◄ Carnets Geol. 23 (5) ►
![]()
Outline:
[1. Introduction]
[2. Geological setting and stratigraphy]
[3. Lithostratigraphy and age of the Kuran Formation]
[4. Study area, material, and methods]
[5. Systematic paleontology]
[6. Discussion and concluding remarks] and ...
[Bibliographic references]
Institute of Ecology and Earth Sciences,
University of Tartu, Ravila 14A, 50411, Tartu (Estonia)
Simon Bolivar Cad. 32B, no:54, Yıldız, Çankaya, 06550, Ankara (Republic of Türkiye)
Published online in final form (pdf) on June 30, 2023
DOI 10.2110/carnets.2023.2305
![]()
[Editor:
Bruno R.C. Granier]
![]()
The Jurassic platform margin succession in the Hezan area, called the "Hezan Unit" (Diyarbakır), on the north of the Arabian platform in southeastern Turkey includes five formations of carbonate-dominated rocks. The most interesting is the upper part of the Hezan unit (the Kuran Formation) that contains unique layers of oolitic and clayey limestone. A first record of the calcareous polychaete tubeworm Propomatoceros lumbricalis is described from the Middle Jurassic lower part of the Kuran Formation of the Hezan area. Most knowledge of fossil serpulids is centered on European material, and little has been done on Middle East fossil calcareous tubes. The taxonomic information recorded as a result of this study will add to our understanding of the biogeographic history of the Middle Jurassic calcareous polychaete associations and help to interpret the structure and paleoecology of its marine communities.
calcareous tubeworms;
Serpulidae;
Bivalvia;
Tethys;
Arabian plate;
biogeography
Vinn O. & Hoşgör İ. (2023).- The first record of Middle Jurassic serpulids from SE Turkey, equatorial Tethys.- Carnets Geol., Madrid, vol. 22, no. 5, p. 97-105.
Première observation de serpulidés du Jurassique moyen dans le sud-est de la Turquie (Téthys équatoriale).- Au nord de la plate-forme arabe, dans le sud-est de la Turquie, les séries jurassiques de bordure de plate-forme de la région de Hezan, appelée "unité de Hezan" (Diyarbakir), sont constituées de cinq formations sédimentaires, essentiellement des roches carbonatées. La partie supérieure de l'unité de Hezan, la Formation de Kuran, est particulièrement intéressante car elle présente des niveaux constitués de calcaires oolithiques et argileux. Le ver tubicole polychète calcaire Propomatoceros lumbricalis est signalé pour la première fois dans la partie inférieure du Jurassique moyen de la Formation de Kuran de la région de Hezan. La plupart des connaissances sur les serpulidés fossiles sont fondées sur du matériel européen tandis que peu de travaux concernent les tubes calcaires fossiles du Moyen-Orient. Les informations taxonomiques résultant de notre étude devraient donc permettre de mieux appréhender l'histoire biogéographique des associations de polychètes calcaires du Jurassique moyen, de même que la structure et la paléoécologie de ces communautés marines.
vers tubicoles calcaires ;
serpulidés ;
bivalves ;
Téthys ;
plaque arabique ;
biogéographie
The family Serpulidae is a group of sedentary calcareous polychaete tubeworms within the large order Sabellida (Hove & Kurpiyanova, 2009). Serpulids build a calcareous tube that from its embryonic state on is open at both ends. When the worm is active, branchiae protrude from the anterior aperture of the tube. The serpulid tubes usually have a smooth interior but a few species can form tabulae inside the lumen, oriented perpendicular to the tube's longitudinal axis. They may alternatively form internal longitudinal keels (Spiraserpula; Pillai 1993; Pillai & Hove, 1994; Vinn et al., 2008), but this is rare. The external morphology of serpulid tubes can vary considerably; tubes can show faint growth striations or variably developed ornamentations, such as transverse and longitudinal ridges and peristomes (Hove & Kupriyanova, 2009). In addition to Serpulidae, calcareous species occur in few other polychaetes, such as Sabellidae and Cirratulidae, where they are restricted to single genera in each family, Glomerula and Dodecaceria, respectively (Perkins, 1991; Fischer et al., 1989, 2000; Vinn et al., 2008). The extinct cirratulid genus Diplochaetetes dwelled in partly calcareous tubes (Kočí et al., 2021).
The serpulids appeared in the middle Permian (Sanfilippo et al., 2017, 2018) and became widespread since the Early Jurassic. Their major diversification took place during the Middle and Late Jurassic, and continued during the Cretaceous (Ware, 1975; Jäger, 1983, 2005; Ippolitov et al., 2014). Among fossil serpulids, faunas from the Upper Cretaceous are best known (e.g., Brünnich Nielsen, 1931; Jäger, 1983, 2005, 2011; Macellari, 1984; Radwańska, 1996; Kočí & Jäger, 2015a, 2015b; Kočí et al., 2017). Recently, Jurassic serpulids and sabellids have been in focus of several studies (e.g., Jäger et al., 2001; Radwańska, 2004; Ippolitov, 2007a, 2007b, 2010; Jäger & Schubert, 2008; Schögl et al., 2008; Vinn & Wilson, 2010; Kočí et al., 2019; Kočí & Főzy 2022). The most thorough taxonomic and paleoecological treatments of Jurassic serpulids have recently been published by Słowiński et al. (2022).
