12th Meeting of the International Nannoplankton Association
(Lyon, September 7-10, 2008)
Guidebook for the post-congress fieldtrip
in the Vocontian Basin, SE France
(September 11-13, 2008)
_
Emanuela (special editor),
Silvia , Fabienne , Davide ,
Bernard & Stéphane
Davide with the contribution of Emanuela
The Digne area is located in southeastern France (Fig. 1.1 ![]() ), in the
department of "Alpes de Haute-Provence". French authors classically name this region
the "Dauphinois Basin", for the major portion of the Jurassic period, and
the "Vocontian Trough" (or Basin) for the Late Jurassic - Late Cretaceous interval.
For the Jurassic succession discussed here the name "French Subalpine Basin" is
preferred. In Jurassic and Cretaceous times the French Subalpine Basin was a gulf located along the
northwestern margin of the Tethys (Fig. 1.2
), in the
department of "Alpes de Haute-Provence". French authors classically name this region
the "Dauphinois Basin", for the major portion of the Jurassic period, and
the "Vocontian Trough" (or Basin) for the Late Jurassic - Late Cretaceous interval.
For the Jurassic succession discussed here the name "French Subalpine Basin" is
preferred. In Jurassic and Cretaceous times the French Subalpine Basin was a gulf located along the
northwestern margin of the Tethys (Fig. 1.2 ![]() ). The
gulf was bounded to the West by the Paleozoic "Massif Central" and to the East by the Alpine chain. To the South,
was a land mass represented now by the Corsica-Sardinia and Maures-Esterel massifs.
). The
gulf was bounded to the West by the Paleozoic "Massif Central" and to the East by the Alpine chain. To the South,
was a land mass represented now by the Corsica-Sardinia and Maures-Esterel massifs.
Some authors (, 1984, 1985) consider the French Subalpine Basin as an analog of the present European margin of the Atlantic Ocean, for it was formed by a series of tilted blocks that deepened eastward toward the open Tethyan Ocean. (1990) suggests that the evolution of the area was in relation with an aborted rift basin. In this view, the French Subalpine Basin may be considered as a sort of pull-apart basin. In Middle Jurassic times, the French Subalpine Basin was a transitional area between the epicontinental sea of the Paris Basin and the deeper water Piedmont domain, where an oceanic ridge was active until the Early Cretaceous. The basin probably attained its maximum depth (~500 - 800 m) in the Early Cretaceous (Hauterivian-Barremian; , personal communication).
The French Subalpine Basin began its development during the Middle to Late Triassic, with the formation of a Germanic-type salt basin along the fractured Alpine margin of the Tethys in which evaporites predominate (, 1990). During Jurassic times the shallow sea deepened after a general marine transgression (, 2001). In the Early Jurassic, a threshold (the Verdon sill) separated a shelf area (the Provence Platform) to the north from a deeper facies (dolomitic limestones) to the south. On this threshold is a sedimentary succession consisting of mixed carbonate-silicoclastic sediments that is reduced in thickness northward, with many hiatuses (, 1984). Basinal facies, represented by marl-limestone alternations, are recorded in the southernmost areas but in the sector south of the Verdon River a shallow-water platform developed, documented by the occurrence of bioclastic limestones with corals. This development is evidence of a progressive extension to the Provence platform southward.
In the Middle Jurassic, the central part of the basin (Laragne area, North of Sisteron; Fig. 1.2 ![]() ) was
subject to strong subsidence, as evinced by dark-colored marls formed under dysoxic conditions ("Terres Noires" Formation). Conversely, along the southernmost edge of the basin, shallower facies and a dominantly carbonate sedimentation are documented (,
1984). A marked regression occurred during the Late Jurassic (,
1984), when calcareous sediments indicative of a shelf environment were deposited
over the entire basin (Tithonian cliffs).
) was
subject to strong subsidence, as evinced by dark-colored marls formed under dysoxic conditions ("Terres Noires" Formation). Conversely, along the southernmost edge of the basin, shallower facies and a dominantly carbonate sedimentation are documented (,
1984). A marked regression occurred during the Late Jurassic (,
1984), when calcareous sediments indicative of a shelf environment were deposited
over the entire basin (Tithonian cliffs).
From Berriasian through Aptian times, in a general regime of extensional tectonics, the depth of the basin increased progressively. Limestone-marl alternations were deposited on the Tithonian older strata (, 1984). A maximum depth was probably attained during the Aptian-Albian interval. At the end of the Albian, the basin began to be infilled; general emergence took place during the Santonian. This uplift is contemporaneous with the first phase of Alpine tectonics: the "Pyrénéo-Provençale" phase. This phase involved the formation of structures with a general W-to-E trend.
The Aalenian-Bajocian interval in the Digne area is represented by
a thick series of marl-limestone alternations. Limestones are both packstones (with recrystallised Bositra, rare benthic foraminifers and siliceous sponge spicules) and wackestones with radiolarians, rare Bositra,
lagenid foraminifers and sponge spicules (,
1983). Marls are locally very shaly and finely laminated. This stratigraphic succession
is the equivalent of the "Calcaires à Cancellophycus" Formation, which in the Digne area is dated Middle Aalenian - end of Bajocian or
basal Bathonian. These limestones are overlain by the dark-grey to black marls of the
"Terres Noires" Formation. The thickness of the "Terres Noires" Fm.
ranges widely in the French Subalpine Basin: it may reach 700 metres in the section near Les Dourbes (Figs. 1.1b and c ![]() ). The age of the Terres Noires Formation is Late Bathonian to Middle Oxfordian, with a hiatus at its base.
). The age of the Terres Noires Formation is Late Bathonian to Middle Oxfordian, with a hiatus at its base.
Cephalopods (ammonoids, belemnoids and nautiloids) are locally abundant both in the "Calcaires à Cancellophycus" and in the "Terres Noires" formations, their presence thus allowing detailed biostratigraphical studies (, 1967; , 1973, 1983). Ichnofossils are also common, represented mainly by Chondrites and Zoophycos (Cancellophycus of French authors). Locally Zoophycos can be very abundant (, 2003). These marl and limestone alternations were deposited on the continental slope, at depths ranging between 300 and 500 metres (, 2003).
In the Digne area, an uninterrupted and beautifully exposed section covering the entire Middle Jurassic
exists in the "Ravine du Feston", near Les Dourbes, 5 km east-southeast of Digne (Fig. 1.1b ![]() ). The section
is exposed on the norteast side of the valley (Fig. 1.1c
). The section
is exposed on the norteast side of the valley (Fig. 1.1c ![]() ). Here, the Bajocian
succession is 250 metres thick
(Figs. 1.3
). Here, the Bajocian
succession is 250 metres thick
(Figs. 1.3 ![]() and 1.4
and 1.4 ![]() ). Several
biostratigraphical studies have been carried out (,
1973, 1983; & ,
1968), and a sequence stratigraphy has been developed (,
1990, 1991; & ,
1995). The base of the section studied by
(1973) is at the floor of the valley, and can be followed up to the ridge
from an elevation of 874 metres to the Serre d'Enchas, a few tens of metres south of the village
of Condamine (Fig. 1.1c
). Several
biostratigraphical studies have been carried out (,
1973, 1983; & ,
1968), and a sequence stratigraphy has been developed (,
1990, 1991; & ,
1995). The base of the section studied by
(1973) is at the floor of the valley, and can be followed up to the ridge
from an elevation of 874 metres to the Serre d'Enchas, a few tens of metres south of the village
of Condamine (Fig. 1.1c ![]() ).
).
The lower part of the section is made up of an alternation of limestones and shaly marls dated as
the Humphresianum
ammonite Zone. After a clear break in the topographic profile (Fig. 1.3 ![]() ), the marl-limestone alternations become richer in clays, especially in the upper
part of the section. This interval corresponds to the Niortense (42 m) and Garantiana
ammonite zones, and to the Tetragona Subzone (43 m), where a slump of 8 metres is clearly visible (Fig. 1.4
), the marl-limestone alternations become richer in clays, especially in the upper
part of the section. This interval corresponds to the Niortense (42 m) and Garantiana
ammonite zones, and to the Tetragona Subzone (43 m), where a slump of 8 metres is clearly visible (Fig. 1.4 ![]() ). The upper part of the section
(Parkinsoni ammonite Zone) is characterized by thin calcareous beds in alternation with thick
marls. In time terms this interval corresponds roughly to the base of the "Terres Noires" Formation. A hiatus followed by a flooding surface (,
1990) separates the Bajocian from the Bathonian stage.
The basal Bathonian is represented by the Zigzag
ammonite Zone (Convergens Subzone) in a "Terres Noires" facies.
). The upper part of the section
(Parkinsoni ammonite Zone) is characterized by thin calcareous beds in alternation with thick
marls. In time terms this interval corresponds roughly to the base of the "Terres Noires" Formation. A hiatus followed by a flooding surface (,
1990) separates the Bajocian from the Bathonian stage.
