◄ Carnets Geol. 25 (2) ►
![]()
Outline:
[1. Introduction][2. Geological context]
[3. History of previous geological work]
[4. Material and methods] [5. Biostratigraphic data]
[6. Are Busot calpionellids reworked or not?] [7. Conclusions]
[Bibliographic references] [Plates] and ... [Appendix]
D�pt. STU, Fac. Sci. Tech., UBO, 6 avenue Le Gorgeu, CS 93837, F-29238 Brest (France);
Membre correspondant, Mus�um d'Histoire Naturelle (MHN), Route de Malagnou 1, 1208 Gen�ve (Switzerland);
2 impasse Charles Martel, F-29217 Plougonvelin (France)
Ciencias de la Tierra y del Medio Ambiente, Universidad de Alicante, Ap. de correos, 99, 03080 Alicante (Spain)
Department of Geology, Moulay Ismail University, Meknes (Morocco)
Feldbergstrasse 5, 61191 Rosbach-Rodheim (Germany)
24 Church Street, P.O. Box 700, Gobabis (Namibia)
Ciencias de la Tierra y del Medio Ambiente, Universidad de Alicante, Ap. de correos, 99, 03080 Alicante (Spain)
2 rue Guy de Maupassant, F-76800 Saint �tienne du Rouvray (France)
Soci�t�
Axonaise de Pal�ontologie, 43 rue du ch�teau, F-02110 Bohain-en-Vermandois (France)
via Venezia n 42 - 61043 Cagli Pu (Italy)
TimeMatters Espa�a, Calle Costa Esmeralda 47, 03590 Altea (Spain)
Universit� Lyon, Univ. Lyon 1, ENSL, CNRS, LGL-TPE, F-69622, Villeurbanne (France)
OT/TL/G&R/GT/CARB,
CST-JF, Office: BA 2049, 18 Avenue Larribau, 64018 Pau Cedex (France)
Published online in final form (pdf) on December 25, 2024
DOI 10.2110/carnets.2025.2502
![]()
[Editor:
Michel Moullade; language editor: Phil Salvador;
technical editor: Bruno R.C. Granier]
![]()
Earlier misidentifications of ammonites from Busot (SE Spain) led to two incorrect hypotheses regarding calpionellids, which are today attributed to the Calpionellopsis Zone (D) of the upper Berriasian. The first hypothesis suggested that some calpionellid species survived into Hauterivian times, while the second hypothesis proposed that these calpionellids were reworked into Hauterivian strata. However, the revision of the ammonitic material collected in 1993-1994, along with the study of new material collected in 2024, suggests a late Berriasian age, as the ammonite assemblages likely correspond to the Fauriella boissieri and Tirnovella alpillensis Standard Ammonites zones. Nannofossil assemblages and the presence of Octahedronoides tethysianus further support this dating. This reassessment calls for a reexamination of claims regarding the occurrence of calpionellids in strata younger than the Valanginian in the literature.
� biostratigraphy;
� biozones;
� ammonites;
� calpionellids;
� nannofossils;
� Berriasian;
� Hauterivian
Granier B.R.C. (coordinator), Andreu Rodes J.M., Benzaggagh M., Blau J., B�ker �., Est�vez Rubio A., Ferr� B., Lambert B.C., Marini A., Monteil E., Reboulet S. & Virgone A. (2024).- Rectifying a substantial biostratigraphic error: There are no Hauterivian calpionellids in Busot (Alicante, Spain).- Carnets Geol., Madrid, vol. 25, no. 2, p. 29-72. DOI: 10.2110/carnets.2025.2502
Correction d'une erreur biostratigraphique majeure : Il n'y a pas de calpionelles hauteriviennes � Busot (Alicante, Espagne).- Les identifications ant�rieures d'ammonites provenant de Busot (sud-est de l'Espagne) �taient erron�es ; elles ont conduit � deux hypoth�ses incorrectes concernant des calpionelles qui sont aujourd'hui attribu�es � la Zone � Calpionellopsis (Zone D) du Berriasien sup�rieur. La premi�re hypoth�se sugg�rait que certaines esp�ces aient surv�cu jusqu'� l'Hauterivien, tandis que la seconde proposait que ces calpionelles aient �t� remani�es dans des strates hauteriviennes. Cependant, la r�vision des ammonites r�colt�es en 1993-1994 ainsi que l'�tude de nouveaux sp�cimens r�colt�s en 2024 sugg�rent un �ge Berriasien sup�rieur, car ces associations d'ammonites correspondraient aux zones standards � Fauriella boissieri et Tirnovella alpillensis. Les associations de nannofossiles et la pr�sence d'Octahedronoides tethysianus confirment �galement cette datation. Cette r��valuation appelle � une remise en question de la plupart des signalements de calpionelles dans des strates post�rieures au Valanginien mentionn�s dans la litt�rature.
� biostratigraphie ;
� biozones ;
� ammonites ;
� calpionelles ;
� nannofossiles ;
� Berriasien ;
� Hauterivien
As with any other component of the Tethys plankton (Granier, 2022), upon the death of the microorganisms, the calpionellid loricae sank from the upper layers of the water column down to the sea floor to contribute to pelagic and hemipelagic sedimentation. The specialists who initially examined the calpionellids from Busot (Alicante, SE Spain) attributed these microfossils to a Berriasian age. Then, upon discovering their association with supposedly Hauterivian ammonites, the same specialists promptly shifted their interpretation in favor of a reworking hypothesis. However, the abundance and relatively good preservation of the delicate calpionellid loricae found in the Busot material pleaded in favor of their post-mortem sinking as a part of the 'marine snow', rather than the downdip resedimentation hypothesis. Therefore, taking into consideration attested earlier records of late Valanginian to early Barremian calpionellids in the literature (see Granier et al., 1995, for bibliographic references), Granier et al. (1995) advocated for a third option, suggesting that the stratigraphic ranges of some calpionellid taxa may have been longer than previously thought. However, both the reworking option and the Granier's 1995 hypothesis will be demonstrated as erroneous in this publication because today the original identifications of the Busot ammonites have proved to be wrong. According to one of us (S.R.), these ammonites are of Berriasian age and not Hauterivian, as initially suggested by the late L.G. Bulot and the late R. Busnardo in Granier et al. (1995), an opinion still upheld by Bulot (personal communication to the first author dated September 4, 2020). Similarly, the assemblage of calpionellids from Busot will be shown in this publication to be late Berriasian in age.
The
village of Busot (Fig. 1.A ![]() ) is sited on a structural anomaly, a small tectonic
unit bordered by faults on all sides (Figs. 1.E
) is sited on a structural anomaly, a small tectonic
unit bordered by faults on all sides (Figs. 1.E ![]() ,
2
,
2 ![]() ). This spot offers a unique
exposure of uppermost Jurassic to lowermost Cretaceous strata contrasting with
surrounding strata that range from the Middle Cretaceous to the Eocene. From a
paleogeographic viewpoint, it was located on a slope in the transition zone from
1) the 'Jurassic' Prebetic carbonate platform, drowned and later dislocated in
Valanginian times, to 2) the Pre- and Subbetic basinal domain. In earlier
publications (Granier, 1987; Granier & Perthuisot,
2009), the sedimentary contrasts in
both thicknesses and facies between a) the thin, condensed, or missing upper
Valanginian to lower Barremian strata not only on the crests and slopes of
paleotopographic highs [i.e., in Serra Gelada, Puig Campana, Cabe��
d'Or (Granier, 1987)], and b) the slightly
thicker, but still condensed coeval strata in nearby starved basinal areas [e.g.,
Serra de Fontcalent (Rasplus & Fourcade,
1987)], were
interpreted as resulting from the Neocimmerian distention (Granier,
1987; Granier & Perthuisot,
2009: Figs. 2-5) that affected both the Prebetic and Subbetic domains.
). This spot offers a unique
exposure of uppermost Jurassic to lowermost Cretaceous strata contrasting with
surrounding strata that range from the Middle Cretaceous to the Eocene. From a
paleogeographic viewpoint, it was located on a slope in the transition zone from
1) the 'Jurassic' Prebetic carbonate platform, drowned and later dislocated in
Valanginian times, to 2) the Pre- and Subbetic basinal domain. In earlier
publications (Granier, 1987; Granier & Perthuisot,
2009), the sedimentary contrasts in
both thicknesses and facies between a) the thin, condensed, or missing upper
Valanginian to lower Barremian strata not only on the crests and slopes of
paleotopographic highs [i.e., in Serra Gelada, Puig Campana, Cabe��
d'Or (Granier, 1987)], and b) the slightly
thicker, but still condensed coeval strata in nearby starved basinal areas [e.g.,
Serra de Fontcalent (Rasplus & Fourcade,
1987)], were
interpreted as resulting from the Neocimmerian distention (Granier,
1987; Granier & Perthuisot,
2009: Figs. 2-5) that affected both the Prebetic and Subbetic domains.
|
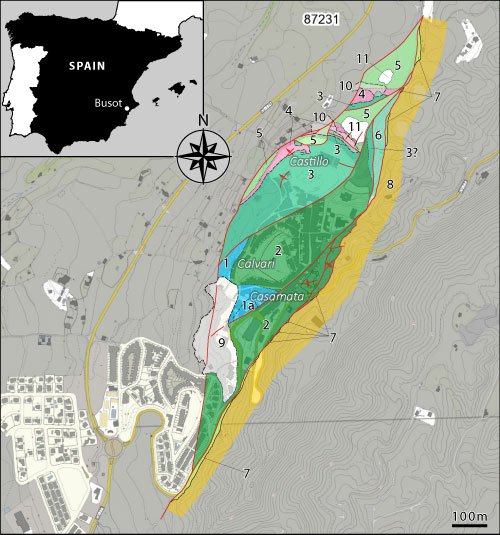
Figure 1: A)
general
view of Busot, with the Cabe�� d'Or in the background, and its three hills
from left to right: Calvari, Castell, and Casamata; B) Calvari section
viewed from the southwest; C) Casamata section viewed from the east; D)
photo of the late Robert Busnardo during the 2010 campaign; E) contact
between Berriasian strata (left) and Upper Cretaceous (Senonian) strata (right)
along the road to Aig�es, near the Casamata section. Fauriella latecostata
bundle arrowed in B and C. |
In
the geological map of Alicante at scale 1:50,000 (Leret Verd� & Lend�nez
Gonz�lez, 1978), the small tectonic unit comprising three hills: Calvari (Figs.
1.A-B ![]() ,
2
,
2 ![]() ), Casamata
(Figs. 1.A, 1.C
), Casamata
(Figs. 1.A, 1.C ![]() ,
2
,
2 ![]() ), and Castell
(Figs. 1.A
), and Castell
(Figs. 1.A ![]() ,
2
,
2 ![]() ), with the
latter located at the northern edge of the village, is labeled as C11-14,
i.e., "Neocomiense-Barremiense". Strata in the Castell hill dip
northwestward, whereas strata in the Calvari and Casamata hills form anticlines,
and the detailed geological map presented here clarifies the situation (Fig.
2
), with the
latter located at the northern edge of the village, is labeled as C11-14,
i.e., "Neocomiense-Barremiense". Strata in the Castell hill dip
northwestward, whereas strata in the Calvari and Casamata hills form anticlines,
and the detailed geological map presented here clarifies the situation (Fig.
2 ![]() ).
Considering our knowledge of the local stratigraphic succession with, from base
to top: 1) a thick calcarenitic unit corresponding to the lower Valanginian (forming
the core of the Castell hill), 2) a condensed section spanning the upper
Valanginian to Barremian and made of limestones, locally consisting of Barremian
ammonite-rich ferruginous oolites, and 3) Aptian ammonite marls, it was first
assumed that the Calvari section (Fig. 3
).
Considering our knowledge of the local stratigraphic succession with, from base
to top: 1) a thick calcarenitic unit corresponding to the lower Valanginian (forming
the core of the Castell hill), 2) a condensed section spanning the upper
Valanginian to Barremian and made of limestones, locally consisting of Barremian
ammonite-rich ferruginous oolites, and 3) Aptian ammonite marls, it was first
assumed that the Calvari section (Fig. 3 ![]() ) could expose strata older than those
of the Castell hill, i.e., older than the Valanginian. However, the find
of two supposedly Hauterivian ammonites, later identified by L.G. Bulot
and R.
Busnardo
as "Lyticoceras gr. nodosoplicatum (Kilian et Reboul)"
[UCBL-FSL 88816] (Granier et al., 1995: Fig. 4.5) and "Pseudothurmania grandis Busnardo" [UCBL-FSL
88817] (Granier et al., 1995: Fig.
4.7), altered our perspective regarding the calpionellid stratigraphic
distribution, among other related issues.
) could expose strata older than those
of the Castell hill, i.e., older than the Valanginian. However, the find
of two supposedly Hauterivian ammonites, later identified by L.G. Bulot
and R.
Busnardo
as "Lyticoceras gr. nodosoplicatum (Kilian et Reboul)"
[UCBL-FSL 88816] (Granier et al., 1995: Fig. 4.5) and "Pseudothurmania grandis Busnardo" [UCBL-FSL
88817] (Granier et al., 1995: Fig.
4.7), altered our perspective regarding the calpionellid stratigraphic
distribution, among other related issues.
|
Figure 2:
Geological map of Busot. Legend: |
On July 25,
1993, a section (Fig. 3 ![]() , referred to as Busot II section in Granier et al.,
1995) on a small hill in the village center,
the Mont del Calvari (Figs. 1.A-B
, referred to as Busot II section in Granier et al.,
1995) on a small hill in the village center,
the Mont del Calvari (Figs. 1.A-B ![]() ,
2
,
2 ![]() ), was measured by two junior geologists (�.B.
and A.V.), working under the supervision of the first author (B.R.C.G.).
), was measured by two junior geologists (�.B.
and A.V.), working under the supervision of the first author (B.R.C.G.).
|
Figure 3:
Mont del Calvari section referred to as Busot II section in Granier
et al., 1995. Red stars: nannofossil analyses. |
Two small hills
on the eastern side of the village (and on the western side of the road to Aig�es
/ Aguas de Busot) exhibit some similarities with the Calvari outcrop. The more
southern of these two hills is called Casamata. We measured section on Casamata
hill (Figs. 1.A,
1.C ![]() ,
2
,
2 ![]() , 4). There,
additional ammonites were collected including a "Ps. n. sp." fide
Busnardo [UCBL-FSL 88814], the specimen of which is unfortunately missing
in the Granier Collection.