Records of Jurassic calcareous tubeworms from Turkey are very rare, usually limited to a few specimens. Only a few taxa, represented mostly by single specimens, have been reported so far (Okan & Hoşgör, 2007). Thus, any addition to the faunas of poorly known regions, such as parts of southeastern Turkey belonging to the northern Arabian Plate, provides new and interesting material for paleobiogeographical analyses.
The aims of the paper are: 1) to describe the first serpulids from the Jurassic of southeastern Turkey (Hezan Unit); 2) to compare Turkish serpulids with those of the European Middle Jurassic (south Germany, Hungary, Slovakia, Czechia, Poland and Russia); and 3) to discuss ecology and paleobiogeography of Turkish serpulids.
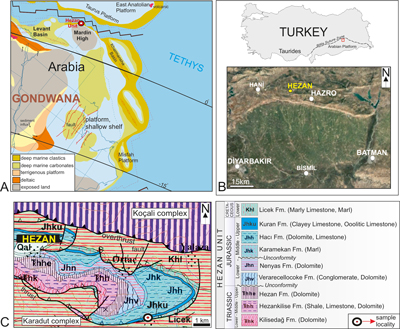
The northern
margin of the Arabian platform (southeastern Turkey) and its neighboring regions
were situated at the margin of the Tethys during the Middle Jurassic and
represented wide carbonate-siliciclastic platform margin and slope settings (Barrier
& Vrielynck, 2008; Énay, 2011) (Fig.
1A ![]() ). The platform
margin sequences (Hezan Unit) display carbonate facies deposited near the
paleoequator in a coastal and often inter- to supra-tidal environment ranging
in age from Lower Triassic to Middle Jurassic on the southern margin of the
Tethys (Fontaine et al., 1989; Golonka, 2004; Énay, 2011).
). The platform
margin sequences (Hezan Unit) display carbonate facies deposited near the
paleoequator in a coastal and often inter- to supra-tidal environment ranging
in age from Lower Triassic to Middle Jurassic on the southern margin of the
Tethys (Fontaine et al., 1989; Golonka, 2004; Énay, 2011).
In
southeastern Turkey, the Arabian platform consists of lower and upper
autochthonous and intervening allochthonous units (Fig. 1B ![]() ) (Righo de Righi
& Cortesini, 1964; Şengör & Yılmaz,
1981). The allochthonous Hezan Unit, consisting of carbonate and siliciclastic
rocks, crops out in the Late Cretaceous foothills belt north of Hazro in the
Border Fold zone (Bitlis Suture Zone) of SE Turkey (Fig. 1C
) (Righo de Righi
& Cortesini, 1964; Şengör & Yılmaz,
1981). The allochthonous Hezan Unit, consisting of carbonate and siliciclastic
rocks, crops out in the Late Cretaceous foothills belt north of Hazro in the
Border Fold zone (Bitlis Suture Zone) of SE Turkey (Fig. 1C ![]() ) (Tolun,
1949; Altıner, 1989; Fontaine et al., 1989;
Sütçü,
2008).The Arabian
carbonate platform is overthrusted by the Hezan allochthonous unit consisting of
Early Triassic to Early Cretaceous sediments, in which the transition between
neritic carbonates and the pelagic and detrital facies is indicated by an
ammonite-bearing succession of Jurassic age (Fontaine et
al., 1989; Günay et al.,
1990). Despite intensive study of the area, especially in the last thirty-five
years, the fossiliferous content of the outcropping
deposits in the Hezan is historically rare. For a long time it was essentially
represented by some cephalopods, foraminifers and calpionellids from the Hezan
unit succession providing lower Triassic to lower Cretaceous ages (Fig.
1C
) (Tolun,
1949; Altıner, 1989; Fontaine et al., 1989;
Sütçü,
2008).The Arabian
carbonate platform is overthrusted by the Hezan allochthonous unit consisting of
Early Triassic to Early Cretaceous sediments, in which the transition between
neritic carbonates and the pelagic and detrital facies is indicated by an
ammonite-bearing succession of Jurassic age (Fontaine et
al., 1989; Günay et al.,
1990). Despite intensive study of the area, especially in the last thirty-five
years, the fossiliferous content of the outcropping
deposits in the Hezan is historically rare. For a long time it was essentially
represented by some cephalopods, foraminifers and calpionellids from the Hezan
unit succession providing lower Triassic to lower Cretaceous ages (Fig.
