◄ Carnets Geol. 24 (6) ►
![]()
Outline:
[1. Introduction]
[2. Material and methods]
[3. Results]
[4. Discussion]
[5. Conclusion]
[Bibliographic references]
and ...
[Appendix]
Corresponding author;
State Key
Laboratory of Palaeobiology and Oil Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese
Academy of Sciences, Nanjing 210008 (China);
Lebanese University, Faculty of Science II, Natural Sciences
Department, Fanar - El-Matn, PO box 26110217 (Lebanon)
State Key
Laboratory of Palaeobiology and Oil Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese
Academy of Sciences, Nanjing 210008 (China);
Lebanese University, Faculty of Science II, Natural Sciences
Department, Fanar - El-Matn, PO box 26110217 (Lebanon)
State Key
Laboratory of Palaeobiology and Oil Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese
Academy of Sciences, Nanjing 210008 (China)
Mazraat Yachouh, El-Maten (Lebanon)
State Key
Laboratory of Palaeobiology and Oil Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese
Academy of Sciences, Nanjing 210008 (China)
Published online in final form (pdf) on April 1, 2024
DOI 10.2110/carnets.2024.2406
![]()
[Editor:
Jordi P�rez-Cano; language editor: Simon F. mitchell; technical editor: Bruno R.C. Granier]
![]()
A new species, Nannotanyderus granieri sp. nov., belonging to the Tanyderidae (Diptera, Nematocera), from Lower Barremian amber of Bqaatouta (Lebanon), is characterized, described, figured, and its taxononomic position discussed. The possible confusion regarding the attribution of new fossils to either the Tanyderidae or the Psychodidae, in addition to the results of recent molecular phylogenies, have led us to re-evaluate the taxonomic position of the Tanyderidae and to consider it as a subfamily within the Psychodidae. The fossil described herein is a tiny tanyderine sensu nov. species, and the second one belonging to the genus Nannotanyderus from Lebanese amber. A tentative molecular phylogeny of recent Psychodidae and an exhaustive catalogue of fossil Tanyderinae stat. nov. are provided.
� Diptera;
� Tanyderidae;
� Psychodidae;
� Tanyderinae stat.
nov.;
� Tanyderini;
� Nannotanyderini;
� Lower Cretaceous;
� Lebanon
Azar D., Maksoud S., Huang D.-Y., Maalouf M. & Cai C.-Y. (2024).- A new fossil psychodomorphan fly from Lower Barremian Lebanese amber elucidates the relationship of the Tanyderinae stat. nov. within the Psychodidae.- Carnets Geol., Madrid, vol. 24, no. 6, p. 113-125.
Une nouvelle mouche psychodomorphe fossile de l'ambre Barr�mien inf�rieur du Liban clarifie la relation des Tanyderinae stat. nov. au sein des Psychodidae.- Une nouvelle esp�ce, Nannotanyderus granieri sp. nov., appartenant aux Tanyderidae (Diptera, Nematocera), provenant de l'ambre du Barr�mien inf�rieur de Bqaatouta (Liban), est caract�ris�e, d�crite et illustr�e ; sa position taxonomique est discut�e. En plus des r�sultats des phylog�nies mol�culaires r�centes, la possible confusion concernant l'attribution de nouveaux fossiles soit aux Tanyderidae, soit aux Psychodidae, nous a amen�s � r��valuer la position taxonomique des Tanyderidae et � les consid�rer comme une sous-famille au sein des Psychodidae. Le fossile d�crit ici est une esp�ce tanyd�rine sensu nov. et le deuxi�me appartenant au genre Nannotanyderus de l'ambre libanais. Une phylog�nie mol�culaire provisoire des Psychodidae actuels et un catalogue exhaustif des Tanyderinae stat. nov. fossiles sont fournis.
� Diptera ;
�
Tanyderidae ;
�
Psychodidae ;
�
Tanyderinae stat.
nov. ;
�
Tanyderini ;
�
Nannotanyderini ;
�
Cr�tac� inf�rieur ;
�
Liban
The Tanyderidae Osten-Sacken, 1880, colloquially known as primitive crane flies, are a small relictual cryptic cosmopolitan family of mostly aquatic flies, with 39 known living species worldwide (Madriz et al., 2018) across ten genera, i.e., Araucoderus Alexander, 1929, Eutanyderus Alexander, 1928, Mischoderus Handlirsch, 1909, Neoderus Alexander, 1928, Nothoderus Alexander, 1928, Peringueyomyina Alexander, 1921, Protanyderus Handlirsch, 1909, Protoplasa Osten-Sacken, 1859, Radinoderus Handlirsch, 1909, and Tanyderus Philippi, 1865. The genus Protanyderus Handlirsch, 1909, is represented in both fossil and extant faunas (Alexander, 1932; Krzemiński & Judd, 1997; Exner & Craig, 1976). Immature stages of extant species are aquatic to semiaquatic, often associated with wet sandy soil and the outer layers of submerged rotting logs in streams (Krzemiński & Judd, 1997). Adults are small to large flies (5-35 mm) and typically exhibit conspicuous mouthparts, with an elongated rostrum in some species. The family is characterized by elongate cervical sclerites in some species, creating the appearance of a long neck, which serves as the basis for the family name ['tany' from Greek (τανύω) meaning 'stretch', and 'dero' from Greek (δέρω) meaning skin]. Their legs are as long as, or longer than, the body. Males sometimes form large swarms in the evening, while spending daylight hours among riparian vegetation near stream borders. Little is known about the biology of these unusual flies.
The Tanyderidae have been understudied, which poses challenges for their placement within an evolutionary context (Madriz Villanueva, 2017). Considered among the rarest of Diptera (Crampton, 1926b; Krzemiński & Judd, 1997; Judd, 2004), tanyderids have long been described as 'primitive' (Crampton, 1930; Alexander, 1930; Williams, 1933; Colless & McAlpine, 1991; Rohdendorf, 1974; Exner & Craig, 1976). Phylogenetic hypotheses based on morphological characters of immature stages (Alexander, 1930), adult thoracic sclerites (Crampton, 1926a, 1926b, 1926c, 1930), and wing venation (Rohdendorf, 1974) have yielded inconsistent results regarding sister-group relationships, leading to a long-standing unresolved phylogenetic placement. Morphological phylogenetic analyses have suggested a close relationship between the Tanyderidae and Ptychopteridae (Hennig, 1973; Wood & Borkent, 1989; Oosterbroek & Courtney, 1995) and, more recently, the Blephariceridae (Lambkin et al., 2013). In contrast, recent analyses based on molecular data strongly support a sister group relationship between the Blephariceridae and Tanyderidae + Psychodidae (Bertone et al., 2008; Wiegmann et al., 2011; Curler & Moulton, 2012).