, 4). There,
additional ammonites were collected including a "Ps. n. sp." fide
Busnardo [UCBL-FSL 88814], the specimen of which is unfortunately missing
in the Granier Collection.
|
Figure 4:
Casamata
section, referred to as Busot III section in Granier
et al., 1995. Red star: nannofossil analysis. |
On June 28,
1994, the same Casamata section (Fig. 4 ![]() , referred to as Busot III section in Granier
et al., 1995) was measured by one of the two junior geologists (A.V.)
under the supervision of the first author (B.R.C.G.). Thin sections were
prepared exclusively from this section. The discovery of abundant and
well-preserved calpionellids in association with supposedly late Hauterivian
ammonites suggested that these calpionellids survived the Valanginian, leading
to the publication of a preliminary paper the following year (Granier
et al., 1995). A subsequent paper (Ferr� & Granier,
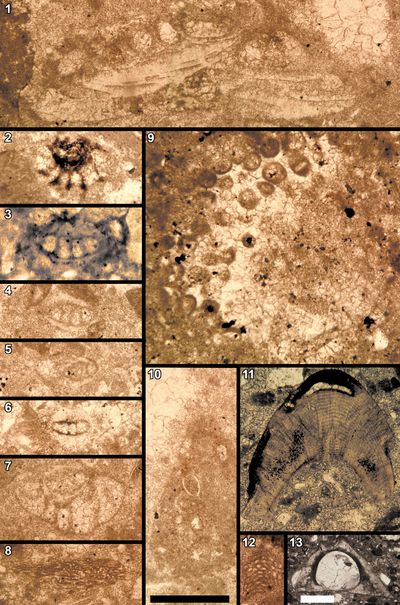
2000) included the description of a new Roveacrinus species (Pl. 13
, referred to as Busot III section in Granier
et al., 1995) was measured by one of the two junior geologists (A.V.)
under the supervision of the first author (B.R.C.G.). Thin sections were
prepared exclusively from this section. The discovery of abundant and
well-preserved calpionellids in association with supposedly late Hauterivian
ammonites suggested that these calpionellids survived the Valanginian, leading
to the publication of a preliminary paper the following year (Granier
et al., 1995). A subsequent paper (Ferr� & Granier,
2000) included the description of a new Roveacrinus species (Pl. 13 ![]() , fig.
1) from the Calvari section (Fig. 3
, fig.
1) from the Calvari section (Fig. 3 ![]() , referred to as Busot II section in Granier
et al., 1995), which was then assigned an early Hauterivian age. The scale bars of the logs represent 5 m, not 10 m as
erroneously indicated in both papers.
, referred to as Busot II section in Granier
et al., 1995), which was then assigned an early Hauterivian age. The scale bars of the logs represent 5 m, not 10 m as
erroneously indicated in both papers.
On September
24, 2010, two of us (B.R.C.G. and A.E.R.), along with the late R. Busnardo
(Fig. 1.D ![]() ) and the late B. Clavel, visited the eastern outcrops,
including the Casamata section (Fig. 4
) and the late B. Clavel, visited the eastern outcrops,
including the Casamata section (Fig. 4 ![]() , referred to as the Busot III section in Granier
et al., 1995), and the Calvari section (Fig. 3
, referred to as the Busot III section in Granier
et al., 1995), and the Calvari section (Fig. 3 ![]() , referred to as the Busot
II section in Granier et al., 1995).
New rock samples were collected, and thin sections were later prepared. However,
no new ammonite findings were made. Additionally, outcrops located along 'Calleja
Almazara' (38�28'56.3"N, 0�25'09.9"W) and at a bend of 'Carrer Sant
Josep' (38�28'54.7"N, 0�25'10.8"W, with a dip of N15�E, 25�W),
both in the southern part of the village (refer to the detailed map), were also
sampled. These outcrops are composed of 'Jurassic' limestone (undifferentiated
Tithonian-lower Berriasian).
, referred to as the Busot
II section in Granier et al., 1995).
New rock samples were collected, and thin sections were later prepared. However,
no new ammonite findings were made. Additionally, outcrops located along 'Calleja
Almazara' (38�28'56.3"N, 0�25'09.9"W) and at a bend of 'Carrer Sant
Josep' (38�28'54.7"N, 0�25'10.8"W, with a dip of N15�E, 25�W),
both in the southern part of the village (refer to the detailed map), were also
sampled. These outcrops are composed of 'Jurassic' limestone (undifferentiated
Tithonian-lower Berriasian).
Following the 11th International Symposium on the Cretaceous System (August 22-26, 2022), Roque Aguado Merlo and one of us (B.C.L.) were invited to examine the nannofossil contents of Busot hemipelagites. The first (R.A.M.) did not identify any age-diagnostic species and provided an inconclusive report whereas the second (B.C.L.) focusing on Nannoconus spp. and some other nannofossils attributed the samples studied to the Berriasian (most likely zones NK-1 and NK-2 of Bralower et al., 1989), definitely excluding an Hauterivian age.
On April 20-22
and 24, 2024, two of us (B.R.C.G. and E.M.) visited the eastern outcrops (Fig.
4 ![]() , Busot III section in Granier et al.,
1995) and the Calvari section (Fig. 3
, Busot III section in Granier et al.,
1995) and the Calvari section (Fig. 3 ![]() , Busot II section in Granier et al.,
1995), as well as the 'Calleja Almazara' and
the 'Carrer Sant Josep' outcrops. The sixth author (A.E.R.) accompanied them on
April 24, 2024. New rock samples and a few new ammonites were collected from
both sections.
, Busot II section in Granier et al.,
1995), as well as the 'Calleja Almazara' and
the 'Carrer Sant Josep' outcrops. The sixth author (A.E.R.) accompanied them on
April 24, 2024. New rock samples and a few new ammonites were collected from
both sections.
Finally, the
first author (B.R.C.G.) introduced Octahedronoides tethysianus Granier,
2024b, to identify fossil coenobia consisting of clusters of leiospheres (Pl. 9 ![]() ,
fig. 26; Pl. 11
,
fig. 26; Pl. 11 ![]() , figs. 22-25), formerly known as Cadosina minuta Borza,
1980, which are very common in the sections studied.
, figs. 22-25), formerly known as Cadosina minuta Borza,
1980, which are very common in the sections studied.
Ammonites appear to be rare in the Calvari and Casamata sections. Only a few specimens were collected by three of us (�.B., A.V., and the first author, B.R.C.G.) during the 1993 and 1994 campaigns. These specimens were initially identified by the late L.G. Bulot and the late R. Busnardo (see Granier et al., 1995). However, in the current study, these ammonites were re-studied by one of us (S.R.). The collection has been recently supplemented by five additional specimens, collected by two of us (B.R.C.G. and E.M.) in spring 2024. In the ammonite descriptions below, the year of sampling is indicated in brackets next to the collection number.
The standard zonation defined by the ICS (International Commission on Stratigraphy) Kilian Group is applied here (cf. Reboulet et al., 2018; Szives et al., 2024; see also Kenjo et al., 2021, for discussion on the zonal scheme on the interval around the Berriasian/Valanginian boundary). Standard zones and subzones are referred to as StZ and StSz, respectively. Zones and subzones used by other authors cited in this work are written in quotation marks as they appear in their original works, to avoid confusion between previous zonations and the Standard Zonation. Except for three specimens that retain the body chamber and a substantial portion of the phragmocone, most of the ammonites collected are relatively poorly preserved, exhibiting fragmentation and the loss of inner whorls. Some of these specimens were identified only at the family level. Specimens that are doubtfully identified at the generic or specific level are marked with a question mark. For certain individuals, measurements were taken at the maximum preserved shell size and indicated as follows: D = diameter of the shell; U = width of the umbilicus; H = height of the whorl; W = width of the whorl (the ammonites are not crushed). The ratios of U/D and H/D are also provided.
Due to the nature of the preservation, observing the suture lines is difficult. Shell dissolution is common, and the ammonites are preserved as internal calcareous molds. The studied and illustrated material is housed and cataloged as "Collection Granier" with the label UCBL-FSL, which stands for "Universit� Claude Bernard Lyon 1 - Facult� des Sciences de Lyon" (France).
The ammonite findings referenced in the text are associated with UCBL-FSL numbers, corresponding to specimens housed in the 'Collections de l'Universit� Claude Bernard Lyon 1'. Part of this material was initially pre-registered by R. Busnardo as UCBL-FSL 88813-88822. However, Busnardo retained the specimens at his home, and following his passing on March 24, 2018, it appears that specimen UCBL-FSL 88814 has likely been lost. In addition to the original collection, five new ammonite specimens were gathered in 2024, with four now registered under UCBL-FSL numbers 88892 to 88895.
The total set of petrographic thin sections studied consists of
47 thin sections, including 2 prepared from the matrix of ammonites collected in
1993, 26 from material collected in 1994, 11 from material collected in 2010,
and 8 from material collected in 2024 (see Appendix). All
these thin sections are now registered in the collections of the 'Mus�e
d'Histoire naturelle de Gen�ve' under the numbers MHNG-GEPI-2024-10031 to 10057
and MHNG-GEPI-2024-10060 to 10079. A few more thin sections, belonging to the
collection of the sixth author (A.E.R.), Universidad de Alicante, are also used
in this study: no. 6521-CALV-1 (Pl. 11 ![]() , fig. 24;
Pl. 15
, fig. 24;
Pl. 15 ![]() , figs. 6, 9), GPS coordinates:
38�28'59.61"N,
0�25'8.98"W; no. 6521-BAR-1 (Pl. 15
, figs. 6, 9), GPS coordinates:
38�28'59.61"N,
0�25'8.98"W; no. 6521-BAR-1 (Pl. 15 ![]() , figs. 1-5, 8), GPS coordinates:
38�28'54.7"N, 0�25'11.0"W. These
thin sections were used for identification of calpionellids, cadosinids, and
other calcareous dinoflagellate cysts, as well as benthic foraminifers and
calcareous algae; they were also used for identification of other allochems and
the microfacies. Photomicrographs of the petrographic thin sections were taken using a
digital MU900 AmScope camera on a Leiz Diaplan microscope. Regarding the calpionellid
zonations, although there are some differences among the schemes proposed by Remane
(1970), Gr�n
and Blau (1997), and Benzaggagh
(2020), all concur on the extent of Zone D, also known as the Calpionellopsis
Zone, as well as on its subdivisions.
, figs. 1-5, 8), GPS coordinates:
38�28'54.7"N, 0�25'11.0"W. These
thin sections were used for identification of calpionellids, cadosinids, and
other calcareous dinoflagellate cysts, as well as benthic foraminifers and
calcareous algae; they were also used for identification of other allochems and
the microfacies. Photomicrographs of the petrographic thin sections were taken using a
digital MU900 AmScope camera on a Leiz Diaplan microscope. Regarding the calpionellid
zonations, although there are some differences among the schemes proposed by Remane
(1970), Gr�n
and Blau (1997), and Benzaggagh
(2020), all concur on the extent of Zone D, also known as the Calpionellopsis
Zone, as well as on its subdivisions.
Calcareous nannofossils were studied using smear slides without any special processing, i.e., a small piece of sediment was crushed in a drop of distilled water, dried, and then sealed with Canada balsam. The analysis of the nannofossils encompasses only three samples (Busot-1994-03 = CLV_3; Busot-1994-05 = CLV_5; Busot-1994-15 = CLV_15). Preservation is moderate, and the concentration is notably low, requiring the preparation of multiple slides to secure a minimum count of 100 specimens. The nannofossil zonation used in this work follows Bralower et al., 1989. For further comparison with other existing zonations, refer to the 'Nannofossils' chapter in the 2021 contribution by Kenjo et al.
Three large ammonites are identified as Fauriella latecostata (Mazenot, 1939), and they are briefly described here. Their measurements are given in Table 1. According to Le H�garat (1973: p. 161), a thin section was made from the rock containing the holotype (from Berrias, UCBL-FSL 13398; figured in Mazenot, 1939: Pl. 17, fig. 1). The analysis allowed this sample to be dated to an interval covering the transition from D2 to D3 of the calpionellid subzones. For Le H�garat (1973: Table 10, p. 148), the range of F. latecostata spans the "Callisto" ammonite Subzone, which approximatively corresponds to the current Tirnovella alpillensis Standard Subzone (StSz).
|
Figure 5:
Correlation
of Calvari and Casamata sections. Red stars: nannofossil analyses. |
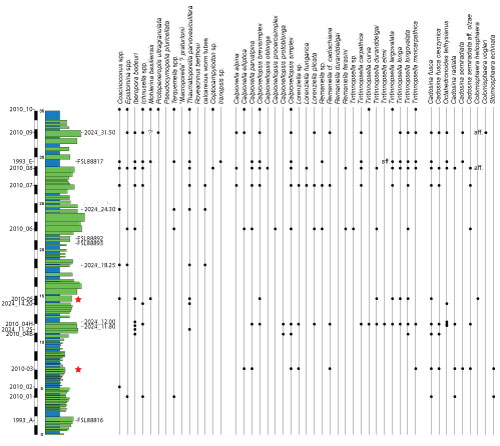
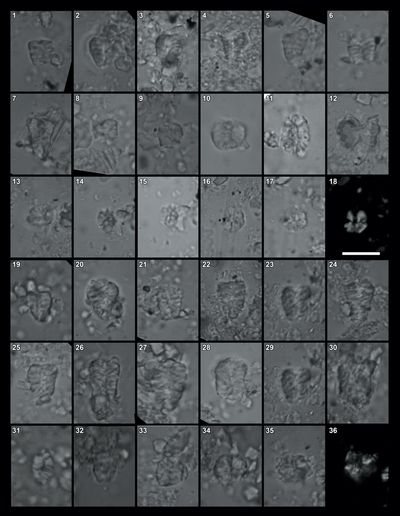
The
best-preserved specimen is the UCBL-FSL 88892 (2024; Pl. 1 ![]() ), located at 21.15
meters in the Calvari section (Fig. 5
), located at 21.15
meters in the Calvari section (Fig. 5 ![]() , formerly
the Busot II section). Compared to the holotype, the ribbing of the phragmocone
is thicker and less dense. In the poorly preserved early inner whorls, most of
the radial ribs appear to originate as simple ribs from the umbilicus. In the
later inner whorls, the ribs are curved, slightly inclined toward the peristome,
and fasciculate (in pairs) from small bullate umbilical tubercules. Toward the
end of the phragmocone, the ribs and their tubercules progressively become
coarser, and intermediate ribs occasionally occur. It seems that all ribs are
interrupted on the venter. In these terms, the ornamentation of the phragmocone
of the Spanish specimen is closer to that of the Moroccan ammonite figured by Wippich
(2001: Pl. 14, fig. 1), dated to his "Fauriella boissieri Zone", "Tirnovella
alpillensis Subzone", which likely corresponds to the Tirnovella
alpillensis Standard Zone (StZ). For the three specimens mentioned above,
the ribbing on the body chamber is similar. Umbilical tubercles become stronger
and more widely spaced, with generally one rib originating from each tubercle.