1C ![]() ) (Fontaine,
1981; Atalay & Bayram, 1987; Fontaine
et al., 1989; Günay et
al., 1990). Fontaine (1981) subdivided the Jurassic rocks of the
Hezan region into five formations (in upward succession): Verarecellocoke,
Nenyas, Karamekan, Hacı and Kuran. The Verarecelloke and Nenyas formations
are Early Jurassic in age; the Karamekan, Hacı and lower part of the Kuran
formations belong to the Middle Jurassic, and the upper part of the Kuran
Formation belongs to the Upper Jurassic (Fig. 1C
) (Fontaine,
1981; Atalay & Bayram, 1987; Fontaine
et al., 1989; Günay et
al., 1990). Fontaine (1981) subdivided the Jurassic rocks of the
Hezan region into five formations (in upward succession): Verarecellocoke,
Nenyas, Karamekan, Hacı and Kuran. The Verarecelloke and Nenyas formations
are Early Jurassic in age; the Karamekan, Hacı and lower part of the Kuran
formations belong to the Middle Jurassic, and the upper part of the Kuran
Formation belongs to the Upper Jurassic (Fig. 1C ![]() ). The calcareous tube worm
material encrusting anomalodesmatan bivalves (Pholadomya sp.) described
here was collected from the upper part of the Hezan Unit (Kuran Formation).
). The calcareous tube worm
material encrusting anomalodesmatan bivalves (Pholadomya sp.) described
here was collected from the upper part of the Hezan Unit (Kuran Formation).
|
Figure 1:
A. Middle Jurassic
paleogeographic sketch map showing the position of the Hezan Unit and its
neighboring regions (Barrier & Vrielynck, 2008), B. Geographic
map of the south-east Turkey and Hezan area, C. Geological sketch map (Fontaine
et al., 1989; Sütçü, 2008) of the
Hezan area in northwestern Hazro showing the locations of the study area. |
The Kuran
Formation marks the beginning of an open marine environment. At the base are
several oolitic limestone layers with benthonic foraminifers and fragments of
reptile bones (Fontaine et al., 1989). Then follows a few metres of bio-lithoclastic
packstones, the most conspicuous horizon being a nodular red limestone in which
ammonites, belemnites and aptychi are abundant (Fig. 2 ![]() ) (Günay et
al., 1990). Occurrences of Kuran limestones, stated in the literature as
stratigraphically younger than Bajocian, are usually quite different
lithologically and extend partly into the early Valanginian. They can also be
found in places a considerable distance from the type area of Hezan (Atalay
& Bayram, 1987; Fontaine et
al., 1989). The age of the Kuran Formation at the Licek section is Bathonian
to Late Tithonian, which is a rather long interval (Günay et
al., 1990). Locally it contains a rather rich ammonite fauna, often
concentrated in few condensed beds. In his preliminary evaluation, Günay
et al. (1990) recorded diagnostic
ammonoid taxa of the Cadomites and Bullatimorphites
horizons which, in the current Submediterranean ammonoid zonal subdivisions (Mangold,
1984), corresponds to the middle Bathonian Cadomites
bremeri Zone. According to the lithological subdivision of the study area,
our material occurs in the middle Jurassic (Bathonian) Kuran Formation.
) (Günay et
al., 1990). Occurrences of Kuran limestones, stated in the literature as
stratigraphically younger than Bajocian, are usually quite different
lithologically and extend partly into the early Valanginian. They can also be
found in places a considerable distance from the type area of Hezan (Atalay
& Bayram, 1987; Fontaine et
al., 1989). The age of the Kuran Formation at the Licek section is Bathonian
to Late Tithonian, which is a rather long interval (Günay et
al., 1990). Locally it contains a rather rich ammonite fauna, often
concentrated in few condensed beds. In his preliminary evaluation, Günay
et al. (1990) recorded diagnostic
ammonoid taxa of the Cadomites and Bullatimorphites
horizons which, in the current Submediterranean ammonoid zonal subdivisions (Mangold,
1984), corresponds to the middle Bathonian Cadomites
bremeri Zone. According to the lithological subdivision of the study area,
our material occurs in the middle Jurassic (Bathonian) Kuran Formation.
|
Figure 2:
Lithostratigraphy
of the Kuran Formation (Licek section, Günay et
al., 1990), field views of the studied section and bivalve-bearing beds
recognized in the new stratigraphic section (Licek-2). |
The studied
serpulids described and illustrated herein came from field collecting in 2019.
They were found on an articulated pholadomyid bivalve shell from the lower part
of the Kuran Formation (Fig. 2 ![]() ), recovered from the south of Hezan village (GPS
coordinates: N38°19'51.68" E40°38'21.25"), about
15 km of northwest of Hazro area (Fig. 1B
), recovered from the south of Hezan village (GPS
coordinates: N38°19'51.68" E40°38'21.25"), about
15 km of northwest of Hazro area (Fig. 1B ![]() ).
).
