◄ Carnets Geol. 24 (1) ►
![]()
Outline:
[1. Introduction] [2.
Geographical and geological context of the Melilla-Nador basin]
[3. Material and methods]
[4. Biostratigraphic results (Foraminifera)]
[5. Palynological results]
[6. Discussion] [7. Conclusions] [Bibliographic references] and ... [Plates]
Hassan II University of Casablanca, Faculty of Sciences Ben M'sik, Geology Department, Geosciences and Applications Laboratory, B.P 7955, Sidi Othmane Casablanca
(Morocco)
Hassan II University of Casablanca, Faculty of Sciences Ben M'sik, Geology Department, Geosciences and Applications Laboratory, B.P 7955, Sidi Othmane Casablanca
(Morocco)
Hassan II University of Casablanca, Faculty of Sciences Ben M'sik, Geology Department, Geosciences and Applications Laboratory, B.P 7955, Sidi Othmane Casablanca (Morocco)
Hassan II University of Casablanca, Faculty of Sciences Ben M'sik, Geology Department, Geosciences and Applications Laboratory, B.P 7955, Sidi Othmane Casablanca (Morocco)
Hassan II University of Casablanca, Faculty of Sciences Ben M'sik, Geology Department, Geosciences and Applications Laboratory, B.P 7955, Sidi Othmane Casablanca (Morocco)
Hassan II University of Casablanca, Faculty of Sciences Ben M'sik, Geology Department, Geosciences and Applications Laboratory, B.P 7955, Sidi Othmane Casablanca (Morocco)
Sorbonne Universitť, CNRS-INSU, Institut des Sciences de la Terre Paris, ISTeP UMR 7193, 75005 Paris (France)
Geobiostratdata.consulting, 385 Rte du Mas Rillier, 69140 Rillieux-la-Pape (France)
Published online in final form (pdf) on January 17, 2024
DOI:
10.2110/carnets.2024.2401
![]()
[Editor: Bruno
R.C. Granier; language editor: Stephen Eagar]
![]()
In order to contribute to the understanding of the evolution of marine and continental
environments, preceding the onset of the Messinian salinity crisis in the western
Mediterranean, we conducted an integrated study of the pre-evaporitic Messinian sedimentary series in the Melilla-Nador basin. Three sections have been carried out in the marl-diatomite series and were the subject of a detailed biostratigraphic and palynological
study. The study of planktonic foraminifera, pollen, dinocysts, and palynofacies allowed us to characterize the evolution of these
environments. From 6.83 to 6.52 Ma, the marine environment was relatively open,
calm, probably subject to the action of upwellings and received periodic continental inputs. Starting 6.52 Ma, the abundance and diversity of planktonic foraminifera
decreased. Continental inputs gradually dominate, alternating with marine ones, and reflecting a succession between proximal and distal neritic
environments. The surface water conditions were warm. After 6.35 Ma, began the degradation of marine conditions. The continental environment shows an open vegetal landscape dominated by herbaceous plants, reflecting a tropical to arid subtropical
climate, slightly less dry than that of the South Rifian Corridor.
This study confirms the existence of several parameters that contributed to the deposition of the cyclical marl-diatomite
series: on the one hand, the hot and dry climate favored the reduction of the plant landscape and therefore erosion (continental inputs); on the
other, the tectonics (volcanism and uplift).
• Melilla-Nador basin;
• Morocco;
• Western
Mediterranean;
• pre-evaporitic Messinian;
• marine environment;
•
continental environment;
• Foraminifera;
• pollen;
• dinocysts
Bahaj H., Barhoun N., Bachiri Taoufiq N., Rahmouna J., Targhi S., Berry N., Suc J.-P. & Popescu S.-M. (2024).- Paleoenvironmental changes preceding the onset of the Messinian salinity crisis in the western Mediterranean Sea (pre-evaporitic Messinian of the Melilla-Nador Basin, NE Morocco).- Carnets Geol., Madrid, vol. 24, no. 1, p. 1-27. DOI: 10.2110/carnets.2024.2401
Changements palťoenvironnementaux prťcťdant le
dťbut de la crise de salinitť du Messinien dans la mer Mťditerranťe
occidentale (Messinien prťťvaporitique du bassin de Melilla-Nador, Maroc NE).-
Afin de contribuer ŗ la comprťhension de l'ťvolution des environnements marins et continentaux, prťcťdant le dťbut de la crise de salinitť messinienne dans la Mťditerranťe occidentale, nous avons menť une ťtude intťgrťe de la sťrie sťdimentaire messinienne prťťvaporitique dans le bassin de Melilla-Nador. Trois coupes ont ťtť rťalisťes dans la sťrie marno-diatomique et ont fait l'objet d'une ťtude biostratigraphique et palynologique dťtaillťe. L'ťtude des foraminifŤres planctoniques, du pollen, des dinokystes et du palynofaciŤs a permis de caractťriser l'ťvolution de ces environnements. De 6,83 ŗ 6,52 Ma, le milieu marin est relativement ouvert, calme, probablement soumis ŗ l'action des upwellings et reÁoit pťriodiquement des apports continentaux. ņ partir de 6,52 Ma, l'abondance et la diversitť des foraminifŤres planctoniques diminuent. Les apports continentaux, alternant avec des apports marins, deviennent progressivement dominants, l'ensemble reflťtant une succession cyclique entre des environnements nťritiques proximal et distal. Les conditions de l'eau de surface sont chaudes. AprŤs 6,35 Ma, commence la dťgradation des conditions marines. L'environnement continental montre un paysage vťgťtal ouvert dominť par des plantes herbacťes, reflťtant un climat tropical ŗ subtropical aride, lťgŤrement moins sec que celui du Corridor Sud
Rifain.
Cette ťtude confirme l'existence de plusieurs paramŤtres qui ont contribuť au dťpŰt de la sťrie cyclique marno-diatomitique avec, d'une part, le climat chaud et sec favorisant l'ouverture du paysage vťgťtal et donc l'ťrosion (apports continentaux) et, d'autre part, la tectonique (volcanisme et soulŤvement).
• bassin de Melilla-Nador ;
• Maroc ;
•
Mťditerranťe occidentale ;
• Messinien prťťvaporitique ;
• environnement marin ;
•
environnement continental ;
• foraminifŤres ;
• pollen ;
•
dinokystes
The Messinian salinity crisis -MSC- (HsŁ et al., 1973, 1977) stands as an exceptional event that profoundly influenced the recent history of the Mediterranean basin. It resulted from the successive closure of the Betic and Rifian corridors (Krijgsman et al., 1999a, 1999b; MartŪn et al., 2001; Rouchy et al., 2003; Warny et al., 2003; CIESM, 2008; Bache et al., 2015; Gorini et al., 2015a, 2015b; Flecker et al., 2015; Tulbure et al., 2017; Capella et al., 2018, 2019; Targhi et al., 2021), which previously served as gateways for water exchange between the Atlantic Ocean and the Mediterranean Sea during the Late Miocene (Flecker et al., 2015; Achalhi et al., 2016; Capella et al., 2018, 2019; Krijgsman et al., 2018; Suc et al., 2019).
The progressive isolation of the Mediterranean has caused an important environmental change that has had an impact on the terrestrial and marine ecosystems of the Mediterranean region (Saint-Martin et al., 2003; Vasiliev et al., 2013, 2019; Roveri et al., 2014; Flecker et al., 2015; Agiadi et al., 2017; Karakitsios et al., 2017a, 2017b; Moissette et al., 2018; Suc & Frizon de Lamotte, 2019; Kontakiotis et al., 2019, 2020). The pre-evaporitic Messinian series of the peripheral Mediterranean basins are characterized by a typical succession of clays/marls surmounted by a cyclic series of marls and diatomites or sapropels (Sierro et al., 1993, 1999; Saint-Martin et al., 2003; Assen et al., 2006; Achalhi et al., 2016). These sediments have registered the changes that preceded the Messinian salinity crisis.
The Melilla-Nador peripheral basin, located in the northeast of Morocco, represents the eastern segment of the southern Rifian corridor. The sedimentary sequence of this basin consists of an Upper Miocene carbonate platform exhibiting reef characteristics, laterally passing to clays and marl-diatomite alternations (Saint-Martin & Rouchy, 1990; Cornťe et al., 1996; Saint-Martin et al., 2003).
Extensive research has focused on the Messinian sedimentation in this basin, encompassing stratigraphical, paleontological, and sedimentological investigations (Guillemin et Houzay, 1982; Rouchy, 1982; Saint-Martin & Rouchy, 1986; Saint-Martin, 1990; Saint-Martin et al., 1991; Gaudant et al., 1994; Cunningham et al., 1994; Saint-Martin & Cornťe, 1996; Cornťe et al., 1996, 2002, 2006; Rachid et al., 1997; MŁnch et al., 2001). However, the vegetation and, consequently, the climatic context in the Melilla-Nador basin before the Messinian salinity crisis remains poorly known. The present work is based on the detailed analyses of planktonic foraminifera, spores, pollen, dinocysts, and palynofacies from three selected sections in the study basin. The primary objectives of this study are, on one hand, to provide a better knowledge of the evolution of the marine environments and, on the other hand, to address the information gap regarding the continental environments by reconstructing the vegetation and climate of this basin during the pre-evaporitic Messinian.
Building upon a recently developed high-resolution biostratigraphic analysis for the Mediterranean (Sierro et al., 1993, 2001; Lourens et al., 2004; Assen et al., 2006; CorbŪ & Soria, 2016; Karakitsios et al., 2017a, 2017b; Kontakiotis et al., 2019; Lirer et al., 2019), a precise biostratigraphic framework has been established for the studied sediments. The study represents a new contribution to the reconstruction of the paleoclimates and the evolution of marine and continental environments during the pre-evaporitic Messinian in the Melilla-Nador basin, within a well-defined chronostratigraphic context. The aim is to enhance our understanding of the impact of the narrowing of the southern Rifian corridor on the climate and environment during a critical period in the history of the Mediterranean.
The Melilla-Nador basin, situated in the northeastern Rif near the Cap des Trois-Fourches
(Fig. 1
![]() ), is bordered to the south by the metamorphic terrains of the Rif
foreland. Neogene sediments were deposited unconformably on metamorphic rocks or tectonized Tortonian formations (Guillemin & Houzay,
1982). South of the Trois-Fourches cape, this basin is characterized by the development of a carbonate reef platform passing laterally into a basal marl-diatomite
sequence, which includes cinerite intercalations (Saint-Martin & Cornťe,
1996; MŁnch et al.,
2001; Saint-Martin et al.,
2003; Assen et al.,
2006; MŁnch et al., 2006; Cornťe et al.,
2006). The cyclical succession of marl and diatomite was deposited in shallow marine conditions registering precession cycles (Assen et al.,
2006). The carbonate platform is initially characterized by conglomeratic to bioclastic retrogradational
deposits, followed by bioclastic and reef carbonate prograding deposits (Saint-Martin & Cornťe,
1996; Saint-Martin et al.,
2003; Cornťe et al.,
2006). The initiation of cyclic sedimentation in the Melilla-Nador basin commenced around 6.84 Ma, marking a restriction of paleo-circulation across the basin and the onset of bioclastic platform progradation along the
shoreline. The restriction of paleo-circulation is linked to the reduction of the southern Rif corridor (Assen et al.,
2006). The commencement of diatomite deposition in the Melilla Basin aligned with the tectonic subsidence of the Taza-Guercif Basin and potentially with the initiation of siphoning of intermediate Atlantic waters through the narrowing Rifian corridor (Assen et al.,
2006). A change in diatom species from boreal to tropical coincided with the appearance of prograding coral reefs along the shelf (Assen et al.,
2006). This event indicates a decline in the inflow of cold water from the Atlantic
(ending the siphoning, according to Benson et al.,
1991). The end of diatomite deposition marks the closure of the open marine environment of the Melilla Basin (Assen et al.,
2006; Flecker et al., 2015).
), is bordered to the south by the metamorphic terrains of the Rif
foreland. Neogene sediments were deposited unconformably on metamorphic rocks or tectonized Tortonian formations (Guillemin & Houzay,
1982). South of the Trois-Fourches cape, this basin is characterized by the development of a carbonate reef platform passing laterally into a basal marl-diatomite
sequence, which includes cinerite intercalations (Saint-Martin & Cornťe,
1996; MŁnch et al.,
2001; Saint-Martin et al.,
2003; Assen et al.,
2006; MŁnch et al., 2006; Cornťe et al.,
2006). The cyclical succession of marl and diatomite was deposited in shallow marine conditions registering precession cycles (Assen et al.,
2006). The carbonate platform is initially characterized by conglomeratic to bioclastic retrogradational
deposits, followed by bioclastic and reef carbonate prograding deposits (Saint-Martin & Cornťe,
1996; Saint-Martin et al.,
2003; Cornťe et al.,
2006). The initiation of cyclic sedimentation in the Melilla-Nador basin commenced around 6.84 Ma, marking a restriction of paleo-circulation across the basin and the onset of bioclastic platform progradation along the
shoreline. The restriction of paleo-circulation is linked to the reduction of the southern Rif corridor (Assen et al.,
2006). The commencement of diatomite deposition in the Melilla Basin aligned with the tectonic subsidence of the Taza-Guercif Basin and potentially with the initiation of siphoning of intermediate Atlantic waters through the narrowing Rifian corridor (Assen et al.,
2006). A change in diatom species from boreal to tropical coincided with the appearance of prograding coral reefs along the shelf (Assen et al.,
2006). This event indicates a decline in the inflow of cold water from the Atlantic
(ending the siphoning, according to Benson et al.,
1991). The end of diatomite deposition marks the closure of the open marine environment of the Melilla Basin (Assen et al.,
2006; Flecker et al., 2015).
Bioclastic limestones and prograding Porites fringing reefs characterize the Messinian transgressive system of the studied basin. These sediments transition basinwards to open marine marl, diatomite, and volcano-clastic deposits (Cornťe et al., 2006). The age of prograding carbonate deposits varies, ranging from around 6.87Ī0.02 Ma (Roger et al., 2000) to 5.95-5.99 Ma (Cornťe et al., 2006). The conclusion of this transgressive cycle (Cycle 1) is marked by the development of Halimeda algal beds around 6.08 Ma. This Halimeda-rich level serves as a reference point across the entire basin.