The Tanyderidae are relatively uncommon as fossils, yet they are well represented and diverse compared to recent extant tanyderids. The oldest known representative is Nannotanyderus oliviae Skibińska et al., 2014, which was described from the Lower Jurassic of England (Sinemurian, ca. 199.5 192.9 Ma). To date, 34 fossil species, representing ten genera, have been described, including Coramus Skibińska, 2016, Dacochile Poinar & Brown, 2004, Espanoderus Skibińska et al., 2019, Macrochile Loew, 1850, Podemacrochile Krzemiński & Kania in Krzemiński et al., 2013, Nannotanyderus Ansorge, 1994, Praemacrochile Kalugina in Kalugina & Kovalev, 1985, Protanyderus Handlirsch, 1909, and Similinannotanyderus Dong et al., 2015. A complete list of fossil Tanyderidae (or Tanyderinae stat. nov.) is provided in the appendix. The Psychodidae Newman, 1834, is a large family of small, hairy nematoceran dipterans, encompassing more than 2,600 described extant species (Azar & Maksoud, 2022a, 2022b). Presently, it is subdivided into seven subfamilies: Bruchomyiinae Alexander, 1920, Horaiellinae Enderlein, 1936, Phlebotominae Rondani, 1840, Psychodinae Newman, 1834, Sycoracinae Rondani, 1856, Trichomyiinae Tonnoir, 1922, and the extinct subfamily Datziinae (replacing Protopsychodinae Stebner et al., 2015a) known thus far only from mid-Cretaceous Burmese amber (Stebner et al., 2015a, 2015b).
Herein, we describe a new tiny tanyderine species (sensu the new classification proposed) that we assign to Nannotanyderus (based on wing shape and venation) from Lower Cretaceous amber of Bqaatouta. This marks the second species belonging to the genus Nannotanyderus recorded from Lebanese amber.
The material described from Lebanese amber herein originates from the Bqaatouta outcrop (Maksoud et al., 2022) in the Caza (District) of Kesserouan, Central Lebanon, from the Maalouf collection. Syn-inclusions include a spider and a male chironomid dipteran. The amber piece containing the inclusion was cut, shaped, and polished before being placed between two cover slips in Canada balsam (following the method described by Azar et al., 2003).
Images in reflected light were captured using an AxioCam MRc 5 camera attached to a Zeiss Discovery V16 stereo microscope. Widefield fluorescence images were obtained using a Zeiss Axio Imager 2 light microscope combined with a fluorescence imaging system. Confocal images were obtained with a Zeiss LSM710 confocal laser scanning microscope with 488 nm Argon laser excitation line (Cai & Huang, 2014; Fu et al., 2021). Photomicrographs were stacked using Helicon Focus 8.1.0. and processed with Adobe Photoshop CC 2019. The recent standard wing venation nomenclature of Kvifte and Wagner (2017) from the 'Manual of Afrotropical Diptera' was followed. All measurements are in micrometres ( m). The type specimen is housed in the collections of Natural History Museum of the Lebanese University, Faculty of Sciences II, located in Fanar, Lebanon.
In order to assess the interrelationships among the subfamilies of the Psychodidae and its relationship with the closely related Tanyderidae, we conducted a phylogenetic analysis utilizing Sanger-sequencing data available in GenBank. This dataset included 3 outgroup taxa (each species for Limoniidae, Ptychopteridae, and Tanyderidae) and 43 ingroup taxa representing each subfamily with the Psychodidae (Curler & Moulton, 2012). Two nuclear genes, the large nuclear ribosomal subunit (18S rDNA) and bromodomain and PHD finger-containing protein (Peregrin), were selected and utilized in our analyses. GenBank sequence numbers mostly follow those provided in Curler and Moulton (2012), with the exception of the following gene sequences: 1) the 18S rDNA gene of Ptychoptera albimana (KX453704) was incorporated with Ptychoptera sp., and 2) the problematic Peregrin gene of Trichomyia sp. nov. 2 (JF714438), which cannot be correctly aligned with the remaining species, was excluded from our analyses. All sequences were retrieved from GenBank using the Batch Entrez tool. The protein-coding gene (Peregrin) was aligned with respect to their codon structure using MACSE v2.06 (Ranwez et al., 2018). The ribosomal 18S gene was aligned using the Q-INS-I algorithm in MAFFT (Katoh et al., 2002) available through the MAFFT online service (Katoh et al., 2019). The concatenated matrix included 46 taxa and 5,206 nucleotide sites.
The selection of evolutionary models for tree inference can significantly impact phylogenetic analyses. Previous studies (Tihelka et al., 2020, 2021) have demonstrated that different models make various assumptions about the nature of the data, and violations of these assumptions can lead to misleading topologies, even with high statistical support. Therefore, we employed the site-heterogeneous CAT-GTR model to analyze our data. This model accounts for compositional heterogeneity across sites and can mitigate common sources of phylogenetic error such as long-branch attraction (Lartillot et al., 2007; Tihelka et al., 2021). The site-heterogeneous model CAT-GTR was implemented in PhyloBayes MPI1.7 (Lartillot et al., 2009), run at the Beijing Super Cloud Computing Center. Two independent Markov chain Monte Carlo (MCMC) chains were run until convergence. The bpcomp program was utilized to generate the largest (maxdiff) and mean (meandiff) values. The chains were considered to have converged when the maximum difference (maxdiff) was less than 0.3, with a recorded value of maxdiff = 0.13.
Order DIPTERA Linnaeus, 1758
Infraorder PSYCHODOMORPHA Hennig, 1968
Family PSYCHODIDAE Newman, 1834
former family Tanyderidae Osten-Sacken, 1880
Subfamily TANYDERINAE Osten-Sacken, 1880, stat. nov.
former family Tanyderidae Osten-Sacken, 1880
Tribe NANNOTANYDERINI Skibińska, 2016, stat. nov.
former subfamily Nannotanyderinae Skibińska, 2016
Genus NANNOTANYDERUS Ansorge, 1994
Type species. Nannotanyderus krzeminskii Ansorge, 1994, Lower Jurassic (Toarcian), Grimmen, Germany.
Nannotanyderus granieri sp. nov.
Material. Holotype (specimen BKT-11A), female, part of the Maalouf collection, Lebanese Lower Cretaceous amber found at the Bqaatouta outcrop, in the Caza (District) of Kesserouan, Central Lebanon. Syn-inclusions include a spider and a male chirbnomid dipteran. This material is housed in the collections of Natural History Museum of the Lebanese University, Faculty of Sciences II, located in Fanar, Lebanon.
Diagnosis. Female only. Head lacking an elongated neck; mouthparts well developed, longer than head, with well-developed sclerotized maxillae. R2 very short, slightly longer than half of R3, with R2+3 about 3 times longer than R2, hook-like at its basal part. Presence of cross-veins m-m and m-cu, as well as cell d.
N. granieri and N. ansorgei are the smallest members of the Tannyderidae, with female terminalia being simple, cerci terminating in a single lobe. The wing venation of N. granieri is very similar to that of N. ansorgei with the following differences: 1) R1 reaching costal margin is slightly beyond the fork of R2+3 into R2 and R3 in N. granieri, whereas it is opposite to it in N. ansorgei; 2) R2 slightly longer than half of R3 in N. granieri, whereas it is equal to half of R3 in N. ansorgei; 3) the bifurcation of R2+3 from Rs is slightly distal to the level where Sc meets the costal margin in N. granieri, whereas it is slightly basal to the level where Sc meets the costal margin in N. ansorgei; 4) the distance between the bifurcation of R2+3 from Rs to the base of R4 is smaller in N. granieri.
|
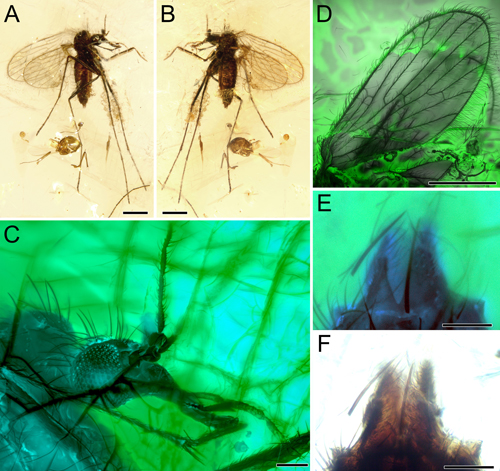
Figure 1:
Nannotanyderus
granieri sp. nov., holotype, female, specimen number BKT-11A. A)
Habitus, right lateral side. Scale bar: 500 m. B) Habitus, left lateral
side.