These main ribs are coarser on the lower part of the flank, with one
intermediate rib arising low from the basal part of the flank between each pair
of main ribs, sometimes merging with the main rib. All ribs are radial to weakly
prosiradiate, rigid to slightly flexuous, and most bifurcate in the upper third
part of the flank, continuing across the venter.
, formerly
the Busot II section). Compared to the holotype, the ribbing of the phragmocone
is thicker and less dense. In the poorly preserved early inner whorls, most of
the radial ribs appear to originate as simple ribs from the umbilicus. In the
later inner whorls, the ribs are curved, slightly inclined toward the peristome,
and fasciculate (in pairs) from small bullate umbilical tubercules. Toward the
end of the phragmocone, the ribs and their tubercules progressively become
coarser, and intermediate ribs occasionally occur. It seems that all ribs are
interrupted on the venter. In these terms, the ornamentation of the phragmocone
of the Spanish specimen is closer to that of the Moroccan ammonite figured by Wippich
(2001: Pl. 14, fig. 1), dated to his "Fauriella boissieri Zone", "Tirnovella
alpillensis Subzone", which likely corresponds to the Tirnovella
alpillensis Standard Zone (StZ). For the three specimens mentioned above,
the ribbing on the body chamber is similar. Umbilical tubercles become stronger
and more widely spaced, with generally one rib originating from each tubercle.
These main ribs are coarser on the lower part of the flank, with one
intermediate rib arising low from the basal part of the flank between each pair
of main ribs, sometimes merging with the main rib. All ribs are radial to weakly
prosiradiate, rigid to slightly flexuous, and most bifurcate in the upper third
part of the flank, continuing across the venter.
The
ammonite UCBL-FSL 88813 (1994; Pl. 2 ![]() ), found at 10.90 meters in the median part
of the Casamata section (Fig. 5
), found at 10.90 meters in the median part
of the Casamata section (Fig. 5 ![]() , formerly the Busot III section),
was initially identified as Pseudothurmannia grandis (within the
framework of Granier et al., 1995). However, this identification can be
excluded, as the ornamentation of the body chamber is similar to that of the
specimen UCBL-FSL 88892. Additionally, the ribs only cross the venter on the
second part of the body chamber, not from the end part of the phragmocone, as
would be expected for Pseudothurmannia.
, formerly the Busot III section),
was initially identified as Pseudothurmannia grandis (within the
framework of Granier et al., 1995). However, this identification can be
excluded, as the ornamentation of the body chamber is similar to that of the
specimen UCBL-FSL 88892. Additionally, the ribs only cross the venter on the
second part of the body chamber, not from the end part of the phragmocone, as
would be expected for Pseudothurmannia.
The
specimen UCBL-FSL 88815 (1994; Pl. 3 ![]() ), sampled at 12.75 meters in the Casamata
section (Fig. 5
), sampled at 12.75 meters in the Casamata
section (Fig. 5 ![]() , formerly the Busot III section), is less well-preserved and
has been tentatively identified as Fauriella latecostata?. Its
ribbing resembles that observed on the previous Spanish specimens, though a few
constrictions occur. It was first identified as Lyticoceras sp. and later
as Pseudothurmannia pseudomalbosi (within the framework of Granier et al.,
1995).
, formerly the Busot III section), is less well-preserved and
has been tentatively identified as Fauriella latecostata?. Its
ribbing resembles that observed on the previous Spanish specimens, though a few
constrictions occur. It was first identified as Lyticoceras sp. and later
as Pseudothurmannia pseudomalbosi (within the framework of Granier et al.,
1995).
Table 1: Measurements (D, U, H, W in mm) of three Fauriella latecostata specimens.
| UCBL-FSL | D | U | H | W | U/D | H/D |
| 88892 | 191 | 80 | 62 | 35 | 0.42 | 0.32 |
| 88813 | 217 | 92 | 73 | 48 | 0.42 | 0.37 |
| 88815 | 209 | 86 | 68 | 40 | 0.41 | 0.33 |
The
ammonite UCBL-FSL 88817 (1993; Pl. 4.A ![]() ), sampled at 29.35 meters in the Calvari
section (Fig. 5
), sampled at 29.35 meters in the Calvari
section (Fig. 5 ![]() , formerly
the Busot II section), was originally identified as Pseudothurmannia gr. grandis
(Granier et al., 1995: Fig. 4.7). However, the ornamentation does not match
that of the genus. This fragment is mostly represented by the body chamber. The
ribs appear to be isolated at the umbilicus edge, bifurcating approximatively
two-thirds up the whorl height. In the lower half of the flank, the ribs are
thicker, and on the last part of the body chamber, they cross the venter. Based
on this description, the specimen is tentatively identified as "Thurmanniceras"
cf. gratianopolitense (Sayn, 1907).
, formerly
the Busot II section), was originally identified as Pseudothurmannia gr. grandis
(Granier et al., 1995: Fig. 4.7). However, the ornamentation does not match
that of the genus. This fragment is mostly represented by the body chamber. The
ribs appear to be isolated at the umbilicus edge, bifurcating approximatively
two-thirds up the whorl height. In the lower half of the flank, the ribs are
thicker, and on the last part of the body chamber, they cross the venter. Based
on this description, the specimen is tentatively identified as "Thurmanniceras"
cf. gratianopolitense (Sayn, 1907).
The systematics and stratigraphic distribution of this species are thoroughly discussed by Kenjo (2014), Kenjo et al. (2021), and other references therein. In Company and Tavera (2015), and Reboulet et al. (2022), the range of "Thurmanniceras" gratianopolitense is restricted at the lower part of the "Thurmanniceras" pertransiens StZ (lower Valanginian). However, according to the discussions by Kenjo (2014), Kenjo et al. (2021), and Reboulet et al. (2022), its range could begin in the upper part of the Tirnovella alpillensis StZ (see also Wippich, 2001, 2003; Ettachfini, 2004).
The
specimen UCBL-FSL 88893 (2024; Pl. 4.B ![]() ), collected at 20.70 meters in the Calvari
section (Fig. 5
), collected at 20.70 meters in the Calvari
section (Fig. 5 ![]() , formerly
the Busot II section), is attributed to the genus Tirnovella. The ribs
are thin, dense, and nearly straight, slanting towards the peristome. Although
the preservation of the umbilicus edge is poor, the ribs appear fasciculate (grouped
by twos; as seen on the outer mold cast) from weakly bullate umbilical tubercles.
Some ribs bifurcate around mid-flank. The ribs seem interrupted on the venter,
which is crushed. Typical representatives of Tirnovella show more
flexuous ribbing with more frequent bifurcations on the flank.
, formerly
the Busot II section), is attributed to the genus Tirnovella. The ribs
are thin, dense, and nearly straight, slanting towards the peristome. Although
the preservation of the umbilicus edge is poor, the ribs appear fasciculate (grouped
by twos; as seen on the outer mold cast) from weakly bullate umbilical tubercles.
Some ribs bifurcate around mid-flank. The ribs seem interrupted on the venter,
which is crushed. Typical representatives of Tirnovella show more
flexuous ribbing with more frequent bifurcations on the flank.
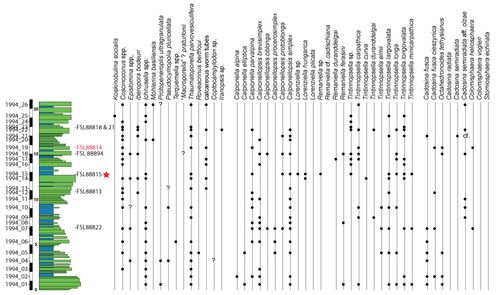
The
specimen UCBL-FSL 88818 (1994; Pl. 5.A ![]() ), found at 18.00 meters in the Casamata
section (Fig. 5
), found at 18.00 meters in the Casamata
section (Fig. 5 ![]() , formerly the Busot III section), was initially attributed to Pseudothurmannia
sp. ind. (within the framework of Granier et al.,
1995). This
fragment, a part of the body chamber, is here identified with uncertainty as Erdenella
cf. paquieri (Simionescu,
1899). The umbilicus appears wide. There are single, thick, relatively straight
and widely spaced ribs, with weak umbilical and lateral tubercles. Between these
main ribs, weaker ribs occur (without tubercules), but they tend to disappear in
the lower part of the flank. The ribs bifurcate in the upper third of the whorl
height. On the venter, the ribs are initially interrupted but then eventually
seem to cross it. This ornamentation matches the pattern observed at the end of
the shell figured by Le H�garat
(1973: Pl. 50, fig. 1; specimen from his "Callisto Subzone" of the
Ginestous-la-Garenne section, H�rault, France). In his monography, Le H�garat
(1973: p. 200-201) noted the significant changes in ribbing observed on the body
chamber of large specimens: "In these specimens, which reach a size of
180-220 mm, the rib bundles tend to separate towards the end of the last whorl.
Gradually, the ornamentation is composed only of large ribs, bifurcated or not,
which emerge individually from the umbilicus. The ribs that were part of the
bundles still exhibit umbilical and lateral thickenings" [translated from
the French: "Chez ceux-ci, dont la taille atteint 180-220 mm, les faisceaux
tendent � se dissocier � la fin du dernier tour; progressivement
l'ornementation ne se trouve plus form�e que de grosses c�tes, bifurqu�es ou
non, qui partent isol�ment de l'ombilic. Celles qui participaient aux faisceaux
portent encore des �paississements ombilicaux et lat�raux"]. E.
paquieri is recorded in the "Fauriella boissieri Zone",
specifically in the "Picteti" and "Callisto" subzones (Le H�garat,
1973: Table 13, p 191). Wippich
(2001) and Ettachfini (2004) also documented
the occurrence of Erdenella paquieri in their "Fauriella boissieri
Zone". Additionally, Company (1987), Bulot
(1995), and Aguado et al. (2000) noted its presence in
their "Otopeta Zone" (currently the "Thurmanniceras" otopeta
StSz). This observation was later confirmed by Kenjo
(2014) and Kenjo et al. (2021), who identified Erdenella paquieri in both
subzones of the Tirnovella alpillensis StZ.
, formerly the Busot III section), was initially attributed to Pseudothurmannia
sp. ind. (within the framework of Granier et al.,
1995). This
fragment, a part of the body chamber, is here identified with uncertainty as Erdenella
cf. paquieri (Simionescu,
1899). The umbilicus appears wide. There are single, thick, relatively straight
and widely spaced ribs, with weak umbilical and lateral tubercles. Between these
main ribs, weaker ribs occur (without tubercules), but they tend to disappear in
the lower part of the flank. The ribs bifurcate in the upper third of the whorl
height. On the venter, the ribs are initially interrupted but then eventually
seem to cross it. This ornamentation matches the pattern observed at the end of
the shell figured by Le H�garat
(1973: Pl. 50, fig. 1; specimen from his "Callisto Subzone" of the
Ginestous-la-Garenne section, H�rault, France). In his monography, Le H�garat
(1973: p. 200-201) noted the significant changes in ribbing observed on the body
chamber of large specimens: "In these specimens, which reach a size of
180-220 mm, the rib bundles tend to separate towards the end of the last whorl.
Gradually, the ornamentation is composed only of large ribs, bifurcated or not,
which emerge individually from the umbilicus. The ribs that were part of the
bundles still exhibit umbilical and lateral thickenings" [translated from
the French: "Chez ceux-ci, dont la taille atteint 180-220 mm, les faisceaux
tendent � se dissocier � la fin du dernier tour; progressivement
l'ornementation ne se trouve plus form�e que de grosses c�tes, bifurqu�es ou
non, qui partent isol�ment de l'ombilic. Celles qui participaient aux faisceaux
portent encore des �paississements ombilicaux et lat�raux"]. E.
paquieri is recorded in the "Fauriella boissieri Zone",
specifically in the "Picteti" and "Callisto" subzones (Le H�garat,
1973: Table 13, p 191). Wippich
(2001) and Ettachfini (2004) also documented
the occurrence of Erdenella paquieri in their "Fauriella boissieri
Zone". Additionally, Company (1987), Bulot
(1995), and Aguado et al. (2000) noted its presence in
their "Otopeta Zone" (currently the "Thurmanniceras" otopeta
StSz). This observation was later confirmed by Kenjo
(2014) and Kenjo et al. (2021), who identified Erdenella paquieri in both
subzones of the Tirnovella alpillensis StZ.
The
ammonite UCBL-FSL 88821 (1994; Pl. 5.B ![]() ), collected at 18.00 meters in the
Casamata section (Fig. 5
), collected at 18.00 meters in the
Casamata section (Fig. 5 ![]() , formerly the Busot III section), was initially identified as Abrytusites
cf. neumayri (within the framework of Granier et al.,
1995).
However, it somewhat resembles Luppovella, a genus from the lower
Valanginian, particularly its species Luppovella superba (Sayn,
1907), which is only known from the middle part of the substage (see details in Kenjo
et al., 2021). Despite some
similarities, the comparison with Luppovella is weak and unsatisfying.
The specimen has a very large umbilicus and a subquadratic section. The
ornamentation is strong and rigid, with two ribs arising from the umbilical
tubercules. One of these ribs may bifurcate in the upper part of the flank,
featuring a large lateral tubercle. Although the ribbing is coarser, this
description can be compared to that provided by Bulot
(1995: p. 163) for Fuhriella, specifically during the "Erdenella
stage". On the Spanish specimen, only a single rib is visible due to the poor
preservation of the venter. It is not possible to determine whether the ribs
cross the venter continuously or with interruption. According to Bulot
(1995) and Bulot and Thieuloy
(1995),
representatives of Fuhriella are recorded in the lower Valanginian, from
the upper part of "Thurmanniceras" pertransiens StZ to the
upper part of Karakaschiceras inostranzewi StZ (with further details
provided by Ettachfini, 2004). Wippich
(2001) noted the presence of Fuhriella sp. in this
substage (lower part of "Thurmanniceras" pertransiens StZ),
but also in the Berriasian, in the upper part of his "Fauriella boissieri
Zone", which corresponds to the Tirnovella alpillensis StZ.