The second carbonate complex, known as the Terminal Carbonate Complex (TCC) or Cycle 2, is of Messinian age. It exhibits a flat aggrading-retrograding geometry (Cornťe et al., 2002, 2006; MŁnch et al., 2006) and comprises reefs, giant microbial accumulations, and a combination of siliciclastic-oolitic coastal deposits associated with stromatolites. Two major unconformities border this complex (MŁnch et al., 2006). The 40Ar/39Ar dating places the base of the TCC (Terminal Carbonate Complex), coinciding with the onset of the Messinian salinity crisis (MŁnch et al., 2006), at an average of 5.95 to 5.99 Ma. Previous interpretations suggested that Cycle 2 deposits could be partially identified as continental Lago Mare-type deposits (Rouchy et al., 2003). However, taking into account that the Melilla-Nador basin emerged after the deposition of Upper Messinian sediments from marine Cycle 2 (Cornťe et al., 2006), MŁnch et al. (2006) demonstrated that these continental deposits evolved laterally into marine deposits, emphasizing the link between the emersion of the Melilla platform and tectono-magmatic activity.
The second major unconformity, corresponding to the major surface of the Upper Messinian, caps the CCT deposits in the Melilla basin. This erosion surface is associated to the primary Messinian drawdown in the Mediterranean. The duration of the hiatus does not exceed 450 kyr (MŁnch et al., 2006). The TCC can be considered as the marginal equivalent of the Messinian Salinity Crisis evaporites (Assen et al., 2006). Re-flooding deposits (Cycle 3) have primarily developed only in the southwestern part of the basin (Cornťe et al., 2006). They consist of yellowish marine sandstones and Isognomon limestones lying on the Messinian erosional surface. They constitute the second index level in the series. The sediments of Cycle 3 have been attributed to the Pliocene (Choubert et al., 1966; Guillemin & Houzay, 1982; MŁnch et al., 2001, 2006; Cornťe et al., 2002, 2006).
|
|
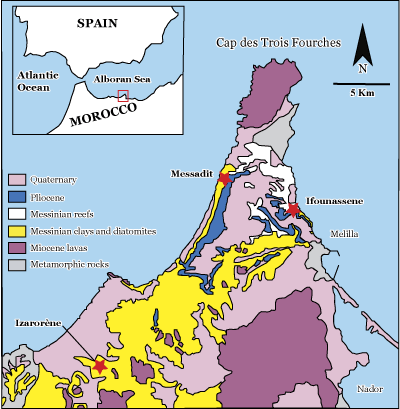
Figure 1: Simplified geological map of the Melilla-Nador basin (Choubert et al.,
1966) and the location of the studied
sections. |
The material studied is derived from three strategically located sections in the Melilla-Nador basin, namely the
Messadit, IzarorŤne, and Ifounassene sections
(Fig. 1
![]() ). These sections were meticulously sampled, resulting in the collection of more than 139 samples. However, only 103 samples exhibiting well-preserved microfauna and suitable for micropaleontological and biostratigraphical study are represented on the stratigraphic columns
(Figs. 2
). These sections were meticulously sampled, resulting in the collection of more than 139 samples. However, only 103 samples exhibiting well-preserved microfauna and suitable for micropaleontological and biostratigraphical study are represented on the stratigraphic columns
(Figs. 2
![]() -
3
-
3
![]() -
4
-
4
![]() ). The sampling interval ranges between 0.5 and 1.5 m.
We aimed for dense sampling to effectively capture variations in microfaunal associations and to account for lithological changes.
). The sampling interval ranges between 0.5 and 1.5 m.
We aimed for dense sampling to effectively capture variations in microfaunal associations and to account for lithological changes.
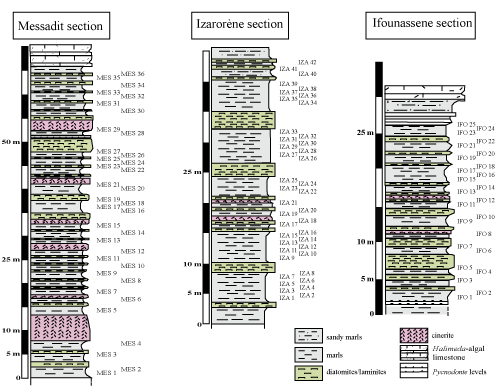
The Messadit section is situated in the western part of the peninsula, in the south of the village of Messadit
(Fig. 1
![]() ). The section has a thickness of 70 m and is characterized by marl-diatomite alternations containing several cinerite levels. This section is the most comprehensive, encompassing the key events of the Late Messinian in Morocco (Gaudant et al.,
1994; Saint-Martin & Cornťe,
1996; Rachid et al.,
1997; Saint-Martin et al.,
2003; Assen et al.,
2006)
(Fig. 2
). The section has a thickness of 70 m and is characterized by marl-diatomite alternations containing several cinerite levels. This section is the most comprehensive, encompassing the key events of the Late Messinian in Morocco (Gaudant et al.,
1994; Saint-Martin & Cornťe,
1996; Rachid et al.,
1997; Saint-Martin et al.,
2003; Assen et al.,
2006)
(Fig. 2
![]() ). At the base of the section, there is a volcano-sedimentary level that unconformably lies on glauconitic sands. Some authors (Gaudant et al.,
1994; Rachid et al., 1997) have described this level as a glauconitic conglomerate level with Messinian age overlying sedimentary Tortonian
units. The marl/diatomite alternations contain both planktonic and benthic foraminifera, ostracods, bivalves, bryozoans, sponge spicules, diatoms, and fish remains (Gaudant et al.,
1994; Rachid et al., 1997; Saint-Martin et al.,
2003). Indurated silicified laminites are intercalated in some places within this sedimentary series. Toward the upper part of the section, two oyster beds (Pycnodonte) were
observed. Assen et al. (2006) conducted a cyclostratigraphic study revealing 35 sedimentary cycles well calibrated on the astronomical time scale. Some cinerite layers have been dated using the isotope method (Roger et al.,
2000; Assen et al., 2006), providing precise dating for the carbonate sequences of the Melilla-Nador basin. Halimeda algae limestone banks, which represents the lateral equivalent of the last prograding coral reefs (Roger et al.,
2000; Saint-Martin et al., 2003; Cornťe et al.,
2006), cap the
section. A total of thirty-six samples
(Fig. 2
). At the base of the section, there is a volcano-sedimentary level that unconformably lies on glauconitic sands. Some authors (Gaudant et al.,
1994; Rachid et al., 1997) have described this level as a glauconitic conglomerate level with Messinian age overlying sedimentary Tortonian
units. The marl/diatomite alternations contain both planktonic and benthic foraminifera, ostracods, bivalves, bryozoans, sponge spicules, diatoms, and fish remains (Gaudant et al.,
1994; Rachid et al., 1997; Saint-Martin et al.,
2003). Indurated silicified laminites are intercalated in some places within this sedimentary series. Toward the upper part of the section, two oyster beds (Pycnodonte) were
observed. Assen et al. (2006) conducted a cyclostratigraphic study revealing 35 sedimentary cycles well calibrated on the astronomical time scale. Some cinerite layers have been dated using the isotope method (Roger et al.,
2000; Assen et al., 2006), providing precise dating for the carbonate sequences of the Melilla-Nador basin. Halimeda algae limestone banks, which represents the lateral equivalent of the last prograding coral reefs (Roger et al.,
2000; Saint-Martin et al., 2003; Cornťe et al.,
2006), cap the
section. A total of thirty-six samples
(Fig. 2
![]() ) were
palynogically and biostratigraphically analyzed with planktonic foraminifera.
) were
palynogically and biostratigraphically analyzed with planktonic foraminifera.
The Ifounassene section is situated in the eastern part of the peninsula
(Fig. 1
![]() ) and corresponds to the lower part of the Rostrogordo section (Saint-Martin & Rouchy,
1986; Cunningham et al.,
1994; Gaudant et al., 1994; Rachid et al.,
1997). The visible Messinian series is characterized by marl-diatomite alternations incorporating volcano-sedimentary
(cinerites) and bioclastic levels
(Fig. 2
) and corresponds to the lower part of the Rostrogordo section (Saint-Martin & Rouchy,
1986; Cunningham et al.,
1994; Gaudant et al., 1994; Rachid et al.,
1997). The visible Messinian series is characterized by marl-diatomite alternations incorporating volcano-sedimentary
(cinerites) and bioclastic levels
(Fig. 2
![]() ). The section is 30 m thick and overlaid by Halimeda limestone, representing the lateral equivalent of the prograding coral constructions of Unit 3 (Roger et al.,
2000; Saint-Martin et al., 2003; Cornťe et al.,
2006; Cunningham et al.,
1994). The section commences with homogeneous gray marls containing a Neopycnodont (Neopycnodonte navicularis) accumulation level (Rachid et al.,
1997) and resting on a volcano-sedimentary base
level. Assen et al. (2006) identified seventeen sedimentary cycles in this section, comprising alternating diatomite levels and homogeneous, partly silty marls containing bryozoans, bivalves, and
echinoderms. Benthic foraminifera, planktonic foraminifera, and ostracods constitute the primary microfauna in the sediments. The upper part of the section is composed of homogeneous sandy marls and two silicified and laminated levels with fish remains (Gaudant et al.,
1994). Twenty-five samples
(Fig. 2
). The section is 30 m thick and overlaid by Halimeda limestone, representing the lateral equivalent of the prograding coral constructions of Unit 3 (Roger et al.,
2000; Saint-Martin et al., 2003; Cornťe et al.,
2006; Cunningham et al.,
1994). The section commences with homogeneous gray marls containing a Neopycnodont (Neopycnodonte navicularis) accumulation level (Rachid et al.,
1997) and resting on a volcano-sedimentary base
level. Assen et al. (2006) identified seventeen sedimentary cycles in this section, comprising alternating diatomite levels and homogeneous, partly silty marls containing bryozoans, bivalves, and
echinoderms. Benthic foraminifera, planktonic foraminifera, and ostracods constitute the primary microfauna in the sediments. The upper part of the section is composed of homogeneous sandy marls and two silicified and laminated levels with fish remains (Gaudant et al.,
1994). Twenty-five samples
(Fig. 2
![]() ) were analyzed
palynogically and biostratigraphically (with planktonic foraminifera).
) were analyzed
palynogically and biostratigraphically (with planktonic foraminifera).
|
|
Figure 2:
Lithostratigraphy of the studied sections (MES : Messadit section, IZA : IzarorŤne section, IFO : Ifounassene section). |
It is located in the east of the village of IzarorŤne, about 5 km to the ESE of the confluence of the oued Kert with the Mediterranean
(Fig. 1
![]() ). This is a thick series of marls where laminated diatomite levels are intercalated in the upper part of the section (Barhoun et al.,
1999). The section begins with homogeneous blue to gray clayey marl, poorly stratified, overlying a level of white cinerite
(Fig. 2
). This is a thick series of marls where laminated diatomite levels are intercalated in the upper part of the section (Barhoun et al.,
1999). The section begins with homogeneous blue to gray clayey marl, poorly stratified, overlying a level of white cinerite
(Fig. 2
![]() ). In this section, Assen et al.
(2006) identified six cycles consisting of alternating diatomites and homogeneous marls. Above the third diatomite level, two cinerite levels are intercalated in the homogeneous marls. The section was densely sampled from the top of the first diatomite level. Forty-two samples
(Fig. 2
). In this section, Assen et al.
(2006) identified six cycles consisting of alternating diatomites and homogeneous marls. Above the third diatomite level, two cinerite levels are intercalated in the homogeneous marls. The section was densely sampled from the top of the first diatomite level. Forty-two samples
(Fig. 2
![]() ) were analyzed
palynogically and biostratigraphically (with planktonic foraminifera).
) were analyzed
palynogically and biostratigraphically (with planktonic foraminifera).
The samples underwent washing and sieving using the standard method (Barhoun, 2000). Qualitative and quantitative analyses of planktonic foraminiferal associations were conducted on residues containing a minimum of 300 individuals of the 150 Ķm fraction. To gain insights into paleoenvironments, two ecological indices were calculated:
1) the Planktonic-Benthic Ratio (P/P+B). This ratio, expressed as P/(P+B) x 100 (the percentage of planktonic foraminifera to the total foraminifera population), is commonly used to reconstruct paleobathymetry (Gibson, 1989). It also provides information about upwelling phenomena that favor high productivity (Mathieu, 1986, 1988; Bellier et al., 2010);
2) the Diversity Index. The species diversity in planktonic foraminiferal assemblages is based on the Shannon-Weaver function (Shannon & Weaver, 1949). It is calculated from the number of individuals of each species and the number of species for each sample. The estimation of the diversity index considers the proportion of each species in the total population. The Shannon index considers both the abundance and the uniformity of the species present (Daget, 1979; Murray, 1991).
Isc = - ∑ ( qi / Q x Ln qi / Q)
qi = the number of individuals counted for each species
Q = the total number of individuals for each level
The distribution of planktonic foraminifera is presented in tables. The number of specimens corresponding to abundant, common, and present is: Abundant > 20%; Common = 5-20%, and Present = 1-5%.
The standard palynological method, commonly employed for sediment treatment, was utilized for sample preparation following the procedure outlined by Bachiri Taoufiq
(2000). It involved HCl and HF treatments, followed by sieving of the residues between 10 and 180 Ķm, and an organic matter enrichment procedure using ZnCl2. Samples from IzarorŤne and Messadit sections
are poor in pollen and
dinocysts. Only 13 samples from the Ifounassene section yielded sufficient pollen grains and dinocysts (at least 160 pollen grains without Pinaceae and 100 dinocysts were counted). Based on the ecology of their representatives, the pollen taxa were grouped in a synthetic diagram (Suc et al.,
2018;
Fig. 7
![]() ), with the frequencies of each taxon represented in a detailed
diagram.
), with the frequencies of each taxon represented in a detailed
diagram.