Scale bar: 500 m. C) Head photomicrograph with confocal microscope. Scale
bar: 100 m. D) Wing photomicrograph with confocal microscope. Scale bar: 500
m. E) Female terminalia, photomicrograph with confocal microscope. Scale
bar: 50 m. F) Female terminalia, photomicrograph with compound microscope.
Scale bar: 50 m. |
Etymology. The species is named after Prof. Bruno Granier, whose research has significantly advanced the dating of amber outcrops in Lebanon (e.g., Granier et al., 2015, 2016).
Description. The specimen is very small, measuring 1571 m in length from the tip of
the female genitalia to the thorax tip, with the head positioned beneath the
thorax at 1890 m (Fig. 1 ![]() ). The head is 411 m wide and 211 m high,
featuring large eyes with a slight eye bridge, measuring 178 m in diameter.
The antennae are delicate and incomplete. The palpi are long, with the right
palpus having two incomplete palpomeres preserved and the left palpus with
four incomplete palpomeres preserved. The thorax is 548 m long. The wing is
hyaline, lacking a stigma (Fig. 2
). The head is 411 m wide and 211 m high,
featuring large eyes with a slight eye bridge, measuring 178 m in diameter.
The antennae are delicate and incomplete. The palpi are long, with the right
palpus having two incomplete palpomeres preserved and the left palpus with
four incomplete palpomeres preserved. The thorax is 548 m long. The wing is
hyaline, lacking a stigma (Fig. 2 ![]() ), measuring 1788 m in length and 712 m
in width. The humeral cross vein (h) reaches the wing margin at 224 m from
the wing base. The subcosta (Sc) is short, measuring 800 m in length, with
the cross-vein sc-r present. R1 ends in the wing margin at about 1368 m from
the wing base, slightly beyond the fork of R2+3 into R2 and R3. R2 and R3
reach the costal margin at 1504 m and 1680 m from the wing base,
respectively. R4 is more than 5 times longer than R4+5, and both R4 and R5 are
straight reaching the wing apex at 1752 m and 1760 m, respectively. M1 is
longer than the discal cell (d) with the d cell closed. M1-M4 reach the wing
margin at 1680 m, 1544 m, 1312 m, and 1104 m apically, respectively.
The cup cell is narrow. CuA and CuP reach the wing posterior margin at 768 m
and 632 m, respectively. The legs are longer than the body and feature two
apical tibial spurs. The abdomen is 1071 m long, with simple female
terminalia (typical of most Psychodidae), and the cerci terminate in a single
lobe measuring 62 m in length.
), measuring 1788 m in length and 712 m
in width. The humeral cross vein (h) reaches the wing margin at 224 m from
the wing base. The subcosta (Sc) is short, measuring 800 m in length, with
the cross-vein sc-r present. R1 ends in the wing margin at about 1368 m from
the wing base, slightly beyond the fork of R2+3 into R2 and R3. R2 and R3
reach the costal margin at 1504 m and 1680 m from the wing base,
respectively. R4 is more than 5 times longer than R4+5, and both R4 and R5 are
straight reaching the wing apex at 1752 m and 1760 m, respectively. M1 is
longer than the discal cell (d) with the d cell closed. M1-M4 reach the wing
margin at 1680 m, 1544 m, 1312 m, and 1104 m apically, respectively.
The cup cell is narrow. CuA and CuP reach the wing posterior margin at 768 m
and 632 m, respectively. The legs are longer than the body and feature two
apical tibial spurs. The abdomen is 1071 m long, with simple female
terminalia (typical of most Psychodidae), and the cerci terminate in a single
lobe measuring 62 m in length.
|
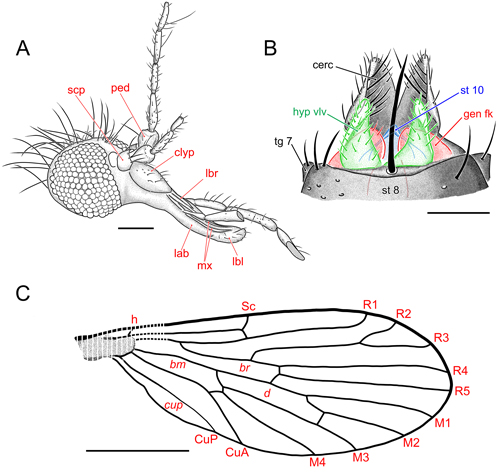
Figure 2:
Nannotanyderus
granieri sp. nov., holotype, female, specimen number BKT-11A, head,
line drawings. A) Head; abbreviations: clyp = clypeus; lab = labium; lbl =
labellum; lbr = labrum; mx = maxilla; ped = pedicel; scp = scape. Scale bar:
100 m. B) Wing, abbreviations: bc = basal costal cell; bm = basal medial
cell; br = basal radial cell; CuA = anterior branch of cubitus vein; cup =
posterior cubital cell; CuP = posterior branch of cubitus vein; d = discal
cell; h = humeral crossvein; M1, M2, M3, M4 = distal branches of medial vein;
R1, R2, R3, R4, R5 = distal branches of radius; Sc = ubcostal vein. Scale bar:
500 m. C) Female terminalia; abbreviations: cerc = cercus; gen fk = genital
fork; hyp vlv = hypogynial valve; st 8 = sternite 8; st 10 = sternite 10; tg 7
= tergite 7. Scale bar: 50 m. |
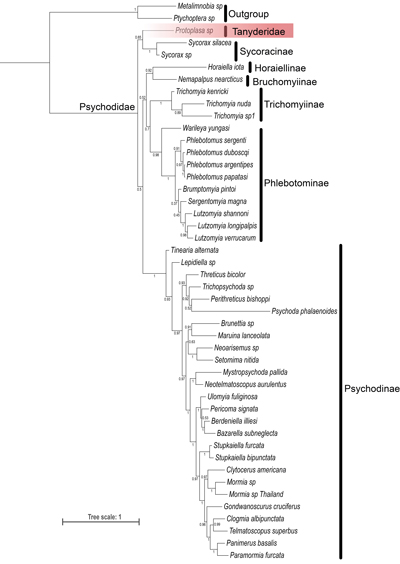
Our phylogenetic analysis of the Tanyderidae and Psychodidae
utilized a site-heterogeneous CAT-GTR model (Fig. 3 ![]() )
in PhyloBayes. This model yielded a moderately resolved topology with strong
statistical support at shallow nodes. Consistent with Curler and Moulton
(2012), the monophyly of the Psychodidae was not upheld in relation to the
Tanyderidae, with the Tanyderidae being identified as a sister group to the
subfamily Sycoracinae. However, contrary to Curler and Moulton
(2012), the Horaiellinae were determined to be a sister group to the subfamily
Bruchomyiinae (Bayesian posterior probability [BPP] = 0.92), rather than to the
remaining subfamilies of the Psychodidae + Tanyderidae. Within the monophyletic
Psychodinae, the genus Tinearia was found to be a sister group to the
remaining sampled genera, with Lepidiella represented a second diverging
lineage within the subfamily (BPP = 0.93).