Unfortunately, the inner whorls on the large specimen figured by this author (Wippich,
2001: Pl. 31, figs. 1-2) are not preserved, making it impossible to directly
compare it with our small specimen. Wippich
(2003) places the name of the genus in quotation marks, suggesting uncertainty.
Thus, based on this discussion, the ammonite UCBL-FSL 88821 is not attributed to
Fuhriella (a genus restricted to the middle part of the lower
Valanginian; see Bulot, 1995) but is tentatively identified as Erdenella?
sp. (cf. the 'Erdenella ornamental stage' mentioned previously).
, formerly the Busot III section), was initially identified as Abrytusites
cf. neumayri (within the framework of Granier et al.,
1995).
However, it somewhat resembles Luppovella, a genus from the lower
Valanginian, particularly its species Luppovella superba (Sayn,
1907), which is only known from the middle part of the substage (see details in Kenjo
et al., 2021). Despite some
similarities, the comparison with Luppovella is weak and unsatisfying.
The specimen has a very large umbilicus and a subquadratic section. The
ornamentation is strong and rigid, with two ribs arising from the umbilical
tubercules. One of these ribs may bifurcate in the upper part of the flank,
featuring a large lateral tubercle. Although the ribbing is coarser, this
description can be compared to that provided by Bulot
(1995: p. 163) for Fuhriella, specifically during the "Erdenella
stage". On the Spanish specimen, only a single rib is visible due to the poor
preservation of the venter. It is not possible to determine whether the ribs
cross the venter continuously or with interruption. According to Bulot
(1995) and Bulot and Thieuloy
(1995),
representatives of Fuhriella are recorded in the lower Valanginian, from
the upper part of "Thurmanniceras" pertransiens StZ to the
upper part of Karakaschiceras inostranzewi StZ (with further details
provided by Ettachfini, 2004). Wippich
(2001) noted the presence of Fuhriella sp. in this
substage (lower part of "Thurmanniceras" pertransiens StZ),
but also in the Berriasian, in the upper part of his "Fauriella boissieri
Zone", which corresponds to the Tirnovella alpillensis StZ.
Unfortunately, the inner whorls on the large specimen figured by this author (Wippich,
2001: Pl. 31, figs. 1-2) are not preserved, making it impossible to directly
compare it with our small specimen. Wippich
(2003) places the name of the genus in quotation marks, suggesting uncertainty.
Thus, based on this discussion, the ammonite UCBL-FSL 88821 is not attributed to
Fuhriella (a genus restricted to the middle part of the lower
Valanginian; see Bulot, 1995) but is tentatively identified as Erdenella?
sp. (cf. the 'Erdenella ornamental stage' mentioned previously).
The
specimen UCBL-FSL 88816 (1993; Pl. 6 ![]() ), found at 1.40 meters, near the base of
the Calvari section (Fig. 5
), found at 1.40 meters, near the base of
the Calvari section (Fig. 5 ![]() , formerly
the Busot II section), was initially identified as Lyticoceras gr. nodosoplicatum
(Granier et al., 1995: Fig. 4.5). However, the ornamentation of the Spanish specimen,
which is mainly represented by the body chamber, does not match that of this
Hauterivian species. Several features highlight this discrepancy. The umbilicus
is very large (Table 2), and the whorl section is ogival-shaped with a rounded
venter. The ornamentation consists of relatively thick, radially elongated
umbilical tubercles, while the flank is smooth. In the first part of the body
chamber, some residual ribs crossing the venter can be faintly observed. This
specimen could be identified as Spiticeras ducale (Matheron, 1889). In the
body chamber of the specimen figured by Djan�lidz�
(1922: Pl. 17, fig.
1), the strength of the ribs tends to weaken around the mid-flank. A comparison
can also be made with a large specimen illustrated by Wippich
(2001: Pl. 7, fig. 1), collected in the Tirnovella
occitanica Zone. The ribbing of the last whorl in that specimen is only
represented by umbilical tubercles, and the flank appears smooth. Wippich
(2001) indicated the presence of Spiticeras ducale in the lower part of
his "Fauriella boissieri Zone" (a stratigraphic interval that should
correspond to the Fauriella boissieri StZ, and with some uncertainty in
his "Tirnovella occitanica Zone"). According to Le H�garat
(1973), the range of Spiticeras aff. ducale is restricted to his "Fauriella boissieri Zone", specifically within the
"Picteti"
Subzone, which roughly corresponds to the upper part of the Fauriella
boissieri StZ.
, formerly
the Busot II section), was initially identified as Lyticoceras gr. nodosoplicatum
(Granier et al., 1995: Fig. 4.5). However, the ornamentation of the Spanish specimen,
which is mainly represented by the body chamber, does not match that of this
Hauterivian species. Several features highlight this discrepancy. The umbilicus
is very large (Table 2), and the whorl section is ogival-shaped with a rounded
venter. The ornamentation consists of relatively thick, radially elongated
umbilical tubercles, while the flank is smooth. In the first part of the body
chamber, some residual ribs crossing the venter can be faintly observed. This
specimen could be identified as Spiticeras ducale (Matheron, 1889). In the
body chamber of the specimen figured by Djan�lidz�
(1922: Pl. 17, fig.
1), the strength of the ribs tends to weaken around the mid-flank. A comparison
can also be made with a large specimen illustrated by Wippich
(2001: Pl. 7, fig. 1), collected in the Tirnovella
occitanica Zone. The ribbing of the last whorl in that specimen is only
represented by umbilical tubercles, and the flank appears smooth. Wippich
(2001) indicated the presence of Spiticeras ducale in the lower part of
his "Fauriella boissieri Zone" (a stratigraphic interval that should
correspond to the Fauriella boissieri StZ, and with some uncertainty in
his "Tirnovella occitanica Zone"). According to Le H�garat
(1973), the range of Spiticeras aff. ducale is restricted to his "Fauriella boissieri Zone", specifically within the
"Picteti"
Subzone, which roughly corresponds to the upper part of the Fauriella
boissieri StZ.
Table 2: Measurements (D, U, H, W in mm) of the Spiticeras ducale specimen.
| UCBL-FSL | D | U | H | W | U/D | H/D |
| 88816 | 193 | 90 | 55 | 37 | 0.47 | 0.28 |
For
the following specimens, identifications are made at the family level: one
Olcostephanid ind. for UCBL-FSL 88822 (1994, from the Casamata section, formerly
the Busot III at 7.05 meters: Fig. 5 ![]() ),
previously identified as "Olcostephanus juv. gr. astieri", and
two Neocomitids ind. for UCBL-FSL 88894 (2024; from Casamata section, formerly
the Busot III at 15.10 meters: Fig. 5
),
previously identified as "Olcostephanus juv. gr. astieri", and
two Neocomitids ind. for UCBL-FSL 88894 (2024; from Casamata section, formerly
the Busot III at 15.10 meters: Fig. 5 ![]() ) and UCBL-FSL 88895 (2024; from a short
section near the Casamata section at GPS coordinates 38�28'59.6"N, 0�25'00.0"W).
) and UCBL-FSL 88895 (2024; from a short
section near the Casamata section at GPS coordinates 38�28'59.6"N, 0�25'00.0"W).
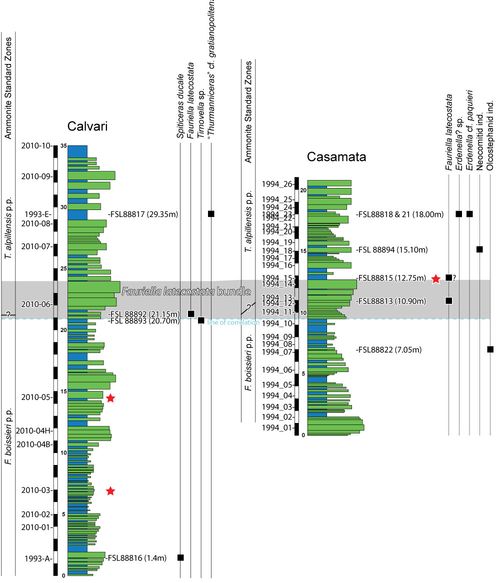
Contrary
to the late L.G. Bulot and the late R. Busnardo
(in Granier et al., 1995), who suggested a Hauterivian age for the Busot
sections (from the Lyticoceras nodosoplicatum Zone to the Pseudothurmannia
ohmi Zone), the revision of the ammonite fauna made by one of us (S.R.) in
the present work has allowed us to date the studied interval to the late
Berriasian. Due to the presence of Spiticeras ducale, the basal (or
possibly even lower) part of the Calvari section can be dated to the Fauriella
boissieri StZ (Fig. 5 ![]() ). Unfortunately, no other ammonites were found in the
lower part of the section. However, the upper parts of the Calvari and Casamata
sections could be dated to the Tirnovella alpillensis StZ, based on the
presence of Fauriella latecostata. At present, in terms of ammonites, the
First Occurrence (FO) of this species could serve to mark the base of the Tirnovella
alpillensis StZ. The uppermost part of the Calvari section may correspond to
the upper part of this zone due to the occurrence of "Thurmanniceras" cf. gratianopolitense. In the Casamata section, the base of the Tirnovella
alpillensis StZ can be placed at the base of the bed where the FO of Fauriella
latecostata is recorded (10.90 m). However, based on correlations with
the Calvari section, the basal boundary of this zone (at 10.80 m) could be
slightly lowered at Casamata (at 9.50 m), considering that Fauriella
latecostata is observed in the basal part of the calcareous bundle (referred
to here as the F. latecostata bundle: Fig. 5
). Unfortunately, no other ammonites were found in the
lower part of the section. However, the upper parts of the Calvari and Casamata
sections could be dated to the Tirnovella alpillensis StZ, based on the
presence of Fauriella latecostata. At present, in terms of ammonites, the
First Occurrence (FO) of this species could serve to mark the base of the Tirnovella
alpillensis StZ. The uppermost part of the Calvari section may correspond to
the upper part of this zone due to the occurrence of "Thurmanniceras" cf. gratianopolitense. In the Casamata section, the base of the Tirnovella
alpillensis StZ can be placed at the base of the bed where the FO of Fauriella
latecostata is recorded (10.90 m). However, based on correlations with
the Calvari section, the basal boundary of this zone (at 10.80 m) could be
slightly lowered at Casamata (at 9.50 m), considering that Fauriella
latecostata is observed in the basal part of the calcareous bundle (referred
to here as the F. latecostata bundle: Fig. 5 ![]() ).
).
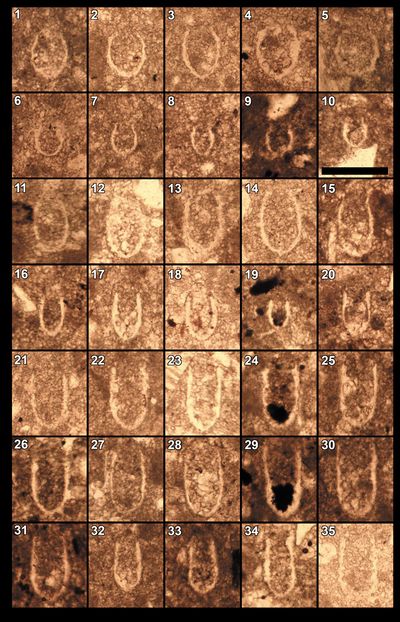
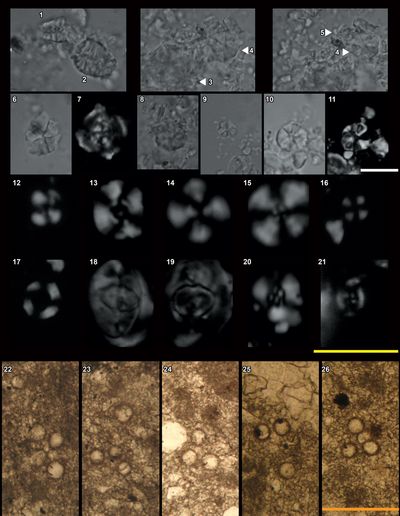
The specific assemblage of calpionellids from Busot, comprising Calpionellopsis gr. simplex/C. gr. oblonga, is characteristic of a narrow time interval. More specifically, it corresponds to the subzones D1-D2, also referred to as the Simplex-Oblonga subzones of the Calpionellopsis Zone (Zone D).
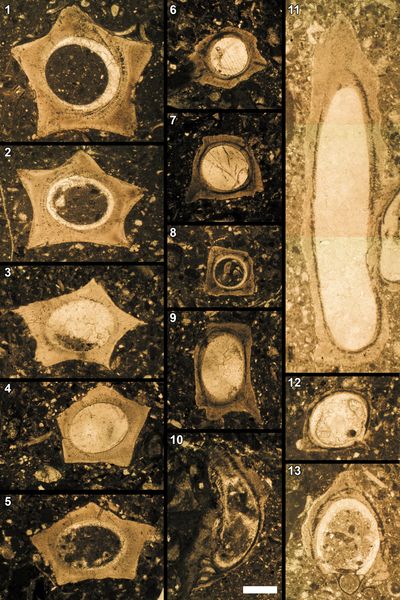
The
D1 (Simplex) subzone contains Calpionellopsis brevisimplex Nagy,
1986 (Pl. 7 ![]() , figs. 16-20), C. simplex (Colom, 1939)
(Pl. 7
, figs. 16-20), C. simplex (Colom, 1939)
(Pl. 7 ![]() , figs.
21-26), C. procerosimplex Nagy, 1986 (Pl. 7
, figs.
21-26), C. procerosimplex Nagy, 1986 (Pl. 7 ![]() , figs. 27-30), Calpionella
grandalpina Nagy,
1986 (Pl. 7
, figs. 27-30), Calpionella
grandalpina Nagy,
1986 (Pl. 7 ![]() , figs. 1-5), and C. elliptica Cadisch, 1932
(Pl. 7
, figs. 1-5), and C. elliptica Cadisch, 1932
(Pl. 7 ![]() ,
figs. 11-15).
In the Calvari section (Fig. 3
,
figs. 11-15).
In the Calvari section (Fig. 3 ![]() ), Calpionellopsis gr. simplex
occurs from thin section no. BR2757/no. 3 (2010) at 7.85 meters to BR2765/10
(2010) above 35 meters while, in the Casamata section (Fig. 4
), Calpionellopsis gr. simplex
occurs from thin section no. BR2757/no. 3 (2010) at 7.85 meters to BR2765/10
(2010) above 35 meters while, in the Casamata section (Fig. 4 ![]() ), it is found from
thin section no. 94.1 (1994) at 0.5 meter to 94.22 (1994) at 17.75 meters. This
subzone is also rich in Cadosina fusca Wanner, 1940
(Pl. 9
), it is found from
thin section no. 94.1 (1994) at 0.5 meter to 94.22 (1994) at 17.75 meters. This
subzone is also rich in Cadosina fusca Wanner, 1940
(Pl. 9 ![]() , figs.