To deduce global climate parameters, several methods were applied. The mean annual temperature (Tann) was estimated using current ecological conditions for plant groupings (Nix, 1982): megathermal elements (24įC), megamesothermal elements (20-24įC), mesothermal elements (14-24įC), and microthermal elements (10-14įC). For humidity assessment, the ratio Poaceae/Asteraceae (Cour & Duzer, 1978) was used. According to Cour and Duzer (1978), a Poaceae/Asteraceae ratio > 20 indicates a relatively humid climate, with precipitation exceeding 500 mm/year; conversely, a Poaceae/Asteraceae ratio < 20 indicates an arid climate with annual precipitation not exceeding 500 mm/year. Dry conditions favor the development of Asteraceae, whereas wet conditions favor Poaceae.
To reconstruct the marine environment, we applied indices previously used in the Neogene of Morocco by Warny (1999), Bachiri Taoufiq (2000), and Targhi et al. (2021, 2023). These indices are derived from the environmental preferences of dinocysts as recognized in the literature (Vernal et al., 1994, 1998, 2001; Vernal & Marret, 2007; Marret et al., 2001; Zonneveld, 1995, 1996; Zonneveld et al., 2013; Londeix et al., 2007). The curve resulting from the D/S ratio (D: Dinocysts and S: Spores + Pollen) is used to appreciate the marine influence versus the continental influence (Morzadec-Kerfourn, 1976; Tyson, 1993; Warny, 1999). The curve resulting from the G/P ratio (Tyson, 1993; Warny, 1999) is used to detect the inshore-offshore direction (G: Gonyaulacoid family, P: Peridinoid family). The IN/ON distality index is the ratio of inner platform neritic assemblages to outer platform neritic assemblages (Versteegh, 1994; Warny, 1999). The temperature index of surface waters is evaluated by the ratio of warm surface water dinocysts (W) to cool surface water dinocysts (C) (Warny, 1999). For the analysis of palynofacies, four standard groups have been identified in this work (Poumot & Suc, 1994; Combaz, 1991): woody organic matter (WOM), cellular organic matter (COM), black organic matter (BOM), and amorphous organic matter (AOM).
In light of recent work carried out on the Upper Neogene in the Mediterranean (CorbŪ & Soria, 2016; Karakitsios et al., 2017a, 2017b; Kontakiotis et al., 2019; Lirer et al., 2019), we conducted a detailed analysis of planktonic foraminiferal associations to revise the biostratigraphic framework of the studied sections. Three carefully chosen sections in the Melilla-Nador basin, where the Messinian deposits are thick and well represented, have been investigated (Messadit, Ifounassene, and IzarorŤne sections). Each section underwent intensive sampling to identify all the biostratigraphic and paleoenvironmental events characteristic of the Messinian. The analysis and detailed study of 103 samples collected from the three sections allowed the identification of 28 species of planktonic foraminifera, classified into 12 genera. To highlight biostratigraphic events, a complete count was performed on a portion containing about 300 individuals from the 150 μm fraction. In this work, we adopted high-resolution biostratigraphy, astronomically calibrated, recently utilized in the Mediterranean area, favoring precise dating and correlation (Hilgen et al., 1995; Krijgsman et al., 1994, 1995, 1999a, 1999b; Sierro et al., 1993, 2001; Lourens et al., 2004; CorbŪ et Soria, 2016; Karakitsios et al., 2017a, 2017b; Kontakiotis et al., 2019; Lirer et al., 2019). The main biostratigraphic events characterizing the Messinian in the Mediterranean area are indicated in Table 1.
Table 1: Main bioevents of planktonic foraminifera characterizing the Messinian in the Mediterranean area.
|
Planktonic foraminifera bioevents |
Chronostratigraphy (Ma) |
References |
|
Last Influx Turborotalita multiloba |
6.07 |
Sierro et al. (2001); Blanc-Valleron et al. (2002); Lourens et al. (2004) |
|
II Influx sinistral Neogloboquadrina acostaensis (40%) |
6.08 |
|
|
I Influx sinistral Neogloboquadrina acostaensis (90%) |
6.12 |
|
|
II Influx Turborotalita multiloba |
6.23 |
|
|
Influx dextral Globorotalia scitula |
6.29 |
|
|
S/D Neogloboquadrina acostaensis |
6.35-6.37 |
Sierro et al. (2001); Lourens et al.
(2004); |
|
FCO Turborotalita multiloba |
6.42 |
|
|
LO Globorotalia miotumida gr. |
6.52 |
|
|
LO Globorotalia nicolae |
6.72 |
Sierro et al. (2001); Blanc-Valleron et al.
(2002); |
|
FO Globorotalia nicolae |
6.83 |
|
|
Top dominant sinistral (Globorotalia) scitula gr. |
7.08 |
Lourens et al. (2004) |
|
influx conical Globorotalia miotumida |
7.18-7.22 |
Lourens et al. (2004) |
|
LCO Globorotalia menardii |
7.23 |
Hilgen et al. (2000a,
2000b); Lourens et al. |
|
FCO Globorotalia miotumida gr. |
7.24 |
Hilgen et al. (2000a,
2000b); Lourens et al. |
The micropaleontological study of sediments from the Messadit section reveals a remarkable richness in species and individuals of planktonic foraminifera, mainly in its lower part
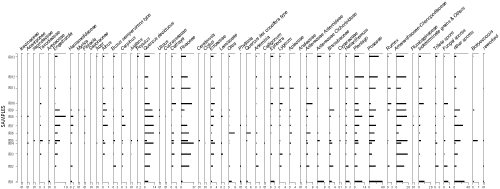
(Table 2). The abundance of planktonic assemblages diminishes from moderate to low towards the upper part of the section. This study identified 28 species and 10 genera. The microfauna also includes benthic foraminifera, some ostracods, fragments of bivalve shells, sponge spicules, and urchin
radioles. In the marl and marl-diatomite levels, the planktonic microfauna is well-preserved, enabling a qualitative and quantitative biostratigraphic study, whereas planktonic foraminifera are deformed and poorly preserved in some diatomite levels, making their identification
challenging. The evolution of planktonic foraminiferal associations, diversity, and the plankton-benthic ratio in the Messadit section allow us to distinguish two intervals
(Fig. 3
![]() ):
):
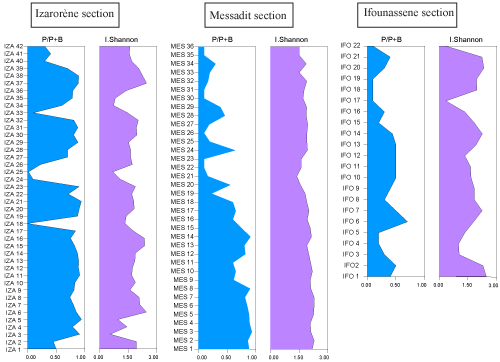
The lower part of the section from MES 1 to MES 18 is characterized by high diversity with a Shannon Diversity Index between 1.9 and 2.28. The Planktonic/Benthic ratio varies between 60 and 90%. Planktonic foraminifera assemblages are dominated by globular forms such as Globigerinoides, Globigerina, Globoturborotalita, Neogloboquadrina, etc., compared to the keeled forms of Globorotalia, represented by Globor. miotumida and Globor. conomiozea. These deposits are characterized by a notable abundance of Globigerina bulloides and Globig. falconensis in some levels (MES 2-MES 6 and MES 9-MES 15).
The upper part of the section from MES 19 to MES 36: The microfauna in these sediments is characterized by average diversity, with a Shannon Diversity Index between 1.5 and 1.9. The Planktonic/Benthic ratio is average to low, fluctuating between 10 and 60%. Benthic foraminifera are globally abundant and well represented compared to planktonic foraminifera.
Table 2: Distribution of the main planktonic foraminifera species in Messadit section.

|
|
Figure 3:
Planktonic/Benthic ratio and Diversity Index of the studied sections. |
Biostratigraphic analysis and dating: The stratigraphic distribution and quantitative analysis of planktonic foraminifera species, especially markers (Globorotalia nicolae, Globor. miotumida group, Turborotalita multiloba, and Neogloboquadrina acostaensis), along the series allows us to highlight the succession of the following events:
Event 1: It corresponds to the first presence of Globorotalia nicolae (FO Globorotalia nicolae). It was observed at the bottom of the section, precisely in the sample MES 3. Its astronomical age is 6.83 Ma (Sierro et al., 2001; Blanc-Valleron et al., 2002; Lourens et al., 2004; Lirer et al., 2019);
Event 2: The disappearance of Globorotalia nicolae (LO Globorotalia nicolae) is identified in sample MES 8. This event, dated at 6.712 Ma, has been documented by Sierro et al. (2001), Blanc-Valleron et al. (2002), Lourens et al. (2004), and Lirer et al. (2019);
Event 3: The Globorotalia miotumida group is present and sometimes abundant from the base of the section. The event 3 is defined by its last occurrence (LO Globorotalia miotumida gr.), located in sample MES 18. The astronomical age of LO Globorotalia miotumida gr. is 6.52 Ma (Sierro et al., 2001; Lourens et al., 2004);
Event 4: The first presence of Turborotalita multiloba (FCO T. multiloba) is observed in the MES 24 sample. Sierro et al. (2001), Lourens et al. (2004), and Lirer et al. (2019) have reported this event, with an astronomical age of 6.42 Ma;
Event 5: It is characterized by the sinister to dextral coiling change of Neogloboquadrina acostaensis (S/D N. acostaensis). It was identified in sample MES 28. This event, astronomically dated at 6.358 Ma, has been observed in many pre-evaporitic Messinian sedimentary series in the Mediterranean (Krijgsman et al., 1999a, 1999b; Hilgen & Krijgsman, 1999; Sierro et al., 2001; Lourens et al., 2004; Drinia et al., 2007; HŁsing et al., 2009; CorbŪ & Soria, 2016; Lirer et al., 2019);
Event 6: The last identified is marked by the first significant reappearance of the sinisterly coiled Neogloboquadrina acostaensis ("sinistral Neogloboquadrina acostaensis influx"). This event was located in sample MES 34, and its astronomical age is 6.12 Ma (Sierro et al., 2001; Lourens et al., 2004).
From the succession of these biostratigraphic bioevents, we could date this section to the pre-evaporitic Messinian with an estimated age of 7.08 to 6.08 Ma. The age of 6.08 Ma corresponds to the base of the Halimeda Limestone (Assen et al., 2006). This biostratigraphic analysis identified new biostratigraphic events and confirmed the results of previous work (MŁnch et al., 2001; Assen et al., 2006).
Microfaunal analysis from the Ifounassene section reveals an average to low abundance and diversity of planktonic foraminifera. Nineteen species and eight genera were identified. The washed residues are composed of planktonic and benthic foraminifera, some ostracods, mollusc fragments, sponge spicules, and urchin
radioles. Towards the top of the section the microfauna becomes rare to absent
(from IFO 22 to IFO 25). The vertical distribution of foraminiferal species is illustrated in Table
3. Planktonic foraminifera assemblages in the Ifounassene section are characterized by average to low diversity, with a Shannon
Diversity Index of approximately 1.6
(Fig. 3
![]() ). Some samples showed a high diversity in the order of
2.3. The Planktonic/Benthic ratio oscillates between 10 and 60%. The planktonic foraminifera association is mainly composed of globular forms such as Globigerinoides, Neogloboquadrina, and Turborotalita.
). Some samples showed a high diversity in the order of
2.3. The Planktonic/Benthic ratio oscillates between 10 and 60%. The planktonic foraminifera association is mainly composed of globular forms such as Globigerinoides, Neogloboquadrina, and Turborotalita.
Table 3: Distribution of planktonic foraminifera in the Ifounassene section.
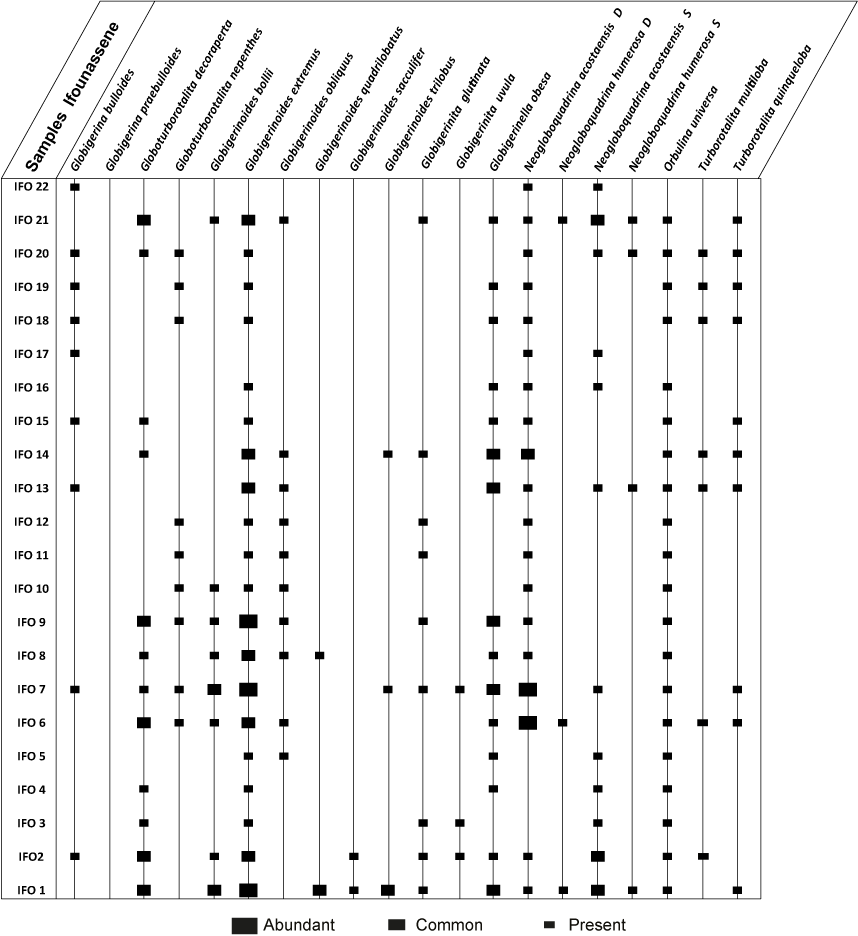
Biostratigraphic analysis and dating: Based on the vertical evolution of the marker species and the qualitative and quantitative analyses, the first presence of Turborotalita multiloba was observed from IFO 2, always associated with Turborotalita quinqueloba. Therefore, the FCO T. multiloba event is recorded at the IFO 2 sample. Furthermore, the vertical evolution of the percentage of Neogloboquadrina acostaensis sinistral and dextral forms of the studied samples expresses that the biostratigraphic event "coiling change of Neogloboquadrina acostaensis sin/dex (S/D N. acostaensis)" is located in sample IFO 6. The first significant reappearance of the sinistral coiling of Neogloboquadrina acostaensis (sinistral influx Neogloboquadrina acostaensis) was observed in sample IFO 21. From these biostratigraphic results, the Ifounassene section is dated to the pre-evaporitic Messinian with an estimated age of 6.52 to 6.08 Ma. This dating confirms the results of Assen et al. (2006).