)
in PhyloBayes. This model yielded a moderately resolved topology with strong
statistical support at shallow nodes. Consistent with Curler and Moulton
(2012), the monophyly of the Psychodidae was not upheld in relation to the
Tanyderidae, with the Tanyderidae being identified as a sister group to the
subfamily Sycoracinae. However, contrary to Curler and Moulton
(2012), the Horaiellinae were determined to be a sister group to the subfamily
Bruchomyiinae (Bayesian posterior probability [BPP] = 0.92), rather than to the
remaining subfamilies of the Psychodidae + Tanyderidae. Within the monophyletic
Psychodinae, the genus Tinearia was found to be a sister group to the
remaining sampled genera, with Lepidiella represented a second diverging
lineage within the subfamily (BPP = 0.93).
|
Figure 3:
Phylogenetic relationships of Tanyderidae (highlighted with red background)
and subfamilies of Psychodidae based on data mainly from Curler and Moulton
(2012) analyzed under a site-heterogeneous CAT-GTR model in PhyloBayes.
Posterior probabilities are reported aside each node. |
Specimens of the Tanyderidae are rare in entomological collections, and this group has been consistently understudied. Placing these dipterans in their evolutionary context presents a formidable challenge, with various evolutionary placements proposed by different authors. These placements have primarily relied on larval features, but especially on adult wing characters, which we consider to be plesiomorphic. Unfortunately, the Tanyderidae remains the only family of extant Diptera for which species-level identification is routinely achievable solely through wing characters (Alexander, 1927). With the exception of one genus (Peringueyomyina Alexander, 1921), all extant tanyderids (in the usual sense) exhibit patterned wings. Existing keys have traditionally been constructed based on the notion that these flies exhibit one of the most 'primitive' conditions of wing venation (MacGillivray, 1923; Rohdendorf, 1974; Krzemiński, 1992). However, there has been no concerted effort to incorporate other structures, such as terminalia, for species-level discrimination. On one hand, phylogenetic hypotheses based on morphological characters of immature stages (Alexander, 1930), adult thoracic sclerites (Crampton, 1926a, 1926b, 1926c, 1930), and wing venation (Rohdendorf, 1974) have yielded unreliable results regarding sister-group relationships, leading to an uncertain phylogenetic placement. For instance, a close relationship between the Tanyderidae and Ptychopteridae (Hennig, 1973; Wood & Borkent, 1989; Oosterbroek & Courtney, 1995), based on mesonotal suture and the foldability of the last tarsomere in males, and more recently with the Blephariceridae (Lambkin et al., 2013), has been suggested.
On the
other hand, recent analyses based on molecular data strongly support a sister
group relationship between the Blephariceridae and Tanyderidae + Psychodidae (Bertone
et al., 2008; Wiegmann et al., 2011; Curler & Moulton,
2012). In all molecular analyses, the families Tanyderidae and Psychodidae are
strongly supported as a monophyletic group. Moreover, a possible character
uniting these three families is the presence of mandibles in the adult Bertone
et al., 2008. Indeed, in the lower Diptera, mandibulate adults only occur in the Culicomorpha,
Blephariceridae, Psychodidae, and Tanyderidae (Downes & Colless,
1967). As Mecoptera (the outgroup of Diptera) have mandibles, this character
is likely a symplesiomorphy. Support for a relationship between the
Tanyderidae and Psychodidae is not surprising. For example, fossil Tanyderidae
and Psychodidae are remarkably similar (Ansorge, 1994; Grimaldi & Engel,
2005; Krzemiński et al., 2013), to the extent that
confusion of fossil taxa has occurred (Woodley, 2005). This confusion
is understandable, as some tanyderid (in the usual sense) fossils exhibit more 'plesiomorphic'
characters akin to
the Psychodidae rather than to the Tanyderidae, where features such as size,
wing venation, head shape, absence of neck stretching, and inversion of male
genitalia observed in some fossils are characteristic of the Psychodidae. Some
other fossil psychodids described from Lebanese amber with m-m crossvein could
also be considered by some workers as tanyderids (in the usual sense). Some of
the earliest described tanyderids were incorrectly placed in the
Ptychopteridae (Loew, 1850; Alexander,
1913). In 1879, Osten-Sacken
proposed that Tanyderina included three genera: Macrochile Loew,
1850, Protoplasa Osten-Sacken,
1859, and Tanyderus Philippi,
1865. Alexander (1920) recognized Tanyderina as a family and divided it
into two subfamilies, Tanyderinae and Bruchomyiinae. Later, he acknowledged that the
Bruchomyiinae belong to the family Psychodidae (Alexander,
1965).
Recently, Skibińska (2016) correctly subdivided the Tanyderidae
into two subfamilies, the Tanyderinae and Nannotanyderinae. This last
subfamily comprises tanyderids with unique male genitalia featuring elongate
gonopods without patterned wings. Nannotanyderus granieri sp.
nov. could be assigned to the Tanyderidae in this sense, based on wing venation, arrangement of setae on flagellomeres,
absence of ascoids, presence of cross-vein m-m, presence of m-cu cross-vein,
presence of discal cell (d), and spurs on the tibiae. It is noteworthy to
state that N. granieri sp. nov. can be easily confused with
members of the family Psychodidae, with whom it shares some features, such as,
small size, absence of well-developed anal lobe, inverted genitalia,
immaculate wings, and a general aspect of venation. Within the Tanyderidae in
the usual sense, Nannotanyderus granieri sp. nov. can be
attributed to the Nannotanyderinae in the usual sense, due to the striking
similarity in wing venation with members of the genus Nannotanyderus in
general, and particularly with N. ansorgei Krzemiński
et al., 2013. N. granieri sp. nov. shares with all
members of Nannotanyderus its small size, absence of well-developed anal lobe, and
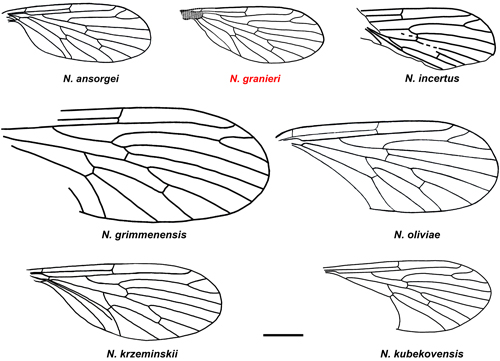
Sc reaching the wing margin at about the half of the wing length (Fig. 4 ![]() ). N.
granieri sp. nov. differs from all known Nannotanyderus species
(except N. ansorgei and N. incertus) by its considerably smaller
wings. N. granieri sp. nov. differs from N. incertus by
the much more proximal position of CuA in the latter. The wing venations of N.
granieri are very similar to those of N. ansorgei, with the
following differences: 1) R1 reaching the costal margin slightly beyond the
fork of R2+3 into R2 and R3 in N. granieri, versus opposite to
it in N. ansorgei; 2) R2 slightly longer than half of R3 in N.
granieri, versus equal to half of R3 in N. ansorgei; 3) R2+3
bifurcation from Rs slightly distal to level where Sc meets costal margin in N.
granieri, versus slightly basal to the level where Sc meets the
costal margin in N. ansorgei; 4) the distance between the bifurcation
of R2+3 from Rs to the base of R4 is smaller in N. granieri. N.
granieri and N. ansorgei are to date the smallest known for
tanyderid flies.