The basal Bathonian is represented by the Zigzag
ammonite Zone (Convergens Subzone) in a "Terres Noires" facies.
Three sections of the Digne area were studied by
(1990): the Beaumont section (close to the Les Dourbes section), Chaudon
(near Barrême), and La Blache (near Castellane, some 10 km south of Barrême; Fig. 1.1b ![]() ). The
several bio-horizons, precisely correlated by
(1990) with the ammonite zones, are shown in
Figure 1.4
). The
several bio-horizons, precisely correlated by
(1990) with the ammonite zones, are shown in
Figure 1.4 ![]() . The most striking pattern
of events in
Figure 1.4
. The most striking pattern
of events in
Figure 1.4 ![]() is the great number of first occurrences (FO) in the vicinity of the Aalenian/Bajocian boundary.
The new occurrences are mainly species of the genus Watznaueria, which experienced a major diversification at that time. In fact, only three species of Watznaueria are known from the Toarcian, namely W. fossacincta, W. colacicchii, and W. contracta. Conversely,
in the interval between the end of the Aalenian and the Early Bajocian, several new species of Watznaueria
make their appearance, in a number of morphotypes.
is the great number of first occurrences (FO) in the vicinity of the Aalenian/Bajocian boundary.
The new occurrences are mainly species of the genus Watznaueria, which experienced a major diversification at that time. In fact, only three species of Watznaueria are known from the Toarcian, namely W. fossacincta, W. colacicchii, and W. contracta. Conversely,
in the interval between the end of the Aalenian and the Early Bajocian, several new species of Watznaueria
make their appearance, in a number of morphotypes.
The FOs shown in Figure 1.4 ![]() may underrepresent their true number. In fact, only the FO of W. britannica and of the large morphotype of W. britannica are
noted by
(1990), repectively in the Discites and Sauzei ammonite
zones. Recently, in the Oxfordian of SW Germany, et alii
(2006) differentiated six morphotypes of W. britannica that
differ in size significantly. Using biometrics, the authors interpret these morphotypes
to be the result of intra-specific variability, in particular in the size of the coccoliths. However, differences
also exist in the structure of the bridge in the central area of the coccoliths of the
several morphotypes. Unfortunately, this structure is too small to be analysed biometrically.
A few of the samples from the Les Dourbes section have been studied, in order to check the occurrence of the six morphotypes
described by et alii
(2006). In the forms of Early Bajocian age, five of the six morphotypes described by et alii
(2006) were recognized: morphotypes A, B, D, E and F (Fig. 1.5
may underrepresent their true number. In fact, only the FO of W. britannica and of the large morphotype of W. britannica are
noted by
(1990), repectively in the Discites and Sauzei ammonite
zones. Recently, in the Oxfordian of SW Germany, et alii
(2006) differentiated six morphotypes of W. britannica that
differ in size significantly. Using biometrics, the authors interpret these morphotypes
to be the result of intra-specific variability, in particular in the size of the coccoliths. However, differences
also exist in the structure of the bridge in the central area of the coccoliths of the
several morphotypes. Unfortunately, this structure is too small to be analysed biometrically.
A few of the samples from the Les Dourbes section have been studied, in order to check the occurrence of the six morphotypes
described by et alii
(2006). In the forms of Early Bajocian age, five of the six morphotypes described by et alii
(2006) were recognized: morphotypes A, B, D, E and F (Fig. 1.5 ![]() ). The morphotypes A and B are recorded from the base of the
Bajocian. Morphotypes D, E and F are in samples of Late Bajocian age. They
appear after the last occurrence (LO) of Carinolithus superbus. In the light of recent study on
cryptic or pseudo-cryptic species ( et alii,
2002), it cannot be completely excluded that the morphotypes shown by et alii
represent discrete biological species.
). The morphotypes A and B are recorded from the base of the
Bajocian. Morphotypes D, E and F are in samples of Late Bajocian age. They
appear after the last occurrence (LO) of Carinolithus superbus. In the light of recent study on
cryptic or pseudo-cryptic species ( et alii,
2002), it cannot be completely excluded that the morphotypes shown by et alii
represent discrete biological species.
The increase in speciation between the Late Aalenian and the Early Bajocian represents the second major pulse of diversification in Jurassic coccolithophores, following the first speciation at the Pliensbachian/Toarcian boundary. Furthermore, the Aalenian/Bajocian diversification is coincident with the emergence of the genus Watznaueria that soon becomes dominant in the assemblages of coccoliths during the remainder of the Jurassic and the entire Cretaceous. This major event in coccolith history is probably a result of an important change in paleoceanographic conditions in Mid Jurassic times. The pattern of oceanic circulation was altered and the surface occupied by epicontinental seas was reduced at that time, probably in relation to the establishment of effective connections between the western Tethys and the central Atlantic. It is however difficult to differentiate any one cause that triggered coccolithophore evolution. A combination of various environmental modifications (such as changes in pCO2, nutrient availability, ocean chemistry, climate, and sea level fluctuations) may together have been responsible for evolutionary innovations (, 2006).
Stéphane
The La Charce section is located in the French department of Drôme (Fig. 1.1a ![]() ). The
biostratigraphy of the section has been well-studied. In the last four decades a
considerable number of works on palaeontology and biostratigraphy have been published: on ammonoids (,
1977a; et alii,
1992; et alii,
1993, 1996; ,
1995; , 1996; & ,
1997, 1999, and references therein),
on belemnites ( & ,
2002), on trace fossils (,
1984; & ,
1987; , 1996),
on foraminifers (,
1966; , 1992; & ,
1994), and on calcareous nannofossils (,
1973; , this
volume). Sedimentological, geochemical and palaeomagnetic data are also available ( et alii,
1980; et alii,
1989; , 1991; et alii,
1999; et alii,
2003; , 2007, and references therein). Here,
we present the lithological evolution of the La Charce section, along with a synthesis of
the biostratigraphic work on ammonoids.
). The
biostratigraphy of the section has been well-studied. In the last four decades a
considerable number of works on palaeontology and biostratigraphy have been published: on ammonoids (,
1977a; et alii,
1992; et alii,
1993, 1996; ,
1995; , 1996; & ,
1997, 1999, and references therein),
on belemnites ( & ,
2002), on trace fossils (,
1984; & ,
1987; , 1996),
on foraminifers (,
1966; , 1992; & ,
1994), and on calcareous nannofossils (,
1973; , this
volume). Sedimentological, geochemical and palaeomagnetic data are also available ( et alii,
1980; et alii,
1989; , 1991; et alii,
1999; et alii,
2003; , 2007, and references therein). Here,
we present the lithological evolution of the La Charce section, along with a synthesis of
the biostratigraphic work on ammonoids.
A detailed lithology of the Valanginian-Hauterivian portion of the La Charce section is presented in
(1996) and &
(1999). So here we discuss only the stratigraphy of
the sequence encompassing the boundary between the stages (Fig. 2.1 ![]() ). There
the lithology is characterized by marl-limestone alternations. The section is intensively bioturbated,
with both limestones and marls affected (,
1984). Zoophycos feeding burrows are common in
the limestones (,
1996). Carbonate-rich marl-limestone alternations predominate the lowest Valanginian
beds (Pertransiens ammonite Zone), then gradually become richer in argillaceous
content in the uppermost part of the Lower Valanginian
(Campylotoxus ammonite Zone). The Verrucosum
ammonite Zone of the Upper Valanginian (Fig. 2.2a
). There
the lithology is characterized by marl-limestone alternations. The section is intensively bioturbated,
with both limestones and marls affected (,
1984). Zoophycos feeding burrows are common in
the limestones (,
1996). Carbonate-rich marl-limestone alternations predominate the lowest Valanginian
beds (Pertransiens ammonite Zone), then gradually become richer in argillaceous
content in the uppermost part of the Lower Valanginian
(Campylotoxus ammonite Zone). The Verrucosum
ammonite Zone of the Upper Valanginian (Fig. 2.2a ![]() )
is also the site of an increase in argillaceous content. The relative abundance
of carbonate in the marl-limestone succession increases gradually upward toward the Valanginian/Hauterivian boundary. The
marls and limestones of Early Hauterivian age show a marked contrast: white limestones quite regularly alternate with dark marls (Fig. 2.2b
)
is also the site of an increase in argillaceous content. The relative abundance
of carbonate in the marl-limestone succession increases gradually upward toward the Valanginian/Hauterivian boundary. The
marls and limestones of Early Hauterivian age show a marked contrast: white limestones quite regularly alternate with dark marls (Fig. 2.2b ![]() ). Their thicknesses
are comparable.
). Their thicknesses
are comparable.