The detailed analysis of the planktonic foraminiferal assemblages in the IzarorŤne section reveals significant richness in both species and individuals. A total of 30 species from 10 genera were
identified. The microfauna consists of planktonic and benthic foraminifera, along with some ostracods, fragments of bivalves, and urchin
radioles. Planktonic foraminiferal assemblages in the IzarorŤne section are characterized by moderate to high diversity, with Shannon Diversity Index varying between 1.4 and 2.4. The Planktonic/Benthic ratio is generally high, varying between 70 and 90%
(Fig. 3
![]() ). Globogerina bulloides, often associated with Globig. falconensis, is well represented in most samples, with significant abundances in some levels. However, periods of high abundance of Globig. bulloides alternate with periods where this species is poorly documented. This information will be utilized in the discussion. The vertical distribution of foraminiferal species is illustrated in
Table 4.
). Globogerina bulloides, often associated with Globig. falconensis, is well represented in most samples, with significant abundances in some levels. However, periods of high abundance of Globig. bulloides alternate with periods where this species is poorly documented. This information will be utilized in the discussion. The vertical distribution of foraminiferal species is illustrated in
Table 4.
Table 4: Distribution of planktonic foraminifera in the IzarorŤne section.

Biostratigraphic analysis and dating: The stratigraphic distribution and quantitative analyses of planktonic foraminiferal species, particularly the markers, reveals the presence of Globorotalia nicolae, the Globor. miotumida group, and Neogloboquadrina acostaensis. The examination highlights the succession of three significant biostratigraphic events. In the lower part of the IzarorŤne section, the first occurrence (FO) of Globorotalia nicolae was situated in sample IZA 3, whereas its last common occurrence (LO Globor. nicolae) is above the last massive diatomite (sample IZA 38). Globor. miotumida group is present from the base of the section, with its last occurrence (LO Globor. miotumida gr.) recorded in sample IZA 40. The succession of these bioevents allows us to attribute the IzarorŤne section to the pre-evaporitic Messinian with an estimated age of 7.24 to 6.35 Ma. These biostratigraphic findings refine and enhance previous dating efforts (Barhoun et al., 1999; Assen et al., 2006).
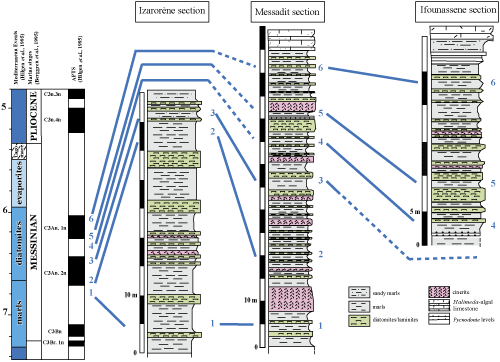
The correlation of the studied sections shows minor variations in thickness and the progressive installation of the degradation in environmental conditions, as evidenced by the decrease in the abundance and diversity of planktonic species and sometimes the microfauna is rare to absent. These unfavorable conditions for the proliferation of planktonic microfauna are related to the Messinian salinity crisis
(Fig. 4
![]() ). Within the interval 6.83-6.52 Ma, the thickness is 36 m in the IzarorŤne section and about 30 m in
Messadit. The environmental conditions indicate a relatively open marine environment probably subjected to the action of upwellings as shown by the
P/P+B ratio and the diversity index as well as the abundance of Globigerina bulloides and Globigerina falconensis. Indeed, Globigerina bulloides is a species that proliferates in cold, rich nutrient waters and is often used as an indicator of upwellings (Thiede,
1983; Prell, 1984). Conversely, from 6.52 Ma to 6.12 Ma, the thickness is around 20 m in the Messadit section. The
P/P+B ratio is average to low, planktonic foraminifera are low in abundance and diversity, particularly after 6.35 Ma, indicating a shallower marine environment (Gibson,
1989). From 6.12 Ma the planktonic microfauna is rare to absent because of deteriorating marine conditions caused by reduced water inflow from the Atlantic Ocean to the Mediterranean Sea resulting in widespread salt precipitation and a decrease in Mediterranean Sea-level due to evaporation. Furthermore, the end of the diatom bloom was reported around 6.11 Ma by Assen et al. (2006), followed by the installation of the algal level of
Halimeda. These events indicate the closure of the open marine environment in the Melilla-Nador basin and the degradation of environmental conditions in relation to the Messinian salinity
crisis.
). Within the interval 6.83-6.52 Ma, the thickness is 36 m in the IzarorŤne section and about 30 m in
Messadit. The environmental conditions indicate a relatively open marine environment probably subjected to the action of upwellings as shown by the
P/P+B ratio and the diversity index as well as the abundance of Globigerina bulloides and Globigerina falconensis. Indeed, Globigerina bulloides is a species that proliferates in cold, rich nutrient waters and is often used as an indicator of upwellings (Thiede,
1983; Prell, 1984). Conversely, from 6.52 Ma to 6.12 Ma, the thickness is around 20 m in the Messadit section. The
P/P+B ratio is average to low, planktonic foraminifera are low in abundance and diversity, particularly after 6.35 Ma, indicating a shallower marine environment (Gibson,
1989). From 6.12 Ma the planktonic microfauna is rare to absent because of deteriorating marine conditions caused by reduced water inflow from the Atlantic Ocean to the Mediterranean Sea resulting in widespread salt precipitation and a decrease in Mediterranean Sea-level due to evaporation. Furthermore, the end of the diatom bloom was reported around 6.11 Ma by Assen et al. (2006), followed by the installation of the algal level of
Halimeda. These events indicate the closure of the open marine environment in the Melilla-Nador basin and the degradation of environmental conditions in relation to the Messinian salinity
crisis.
|
|
Figure 4:
Correlations of the studied sections. |
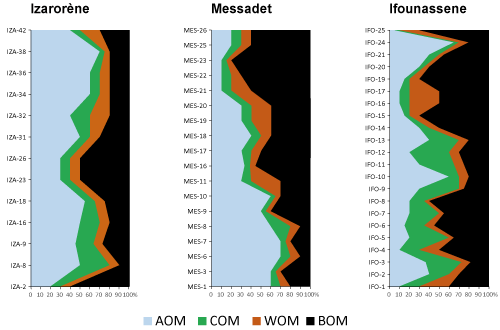
On all the samples processed, 13 from the Ifounassene section contained sufficient pollen, spores, and dinocysts for analysis. The samples from the IzarorŤne and Messadet sections, however, exhibited a low abundance of palynomorphs and were studied in terms of palynofacies.
The palynofacies study of 13 levels from the IzarorŤne section reveals a richness in
AOM, reaching up to 70%, except for levels IZA 2, 23, and 26, where the BOM exceeds 45%
(Fig. 5
![]() ). Samples from the Messadit section were found to be poor in palynomorphs. The palynofacies study
indicates abundant AOM in the lower part of the section, reaching 70%, gradually decreasing towards the upper part, reaching 10%
(Fig. 5
). Samples from the Messadit section were found to be poor in palynomorphs. The palynofacies study
indicates abundant AOM in the lower part of the section, reaching 70%, gradually decreasing towards the upper part, reaching 10%
(Fig. 5
![]() ).
Conversely, BOM follows the opposite trend. In the Messadit section, starting from MES 9, there is an alternation between the abundance of continental
(BOM, WOM, and COM) and marine (AOM) inputs. The content of woody debris (WOM) and cellular organic matter is low, varying between 5 and 20%.
).
Conversely, BOM follows the opposite trend. In the Messadit section, starting from MES 9, there is an alternation between the abundance of continental
(BOM, WOM, and COM) and marine (AOM) inputs. The content of woody debris (WOM) and cellular organic matter is low, varying between 5 and 20%.
|
|
Figure 5:
Palynofacies of the studied sections. |
Pollen grains and spores: Out of 23 samples collected, only 13 yielded palynomorphs. The pollen flora comprises 45 taxa
(Figs. 6
![]() -
7
-
7
![]() ). The floristic image reflected by the pollen spectra is predominantly homogeneous
(Figs. 6
). The floristic image reflected by the pollen spectra is predominantly homogeneous
(Figs. 6
![]() -
7
-
7
![]() ). The synthetic diagram
indicates richness in herbaceous plants, Pinus, mesotherms, and mega-mesothems with sporadic presence of megatherms
(Fig. 7
). The synthetic diagram
indicates richness in herbaceous plants, Pinus, mesotherms, and mega-mesothems with sporadic presence of megatherms
(Fig. 7
![]() ). Pinus constitutes 37% of the total, exhibiting a decrease in the IFO 1 and IFO 8 samples
(Fig. 7
). Pinus constitutes 37% of the total, exhibiting a decrease in the IFO 1 and IFO 8 samples
(Fig. 7
![]() ). Cathaya pollen is consistently present (0 to 6%) whereas Cupressaceae pollen is rare. High-altitude conifer pollen (Picea, Tsuga, Abies) was not
detected. Megatherm elements, including Acantaceae, Icacinaceae, and Sapindaceae, are sporadic, not exceeding 1%. In contrast, Engelhardia dominates (0 to 19%) over the other
mega-mesothermal elements (e.g., Taxodium, Araliaceae,
Hamamelidaceae, Myrica, Arecaceae, Sapotaceae), which are represented by a
lower percentage. Mesothermal elements are characterized by the prevalence of Quercus (3 to 13%),
alongside the presence of Ericaceae, Alnus, Buxus, Populus, Carya, Ulmus, Acer, and Juglans.
Mediterranean xerophytes, including Pistacia, Cistus, Phillyrea, and Ceratonnia, are present with prevailing pollen grains of Quercus coccifera and Olea.
Poaceae (5 to 40%) and Amaranthaceae-Chenopodiaceae (4 to 19.5%) dominate the pollen spectra. Other herbs
(Asteraceae Asteroideae, Asteraceae Cichoroideae, Apiaceae, Lamiaceae, Rumex,
Brassicaceae, Helianthemum, and Cyperaceae) are represented with low frequencies, except for the Plantago genus, reaching
13%. Lygeum, Calligonum, Artemisia, and Ephedra are found in almost all samples without exceeding
4%. Pteridophyte spores vary between 0 and 30%. Botryococcus is present
discontinuously.
). Cathaya pollen is consistently present (0 to 6%) whereas Cupressaceae pollen is rare. High-altitude conifer pollen (Picea, Tsuga, Abies) was not
detected. Megatherm elements, including Acantaceae, Icacinaceae, and Sapindaceae, are sporadic, not exceeding 1%. In contrast, Engelhardia dominates (0 to 19%) over the other
mega-mesothermal elements (e.g., Taxodium, Araliaceae,
Hamamelidaceae, Myrica, Arecaceae, Sapotaceae), which are represented by a
lower percentage. Mesothermal elements are characterized by the prevalence of Quercus (3 to 13%),
alongside the presence of Ericaceae, Alnus, Buxus, Populus, Carya, Ulmus, Acer, and Juglans.
Mediterranean xerophytes, including Pistacia, Cistus, Phillyrea, and Ceratonnia, are present with prevailing pollen grains of Quercus coccifera and Olea.
Poaceae (5 to 40%) and Amaranthaceae-Chenopodiaceae (4 to 19.5%) dominate the pollen spectra. Other herbs
(Asteraceae Asteroideae, Asteraceae Cichoroideae, Apiaceae, Lamiaceae, Rumex,
Brassicaceae, Helianthemum, and Cyperaceae) are represented with low frequencies, except for the Plantago genus, reaching
13%. Lygeum, Calligonum, Artemisia, and Ephedra are found in almost all samples without exceeding
4%. Pteridophyte spores vary between 0 and 30%. Botryococcus is present
discontinuously.
|
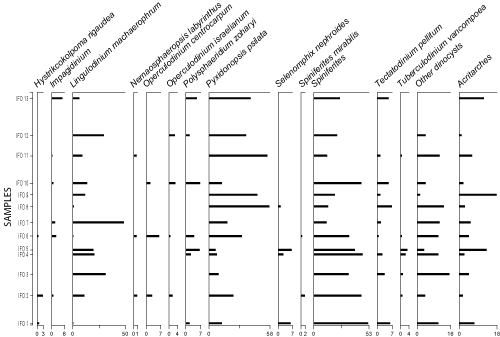
|
Figure 6: Detailed pollen diagram of the Ifounassene
section.
|
|
|
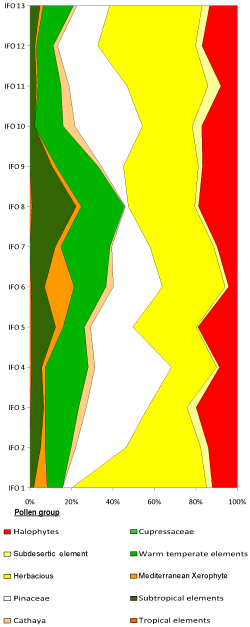
Figure 7:
Synthetic diagram of the Ifounassene section. |
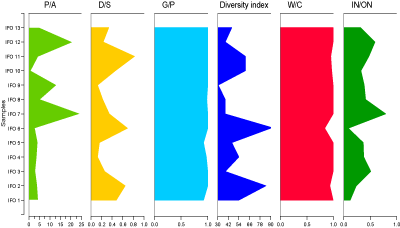
Dinocysts: The D/S curve shows consistently low values, indicative of a continental influence, except in samples IFO 2, IFO 6, and IFO 11, where the ratio exceeds 0.5, suggesting a marine influence
(Fig. 9
![]() ). The G/P curve shows ratios ranging between 0.92 and 1.00 in all samples
(Fig. 9
). The G/P curve shows ratios ranging between 0.92 and 1.00 in all samples
(Fig. 9
![]() ). The prevalence of Gonyaulocaceae indicates an open marine
environment. Dinocyst assemblages consist mainly of neritic taxa, including Spiniferites (13-52%), Pixydinopsis psilata (6-56%), Operculodinium israelianum (0-3.22%), Selenopemphix nephroides (0-6%), Lingulodinium machaerophorum (1-49%), and Polysphaeridium zoharyi (0-8.6%)
(Fig. 8
). The prevalence of Gonyaulocaceae indicates an open marine
environment. Dinocyst assemblages consist mainly of neritic taxa, including Spiniferites (13-52%), Pixydinopsis psilata (6-56%), Operculodinium israelianum (0-3.22%), Selenopemphix nephroides (0-6%), Lingulodinium machaerophorum (1-49%), and Polysphaeridium zoharyi (0-8.6%)
(Fig. 8
![]() ). Oceanic taxa are sparsely represented. These neritic dinocyst associations have been documented during the Early Messinian in Morocco at the Rifian corridor, reflecting a neritic environment (Warny,
1999; Warny et al., 2003, Bachiri Taoufiq,
2000; Targhi et al., 2021,
2023). These dinocyst associations collectively suggest the existence of a neritic marine
environment.