The
Kuran Formation is dominantly a marly limestone unit with thin bedding, and
contains some claystones, oolitic limestones and nodular limestone beds (Licek
section, Günay et al., 1990) (Fig. 2 ![]() ). The macrofaunal content in the new section
(Licek-2) is poor; only the lower levels are fossiliferous, while the upper
levels are mainly composed of oolitic limestone. The fossils described here are
from a marly limestone bed containing bivalve-bearing levels, situated 3 m above
the base of the stratigraphic section (Fig. 2
). The macrofaunal content in the new section
(Licek-2) is poor; only the lower levels are fossiliferous, while the upper
levels are mainly composed of oolitic limestone. The fossils described here are
from a marly limestone bed containing bivalve-bearing levels, situated 3 m above
the base of the stratigraphic section (Fig. 2 ![]() ). Two observations of the bivalves
are noteworthy. First, epifaunal cementing bivalves, in particular gryphaeate
oysters, are abundant. Second, the large and deep infaunal suspension feeding
pholadomyid bivalves are very rare locally. On the pholadomyid bivalve Pholadomya
sp., serpulids are found mostly directed
towards the anterior margin of the bivalve. All the specimens were photographed with a Fujifilm
XH2 digital camera mounted. Studied specimens are housed at the Sivas Cumhuriyet
University Natural History Museum, Turkey (collection number CTF-SET01).
). Two observations of the bivalves
are noteworthy. First, epifaunal cementing bivalves, in particular gryphaeate
oysters, are abundant. Second, the large and deep infaunal suspension feeding
pholadomyid bivalves are very rare locally. On the pholadomyid bivalve Pholadomya
sp., serpulids are found mostly directed
towards the anterior margin of the bivalve. All the specimens were photographed with a Fujifilm
XH2 digital camera mounted. Studied specimens are housed at the Sivas Cumhuriyet
University Natural History Museum, Turkey (collection number CTF-SET01).
Phylum Annelida Lamarck, 1802
Class Polychaeta Grube, 1850
Subclass Sedentaria Lamarck, 1818
Order Sabellida Levinsen, 1883
Family Serpulidae Rafinesque, 1815
Genus Propomatoceros Ware, 1975
Type species: Propomatoceros sulcicarinata Ware, 1975; Aptian (Lower Cretaceous), Faringdon, UK.
Propomatoceros lumbricalis (Schlotheim, 1820)
(Fig. 3A-D ![]() )
)
1820 Serpulites lumbricalis; Schlotheim, p. 96.
1952 Serpula cf. lumbricalis von Schlotheim; Makowski, p. 4, Pl. 2: 2, 3.
1956 Serpula (Dorsoserpula) lumbricalis (von Schlotheim, 1820); Parsch, p. 219, Pl. 20, figs. 18, 20.
2007b Propomatoceros lumbricalis (von Schlotheim, 1820); Ippolitov, p. 432, Pl. 12, figs. 1c, 3, 6-8, 9c, 9d.
2022 Propomatoceros lumbricalis (von Schlotheim, 1820); Słowiński et al., Figs. 2A, 3F, 6C-F, 7A-D.
Material: Eight variably preserved specimens from SE Turkey.
Description: Tubes rather large (up to circa 70 mm long), almost straight to strongly curved. All tubes grow in diameter (up to 4.0 mm) at a moderately fast rate, which seems to constant through ontogeny. Larger tubes show a slightly widened base. A prominent median keel on top of the tube extends along its entire length. The keel is rather straight in the juvenile part of the tube but sometimes tends to undulate in the middle to anterior part. The tube shows no peristomes. Growth lines are weak or absent and better developed only near the median keel. The tube surface is smooth. Shape of the cross section depends on the ontogenetic stage, most often being triangular to subtriangular at early ontogenetic stages, and becoming more subcircular at later stages. The lateral walls are becoming more convex in the adult part of the tube.
Remarks: Propomatoceros is one of the most common fossil serpulids. It has a wide geographic and stratigraphic range (Słowiński et al., 2022). Its occurrence in Jurassic and Cretaceous deposits, combined with its conservative morphology, which varies intraspecifically and depends on paleoenvironment and ontogeny, hampers reliable species determinations (Słowiński et al., 2022). The species name Serpulites lumbricalis is historically the oldest available species name in the genus. It was introduced by Schlotheim (1820) for Middle and Late Jurassic specimens, but without illustration of the type specimen. Nevertheless, as this is the oldest available name, and our specimens resemble the description of the type material, we have decided to assign specimens from SE Turkey to S. lumbricalis. Parsch (1956). Ippolitov (2007b) validated the species status of Promopatoceros lumbricalis, and they considered forma "limax" of Goldfuss (1831) its subjective synonym, a path not followed by Słowiński et al. (2022). Moreover, Propomatoceros lumbricalis sensu stricto differs from the forma "limax" by its more prominent keel, better developed but less common peristomes, and smooth surface, as well as less convex lateral walls due to a usually faster growth, all of which fit well with the Turkish specimens.