The alternation of marl and limestone has been interpreted as the result of cycles in the production of calcareous nannoplankton caused by climatic fluctuations in the frequency band ( et alii, 1980; , 1995). Alternatively, et alii (2003) have proposed for the Vocontian Basin a model of cyclic export of carbonate mud from shallow platform environments towards the basin. Occasionally, synsedimentary slumping and turbidites (rust-coloured calcarenites) occurred in the basin. These calcarenites are particularly well-exposed in portions of the Campylotoxus and Verrucosum zones of the La Charce section (, 1996, and references therein).
Recent research ( C., J.-F., P., J.-P. & S., in progress) on some outcrops in the Vocontian Basin (La Charce and Vergol sections in the Drôme, Montclus in the Hautes-Alpes) has revealed the occurrence of centimetre-thick goethite-rich horizons. These horizons (reddish in colour), already mentioned by et alii (2003), resemble Oxfordian and Aptian bentonites described in other sections of the Vocontian Basin (, 2002; & , 2003).
The La Charce section is well dated because of the important number of ammonoids, which comprise almost all of the macrofauna ( et alii, 1992; & , 1995; , 1996; & , 1999, and references therein). But other nektonic macrofossils are present: belemnites are common and there are a few nautiloids. Bivalves and gastropods are rare.
More than 15,000 ammonoids were collected in a hundred metre section (, 1996). The assemblage consists of six families. Most turnovers took place during the evolution of the Neocomitidae and Olcostephanidae. The ammonoid spectra are often dominated by: Haploceratidae, Bochianitidae, Phylloceratidae, and Lytoceratidae; their abundance indicates a deep-water palaeoenvironment (, 1996).
An ammonoid turnover occurred at the boundary of the Valanginian and Hauterivian, and has been interpreted as the response of nektonic organisms to eustatic and climatic changes ( et alii, 1992; & , 1995; , 1996). The biozonation reflects the evolution of the ammonoid fauna ( & , 1999). The zonal scheme of these authors has been adopted by the Lower Cretaceous Ammonite Working Group (= Group) of the IUGS Subcommission on Cretaceous Stratigraphy and is included in the Cretaceous Standard Zonation ( & (reporters) et alii, 2003; & (reporters) et alii, 2006).
(1874) defined the Hauterivian stage in the Hauterive area (Neuchâtel, Northwest Switzerland). For a long time this locality has been considered to be unsatisfactory as the stratotype due to the condensation of some parts of the section, its poor exposure and the scarcity of ammonoids. For most of the 20th century and thereafter and other French authors have investigated the expanded sections of the Vocontian Basin, where rich ammonoid faunas are recorded in the marl-limestone alternations of the hemi-pelagic successions.
As regards the Valanginian/Hauterivian boundary, the La Charce section is the best documented. It was proposed by (1977a, p. 125) as a candidate for the boundary stratotype. The IUGS retained this proposal during the Copenhagen meeting in 1983 ( et alii, 1984). Because no other supplementary section has been proposed in Spain, the Caucasus or in the Crimea (all are areas in which the Valanginian-Hauterivian is well-documented), during the Brussels meeting in 1995 ( et alii, 1996), the Hauterivian Working Group agreed to recommend the La Charce section as the global boundary stratotype for the base of the Hauterivian. The IUGS-ICS Subcommission on Cretaceous Stratigraphy recommended the La Charce section for the Hauterivian GSSP during the 32nd International Geological Congress at Florence in 2004 (, 2004; et alii, 2004). The members of the Hauterivian Working Group are currently preparing a formal proposal in accordance with this recommendation. The village of La Charce has agreed to preserve the section outcropping along the road.
The base of the Hauterivian in the Tethyan realm was traditionally defined by the first appearance of the index-species Acanthodiscus radiatus (,
1977a). This choice was recommended during the 1st
and 2nd international symposia on the Cretaceous Stage Boundaries (Copenhagen, 1983 and Brussels, 1995; et alii,
1984; et alii,
1996). Due to the scarcity of the index-species in
deep-water distal environments it was also suggested that the base of the Radiatus Zone
be made concomitant with the first appearance of the genus Acanthodiscus (A. radiatus and related species). This proposal was reported in the Geologic Time Scale of et alii
(2004); further discussion can be found in
(1997) and &
(1999). (1996, p. 263 and figure 22) also supports this definition of the Radiatus Zone. This author shows that the genus Acanthodiscus
is probably a biological species with a great variability in macroconchs (A. radiatus,
A. rebouli, A. vaceki, Leopoldia leopoldina), and in microconchs (Breistrofferella peyroulensis,
B. castellanensis and B. varappensis; ,
1996). Therefore, when Acanthodiscus is very rare or absent in deep-water palaeoenvironments, the recognition of the Radiatus Zone is possible using
the species of Breistrofferella that are also generally abundant on platforms.
In addition, the faunal assemblage of the Radiatus Zone is well characterized by other genera,
like Teschenites, Eleniceras, Olcostephanus, Spitidiscus and Oosterella (Fig. 2.1 ![]() ; ,
1996).
; ,
1996).
In accordance with these recommendations and considerations the Golden Spike of the Hauterivian stage (= the base of the Radiatus Zone) is placed at layer 189 of the La Charce type-section (Fig. 2.1 ![]() )
, which is the first occurrence of Acanthodiscus rebouli (,
1996). Bed 189 corresponds to bed 250 in the system
of numbering proposed by et alii
(1993).
)
, which is the first occurrence of Acanthodiscus rebouli (,
1996). Bed 189 corresponds to bed 250 in the system
of numbering proposed by et alii
(1993).
The chronologic age of the base of the Hauterivian is either 123 Ma (+6/-2 Ma, , 1994), 136.4 Ma (+/-2 Ma, et alii, 2004), 124.1 Ma (+/-0.4 Ma, et alii, 2006) or 133.9 Ma (+/-2 Ma, et alii, 2007). It is practically coincident with the base of subchron M10n ( et alii, 1989; et alii, 2007), or with chron M11n ( et alii, 2004).
Conventionally, the base of the Amblygonium Zone of the Boreal Realm is correlated with the base of the Radiatus Zone (, 1973; , 1983, 1993; et alii, 1996; et alii, 1998; et alii, 2004). Recent 87Sr/86Sr data suggest that the base of the Amblygonium Zone may correlate with the uppermost part of the Furcillata Zone (Upper Furcillata Subzone; et alii, 2007). This correlation is in agreement with similar proposals by other authors (, 1977b; et alii, 1981; , 1983; et alii, 1996; figure 6 in & reporters) et alii, 1999).
In southeastern France, Acanthodiscus is common or even relatively abundant in hemipelagic palaeoenvironments (Vivarais/Cévennes area, in et alii, 1989, 1996; , unpublished data) and in shallow-water, proximal environments (Provence platform, et alii, 1990; , 1993; , 1995; , 1996; Jura platform, & , 1989). Conversely, it is reported as rare in deeper-water, distal palaeoenvironments (Vocontian Basin, , 1996; Veveyse de Châtel area, Switzerland, et alii, 2003). A similar distribution is observed in other Tethyan and Atlantic basins (Betic Chains, Spain, , 1987; , 1995; Atlantic High Atlas, Morocco, , 1991).
In the Boreal realm, Acanthodiscus occurs mainly in the shallow-water facies of NW Germany, the Polish seaway and Crimea ( et alii, 1981). In North Germany, Acanthodiscus, which is generally rare in the Endemoceras beds (, 1973; , 1973), seems to be restricted to the Noricum Zone (, 1977b; , 1988) in deep-water environments, but it is recorded in the upper part of the Amblygonium Zone in shallow-water settings ( et alii, 1981; et alii, 1996). The presence versus absence of Acanthodiscus in the upper part of the Amblygonium Zone in the Boreal Realm may be controlled in large part by palaeoenvironmental factors (bathymetry and/or a proximal versus a distal location). The same pattern was observed in southeastern France ( et alii, 2007, and references therein).
Despite the fact that the presence or absence of Acanthodiscus is in part controlled by palaeoenvironmental factors, this genus has a wide palaeobiogeographic distribution and thus is a very good index for the base of the Hauterivian in Europe ( et alii, 1981; et alii, 2007), North Africa (Morocco, , 1991, 2004; , 2001; et alii, 2002), and Chili (, 2007). For further information on the palaeogeographic distribution of Acanthodiscus, see also the synonymies of type species of Acanthodiscus, Leopoldia, and Breistrofferella in (2005).
Silvia
Although there are several publications concerning Lower Cretaceous calcareous nannofossils, the biostratigraphic resolution of nannofossils across the Valanginian/Hauterivian boundary has increased little. In the published literature few datums are proposed to delineate the Valanginian/Hauterivian boundary. (1977) suggested using the FO (first occurrence) of Cretarhabdus loriei as a marker for the Early Hauterivian, though this particular occurrence proved to be much younger (Aptian). (1979, 1985) acknowledged the difficulty of using Cretarhabdus loriei due to problems in recognizing or differentiating this species from other species of Cretarhabdus. (1978, 1983) and (1976) proposed the LOs (last occurrences) of Diadorhombus rectus and Tubodiscus verenae to mark the top of the Valanginian; however, these species have much younger extinctions.
