◄ Carnets Geol. 24 (10) ►
![]()
Outline:
[1. Introduction]
[2. Description of zones]
[3. Future work] and ...
[Bibliographic references]
Department of Geography and Geology, The University of the
West Indies, Mona, Kingston 7 (Jamaica)
Department of Geography and Geology, The University of the
West Indies, Mona, Kingston 7 (Jamaica)
Deceased
Geological Services, 1414 Lumpkin Road Houston, Texas 77043 (U.S.A.)
Department of Geography and Geology, The University of the
West Indies, Mona, Kingston 7 (Jamaica)
Published online in final form (pdf) on October 31, 2024
DOI 10.2110/carnets.2024.2410
![]()
[Editor:
Daniela Basso; technical editor: Bruno R.C. Granier]
![]()
The Cenozoic of the Americas does not have a zonation based on Larger Benthic Foraminifera (LBF) unlike the Tethys where there is a shallow-water benthic zonation. Although there were faunal exchanges, the different evolutionary patterns between the Americas and the Tethys require separate zonations. We have calibrated LBF, stratigraphic distributions and events in the Americas using calcareous nannoflora (Paleocene-Eocene), planktic foraminifers (Oligocene-Miocene) and published strontium (Sr) isotope values. We define 29 zones spanning the Paleocene to Recent based on changing LBF assemblages and evolving lineages, with some zonal boundaries related to global or local palaeoenvironmental events. This will allow correlation across carbonate platforms in the tropical Americas at a higher resolution than previously possible.
Cenozoic;
Larger Benthic Foraminifera;
biostratigraphy;
zonation;
Americas
Mitchell S.F., Robinson E., Özcan E., Jiang M.M. & Robinson N. (2024).- A larger benthic foraminiferal zonation for the Cenozoic of the Americas.- Carnets Geol., Madrid, vol. 24, no. 10, p. 163-172. DOI: 10.2110/carnets.2024.2410
Zonation du Cénozoïque des Amériques à l'aide des foraminifères benthiques de grande taille.- Contrairement au domaine téthysien où, pour la zone néritique au cours du Cénozoïque, les biostratigraphes disposent déjà d'une zonation basée sur les foraminifères benthiques de grande taille (FBGT), il n'en existait pas encore pour les Amériques. Bien qu'il y ait eu des échanges entre les Amériques et la Téthys, les différents schémas évolutifs nécessitent des zonations distinctes. Nous avons calibré les distributions stratigraphiques des FBGT avec des événements dans les Amériques en utilisant le nannoplancton calcaire (Paléocène-Éocène) et les foraminifères planctoniques (Oligocène-Miocène), ainsi que les valeurs publiées des isotopes du strontium (Sr). Nous définissons ainsi 29 zones s'étendant du Paléocène à l'époque actuelle, basées sur les lignées évolutives et les assemblages changeants des FBGT, avec quelques limites de biozones liées à des événements paléoenvironnementaux globaux ou locaux. Ceci devrait permettre des corrélations de meilleure résolution qu'auparavant entre les plates-formes carbonatées des Amériques tropicales.
Cénozoïque ;
foraminifères benthiques de grande taille ;
biostratigraphie ;
zonation ;
Amériques
The
Paleogene to early Neogene of Tethys and the western Pacific can be zoned in
detail using assemblages of larger benthic foraminifers (LBF) (Adams,
1970; Cahuzac & Poignant,
1997; Serra-Kiel et
al., 1998; Less & Özcan,
2012; Papazzoni et
al., 2017, 2023). Although some zonations (often for part of the Paleogene)
have been proposed for the Americas (e.g.,
Robinson, 1977, 2004; Butterlin,
1981; Robinson & Jiang,
1995; Robinson & Mitchell,
1999; Mitchell
et al., 2022; Sierra et
al., 2023; Mello e Sousa et
al., 2023), there is no high-resolution scheme for the Cenozoic. LBF show
significant palaeogeographical differences between the Americas and the Tethys,
and the scheme for the Tethys cannot be used in the Americas. To address this,
we have collected free specimens of LBF and investigated them using orientated
equatorial and axial sections supplemented by data from the literature (CT scans
are also extremely valuable: cf. Sierra et al., 2023). This allows the identification of critical features
that allows us to develop a high-resolution zonation (Fig.
1 ![]() ) that cannot be
achieved using random sections in rock. The zonation is calibrated against the
chronostratigraphy using the NP zonation based on calcareous nannofossils (Martini,
1971; Vandenberghe et al.,
2012; Speijer et al.,
2020) for
the Paleocene-Eocene and the P and N zonation based on planktic foraminifers for
the Oligocene and Miocene (King et al., 2020). We have revised the classification of many of the
forms from the Eocene of Jamaica (Mitchell et al.,
2022) and use this revised taxonomy and other works to erect
a series of zones (based on first and last appearances of taxa) for the
Paleocene to the Recent.
) that cannot be
achieved using random sections in rock. The zonation is calibrated against the
chronostratigraphy using the NP zonation based on calcareous nannofossils (Martini,
1971; Vandenberghe et al.,
2012; Speijer et al.,
2020) for
the Paleocene-Eocene and the P and N zonation based on planktic foraminifers for
the Oligocene and Miocene (King et al., 2020). We have revised the classification of many of the
forms from the Eocene of Jamaica (Mitchell et al.,
2022) and use this revised taxonomy and other works to erect
a series of zones (based on first and last appearances of taxa) for the
Paleocene to the Recent.
American shelf-edge taxa have been chosen for the zonation because they are widely distributed, show less biogeographical restrictions in their distribution than platform interior taxa, and have high rates of evolution. The principal groups include Tremastegina-Helicostegina, the lepidocyclinids, the nummulitids, and the miogypsinids. Helicostegina, the lepidocyclinids, and the miogypsinids all show a continuous endemic evolutionary development in the Americas, whereas the nummulitids show repeated episodes of migration into the region. The present scheme is largely based on the FAD or LAD of a single or two species. As further studies are completed, additional taxa will be added and this will lead to the recognition of assemblages rather than relying on single species or lineages. However, since the Eocene zonation is being applied to successions elsewhere (e.g., Torres-Silva & Hohenegger, 2024, for Cuba), we feel that publishing of the complete scheme at this time is useful. Refinements to the scheme can therefore be added as new information becomes available.
Our biostratigraphy has largely been based on sections in Jamaica, where most zones within the scheme are present (either in shallow-water limestones associated with carbonate platforms, or reworked into deep-water limestones flanking platforms (cf. Mitchell et al., 2022). The Gulf Coast of the United States (Florida and Alabama) contain good developments and some of the Paleocene to Early Miocene zones, whereas Antigua, Puerto Rico, and the Petite Antilles contain intervals associated with the early Oligocene, late Oligocene to Early Miocene, and Pliocene, respectively.