21-25).
, figs.
21-25).
The
D2 (Oblonga) subzone contains Calpionellopsis oblonga (Cadisch, 1932)
(Pl. 8 ![]() , figs. 1-5) and Calpionellopsis
protoblonga Nagy, 1986 (Pl. 7
, figs. 1-5) and Calpionellopsis
protoblonga Nagy, 1986 (Pl. 7 ![]() , figs. 31-35). This calpionellid assemblage
also includes Remaniella spp., primarily known from zones C and D. Remaniella
cf. cadischiana (Colom, 1948) (Pl. 8
, figs. 31-35). This calpionellid assemblage
also includes Remaniella spp., primarily known from zones C and D. Remaniella
cf. cadischiana (Colom, 1948) (Pl. 8 ![]() , figs. 21-25) is found in
the Calvari section (Fig. 3
, figs. 21-25) is found in
the Calvari section (Fig. 3 ![]() ) from thin section no. BR2757/no. 3 (2010) at 7.85
meters to BR2764/9 (2010) at 33.25 meters; it was not identified in the Casamata
section. Other Remaniella representatives, R. ferasini (Catalano,
1965) (Pl. 8
) from thin section no. BR2757/no. 3 (2010) at 7.85
meters to BR2764/9 (2010) at 33.25 meters; it was not identified in the Casamata
section. Other Remaniella representatives, R. ferasini (Catalano,
1965) (Pl. 8 ![]() , figs. 13-17) and R. duranddelgai Pop, 1996
(Pl. 8
, figs. 13-17) and R. duranddelgai Pop, 1996
(Pl. 8 ![]() ,
figs. 18-20), along with Calpionella
alpina Nagy, 1986 (Pl. 7
,
figs. 18-20), along with Calpionella
alpina Nagy, 1986 (Pl. 7 ![]() , figs. 1-5), and C. parvalpina Nagy, 1986
(Pl. 7
, figs. 1-5), and C. parvalpina Nagy, 1986
(Pl. 7 ![]() , figs.
6-10),
are present in the studied sections.
, figs.
6-10),
are present in the studied sections.
The calpionellid assemblage from Busot also includes Lorenziella
spp. and Tintinnopsella spp., which are present in both D and E zones. Lorenziella
hungarica Knauer & Nagy, 1964
(Pl. 8 ![]() , figs. 6-10) is found
in the Calvari section (Fig. 3
, figs. 6-10) is found
in the Calvari section (Fig. 3 ![]() ) from thin section no. BR2757/no. 3 (2010) at
7.85 meters to BR2763/8 (2010) at 29.35 meters while, in the Casamata section (Fig.
4
) from thin section no. BR2757/no. 3 (2010) at
7.85 meters to BR2763/8 (2010) at 29.35 meters while, in the Casamata section (Fig.
4 ![]() ), it is only found in two thin sections no. 94.14 (1994) and no. 94.15
(1994) at 12.30 and 12.80 meters, respectively. L.
plicata Le H�garat & Remane, 1968
(Pl. 8
), it is only found in two thin sections no. 94.14 (1994) and no. 94.15
(1994) at 12.30 and 12.80 meters, respectively. L.
plicata Le H�garat & Remane, 1968
(Pl. 8 ![]() , figs. 11-12) is observed in various thin sections, including no. BR2759/no. 4H (2010) at 12.70 meters and BR2762/no. 7 (2010) at 26.90
meters. Tintinnopsella longa (Colom, 1939)
(Pl. 9
, figs. 11-12) is observed in various thin sections, including no. BR2759/no. 4H (2010) at 12.70 meters and BR2762/no. 7 (2010) at 26.90
meters. Tintinnopsella longa (Colom, 1939)
(Pl. 9 ![]() , figs. 1-5) is found in the Calvari section
(Fig. 3
, figs. 1-5) is found in the Calvari section
(Fig. 3 ![]() ) from thin section no. BR2759/no. 4H (2010) at 12.70 meters to BR2764/9
(2010) at 33.25 meters while, in the Casamata section (Fig. 4
) from thin section no. BR2759/no. 4H (2010) at 12.70 meters to BR2764/9
(2010) at 33.25 meters while, in the Casamata section (Fig. 4 ![]() ), it is found from
thin section no. 94.1 (1994) at 0.5 meter to 94.22 (1994) at 17.75 meters.
), it is found from
thin section no. 94.1 (1994) at 0.5 meter to 94.22 (1994) at 17.75 meters.
Co-occurrences of Lorenziella spp., Remaniella spp., and Tintinnopsella spp., along with the lack of Calpionellites spp., confirm that all the material is characteristic of Zone D.
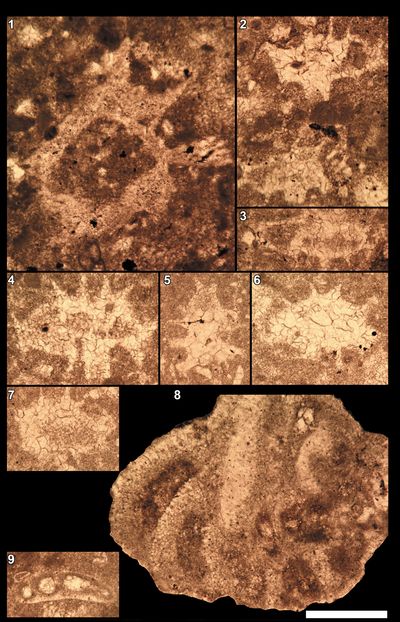
Overall preservation is poor, with significant difficulties in properly identifying the various species. For this reason, we have chosen to present most of the calcareous nannofossils observed, including some doubtful specimens.
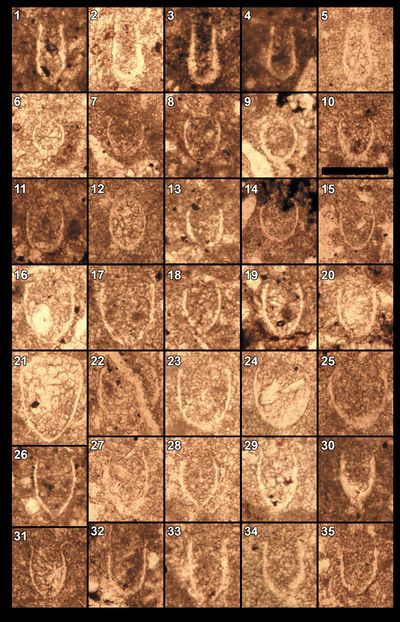
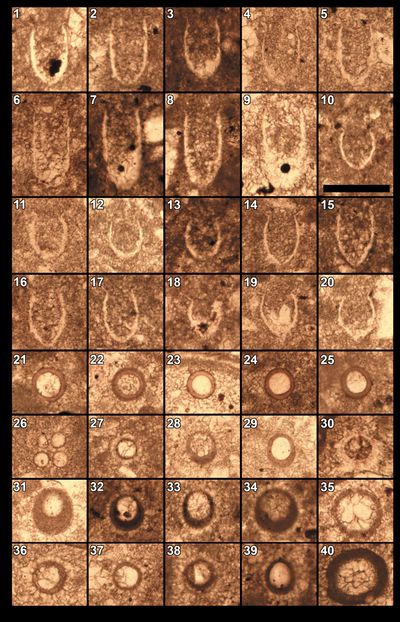
Nannoconus
species are well represented, with N. steinmanni steinmanii Kamptner,
1931
(Pl. 10 ![]() , figs. 10-15;
Pl. 12
, figs. 10-15;
Pl. 12 ![]() , figs. 7, 24-27), and N. steinmanni minor
Deres
& Ach�rit�guy, 1980 (Pl. 10
, figs. 7, 24-27), and N. steinmanni minor
Deres
& Ach�rit�guy, 1980 (Pl. 10 ![]() , figs. 1-9;
Pl. 11
, figs. 1-9;
Pl. 11 ![]() , fig. 1;
Pl. 12
, fig. 1;
Pl. 12 ![]() , figs. 1-6,
19-23) being the dominant species. No wide canal species were identified.
Additionally, N. wintereri Bralower & Thierstein,
1989 (Pl. 10
, figs. 1-6,
19-23) being the dominant species. No wide canal species were identified.
Additionally, N. wintereri Bralower & Thierstein,
1989 (Pl. 10 ![]() , figs. 28, 30-32;
Pl. 12
, figs. 28, 30-32;
Pl. 12 ![]() , fig. 32), N.
compressus Bralower
& Thierstein, 1989 (Pl. 10
, fig. 32), N.
compressus Bralower
& Thierstein, 1989 (Pl. 10 ![]() , figs. 21-23, ? 24;
Pl. 11
, figs. 21-23, ? 24;
Pl. 11 ![]() , fig. 2;
Pl. 12
, fig. 2;
Pl. 12 ![]() , ?
fig. 11), and N. infans Bralower, 1989
(Pl. 12
, ?
fig. 11), and N. infans Bralower, 1989
(Pl. 12 ![]() , figs. 8-9) were observed, though they are present in minor
quantities. These observations are consistent across all nannofossil
assemblages.
, figs. 8-9) were observed, though they are present in minor
quantities. These observations are consistent across all nannofossil
assemblages.
The set of data suggests an age:
not older than NK1 of the nannofossil zonation, Tirnovella occitanica Ammonite Standard Zone, or Calpionella elliptica Subzone (B3) of the calpionellids;
not younger than NK2A of the nannofossil zonation, Fauriella boissieri Ammonite Standard Zone, or Calpionellopsis simplex Subzone (D1) of calpionellids.
Among
the other nannofossils, stratigraphically significant species observed include Diazomatolithus
lehmanii No�l, 1965 (Pl. 11 ![]() , fig. 17), Cruciellipsis cuvillieri (Manivit,
1966) Thierstein, 1971
(Pl. 11
, fig. 17), Cruciellipsis cuvillieri (Manivit,
1966) Thierstein, 1971
(Pl. 11 ![]() , fig. 20), and
Haqius circumradiatus (Stover, 1966) (Pl. 11
, fig. 20), and
Haqius circumradiatus (Stover, 1966) (Pl. 11 ![]() , fig. 15). Polycostella beckmannii Thierstein,
1971 (Pl. 11
, fig. 15). Polycostella beckmannii Thierstein,
1971 (Pl. 11 ![]() , fig. 9;
Pl. 12
, fig. 9;
Pl. 12 ![]() , figs. 14-18), and possibly P. senaria Thierstein,
1971 (Pl. 12
, figs. 14-18), and possibly P. senaria Thierstein,
1971 (Pl. 12 ![]() , ? figs. 35-36) were also identified.
, ? figs. 35-36) were also identified.
These pieces of evidence suggest an age:
not older than NK1 of the nannofossil zonation, Tirnovella occitanica Ammonite Standard Zone, or Calpionella elliptica Subzone (B3) of the calpionellids
not younger than NK3A of the nannofossil zonation, "Thurmanniceras" pertransiens Ammonite Standard Zone, or Calpionellites Subzone (E) of the calpionellids.
The presence of Polycostella beckmannii could indicate a slightly older age, as its last appearances in the literature correspond to the equivalent of Remaniella cadischiana Zone (C) of the calpionellids. However, that is insufficient to challenge the calpionellid and ammonite data. Polycostella beckmannii is characteristic of the Tithonian/Berriasian boundary, abundant in the Crassicolaria Zone (A), with its frequency decreasing in the Calpionella Zone (B), and becoming extinct in Remaniella cadischiana Zone (C). Minor reworking, a common phenomenon in nannofossils due to their small size, cannot be excluded.
Based on the observed nannoflora, there is no evidence supporting a Hauterivian age. A Berriasian age is justifiable and aligns with the revised age given by ammonites and calpionellids.
Octahedronoides
tethysianus
Granier, 2024b (Pl. 11 ![]() , figs. 22-25), are
planktonic organisms forming small colonies, ascribed to the acritarchs. Some
were previously identified as Cadosina minuta Borza,
1980. They
are known from the middle Berriasian Elliptica Subzone of the Calpionella Zone
to the lower Valanginian Calpionellites Zone. The 'Tethysianus zone' is based on
the total range zone of this species and replaces the formerly 'Minuta zone' of Řeh�nek
(1992). These microfossils are commonly found in our thin section material,
especially in thin section no. BR2758/no. 4H (2010) of the Calvari section
(MHNG-GEPI-2024-10065) at 11.70 m (Fig. 3
, figs. 22-25), are
planktonic organisms forming small colonies, ascribed to the acritarchs. Some
were previously identified as Cadosina minuta Borza,
1980. They
are known from the middle Berriasian Elliptica Subzone of the Calpionella Zone
to the lower Valanginian Calpionellites Zone. The 'Tethysianus zone' is based on
the total range zone of this species and replaces the formerly 'Minuta zone' of Řeh�nek
(1992). These microfossils are commonly found in our thin section material,
especially in thin section no. BR2758/no. 4H (2010) of the Calvari section
(MHNG-GEPI-2024-10065) at 11.70 m (Fig. 3 ![]() ).
).