In terms of nannofossil biozones, the Valanginian/Hauterivian boundary (as defined by ammonoid fauna) falls within Biozone CC4a of & (1989) who modified the standard zonation of (1977). This zone is defined by the FAD (first appearance datum) of Eiffellithus striatus and the FAD of Litraphidites bollii. Biozone CC4 also corresponds to the NC4a Zone of (1978), as modified by et alii (1995). It is noteworthy that the last common occurrence of Tubodiscus verenae corresponds approximately with the FO of Eiffellithus striatus in the Late Valanginian.
Splits from the marly intervals were processed using standard preparation techniques and
were examined under a light microscope. All these samples were productive, with abundant calcareous nannofossil assemblages (Fig. 3.1 ![]() );
the number of species is usually high (about 80 species), preservation is moderate. Identification of specimens with the light microscope was not hampered by diagenetic etching and/or overgrowths of
calcite that can affect nannofloral assemblages (Fig. 3.2
);
the number of species is usually high (about 80 species), preservation is moderate. Identification of specimens with the light microscope was not hampered by diagenetic etching and/or overgrowths of
calcite that can affect nannofloral assemblages (Fig. 3.2 ![]() ).
).
Analysis of the complete and expanded Valanginian-Hauterivian sequence at La Charce has allowed a sequence of local events to be evaluated in relation to the established
European sequence. Nannoconus bucheri and Nannoconus wassalli
occur sporadically starting at the Callidiscus ammonoid Zone; their occurrence
is always rare and spotty. These taxa are more abundant and from the Loryi ammonoid
Zone upward are continuously recorded. Tubodiscus verenae and Tubodiscus jurapelagicus occur sporadically up to the
end of the Late Hauterivian
(Ligatus ammonoid Zone). Staurolithithes mitcheneri was first seen in bed 251 of et alii
(1993; Radiatus ammonoid Zone, Castellanensis Horizon). The LO of Eiffellithus windii
is in bed 270, and the FO of Diloma galiciense in bed 272
(Radiatus ammonoid Zone, Buxtorfi Horizon;
Fig. 2.1 ![]() ). These two species are rare but their stratigraphical ranges are reasonably consistent.
). These two species are rare but their stratigraphical ranges are reasonably consistent.
The first Litraphidites bollii was observed in bed 292 (Loryi
/Jeannoti ammonoid Zone) but its occurrence is common only upward from bed 296 in the
Nodosoplicatum/Variegatus ammonoid Zone (Fig. 3.1 ![]() ).
The last occurrence of Rhagodiscus dekaeneli is at the top of this zone
("non-nome" subzone). All these events are summarized in
Figure 3.1
).
The last occurrence of Rhagodiscus dekaeneli is at the top of this zone
("non-nome" subzone). All these events are summarized in
Figure 3.1 ![]() . The
value as markers in other geographical areas must be carefully evaluated. No important
originations or extinctions are coincident with the boundary
(Callidiscus / Radiatus ammonoid Zones). At La Charce, the nannofossil event that best approximates the
Valanginian/Hauterivian boundary is the LO of Eiffellithus windii.
. The
value as markers in other geographical areas must be carefully evaluated. No important
originations or extinctions are coincident with the boundary
(Callidiscus / Radiatus ammonoid Zones). At La Charce, the nannofossil event that best approximates the
Valanginian/Hauterivian boundary is the LO of Eiffellithus windii.
Calcareous nannofloras at La Charce have a predominantly low-latitude (Tethyan) affinity (common Cruciellipsis cuvillieri, Speetonia colligata and Calcicalathina oblongata) though Nannoconids, which are known to prefer low-latitude, warm surface waters, and hemipelagic settings (, 1976; , 1992; , 1994; & , 2000) are very rare. Taxa more commonly associated with higher latitudes such as Sollasites spp., Crucibiscutum salebrosum, Corollithion silvaradion, are consistently present at La Charce although in much lower quantities than in boreal sites. This is due to the northern Tethyan location of the Vocontian Basin, which acted as a "gateway" to boreal domain of northwestern Europe.
Fabienne
The Aptian-Albian interval of the Vocontian Basin is represented
by the Marnes Bleues Formation. It is about 800 metres of rather homogeneous dark-blue marls and
shales with a grayish cast. In southeastern France the OAE 1d level is called the interval (,
1988). Nine laminated horizons containing 1 to 2% wt%TOC (Total Organic Carbon) have been identified in the central part of the Vocontian
Basin, NW of Sisteron (,
1988;
Fig. 1.2 ![]() ).
).
Near Blieux (Alpes de Haute-Provence) near the southern margin of the Vocontian Basin ( et alii,
2003; et alii,
2005;
Fig. 1.2 ![]() )
in the 38 metre succession of dark colored marls intercalated with centimetric-thick
lighter grey marls (Figs. 4.1
)
in the 38 metre succession of dark colored marls intercalated with centimetric-thick
lighter grey marls (Figs. 4.1 ![]() and 4.2
and 4.2 ![]() )
there are changes upward in the abundance of calcareous nannofossils and in the composition
of their assemblages, along with varying proportions of macrofossils, assemblages
of ichnofossils and in the degree of bioturbation. Palaeogeographically this section
is in a key position to record palaeoenvironmental changes involving both proximal areas (platform environments) and the pelagic realm (open marine), and is rich in nektonic/benthic macrofauna. Because of the relatively proximal position of the Blieux section, the interval is
devoid of the typical laminated black-shale horizons in the central part of the Vocontian
Basin (, 1995-1997).
)
there are changes upward in the abundance of calcareous nannofossils and in the composition
of their assemblages, along with varying proportions of macrofossils, assemblages
of ichnofossils and in the degree of bioturbation. Palaeogeographically this section
is in a key position to record palaeoenvironmental changes involving both proximal areas (platform environments) and the pelagic realm (open marine), and is rich in nektonic/benthic macrofauna. Because of the relatively proximal position of the Blieux section, the interval is
devoid of the typical laminated black-shale horizons in the central part of the Vocontian
Basin (, 1995-1997).
Calcareous nannofossils are well preserved and are predominantly
heterococcoliths, but holococcoliths are also present. The range in the number
of species fluctuates from 32 to 50 per sample. As pelagic carbonate production is limited, the carbonate fraction is derived mainly from the tests of nektonic/benthic organisms,
but in part may be allochthonous (export to the basin from adjacent carbonate-platforms). There
is little organic carbon in the interval, and it is not associated with high
productivity in surface waters. Organic matter is mainly terrigenous in origin and its
presence is due to: 1) dysoxic conditions that preserved it and 2) a weak input of allochthonous carbonates. Eustatic fluctuations strongly influenced the changes in nannofossil and macrofaunal abundances.
In the interval there are also distinctive patterns in nannofossil assemblages and macrofaunal
abundance (Figs. 4.2 ![]() and 4.3
and 4.3 ![]() )
that reflect changes in trophic levels. Low diversity in nannoplankton assemblages
and very abundant macrofaunas suggest that mesotrophic conditions prevailed in the lower part of
the interval (Fig.
4.3a
)
that reflect changes in trophic levels. Low diversity in nannoplankton assemblages
and very abundant macrofaunas suggest that mesotrophic conditions prevailed in the lower part of
the interval (Fig.
4.3a ![]() ).
In the upper part there is a greater diversity in nannofossil assemblages, more
abundant ammonoids and a lesser amount of benthic macrofauna (Fig.
4.3b
).
In the upper part there is a greater diversity in nannofossil assemblages, more
abundant ammonoids and a lesser amount of benthic macrofauna (Fig.
4.3b ![]() ). This
divergence might be the result of climatic changes associated with periods of increased precipitation and
runoff during the deposition to the lower part of the interval,
and to a period of drier conditions in the upper part of this interval. The work of et alii
(2003) and et alii
(2005) shows that the deposits did not
indicate an eutrophication of marine surface waters that occurs with the expansion of an oxygen-minimum zone.
). This
divergence might be the result of climatic changes associated with periods of increased precipitation and
runoff during the deposition to the lower part of the interval,
and to a period of drier conditions in the upper part of this interval. The work of et alii
(2003) and et alii
(2005) shows that the deposits did not
indicate an eutrophication of marine surface waters that occurs with the expansion of an oxygen-minimum zone.