). Oceanic taxa are sparsely represented. These neritic dinocyst associations have been documented during the Early Messinian in Morocco at the Rifian corridor, reflecting a neritic environment (Warny,
1999; Warny et al., 2003, Bachiri Taoufiq,
2000; Targhi et al., 2021,
2023). These dinocyst associations collectively suggest the existence of a neritic marine
environment.
Distality index: The abundance of continental inputs indicates that the depositional environment of the Ifounassene section was
epicontinental. The D/S curve consistently shows low values, reflecting a continental influence, except in samples IFO 2, IFO 6, and IFO 11, where the ratio
exceeds 0.5, suggesting a marine influence
(Fig. 9
![]() ). The IN/ON distality index
(Fig. 9
). The IN/ON distality index
(Fig. 9
![]() ) varies between 0.1 and 0.8; indicating an external platform environment, except in levels 3, 7, 4.5, 8, and 12, where the ratio increases,
suggesting a shift towards a proximal neritic environment. This trend aligns well with the Planktonic/Benthic ratio
(Fig. 3
) varies between 0.1 and 0.8; indicating an external platform environment, except in levels 3, 7, 4.5, 8, and 12, where the ratio increases,
suggesting a shift towards a proximal neritic environment. This trend aligns well with the Planktonic/Benthic ratio
(Fig. 3
![]() ).
).
Temperature index: The temperature index varies between 0.8 and 1
(Fig. 9
![]() ), indicating a predominance of warm water taxa and warm thermal conditions at the water surface. In samples 2, 11, and particularly 6, a slight decrease in the W/C ratio is observed due to the appearance of cold taxa such as Operculodinium centrocarpum and Nematospheropsis
labyrinthea.
), indicating a predominance of warm water taxa and warm thermal conditions at the water surface. In samples 2, 11, and particularly 6, a slight decrease in the W/C ratio is observed due to the appearance of cold taxa such as Operculodinium centrocarpum and Nematospheropsis
labyrinthea.
|
|
Figure 8:
Detailed dinocyst diagram of the Ifounassene section. |
|
|
Figure 9:
Curves of P/A, D/S, G/P ratios and diversity, W/C (temperature), IN/ON
(distality) index of the Ifounassene section |
Ifounassene Palynofacies: The palynofacies of Ifounassene section exhibits a recurring shift between the prevalence of amorphous organic matter and the abundance of black organic matter (BOM) and cellular debris (COM)
(Fig. 5
![]() ). The richness of this organic matter indicates a strong continental influence, supporting the dinocyst data suggesting a continental-influenced neritic environment. Presumably, when conditions become less turbulent, they favor the development of
AOM.
). The richness of this organic matter indicates a strong continental influence, supporting the dinocyst data suggesting a continental-influenced neritic environment. Presumably, when conditions become less turbulent, they favor the development of
AOM.
The evolution of the marine environment in the Melilla-Nador basin during the pre-evaporitic Messinian is proposed in three stages based the qualitative and quantitative analyses of planktonic foraminifera, spores, pollen grains, dinocysts, and palynofacies.
Stage 1, from 6.83 to 6.52 Ma: This interval is characterized by three biostratigraphic events: First Occurrence (FO) of Globorotalia nicolae, Last Occurrence (LO) of Globor. nicolae, and Last Occurrence (LO) of Globor. miotumida group. It spans from the onset of cyclic marine sedimentation in the Melilla-Nador basin to the initial changes in diatom associations (Assen et al., 2006). The P/P+B ratio is often high, and the diversity is normal, occasionally marked by an abundance of Globigerina bulloides and Globig. falconensis, indicating a relatively open marine environment likely affected by upwellings. Benthic foraminifera also register episodes of upwellings, characterized by the prevalence of bolivinids and buliminids (Rachid et al., 1997). During this interval (6.83-6.52 Ma), sediment thickness varies between 30 and 36 m. The environmental conditions suggest a relatively open marine setting, as evidenced by the consistently high P/P+B ratio. The presence of Globigerina bulloides and Globig. falconensis implies the impact of upwelling, with Globog. bulloides often considered an indicator of cold, nutrient-rich waters and used as an upwelling indicator (Thiede, 1983; Prell, 1984). The examination of diatom assemblages and benthic foraminifera provides additional insights into the dynamics of Messinian upwellings in this basin (Rachid et al., 1997; Saint-Martin et al., 2003). The palynofacies is characterized by an overall abundance of amorphous organic matter, indicating a distal and calm environment. However, exceptions are noted in levels IZA 2, IZA 23, and IZA 26, where continental inputs (MOX, MOV, and MOB) dominate, reflecting an agitated and proximal environment at these levels. Additionally, the lower part of the Messadit section exhibits calm and distal conditions conducive to the development of amorphous organic matter. These findings align with the diatom data, which feature oceanic and oceanic-neritic species, along with silicoflagellates, confirming the existence of a distinct marine environment connected to the oceanic domain (Saint-Martin et al., 2003). Similar wet and warm conditions with oceanic environments and an external neritic trend have been observed in Salť (NW Morocco) between 7.3 and 6.5 Ma (Warny et al., 2003), the Guadalquivir Basin in Spain between 6.8 and 6.5 Ma (Jimťnez-Moreno et al., 2013), and in the central Mediterranean between 7.6 and 6.5 Ma (Bertini & Menichetti, 2015).
Stage 2, from 6.52 to 6.35 Ma: This stage is characterized by the succession of two biostratigraphic events observed in the Messadit and Ifounassene sections: First Common Occurrence (FCO) of Turborotalia multiloba and the coiling change S/D Neogloboquadrina acostaensis. From 6.52 Ma onwards, the P/P+B ratio becomes average to low, and planktonic foraminifera are less abundant and diversified, suggesting a shallower environment. Continental inputs, including pollen grains, spores, and plant debris dominate, prevail. They indicate a marine environment very close to the continent with significant fluvial inputs. Dinocyst associations in Ifounassene (IFO 1 to IFO 6 samples) suggest a neritic environment with warm surface waters. Distality indices (D/S, ID, Pinaceae) reveal an alternation between proximal and distal neritic environments. Comparisons with sections in Salť, Briqueterie site (NW Morocco, Warny et al., 2003), and the Guadalquivir basin (Spain, Jimťnez-Moreno et al., 2013) indicate that the percentage of pine trees in Ifounassene (not exceeding 35%) is very low. This implies that, from 6.52 Ma, the bathymetry in the Messinian marl-diatomite series gradually decreases due to the reduction of Atlantic water inflow and the uplift of the basin.
Stage 3, from 6.35 Ma to 6.12 Ma: From 6.35 Ma, planktonic foraminifera are poorly represented compared to benthic foraminifera. Regarding palynofacies, from IFO 6 to IFO 25, continental inputs continue to dominate compared to Messadit section. Distality indices (D/S, ID, pins) consistently show an alternation between proximal and distal neritic environments. Despite the closure of the two Rifian Corridors, i.e., the South Rifian Corridor at 7.1-6.9 Ma and the North Rifian Corridor from 7.35 to 7.00 Ma (Cappella et al., 2018, 2019), the basin remains open to the Atlantic and likely receives input through the Strait of Gibraltar (Capella et al., 2018; Krijgsman et al., 2018). From 6.12 Ma, planktonic microfauna becomes rare to absent due to deteriorating marine conditions. Additionally, Assen et al. (2006) reported the end of the diatom bloom around 6.11 Ma, followed by the establishment of the Halimeda algal level, indicating the closure of the open marine environment in the Melilla-Nador basin and the deterioration of environmental conditions associated with the Messinian salinity crisis.
Vegetal landscapes: The pollen study of the pre-evaporitic Messinian deposits of the Ifounassene section reveals the presence of
plant elements living today in the American and Asian regions (Cathaya, Engelhardia,
Sapotaceae, Arecaceae, and Hamamelidaceae), alongside elements currently living in Europe (Quercus,
Ericaceae, Alnus, Buxus, Populus, Carya, Ulmus, Acer, and Juglans) or the Mediterranean regions (Olea, Quercus type ilex-coccifera, Cistus, Pistacia, Phillyrea, and Ceratonnia). These same plant taxa have also been recorded in the Moroccan Messinian at the Rifain corridor (Bachiri Taoufiq,
2000; Bachiri Taoufiq et al.,
2008; Suc et al., 2018; Targhi et al.,
2021) and Boudinar basin (Achalhi et al.,
2016). Sub-desert taxa were also identified (Lygeum, Calligonum, Artemisia, and Ephedra). Taxa representing open plant formations
(Poaceae, Amaranthaceae-Chenopodiaceae, Plantago, etc.)
constitute a significant percentage. The mangrove element Avicennia, encountered in the northwestern Mediterranean from the Early to the Middle Miocene (Bessedik,
1984, 1985; Bessedik et al.,
1984), and in the south and western Mediterranean
(Spain, Sicily, Algeria, and Morocco) until the Messinian (Suc & Bessais,
1990; Chikhi, 1992a,
1992b; Bachiri Taoufiq et al.,
2001a, 2001b; Jimťnez-Moreno et al.,
2013; Targhi et al.,
2021, 2023), has not been recorded in this pre-evaporitic Messinian
section. The presence of a mixture of taxa with different thermal and hydric requirements can be explained by a certain staging of the vegetation (Bessedik,
1985; Suc, 1986,
1989)
and, consequently, a depositional environment relatively near the littoral and the mountain
massifs. Detailed analysis of the diagrams (Figs. 6
![]() - 7
- 7
![]() ) reveals two types of formations that succeeded each other behind the coral formations.
) reveals two types of formations that succeeded each other behind the coral formations.
Open littoral lowland formation: The littoral associations comprised xerophilous and/or halophilous herbaceous plants, including Poaceae, Amaranthaceae-Chenopodiaceae Plantago, and Asteraceae (Cichorioideae and Asteroideae). The prevalence of these pollen grains indicates the presence of a vast littoral plain. The occurrence of sub-desert plants such as Lygeum, Calligonum, Artemisia, and Ephedra confirms the arid nature of the environment. Trees and shrubs such as Arecaceae and Sapotaceae could also coexist in more humid areas. This open vegetation likely transitioned to a formation with Mediterranean elements, including Olea, Quercus type ilex-coccifera, Cistus, Pistacia, Phillyrea, and Ceratonia.
Arboreal formation: In areas relatively distant from the littoral, above the Mediterranean grouping and on the first reliefs, more forested associations of Pinus, Quercus, Ericaceae, Alnus, Buxus, Populus, Carya, Ulmus, Acer, Juglans, and evergreen elements (Sapotaceae, Arecaceae, Ilex) were likely present (Bachiri Taoufiq, 2000; Suc et al., 2018; Targhi et al., 2021). At higher elevations, towards the hinterland, this mixed mesophilic forest probably transitioned to a vegetation dominated by thermophilic elements like Taxodium, Cathaya, Engelhardia, Hamamelidaceae, Quercus, and Cupressaceae. The absence of high-altitude trees (Cedrus, Abies, and Picea) suggests a relatively low relief.
Climate: The pollen flora identified in Ifounassene section indicates a hot and dry
climate,
characterized by the presence of thermophilic elements such as Taxodium, Cathaya, Engelhardia,
Hamamelidaceae, Quercus, and Cupressaceae,
along with the abundance of taxa of dry environments such as herbaceous
sub-desert elements (Artemisia, Ephedra, Lygeum, and Calligonum).
Similar climatic conditions were observed in the southwestern Mediterranean in Morocco and Algeria during the Messinian (Chikhi,
1992a, 1992b; Warny,
1999; Warny et al., 2003; Bachiri Taoufiq et al.,
2001a, 2001b, 2008; Achalhi et al.,
2016; Suc et al., 2018; Targhi et al.,
2021). In the Ifounassene section, the absence of microthermals and the presence of
megathermals, megamesothermals, and mesothermals are notable. This observation leads us to estimate an average temperature
ranging from 20 to 24įC, consistent with the presence of guide taxa such as Lygeum (Fauquette et al.,
1999). At the middle altitude where Engelhardtia lived (Fauquette et al.,
1999), temperatures
consistently remained high (18-21įC), and precipitation levels were higher (600-700 mm per
year). These climatic conditions
bear similarity to those encountered in the Early Messinian in Algeria (Chikhi,
1992a, 1992b); Morocco (Warny,
1999; Bachiri Taoufiq, 2000), Sicily (Suc & Bessais,
1990; Bertini et al., 1998), and Spain (Jimťnez-Moreno et al.,
2013). These estimates
align with results from transfer function results applied to the Upper Miocene (Fauquette et al.,
2007; Suc et al., 2018) and the Lower Pliocene (Fauquette et al.,
1999), indicating a mean annual temperature of
approximately 23įC and annual precipitation around 450 mm. Precipitation values were estimated the ratio P/A was
used. In the Ifounassene section, the Poaceae/ Asteraceae ratio is generally less than 20, except in levels IFO 7 and IFO 12
samples, where it is greater than or equal to 20
(Fig. 9
![]() ). In the former case, rainfall is estimated to be less than 500 mm
while, in both levels, it exceeds 500 mm/year. A comparison of the Ifounassene section ratio with those of the lower Messinian in the southern Rifian corridor (Bachiri,
2000; Bachiri et al.,
2008; Targhi et al., 2021,
2023) indicates that climatic conditions in the Melilla-Nador basin were comparatively less
dry. Moreover, in the Melilla-Nador basin located in the
north, tropical elements such as Avicennia (found in the Tortonian and Lower Messinian along the southern Rifian corridor) are rare, and subtropical elements are less diverse. This implies a slight decrease in temperature and an increase in humidity from the south to the north of
Morocco. These
findings align with quantifications confirming the existence of an increasing latitudinal gradient of temperature and dryness from north to
south,
similar to the current gradient (Jimťnez-Moreno et al.,
2013; Bertini & Menichetti,
2015; Suc et al., 2018).
