fossils includes many examples of Propora conferta which in some cases have very small coralla, and G. Young (pers. comm., 2021) drew our attention to the silicified (?) specimen GIT 740-142 from Porkuni quarry, Porkuni regional stage (Hirnantian). Owing to the size of corallum and corallites, the narrow coenenchyme, the wide septal ridges bearing irregularly displayed small septal spines and the deep calices without obvious tabulae, this specimen appears similar to our material and would be assigned to the new species P. baltica.
It is interesting to note that transverse thin sections of some massive (domical or bulbous) Propora conferta, illustrated in the catalog of Estonian fossils such as the specimens GIT 520-202 and GIT 740-15 from Porkuni quarry, and the specimen illustrated by Lindström (1899, Pl. 9, fig. 1), exhibits low and broad septal ridges alternating with narrow septal furrows similar to those of Pinacopora baltica. A careful examination of the inner outline of the septal ridges of the GIT specimens show that some of them are not smooth and could bear up to three-four pointed knobs. Nevertheless, as stated by Lindström (ibid., p. 94) "the total want of septal spines makes [conferta] distinct from other Proporae", and Dixon (1974), dealing with material from the Late Ordovician of Anticosti, indicates that P. conferta shows a wide range of corallite wall structure but "in none of the specimens are true septal spines developed" (ibid., p. 574).
Lastly, if the opinion of Dixon (1974) about P. (P.) girvanensis (= P. speciosa) is questioned, anyway the morphology of the septal ridges of the specimen illustrated by Lindström (1899, Pl. 9, figs. 36-37) clearly differs from that of the Öjlemyr chert material.
Very small
Propora such as
P. savagei Young & Elias,
1995, share some structural and biometric characters with our material, but no low domical or tabular forms are known and moreover, pronounced exothecal projections are common; these later structures are clearly missing in the Öjlemyr material (cf. Fig. 5
![]() ).
).
As a result, our material exhibits a set of features not recognized in P. grayi and in any Propora and thus, a new species, P. baltica n. sp., is erected.
Heliolitids are usually not described from natural moulds, but this kind of preservation, absolutely devoid of erosion, can provide irreplaceable data on the morphology of some features such as the septal ridges, and about the pattern of development of the colonies, which is better seen at the calicinal base than at the calicinal aperture level.
Similar morphology and structure of the corallum at each stage of growth, strongly suggests that the specimens are not juvenile stages of large species and thus that the genus Pinacopora could be rehabilitated for tabular corals with tabulae absent or few in the corallites, deep calices and coenenchyme of scarce diaphragms.
A new species of Pinacopora, P. baltica n. sp., is fully described. The genus is not monospecific, the assigned species being P. baltica from the Upper Ordovician and P. grayi from the Lower Silurian. Pinacopora baltica occurs in Öjlemyr chert, an erratic Upper Ordovician silicified limestone which is found in Eridanos deposits surfacing in the west and north-east of the Dutch Overijssel-province and the adjoining German area.
The two species (i.e., Pinacopora baltica n. sp. and P. grayi) lived in shallow marine environment, the former, P. baltica, during the Late Ordovician in the south western margin of Baltica and the latter, P. grayi, in the South East of Laurentia during the Early Silurian, i.e., on both sides of the Iapetus Ocean which was narrowing before being closed at the end of the Upper Silurian (Stone et al., 2012). Pinacopora baltica is recorded in the Uppermost Katian and Hirnantian in Baltica whereas P. grayi appears in the Lower Llandovery in Laurentia. This distribution is consistent with an equatorial Iapetus current which facilitated dispersal of organisms from Baltica to Laurentia (Lam & Stigall, 2015).
Much gratitude is expressed first to Graham Young (Manitoba Museum, Canada) for e-discussion about the Proporidae, for providing bibliographic references, for useful comments on the manuscript and for its linguistic review. Thanks to Francis Tourneur (Pierres et Marbres de Wallonie, Belgium), Alain Le Herissé, Rémy Gourvennec (both from Université de Bretagne Occidentale, France), and Paul Copper (Prayssac, France) for various items.
Bassler R.S. (1950).- Faunal lists and descriptions of Paleozoic corals.- The Geological Society of America, Memoirs, Boulder - CO, vol. 44, 315 p.
Beecher C.E. (1891).- The development of a Paleozoic poriferous coral.- Connecticut Academy of Arts and Sciences Transactions, New Haven - CT, vol. 8, p. 207-214.