A high-resolution quantitative analysis of the several microfossil groups and
of the bulk-rock stable isotope data was made on the black shalesof the main
interval in the central part of the Vocontian
Basin (Col de Palluel section, IGN map French Série Bleue 1:25,000 Rosans number 3239
Ouest, Lambert III Zone coordinates 853.750; 3238.425,
Fig. 1.1 ![]() ) by et alii
(2005). This work quantified the absolute abundance of calcareous nannofossils (Fig. 4.4
) by et alii
(2005). This work quantified the absolute abundance of calcareous nannofossils (Fig. 4.4 ![]() ),
using the settling method of et alii
(1999). Nannofossils are well-to-moderately preserved at Col de
Palluel. Like the work of et alii
(2003), this study suggests that the accumulation of organic matter was controlled by preservation rather than by
an increase in its productivity in the photic zone. Paleoclimatic and oceanographic changes
caused by moonsoonal activity were identified. In the interval,
the black shales were laid down by surface waters in a warm and humid climate
under oligotrophic conditions while marlstones were deposited under relatively cool and arid conditions
that favored increased productivity of carbonates.
),
using the settling method of et alii
(1999). Nannofossils are well-to-moderately preserved at Col de
Palluel. Like the work of et alii
(2003), this study suggests that the accumulation of organic matter was controlled by preservation rather than by
an increase in its productivity in the photic zone. Paleoclimatic and oceanographic changes
caused by moonsoonal activity were identified. In the interval,
the black shales were laid down by surface waters in a warm and humid climate
under oligotrophic conditions while marlstones were deposited under relatively cool and arid conditions
that favored increased productivity of carbonates.
Fabienne
The GSSP for the base of the Cenomanian is the Mont Risou section, Hautes-Alpes (IGN map French Série Bleue
1:25,000 Rosans number 3239 Ouest, Lambert II Zone coordinates 852.725; 1937.625,
Figs. 1.1 ![]() and 4.4
and 4.4 ![]() ). This section exposes a continuously accessible succession of nearly 250 m, from the interval in the upper part of the Marnes Bleues Formation to Lower Cenomanian marly limestones and marls, with no evidence of sedimentary breaks or condensation ( et alii,
1996, Fig. 5.1
). This section exposes a continuously accessible succession of nearly 250 m, from the interval in the upper part of the Marnes Bleues Formation to Lower Cenomanian marly limestones and marls, with no evidence of sedimentary breaks or condensation ( et alii,
1996, Fig. 5.1 ![]() ). Ammonites, planktonic foraminifera, and calcareous nannofossils are abundant throughout the succession and allow accurate biostratigraphy (Fig. 5.1
). Ammonites, planktonic foraminifera, and calcareous nannofossils are abundant throughout the succession and allow accurate biostratigraphy (Fig. 5.1 ![]() ). The base of the Cenomanian stage lies 36 m below the top of the Marnes Bleues Formation and
is chosen at the first occurrence of the planktonic foraminifer Rotalipora globotruncanoides.
). The base of the Cenomanian stage lies 36 m below the top of the Marnes Bleues Formation and
is chosen at the first occurrence of the planktonic foraminifer Rotalipora globotruncanoides.
Preservation of calcareous nannofossils is moderate throughout the sequence and the nannofloral assemblages are highly diverse (153 taxa). Biscutum ellipticum, Rhagodiscus achlyostaurion, Tranolithus orionatus, Watznaueria barnesiae and Watznaueria manivitiae are common to abundant. Holococcoliths are unusually common and sometimes well-preserved in particular levels ( in et alii, 1996).
The carbon-isotope curve has a large peak made up of four distinct spikes (A to D, Fig. 5.1 ![]() ). Temperatures of sea-surface water deduced from the oxygen isotope data are comprised between 26 and 27°C. A slight cooling of about 1°C
distinguishes the earliest Cenomanian ( et alii,
1996, Fig. 5.1
). Temperatures of sea-surface water deduced from the oxygen isotope data are comprised between 26 and 27°C. A slight cooling of about 1°C
distinguishes the earliest Cenomanian ( et alii,
1996, Fig. 5.1 ![]() ).
).
Bernard
Today we will analyse the sedimentary evolution of the Cenozoic in the Barrême region. This area is in the second of the foreland basins formed to the west of the Alps during the collision Adria-Europe. The first foreland basin formed during Late Cretaceous times in conjunction with the Pyrénées-Provençal tectonic phase (e.g., deposition of the "Flysh à Helminthoïdes", Campanian-Maastrichtian). The second foreland basin developed in Late Eocene-Early Oligocene (Nummulitic Sea) during the second Alpine tectonic phase, and the third was present during the Miocene in connection with the late Alpine tectonic phase.
Foreland basins are intrinsically related to continent-to-continent collision and the formation of Alpine-type mountain chains. They are narrow, elongated basinal troughs located at the periphery of a mountain chain during its formation. They receive the products of the erosion of the rising chain, and are thus filled mainly by siliciclastic material. They record the major tectonic events that occur during a continent-to-continent collision.
Basically, mountain chains forming during such a collision can be
regarded as a large accretion prism of continental crust. During intensive tectonic activity in the chain, accretion is responsible for a thickening of the crust
(e.g., ~50 km for the Alps, ~70 km for the Himalaya) of the lithosphere.
In isostatic response to this thickening, the mountain chain subsides (Fig. 6.1a ![]() ). Consequently,
along the periphery of the mountain chain subsidence is also important, and creates on both sides of the chain a foreland
basin in which continental or marine sediments are deposited and preserved. In response to the overcharge of the mountain chain, the continental crust (and lithosphere) forms a bulge some few
hundred kms away from the chain (Fig. 6.1a
). Consequently,
along the periphery of the mountain chain subsidence is also important, and creates on both sides of the chain a foreland
basin in which continental or marine sediments are deposited and preserved. In response to the overcharge of the mountain chain, the continental crust (and lithosphere) forms a bulge some few
hundred kms away from the chain (Fig. 6.1a ![]() ). This
rise accounts for the fact that foreland basins are asymmetrical and narrow.
). This
rise accounts for the fact that foreland basins are asymmetrical and narrow.
Because it can be seen as an accretional prism of continental crust, the mountain chain in formation will not only thicken but also enlarge laterally, thus
causing the foreland basin
to migrate away from the zone of accretion at the outer limit of the mountain chain (Fig.
6.1b ![]() ).
Consequently, the oldest sediments deposited in a foreland basin are the closest to the chain and can,
as the basin migrates, become involved in the accretion prism. The youngest sediments are deposited far from the chain, and generally are not deformed.
).
Consequently, the oldest sediments deposited in a foreland basin are the closest to the chain and can,
as the basin migrates, become involved in the accretion prism. The youngest sediments are deposited far from the chain, and generally are not deformed.
The Barrême region was very close to the developing Alpine chain. Consequently, this area records both foreland dynamics (the accommodation space created by subsidence due to thickening of the Alps during intense tectonic pulses) and Alpine dynamics (folding of the sediments deposited in the foreland basin whenever accommodation space was available). These two mechanisms (subsidence and folding) underpin the Cenozoic geological history of the Barrême region.
The Barrême syncline, oriented N-S, exposes the Upper Eocene-Oligocene sediments deposited in the inner part of the Nummulitic-Molasse foreland basin, directly on
previously folded and eroded Mesozoic "deep-water" sediments
(Figs. 1.1 ![]() and 1.2
and 1.2 ![]() ). Consequently, the marine transgression of the Nummulitic Sea in the Late Eocene invaded complex paleoreliefs of different ages (Early to Late Cretaceous). The first sediments attributed to the Cenozoic are
continental alluvial deposits that develop from south to north (Argens Conglomerates; Fig. 6.2
). Consequently, the marine transgression of the Nummulitic Sea in the Late Eocene invaded complex paleoreliefs of different ages (Early to Late Cretaceous). The first sediments attributed to the Cenozoic are
continental alluvial deposits that develop from south to north (Argens Conglomerates; Fig. 6.2 ![]() ) and are present only locally,
thus suggesting that river drainage was already controlled by pre-existing N-S folds. On these alluvial facies (when present), or directly on folded Mesozoic rocks is the Nummulitic Limestone (Fig. 6.2
) and are present only locally,
thus suggesting that river drainage was already controlled by pre-existing N-S folds. On these alluvial facies (when present), or directly on folded Mesozoic rocks is the Nummulitic Limestone (Fig. 6.2 ![]() ), which
consists of shallow-marine bioclastic sediments (nummulites, bivalves mainly oysters) but can locally contain
fluvial conglomerates deposited in the shallow Nummulitic Sea. As transgression
continued deeper-water marls accumulated in the Nummulitic Sea, forming two sub-units, the Lower and the Upper Nummulitic Marls, separated by the Ville Sandstones that are turbiditic lobe deposits. Sedimentary structures (flute casts, prod casts, current ripples) in the
turbidites of the Ville Sandstones indicate a south to north direction for the turbidity currents, thus implying a southern source of the siliciclastic material (Corsica-Sardinia block).