Stratigraphic and geographic range: The material studied herein comes from the Bathonian (Middle Jurassic) of the SE Turkey. This species was also reported from the Middle and Upper Jurassic of Germany (Schlotheim, 1820; Parsch, 1956) and Poland (Słowiński et al., 2022), and the Middle Jurassic of Central Russia (Ippolitov, 2007b).
|
Figure 3:
Propomatoceros
lumbricalis (Schlotheim,
1820) encrusting anomalodesmatan bivalve Pholadomya sp.
from the Kuran Formation, Licek section, SE Turkey. A-B, apertural parts of worm
tubes. C-D, proximal parts of the worm tubes. |
The portion of the Kuran Formation containing this new serpulid record was formed at or near the J30 major maximum flooding surface (168 Ma) of Sharland et al. (2004). The Kuran Formation was deposited on a shallow shelf near the paleoequator on the southwestern margin of the Tethys Ocean. General lithology of lower part of the Kuran Formation is characterized by marly limestones, oolitic limestones and claystones. Thus, the sea bottom of this part of the basin must have been covered with either calcareous mud or clay mud in the Middle Jurassic. Usually shells of epifaunal bivalves serve as benthic islands for serpulids providing them with hard substrate in an otherwise soft bottom environment. In contrast, one would not expect to see encrusting serpulids on deep infaunal bivalves. However, Pholadomya is a deep-infaunal bivalve (Sztajner, 2020). Thus, there are two alternative scenarios. It can be argued that the bivalve was encrusted post mortem, after the shell was exhumed. In this scenario the orientation of the serpulid tubes towards the shell margin is accidental: In seven out of eight larger serpulid tubes, apertures are directed towards the anterior margin of the bivalve. It can be assumed that the orientations of the serpulid apertures were not controlled by the water and food flow generated by the living bivalve. Alternatively, the serpulids must have somehow managed to colonize living bivalves, but this seems very unlikely considering the great burrowing depth of Pholadomya (Sztajner, 2020). The serpulid tubes on the Pholadomya are slightly abraded post mortem, indicating that the waters were somewhat turbulent.
The majority of all known Middle and Upper Jurassic calcareous polychaete communities are described from Europe, including the European part of Russia (e.g., Pugaczewska, 1970; Jäger et al., 2001; Radwańska, 2004; Ippolitov, 2007a, b; Kočí et al., 2019; Breton et al., 2020; Kočí & Főzy, 2022; Słowiński et al., 2022). Except for some older studies (e.g., Parsch, 1956), the majority of reports on Jurassic serpulids dealt with assemblages coming from single stratigraphic intervals (e.g., Ippolitov, 2007a, b). Nine serpulid and sabellid species, mainly encrusting brachiopod shells and remains of sponges, were described by Kočí et al. (2019) from the Oxfordian (Upper Jurassic) of the Czech Republic. The European Jurassic serpulid associations are rather diverse (Parsch, 1956; Słowiński et al., 2022) and also include calcareous sabellids of the genus Glomerula (Słowiński et al., 2022). Propomatoceros is one of the most common serpulid genera in the European associations (Parsch, 1956; Jäger & Lang, 2017; Kočí et al., 2019; Kočí & Főzy, 2022; Słowiński et al., 2022). The calcareous polychaete associations outside Europe which are situated relatively closest to SE Turkey are those of Middle Jurassic (Upper Callovian) Matmor Formation in Israel, which in turn is the closest to the equator among the known Middle Jurassic tubeworm associations (Vinn & Wilson, 2010). In contrast, all European Jurassic basins were and are located in a considerably wide distance north of the equator. The Matmor Formation association differs from European associations by high dominance of the species of sabellid genus Glomerula and the presence of the genus Vermiliopsis. On the one hand, such differences might indicate that warm climate fit especially well with the ecology of the opportunistic Glomerula. On the other hand, Vermiliopsis might have originated in the warm equatorial, shallow seas before its further dispersal towards higher latitudes (Vinn & Wilson, 2010; Słowiński et al., 2022). It is surprising that Glomerula is missing in the studied sample from SE Turkey considering the paleogeographic proximity to Israel. Glomerula has previously been reported from the early Jurassic of the Ankara region of Central Anatolia (Okan & Hoşgör, 2007). Thus, the lack of Glomerula is most likely an artifact of sampling. Alternatively, it could have had a more patchy environmental distribution in the equatorial zone of the Tethys. Nevertheless, Propomatoceros occurs in the Matmor Formation of Israel, though it could not be affiliated with P. lumbricalis with certainty. Thus, it is likely that the genus Propomatoceros was not common only in European tropics but also in the very equatorial zone of Tethys Ocean.
Financial support to O.V. was provided by an Institute of Ecology and Earth Sciences (University of Tartu) Research Grant and a Paleontological Society Sepkoski Grant.
Altıner D. (1989).- An example for the tectonic evolution of the Arabian platform margin (SE Anatolia) during Mesozoic and some criticism of the previously suggested models. In: Şengör A.M.C. (ed.), Tectonic Evolution of the Tethyan Region.- Academic Publishers, Dordrecht, p. 117-129.
Atalay M. & Bayram O. (1987).- Stratigraphy of the allochthonous Triassic-Cretaceous carbonates in the Southeast Anatolia.- 7th Biannual Petroleum Congress of Turkey 131-138.