The
latest Cretaceous and Cenozoic saw major perturbations in marine environments
driven by extra-terrestrial impacts (K-Pg boundary), climatic optima and
Milankovitch-driven temperature/sea-level fluctuations. Shallow-water LBF living
on tropical carbonate platforms were significantly affected by some of these
events, and some of the American zonal boundaries correspond to such
environmental perturbations. The zonation scheme includes immigration events,
evolving lineages, and extinction events (Fig.
1 ![]() ).
).
|
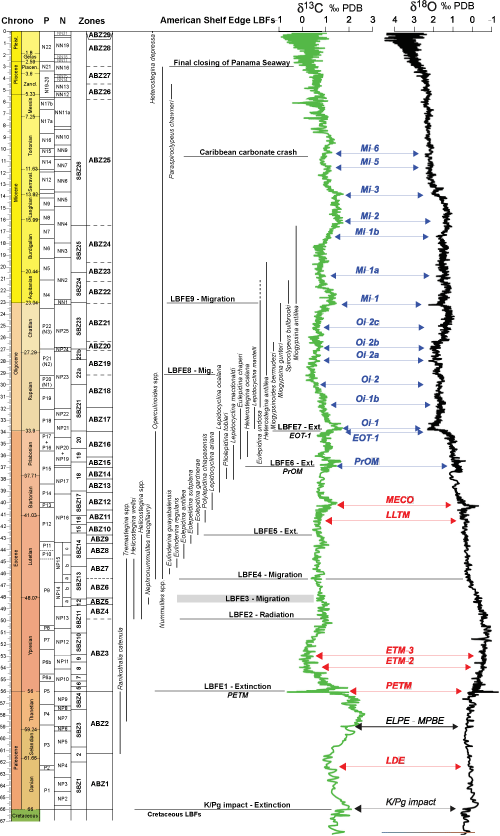
Figure 1:
ABZ for the Cenozoic of the Americas with ranges of selected larger benthic
foraminifers calibrated against calcareous nannoplakton and planktic
foraminifers, foraminiferal events (LBFE), oxygen and carbon isotope
stratigraphies (Speijer et
al., 2020), and global paleoenvironmental events. Standard Benthic Zones
(SBZ) for the Tethys are shown for comparison. EOT-1, Eocene-Oligocene terminal
event; PrOM, Priabonian thermal maximum; PETM, Paleocene-Eocene thermal maximum. |
The K-Pg extinction wiped out many American Cretaceous LBF and it was only in the mid Paleocene that new LBF appeared. These include the orthophragmines (which are not used in the current zonation) and Ranikothalia. The extinction of Ranikothalia at the Paleocene-Eocene boundary (Larger Benthic Foraminiferal Event 1: LBFE1) in the Americas is broadly coincident with the Paleocene-Eocene Thermal Maximum (PETM) suggesting environmental control.
In the mid Ypresian, a radiation involving Helicostegina began (LBFE2), but many of the species are difficult to distinguish in random sections and, other than for the easily recognizable Helicostegina wellsi, they are not used in the zonation. Helicostegina (excluding the unrelated Helicolepidina) disappears at the top of American Benthic Zone (ABZ) 9 (LBFE5).
Nummulitids (Nephronummulites macgillavryi) reappear in the latest Ypresian, but disappear at the boundary with the Lutetian. This short influx of Nephronummulites macgillavryi (LBFE4) with the microspheric form about eight times the size of the megalospheric form, suggests an affinity with Tethys forms (late Ypresian to Bartonian: e.g., BouDagher-Fadel & Price, 2014), which also show similar size differences. However, the embryo of megalospheric Nephronummulites macgillavryi has a large, broadly circular protoconch and a narrow crescent-shaped deteroconch (Mitchell et al., 2022), which contrasts with the sub-equal first two chambers in the embryos of the Tethys forms (BouDagher-Fadel & Price, 2014), and they may not be closely related.
A
major migration event marks the base of ABZ6 in the mid Lutetian (LBFE3), which
sees appearances of many taxa including small nummulitids (variously attributed
to Nummulites and Operculinoides),
Pseudolepidina, Linderina, and the first lepidocyclinids (Eulinderina and Planorbitoinella;
Mitchell et al., 2022). The
lepidocyclinids show a major radiation throughout the rest of the Eocene that is
used to establish a high-resolution zonation. The evolution from Eulinderina
through Eolepidina and Polylepidina
to Lepidocyclina (Fig. 2.7-11 ![]() )
shows a
reduction in the length of the primary spire from 7-9 chambers in Eulinderina
to an embryo of two chambers in Lepidocyclina,
and the development of lateral chambers from Eolepidina
onwards (Adams, 1987; Mitchell et al.,
2022). Populations of specimens show consistent numbers of
chambers in the primary spire and calculated means are used for the zonation.
The subsequent evolution of the lepidocyclinids shows the development of
adauxilary chambers from the deuteroconch and eventually the protoconch, and an
increase in the size of the deuteroconch which progressively enwraps the
protoconch (Fig. 2.15-16
)
shows a
reduction in the length of the primary spire from 7-9 chambers in Eulinderina
to an embryo of two chambers in Lepidocyclina,
and the development of lateral chambers from Eolepidina
onwards (Adams, 1987; Mitchell et al.,
2022). Populations of specimens show consistent numbers of
chambers in the primary spire and calculated means are used for the zonation.
The subsequent evolution of the lepidocyclinids shows the development of
adauxilary chambers from the deuteroconch and eventually the protoconch, and an
increase in the size of the deuteroconch which progressively enwraps the
protoconch (Fig. 2.15-16 ![]() ).
).
Two
LBF extinction events characterize the later Eocene. The top of ABZ14 (LBFE6)
sees the extinction of Pliolepidina (Fig. 2.13 ![]() )
and the platform-interior Yaberinella, and corresponds with the Priabonian oxygen maximum (PrOM). The top of ABZ16
(LBFE7) sees the loss of many typical Eocene taxa, including Heterostegina
ocalana, Fabularia verseyi, Coskinolina cf. douvillei,
Pellatispirella matleyi, Pseudochrysalidina,
Pseudophragmina, Asterocyclina,
Helicolepidina, and
Butterliniana
cubensis (Robinson, 2004) The top of ABZ16 is associated with an
oxygen stable isotope excursion marking the onset of glaciation in the Antarctic
and changes in ocean circulation patterns (Eocene-Oligocene Transition 1/EOT 1
and Oxygen isotope event 1/Oi 1 (Speijer et
al., 2020). We ascribe the occurrence of typical Eocene taxa in turbidites
in the early Oligocene of Cuba (Molina et
al., 2015) as probably due to reworking.