Bondarenko O.B. (1971).- Obyem novogo semeystva Stelliporellidae (Geliolitidy). In: Dubatolov V.N. (ed.), Tabulyaty i geliolitoidei paleozoya SSSR.- Trudy II, Vsesoyuznogo simpoziuma po izucheniyu iskopaemykh korallov SSSR, Moskva, vol. 1, p. 166-178.
Copper P., Jin J. & Desrochers A. (2013).- The Ordovician-Silurian boundary (Late Katian-Hirnantian) of western Anticosti Island: Revised stratigraphy and benthic megafaunal correlations.- Stratigraphy, Flushing - NY, vol. 10, no. 4, p. 213-227.
Dixon O.A. (1974).- Late Ordovician Propora (Coelenterata: Heliolitidae) from Anticosti Island, Quebec, Canada.- Journal of Paleontology, Cambridge - MA, vol. 48, p. 568-585.
Eiserhardt K.-H. (1992).- Die Acritarcha des Öjlemyrflintes.- Palaeontographica B, Stuttgart, vol. 226, 132 p.
Fuchs G. & Plusquellec Y. (1982).- Pleurodictyum problematicum Goldfuss 1829 (Tabulata, Dévonien). Statut, morphologie, ontogénie.- Geologica et Palaeontologica, Marburg, vol. 15, p. 1-26.
Hill D. (1981).- Treatise on Invertebrate Paleontology, Part F: Coelenterata, Supplement 1: Rugosa and Tabulata, vol. 1.- The Geological Society of America, Boulder - CO; The University of Kansas, Lawrence - KS, 378 p.
Hucke K. (1928).- Neue Untersuchungen über das Pliozän in Pommern und Brandenburg.- Zeitschrift für Geschiebeforschung, Berlin, vol. 4, p. 157-183.
Keulen P.S.F. van & Rhebergen F. (2017).- Typology and fossil assemblage of Sandbian (Ordovician) 'baksteenkalk': An erratic silicified limestone of Baltic origin from the northeastern Netherlands and adjacent areas of Germany.- Estonian Journal of Earth Sciences, Tallin, vol. 66, no. 4, p. 198-219.
Lam A.R. & Stigall A.L. (2015).- Pathways and mechanisms of Late Ordovician (Katian) faunal migrations of Laurentia and Baltica.- Estonian Journal of Earth Sciences, Tallin, vol. 64, no. 1, p. 62-67.
Laub R.S. (1979).- The corals of the Brassfield Formation (Mid-Llandovery; Lower Silurian) in the Cincinnati Arch Region.- Bulletins of American Paleontology, Ithaca - NY, vol. 75, no. 305, p. 1-526.
Lindström G. (1899).- Remarks on the Heliolitidć.- Kongl. Svenska Vetenskaps-Akademiens Handlingar, Stockholm, vol. 32, no. 1, 114 p.
Nicholson H.A. (1879).- On the structure and affinities of the "tabulate corals" of the Palaeozoic period.- Wm. Blackwood & Sons, Edinburg, London, XIII + 342 p. (44 Figs., 15 Pls.).
Nicholson H.A. & Etheridge R. (1878).- A monograph of the Silurian fossils of the Girvan district in Ayrshire with special reference to those contained in the "Gray collection".- Wm. Blackwood & Sons, Edinburg, London, vol. 1, fasc. 1, p. 1-135 (Pls. 1-9).
Nicholson H.A. & Etheridge R. (1880).- A monograph of the Silurian fossils of the Girvan district in Ayrshire with special reference to those contained in the "Gray collection".- Wm. Blackwood & Sons, Edinburg, London, vol. 1, fasc. 3, p. 237-341 (Pls. 16-24).
Noble J.P.A. & Lee D.-J. (1990).- Ontogenies and astogenies and their significance in some favositid ans heliolitid corals.- Journal of Paleontology, Lawrence - KS, vol. 64, no. 4, p. 515-523.
Oliver W.A. Jr (1968).- Some aspects of colony development in corals. In: Macurda D.B. (ed.), Paleobiological aspects of growth and development, a symposium.- Journal of Paleontology, Cambridge - MA, vol. 42, no. 5, supplement, p. 16-34.
Plusquellec Y. (2007).- Histoire naturelle des pleurodictyformes (Cnidaria, Tabulata, Dévonien) du Massif armoricain et des régions maghrébo-européennes principalement.- Mémoires de la Société géologique et minéralogique de Bretagne, Rennes, vol. 32, 126 p.