Barrier E. & Vrielynck B. (2008).- "Paleotectonic Maps of the Middle East", Middle East Basins Evolution Programme.- CGMW, Atlas, Maps, p. 1-14.
Breton G., Jäger M. & Kočí T. (2020).- The sclerobionts of the Bajocian Oolithe ferrugineuse de Bayeux Formation from Calvados (Paris Basin, Normandy, France).- Annales de Paléontologie, Paris, vol. 106, 102361.
Brünnich Nielsen K. (1931).- Serpulidae from the Senonian and Danian deposits of Denmark.- Meddelelser fra Dansk Geologisk Forening, Copenhagen, vol. 8, p. 71-113.
Énay R. (2011).- Toarcian and Bajocian ammonites from the Haushi-Huqf Massif of southwestern Oman and the Hawasina Nappes of the Oman Mountains: Implications for paleoecology and paleobiogeography.- GeoArabia, Manama, vol. 16, p. 87-122.
Fischer R., Pernet B. & Reitner J. (2000).- Organomineralization of cirratulid tubes-fossil and Recent examples.- Facies, Erlangen, vol. 42, p. 35-50.
Fischer R., Galli Oliver C. & Reitner J. (1989).- Skeletal structure, growth, and paleoecology of the patch reef-building polychaete worm Diplochaetetes mexicanus Wilson, 1986 from the Oligocene of Baja California (Mexico).- Geobios, Villeurbanne, vol. 22, p. 761-775.
Fontaine J.M. (1981).- La plate-forme arabe et sa marge passive mésozoïque : L'exemple d'Hazro (SE de la Turquie).- Édition Technip, Paris, p. 1-270 (Thèse 3e cycle, Orsay-Paris-11, Institut français du pétrole).
Fontaine J.M., Monod O. & Perinçek D. (1989).- The Hezan units: A fragment of the south Neo Tethyan passive continental margin in SE Turkey.- Journal of Petroleum Geology, Hoboken, vol. 12, p. 29-50.
Goldfuss A. (1831).- Petrefacta Germaniae.- Arnz, Düsseldorf1, Theil 1, Heft 3, p. 165-240 (Pls. LI-LXXI).
Golonka J. (2004).- Plate tectonic evolution of the southern margin of Eurasia in the Mesozoic and Cenozoic.- Tectonophysics, vol. 381, p. 235-273.
Grube A.E. (1850).- Die Familien der Anneliden.- Archiv für Naturgeschichte, Berlin, vol. 16, p. 249-364.
Gunay Y., Fourcade É., Dercourt J., Azéma J., Kozlu H., Bellier J.-P., Cordey F., Cros P., De Wever P., Énay R., Lauer J.P. & Vrielynck B. (1990).- The northern margin of the Arabian platform and its transition to a Tethyan oceanic area in the Mesozoic time in southeastern Turkey (Stratigraphy - Paleogeography).- 8th Petroleum Congress Turkey, Ankara, Turkish Association of Petroleum Geologists, p. 403-434.
Hove H.A. ten & Kupriyanova E.K. (2009).- Taxonomy of Serpulidae (Annelida, Polychaeta): The state of affairs.- Zootaxa, St. Lukes, vol. 2036, p. 1-126.
Ippolitov A.P. (2007a).- Contribution to the revision of some late Callovian serpulids (Annelida, Polychaeta) of central Russia: Part 1.- Paleontological Journal, Moscow, vol. 41, p. 260-267.
Ippolitov A.P. (2007b).- Contribution to the revision of some late Callovian serpulids (Annelida, Polychaeta) of central Russia: Part 2.- Paleontological Journal, Moscow, vol. 41, p. 429-436.
Ippolitov A.P. (2010).- Serpulid (Annelida, Polychaeta) evolution and ecological diversification patterns during Middle-Late Jurassic.- Earth Science Frontiers, Beijing, vol. 17, p. 207-208.
Ippolitov A.P., Vinn O., Kupriyanova E. & Jäger M. (2014).- Written in stone: History of serpulid polychaetes through time.- Memoirs of Museum Victoria, vol. 71, p. 123-159.
Jäger M. (1983).- Serpulidae (Polychaeta sedentaria) aus der norddeutschen höheren Oberkreide - Systematik, Stratigraphie, Ökologie.- Geologisches Jahrbuch, Reihe A, Stuttgart, vol. 68, p. 3-219.
Jäger M. (2005).- Serpulidae und Spirorbidae (Polychaeta sedentaria) aus Campan und Maastricht von Norddeutschland, den Niederlanden, Belgien und angrenzenden Gebieten.- Geologisches Jahrbuch, Reihe A, Stuttgart, vol. 157 (2004), p. 121-249.
Jäger M. (2011).- Sabellidae, Serpulidae and Spirorbinae (Polychaeta sedentaria) from the Barremian (Lower Cretaceous) of the Serre de Bleyton (Drôme, SE France).- Annalen des Naturhistorischen Museums in Wien, Serie A, Vienna, vol. 113, p. 675-733.