)
and the platform-interior Yaberinella, and corresponds with the Priabonian oxygen maximum (PrOM). The top of ABZ16
(LBFE7) sees the loss of many typical Eocene taxa, including Heterostegina
ocalana, Fabularia verseyi, Coskinolina cf. douvillei,
Pellatispirella matleyi, Pseudochrysalidina,
Pseudophragmina, Asterocyclina,
Helicolepidina, and
Butterliniana
cubensis (Robinson, 2004) The top of ABZ16 is associated with an
oxygen stable isotope excursion marking the onset of glaciation in the Antarctic
and changes in ocean circulation patterns (Eocene-Oligocene Transition 1/EOT 1
and Oxygen isotope event 1/Oi 1 (Speijer et
al., 2020). We ascribe the occurrence of typical Eocene taxa in turbidites
in the early Oligocene of Cuba (Molina et
al., 2015) as probably due to reworking.
During
the Oligocene important new taxa appear including Eulepidina
undosa (Fig. 3.2-3 ![]() ) and its allies, Heterostegina
antillea (Fig. 3.6-7
) and its allies, Heterostegina
antillea (Fig. 3.6-7 ![]() ), and the evolutionary development of the miogypsinids
(Fig. 3.4-5, 8-9
), and the evolutionary development of the miogypsinids
(Fig. 3.4-5, 8-9 ![]() ). The miogypsinids show a progressive evolution from a simple
spired (Neorotalia) ancestor with the
progressive development of a fan of equatorial chamberlets, lateral chamberlets, and
the reduction in the length of the primary spire. Some work utilizing random
sections (BouDagher-Fadel & Price,
2010) has suggested a
complex evolution, but the interpretation of random sections is not straight
forward. We presently use Cole's (1957) simplified classification of
the miogypsinids, but future work on assemblages of free specimens, tied to the
chronostratigraphy, rather than in random thin sections of rock is needed for
subsequent refinement of the zonation.
). The miogypsinids show a progressive evolution from a simple
spired (Neorotalia) ancestor with the
progressive development of a fan of equatorial chamberlets, lateral chamberlets, and
the reduction in the length of the primary spire. Some work utilizing random
sections (BouDagher-Fadel & Price,
2010) has suggested a
complex evolution, but the interpretation of random sections is not straight
forward. We presently use Cole's (1957) simplified classification of
the miogypsinids, but future work on assemblages of free specimens, tied to the
chronostratigraphy, rather than in random thin sections of rock is needed for
subsequent refinement of the zonation.
In
the Miocene there is a progressive loss of LBF diversity, with sequential
extinctions of Heterostegina antillea,
Miogypsina, Eulepidina, and Lepidocyclina. This presumably relates to long-term climatic change
linked to the development of nutrient upwelling in the Caribbean region (Edinger
& Risk, 1994) and finally a loss of carbonate productivity (the
Caribbean carbonate crash) (Roth et
al., 2000). Subsequently, in the Late Miocene, the platform margins of the
Americas had relatively low-diversity LBF assemblages dominated by Amphistegina
and soritids, although Nummulites and Paraspiroclypeus chawneri (ABZ26 and ABZ27) occur in the early
Pliocene (Andreieff, 1983). The final extinction of Nummulites might be due to the closure of the Panama Seaway at
around 2.7 Ma (Fig.
1 ![]() ; e.g.,
Lunt et al., 2008; Öğretmen et
al., 2020). Heterostegina depressus
appears in the late Holocene as an immigrant from Tethys and can be taken as a
biostratigraphic proxy close to the base of the Anthropocene (ABZ29).
; e.g.,
Lunt et al., 2008; Öğretmen et
al., 2020). Heterostegina depressus
appears in the late Holocene as an immigrant from Tethys and can be taken as a
biostratigraphic proxy close to the base of the Anthropocene (ABZ29).
The
new zonation (Fig.
1 ![]() ) will allow a correlation of successions across the
tropical Americas with the international chronostratigraphy and allow a better
understanding of the development of carbonate platforms during the Cenozoic.
) will allow a correlation of successions across the
tropical Americas with the international chronostratigraphy and allow a better
understanding of the development of carbonate platforms during the Cenozoic.
|
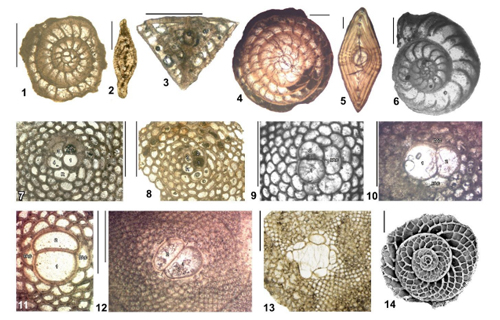
Figure 2:
Cenozoic foraminifera from the Americas (Jamaica unless otherwise stated):
1 (E) - 2 (A), Ranikothalia catenula (Cushman
& Jarvis), EO.WL2062-33, Nonsuch Formation, Nonsuch;
3 (A), Helicostegina
wellsi (Cole & Bermúdez), EO.WL2044-75, Chapelton
Formation, Lilyfield; 4 (E) - 5 (A), Nephronummulites
macgillavryi (Rutten), UWIGM.WL3315-05/03, Palmetto Grove Formation,
Hampstead; 6 (E), Operculinoides sp.,
UWIGM.WL3353-07, Preston Hill Formation, Jeffrey Town; 7 (E), Eolepidina
antillea (Cushman), EO.WL2046-25, Preston Hill Formation,
Lilyfield; 8 (E), Eo. subplana Barker & Grimsdale, EO.WL2044-36),
Chapelton Formation, Lilyfield; 9 (E), Eo.