). N.
granieri sp. nov. differs from all known Nannotanyderus species
(except N. ansorgei and N. incertus) by its considerably smaller
wings. N. granieri sp. nov. differs from N. incertus by
the much more proximal position of CuA in the latter. The wing venations of N.
granieri are very similar to those of N. ansorgei, with the
following differences: 1) R1 reaching the costal margin slightly beyond the
fork of R2+3 into R2 and R3 in N. granieri, versus opposite to
it in N. ansorgei; 2) R2 slightly longer than half of R3 in N.
granieri, versus equal to half of R3 in N. ansorgei; 3) R2+3
bifurcation from Rs slightly distal to level where Sc meets costal margin in N.
granieri, versus slightly basal to the level where Sc meets the
costal margin in N. ansorgei; 4) the distance between the bifurcation
of R2+3 from Rs to the base of R4 is smaller in N. granieri. N.
granieri and N. ansorgei are to date the smallest known for
tanyderid flies.
|
Figure 4:
Line
drawings of wings of all known species of Nannotanyderus (Scale bar =
500 m). |
The discovery of very small Tanyderidae, which share several structural characters with Psychodidae such as the absence of neck, wing venation, and inversion of male genitalia, alongside primitive Psychodidae from the Jurassic and Cretaceous periods, has enhanced our understanding of the close phylogenetic relationship between the Tanyderidae and Psychodidae, as previously stated by Krzemiński (1992), Krzemiński and Evenhuis (2000), and Krzemiński and Krzemińska (2003). It is worth recalling that the Bruchomyiinae, now considered as a psychodid subfamily, was initially classified within the Tanyderidae by Alexander (1920). Furthermore, in all molecular phylogenies conducted since 2008 (Bertone et al., 2008; Wiegmann et al., 2011; Curler & Moulton, 2012), Tanyderids consistently appear as a sister-group to the Psychodidae, forming a monophyletic clade. Considering all these findings, and given the possibility of confusion in the classification of present-day tanyderids and psychodids with the discovery of fossil material, we find it legitimate to downgrade the Tanyderidae to the rank of a subfamily within the Psychodidae. In this case, the two existing subfamilies of the Tanyderidae would be considered as the tribes Tanyderini Osten-Sacken, 1880, and Nannotanyderini Skibińska, 2016, instead of the subfamilies Tanyderinae Osten-Sacken, 1880, and Nannotanyderinae Skibińska, 2016, respectively.
The discovery of new fossils has caused confusion in the classification of modern tanyderids and psychodids. Moreover, all recent molecular phylogenies consistently show a sister-group relationship between the Tanyderidae and Psychodidae within a monophyletic clade. These findings prompt us to consider the Tanyderidae and Psychodidae as a single family, with the Tanyderidae being regarded as a subfamily (Tanyderinae) within the Psychodidae.
The ongoing discovery of new fossils and recent material continues to provide valuable insights into the true evolutionary history of tanyderids and psychodids.
The authors would like to express their gratitude two anonymous reviewers for their valuable comments on a previous version of this paper. This study is a contribution to the activity of the laboratory "Advanced Micropalaeontology, Biodiversity and Evolution Researches" (AMBER) led by Dany Azar at the Lebanese University. Financial support for this work was provided by the National Natural Science Foundation of China (42288201) and the Second Tibetan Plateau Scientific Expedition and Research project (2019QZKK0706).
Alexander C.P. (1920).- A new genus and species net winged midge (Blephariceridae) and an undescribed species of Tanyderidae (Diptera).- Arkiv for Zoologi, Stockholm, vol. 13, p. 1-7.
Alexander C.P. (1921).- A new genus and species of Tanyeridae (Peringueyomyina barnardi) in the South African museum (Diptera).- Annals of the South African Museum, Cape Town, vol. 18, p. 231-234.
Alexander C.P. (1927).- Diptera. Family Tanyderidae.- Genera Insectorum, Bruxelles, vol. 189, p. 1-13.
Alexander C.P. (1928).- The Tanyderidae of Australia (Diptera).- Proceedings of the Linnean Society of New South Wales, Sydney, vol. 4, p. 367-374.
Alexander C.P. (1929).- Diptera of Patagonia and South Chile. Part I. Crane-flies (Tipulidae, Trichoceridae, Tanyderidae).- The British Museum (Natural History), London, Fasc. 1, 240 p.
Alexander C.P. (1930).- Observations on the dipterous family Tanyderidae.- Proceedings of the Linnean Society of New South Wales, Sydney, vol. 55, p. 221-230.
Alexander C.P. (1932).- The Dipterous family Tanyderidae in Japan (Insecta).- Annotationes Zoologicae Japonenses, Tokyo, vol. 13, p. 273-281.
Alexander C.P. (1965).- Superfamily Psychodoidea Family Tanyderidae. In: Stone A., Sabrosky C.W., Wirth W.W., Foote R.H. & Coulson J.R. (eds.), A Catalog of the Diptera of America North of Mexico.- U.S. Government Printing Office, Washington - DC, p. 90.
Ansorge J. (1994).- Tanyderidae and Psychodidae (Insecta: Diptera) from the Lower Jurassic of northeastern Germany.- Pal�ontologische Zeitschrift, vol. 68, p. 199-210.
Ansorge J. & Krzeminski W. (2002).- Lower Jurassic tanyderids (Diptera: Tanyderidae) from Germany.- Studia Dipterologica, Halle an der Saale, vol. 9, p. 21-29.
Azar D., Perrichot V., N�raudeau D. & Nel A. (2003).- New psychodid flies from the Cretaceous ambers of Lebanon and France, with a discussion about Eophlebotomus connectens Cockerell, 1920 (Diptera, Psychodidae).- Annals of the Entomological Society of America, vol. 96, p. 117-127.
Azar D. & Maksoud S. (2022a).- Paleopsychoda jarzembowskii, a new Lower Cretaceous species of moth flies from Lebanese amber (Diptera: Psychodidae: Psychodinae).- Palaeoentomology, Auckland, vol. 5, p. 262-268.
Azar D. & Maksoud S. (2022b).- Libanophlebotomites ramyi, a new genus and species of phlebotomine sandflies from the Lower Cretaceous Lebanese amber (Diptera: Psychodidae: Phlebotominae).- Palaeoentomology, Auckland, vol. 5, p. 340-346.
Bertone M.A., Courtney G.W. & Wiegmann B.M. (2008).- Phylogenetics and temporal diversification of the earliest true flies (Insects: Diptera) based on multiple nuclear genes.- Systematic Entomology, vol. 33, p. 668-687.
Bode A. (1953).- Die Insektenfauna des ostniedersachsischen oberen Lias.- Palaeontographica A, Stuttgart, Band 103, p. 1-375.
Cai C.-Y. & Huang D.-Y. (2014).- The oldest micropepline beetle from Cretaceous Burmese amber and its phylogenetic implications (Coleoptera: Staphylinidae).- Naturwissenschaften, vol. 101, p. 813-817.
Colless D.H. & McAlpine D.K. (1991).- Diptera (flies). In: The insects of Australia, vol. 2.- Cornell University Press, p. 717-786.