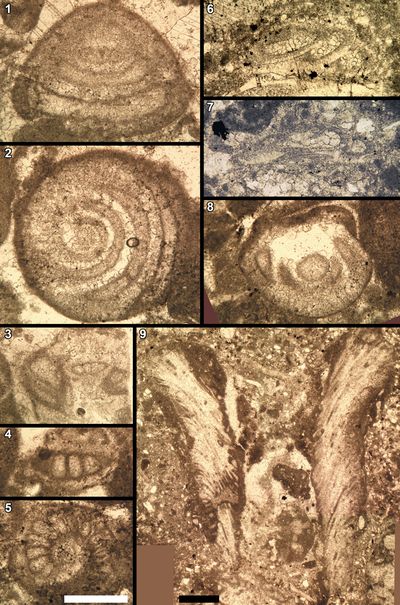
Some
organisms living in the Cabe�� d'Or shallow-water areas have been transported
off the platform and redeposited into the slope sediments at Busot. They consist
of Dasycladales and benthic Foraminifera. Although they are reworked, they may
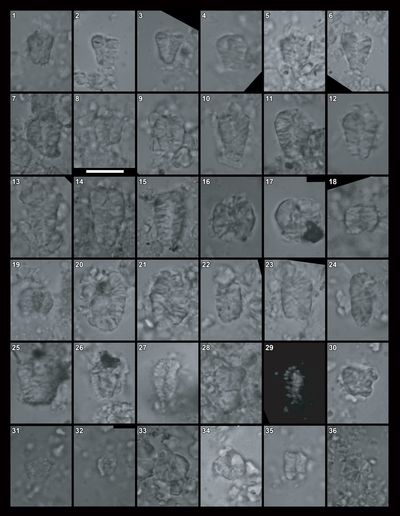
provide some valuable information. For instance, some Dasycladales, such as Macroporella?
praturloni Dragastan, 1978, and Pseudocymopolia pluricellata
Bakalova, 1973 (Pl. 15 ![]() , fig. 9;
Pl. 16
, fig. 9;
Pl. 16 ![]() , fig. 9), are respectively known
from the middle Berriasian - lower Valanginian and upper Berriasian - lower
Valanginian intervals (Granier, 2019). In
addition, some foraminifers, including Mohlerina basiliensis (Mohler,
1938) (Pl. 15
, fig. 9), are respectively known
from the middle Berriasian - lower Valanginian and upper Berriasian - lower
Valanginian intervals (Granier, 2019). In
addition, some foraminifers, including Mohlerina basiliensis (Mohler,
1938) (Pl. 15 ![]() , figs. 6-7;
Pl. 16
, figs. 6-7;
Pl. 16 ![]() , fig. 1) and Iberopora bodeuri Granier
& Berthou, 2002 (Pl. 16
, fig. 1) and Iberopora bodeuri Granier
& Berthou, 2002 (Pl. 16 ![]() , fig. 12), are not know in strata younger
than the Berriasian (see Granier, 2019
and 2024a, respectively).
, fig. 12), are not know in strata younger
than the Berriasian (see Granier, 2019
and 2024a, respectively).
Studies with reworked calpionellids are few in the literature; those with illustrations are even fewer, but demonstrative. In such cases, the reworking can easily be detected because it is documented by occurrences of inframillimetric microlithoclasts or millimetric to centimetric pseudointraclasts containing calpionellids:
1) reworking of calpionellids has been reported in the "Cenomanian Flysch" of the province of Varese, northern Italy (Reggiori, 1958: Pl. II, fig. 3; Cita, 1965: Pl. XLVI, figs. 1-2) in extraclasts whereas the matrix contains planktonic foraminifers;
2) Blau and Gr�n (1992) have also documented examples from the Aptian and/or Albian of northern Austria. One specimen (Blau & Gr�n, 1992: Pl. 1, figs. a-c) is clearly part of a microlithoclast. In other specimens (Blau & Gr�n, 1992: Pl. 1, figs. d-h, m-s), the lorica infills look slightly coarser than the matrix; in such cases, it is suspected that the outer side of the lorica coincides with the microlithoclast border (as it is also the case with the left hand side of the specimen in their Pl. 1, figs. a-c);
3) while studying calciturbidites and associated breccias (debris and mud flows) from the Tithonian-lower Berriasian of SE France, Granier et al. (2020, 2023a, 2023b) documented pseudointraclasts containing calpionellids (Granier et al., 2020: Pl. 3, figs. A-C; 2023a: Figs. 7.A, 7.L, 7.P, 8.C). They also illustrated inframillimetric microlithoclasts, commonly consisting of a single lorica with its micritic 'coating' and infill (Granier et al., 2023a: Fig. 7.M), which are the main components of cryptoturbidites (Granier et al., 2020: Pl. 2, figs. A-B; Pl. 3, fig. A; 2023a: Figs. 7.M, 8.G; 2023b: Figs. 6.L, 6.P).
Calpionellids
are common in the matrices of the Busot limestones. Although the thin loricae of
the calpionellids are assumed to be fragile, very few loricae or even their
collar structures are broken in the studied material. None appear to be
microlithoclasts, and very few are found in pseudointraclasts (Pl. 16 ![]() , fig. 10).
All these observations led Granier et al.
(1995) to state that these microfossils were not reworked, a conclusion that
will prove to be correct.
, fig. 10).
All these observations led Granier et al.
(1995) to state that these microfossils were not reworked, a conclusion that
will prove to be correct.
It is worth mentioning that, until 1995 with the publication of the Busot case (Granier et al., 1995), some species of this Busot assemblage, e.g., Calpionellopsis simplex, have never been reported from strata younger than the Valanginian. However, their association with supposedly Hauterivian ammonites led Granier et al. (1995) to state the stratigraphic ranges of some calpionellid taxa may have been longer than previously thought, a conclusion that will prove to be wrong.
Earlier misidentifications of ammonites have led to two incorrect hypotheses regarding the nature of the calpionellids from Busot:
these calpionellids were Hauterivian in age (Granier et al., 1995);
these Berriasian calpionellids were reworked into Hauterivian strata (Granier et al., 2022).
However, some ammonites are identified at the species or genus level, including Fauriella latecostata, Spiticeras ducale, Tirnovella sp., and others tentatively identified as "Thurmanniceras" cf. gratianopolitense, Erdenella cf. paquieri, and Erdenella? sp. These ammonite assemblages likely correspond to the Fauriella boissieri and Tirnovella alpillensis Standard Ammonite zones, which date to the late Berriasian.
The calpionellids from Busot belong to the Calpionellopsis Zone (D), which corresponds to the late Berriasian. While the nannofossil assemblages are less precise for dating than the ammonites, they indicate an age older than the NK1 zone and predominantly not younger than NK2A zone and, in any case, not younger than NK3A zone, pointing to a Berriasian age. Similarly, the common occurrence of Octahedronoides tethysianus supports the identification of the Tethysianus zone (formerly referred to as the 'Minuta zone' of the Cadosinas), which dates to the late Berriasian-early Valanginian interval.
The revision of part of the original ammonite finds in the 'Collections de l'Universit� Claude Bernard Lyon 1' (Villeurbanne, France), along with the study of additional ammonitic material collected in 2024, the analysis of the nannofossil assemblage, the reevaluation of the calpionellid assemblage (notably their relatively pristine preservation), and the occurrence of Octahedronoides tethysianus Granier, 2024b, all indicate that the studied sections at the Busot locality should be assigned to the upper Berriasian, rather than to the Hauterivian as previously stated (Granier et al., 1995, 2022).
In conclusion, a third hypothesis is validated here. The late Berriasian calpionellids from Busot are not reworked into Hauterivian strata. Most corresponding species did not survive the decline of the group during Valanginian times. Consequently, every earlier reference in the literature (see Granier et al., 1995) of upper Valanginian to lower Barremian occurrences of calpionellids should be reexamined with considerable scrutiny and caution.
This contribution is dedicated to three of our former colleagues and scientific contributors: Roque Aguado Merlo (1961-2023), Robert Busnardo (1926-2018), and Bernard Clavel (1938-2018). Fieldwork in SE Spain (in September 2010 and April 2024) was made possible thanks to a grant from the Association 'Carnets de G�ologie'. The studied thin sections will be deposited in the collections of the 'Mus�um d'Histoire Naturelle de Gen�ve', Switzerland. The UCBL-FSL-numbered ammonites are part of the 'Collections de l'Universit� Claude Bernard Lyon 1' (Granier Collection), Villeurbanne, France. Special thanks are due to Emmanuel Robert, the Geosciences Collection Manager, for locating part of the ammonites collected in 1993-1994 and transferred to these collections from Busnardo's home after his passing in 2018. The authors acknowledge the positive comments and suggestions provided by a group of experts, including Elisabetta Erba (Milano), Daniela Reh�kov� (Bratislava), and Miguel Company (Granada). Finally, the first author (B.R.C.G.) extends his gratitude to Phil Salvador for his assistance in refining the English text of the original manuscript.
Aguado R., Company M. & Tavera J.M. (2000).- The Berriasian/Valanginian boundary in the Mediterranean region: New data from the Caravaca and Ceheg�n sections, SE Spain.- Cretaceous Research, vol. 21, p. 1-21.
Benzaggagh M. (2020).- Discussion on the calpionellid biozones and proposal of a homogeneous calpionellid zonation for the Tethyan Realm. In: Granier B. (ed.), Virtual Special Issue on "The transition of the Jurassic to the Cretaceous: An early XXIth century holistic approach".- Cretaceous Research, vol. 114, article 104184, 24 p.
Black M. (1973).- British Lower Cretaceous coccoliths. I. Gault Clay.- Monographs of the Palaeontographical Society, London, vol. 127, no. 537, p. 49-114.
Blau J. & Gr�n B. (1992).- Calpionellen der tiefen Unterkreide im Apt/Alb der Lienzer Dolomiten: Ein Beispiel f�r umgelagerte Faunen. Revision einiger Calpionella-Arten.- Giessener Geologische Schriften, Gie�en, no. 48 Festschrift Stibane, p. 9-28.
Borza K. (1980).- Cadosina minuta n.sp. aus der unteren Kreide der Westkarpaten. Geologick� Zborn�k - Geologica Carpathica, Bratislava, vol. 31, no. 3, p. 263-266.
Bralower T.J., Monechi S. & Thierstein H.R. (1989).- Calcareous nannofossil zonation of the Jurassic-Cretaceous boundary interval and correlation with the geomagnetic polarity timescale.- Marine Micropaleontology, vol. 14, no. 1-3, p. 153-235.
Bulot L.G. (1995, unpublished).- Les formations � ammonites du Cr�tac� inf�rieur dans le Sud-Est de la France (Berriasien � Hauterivien): biostratigraphie, pal�ontologie et cycles s�dimentaires.- PhD Thesis, Mus�um National d'Histoire Naturelle, Paris, 375 p.
Bulot L.G. & Thieuloy J.-P. (1995).- Les biohorizons du Valanginien du Sud-Est de la France : Un outil fondamental pour les corr�lations au sein de la T�thys occidentale.- G�ologie Alpine, Grenoble, M�moire H.S., vol. 20 (1994), p. 15-41.
Cita M.B. (1965).- Jurassic, Cretaceous and Tertiary microfacies from the Southern Alps (Northern italy).- International Sedimentary Petrographical Series, Leiden, vol. VIII, 99 p. (CXVIII Pls.).
Company M. (1987).- Los ammonites del Valanginiense del sector oriental de las Cordilleras B�ticas (SE de Espa�a).- Universidad de Granada, 294 p.
Company M. & Tavera. J.M. (2015).- Lower Valanginian ammonite biostratigraphy in the Subbetic Domain (Betic Cordillera, southeastern Spain).- Carnets Geol., Madrid, vol. 15, no. 8, p. 71-88. DOI: 10.4267/2042/56745
Deres F. & Ach�rit�guy J. (1980).- Biostratigraphie des Nannoconid�s.- Bulletin des Centres de Recherches Exploration-Production elf-Aquitaine, Pau, vol. 4, no. 1, p. 1-53.
Djan�lidz� M.A. (1922).- Les Spiticeras du Sud-Est de la France.- M�moires pour servir � la l'explication de la carte g�ologique d�taill�e de la France, Paris, vol. 23, 207 p.
Ettachfini M. (2004).- Les ammonites n�ocomiennes dans l'Atlas atlantique (Maroc). Biostratigraphie, pal�ontologie, pal�obiog�ographie et pal�o�cologie.- Strata (s�rie 2), Toulouse, vol. 43, 224 p.
Ferr� B. & Granier B. (2000).- Roveacrinus berthoui nov. sp., Early Hauterivian representative of Roveacrinidae (Roveacrinida, Crinoidea) of Busot (Alicante, Spain).- Geologica carpathica, Bratislava, vol. 51, no. 2, p. 101-107.
Granier B. (1987).- Le Cr�tac� inf�rieur de la Costa Blanca entre Busot et Altea, Alicante (Espagne) : Biostratigraphie, s�dimentologie, �volution tectono-s�dimentaire.- Th�se, Docteur de l'Universit� Paris VI (nouveau r�gime), 23 Novembre 1987; M�moires des Sciences de la Terre, Universit� Pierre et Marie Curie, Paris, no. 87-49, vol. I (text): 281 p.; vol. II (plates): 54 Pls.
Granier B. (2019).- Dual biozonation scheme (benthic foraminifera and "calcareous" green algae) over the Jurassic-Cretaceous transition. Another plea to revert the system boundary to its historical Orbigny's and Oppel's definition. In: Granier B. (ed.), VSI: The transition of the Jurassic to the Cretaceous: An early XXIth century holistic approach.- Cretaceous Research, vol. 93, p. 245-274.
Granier B. (2022).- Tethysian, Tethyan or � Tethys Ocean and Tethys.- Carnets Geol., Madrid, vol. 22, Editorial Note 1, p. 681-683. DOI: 10.2110/carnets.2022.22EN1
Granier B. (2024a).- Reassessment of Iberopora bodeuri, a primitive plurilocular calcareous encrusting foraminifer from the "Upper Jurassic" (including Berriasian) carbonate platforms of the northern and central Tethys.- Cretaceous Research, vol. 155, article 105782, 8 p.
Granier B. (2024b).- Octahedronoides tethysianus n.gen., n.sp., enigmatic clusters of microspheres at the Jurassic-Cretaceous transition.- Carnets Geol., Madrid, vol. 24, no. 7, p. 127-133. DOI: 10.2110/carnets.2024.2407
Granier B., Benzaggagh M. & Ferry S. (2023a).- Revised holostratigraphy of the Tithonian-Berriasian transition at Tr� Maroua (Le Saix, Hautes-Alpes, SE France): Study of a rejected Berriasian GSSP candidate.- Volumina Jurassica, Varsaw, vol. XXI, p. 1-18.
Granier B., Ferry S. & Benzaggagh M. (2020).- A critical look at Tr� Maroua (Le Saix, Hautes-Alpes, France), the Berriasian GSSP candidate section.- Carnets Geol., Madrid, vol. 20, no. 1, p. 1-17. DOI: 10.4267/2042/70714
Granier B., Ferry S. & Benzaggagh M. (2022).- Calpionellid biostratigraphy judged by the yardstick of sedimentology. In: 11th International Cretaceous Symposium (Warsaw, Poland, 2022).- Abstract Volume, p. 176-177
Granier B., Ferry S. & Benzaggagh M. (2023b).- Hiatuses and redeposits in the Tithonian-Berriasian transition at Le Chouet (Les Pr�s, La Dr�me, SE France): Sedimentological and biostratigraphical implications.- Carnets Geol., Madrid, vol. 23, no. 7, p. 123-147. DOI: 10.2110/carnets.2023.2307
Granier B. & Perthuisot V. (2009).- Discussion of: Development of carbonate platforms on an extensional (rifted) margin: The Valanginian-Albian record of the Prebetic of Alicante (SE Spain), by J.M. Castro et al., Cretaceous Research 29 (2008), 848-860.- Cretaceous Research, vol. 30, Issue 4, p. 1019-1026.