gardnerae Cole, UWIGM.Content.3327ft-6, Yellow Limestone, Content
Well (depth 3327ft = 1014 m); 10 (E), Polylepidina
chiapasensis Vaughan, UWIGM.WL2837-08, Preston Hill Formation, Salt
Spring; 11 (E) Lepidocyclina ariana Cole
& Ponton, UWIGM.WL3571-2, Yellow Limestone, Grants
Pen; 12 (E) Lepidocyclina
ocalana Cushman, UWIGM.WL2635-9, Ipswich Formation,
Ipswich; 13 (E)
Pliolepidina tobleri Douvillé,
NHM.PM.P.52831, Nicaragua; 14 (E), Heterostegina
ocalana Cushman, ER176, White Limestone (chalk), Dressikie. E
- equatorial section, A - axial section. Scale bars - 500 μm. NHM,
Natural History Museum London. OZ, Geological Engineering Department of Istanbul
Technical University, Turkey; UWIGM, UWI Geology Museum, Kingston, Jamaica. |
|
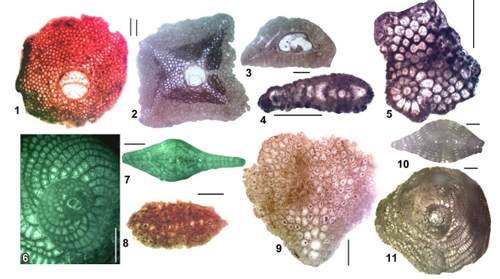
Figure 3:
Cenozoic foraminifera from the Americas (Jamaica): 1 (E) Eulepidina
chaperi (Lemoine & R. Douvillé), UWIGM.WL4996-06, Browns
Town Formation, Browns Town (ABZ17); 2 (E, UWIGM.WL4475B-3) - 3 (A,
UWIGM.WL4475B-10), Eulepidina undosa Cushman, Chalk, Ramble, Westmoreland
(ABZ18); 4-5 (UWIGM.R118), Miogypsinoides
bermudezi Drooger, chalk, Sherwood Content (ABZ20);
6
(UWIGM.WL4498B-2) - 7 (UWIGM.WL4498B-1), Heterostegina
antillea Cushman, chalk, Montego Bay (ABZ23); 8 (E, UWIGM.WL5008.20)
- 9 (A, UWIGM.WL5008.21), Miogypsina
tani Drooger, Montpelier Formation, Hope Bay, Portland
(ABZ22); 10
(UWIGM.WL5008.2) - 11 (UWIGM.WL5008.5), Spiroclypeus
bullbrooki (Vaughan & Cole), Montpelier Formation, Hope
Bay, Portland (ABZ22). Scale
bar = 500 μm. |
ABZ1. From the LAD of Cretaceous LBFs to the FAD of Ranikothalia spp. The end Cretaceous mass extinction wiped out the LBF assemblages of the Americas which were characterized by species including Orbitoides, Omphalocyclus, and Chubbina (e.g., Gunter et al., 2002; Mitchell, 2005). ABZ1 represents a low diversity zone before the appearance of new LBF lineages in the Paleogene. ABZ1 is calibrated with zones NP1 to NP4. The zone is well-developed in the Nonsuch Limestone in Jamaica (Robinson & Wright, 1993) and Costa Rica (Baumgartner-Mora & Baumgartner, 2017).
ABZ2.
From the FAD to
the LAD of Ranikothalia spp. The zone
contains Ranikothalia spp. (Fig. 2.1-2 ![]() ) in association with orthophragmines (Özcan et
al., 2019, 2022) and can be calibrated with NP5 to NP7 in Jamaica (Jiang & Robinson,
1987). Based on Sr-isotopes Ranikothalia
catenula ranges through the Seladian and Thanetian, but could potentially
range into the early Ypresian based on the associated error bars (Baumgartner-Mora
& Baumgartner, 2017). We provisionally place the top of ABZ2 at the
top of the Thanetian (top of NP9), pending further work. The zone marks the
probable first migration of nummulitids into the American province during the
Cenozic. Subdivision of the zone may be possible based on orthophragmines (Özcan
et al., 2019), but further work is needed.
) in association with orthophragmines (Özcan et
al., 2019, 2022) and can be calibrated with NP5 to NP7 in Jamaica (Jiang & Robinson,
1987). Based on Sr-isotopes Ranikothalia
catenula ranges through the Seladian and Thanetian, but could potentially
range into the early Ypresian based on the associated error bars (Baumgartner-Mora
& Baumgartner, 2017). We provisionally place the top of ABZ2 at the
top of the Thanetian (top of NP9), pending further work. The zone marks the
probable first migration of nummulitids into the American province during the
Cenozic. Subdivision of the zone may be possible based on orthophragmines (Özcan
et al., 2019), but further work is needed.
ABZ3. From the LAD of Ranikothalia catenula to the FAD of Helicostegina wellsi. The zone lacks diverse LBF other than for Tremastegina, orthophragmines, and dictyoconids. We calibrate ABZ3 with NP10 to NP12. Localities in the Americas with good representatives of zones ABZ3 to ABZ16 have been given by Mitchell et al. (2022).
ABZ4.
From the FAD of Helicostegina wellsi to the FAD
of Nephronummulites macgillavryi. The
zone sees the appearance of Helicostegina (=Eoconuloides)
in abundance and represents an increase in diversity of LBF. Helicostegina
wellsi (Fig. 2.3 ![]() ) is distinctive, but relatively long ranging. The zone can
be calibrated with lower NP14a (Mitchell et
al., 2022). LBF faunas from NP13 are at present unknown.
) is distinctive, but relatively long ranging. The zone can
be calibrated with lower NP14a (Mitchell et
al., 2022). LBF faunas from NP13 are at present unknown.
ABZ5.
From the FAD to the LAD of Nephronummulites
macgillavryi (Fig. 2.4-5 ![]() ). The zone can be calibrated with upper NP14a (Mitchell et
al., 2022). The zone marks the second migration of nummulitids into the
American province. The top of the zone corresponds with, or is just below, the
Ypresian-Lutetian boundary.
). The zone can be calibrated with upper NP14a (Mitchell et
al., 2022). The zone marks the second migration of nummulitids into the
American province. The top of the zone corresponds with, or is just below, the
Ypresian-Lutetian boundary.
ABZ6. From the LAD of Nephronummulites macgillavryi to the FAD of Operculinoides spp./Nummulites spp. The zone is a relatively low-diversity interval during which nummulitids are absent. The zone can be calibrated with NP14b (Mitchell et al., 2022).
ABZ7.
From the FAD of Operculinoides spp./Nummulites
spp. to the FAD of Eolepidina antillea.
The base of the zone is marked by the appearance of numerous taxa including Operculinoides
spp. (Fig. 2.6 ![]() ), Nummulites ex gr. striatoreticulatus, Pseudolepidina
trimera, Eulinderina guayabalensis,
Linderina floridensis, and others,
which represents a major migration event (the third migration of nummulitids
into the American province). The zone is calibrated with NP15b (Mitchell et
al., 2022). NP15a is represented by a hiatus in Jamaica. Elsewhere, more
primitive taxa of some lineages (without independent calibration) are associated
with Operculinoides spp. and we tentatively place the base of the zone
within NP15a (Mitchell et al., 2022).
), Nummulites ex gr. striatoreticulatus, Pseudolepidina
trimera, Eulinderina guayabalensis,
Linderina floridensis, and others,
which represents a major migration event (the third migration of nummulitids
into the American province). The zone is calibrated with NP15b (Mitchell et
al., 2022). NP15a is represented by a hiatus in Jamaica. Elsewhere, more
primitive taxa of some lineages (without independent calibration) are associated
with Operculinoides spp. and we tentatively place the base of the zone
within NP15a (Mitchell et al., 2022).
ABZ8. From the FAD of Eolepidina antillea to the FAD of Eolepidina subplana. The zone calibrates with upper NP15b to NP15c. ABZ7 to ABZ12 show the progressive evolution of primitive members of the lepidocyclinid lineage and show the appearance of lateral chamberlets and the gradual reduction of coiled equatorial chambers to a Lepidocyclina-type embryo (Mitchell et al., 2022).