Crampton G.C. (1926a).- A phylogenetic study of the thoracic sclerites of the pychodoid Diptera, with remarks on the interrelationships of the Nematocera.- Entomological News, vol. XXXVII, no. 2, p. 33-39. URL: https://www.biodiversitylibrary.org/partpdf/30135
Crampton G.C. (1926b).- A phylogenetic study of the thoracic sclerites of the pychodoid Diptera, with remarks on the interrelationships of the Nematocera.- Entomological News, vol. XXXVII, no. 3, p. 65-70. URL: https://www.biodiversitylibrary.org/partpdf/30139
Crampton G.C. (1926c).- The external anatomy of the primitive tanyderid dipteran Macrochile spectrum Loew, preserved in Baltic Amber.- Brooklyn Entomological Society, New York - NY, vol. XXI, nos. 1-2, p. 1-14. URL: https://archive.org/details/biostor-169514
Crampton G.C. (1930).- A comparison of the more important structural details of the larvae of the archaic tanyderid dipteron Protoplasa fitchii with other Holometabola from the standpoint of phylogeny.- Brooklyn Entomological Society, vol. XXV, no. 5, p. 239-258. URL: https://www.biodiversitylibrary.org/part/177843
Curler G.R. & Moulton J.K. (2012).- Phylogeny of psychodid subfamilies (Diptera: Psychodidae) inferred from nuclear DNA sequences with a review of morphological evidence for relationships.- Systematic Entomology, vol. 37, p. 603-616.
Dong F., Shih C.-K. & Ren D. (2015).- A new genus of Tanyderidae (Insecta: Diptera) from Myanmar amber, Upper Cretaceous.- Cretaceous Research, vol. 54, p. 260-265.
Dong F., Shih C.-K., Skibińska K., Krzemiński W. & Ren D. (2015).- New species of Tanyderidae (Diptera) from the Jiulongshan Formation of China.- Alcheringa: An Australian Journal of Palaeontology, vol. 39, p. 494-507.
Downes J.A. & Colless D.H. (1967).- Mouthparts of the biting and blood-sucking type in Tanyderidae and Chironomidae (Diptera).- Nature, vol. 214, p. 1355-1356.
Enderlein G. (1936).- Klassifikation der Psychodiden (Dipt.).- Deutsche Entomologische Zeitschrift, Berlin, Jahrgang 1936, Heft III/IV, p. 81-112. DOI: 10.1002/mmnd.193619360301
Exner K. & Craig D.A. (1976).- Larvae of Alberta Tanyderidae (Diptera: Nematocera).- Quaestiones Entomologicae, Edmonton, vol. 12, p. 219-237.
Fu Y.-Z., Li Y.-D., Su Y.-T., Cai C.-Y. & Huang D.-Y. (2021).- Application of confocal laser scanning microscopy to the study of amber bioinclusions.- Palaeoentomology, vol. 4, p. 266-278.
Granier B., Azar D., Maksoud S., G�ze R. & Habchi R. (2015).- New fossiliferous sites with Barremian Charophyta in the "Gr�s du Liban" auct. (Lebanon), with a critical perspective regarding the nature of Munieria Deecke, 1883.- Carnets Geol., Madrid, vol. 15, no. 15, p. 199-229. DOI: 10.4267/2042/57947
Granier B., Toland C., G�ze R., Azar D. & Maksoud S. (2016).- Some steps toward a new story for the Jurassic - Cretaceous transition in Mount Lebanon.- Carnets Geol., Madrid, vol. 16, no. 8, p. 247-269. DOI: 10.4267/2042/59924
Grimaldi D.A. & Engel M.S. (2005).- The evolution of insects.- Cambridge University Press, Cambridge, 772 p.
Handlirsch A. (1909).- Zur Phylogenie und Fl�gelmorphologie der Ptychopteriden (Dipteren).- Annalen der Naturhistorischen Hofmuseums, Wien, Band 23, H�ft 3/4, p. 263-272.
Hennig W. (1968).- Kritische Bemerkungen �ber den Bau der Fl�gelwurzel bei den Dipteren und die Frage nach der Monophylie der Nematocera.- Stuttgarter Beitr�ge zur Naturkunde, no. 193, p. 1-23. URL: https://archive.org/details/biostor-233004
Hennig W. (1973).- Ordnung Diptera (Zweifl�gler). In: Helmcke J.-G., Starch, D. & Wermuth H. (eds.), Handbuch der Zoologie. IV. Band: Arthropoda - 2. H�lfte: Insecta (Zweite Auflage). 2. Teil: Spezielles.- W. De Gruyter, Berlin, 337 p.
Judd D.D. (2004).- Insecta: Diptera, Tanyderidae. In: Yule C.M & Yong H.S (eds.), Freshwater Invertebrates of the Malaysian Region.- Academy of Sciences Malaysia, Kuala Lumpur, p. 626-633.
Kalugina N.S. (1985).- Infraorders Psychodomorpha, Tipulomorpha and Culicomorpha. In: Kalugina N.S. & Kovalev V.G. (eds.), Dvukrylye nasekomye Yury Sibiri [Jurassic Diptera of Siberia]. Akademia Nauk, Moscow, p. 33-113 [in Russian].
Kalugina N.S. (1988).- Mesozoic Diptera Psychodomorpha and Tipulomorpha (Tanyderidae, Eoptychopteridae, Limoniidae: Diptera). In: New species of fossil invertebrates of Mongolia.- Trudy Sovmestnoy Sovetsko-Mongol'skoy Paleontologicheskoy Ehkspeditsii, vol. 33, p. 81-89 [in Russian].
Kalugina N.S. (1992).- Psychodomorphian Diptera from the Jurassic of Mongolian Altai (Diptera: Tanyderidae, Eoptychopteridae).- Paleontological Journal, vol. 26, p. 110-113.
Kalugina N.S. & Kovalev V.G. (1985).- Dvukryliena sekomye Yury Sibirii [Jurassic Diptera of Siberia].- Moscow, 198 p. [in Russian]
Katoh K., Misawa K., Kuma K.I. & Miyata T. (2002).- MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform.- Nucleic acids research, vol. 30, p. 3059-3066.
Katoh K., Rozewicki J. & Yamada K.D. (2019).- MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization.- Briefings in bioinformatics, vol. 20, p. 1160-1166.
Krzemiński W. (1992).- Triassic and Lower Jurassic stage of Diptera evolution.- Mitteilungen der Schweizerischen Entomologischen Gesellschaft, vol. 65, p. 39-59.
Krzemiński W., Azar D. & Skibińska K. (2013).- Nannotanyderus ansorgei sp. n., the first member of the family Tanyderidae from Lebanese amber (Lower Cretaceous). In: Azar D., Engel M.S., Jarzembowski E., Krogmann L., Nel A. & Santiago-Blay J. (eds.), Insect evolution in an amberiferous and stone alphabet, Proceedings of the 6th International Congress on Fossil Insects, Arthropods and Amber.- Brill, Leiden, p. 131-143.
Krzemiński W. & Evenhuis N.L. (2000).- Review of Diptera palaeontological records. In: Papp L. & Darvas B. (eds.), Manual of Palaearctic Diptera, vol. 1. General and Applied Dipterology.- Science Herald, Budapest, p. 535-564.
Krzemiński W. & Judd D.D. (1997).- Family Tanyderidae. In: Papp L. & Darvas B. (eds.), Contributions to a manual of Palaearctic Diptera, vol. 2.- Science Herald, Budapest, p. 281-289.
Krzemiński W. & Krzemińska E. (2003).- Triassic Diptera: descriptions, revisions and phylogenetic relations.- Acta Zoologica Cracoviensia, vol. 46 (Suppl. Fossil Insects), p. 153-184.