Granier B., Virgone A., Busnardo R. & Bulot L.G. (1995).- Des calpionelles dans l'Hauterivien sup�rieur. D�couverte exceptionnelle � Busot (Alicante, Espagne).- Comptes-Rendus de l'Acad�mie des Sciences (S�rie II a), Paris, t. 321, p. 1179-1186.
Gr�n B. & Blau J. (1997).- New aspects of calpionellid biochronology: Proposal for a revised calpionellid zonal and subzonal division.- Revue de Pal�obiologie, Gen�ve, vol. 16, p. 197-214.
Kamptner E. (1931).- Nannoconus steinmanni novo gen., novo spec., ein merkwurdiges gesteinbildendes Mikrofossil aus dem jungeren Mesozoikum der Alpen.- Pal�ontologische Zeitschrift, vol. 13, p. 288-298.
Kenjo S. (2014, unpublished).- Biostratigraphie int�gr�e � nannofossiles calcaires et ammono�des: D�veloppement et implications pour la d�finition et la valorisation des stratotypes d'unit� et de limite. L'exemple des �tages Berriasien et Valanginien et de leur limite (~140 Millions d'ann�es).- PhD Thesis, Universit� Claude Bernard, Lyon, 226 p.
Kenjo S., Reboulet S., Mattioli E. & Ma'louleh K. (2021).- The Berriasian-Valanginian boundary in the Mediterranean Province of the Tethyan Realm: Ammonite and calcareous nannofossil biostratigraphy of the Vergol section (Montbrun-les-Bains, SE France), candidate for the Valanginian GSSP.- Cretaceous Research, vol. 121, article 104738, 27 p.
Le H�garat G. (1973).- Le Berriasien du Sud-Est de la France.- Documents des Laboratoires de G�ologie de la Facult� des Sciences de Lyon, Villeurbanne,vol. 43, 576 p.
Leret Verd� G. & Lend�nez Gonz�lez A. (1978).- Hoja 872 (Alicante).- Mapa geologico de Espa�a, Mardid, E. 1:50.000, segunda serie, primera edici�n, 33 p. URL: http://info.igme.es/cartografiadigital/geologica/Magna50Hoja.aspx?Id=872&language=es#memoria_y_metadatos
Manivit H. (1966).- Sur quelques coccolithes nouveaux du N�ocomien.- Compte Rendu sommaire des S�ances de la Soci�t� g�ologique de France, Paris, fasc. 7, p. 267-268.
Mazenot G. (1939).- Les Pal�ohoplitidae tithoniques et berriasiens du Sud-Est de la France.- M�moires de la Soci�t� g�ologique de France (nouvelle s�rie), Paris, vol. 41, 303 p.
Mi��k M., Sot�k J. & Ziegler V. (1999).- Serpulid worms Mercierella Fauvel, Durandella Dragastan and Carpathiella nov. gen. from the Jurassic, Cretaceous and Paleogene of the Western Carpathians.- Geologica Carpathica, Bratislava, vol. 50 no. 4, p. 305-312.
No�l D. (1965).- Sur les coccolithes du Jurassique europ�en et d'Afrique du Nord.- �ditions du CNRS, Paris, 209 p.
Rasplus L. & Fourcade �. (coords.), Ambroise D., And�ol B., Az�ma J., Blanc P., Busnardo R., Clerc-Renaud T., Damotte R., Dercourt J., Foucault A., Galbrun B., Granier B., Lachkar G., Le H�garat G., Magn� J., Manivit H., Mangin A.-M., Masure E., Mazaud A., Michaud F., Morand F., Renard M., Schuber N. & Taugourdeau J. (1987).- Stratigraphie int�gr�e du sillon citrab�tique (Sierra de Foncalent, Province d'Alicante, Espagne).- Geobios, Villeurbanne, vol. 20, no. 3, p. 337-387.
Reboulet S., Szives O. (reporters), Aguirre-Urreta M.B., Barrag�n R., Company M., Frau C., Kakabadze M.V., Klein J., Moreno-Bedmar J.A., Lukeneder A., Pictet A., Ploch I., Raisossadat S.N., Va��ček Z., Baraboshkin E.J. & Mitta V.V. (2018).- Report on the 6th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the Kilian Group (Vienna, Austria, 20th August 2017).- Cretaceous Research, vol. 91, p. 100-110.
Reboulet S., Jaillard �., Shmeit M., Giraud F., Masour M. & Spangenberg J. (2022).- Biostratigraphy, carbon isotope and sequence stratigraphy of South Tethyan Valanginian successions in the Essaouira-Agadir Basin (Morocco).- Cretaceous Research, vol. 140, article 105341, 27 p.
Reggiori G. (1958).- Gli affioramenti cretacei della zona nord-occidentale del Lago di Varese.- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. LXIV, fasc. 1, p. 27-46.
Remane J. (1970).- Die Entstehung der resediment�ren Breccien im Obertithon der subalpinen Ketten Frankreichs.- Dissertation (1969), G�ttingen; Eclogae Geologicae Helvetiae, Basel, vol. 63, no. 3, p. 685-740.
Řehanek J. (1992).- Valuable species of cadosinids and stomiosphaerids for determination of the Jurassic-Cretaceous boundary (vertical distribution, biozonation).- Scripta (Geology), Brno, vol. 22, p. 117-122.
Samuel O., Borza K. & K�hler E. (1972).- Microfauna and lithostratigraphy of the Paleogene and adjacent Cretaceous of the Middle Vah Valley (West Carpathian).- Geologick� �stav Dion�za �t�ra, Bratislava, 246 p. (CLXXX Pls.).
Stover L.E. (1966).- Cretaceous coccoliths and related nannofossils from France and Netherlands.- Micropaleontology, Flushing - NY, vol. 12, p. 133-167.
Szives O., Moreno-Bedmar J.A. (reporters), Aguirre-Urreta M.B., Company M., Frau C., L�pez-Horgue M., Pictet A., Ploch I., Salazar C., Barrag�n R., Latil J.-L., Lehmann J., Robert E. & Reboulet S. (2024).- Report on the 7th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the Kilian Group (Warsaw, Poland, 21st August 2022): State of the art on the current Standard Ammonite Zonation of the Western Tethyan Mediterranean Province.- Cretaceous Research, vol. 153, article 105716, 14 p.
Thierstein H.R. (1971).- Tentative Lower Cretaceous nannoplankton zonation.- Eclogae geologicae Helvetiae, Basel, vol. 64, no. 3, p. 459-488 (Pls. I-VIII).
Wippich M.G.E. (2001, unpublished).- Die tiefe Unter-Kreide (Berrias bis Unter-Hauterive) im s�dwestmarokkanischen Becken: Ammonitenfauna, Bio-und Sequenzstratigraphie.- PhD thesis, Universit�t Bochum, 142 p.
Wippich M.G.E. (2003).- Valanginian (Early Cretaceous) ammonite faunas from the western High Atlas, Morocco, and the recognition of western Mediterranean 'standard' zones.- Cretaceous Research, vol. 24, no. 4, p. 357-374.
Plate
1: Fauriella latecostata
(Mazenot, 1939), UCBL-FSL 88892, 21.15
meters, Calvari section (formerly the Busot II section). Scale bar = 1 cm. Arrow
= end of the phagmocone. |
|
Plate
2: Fauriella latecostata
(Mazenot, 1939), UCBL-FSL 88813, 10.90
meters, Casamata section (formerly the Busot III section). Scale bar = 1 cm.
Arrow = end of the phagmocone. |
|
Plate
3: Fauriella latecostata? (Mazenot, 1939), UCBL-FSL 88815, 12.75 meters, Casamata section (formerly the Busot III
section). Scale bar = 1 cm. Arrow = end of the phagmocone. |
|
Plate
4: A) "Thurmanniceras" cf. gratianopolitense (Sayn, 1907), UCBL-FSL 88817, 29.35 meters,
Calvari section (formerly the Busot II section); B) Tirnovella sp.,
UCBL-FSL 88893, 20.70 meters, Calvari section (formerly the Busot II
section). Scale bars = 1 cm. |
|
Plate
5: A) Erdenella cf. paquieri (Simionescu,
1899), UCBL-FSL 88818, 18.00 meters, Casamata section (formerly the Busot III
section); B) Erdenella? sp., UCBL-FSL 88821, 18.00 meters, Casamata section (formerly
the Busot III section). Scale bars = 1 cm. |
|
Plate
6: Spiticeras ducale
(Matheron, 1889),
UCBL-FSL 88816, 1.40 meters, Calvari section (formerly the Busot II section).
Scale bar = 1 cm. Arrow = end of the phagmocone. |
|
Plate 7: 1-5) Calpionella alpina Lorenz, 1902: 1) no. 94.2 (1994), 2) no. BR2764/no. 9 (2010), 3) no. BR2763/no. 8 (2010), 4-5) no. BR2762/no. 7 (2010); 6-10) Calpionella parvalpina Nagy, 1986: 6) no. BR2762/no. 7 (2010), 7) no. BR2759/no. 4H (2010), 8) no. 94.15 (1994), 9) no. 94.22 (1994), 10) no. BR2761/no. 6 (2010); 11-15) Calpionella elliptica Cadisch, 1932: 11) no. 94.1 (1994), 12) no. BR2765/no. 10 (2010), 13) no. BR2757/no. 3 (2010), 14) no. BR2764/no. 9 (2010), 15) no. 94.5 (1994); 16-20) Calpionellopsis brevisimplex Nagy, 1986: 16) no. 94.7 (1994), 17-18) no. BR2759/no. 4H (2010), 19) no. 94.18 (1994), 20) no. 94.15 (1994); 21-26) Calpionellopsis simplex (Colom, 1939): 21) no. BR2764/no. 9 (2010), 22) no. BR2657/no. 93.D (1993), 23) no. 94.18 (1994) (= Tintinnopsella longa in Granier et al., 1995: Fig. 4.15), 24) no. 94.10 (1994), 25) no. 94.22 (1994), 26) no. 94.7 (1994); 27-30) Calpionellopsis procerosimplex Nagy, 1986: 27) no. BR2761/no. 6 (2010), 28) no. 94.7 (1994), 29) no. 94.6 (1994), 30) no. 94.5 (1994); 31-35) Calpionellopsis protoblonga Nagy, 1986: 31) no. 94.15 (1994), 32) no. BR2759/no. 4H (2010), 33) no. 94.21 (1994), 34) no. 94.7 (1994), 35) no. BR2758/no. 4B (2010). 1, 8-9, 11, 15-16, 19-20, 23-26, 28-31, 33-34) Casamata section, 2-7, 10, 12-14,
17-18, 21-22, 27, 32, 35) Calvari section, Busot, Alicante. All photos with the
same scale bar = 100 �m, on Photo 10. |
|
Plate 8: 1-5) Calpionellopsis oblonga (Cadisch, 1932): 1-2) no. 94.18 (1994), 3-4) no. 94.7 (1994), 5) no. BR2763/no. 8 (2010); 6-10) Lorenziella hungarica Knauer & Nagy, 1964: 6) no. BR2757/no. 3 (2010), 7) no. 94.14 (1994), 8) no. 94.15 (1994), 9) no. BR2759/no. 4H (2010), 10) no. BR2762/no. 7 (2010); 11-12) Lorenziella plicata Le H�garat & Remane, 1968: 11) no. BR2762/no. 7 (2010), 12) no. BR2759/no. 4H (2010); 13-17) Remaniella ferasini (Catalano, 1965): 13) no. 94.18 (1994), 14-15) no. BR2763/no. 8 (2010), 16) no. 94.18 (1994), 17) no. BR2761/no. 6 (2010); 18-20) Remaniella duranddelgai Pop, 1996: 18-20) no. 94.17 (1994); 21-25) Remaniella cf. cadischiana (Colom, 1948): 21) no. BR2757/no. 3 (2010), 22, 25) no. BR2762/no. 7 (2010), 23) no. BR2759/no. 4H (2010), 24) no. BR2764/no. 9 (2010); 26-30) Tintinnopsella carpathica (Murgeanu & Filipescu, 1933): 26) no. 94.17 (1994), 27) no. BR2764/no. 9 (2010), 28) no. BR2763/no. 8 (2010), 29) no. BR2759/no. 4H (2010), 30) no. BR2660/no. 93.D (1993); 31) Tintinnopsella curva Benzaggagh et al., 2012: no. BR2759/no. 4H (2010);32-35) Tintinnopsella duranddelgai Benzaggagh et al., 2012: 32) no. 94.22 (1994), 33) BR2761/no. 6 (2010), 34) no. BR2763/no. 8 (2010), 35) no. BR2759/no. 4H (2010). 1-4, 7-8, 13, 16, 18-20, 26, 32) Casamata section, 5-6, 9-12, 14-15, 17, 21-25,
27-31, 33-35) Calvari section, Busot, Alicante. All photos with the same scale
bar = 100 �m, on Photo 10. |
|
Plate 9: 1-5) Tintinnopsella longa (Colom, 1939): 1) no. 94.10 (1994), 2) no. BR2660/no. 93.D (1993), 3) no. 94.22 (1994), 4) no. BR2763/no. 8 (2010), 5) no. BR2764/no. 9 (2010); 6-9) Tintinnopsella elmii Benzaggagh et al., 2012: 6) no. BR2759/no. 4H (2010), 7) no. 94.7 (1994), 8) no. 94.10 (1994) (= Tintinnopsella longa in Granier et al., 1995: Fig. 4.13), 9) no. 94.5 (1994); 10-13) Tintinnopsella largovalata Benzaggagh et al., 2012: 10) no. 94.10 (1994), 11) no. BR2763/no. 8 (2010), 12) no. BR2759/no. 4H (2010), 13) no. 94.21 (1994); 14-17) Tintinnopsella longovalata Benzaggagh et al., 2012: 14, 16) no. BR2759/no. 4H (2010), 15, 17) no. 94.2 (1994); 18-20) Tintinnopsella minicarpathica Benzaggagh et al., 2012: 18) no. 94.15 (1994), 19) no. BR2763/no. 8 (2010), 20) no. BR2765/no. 10 (2010); 21-25) Cadosina fusca Wanner 1940: 21) no. 94.7 (1994), 22) no. BR2757/no. 3 (2010), 23) no. BR2760/no. 5 (2010), 24) no. BR2759/no. 4H (2010), 25) no. BR2657/no. 93.D (1993); 26) Octahedronoides tethysianus Granier, 2024b: no. BR2763/no. 8 (2010); 27) Colomisphaera heliosphaera (Vogler, 1941): no. BR2760/no. 5 (2010); 28-29) Cadosina fusca cieszynica Nowak, 1966: 28) no. 94.7 (1994), 29) no. BR2758/no. 4B (2010); 30) Stomiosphaera echinata Nowak, 1968: no. BR2755/no. 1 (2010); 31) Cadosina semiradiata olzae Nowak, 1968: no. BR2757/no. 3 (2010); 32-34) Cadosina semiradiata aff. olzae Nowak, 1968: 32) no. 94.22 (1994), 33) BR2761/no. 6 (2010), 34) no. 94.18 (1994); 35-39) Cadosina semiradiata Wanner 1940: 35) no. BR2757/no. 3 (2010), 36-37) no. BR2764/no. 9 (2010), 38) no. BR2660/no. 93.D (1993), 39) no. 94.21 (1994); 40) Cadosina radiata Vogler, 1941: no. BR2755/no. 1 (2010). 1, 3, 7-10, 13, 15, 17-18, 21, 28, 32, 34, 39) Casamata section, 2, 4-6, 11-12,
14, 16, 19-20, 22-27, 29-31, 33, 35-38, 40) Calvari section, Busot, Alicante.