ABZ9. From the FAD of Eolepidina subplana to the FAD of Eolepidina gardnerae. The zone is calibrated with lower NP16 (Mitchell et al., 2022).
ABZ10. From the FAD of Eolepidina gardnerae to the FAD of Polylepidina chiapasensis. The zone is calibrated with lower mid NP16 (Mitchell et al., 2022).
ABZ11. From the FAD of Polylepidina chiapasensis to the FAD of Lepidocyclina ariana. The zone can be calibrated with upper mid NP16 (Mitchell et al., 2022).
ABZ12. From the FAD of Lepidocyclina ariana to the FAD of Lepidocyclina ocalana. The zone is calibrated with upper NP16 to possibly lower NP17 (Mitchell et al., 2022).
ABZ13. From the FAD of Lepidocyclina ocalana to the FAD of Pliolepidina tobleri. The assemblage is characterised by the appearance of species of Lepidocyclina with adauxillary chambers. We calibrate ABZ13 primarily with lower-mid NP17 (Mitchell et al., 2022).
ABZ14. From the FAD to the LAD of Pliolepidina tobleri. The zone is characterized by the distinctive species Pliolepidina tobleri. We calibrate ABZ14 with upper NP17 (Mitchell et al., 2022).
ABZ15. From the LAD of Pliolepidina tobleri to the FAD of Heterostegina ocalana. We calibrate ABZ15 with NP18 and lower NP19/20 (sensu Martini, 1971).
ABZ16. From the FAD to the LAD of Helicostegina ocalana. The zone corresponds to the migration of Helicostegina ocalana into the Americas. We assign ABZ16 to NP19/20 through lower NP21 (Mitchell et al., 2022).
ABZ17.
From the LAD of Heterostegina ocalana
to the FAD of Eulepidina undosa. It is
characterized by a relatively low diversity assemblage of LBFs including Eulepidina
chaperi (Fig. 3.1 ![]() ), Lepidocyclina yurnagunensis, and Orbitoina sp. in the Walderston
and lower Browns Town limestones of Jamaica and Lepidocyclina
mantelli and Nummulites dia in the
Marianna Limestone of the Gulf Coast (Vaughan, 1927; Cole & Ponton,
1930). We calibrate ABZ17 with mid NP21 to NP22.
), Lepidocyclina yurnagunensis, and Orbitoina sp. in the Walderston
and lower Browns Town limestones of Jamaica and Lepidocyclina
mantelli and Nummulites dia in the
Marianna Limestone of the Gulf Coast (Vaughan, 1927; Cole & Ponton,
1930). We calibrate ABZ17 with mid NP21 to NP22.
ABZ18.
From the FAD of Eulepidina undosa to
the FAD of Heterostegina antillea. The
LBF are characterized by Eulepidina undosa (Fig. 3.2-3 ![]() ), Eu.
favosa, Nephrolepidina spp., Lepidocyclina
supera, and Nummulites spp. Sr
ratios determined from screened calcite in Kuphus
tubes from ABZ18 indicate ages ranging from 28.5 to 31.5 Ma (late Rupelian). In
Alabama Eulepidina undosa appears in lower
P19/lower O2 (Gravel & Hanna,
1937; Miller et
al., 2008) and we place the base of ABZ18 at this level. The zone is widely
distributed in the Americas from northern Florida and Jamaica.
), Eu.
favosa, Nephrolepidina spp., Lepidocyclina
supera, and Nummulites spp. Sr
ratios determined from screened calcite in Kuphus
tubes from ABZ18 indicate ages ranging from 28.5 to 31.5 Ma (late Rupelian). In
Alabama Eulepidina undosa appears in lower
P19/lower O2 (Gravel & Hanna,
1937; Miller et
al., 2008) and we place the base of ABZ18 at this level. The zone is widely
distributed in the Americas from northern Florida and Jamaica.
ABZ19.
From the FAD of Heterostegina antillea
to the FAD of Miogypsinoides bermudezi.
This zone contains a LBF assemblage consisting of Heterostegina
antillea
(Fig. 3.6-7 ![]() ), Eulepidina undosa, Eu. favosa, Eu.
gigas, Nephrolepidina spp., Nummulites
spp.,, and Neorotalia mexicana. We
place the base of ABZ19 at the base of zone P21 (Robinson,
2004). The
zone is well developed in the upper part of the Browns Town Formation in Jamaica
(Mitchell, 2013).
), Eulepidina undosa, Eu. favosa, Eu.
gigas, Nephrolepidina spp., Nummulites
spp.,, and Neorotalia mexicana. We
place the base of ABZ19 at the base of zone P21 (Robinson,
2004). The
zone is well developed in the upper part of the Browns Town Formation in Jamaica
(Mitchell, 2013).
ABZ20.
From the FAD of Miogypsinoides bermudezi
to the FAD of Miogypsina gunteri. The
base of the zone corresponds to the evolutionary development of Miogypsinoides (Fig.
3.4-5 ![]() )
from Neorotalia. Earlier work (Drooger, 1952,
1993) suggested an idealized evolutionary scheme for the miogypsinids of
the Americas, however they were poorly stratigraphically constrained and
subsequent work has not confirmed this (Baumgartner-Mora et
al., 2008). The LBF includes Heterostegina, Eulepidina undosa, and Miogypsinoides.
In Antigua Miogypsinoides first
appears within zone NP25 near the base of P22 and this is confirmed based on
Sr-isotope ratios (Robinson et al., 2017). The zone is well-represented in the Antigua Formation of Antigua (Robinson
et al., 2017) and in deep-water chalks
in northern Jamaica (unpublished).
)
from Neorotalia. Earlier work (Drooger, 1952,
1993) suggested an idealized evolutionary scheme for the miogypsinids of
the Americas, however they were poorly stratigraphically constrained and
subsequent work has not confirmed this (Baumgartner-Mora et
al., 2008). The LBF includes Heterostegina, Eulepidina undosa, and Miogypsinoides.
In Antigua Miogypsinoides first
appears within zone NP25 near the base of P22 and this is confirmed based on
Sr-isotope ratios (Robinson et al., 2017). The zone is well-represented in the Antigua Formation of Antigua (Robinson
et al., 2017) and in deep-water chalks
in northern Jamaica (unpublished).
ABZ21.
From the FAD of Miogyspina gunteri to
the FAD of Spiroclypeus bullbrooki.
The zone is characterized by Miogyspina (Fig. 3.9 ![]() )
with long primary spires of more than one whorl (Robinson
et al., 2017).
The LBF assemblage includes Eulepidina undosa, Miogyspina gunteri, and Heterostegina antillea.