). In the former case, rainfall is estimated to be less than 500 mm
while, in both levels, it exceeds 500 mm/year. A comparison of the Ifounassene section ratio with those of the lower Messinian in the southern Rifian corridor (Bachiri,
2000; Bachiri et al.,
2008; Targhi et al., 2021,
2023) indicates that climatic conditions in the Melilla-Nador basin were comparatively less
dry. Moreover, in the Melilla-Nador basin located in the
north, tropical elements such as Avicennia (found in the Tortonian and Lower Messinian along the southern Rifian corridor) are rare, and subtropical elements are less diverse. This implies a slight decrease in temperature and an increase in humidity from the south to the north of
Morocco. These
findings align with quantifications confirming the existence of an increasing latitudinal gradient of temperature and dryness from north to
south,
similar to the current gradient (Jimťnez-Moreno et al.,
2013; Bertini & Menichetti,
2015; Suc et al., 2018).
The detailed study of planktonic foraminifera, spores, pollen grains, dinocysts, and palynofacies on three marl-diatomite sedimentary series in the Melilla-Nador Basin enhances our understanding of the evolution of marine and continental environments during the pre-evaporitic Messinian in the Western Mediterranean. During the interval 6.83-6.52 Ma, the marine environment in the basin was relatively deep and open, likely influenced by upwellings, rich in planktonic foraminifera, oceanic and oceanic-meridian diatoms, and MOA. Periodically, continental inputs occurred, suggesting a shift towards a proximal environment. Warm surface water conditions are indicated by the presence of coral reefs.
Starting from 6.52 Ma, the significance and diversity of planktonic foraminifera decreased, boreal diatoms transitioned to tropical diatoms, and carbonate platform reefs prograded. Alternating with amorphous organic matter, continental inputs gradually dominated, indicating a marine environment close to the continent with significant fluvial inputs. Distality indexes suggest alternation between proximal and distal neritic environments. Surface water conditions remained warm and tropical, and the continental environment exhibited an open vegetation landscape dominated by herbaceous plants, reflecting a tropical to subtropical arid climate conducive to erosion.
Post 6.35 Ma, the environment evolved towards the degradation of marine conditions, as evidenced by the rarity of the planktonic microfauna. At 6.08 Ma, the formation of Halimeda algal beds commenced, overlying the marl-diatomite cyclic sequence.
In summary, during the pre-evaporitic Messinian, the Melilla-Nador Basin underwent the influence of two main factors contributing to the formation of the marl-diatomite series: 1) tectonic activity causing a decrease in bathymetry and progressive uplift and 2) global parameters including 2a) a warm, arid climate favoring herbaceous plant spread, and 2b) episodic oceanographic phenomena such as upwelling. These findings support and align with previous results reported by Pelligrino et al. (2018).
This work benefited from the technical equipment of the Geosciences and Applications Laboratory, Geology Department, Faculty of Sciences Ben M'sik, Hassan II University of Casablanca (Morocco). Many thanks are extended to the individuals (reviewers and editors) who provided constructive comments to enhance the quality of this paper.
Achalhi M., MŁnch P., Cornťe J.-J., Azdimousa A., Melinte-Dobrinescu M., Quillťvťrť F., Drinia H., Fauquette S., Jimťnez-Moreno G., Merzeraud G., Moussa A.B., El Karim Y. & Feddi N. (2016).- The late Miocene Mediterranean-Atlantic connections through the North Rifian Corridor: New insights from the Boudinar and Arbaa Taourirt basins (northeastern Rif, Morocco).- Palśogeography, Palśoclimatology, Palśoecology, vol. 459, p. 131-152.
Agiadi K., Antonarakou A., Kontakiotis G., Kafousia N., Moissette P., Cornťe J.-J., Manoutsoglou E. & Karakitsios V. (2017).- Connectivity controls on the late Miocene eastern Mediterranean fish fauna.- International Journal of Earth Sciences, vol. 106, p. 1147-1159.
Assen E. van, Kuiper K.F., Barhoun N., Krijgsman W. & Sierro F.J. (2006).- Messinian astrochronology of the Melilla Basin: Stepwise restriction of the Mediterranean-Atlantic connection through Morocco.- Palśogeography, Palśoclimatology, Palśoecology, vol. 238, p. 15-31.
Bache F., Gargani J., Suc J.-P., Gorini C., Rabineau M., Popescu S.M., Leroux E., Do Couto D., Jouannic G., Rubino J.-L., Olivet J.-L., Clauzon G., Dos Reis A.T. & Aslanian D. (2015).- Messinian evaporite deposition during sea level rise in the Gulf of Lions (Western Mediterranean).- Marine and Petroleum Geology, vol. 66, no. 1, p. 262-277.
Bachiri Taoufiq N. (2000, unpublished).- Les environnements marins et continentaux du corridor rifain au MiocŤne supťrieur d'aprŤs la palynologie.- PhD Thesis, University Hassan II-Mohammedia, Casablanca, 212 p.
Bachiri Taoufiq N. & Barhoun N. ( 2001a).- La vťgťtation, le climat et l'environnement marin au Tortonien dans le bassin de Taza-Guercif (Corridor rifain, Maroc oriental) d'aprŤs la palynologie.- Gťobios, Villeurbanne, vol. 34, no. 1, p. 13-24.
Bachiri Taoufiq N., Barhoun N. & Suc J.-P. (2008).- Les environnements continentaux du corridor rifain (Maroc) au MiocŤne supťrieur d'aprŤs la palynologie.- Geodiversitas, Paris, vol. 30, no. 1, p. 41- 58.
Bachiri Taoufiq N., Barhoun N., Suc J.-P., Mťon H., Elaouad Z. & Benbouziane A. (2001b).- Environnement, vťgťtation et climat du Messinien au Maroc.- Paleontologia i Evoluciů, Sabadell, no. 32-33, p. 127-138.
Barhoun N. (2000).- Biostratigraphie et palťoenvironnement du MiocŤne supťrieur et du PliocŤne infťrieur du Maroc septentrional : Apport des foraminifŤres planctoniques.- ThŤse d'…tat Ťs Sciences, Universitť Hassan II-Mohammedia, Casablanca, Maroc, 272 p.
Barhoun N., Sierro F.J., El Hajjaji K. & Ben Bouziane A. (1999).- Biostratigraphie et palťoenvironnement du MiocŤne supťrieur du bassin de Zťghanghane (Rif nord-oriental, Maroc) : Apport des foraminifŤres planctoniques.- Revista EspaŮola de Micropaleontologia, Madrid vol. 31, no. 2, p. 279-287.
Bellier J.-P., Mathieu R. & Granier B. (2010).- Short Treatise on Foraminiferology (Essential on modern and fossil Foraminifera) [Court traitť de foraminifťrologie (L'essentiel sur les foraminifŤres actuels et fossiles)].- Carnets de Gťologie - Notebooks on Geology, Brest, Book 2010/02 (CG2010_B02), 104 p. (10 pls.). DOI: 10.4267/2042/33629
Benson R.H., Rakic El Bied K. & Bonaduce G. (1991).- An important current reversal in the Rifian Corridor (Morocco) at the Tortonian-Messinian boundary: The end of the Tethys Ocean.- Paleoceanography, vol. 6, p. 165-192.
Bertini A., Londeix L., Maniscalco R., Di Stefano A., Suc J.-P., Clauzon G., Gautier F. & Grasso M. (1998).- Paleobiological evidence of depositional conditions in the Salt Member, Gessoso-Solfifera Formation (Messinian, Upper Miocene) of Sicily.- Micropaleontology, vol. 44, no. 4, p. 413-433.
Bertini A. & Menichetti E. (2015).- Palaeoclimate and palaeoenvironments in central Mediterranean during the last 1.6 Ma before the onset of the Messinian Salinity Crisis: A case study from the Northern Apennine foredeep basin.- Review of Palaeobotany and Palynology, vol. 218, p. 218-106.
Bessedik M. (1984).- The early Aquitanian and upper Langhian-lower Serravalian environments in the northwestern Mediterranean region.- Paleobiology, vol. 14, no. 2, p. 153-179.
Bessedik M. (1985).- Reconstitution des environnements miocenes des rťgions nord-ouest mťditerranťennes ŗ partir de la palynologie.- ThŤse Doctorat Sciences, U.S.T.L., Montpellier, vol. 18, no. 6, 162 p.
Bessedik M., Guinet P. & Suc J.-P. (1984).- Donnťes palťofloristiques en Mťditerranťe nord-occidentale depuis l'Aquitanien.- Revue de Palťobiologie, GenŤve, volume special, p. 25-31.
Blanc-Valleron M.-M., Pierre C., Caulet J.-P., Caruso A., Rouchy J.-M., Gespuglio G., Sprovieri R., Pestrea S. & Di Stefano E. (2002).- Sedimentary, stable isotope and micropaleontological records of paleoceanographic change in the Messinian Tripoli Formation (Sicily, Italy).- Palśogeography, Palśoclimatology, Palśoecology, vol. 185, p. 255-286.
Capella W., Barhoun N., Flecker R., Hilgen F., Kouwenhoven T., Matenco L.C., Sierro F.J., Tulbure M.A., Yousfi M.Z. & Krijgsman W. (2018).- Palaeogeographic evolution of the late Miocene Rifian Corridor (Morocco): Reconstructions from surface and subsurface data.- Earth Science Reviews, vol. 180, p. 37-59.
Capella W., Flecker R., Hernandez-Molina F.J., Simon D., Meijer P.T., Rogerson M., Sierro F.J. & Krijgsman W. (2019).- Mediterranean isolation preconditioning the Earth System for late Miocene climate cooling.- Scientific Reports, vol. 9, article 3795, 8 p.
Chikhi H. (1992a).- Palynoflore du Messinien infra-ťvaporitique de la sťrie marno-diatomitique de Sahaouria (Beni-Chougrane) et de Chabet Bou Seter (Tessala), bassin du Chelif, Algťrie.- ThŤse, Universitť d'Oran, 169 p.
Chikhi H. (1992b).- Une palynoflore mťditerranťenne ŗ subtropicale au Messinien prť-ťvaporitique en Algťrie.- Gťologie Mťditerranťenne, Marseille, t. XIX, no. 1, p.19-30.
Choubert G., Faure-Muret A., Hottinger L. & LecoÓntre G. (1966).- Le Nťogene du bassin Melilla (Maroc septentrional) et sa signification pour definir la limite Mio-PliocŤne au Maroc.- Committee on Mediterranean Neogene stratigraphy, ICS-IUGS, p. 238-249.
CIESM (2008).- The Messinian Salinity Crisis from Mmega-deposits to m.- A Consensus Report, Monaco, vol. 30, p. 168.
Combaz A. (1980).- Les kťrogŤnes vus au microscope.- …dition Technip, Paris, 519 p.
CorbŪ H. & Soria J.M. (2016).- Late Miocenee early Pliocene planktonic foraminifer event stratigraphy of the Bajo Segura basin: A complete record of the western Mediterranean.- Marine and Petroleum Geology, vol. 77, p. 1010-1027.
Cornťe J.-J., Ferrandini M., Saint-Martin J.-P., MŁnch P., Moullade M., Ribaud-Laurenti A., Roger S., Saint-Martin S. & Ferrandini J. (2006).- The Late Messinian erosional surface and the subsequent reflooding in the Mediterranean: New insights from the Melilla-Nador Basin (Morocco).- Palśogeography, Palśoclimatology, Palśoecology, vol. 230, no. 1-2, p. 129-154.
Cornťe J.-J., Roger S., MŁnch P., Saint-Martin J.-P., Fťraud G., Conesa G., Pestrea-Saint-Martin S. (2002).- Messinian events: New constraints from sedimentological investigations and new 40ArR/39Ar ages in the Melilla-Nador Basin (Morocco).- Sedimentary Geology, vol. 151, no. 1-2, p. 127-147.
Cornťe J.-J., Saint-Martin J.-P., Conesa G., Muller J. & Andrť J.-P. (1996).- Anatomie de quelques plates-formes progradantes messiniennes de Mťditerranťe occidentale.- Bulletin de la Sociťtť gťologique de France, Paris, vol. 167, no. 4, p. 495-507.
Cour P. & Duzer D. (1978).- La signification climatique, ťdaphique et sťdimentologique des rapports entre taxons en analyse pollinique.- Annales des Mines de Belgique, Bruxelles, no. 7-8, p. 155-164.
Cunningham K.J., Farr M.R. & Rakic-El Bied K. (1994).- Magnetostratigraphic dating of an Upper Miocene shallow-marine and continental sedimentary succession in northeastern Morocco.- Earth Planetary Science Letters, vol. 127, p. 77-93.
Daget J. (1979).- Les modťles mathťmatiques en ťcologie.- Masson, Paris, 172 p.
Drinia H., Antonarakou A., Tsaparas N. & Kontakiotis G. (2007).- Palaeoenvironmental conditions preceding the Messinian salinity crisis: A case study from Gavdos Island.- Gťobios, Villeurbanne, vol. 40, p. 251-265.
Fauquette S., Suc J.-P., Guiot J., Filomena D., Feddi N., Zheng Z., Bessais E. & Drivaliari A. (1999).- Climate and biomes in the West Mediterranean area during the Pliocene.- Palśogeography, Palśoclimatology, Palśoecology, vol. 152, p. 15-36.