Jäger M., Kapitzke M. & Rieter M. (2001).- Neufunde von Pannoserpula pannosa (Quenstedt, 1857) (Polychaeta, Serpulidae) aus den Korallenkalken (Ober-Kimmeridgium) von Nattheim und Gerstetten (Schwäbische Alb).- Stuttgarter Beiträge zur Naturkunde (Serie B), Stuttgart, vol. 308, p. 1-17.
Jäger M. & Lang F. (2017).- Serpuliden und Sabelliden aus dem oberen Kimmeridgium von Saal bei Kelheim. In: Lang F. & Simonsen S. (eds.), Fossilien aus dem Riffschuttkalk des Kimmeridgium (Oberjura) von Saal a. d. Donau bei Kelheim (Bayern).- Der Steinkern, Bielefeld, vol. 30, p. 76-83.
Jäger M. & Schubert S. (2008).- Das Ober-Pliensbachium (Domerium) der Herforder Liasmulde; Teil 2, Serpuliden (Kalkröhrenwürmer).- Geologie und Paläontologie in Westfalen, Münster, vol. 71, p. 47-75.
Kočí T., Bosio G., Collareta A., Sanfilippo R., Ekrt B., Urbina M. & Malinverno E. (2021).- First report on the cirratulid (Annelida, Polychaeta) reefs from the Miocene Chilcatay and Pisco Formations (East Pisco Basin, Peru).- Journal of South American Earth Sciences, vol. 107, article 103042, 14 p.
Kočí T. & Főzy I. (2022).- Late Jurassic-Early Cretaceous tube-dwelling worms (Sabellidae, Serpulidae) from the Bakony Mountains. In: Főzy I. (ed.), Fauna, biostratigraphy, facies and paleotectonic evolution of the Late Jurassic-Early Cretaceous formations in the Bakony Mountains (Transdanubian Range, Hungary).- Institute of Geosciences, University of Szeged, GeoLitera Publishing House, p. 191-199.
Kočí T. & Jäger M. (2015a).- Filogranula cincta (Goldfuss, 1831), a serpulid worm (Polychaeta, Sedentaria, Serpulidae) from the Bohemian Cretaceous Basin.- Sborník Národního Muzea v Praze, řada B, Přírodnívédy, Acta Musei Nationalis Pragae (Series B, Historia Naturalis), vol. 71, p. 293-300.
Kočí T. & Jäger M. (2015b).- Sabellid and serpulid worms (Polychaeta, Canalipalpata, Sabellida, Sabellidae, Serpulidae) from the rocky coast facies (Late Cenomanian) at Předboj near Prague.- Sborník Národního Muzea v Praze, řada B, Přírodnívédy, Acta Musei Nationalis Pragae (Series B, Historia Naturalis), vol. 71, p. 31-50.
Kočí T., Jäger M. & Morel N. (2017).- Sabellid and serpulid worm tubes (Polychaeta, Canalipalpata, Sabellida) from the historical stratotype of the Cenomanian (Late Cretaceous; Le Mans region, Sarthe, France).- Annales de Paléontologie, Paris, vol. 103, p. 45-80.
Kočí T., Jäger M., amánek J. & Hyk P. (2019).- Tube dwelling polychaetes from the Oxfordian (Late Jurassic) of Hády Quarry at Brno (Moravia, Czech Republic).- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 294, no. 3, p. 311-332.
Lamarck, J.-B. de (1802 [privately published, reprinted 1906]).- Discours d'ouverture, prononcé le 27 floréal An 10, au Muséum d'Histoire naturelle. Recherches sur l'organisation des corps vivans.- Bulletin Scientifique de la France et de la Belgique (5e série), Paris, vol. 40, p. 483-517.
Lamarck J.B. de (1818).- Histoire naturelle des animaux sans vertèbres.- Deterville and Verdière, Paris, 612 p. URL: https://www.biodiversitylibrary.org/bibliography/12712
Levinsen G.M.R. (1883).- Systematisk-geografisk Oversigt over de nordiske Annulata, Gephyrea, Chaetognathi og Balanoglossi.- Videnskabelige Meddelelser fra Dansk naturhistorisk Forening i Kjøbenhaven, Copenhagen, vol. 1882, p. 160-251.
Macellari C.E. (1984).- Revision of serpulids of the genus Rotularia (Annelida) at Seymour Island (Antarctic Peninsula) and their value in stratigraphy.- Journal of Paleontology, New York - NY, vol. 58, p. 1098-1116.
Makowski H. (1952).- Callovian fauna from Łuków in Poland.- Palaeontologia Polonica, Warsaw, vol. 4, p. 1-64.
Mangold C. (1984).- Report of the Bathonian Working Group.- 1st International Symposium on Jurassic Stratigraphy, 1985, Copenhagen, 67-75.
Okan Y. & Hoşgör I. (2007).- The Bivalvia and Gastropoda fauna of the Ammonitico Rosso facies of Late Sinemurian-Early Pliensbachian of the Kösrelik Region (NE Ankara-Turkey); first record of Annelid Polychaete species and their paleogeographic and paleoecologic characteristics.- Mineral Research and Exploration Bulletin, Ankara, vol. 135, p. 19-29.