All photos with the same scale bar = 100 �m, on Photo 10. |
|
Plate 10: Due to the relatively poor preservation of the specimens, we have chosen to illustrate most of the observed nannofossils, including those that are non-diagnostic. 1-9) Nannoconus steimannii subsp. minor Deres & Ach�rit�guy, 1980; 10-15) Nannoconus steinmannii subsp. steinmannii Kamptner, 1931; 16-17) Nannoconus globulus subsp. minor Bralower, 1989; 18-19) Nannoconus sp.; 20) Nannoconus sp. (? group compactus); 21-24) Nannoconus compressus Bralower & Thierstein, 1989 (24, doubtfull specimen); 25-26) Nannoconus kamptnerii subsp. minor Deres & Ach�rit�guy, 1980; 27, 29) Faviconus ? sp., 27) LN, 29) LP (doubtfull specimen); 28, 30-32) Nannoconus wintereri Bralower & Thierstein, 1989; 35) Nannoconus sp.; 33-34, 36) Assipetra or ? Nannoconid. 1-36)
All specimens from no. 94.15 (1994), 11.95 m, Casamata section, Busot, Alicante; 22-23) no. BR2764/no. 9
(2010), Calvari section, Busot, Alicante. All
photos with the same scale bar = 10 �m, on Photo 8.
|
|
Plate 11: 1-2) Cluster 1: 1) Nannoconus steimannii subsp. minor Deres & Ach�rit�guy, 1980; 2) Nannoconus sp. cf. compressus Bralower & Thierstein, 1989; 3-5) Cluster 2: 3-4) Polycostella beckmannii Thierstein, 1971 (doubtfull specimens); 5) Rucinolithus irregularis Thierstein, 1972; 6) Braarudosphaera regularis (Black, 1973);7) Watznaueria sp.; 8-11) Polycostella beckmannii Thierstein, 1971, 10) LN, 11) LP (8 & 10-11 doubtfull larger specimens); 12) Watznaueria barnesiae (Black, 1959); 13) Watznaueria biporta Bukry, 1969; 14) ? Cyclagelosphaera margerelii No�l, 1965 (large form); 15) Haqius circumradiatus (Stover, 1966); 16) Cyclagelosphaera sp., small form; 17) Diazomatolithus lehmanii No�l, 1965; 18-19) Zeugrhabdotus embergeri (No�l, 1958); 20) Cruciellipsis cuvillieri (small form) (Manivit, 1966); 21) Rhagodiscus sp.; 22-25) Octahedronoides tethysianus Granier, 2024b. 1-21) no. 94.15
(1994), 11.95 m, Casamata section, Busot, Alicante; 22-23) no. BR2764/no. 9
(2010), Calvari section, Busot, Alicante;
24) 6521-CALV-1 (Antonio Est�vez Rubio Collection,
Alicante), Calvari section, Busot, Alicante;
24-25) no. 94.2 (1994), Casamata section, Busot, Alicante. Photos 1-11
with the same white scale bar = 10 �m, on Photo 11; photos 12-21 with the same
yellow scale bar = 10 �m, on Photo 21; photos 22-25 with the same orange scale
bar = 100 �m, on Photo 25. |
|
Plate 12: 1-6, 19-23) Nannoconus steimannii subsp. minor Deres & Ach�rit�guy, 1980; 7, 24-27) Nannoconus steinmannii subsp. steinmannii Kamptner, 1931; 8-9) Nannoconus infans Bralower, 1989; 10) Nannoconus sp. (aff. globulus minor); 11) ? Nannoconus compressus Bralower & Thierstein, 1989; 12) Nannoconus cornuta Deres & Ach�rit�guy, 1980; 13) Assipetra sp.; 14-18) Polycostella beckmannii Thierstein, 1971, 14-15) same specimen LN; 16-18) same specimen, 16-17) LN, 18) LP; 28-31) indeterminate Nannoconus; 32) Nannoconus wintereri Bralower & Thierstein, 1989; 33) Assipetra sp. (doubtfull specimens); 34) Polycostella beckmannii Thierstein, 1971; 35-36) ? Polycostella senaria Thierstein, 1971, 35) LN, 36) LP. 1-18) no. 3
(2010), (CLV_3), 6.95 m; 19-36) no. 5 (2010), (CLV_5), 14.45 m, Calvari section,
Busot, Alicante. All photos with
the same scale bar = 10 �m, on Photo 18. |
|
Plate 13: 1) Roveacrinus berthoui Ferr� & Granier, 2000, holotype, no. 94.5 (1994); 2-7) Epistomina spp., 2) no. 94.14 (1994), 3) no. 94.18 (1994), 4) no. BR2761/no. 6 (2010), 5, 7) no. BR2763/no. 8 (2010), 6) no. BR2764/no. 9 (2010); 8-9) Mohlerina basiliensis (Mohler, 1938), 8) no. 94.20 (1994), 9) no. BR2657/no. 93.D (1993). 1-3, 8) Casamata section, 4-7, 9) Calvari section, Busot, Alicante.
All photos with the same scale bar = 250 �m, on Photo 8. |
|
Plate 14: 1-24) Ichnusella gr. infragranulata (Noth, 1951) - reicheli (Neagu, 1995), 1-2) no. BR2660/no. 93.D (1993); 3, 9, 13) no. 94.25 (1994); 4) no. 94.26 (1994); 5, 21) no. 94.7 (1994); 6) no. 94.8 (1994); 7) no. 94.10 (1994); 8) no. 94.15 (1994); 10, 22, 26) no. 94.26 (1994); 11) no. BR2755/no. 1 (2010); 12) no. 94.12 (1994); 14-15, 18-20, 24) no. BR2765/no. 10 (2010); 16) no. BR2764/no. 9 (2010); 17) no. BR2763/no. 8 (2010); 23) no. BR2762/no. 7 (2010); 25) Coscinoconus sp. as the nucleus of a superficial ooid, no. BR2765/no. 10 (2010); 26) broken ooid or more likely a small lithoclast, no. BR2765/no. 10 (2010); 27) bored ooid, no. BR2755/no. 1 (2010); 28) bored lithoclast with ooids, no. BR2755/no. 1 (2010). 1-2, 11, 14-20, 23-28) Calvari section, 3-10, 12-13, 21-22, 26) Casamata
section, Busot, Alicante. All photos with the same white scale bar (on Photo 26)
= 250 �m, except for photos 27-28 with the same black scale bar (on Photo 27) =
250 �m. |
|
Plate 15: 1) Coscinoconus sp. 6521-BAR-1; 2, 8) Frentzenella sp.; 3-5) Protopeneroplis ultragranulata (Gorbachik, 1971); 6-7) Mohlerina basiliensis (Mohler, 1938); 9) Pseudocymopolia pluricellata Bakalova, 1973. 1-5) no. 6521-BAR-1,
Antonio Est�vez Rubio Collection,
Alicante; 6, 9) no. 6521-CALV-1,
Calvari section, Antonio Est�vez Rubio Collection,
Alicante; 8) no.
BR2765/no. 10 (2010), Calvari section, Busot, Alicante. All photos with the same
white scale bar (on Photo 5) = 250 �m, except for photo 9 with black scale bar
= 500 �m. |
|
Plate 16: 1) Mohlerina basiliensis (Mohler, 1938); 2-6) Protopeneroplis ultragranulata (Gorbachik, 1971); 8) "Girvanella"-like microbial structure; 9) Pseudocymopolia pluricellata Bakalova, 1973; 10) pseudointraclast with a calpionellid; 11) rhyncholite; 12) Iberopora bodeuri Granier & Berthou, 2002; 13) calcareous tubeworm, Carpathiella triangulata Mi��k et al., 1999 ("cf. Haliotus" sensu Samuel et al., 1972). 1) no. BR2763/no. 8 (2010); 2) no. 94.25 (1994); 3) no. 94.1 (1994); 4-5,
7) no. BR2764/no. 9 (2010); 6) no. BR2765/no. 10 (2010); 8) no. BR2762/no. 7
(2010); 9) no. 94.10 (1994); 10) no. BR2764/no. 9 (2010); 11) no. BR2756/no. 2
(2010); 12) no. 94.26 (1994). 1, 4-8, 10-11) Calvari section; 2-3, 9, 12)
Casamata section, Busot, Alicante. All photos with the same black scale bar (on
Photo 9) = 250 �m, except for photo 13 with white scale bar = 500 �m. |
|
Plate 17: 1-13) calcareous tubeworms, serpulids. 1-2, 12) no. BR3427/no. 18.25 (2024); 3) no. 94.19 (1994); 4-5) no. no. 94.13 (1994); 6) no. 94.22 (1994); 7) no. 94.17 (1994); 8) no. 94.14 (1994); 9) no. 94.16 (1994); 10) no. 94.21 (1994); 11, 13) no. BR2762/no. 7 (2010). 1-2, 11-13) Calvari section; 3-10)
Casamata section, Busot, Alicante. All photos with the same black scale bar (on
Photo 10) = 500 �m. |
|
List of thin sections housed at the 'Mus�um d'Histoire Naturelle de Gen�ve', Switzerland.
| MHNG-GEPI-2024-10031: no. 94.1 (1994), Casamata section |
| MHNG-GEPI-2024-10032: no. 94.2 (1994), Casamata section |
| MHNG-GEPI-2024-10033: no. 94.3 (1994), Casamata section |
| MHNG-GEPI-2024-10034: no. 94.4 (1994), Casamata section |
| MHNG-GEPI-2024-10035: no. 94.5 (1994), Casamata section |
| MHNG-GEPI-2024-10036: no. 94.6 (1994), Casamata section |
| MHNG-GEPI-2024-10037: no. 94.7 (1994), Casamata section |
| MHNG-GEPI-2024-10038: no. 94.8 (1994), Casamata section |
| MHNG-GEPI-2024-10039: no. 94.9 (1994), Casamata section |
| MHNG-GEPI-2024-10040: no. 94.10 (1994), Casamata section |
| MHNG-GEPI-2024-10041: no. 94.11 (1994), Casamata section |
| MHNG-GEPI-2024-10042: no. 94.12 (1994), Casamata section |
| MHNG-GEPI-2024-10043: no. 94.13 (1994), Casamata section |
| MHNG-GEPI-2024-10044: no. 94.14 (1994), Casamata section |
| MHNG-GEPI-2024-10045: no. 94.15 (1994), Casamata section |
| MHNG-GEPI-2024-10046: no. 94.16 (1994), Casamata section |
| MHNG-GEPI-2024-10047: no. 94.17 (1994), Casamata section |
| MHNG-GEPI-2024-10048: no. 94.18 (1994), Casamata section |
| MHNG-GEPI-2024-10049: no. 94.19 (1994), Casamata section |
| MHNG-GEPI-2024-10050: no. 94.20 (1994), Casamata section |
|
MHNG-GEPI-2024-10051: no. 94.21 (1994), Casamata section |
| MHNG-GEPI-2024-10052: no. 94.22 (1994), Casamata section |
| MHNG-GEPI-2024-10053: no. 94.23 (1994), Casamata section |
| MHNG-GEPI-2024-10054: no. 94.24 (1994), Casamata section |
| MHNG-GEPI-2024-10055: no. 94.25 (1994), Casamata section |
| MHNG-GEPI-2024-10056: no. 94.26 (1994), Casamata section |
| MHNG-GEPI-2024-10057: no. BR2657/no. 93.D (1993), Calvari section |
| MHNG-GEPI-2024-10060: no. BR2660/no. 93.D (1993), Calvari section |
| MHNG-GEPI-2024-10061: no. BR2755/no. 1 (2010), Calvari section |
| MHNG-GEPI-2024-10062: no. BR2756/no. 2 (2010), Calvari section |
| MHNG-GEPI-2024-10063: no. BR2757/no. 3 (2010), Calvari section |
| MHNG-GEPI-2024-10064: no. BR2758/no. 4B (2010), Calvari section |
| MHNG-GEPI-2024-10065: no. BR2759/no. 4H (2010), Calvari section |
| MHNG-GEPI-2024-10066: no. BR2760/no. 5 (2010), Calvari section |
| MHNG-GEPI-2024-10067: no. BR2761/no. 6 (2010), Calvari section |
| MHNG-GEPI-2024-10068: no. BR2762/no. 7 (2010), Calvari section |
| MHNG-GEPI-2024-10069: no. BR2763/no. 8 (2010), Calvari section |
| MHNG-GEPI-2024-10070: no. BR2764/no. 9 (2010), Calvari section |
| MHNG-GEPI-2024-10071: no. BR2765/no. 10 (2010), Calvari section |
| MHNG-GEPI-2024-10072: BR3435/route (2024), below Calvari section |
| MHNG-GEPI-2024-10073: BR3423/C11.25 (2024), Calvari section |
| MHNG-GEPI-2024-10074: BR3424/C11.80 (2024), Calvari section |
| MHNG-GEPI-2024-10075: BR3425/C12.00 (2024), Calvari section |
| MHNG-GEPI-2024-10076: BR3426/C14.20 (2024), Calvari section |
| MHNG-GEPI-2024-10077: BR3427/C18.25 (2024), Calvari section |
| MHNG-GEPI-2024-10078: BR3428/C24.30 (2024), Calvari section |
| MHNG-GEPI-2024-10079: BR3429/C31.50 (2024), Calvari section |