In Puerto Rico Miogyspina gunteri occurs in
the San Sabastian Formation (Sachs, 1959) that is calibrated with the
late Oligocene based on Sr isotopes (Ortega-Ariza et al., 2015). This is consistent with Sr-isotope and
biostratigraphic data from Carriacou (Baumgartner-Mora et
al., 2008), and we place the base of ABZ21 low in zones P23 and NP25. The
zone is represented in Puerto Rico and Caaiacou (Sachs, 1959; Baumgartner-Mora
et al., 2008), but has not yet been recorded in Jamaica.
)
with long primary spires of more than one whorl (Robinson
et al., 2017).
The LBF assemblage includes Eulepidina undosa, Miogyspina gunteri, and Heterostegina antillea.
In Puerto Rico Miogyspina gunteri occurs in
the San Sabastian Formation (Sachs, 1959) that is calibrated with the
late Oligocene based on Sr isotopes (Ortega-Ariza et al., 2015). This is consistent with Sr-isotope and
biostratigraphic data from Carriacou (Baumgartner-Mora et
al., 2008), and we place the base of ABZ21 low in zones P23 and NP25. The
zone is represented in Puerto Rico and Caaiacou (Sachs, 1959; Baumgartner-Mora
et al., 2008), but has not yet been recorded in Jamaica.
ABZ22.
From the FAD
to the LAD of Spiroclypeus bullbrooki. The zone
is characterized by Miogypsina with a spire of less than one whorl in association with Nummulites,
Lepidocyclina canellei, and small specimens of Miogyspina tani. Spiroclypeus bullbrooki
(Fig. 3.10-11 ![]() )
appears suddenly at the base of the zone as an immigrant. This LBF assemblage
has previously been assigned to the late Oligocene due to the higher placement
of the Oligocene-Miocene boundary in the Americas compared with Europe (Vaughan & Cole,
1943; Akers & Drooger,
1957; Cole, 1957). However Spiroclypeus bullbrookei
and Miogyspina tani occur in zone N4 (=M1) in eastern Jamaica (Robinson,
2004)
and we place the base of ABZ22 at the base of N4. The zone is well-developed in
eastern Jamaica (Robinson, 2004).
)
appears suddenly at the base of the zone as an immigrant. This LBF assemblage
has previously been assigned to the late Oligocene due to the higher placement
of the Oligocene-Miocene boundary in the Americas compared with Europe (Vaughan & Cole,
1943; Akers & Drooger,
1957; Cole, 1957). However Spiroclypeus bullbrookei
and Miogyspina tani occur in zone N4 (=M1) in eastern Jamaica (Robinson,
2004)
and we place the base of ABZ22 at the base of N4. The zone is well-developed in
eastern Jamaica (Robinson, 2004).
ABZ23. From the LAD of Spiroclypeus bullbrooki to the LAD of Heterostegina antillea. The last representatives of Eulepidina and Lepidocyclina also occur in this zone. The top of ABZ23 is placed within N5 (Robinson, 2004). Zones ABZ23 to ABZ28 are developed only locally, and further work is needed to understand their foraminiferal assemblages.
ABZ24. From the LAD of Heterostegina antillea to the LAD of Miogypsina. This marks the final extinction of Miogypsina in the Americas. The top of ABZ24 is placed at the top of N7 (Robinson, 2004).
ABZ25. From the LAD of Miogypsina to the FAD of Paraspiroclypeus chawneri. The zone is characterized by the presence of Nummulites, but cannot at present be distinguished from ABZ27 unless ABZ26 is present.
ABZ26. From the FAD to the LAD of Paraspiroclypeus chawneri. In the Petites Antilles the range of Paraspiroclypeus chawneri is calibrated with zones N18 and N19 (Andreieff, 1983).
ABZ27.
From the LAD of Paraspiroclypeus chawneri to the LAD
of Nummulites. In the Petites Antilles
Nummulites ranges up into the middle
part of N21 (Andreieff, 1983). This might be related to the final closure
of the Panama Seaway (Fig.
1 ![]() ; e.g., Lunt et al.,
2008; Öğretmen et al.,
2020).
; e.g., Lunt et al.,
2008; Öğretmen et al.,
2020).
ABZ28. From the LAD of Nummulites to the FAD of Heterostegina depressa. The zone is characterized by the presence of Amphistegina in the absence of Nummulites.
ABZ29. From the FAD of Heterostegina depressa to the present. The zone is characterized by the presence of Heterostegina depressa (probably a Tethys immigrant brought in via ships) in association with Amphistegina and is a proxy for the Anthropocene. This foraminiferan is found in modern beach sands in the Caribbean.
We are currently studying selected sections in the Americas to provide additional information on foraminiferal assemblages associated with each zone. We will publish these updates separately which will lead to a more robust understanding of the foraminiferal succession through the Paleogene and Neogene.
We thank Julian Williams and Roshaun Brown for providing field support to SFM while mapping, collecting material and processing material for LBFs. EÖ thanks İstanbul Technical University BAP Unit for financial support for a visit to the University of the West Indies, Mona, for 3-months duration in 2012. We thank the two reviewers (Giovanni Coletti and one anonymous) for their very useful comments that helped improve the manuscript and the editor (Daniela Maria Basso) for her help in completing the manuscript.
S.F.M. designed the project, collected and cut material, prepared photographs and taxonomic reviews, prepared the initial draft, edited the initial draft. E.R. provided photographs and taxonomic reviews, edited the initial draft. E.Ö. collected and cut material, provided photographs and taxonomic reviews, edited the initial draft. M.M.J. undertook nannofossil identification on split samples, edited the initial draft. N.R. prepared thin sections, edited the initial draft.
All data are available in the main text (specimens are deposited in the UWI Geological Museum, Jamaica, and the Geological Engineering Department of Istanbul Technical University, Turkey)
Adams C.G. (1970).- A reconsideration of the East Indian letter classification of the Tertiary.- Bulletin of the British Museum (Natural History), Geology, London, vol. 19, no. 3, p. 85-137.
Adams C.G. (1987).- On the classification of the Lepidocyclinidae (Foraminiferida) with redescriptions of the unrelated Paleocene genera Actinosiphon and Orbitosiphon.- Micropaleontology, Flushing - NY, vol. 33, no. 4, p. 289-317.
Akers W.H. & Drooger C.W. (1957).- Miogypsinids, planktonic foraminifera, and Gulf Coast Oligocene-Miocene correlation.- AAPG Bulletin, Tulsa - OK, vol. 41, no. 4, p. 656-678.
Andreieff P. (1983).- Extension stratigraphique des grands Foraminifères néogènes de la région caraïbe : Paraspiroclypeus chawneri (Palmer) et Operculinoides cojimarensis (Palmer).- Bulletin de la Société Géologique de France (7e série), Paris, t. XXV, no. 6, p. 885-888.
Baumgartner-Mora C. & Baumgartner P.O. (2017).- Paleocene-earliest Eocene Larger Benthic Foraminifera and Ranikothalia-bearing carbonate paleo-environments of Costa Rica (South Central America).- Micropaleontology, Flushing - NY, vol. 62, no. 6, p. 453-508.