), which
consists of shallow-marine bioclastic sediments (nummulites, bivalves mainly oysters) but can locally contain
fluvial conglomerates deposited in the shallow Nummulitic Sea. As transgression
continued deeper-water marls accumulated in the Nummulitic Sea, forming two sub-units, the Lower and the Upper Nummulitic Marls, separated by the Ville Sandstones that are turbiditic lobe deposits. Sedimentary structures (flute casts, prod casts, current ripples) in the
turbidites of the Ville Sandstones indicate a south to north direction for the turbidity currents, thus implying a southern source of the siliciclastic material (Corsica-Sardinia block).
The Nummulitic deposits, both limestone and marls, were folded
in some localities before the deposition of the Clumanc Conglomerates (Fig. 6.2 ![]() ). These conglomerates contain
Alpine-derived gravels. They demonstrate that the centre of the Alps (oceanic crust and deep-ocean sediments,
i.e. serpentinites and radiolarites) was already exposed some 32 Ma ago.
Here at Clumanc the conglomerates are remarkable because they are formed by successive flows
of phreato-magmatic debris that followed the river system to be deposited at the mouth of a
delta on the shore of the Nummulitic Sea. The genesis of phreato-magmatic flows is
demonstrated by their andesitic matrix and the presence of andesite gravels. A
rapid forceful discharge of material is evinced by the presence of thick (3080 cm) layers containing mud boulders that testify
to a mass-flow origin for these deposits.
). These conglomerates contain
Alpine-derived gravels. They demonstrate that the centre of the Alps (oceanic crust and deep-ocean sediments,
i.e. serpentinites and radiolarites) was already exposed some 32 Ma ago.
Here at Clumanc the conglomerates are remarkable because they are formed by successive flows
of phreato-magmatic debris that followed the river system to be deposited at the mouth of a
delta on the shore of the Nummulitic Sea. The genesis of phreato-magmatic flows is
demonstrated by their andesitic matrix and the presence of andesite gravels. A
rapid forceful discharge of material is evinced by the presence of thick (3080 cm) layers containing mud boulders that testify
to a mass-flow origin for these deposits.
In the Clumanc area, the Nummulic Sea includes a succession of 3 phreato-magmatic episodes that alternate with "normal" nummulitic marls to fill this part of the basin. The Barrême syncline (still in formation) then served as a river valley. This river formed a new delta in St-Lions, some 7 km south of Clumanc. The progradation of the delta from Clumanc to St-Lions attests that the river flowed from north to south at that time.
The Red Molasse that overlies the Clumanc and St-Lions conglomerates is slightly discordant, suggesting that tectonic movements occurred after deposition of the Clumanc/St-Lyons
conglomerates ended, and before (or at the beginning of) the deposition of the Red Molasse. Alluvial fan deposits
comprise this new sedimentary unit. In the Barrême syncline, the Red Molasse is dominantly argillaceous (alluvial plain red-clays) interrupted by
small channels, which are infilled by conglomerates. The conglomerates are for
the most part of local origin (Mesozoic carbonates from the flanks of the Barrême syncline). Southwards, in the Senez region (some 6 km farther from
Barrême; Fig. 1.2 ![]() ), the Red Molasse is dominantly conglomeratic, and clays are much
rarer. This implies that the alluvial fan developed from south to north.
), the Red Molasse is dominantly conglomeratic, and clays are much
rarer. This implies that the alluvial fan developed from south to north.
Above the Red Molasse and concordant with it is the Grey Molasse. Its deposits are mainly argillaceous (whitish to light-greenish alluvial plain sediments), with a few sandstone levels and common palustrine-lacustrine limestones. The evolution from the Red to the Grey Molasse is thought to reflect a flattening of the relief in the back-land due to increased subsidence. The change in colour from one formation to the other probably has a climatic origin.
This section, along the road from Les Sauzeries Basses to La Poste (1 km to the south-east; Fig. 6.3 ![]() ),
exposes a continuous sedimentary succession, approximately 150-200 m thick that
occupies the interval between the Nummulitic Limestone and the Clumanc Conglomerates. The Nummulitic Limestone in this area was deposited in unconformity
on the Blue Marls (Fig. 6.4a
),
exposes a continuous sedimentary succession, approximately 150-200 m thick that
occupies the interval between the Nummulitic Limestone and the Clumanc Conglomerates. The Nummulitic Limestone in this area was deposited in unconformity
on the Blue Marls (Fig. 6.4a ![]() ),
which are earliest Albian in age to the north and latest Late Aptian to the south. Two plurimetric limestone beds separated by nummulitic marls
represent the Nummulitic Limestone here. The limestones are fine-grained
bioclasts, with abundant oyster fragments and locally with nummulites. The Lower Nummulitic Marls deposited above
the limestones have a thickness of 80m to about 100m. The Ville Sandstones are a succession of fine-grained turbiditic lobes. The turbidites typically
have plane-parallel laminations with few occurrences of current ripples. An erosional
contact at the base of the turbidites is common. Looking south from the road, three units of the Clumanc Conglomerates can be
seen, as intercalations into the Upper Nummulitic Marls. To the southeast, below the Clumanc castle (Stop 2), these 3 units are amalgamated. A geometry of the deltaic system
of the Clumanc Conglomerates can be reconstructed by geological mapping.
),
which are earliest Albian in age to the north and latest Late Aptian to the south. Two plurimetric limestone beds separated by nummulitic marls
represent the Nummulitic Limestone here. The limestones are fine-grained
bioclasts, with abundant oyster fragments and locally with nummulites. The Lower Nummulitic Marls deposited above
the limestones have a thickness of 80m to about 100m. The Ville Sandstones are a succession of fine-grained turbiditic lobes. The turbidites typically
have plane-parallel laminations with few occurrences of current ripples. An erosional
contact at the base of the turbidites is common. Looking south from the road, three units of the Clumanc Conglomerates can be
seen, as intercalations into the Upper Nummulitic Marls. To the southeast, below the Clumanc castle (Stop 2), these 3 units are amalgamated. A geometry of the deltaic system
of the Clumanc Conglomerates can be reconstructed by geological mapping.
A perpective of the hill on which the Clumanc castle was
constructed will be examined at this stop (Fig. 6.4b ![]() ). The three amalgamated units of the Clumanc Conglomerates constitute the
hilltop. They rest unconformably on the previously folded synclinal Nummulic Limestone and Marls. Finally, both the Nummulitic Limestone and Marls and the Clumanc Conglomerates were again folded to form an anticline. This
sequence of events is a good illustration of the tectonic activity in the Alps
as it was recorded in the Barrême syncline, for the sequence described is
repeated generally: 1) subsidence creates a depressed space to accommodate marine or continental sediments; 2) these sediments are folded; 3) a new phase of accommodation is created by subsidence, and
more sediments are deposited; 4) folding, etc.
Note that both folding and the creation of accommodation can occur at the same time.
). The three amalgamated units of the Clumanc Conglomerates constitute the
hilltop. They rest unconformably on the previously folded synclinal Nummulic Limestone and Marls. Finally, both the Nummulitic Limestone and Marls and the Clumanc Conglomerates were again folded to form an anticline. This
sequence of events is a good illustration of the tectonic activity in the Alps
as it was recorded in the Barrême syncline, for the sequence described is
repeated generally: 1) subsidence creates a depressed space to accommodate marine or continental sediments; 2) these sediments are folded; 3) a new phase of accommodation is created by subsidence, and
more sediments are deposited; 4) folding, etc.
Note that both folding and the creation of accommodation can occur at the same time.
A short stop here is devoted to the alluvial fan facies of the Red Molasse. An outcrop along road D19 epitomizes the typical characteristics of alluvial fans: 1) ephemeral conglomeratic channels filled by local material; 2) sheet flood deposits that coarsen upward, thickening-upward silt-to-sand sediments; and 3) flooding clays deposited at considerable distances from the channels during the partial or entire flooding of the fan. In this outcrop the predominance of clays indicates that this area was in a distal position on the alluvial fan.
The authors wish to warmly thank Bruno for his precious help in editing this guidebook and for his useful advices. Special thanks are due Nestor for language corrections and improved readability of the text.
J.L. & J.A. (1989).- 20. Cretaceous calcareous nannofossil biostratigraphy of sediments recovered from the Galicia margin, ODP Leg 103.- In: G., E.L. & (eds.), Galicia Margin, Sites 637-641.- Proceedings of the Ocean Drilling Project, Scientific Results, College Station, vol. 103, p. 293-348.
H. & M. (1993).- Structure and Mesozoic-Cenozoic evolution of the South-East France Basin (SFB).- Géologie Alpine, Grenoble, (série spéciale "Colloques et Excursions"), n° 3, p. 53-58.
F. (1984).- 2.7. Chaînes subalpines (Jurassique supérieur : Malm). In: D.S. (ed.), Synthèse géologique du Sud-Est de la France.- Mémoire du Bureau des Recherches Géologiques et Minières, Orléans, n° 125, vol. 1 (Stratigraphie et paléogéographie), p. 255-257.