Parsch K.O.A. (1956).- Die Serpuliden-Fauna des südwestdeutschen Jura.- Palaeontographica A, Paläozoologie, Stratigraphie, Stuttgart, vol. 107, p. 211-240.
Perkins T.H. (1991).- Calcisabella piloseta, a new genus and species of Sabellinae (Polychaeta: Sabellidae).- Bulletin of Marine Science, Miami, vol. 48, p. 261-267.
Pillai T.G. (1993).- A review of some Cretaceous and Tertiary serpulid polychaetes of the genera Cementula and Spiraserpula Regenhardt 1961, Laqueoserpula Lommerzheim 1979 and Protectoconorca Jäger 1983.- Paläontologische Zeitschrift, Heidelberg, vol. 67, p. 69-88.
Pillai T.G. & Hove H.A. ten (1994).- On recent species of Spiraserpula Regenhardt, 1961, a serpulid polychaete genus hitherto known only from Cretaceous and Tertiary fossils.-Bulletin Natural History Museum London (Zoology), London, vol. 60, p. 39-104.
Pugaczewska H. (1970).- Traces of the activity of bottom organisms on the shells of the Jurassic ostreiform pelecypods of Poland.- Acta Palaeontologica Polonica, Warsaw, vol. 15, p. 425-440.
Radwańska U. (1996).- Tube-dwelling polychaetes from some Upper Cretaceous sequences of Poland.- Acta Geologica Polonica, Warsaw, vol. 46, p. 61-80.
Radwańska U. (2004).- Tube-dwelling polychaetes from the Upper Oxfordian of Wapienno/Bielawy, Couiavia region, north-central Poland.- Acta Geologica Polonica, Warsaw, vol. 54, p. 35-52.
Rafinesque C.S. (1815).- Analyse de la nature ou tableau de l'univers et des corps organisés.- Aux dépens de l'Auteur [privately published], Palerme, 224 p. URL: https://www.biodiversitylibrary.org/bibliography/106607
Righo de Righi M.& Cortesini A. (1964).- Gravity tectonics in foothills structure belt of southeast Turkey.- American Association of Petroleum Geologists, Bulletin, Tulsa, vol. 48, p. 1911-1937.
Sanfilippo R., Rosso A., Reitano A. & Insacco G. (2017).- First record of sabellid and serpulid polychaetes from the Permian of Sicily.- Acta Palaeontologica Polonica, Warsaw, vol. 62, p. 25-38.
Sanfilippo R., Rosso A., Reitano A., Viola A. & Insacco G. (2018).- New serpulid polychaetes from the Permian of western Sicily.- Acta Palaeontologica Polonica, Warsaw, vol. 63, p. 579-584.
Schlotheim E.F. von (1820).- Die Petrefactenkunde auf ihrem jetzigen Standpunkte.- Becker, Gotha, 437 p.
Schlögl J., Michalík J., Zágorŝek K. & Atrops F. (2008).- Early Tithonian serpulid-dominated cavity-dwelling fauna, and the recruitment pattern of the serpulid larvae.- Journal of Paleontology, New York - NY, vol. 82, p. 351-361.
Sharland P.R., Casey D.M., Davies R.B., Simmons M.D. & Sutcliffe O.E. (2004).- Arabian Plate sequence stratigraphy.- GeoArabia, Manama, vol. 9, p. 199-214.
Słowiński J., Vinn O., Jäger M. & Zatoń M. (2022).- Middle and Late Jurassic tube-dwelling polychaetes from the Polish Basin: Diversity, palaeoecology and comparisons with other assemblages.- Acta Palaeontologica Polonica, Warsaw, vol. 67, p. 827-864.
Sütçü Y.F. (2008).- 1/100 000 ölçekli Muş-L45 Paftası, no. 72, 54 p.
Sztajner P. (2020).- The mode of life in the genus Pholadomya as inferred from the fossil record.- Geosciences, Basel, vol. 10, no. 10, 23 p.
Şengör A.M.C. & Yılmaz Y. (1981).- Tethyan evolution of Turkey: A plate tectonic approach.- Tectonophysics, vol. 75, p. 181-241.
Tolun N. (1949).- Geologic notes on the Silvan ve Hazru area.- Bulletin of the Geological Society of Turkey, Ankara, vol. 2, p. 65-89.
Vinn O., Hove H.A. ten & Mutvei H. (2008).- On the tube ultrastructure and origin of calcification in sabellids.- Palaeontology, Hoboken, vol. 51, p. 295-301.
Vinn O. & Wilson M.A. (2010).- Sabellid-dominated shallow water calcareous polychaete tubeworm association from the equatorial Tethys Ocean (Matmor Formation, Middle Jurassic, Israel).- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 258, p. 31-38.
Ware S. (1975).- British Lower Greensand Serpulidae.- Palaeontology, Hoboken, vol. 18, p. 93-116.