Baumgartner-Mora C., Baumgartner P.O. & Tschudin P. (2008).- Late Oligocene larger foraminifera from Nosara, Nicoya Península (Costa Rica) and Windward, Carriacou (Lesser Antilles), calibrated by 87Sr/86Sr isotope stratigraphy.- Revista Geológica de América Central, San José, Costa Rica, vol. 38, p. 33-52. URL: https://revistas.ucr.ac.cr/index.php/geologica/article/view/4215/4039
BouDagher-Fadel M.K. & Price G.D. (2010).- American Miogypsinidae: An analysis of their phylogeny and biostratigraphy.- Micropaleontology, Flushing - NY, vol. 56, no. 6, p. 567-586.
BouDagher-Fadel M.K. & Price, G.D. (2014).- The phylogenetic and palaeogeographic evolution of the nummulitoid larger benthic foraminifera.- Micropaleontology, Flushing - NY, vol. 60, no. 6, p. 483-508.
Butterlin J. (1981).- Claves para la determinación de macroforaminiferos de Mexico y del Caribe, del Cretacio superior al Mioceno medio.- Instituto Mexicano del Petroleo, Mexico D.F., 219 p.
Cahuzac B. & Poignant A. (1997).- Essai de biozonation de l'Oligo-Miocène dans les bassins européens à l'aide des grands foraminifères néritiques.- Bulletin de la Société Géologique de France, Paris, t. 168, no. 2, p. 155-169.
Cole W.S. (1957).- Late Oligocene larger foraminifera from Barro Colorado Island, Panama Canal Zone.- Bulletins of American Paleontology, Ithaca - NY, vol. 37, p. 313-338. URL: https://www.biodiversitylibrary.org/item/40446#page/7/mode/1up
Cole W.S. & Ponton G.M. (1930).- The Foraminifera of the Marianna Limestone of Florida.- Florida State Geological Survey, Bulletin, Tallahassee - FL, vol. 5, p. 19-69. URL: https://ufdc.ufl.edu/UF00000436/00001/19j
Drooger C.W. (1952).- Study of American Miogypsinidae.- Vonk & Co's Drukkeeij, Zeist, p. 1-80.
Drooger C.W. (1993).- Radial Foraminifera: Morphometrics and evolution.- Verhandelingen der Koninklijke Nederlandse Akademie van Wetenschappen (Afd. Natuurkunde), Amsterdam, vol. 41, p. 1-242.
Edinger E.N. & Risk M.J. (1994).- Oligocene-Miocene extinction and geographic restriction of Caribbean corals: Roles of turbidity, temperature, and nutrients.- Palaios, Tulsa - OK, vol. 9, no. 6, p. 576-598.
Gravel D.W. & Hanna M.A. (1937).- The Lepidocyclina texana horizon in the Heterostegina zones, upper Oligocene, of Texas and Louisiana.- Journal of Paleontology, Tulsa - OK, vol. 11, no. 6, p. 517-529.
Gunter G.C., Robinson E. & Mitchell S.F. (2002).- A new species of Omphalocyclus (Foraminiferida) from the Upper Cretaceous of Jamaica and its stratigraphical significance.- Journal of Micropaleontology, London, vol. 21, no. 2, p. 149-153. DOI: 10.1144/jm.21.2.149
Jiang M.-J. & Robinson E. (1987).- Calcareous nannofossils and larger Foraminifera in Jamaican rocks of Cretaceous to early Eocene age. In: Ahmad R. (ed.), Proceedings of a workshop on the status of Jamaican geology.- Geological Society of Jamaica, Kingston, Jamaica, p. 24-51.
King D.J., Wade B.S., Liska R.D. & Miller C.G. (2020).- A review of the importance of the Caribbean region in Oligo-Miocene low latitude planktonic foraminiferal biostratigraphy and the implications for modern biogeochronological schemes.- Earth Science Reviews, vol. 202, article #102968, 27 p.
Less G. & Özcan E. (2012).- Bartonian-Priabonian larger benthic foraminiferal events in the Western Tethys.- Austrian Journal of Earth Sciences, Vienna, vol. 105, no. 1, p. 129-140.
Lunt D.J., Valdes P.J., Haywood A. & Rutt I.C. (2008).- Closure of the Panama Seaway during the Pliocene: implications for climate and Northern Hemisphere glaciation.- Climate Dynamics, vol. 30, p. 1-18.
Martini E. (1971).- Standard Tertiary and Quaternary calcareous nannoplankton zonation. In: Farinacci A. (ed.), Proceedings of the II Planktonic Conference, Roma, 1969.- Edizioni Tecnoscienza, Roma, p. 739-785.
Mello e Sousa S.H. de, Fairchild T.R. & Tibana P. (2003).- Cenozoic biostratigraphy of larger foraminifera from the Foz do Amazonas Basin, Brasil.- Journal of Micropaleontology, London, vol. 49, no. 3, p. 253-266.
Miller K.G., Browning J.V., Aubry M.-P., Wade B.S., Katz M., Kulpecz A.A. & Wright J.D. (2008).- Eocene-Oligocene global climate and sea-level changes: St. Stephens Quarry, Alabama.- GSA Bulletin, Denver - CO, vol. 120, no. 1-2, p. 34-53.
Mitchell S.F. (2005).- Biostratigraphy of Late Maastrichtian larger foraminifers in Jamaica and the importance of Chubbina as a late Maastrichtian index fossil.- Journal of Micropalaeontology, London, vol. 24, p. 123-130.
Mitchell S.F. (2013).- Stratigraphy of the White Limestone of Jamaica.- Bulletin de la Société Géologique de France, Paris, t. 184, no. 1-2, p. 111-118.
Mitchell S.F., Robinson E., Özcan E., Jiang M.M. & Robinson N. (2022).- A larger benthic foraminiferal zonation for the Eocene of the Caribbean and central American region.- Carnets Geol., Madrid, vol. 22, no. 11, p. 409-565. DOI: 10.2110/carnets.2022.2211
Molina E., Torres-Silva A.I., Ćorić S. & Briguglio A. (2015).- Integrated biostratigraphy across the Eocene/Oligocene boundary at Noroña, Cuba, and the question of the extinction of orthophragminids.- Newsletters on Stratigraphy, Stuttgart, vol. 49, p. 27-40.
Öğretmen N., Schiebel R. Jochum K.P., Stoll B., Weis U., Repschläger J., Jentzen A., Galer S. & Haug G.H. (2020).- Deep thermohaline circulation across the closure of the Central American Seaway.- Paleoceanography and Paleoclimatology, vol. 35, no. 12, article e2020PA004049, 15 p. DOI: 10.1029/2020PA004049
Ortega-Ariza D., Franseen E.K., Santos-Mercado H., Ramírez-Martínez W.R. & Core-Suárez E.E. (2015).- Strontium isotope stratigraphy for Oligocene-Miocene carbonate systems in Puerto Rico and the Dominican Republic: Implications for Caribbean processes affecting depositional history.- Journal of Geology, Chicago - IL, vol. 123, no. 6, p. 539-560.