F., M., B. & M. (2002).- Biostratigraphie par ammonites, au passage Valanginien-Hauterivien, dans le Haut Atlas occidental (région d'Agadir, Maroc). In: C. & P. (eds.), Strati 2002 : 3ème congrès français de stratigraphie, Lyon (8-10 juillet 2002).- Documents des Laboratoires de Géologie de Lyon, p. 22.
F. & S. (1995).- Le Valanginien-Hauterivien basal du bassin vocontien et de la bordure provençale: zonation et corrélations.- Comptes Rendus de l'Académie des Sciences, Paris, t. 320, (série II), n° 10, p. 985-992.
G. (1993).- L'évolution de la marge Nord-Est provençale (Arc de Castellane) du Valanginien moyen à l'Hauterivien à travers l'analyse biostratigraphique des séries de la région de Peyroules: séries condensées, discontinuités et indices d'une tectogenèse distensive. Paléobiologie.- Annales du Muséum d'Histoire Naturelle de Nice, t. X, 240 p.
B., J., M., F., O., L., M., L., S., G., F., O., M., D., V., O., C., M., S., F., G., A., P., B. & G. (2003).- Les bentonites du Crétacé inférieur du SE de la France. In: Bassins crétacés de France et d'Europe occidentale, Paris (6-7 Novembre 2003).- Société Géologique de France, Paris, p. 8-9.
T., J.M., M.B., P.F., J., F., F. & F. (1984).- Cretaceous stage boundaries proposals.- Bulletin of the Geological Society of Denmark, Copenhagen, vol. 33, p. 3-20.
A., J., K., J.O., C. & J. (2005).- Reconstruction of short-term palaeoceanographic changes during the formation of the Late Albian 'Niveau ' black shales (Oceanic Anoxic Event 1d, SE France).- Journal of the Geological Society, London, vol. 162, p. 101-114.
T.J., R.M., W.V. & H.R. (1995).- An integrated Cretaceous microfossil biostratigraphy. In: W.A., D.V., M.-P. & J. (eds.), Geochronology, time scales, and global stratigraphic correlation.- Society of Economic Paleontologists and Mineralogists (SEPM) Special Publication, Tulsa, n° 54, p. 65-79.
J.G. (1988).- Épisodes de sédimentation riche en matière organique dans les Marnes Bleues d'âge aptien et albien de la partie pélagique du bassin vocontien.- Bulletin de la Société Géologique de France, Paris, (Série 8), t. IV, n° 2, p. 349-356.
J.G. (1995).- L'Aptien et l'Albien de la Fosse vocontienne (des bordures au bassin). Évolution de la sédimentation et enseignements sur les événements anoxiques.- Thèse Doctorat de l'Université François Rabelais, Tours, 564 p.; Publication de la Société Géologique du Nord, Lille, t. 25 (1997), 614 p.
L.G. (1995).- Les formations à ammonites du Crétacé inférieur dans le Sud-Est de la France (Berriasien à Hauterivien): biostratigraphie, paléontologie et cycles sédimentaires.- Unpublished Ph.D. thesis, Muséum National d'Histoire Naturelle, Paris, 398 p.
L.G., J.-P., H. & G. (1996).- The Lower Cretaceous of the South Vocontian Basin and margins.- Géologie Alpine, Grenoble, (Mémoire H.S.), t. 20, p. 383-399.
L.G., J.-P., É. & J. (1993).- Le cadre stratigraphique du Valanginien supérieur et de l'Hauterivien du Sud-Est de la France: définition des biochronozones et caractérisation de nouveaux biohorizons.- Géologie Alpine, Grenoble, t. 68, p. 13-56.
R., J., M. & B. (2003).- Le Crétacé inférieur de la Veveyse de Châtel (Ultrahelvétique des Préalpes externes; canton de Fribourg, Suisse).- Revue de Paléobiologie, Genève, vol. 22, n° 1, p. 1-174.
R. & J.-P. (1989).- Les ammonites de l'Hauterivien jurassien: révision des faunes de la région du stratotype historique de l'étage hauterivien.- Mémoire de la Société neuchâteloise de Sciences naturelles, vol. 11, p. 101-147.
M. (1987).- Los ammonites del Valanginense del sector oriental de las Cordilleras Béticas (SE de España).- Tesis Doctoral de la Universidad de Granada, 294 p.
P., S., C., E., G. & M. (1980).- Fluctuation des paramètres du milieu marin dans le domaine vocontien (France Sud-Est) au Crétacé inférieur: mise en évidence par l'étude des formations marno-calcaires alternantes.- Bulletin de la Société Géologique de France, Paris, (Série 7), t. XXII, n° 5, p. 735-744.
L. (2002).- Litho-, bio- et chronostratigraphie comparées dans le Bassin Vocontien, à l'Aptien.- Unpublished Ph.D. thesis, Université des Sciences et Techniques Lille 1, Lille, 516 p.
S. (1984).- 2.8 Causses -Bordure vivaro-cévénole s.l. (Jurassique moyen : Dogger). In: D.S. (ed.), Synthèse géologique du Sud-Est de la France.- Mémoire du Bureau des Recherches Géologiques et Minières, Orléans, n° 125, vol. 1 (Stratigraphie et paléogéographie), p. 205-211.
S., P., G., H., R., P.& C. (1989).- Notice explicative, Carte Géologique de France 1/50000, feuille Bessèges (888).- BRGM, Orléans, 115 p.
S., R., B., G., G., P.& B. (1996).- Notice explicative, Carte Géologique France 1/50000, feuille Aubenas (865).- BRGM, Orléans, 170 p.
E. (1990).- Calcareous nannofossil biostratigraphy of some Bajocian sections from the Digne area (SE France).- Memorie Descrittive della Carta Geologica d'Italia, Roma, vol. XL, p. 237-255.
E. (1994).- Nannofossils and superplumes: the Early Aptian "nannoconid crisis".- Paleoceanography, Washington, vol. 9, n° 3, p. 483-501.
E. (2006).- The first 150 million years history of calcareous nannoplankton: Biosphere-geosphere interactions.- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, vol. 232, n° 2-4, p. 237-250.
M. (1991).- Le Valanginien de l'Atlas atlantique (Maroc) : stratigraphie et ammonitofaune.- Unpublished Ph.D. thesis, Université Cadi Ayyad, Faculté des Sciences, Marrakech, 153 p.
M. (2004).- Les ammonites néocomiennes dans l'Atlas atlantique (Maroc). Biostratigraphie, paléontologie, paléobiogéographie et paléoécologie.- Strata, Toulouse, vol. 43, (série 2 : mémoires), 223 p.
S. (1990).- Post-field comments. Mesozoic eustacy record on western Tethyan margins. Post-meeting field-trip in the Vocontian Trough.- Publication de l'Association des Sédimentologistes français, vol. 12, p. 121-140.
S. (1991).- Une alternative au modèle de stratigraphie séquentielle d'Exxon : la modulation tectono-climatique des cycles orbitaux.- Géologie Alpine, Grenoble, (Mémoire H.S.), t. 18, p. 47-99.
S. & C. (1995).- Faciès de dépôt et stratigraphie séquentielle des calcaires bajociens du Jura méridional.- Documents des Laboratoires de Géologie de Lyon, n° 133, 96 p.
S., J., J.L. & F. (1989).- Inversions magnétiques et cycles sédimentaires: un premier résultat dans le Crétacé de la fosse vocontienne (Sud-Est de la France).- Comptes Rendus de l'Académie des Sciences, Paris, t. 308, (série II), n° 2, p. 773-780.
N., X., Parize O., L.G. & P.Y. (2006).- Lower Cretaceous stage durations combining radiometric data and orbital chronology: Towards a more stable relative time scale?- Earth and Planetary Science Letters, Amsterdam, vol. 246, n° 3-4, p. 407-417.
C. (1984).- Bioturbation des sédiments pélagiques du Crétacé inférieur dans le bassin vocontien (Chaînes subalpines méridionales, France).- Geobios, (Mémoire spécial), n° 8, p. 205-214.
C. & E. (1987).- The use of burrows to detect compaction and sliding in fine-grained sediments: an example from the Cretaceous of SE France.- Sedimentology, Oxford, vol. 34, n° 4, p. 585-593.
A.S., W.J., J.A, M. & B.E. (1996).- The Late Albian to Early Cenomanian succession at Mont Risou near Rosans (Drôme, SE France): an integrated study (ammonites, inoceramids, planktonic foraminifera, nannofossils, oxygen and carbon isotopes).- Cretaceous Research, London, vol. 17, n° 5, p. 515-606.
M., C., A.T.C., L., I. & J.R. (2002).- Life-cycle associations involving pairs of holococcolithophorid species: intraspecific variation or cryptic speciation?- European Journal of Phycology, London, vol. 37, n° 4, p. 531-550.