Krzemiński W., Krzemińska E., Kania I. & Ross A.J. (2013).- New taxa of Tanyderidae (Diptera) from Eocene Baltic amber.- Zootaxa, Auckland, vol. 3599, p. 59-66.
Krzemiński W. & Ren D. (2001).- Praemacrochile chinensis sp. n. from the Middle Jurassic of China (Diptera: Tanyderidae).- Polskie pismo entomologiczne, Lublin, vol. 70, p. 127-129.
Kvifte G.M. & Wagner R. (2017).- Psychodidae (sand flies, moth flies or owl flies). In: Kirk-Spriggs A.H. & Sinclair B.J. (eds.), Manual of Afrotropical Diptera, vol. 2. Nematocerous Diptera and lower Brachycera. Suricata 5.- South African National Biodiversity Institute, Pretoria, p. 607-632.
Lambkin C., Sinclair B.J., Pape T., Courtney G.W., Skevington J.H., Meier R., Yeates D., Blagoderov V. & Wiegmann B.M. (2013).- The phylogenetic relationships among infraorders and superfamilies of Diptera based on morphological evidence.- Systematic Entomology, vol. 38 p. 164-179.
Lartillot N., Lepage T. & Blanquart S. (2009).- PhyloBayes 3: A Bayesian software package for phylogenetic reconstruction and molecular dating.- Bioinformatics, vol. 25, p. 2286-2288.
Lartillot N., Brinkmann H. & Philippe H. (2007).- Suppression of long-branch attraction artefacts in the animal phylogeny using a site-heterogeneous model.- BMC Evolutionary Biology, vol. 7, suppl. 1, article S4, 14 p. URL: https://bmcecolevol.biomedcentral.com/articles/10.1186/1471-2148-7-S1-S4
Linnaeus C. (1758).- Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, (ed.), 10.- Salviae, Holmiae (Stockholm) [in Latin].
Loew H. (1850).- Dipterologische Beitr�ge. Vierter Beitrag. Die Gallm�cken. (Progr).- Kiessling F.W.G., Posen, p. 1-40.
Lukashevich E.D. (2011).- New Nematocerans (Insecta: Diptera) from the Late Jurassic of Mongolia.- Paleontological Journal, vol. 45, p. 620-628.
Lukashevich E.D. & Krzemiński W. (2009).- New Jurassic Tanyderidae (Diptera) from Asia with first find of larvae.- Zoosymposia, Auckland, vol. 3, p. 155-172.
MacGillivray A.D. (1923).- External insect-anatomy: A guide to the study of insect anatomy and an introduction to systematic entomology.- Flanigan-Pearson Company, Champaign IL, p. 319-333.
Madriz R.I., Astorga A., Lindsay T. & Courtney G.W. (2018).- A new species of Neoderus Alexander, 1927 (Diptera: Tanyderidae) from southern Chile, with a first description of a male and key to extant genera of the family.- Aquatic Insects, vol. 39, p. 155-179.
Madriz Villanueva R.I. (2017).- The primitive crane flies (Diptera: Tanyderidae).- Graduate Theses and Dissertations, Iowa State University, Ames IA, ix + 454 p. URL: https://dr.lib.iastate.edu/handle/20.500.12876/30356
Maksoud S., Maalouf M., Maalouf R. & Azar D. (2021).- Baskinta and Bqaatouta: Two new early Barremian fossiliferous amber outcrops from Central Lebanon.- Palaeoentomology, vol. 4, p. 367-373.
Men Q.-L., Hu Z.-K. & Xu L.-Y. (2020).- The first representative of Espanoderus and one new species of Similinannotanyderus (Diptera: Tanyderidae) from mid-Cretaceous amber of northern Myanmar.- Cretaceous Research, vol. 111, article 104442, 7 p.
Newman E. (1834).- Attempted division of British insects into natural orders.- Entomological Magazine, vol. 2, p. 379-431.
Oosterbroek P. & Courtney G.W. (1995).- Phylogeny of the nematocerous families of Diptera (Insecta).- Zoological Journal of the Linnean Society, vol. 115, p. 267-311.
Osten-Sacken C.R. (1859).- New genera and species of North America Tipulidae with short palpi, with an attempt at a new classification of the tribe.- Proceedings of the Academy of Natural Sciences of Philadelphia, vol. 1859, p. 197-254.
Osten-Sacken C.R. (1880).- Die Tanyderina, eine merkw�rdige gruppe der Tipuliden.- Verhandlungen der Kaiserlich-K�eniglichen zoologisch-botanischen Gesellschaft in Wien, Band XXIX , p. 517-522.
Philippi R.A. (1865).- Aufz�hlung der chilenischen Dipteren.- Verhandlungen der kaiserlich-koeniglichen zoologisch-botanischen Gesellschaft in Wien, Band XV, p. 595-782.
Podenas S. (1997).- New Macrochile Loew, 1850 (Diptera, Tanyderidae) from the Baltic amber.- Mitteilungen des Geologisch-Pal�ontologischen Instituts der Universit�t Hamburg, vol. 80, p. 173-177.
Poinar G.O. Jr & Brown A.E. (2004).- A new genus of primitive crane flies (Diptera: Tanyderidae) in Cretaceous Burmese amber, with a summary of fossil tanyderids.- Proceedings of the Entomological Society of Washington, vol. 106, p. 339-345.
Ranwez V., Douzery E.J., Cambon C., Chantret N. & Delsuc F. (2018).- MACSE v2: Toolkit for the alignment of coding sequences accounting for frameshifts and stop codons.- Molecular Biology and Evolution, vol. 35, p. 2582-2584.
Rohdendorf B. (1974).- The historical development of Diptera.- The University of Alberta Press, Edmonton, 360 p.
Rondani C. (1840).- Sopra una specie di insetto dittero. In: Memoria prima per servire alla ditterologia italiana.- Stamperia Donati, Parma, 16 p. (1 Pl.)
Rondani C. (1856).- Dipterologiae Italicae Prodromus. Vol. 1. Genera italica ordinis dipterorum ordinatim disposita et distincta et in familias et stirpes aggregata.- Tipographia Alexandri Stoschi, Parmae, 226 p. [in Latin]. DOI: 10.5962/bhl.title.8160
Skibińska K. (2016).- Nannotanyderinae: A new subfamily of Tanyderidae (Diptera).- Palaeontologia Electronica, article 19.3.56A, p. 1-16. DOI: 10.26879/551
Skibińska K. & Krzemiński W. (2013).- Nannotanyderus kubekovensis sp. nov. (Diptera: Tanyderidae) from the Middle Jurassic of Kubekovo (Russia).- Annales Zoologici, Warszawa, vol. 63, no. 3, p. 409-412.
Skibińska K. & Krzemiński W. (2018).- Two new species of the genus Dacochile (Diptera, Tanyderidae) from Burmese amber.- Historical Biology, vol. 32, p. 356-361.
Skibińska K., Krzemiński W. & Arillo A. (2019).- The first Tanyderidae (Diptera) from Lower Cretaceous lava amber (Spain).- Historical Biology, vol. 31, p. 872-878.
Skibińska K., Krzemiński W. & Coram R. (2014).- Discovery of the most ancient member of the family Tanyderidae (Diptera: Nematocera) from the Lower Jurassic (Sinemurian) of England.- Zootaxa, Auckland, vol. 3857, no. 1, p. 125-130.