Fauquette S., Suc J.-P., Jimťnez-Moreno G., Micheels A., Jost A., Favre E., Bachiri-Taoufiq N., Bertini A., Clet-Pellerin M., Diniz F., Farjanel G., Feddi N. & Zheng Z. (2007).- Latitudinal climatic gradients in western European and Mediterranean regions from the Mid-Miocene (15 Ma) to the Mid-Pliocene (3.5 Ma) as quantified from pollen data. In: Williams M., Haywood A., Gregory J. & Schmidt D. (eds.), Deep time perspectives on climate change: Marrying the signal from computer models and biological proxies.- The Micropaleontological Society Special Publications, London, p. 481-502.
Flecker R., Krijgsman W., Capella W., Castro MartŪns C. de, Dmitrieva E., Mayser J.-P., Marzocchi A., Modestu S., Ochoa D., Simon D., Tulbure M., Berg B. van den, Schee M. van der, Lange G. de, Ellam R., Govers R., Gutjahr M., Hilgen F., Kouwenhoven T., Lofi J., Meijer P., Sierro F.J., Bachiri N., Barhoun N., Alami A.C., Chacon B., Flores J.A., Gregory J., Howard J., Lunt D., Ochoa M., Pancost R., Vincent S. & Yousfi M.Z. (2015).- Evolution of the Late Miocene Mediterranean-Atlantic gateways and their impact on regional and global environmental change.- Earth Science Reviews, vol. 150, p. 365-392.
Gaudant J., Saint-Martin J.-P., Benmoussa A., Cornťe J.-J., Hajjaji K.El. & Muller J. (1994).- L'ichtyofaune mesinienne ŗ la pťriphťrie de la plateforme carbonatťe de Melilla-Nador (Nord-Est du Maroc).- Gťologie Mťditerranťenne, vol. 21, no. 1-2, p. 25-35.
Gibson T.G. (1989).- Planktonic benthonic foraminiferal ratios: Modern patterns and Tertiary applicability.- Marine Micropaleontology, vol. 15, no. 1-2, p. 29-52.
Gorini C., Montadert L. & Rabineau M. (2015a).- New imaging of the salinity crisis: Dual Messinian lowstand megasequences recorded in the deep basin of both the eastern and western Mediterranean.- Marine and Petroleum Geology, vol. 66, p. 1-17.
Gorini C., Suc J.-P. & Rabineau M. (2015b).- Le dťluge et la Crise messinienne. In: Euzen A., Jeandel C.& Mosseri R. (eds.), L'eau ŗ dťcouvert.- CNRS …ditions, Paris, p. 98-99.
Guillemin M. & Houzay J.-P. (1982).- Le NťogŤne post-nappe et le Quaternaire du Rif nord-oriental (Maroc). Stratigraphie et tectonique des bassins de Melilla, du Kert, de Boudinar et du piedmont des Kebdena.- Notes et Mťmoires du Service Gťologique du Maroc, Rabat, no. 314, p. 7-238.
Hilgen F.J., Bissoli L., Iaccarino S., Krijgsman W., Meijer R., Negri A. & Villa G. (2000a).- Integrated stratigraphy and astrochronology of the Messinian GSSP at Oued Akrech (Atlantic Morocco).- Earth and Planetary Science Letters, vol. 182, no. 3-4, p. 237-251.
Hilgen F.J., Iaccarino S., Krijgsman W., Villa G., Langereis C.G. & Zachariasse W.J. (2000b).- The global boundary stratotype section and point (GSSP) of the Messinian Stage (uppermost Miocene).- Episodes, Beijing, vol. 23, no. 3, p. 172-178.
Hilgen F.J. & Krijgsman W. (1999).- Cyclostratigraphy and astrochronology of the Tripoli diatomite formation pre-evaporite Messinian, Sicily Italy.- Terra Nova, vol. 11, p. 16-22.
Hilgen F.J., Krijgsman W., Langereis C.G., Lourens L.J., Santarelli A. & Zachariasse W.J. (1995).- Extending the astronomical polarity time scale into the Miocene.- Earth and Planetary Science Letters, vol. 136, p. 495-510.
Hodell D.A., Benson R.H., Kent D.V., Boersma A. & Rakic-El Bied K. (1994).- Magnetostratigraphy, biostratigraphy, and stable isotope stratigraphy of a late Miocene drill core from the Salť Briqueterie Northwestern Morocco: A high-resolution chronology for the Messinian Stage.- Paleoceanography, vol. 9, p. 835-855.
Hodell D.A., Curtis J., Sierro F.J. & Raymo M. (2001).- Correlation of late Miocene to early Pliocene sequences between the Mediterranean and North Atlantic.- Paleoceanography, vol. 16, no. 2, p. 164-178.
HsŁ K.J., Cita M.B. & Ryan W.B.F. (1973).- The origin of the Mediterranean evaporites. In: Ryan W.B.F. & Hsu K.J. (eds.), Initial Reports of the Deep Sea Drilling Project 12.- U.S. Government Printing Office, Washington - DC, p. 1203-1231.
HsŁ K.J., Montadert L., Bernoulli D., Cita M.B., Erickson A., Garrison R.E., Kidd R.B., MťliŤres F., MŁller C. & Wright R.C. (1977).- History of the Mediterranean salinity crisis.- Nature, vol. 267, p. 399-403.
HŁsing S.K., Kuiper K.F., Link W., Hilgen F.J. & Krijgsman W. (2009).- The upper TortonianĖlower Messinian at Monte dei Corvi (Northern Apennines, Italy): Completing a Mediterranean reference section for the Tortonian Stage.- Earth and Planetary Science Letters, vol. 282, no. 1Ė4, p. 140-157.
Jimťnez-Moreno G., Pťrez-Asensio J.N., LarrasoaŮa J.C., Aguirre J., Civis J., Rivas-Carballo M.R., Valle-Hernŗndez M.F. & Gonzŗlez-Delgado J.A. (2013).- Vegetation, sea-level and climate changes during the Messinian salinity crisis.- Geological Society of America Bulletin, Boulder - CO, vol. 125, p. 432-444.
Karakitsios V., Cornťe J.-J., Tsouroua T., Moissette P., Kontakiotis G., Agiadia K., Manoutsogloud E., Triantaphylloua M., Koskeridoua E., Drinia H. & Roussosa D. (2017b).- Messinian salinity crisis record under strong freshwater input in marginal, intermediate, and deep environments: The case of the North Aegean.- Palśogeography, Palśoclimatology, Palśoecology, vol. 485, p. 316-335.
Karakitsios V., Roveri M., Lugli S., Manzi V., Gennari R., Antonarakou A., Triantaphyllou M., Konstantina Agiadi K., Kontakiotis G., Kafousia N. & Marc R. (2017a).- A record of the Messinian salinity crisis in the eastern Ionian tectonically active domain (Greece, eastern Mediterranean).- Basin Research, vol. 29, p. 203-233.
Kontakiotis G., Besiou E., Antonarakou A., Zarkogiannis S.D., Kostis A., Mortyn P.G., Moissette P., Cornťe J.J., Schulbert C., Drinia H., Anastasakis G. & Karakitsios V. (2019).- Decoding sea surface and paleoclimate conditions in the eastern Mediterranean over the Tortonian-Messinian transition.- Palśogeography, Palśoclimatology, Palśoecology, vol. 534, article 109312, 16 p.
Kontakiotis G., Karakitsios V., Maravelis A.G., Zarkogiannis S.D., Agiadi K., Antonarakou A., Pasadakis N. & Zelilidis A. (2020).- Integrated isotopic and organic geochemical constraints on the depositional controls and source rock quality of the Neogene Kalamaki sedimentary successions (Zakynthos Island, Ionian Sea).- Mediterranean Geoscience Reviews, vol. 3, p. 193-217.
Krijgsman W., Capella W., Simon D., Hilgen F.J., Kouwenhoven T.J., Meijer P.T., Sierro F.J., Tulbure M.A., Berg B.C.J. van den, Schee M. van der & Flecker R. (2018).- The Gibraltar corridor: Watergate of the Messinian salinity crisis.- Marine Geology, vol. 403, p. 238-246.
Krijgsman W., Hilgen F.J., Langereis C.G., Lourens L.J., Santarelli A. & Zachariasse W.J. (1995).- Late Miocene magnetostratigraphy, biostratigraphy and cyclostratigraphy in the Mediterranean.- Earth and Planetary Science Letters, vol. 136, p. 475-494.
Krijgsman W., Hilgen F., Langereis C.G. & Zachariasse W.J. (1994).- The age of the Tortonian/Messinian boundary.- Earth and Planetary Science Letters, vol. 121, p. 533-547.
Krijgsman W., Hilgen F.J., Raffi I., Sierro F.J. & Wilson D.S. (1999a).- Chronology, causes and progression of the Messinian Salinity Crisis.- Nature, vol. 400, p. 652-655.
Krijgsman W., Langereis C.G., Zachariasse W.J., Boccaletti M., Moratti G., Gelati R., Iaccarino S., Papani G. & Villa G. (1999b).- Late Neogene evolution of the Taza-Guercif Basin (Rifian Corridor, Morocco) and implications for the Messinian Salinity Crisis.- Marine Geology, vol. 153, p. 147-160.
Lirer F., Foresi L.M., Iaccarino S.M., Salvatorini G., Turco E., Cosentino C., Sierro F.J. & Caruso A. (2019).- Mediterranean Neogene planktonic foraminifer biozonation and biochronology.- Earth Science Reviews, vol. 196, article 102869, 36 p.
Londeix L., Benzakour M., Suc J.-P. & Turon J.-L. (2007).- Messinian palaeoenvironments and hydrology in Sicily (Italy): The dinoflagellate cyst record.- Gťobios, Villeurbanne, vol. 40, no. 3, p. 233-250.
Lourens L.J., Hilgen F.J., Laskar J., Shackleton N.J. & Wilson D.S. (2004).- The Neogene Period. In: Gradstein F.M., Ogg J.G. & Smith A.G. (eds.), A Geologic Time Scale 2004.- Cambridge University Press, Cambridge (UK), p. 409-440.
Marret F., Vernal A. de, Benderra F. & Harland R. (2001).- Late Quaternary sea-surface conditions at DSDP Site 594 in the southwest Pacific Ocean based on dinoflagellate cyst assemblages.- Journal of Quaternary Science, vol. 16, no. 7, p. 739-749.
MartŪn J.M., Braga J.C. & Betzler C. (2001).- The Messinian Guadalhorce corridor: The last northern, Atlantic-Mediterranean gateway.- Terra Nova, vol. 13, p. 418-424.
Mathieu R. (1986).- Sťdiments et foraminifŤres actuels de la marge continentale atlantique du Maroc.- ThŤse de Doctorat d'…tat Ťs Sciences naturelles, Universitť Pierre et Marie Curie - Paris 6; Mťmoires des Sciences de la Terre, Paris, no. 86-14, 419 p.
Mathieu R. (1988).- ForaminifŤres actuels et rťsurgences cŰtiŤres sur la marge continentale atlantique du Maroc. In: Benthos'86.- Revue de Palťobiologie, GenŤve, vol. spťcial, no. 2, Partie II, p. 845-850.
Moissette A.P., Cornťe C., Antonarakoua A., Kontakiotisa G., Driniaa H, Koskeridoua E., Tsouroua T., Agiadia K. & Karakitsiosa V. (2018).- Palaeoenvironmental changes at the Tortonian/Messinian boundary: A deep-sea sedimentary record of the eastern Mediterranean Sea.- Palśogeography, Palśoclimatology, Palśoecology, vol. 505, p. 217 -233.
Morzadec-Kerfourn M.T. (1976).- La signification ťcologique des dinoflagellťs et leur intťrÍt pour l'ťtude des variations du niveau marin.- Revue de Micropalťontologie, Paris, vol. 18, no. 4, p. 229-235.
MŁnch P., Cornťe J.-J., Fťraud G., Saint-Martin J.-P., Ferrandini M., Garcia F., Conesa G., Moullade M. & Roger S. (2006).- Precise 40Ar/39Ar chronostratigraphy of the upper Messinian Melilla carbonate complex (NE Morocco): Implications for the Messinian Salinity Crisis.- International Journal of Earth Sciences, vol. 95, p. 491-503.
MŁnch P., Roger S., Cornťe J.-J., Saint-Martin J.-P., Fťraud G. & Ben Moussa A. (2001).- Restriction des communications entre l'Atlantique et la Mťditerranťe au Messinien : Apports de la tťphrochronologie dans la plate-forme carbonatťe et le bassin de Melilla-Nador (Rif nord-oriental, Maroc).- Comptes Rendus de l'Acadťmie des Sciences, Paris, vol. 332, p. 569-576.
Murray J.W. (1991).- Ecology and palaeoecology of benthic foraminifera.- Longman Scientific & Technical, Harlow, 397 p.
Nix H. (1982).- Environmental determinant of biogeography and evolution in Terra Australis. In: Barker W.R. & Greenslade P.J.M. (eds.), Evolution of the flora and fauna of arid Australia.- Peacock Publications, Adelaide, p. 47-66.
Pellegrino L., Dela Pierre F., Natalicchio M. & Carnevale G. (2018).- The Messinian diatomite deposition in the Mediterranean region and its relationships to the global silica cycle.- Earth-Science Reviews, vol. 178, p. 154-176.
Poumot C. & Suc J.-P. (1994).- PalynofaciŤs et dťpŰts sťquentiels dans des sťdiments marins du NťogŤne.- Bulletin des Centres de Recherches Exploration-Production elf-Aquitaine, Pau, vol. 18, p. 107-119.
Prell W.L. (1984).- Variation of monsoonal upwelling: A response to changing solar radiation. In: Hansen J.E. & Takahashi T. (eds.), Climate processes and climate sensitivity.- M. Ewing - Geophysical Monograph 29, Washington - DC, vol. 5, p. 48-57.
Rachid A., El Hajjaji K. & Ben Moussa A. (1997).- Les associations foraminifŤres benthiques des sťries marno-diatomitiques messiniennes du bassin de Nador-Melilla (Maroc nord oriental).- Gťologie Mťditerranťenne, Marseille, T. XXIV 24, no. 1-2, p. 29-49.