Rhebergen F. (2009).- Ordovician sponges (Porifera) and other silicifications from Baltica in Neogene and Pleistocene fluvial deposits of the Netherlands and northern Germany.- Estonian Journal of Earth Sciences, Tallin, vol. 58, no. 1, p. 24-37.
Schallreuter R. (1984).- Geschiebe-Ostrakoden I.- Neues Jahrbuch für Geologie und Paläontologie (Abh.), Stuttgart, vol. 169, p. 1-40.
Schallreuter R. (1987[a]).- Ostrakoden aus Öjlemyrflint-Geschieben von Sylt. In: Hacht U. von (ed.), Fossilien von Sylt II.- Verlag und Verlagsbuchhandlung Inge-Maria von Hacht, Hamburg, p. 203-232.
Schallreuter R. (1987[b]).- Bryozoen aus Öjlemyrflint-Geschieben von Sylt. In: Hacht U. von (ed.), Fossilien von Sylt II.- Verlag und Verlagsbuchhandlung Inge-Maria von Hacht, Hamburg, p. 233-247.
Schmidt F. (1881).- Revision der ostbaltischen silurischen Trilobiten nebst geognostischer Übersicht des ostbaltischen Silurgebiets. Abtheilung I.- Mémoires de l'Académie Impériale des Sciences de St-Pétersbourg (VIIe série), tome XXX, no. 1, 237 p.
Scrutton C.T. (1993).- Growth-form variation and control in two British Silurian species of Propora.- Courier Forschunginstitut Senckenberg, Frankfurt am Main, vol. 164, p. 273-281.
Scrutton C.T. (1997).- Growth strategies and colonial form in tabulate corals.- Bolletin de la Real Sociedad Espańola de Historia Natural (Sec. Geol.), Madrid, vol. 91, no. 1/4, p. 179-191.
Sokolov B.S. (1962).- Osnovy paleontologii. gubki, arkheotsiaty, kishechnopolostnye, chervi.- Izdatelstvo akademii nauk SSSR, Moscow, 485 p.
Stel J.H. (1978).- Studies on the paleobiology of favositids.- Rijkuniversiteit te Groningen, 247 p.
Stone P., McMillan S.P., Floyd A.A., Barnes J.D. & Phillip E.R. (2012).- British regional geology: South Scotland.- British Geological Survey, fourth edition, Keyworth, Nottingham, not paginated.
Swann D.H. (1947).- The Favosites alpenensis lineage in the middle Devonian Traverse Group of Michigan.- Contributions from the Museum of Paleontology University of Michigan, Ann Harbor - MI, vol. 6, no. 9, p. 235-318.
Vinn O. & Eyzenga J. (2021).- When did spines appear in cornulitids - A new spiny Cornulites from the Upper Ordovician of Baltica.- Neues Jahrbuch für Geologie und Paläontologie (Abh.), Stuttgart, vol. 299, no. 1, p. 99-105.
Webby B.D. (1975).- Patterns of increase in coenosteoid halysitid corals.- Alcheringa: An Australasian Journal of Palaeontology, vol. 1, no. 1, p. 31-36.
Webby B.D. & Cruse P.D. (1984).- The earliest heliolitines: A diverse fauna from the Ordovician of New South Wales.- Palaeontographica americana, Ithaca - NY, vol. 54, p. 164-168.
Webby B.D. & Semeniuk V. (1969).- Ordovician halysitid corals from New South Wales.- Lethaia, Oslo, vol. 2, p. 345-360.
Wiman C. (1901).- Über die Borkholmer Schicht im Mittelbaltischen Silurgebiet.- Bulletin of the Geological Institution of the University of Upsala, vol. 5, no. 10, Part. II, p. 149-222.
Young G.A. & Scrutton C.T. (1991).- Growth form in Silurian heliolitid corals: The influence of genetics and environment.- Paleobiology, Cambridge - MA, vol. 17, no. 4, p. 369-387.
Young G.A. & Elias R.J. (1995).- Latest Ordovician to earliest Silurian colonial corals of the East-Central United States.- Bulletins of American Paleontology, Ithaca - NY, vol. 108, no. 347, p. 7-148.
| Nomenclatural note: Life Sciences Identifier (LSID) https://zoobank.org/References/411A5853-DDF5-4962-BE0E-0764EB29AE19 Species Group • Pinacopora balticaPlusquellec, Eyzenga & Keulen, 2022 https://zoobank.org/NomenclaturalActs/A35D8F00-DCDF-4CBE-AA2B-76252461B87C |