Stebner F., Sol rzano Kraemer M.M., Ib ez-Bernal S. & Wagner R. (2015a).- Moth flies and sand flies (Diptera: Psychodidae) in Cretaceous Burmese amber.- PeerJ, vol. 3, no. 1254, 34 p. DOI: 10.7717/peerj.1254
Stebner F., Sol rzano Kraemer M.M., Ib ez-Bernal S. & Wagner R. (2015b).- Datziinae as a new subfamily name for the unavailable name Protopsychodinae (Diptera: Psychodidae).- PeerJ, vol. 3, no. e1423, 3 p. DOI: 10.7717/peerj.1423
Tihelka E., Thayer M.K., Newton A.F. & Cai C.-Y. (2020).- New data, old story: molecular data illuminate the tribal relationships among rove beetles of the subfamily Staphylininae (Coleoptera: Staphylinidae).- Insects, vol. 11, no. 3, article 164, 12 p.
Tihelka E., Cai C., Giacomelli M., Lozano-Fernandez J., Rota-Stabelli O., Huang D., Engel M.S., Donoghue P.C.J. & Pisani D. (2021).- The evolution of insect biodiversity.- Current Biology, vol. 31, p. R1299-R1311.
Tonnoir A.L. (1922).- Note sur le genre Nemopalpus (Psychodidae) et description d'une esp�ce nouvelle.- Annales de la Soci�t� Entomologique de Belgique, Bruxelles, vol. 62, p. 125-136.
Wiegmann B.M., Trautwein M.D., Winkler I.S., Barr N.B., Kim J., Lambkin C., Bertone M.A., Cassel B.K., Bayless K.M., Heimberg A.M., Wheeler B.M., Peterson K.J., Pape T., Sinclair B.J., Skevington J.S., Blagoderov V., Caravas J., Kutty S.N., Schmidt-Ott U., Kampmeier G.E., Thompson F.C., Grimaldi D.A., Beckenbach A.T., Courtney G.W., Friedrich M., Meier R. & Yeates D.K. (2011).- Episodic radiations in the fly tree of life.- Proceedings of the National Academy of Sciences United States, Washington DC, vol. 108, p. 5690-5695.
Williams I.W. (1933).- The external morphology of the primitive tanyderid dipteron Protoplasa fitchii O.S., with notes on the other Tanyderidae.- Journal of the New York Entomological Society, vol. 41, p. 1-35.
Wood D.M. & Borkent A. (1989).- 114. Phylogeny and classification of the Nematocera. In: McAlpine J.F. (ed.), Manual of Nearctic Diptera. Volume 3.- Research Branch, Agriculture Canada, Monograph, Ottawa, no. 32, p. 1333-1370.
Woodley N.E. (2005).- Dacochile microsoma Poinar & Brown, not a tanyderid but a bruchomyiine psychodid (Diptera: Psychodidae, Tanyderidae).- Zootaxa, Auckland, vol. 1012, p. 53-60.
Zhang J.-F. (2004).- Nematoceran Dipterans from the Jurassic of China (Insecta, Diptera: Limoniidae, Tanyderidae).- Paleontologicheskii Zhurnal, vol. 5, p. 53-57 [in Russian, translated in English, Paleontological Journal, vol. 38, p. 522-527].
List of all 34 described fossil Tanyderinae stat. nov.
| Genus | Species | Author(s) | Age | Origin |
| Coramus | gedanensis | Skibińska, 2016 | Upper Eocene | Baltic amber |
| Dacochile | browni | Skibińska & Krzemiński, 2018 | mid-Cretaceous | Burmese amber |
| D. | microsoma | Poinar & Brown, 2004 | mid-Cretaceous | Burmese amber |
| D. | poinari | Skibińska & Krzemiński, 2018 | mid-Cretaceous | Burmese amber |
| Espanoderus | barbarae | Skibińska et al., 2019 | Lower Cretaceous | lava, Spanish amber |
| Esp. | orientalis | Men & Hu in Men et al., 2020 | mid-Cretaceous | Burmese amber |
| Macrochile | hornei | Krzemiński et al., 2013 | Upper Eocene | Baltic amber |
| M. | spectrum | Loew, 1850 | Upper Eocene | Baltic amber |
| Nannotanyderus | ansorgei | Krzemiński et al., 2013 | Lower Cretaceous | Bkassine, Lebanese amber |
| N. | granieri | This work | Lower Cretaceous | Bqaatouta, Lebanese amber |
| N. | grimmensis | Ansorge & Krzemiński, 2002 | Lower Jurassic | Western Pomerania, Germany |
| N. | incertus | Lukashevich, 2011 | Upper Jurassic | Share Teg, Mongolia; Durlstone Bay, England |
| N. | krzeminskii | Ansorge, 1994 | Lower Jurassic | Germmen, Germany; Dobbertin, Germany |
| N. | kubekorensis | Skibińska & Krzemiński, 2013 | Middle Jurassic | Kubekovo, Russia |
| N. | oliviae | Skibińska et al., 2014 | Lower Jurassic | Dorset, England |
| Podemacrochile | baltica | (Podenas, 1997) | Upper Eocene | Baltic amber |
| Praemacrochile | ansorgei | Lukashevich & Krzemiński, 2009 | Middle or Upper Jurassic | Karatau, Kazakhstan; Shar-Teg, Mongolia |
| Praem. | chinensis | Krzemiński & Ren, 2001 | Middle Jurassic | Daohugou, China |
| Praem. | decipiens | (Bode, 1953) | Lower Jurassic | Dobbertin, Germany |
| Praem. | dobbertinensis | Ansorge & Krzemiński, 2002 | Lower Jurassic | Dobbertin, Germany |
| Praem. | dryasis | Dong et al., 2015 | Middle Jurassic | Daohugou, China |
| Praem. | kaluginae | Lukashevich & Krzemiński, 2009 | Middle or Upper Jurassic | Karatau, Kazakhstan |
| Praem. | stackelbergi | Kalugina, 1985 | Lower or Middle Jurassic | Siberia, Russia |
| Protanyderus | astictum | Dong et al., 2015 | Middle Jurassic | Daohugou, China |
| Prot. | invalidus | Lukashevich & Krzemiński, 2009 | Middle Jurassic | Siberia and Kubekovo, |
| Prot. | mesozoicus | Kalugina, 1988 | Upper Jurassic or Upper Cretaceous | Khutel Khara, Mongolia |
| Prot. | nebulosus | Lukashevich & Krzemiński, 2009 | Upper Jurassic | Shar-Teg, Mongolia |
| Prot. | savtchenkoi | Lukashevich & Krzemiński, 2009 | Middle or Upper Jurassic | Karatau, Kazakhstan |
| Prot. | senilis | Kalugina, 1992 | Upper Jurassic | Shar-Teg, Mongolia |
| Prot. | vetus | Kalugina, 1992 | Upper Jurassic | Shar-Teg, Mongolia |
| Prot. | vulcanium | Zhang, 2004 | Middle Jurassic | Daohugou, China |
| Similinannotanyderus | lii | Dong et al., 2015 | mid-Cretaceous | Burmese amber |
| S. | longitergata | Men & Hu in Men et al., 2020 | mid-Cretaceous | Burmese amber |
| S. | zbigniewi | Skibińska & Krzemiński, 2018 | mid-Cretaceous | Burmese amber |
| Nomenclatural note: Life Sciences Identifier (LSID) https://zoobank.org/References/86D9B493-1246-4427-915E-2AF0EC0286BC Species Group |