Roger S., MŁnch P., Cornťe J.-J., Saint-Martin J.-P., Fťraud G., Pestrea S., Conesa G. & Ben Moussa A. (2000).- 40Ar/39Ar dating of the pre-evaporitic Messinian marine sequences of the Melilla basin (Morocco): A proposal or some bio-sedimentary events as isochrons around the Alboran Sea.- Earth and Planetary Science Letters, vol. 179, p. 101-113.
Rouchy J.M. (1982).- La genŤse des ťvaporites messiniennes de Mťditerranťe.- Mťmoires du Musťum National d'Histoire Naturelle (sťrie C), Paris, vol. 50, p. 1-267.
Rouchy J.-M., Pierre C., Et-Touhami M., Kerzazi K., Caruso A. & Blanc-Valleron M.M. (2003).- Late Messinian to Early Pliocene paleoenvironmental changes in the Melilla Basin (NE Morocco) and their relation to Mediterranean evolution.- Sedimentary Geology, vol. 163, p. 1-27.
Roveri M., Flecker R., Krijgsman W., Lofi J., Lugli S., Manzi V., Sierro F.J., Bertini A., Camerlenghi A., Lange G. de, Govers R., Hilgen F.J., HŁbscher C., Meijer P.T. & Stoica M. (2014).- The Messinian Salinity Crisis: Past and future of a great challenge for marine sciences.- Marine Geology, vol. 352, p. 25-58.
Saint-Martin J.-P. (1990).- Les formations rťcifales coralliennes du MiocŤne supťrieur d'Algťrie et du Maroc.- Mťmoires du Musťum National d'Histoire Naturelle, (sťrie C), Paris, t. 56, 366 p.
Saint-Martin J.-P. & Cornťe J.-J. (1996).- The Melilla reef complex (Northeastern Rif, Morocco). In: Franseen E. et al. (eds.), Models for Carbonate Stratigraphy from Miocene Reef Complexes of the Mediterranean Regions.- Concepts in Sedimentology and Paleontology, vol. 5, p. 227-237.
Saint-Martin J.-P., Cornťe J.-J., Muller J., Camoin G., Andrť J.-P., Rouchy J.-M. & Benmoussa A. (1991).- ContrŰles globaux et locaux dans l'ťdification d'une plate-forme carbonatťe messinienne (bassin de Melilla, Maroc) : Apport de la stratigraphie sťquentielle et de l'analyse tectonique.- Comptes Rendus de l'Acadťmie des Sciences (Sťrie II), Paris, t. 312, p. 1573-1579.
Saint-Martin J.-P. & Rouchy J.-M. (1986).- IntťrÍt du complexe rťcifal du Cap des Trois Fourches (Bassin de Nador, Maroc septentrional) pour l'interprťtation palťogťographique des ťvťnements messiniens en Mťditerranťe occidentale.- Comptes Rendus de l'Acadťmie des Sciences (Sťrie II), Paris, t. 302, p. 957-962.
Saint-Martin J.-P. & Rouchy J.-M. (1990).- Les plates-formes carbonatťes messiniennes en Mťditerranťe occidentale : Leur importance pour la reconstitution des variations du niveau marin au MiocŤne terminal.- Bulletin de la Sociťtť gťologique de France, Paris, vol. 8, no. 1, p. 83-94.
Saint-Martin S., Conesa G. & Saint-Martin J.-P. (2003).- Signification palťoťcologique des assemblages de diatomťes du Messinien dans le bassin de Melilla-Nador (Rif Nord-Oriental, Maroc).- Revue de Micropalťontologie, Paris, vol. 46, p. 161-190.
Shannon C.E. & Weaver W. (1949).- The mathematical theory of communication.- University of Illinois Press, Urbana - IL, 144 p.
Sierro F.J. (1985).- The replacement of the "Globorotalia menardii" group by the Globorotalia miotumida group: An aid to recognizing the Tortonian/Messinian boundary in the Mediterranean and adjacent Atlantic.- Marine Micropaleontology, vol. 9, p. 525-535.
Sierro F.J., Flores J.A., Civis J., GonzŠlez-Delgado J.A. & Frances G. (1993).- Late Miocene Globorotaliid event-stratigraphy and biogeography in the NE-Atlantic and Mediterranean.- Marine Micropaleontology, vol. 21, p. 143-167.
Sierro F.J., Flores J.A., Zamarreno I., Vasquez A., Utrilla R., Francťs G., Hilgen F.J. & Krijgsman W. (1999).- Messinian pre-evaporite sapropels and precession-induced oscillations in western Mediterranean climate.- Marine Geology, vol. 153, p. 137-146.
Sierro F.J., Hilgen F.J., Krijgsman W. & Flores J.A. (2001).- The Abad composite SE Spain: A Messinian reference section for the Mediterranean and the APTS.- Palśogeography, Palśoclimatology, Palśoecology, vol. 168, p. 141-169.
Suc J.-P. (1986).- Flores NťogŤnes de mťditerranťe occidentale. Climat et palťogťographie.- Bulletin des Centres de Recherches Exploration-Production elf-Aquitaine, Pau, vol. 10, no. 2, p. 477-488.
Suc J.-P. (1989).- Distribution latitudinale et ťtagement des associations vťgťtales au CťnozoÔque supťrieur dans l'aire ouest-mťditerranťenne.- Bulletin de la Sociťtť gťologique de France, Paris (8e Sťrie), vol. 3, p. 541-550.
Suc J.-P. & Bessais E. (1990).- Pťrennitť d'un climat thermo-xťrique en Sicile avant, pendant, aprŤs la crise de salinitť messinienne.- Comptes rendus de l'Acadťmie des Sciences (Sťrie II), Paris, vol. 310, p. 1701-1707.
Suc J.-P. & Frizon de Lamotte D. (2019).- Palťoenvironnements mťditerranťens : Changements et bouleversements environnementaux depuis 23 Ma.- Gťochronique, Paris, no. 151, p. 16-17.
Suc J.-P., Popescu S.M., Fauquette S., Bessedik M., Jimťnez-Moreno G., Bachiri Taoufiq N., Zheng Z., Mťdail F. & Klotz S. (2018).- Reconstruction of Mediterranean flora, vegetation and climate for the last 23 million years based on an extensive pollen dataset.- Ecologia mediterranea, Avignon, vol. 44, no. 2, p. 53-85.
Suc J.-P., Saint-Martin J.-P., Saint-Martin S., Popescu S.M. & Fauquette S. (2019).- …volution des environnements marins et continentaux depuis 23 Ma dans le domaine mťditerranťen.- Gťochronique, Paris, no. 151, p. 18-23.
Targhi S., Barhoun N., Taoufiq N.B., Achab M., Ait Salem A. & Yousfi M.Z. (2021).- Vegetation climate and marine environmental reconstruction in the western Mediterranean (southern Rifian corridor, Morocco) over the Tortonian-Messinian transition.- Heliyon, vol. 7, no. 12, article e08569, 19 p.
Targhi S., Barhoun N., Taoufiq N.B., Achab M., Essamoud R., Rahmouna J. & Berry N. (2023).- Late Miocene biostratigraphy and paleoenvironment of the Sais Basin (southern Rifian Corridor, Morocco): New insights from the Moulay Yakoub area.- Carnets Geol., Madrid, vol. 23, no. 3, p. 53-75. DOI: 10.2110/carnets.2023.2303
Thiede J. (1983).- Skeletal plankton and nekton in upwelling water masses off northwestern South America and northwest Africa. In: Suess E. & Thiede J. (eds.), Coastal Upwelling.- Plenum, New York - NY, p. 183-207.
Tulbure M.A., Capella W., Barhoun N., Flores J.A., Hilgen F.J., Krijgsman W., Kouwenhoven T., Sierro F.J. & Yousfi M.Z. (2017).- Age refinement and basin evolution of the North Rifian Corridor (Morocco): No evidence for a marine connection during the Messinian salinity crisis.- Palśogeography, Palśoclimatology, Palśoecology, vol. 485, p. 416-432.
Tyson R.V. (1993).- Palynofacies analysis. In: Applied micropalaeontology.- Springer, Dordrecht, p. 153-191.
Vasiliev I., Karakitsios V., Bouloubassi I., Agiadi K., Kontakiotis G., Antonarakou A., Triantaphyllou M., Gogou A., Kafousia N., Rafťlis M. de, Zarkogiannis S., Kaczmar F., Parinos C. & Pasadakis N. (2019).- Large sea surface temperature, salinity, and productivity-preservation changes preceding the onset of the Messinian Salinity Crisis in the eastern Mediterranean Sea.- Paleoceanography and Paleoclimatology, vol. 34, no. 2, p. 182-202.
Vasiliev I., Reichart G.J. & Krijgsman W. (2013).- Impact of the Messinian Salinity Crisis on Black Sea hydrology - Insights from hydrogen isotopes analysis on biomarkers.- Earth and Planetary Science Letters, vol. 362, p. 272-282.
Vernal A. de, Henry M., Matthiessen J., Mudie P.J., Rochon A., Boessenkool K.P., Eynaud F., Grosfjeld K., Guiot J., Hamel D., Harland R., Head M.J., Kunz-Pirrung M., Levac E., Loucheur V., Peyron O., Pospelova V., Radi T., Turon J.-L. & Voronina E. (2001).- Dinoflagellate cyst assemblages as tracers of sea-surface conditions in the northern North Atlantic, Arctic and sub-Arctic seas: The new 'n = 677' data base and its application for quantitative palaeoceanographic reconstruction.- Journal of Quaternary Science, vol. 16, no. 7, p. 681-698.
Vernal A. de & Marret F. (2007).- Organic-walled dinoflagellate cysts: Tracers of sea-surface conditions.- Developments in Marine Geology, vol. 1, p. 371-408.
Vernal A. de, Rochon A., Turon J.-L. & Matthiessen J. (1998).- Organic walled dinoflagellate cysts: Palynological tracers of sea-surface conditions in middle to high lattitude marine environments.- Gťobios, Villeurbanne, vol. 30, p. 905-920.
Vernal A. de, Turon J.-L. & Guiot J. (1994).- Dinoflagellate cyst distribution in high latitude marine environments and quantitative reconstruction of sea surface salinity, temperature and seasonality.- Canadian Journal of Earth Sciences, vol. 31, no. 1, p. 48-62.
Versteegh G.J.M. (1994).- Recognition of cyclic and non-cyclic environmental changes in the Mediterranean Pliocene; a palynological approach.- Marine Micropaleontology, vol. 23, p. 147-171.
Warny S. (1999).- Marine and continental environmental changes in the Gibraltar Arc Area during the late Neogene (8-2.7 Ma) linked to the evolution of global climate and to Atlantic Ocean-Mediterranean Sea relationship. A palynological contribution to the Mediterranean Messinian Salinity Crisis through dinoflagellate cysts and pollen analysis.- ThŤse, Universitť Catholique de Louvain, 295 p.
Warny S., Bart P.J. & Suc J.-P. (2003).- Timing and progression of climatic tectonic and glacioeustatic influences on the Messinian salinity crisis.- Palśogeography, Palśoclimatology, Palśoecology, vol. 202, p. 59-66.
Zonneveld K.A.F. (1995).- Palaeoclimatical and palaeo-ecological changes during the last deglaciation in the Eastern Mediterranean; implications for dinoflagellate ecology.- Review of Palaeobotany and Palynology, vol. 84, p. 221-253.
Zonneveld K.A.F. (1996).- Palaeoclimatic and palaeo-ecologic changes in the Eastern Mediterranean and Arabian Sea regions during the last deglaciation: A palynological approach to land-sea correlation.- Universiteit Utrecht, Series 3, p. 1-199.
Zonneveld K.A.F., Marret F., Versteegh G.J.M., Bogus K., Bonnet S., Bouimetarhan I., Crouch E., Vernal A. de, Elshanawany R., Edwards L., Esper O., Forke S., GrÝsfjeld K., Henry M., Holzwarth U., Kielt J.F., Kim S.Y., Ladouceur S., Ledu D. & Young M. (2013).- Atlas of modern dinoflagellate cyst distribution based on 2405 data points.- Review of Palaeobotany and Palynology, vol. 191, p. 1-197.
Plate
1: a-b:
Globorotalia miotumida (Jenkins, 1960),
SHY-3, Sidi El Haj Youssef section, Boudinar basin;
c-d: Globorotalia conomiozea (Kennett, 1966), SHY-3, Sidi El Haj Youssef
section, Boudinar basin;
e: Globorotalia nicolae Catalano & Sprovieri, 1971,
IZA 22, IzarorŤne section, Melilla-Nador basin; f-g: Globorotalia nicolae
Catalano & Sprovieri, 1971,
IZA 23, IzarorŤne section, Melilla-Nador basin;
h: Sphaeroidinellopsis semilunata (Schwager,
1866), IZA 3, IzarorŤne section, Melilla-Nador basin; i-j: Neogloboquadrina acostaensis (Blow, 1959) with dextral coiling, MES 28, Messadit
section, Melilla-Nador basin; k: Neogloboquadrina acostaensis (Blow, 1959) with
sinistral coiling, MES 14, Messadit section, Melilla-Nador basin;
l: Turborotalita multiloba (Romeo, 1965),
IFO 14, Ifounassene section, Melilla-Nador basin; m: Turborotalita quinquleba (Natland, 1938), SHY-17, Sidi El Haj Youssef
section, Boudinar basin. All photomicrographs have the same scale (scale bar = 100 Ķm). |
|
Plate
2:
Pollen: a: Cathaya, IFO 10; b: Lygeum,
IFO 11, subdesertic; c: Quercus, IFO 6, mesotherm; d:
Engelhardia, IFO 4, mega-mesotherm; e: Poaceae, IFO 1, herb; f: Olea,
IFO 4, Mediterranean xerophyte; g: Ephedra, IFO 11, subdesertic;
h-i: Asteraceae, IFO 10, herb; j: Amaranthaceae - Chenopodiaceae, IFO
5, halophyte. Dinocysts: k: Impagidinium
spp., IFO 13; l: Spiniferites spp., IFO 4; m: Nematospheropsis labyrinthus
(Ostenfeld, 1903), IFO 2; n: Operculodinium israelianum
(Rossignol, 1962), IFO 10; o: Lingulodinium machaerophorum (Deflandre &
Cookson, 1955), IFO 7. |
|