Özcan E., Mitchell S.F., Less G., Robinson E., Bryan J.R., Pignatti J. & Yücel A.O. (2019).- A revised suprageneric classification of American orthophragminids with emphasis on late Paleocene representatives from Jamaica and Alabama.- Journal of Systematic Palaeontology, vol. 17, no. 18, p. 1551-1579.
Özcan E., Yücel A.O., Erkizan L.S., Gültekin M.N., Kayğılı S. & Yurtsever S. (2022).- Atlas of the Tethyan orthophragmines.- Mediterranean Geoscience Reviews, vol. 4, p. 3-213.
Papazzoni C.A., Ćosović V., Briguglio A. & Drobne K. (2017).- Towards a calibrated larger Foraminifera biostratigraphic zonation: Celebrating 18 years of the application of shallow benthic zone.- Palaios, Lawrence - KS, vol. 32, no. 1, p. 1-5.
Papazzoni C.A., Fornaciari B., Giusberti L., Simonato M. & Fornaciari E. (2023).- A new definition of the Paleocene Shallow Benthic Zones (SBP) by means of larger foraminiferal biohorizons, and their calibration with calcareous nannofossil biostratigraphy.- Micropaleontology, Flushing - NY, vol. 69, no. 4-5, p. 363-399.
Robinson E. (1977).- Larger imperforate foraminiferal zones of the Eocene of central Jamaica. In: Segundo Congreso Latinoamericano de Geología, Caracas, Venezuela, 11 al 16 de Noviembre de 1973, Tomo III.- Ministerio de Minas e Hidrocarburos, Caracas, p. 1413-1421.
Robinson E. (2004).- Zoning the White Limestone Group of Jamaica using larger foraminiferal genera: A review and proposal. In: Donovan S.K. (ed.), The mid-Cainozoic White Limestone Group of Jamaica.- Cainozoic Research, Leiden, vol. 3, no. 1-2, p. 39-75. URL: https://natuurtijdschriften.nl/pub/541702/CR2004003001004.pdf
Robinson E. & Jiang M.-J. (1995).- Evolution of the foraminiferal genus Lepidocyclina in the Middle Eocene and its implication for Gulf Coast stratigraphy.- Gulf Coast Association of Geological Societies, Transactions, Baton Rouge - LA, vol. 45, p. 509-517.
Robinson E. & Mitchell S.F. (1999).- Middle Eocene to Oligocene stratigraphy and palaeogeography in Jamaica: A window on the Nicaragua Rise.- Contributions to Geology, UWI, Mona, Kingston, Jamaica, no. 4, 47 p. URL: http://www.sfmgeology.com/pdfs/MiddleEocenetoOligoceneStratigraphy.pdf
Robinson E. & Wright R.M. (1993).- Jamaican Paleogene larger foraminifera. In: Wright R.M. & Robinson E. (eds.), Biostratigraphy of Jamaica.- Geological Society of America, Memoir, Tulsa - OK, no. 182, p. 283-345.
Robinson E., Paytan A. & Chien C.-T. (2017).- Strontium isotope dates for the Oligocene Antigua Formation, Antigua, W.I.- Caribbean Journal of Earth Science, Kingston, Jamaica, vol. 50, p. 11-18. URL: http://caribjes.com/CJESpdf/CJES50-3-Robinson-Antigua.pdf
Roth J.M., Droxler A.W. & Kameo K. (2000).- 17. The Caribbean carbonate crash at the middle to late Miocene transition: Linkage to the establishment of the modern global ocean conveyor. In: Leckie R.M., Sigurdsson H., Acton G.D. & Draper G. (eds.), Caribbean ocean history and the Cretaceous/tertiary boundary event.- Proceedings of the Ocean Drilling Program, Scientific Results, College Station - TX, vol. 165, p. 249-273. URL: http://www-odp.tamu.edu/publications/165_SR/VOLUME/CHAPTERS/SR165_17.PDF
Sachs K.N. Jr (1959).- Puerto Rican upper Oligocene larger Foraminifera.- Bulletins of American Paleontology, Ithaca - NY, vol. 39, p. 399-416. URL: https://www.biodiversitylibrary.org/item/40436
Serra-Kiel J., Hottinger L., Caus E., Drobne K., Ferràndez C., Kumar Jauhri A., Less G., Pavlovec R., Pignatti J., Samsó J.M., Schaub H., Sirel E., Strougo A., Tambareau Y., Tosquella J. & Zakrevskaya E. (1998).- Larger foraminiferal biostratigraphy of the Tethyan Paleocene and Eocene.- Bulletin de la Société Géologique de France, Paris, t. 169, no. 2, p. 281-299.
Sierra D.A., Aguilera O.M., Araújo O.M.O. de, Lopes R.T., Geraldes M., Martins M.V.A., Coletti G., Guimarães B.T., Linhares A.P. & Kütter V.T. (2023).- Cenozoic biostratigraphy of larger foraminifera from equatorial carbonate platform of northwestern Brazil.- Marine and Petroleum Geology, vol. 156, article 106458, 16 p.
Speijer R.P., Pälike H., Hollis C.J., Hooker J.J. & Ogg G. (2020).- The Paleogene Period.- In: Gradstein F.M., Ogg J.G., Schmitz M. & Ogg G. (eds.), The Geologic Time Scale 2012.- Elsevier, Amsterdam, p. 1087-1140.
Torres-Silva A.I. & Hohenegger J. (2024).- Objective identification of Lepidocyclina (Foraminifera) species from the Eocene of Cuba based on growth-invariant morphometric characters.- Journal of Systematic Palaeontology, London, vol. 22, no. 1, article 2287728, 42 p. DOI: 10.1080/14772019.2023.2287728
Vandenberghe N., Hilgen F.L. & Speijer R.P. (2012).- The Paleogene Period. In: Gradstein F.M., Ogg J.G., Schmitz M. & Ogg G. (eds.), The Geologic Time Scale 2012.- Elsevier, Amsterdam, p. 855-921.
Vaughan T.W. (1927).- Notes on the types of Lepidocyclina mantelli (Morton) Gümbel and on topotypes of Nummulites floridanus Conrad.- Proceedings of the Academy of Natural Sciences Philadelphia, vol. LXXIX, p. 299-303.
Vaughan T.W. & Cole W.S. (1943).- Preliminary report of the Cretaceous and Tertiary larger Foraminifera of Trinidad British West Indies.- GSA Special Publications, Boulder - CO, vol. 30, 137 p.