M., J., J.O., J. & J.R. (1999).- Calibration of the random settling technique for calculation of absolute abundances of calcareous nannoplankton.- Micropaleontology, New York, vol. 45, n° 4, p. 437-442.
F. (1995).- Recherche des périodicités astronomiques et des fluctuations du niveau marin à partir de l'étude du signal carbonaté des séries pélagiques alternantes. Application au crétacé inférieur du Sud Est de la France (bassin vocontien), de l'Atlantique central (Site 534 DSDP) et du golfe du Mexique (Site 535 DSDP).- Documents des Laboratoires de Géologie de Lyon, n° 134, 279 p.
F., D., F., S., B. & O. (2003).- Minor changes in surface water fertility across the oceanic anoxic event 1d (latest Albian, SE France) evidenced by calcareous nannofossils.- International Journal of Earth Sciences (Geologische Rundschau), Stuttgart, vol. 92, n° 2, p. 267-284.
F., B., E., & V. (2006).- Paleoenvironmental controls on the morphology and abundance of the coccolith Watznaueria britannica (Late Jurassic, southern Germany).- Marine Micropaleontology, Amsterdam, vol. 60, n° 3, p. 205-225.
B. (2007).- Impact des variations paléoclimatiques sur la sédimentation carbonatée au Valanginien.- Unpublished Ph.D. thesis, Université de Lyon, 154 p.
A. (2001).- A review of the broad pattern of Jurassic sea-level changes and their possible causes in the light of current knowledge.- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, vol. 167, n° 1-2, p. 2337.
S., H. & L. (1999).- C-isotope stratigraphy, a calibration tool between ammonite and magnetostratigraphy: the Valanginian-Hauterivian transition.- Geologica Carpathica, Bratislava, vol. 50, n° 1, p. 91-96.
P.J. (1995).- Ammonite distribution around the Hauterivian-Barremian boundary along the Río Argos (Caravaca, SE Spain).- Géologie Alpine, Grenoble, (Mémoire H.S.), t. 20, p. 219-277.
P.J. & S. (reporters), M.B., P., M., F., R., M., C., J., A., I., N., P.F., P., Z., J. & M.G.E. (2003).- Report on the 1st International Workshop of the IUGS Lower Cretaceous Ammonite Working Group, the ' Group' (Lyon, 11 July 2002).- Cretaceous Research, London, vol. 24, n° 1, p. 89-94 ; erratum, vol. 24, n° 6, p. 805.
T., G., F., P.C. de & F. (1998).- The north atlantic cycle: an overview of 2nd-order transgressive/regressive facies cycles in the lower Cretaceous of western Europe.- In: P.C. de, J., T. & P.R. (eds.), Mesozoic and Cenozoic sequence stratigraphy of European basins.- Society of Economic Paleontologists and Mineralogists (SEPM) Special Publication, Tulsa, n° 60, p. 397-409.
N.M.M. & A. (2002).- Extinction and renewal patterns among Tethyan belemnites in the Verrucosum Subzone (Valanginian) of southeast France.- Cretaceous Research, London, vol. 23, n° 4, p. 509-522.
E. (1984).- Écailles de l'Arc de Castellane (Jurassique inférieur : Lias). In: D.S. (ed.), Synthèse géologique du Sud-Est de la France.- Mémoire du Bureau des Recherches Géologiques et Minières, Orléans, n° 125, vol. 1 (Stratigraphie et paléogéographie), p. 157-158.
E. (1973).- The Valanginian and Hauterivian stages in northwest Germany.- In: R. & P.F. (eds.), The Boreal Lower Cretaceous.- Geological Journal, Liverpool, special issue, n° 5, p. 327-344.
E., P.F. & J.-P. (1981).- Ammonites of Tethyan ancestry in the early Lower Cretaceous of north-west Europe.- Palaeontology, vol. 24, p. 251-311.
W.J., A.S., J.A. & M. (2004).- The Global Boundary Stratotype Section and Point (GSSP) for the base of the Cenomanian Stage, Mont Risou, Hautes-Alpes, France.- Episodes, Beijing, vol. 27, n° 1, p. 21-32.
J. (1997).- Ammonite stratigraphy for the Valanginian and Lower Hauterivian of the Mediterranean region.- Tethyan and Boreal Cretaceous Correlation - Newsletter, n° 9, p. 6-15.
J. (2005).- Lower Cretaceous Ammonites I, Perisphinctaceae 1: Himalayitidae, Olcostephanidae, Holcodiscidae, Neocomitidae, Oosterellidae. In: W. (ed.), Fossilium Catalogus I: Animalia.- Backhuys Publishers (Leiden), part 139, 484 p.
S. & J. (2000).- Coccoliths under stress? Lower Cretaceous calcareous nannofossil assemblages of nearshore and pelagic settings (NW Africa).- Journal of Nannoplankton Research, London, vol. 22, n° 2, p. 171.
M. (1984).- Mesozoic evolution of the western Alps.- Annales Geophysicae, vol. 2, n° 2, p. 171-172.
M. (1985).- Structuration jurassique des Alpes occidentales et palinspastiques de la Téthys ligure.- Bulletin de la Société Géologique de France, Paris, (Série 8), t. I, n° 1, p. 126-137.
J.M., N.M.M., S., M.J., M.F. & B. van de (2007).- Palaeotemperatures, polar ice-volume, and isotope stratigraphy (Mg/Ca, δ18O, δ13C, 87Sr/86Sr): The Early Cretaceous (Berriasian, Valanginian, Hauterivian).- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, vol. 248, n° 3-4, p. 391-430.
F. (1992).- Enregistrement de l'eustatisme par les foraminifères dans les séquences de dépôt du Crétacé inférieur du bassin vocontien (Sud-Est de la France).- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, vol. 91, n° 3-4, p. 247-262.
F. & J.L. (1994).- Foraminifères vs. ammonites en fosse vocontienne vers la limite Valanginien-Hauterivien.- Comptes Rendus de l'Académie des Sciences, Paris, t. 319, ( série II), n° 2, p. 957-962.
E. & E. (1999).- Synthesis of calcareous nannofossil events in Tethyan Lower and Middle Jurassic successions.- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 105, n° 3, p. 343-376.
M. (1966).- Étude stratigraphique et micropaléontologique du Crétacé inférieur de la fosse vocontienne.- Documents des Laboratoires de Géologie de Lyon, n° 15, 369 p.
A. (2007).- La transgression du Crétacé inférieur au Nord du Chili. Biostratigraphie, Paléontologie (Ammonites), Statigraphie séquentielle et tectonique synsédimentaire.- Unpublished Ph.D. thesis, Université Paul Sabatier, Toulouse, 293 p.
J. (1992).- Biostratigraphy and palaeobiogeography of early Cretaceous calcareous nannofossils.- Cretaceous Research, London, vol. 13, n° 2, p. 167-189.
J. (reporter), G., E.J., F., E., S., W., P., M., J., H., P.F., P., Z. & and K. von (1996).- The Hauterivian Stage.- Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre, n° 66 suppl., p. 19-24.
G.S. (1994).- Geological time scale (1994).- Comptes Rendus de l'Académie des Sciences, Paris, t. 318, (série II), n° 1, p. 59-71.
J.G., F.P.& F.M. (2004).- The Cretaceous Period. In: F., J., A. (eds.), A geological time scale.- Cambridge University Press, Cambridge, p. 344-383.
D. (1996).- Zoophycos distribution and sequence stratigraphy. Examples from the Jurassic and Cretaceous deposits in south-eastern France.- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, vol. 123, n° 1-4, p. 273-287.
D. (2003).- Early Jurassic to Late Cretaceous evolution of Zoophycos in the French Subalpine Basin (southeastern France).- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, vol. 192, n° 1-4, p. 59-78.
G. (1973).- Ammoniti del Baiociano superiore di Digne (Francia SE, dip. Basses-Alpes).- Bollettino della Società Paleontologica Italiana, Modena, vol. 10, n° 2, p. 75-142.
G. (1983).- Ammoniti e biostratigrafia del Baiociano inferiore di Digne.- Bollettino del Museo Regionale di Scienze Naturali di Torino, Monografie, II, 257 p.
G. & C. (1968).- Étude biostratigraphique du Bajocien des Chaines Subalpine aux environs de Digne (Basses Alpes).- Bollettino della Società Geologica Italiana, Roma, vol. LXXXVII, p. 305-316.
P., J.-F., W.D., J., D., D. & A. (2003).- Characterization and correlation of Upper Jurassic (Oxfordian) bentonite deposits in the Paris Basin and the Subalpine Basin, France.- Sedimentology, vol. 50, n° 6, p. 1035-1060.
K. (1979).- Calcareous nannofossils from the Cretaceous between the North Sea and the Mediterranean.- Aspekte der Kreide Europas, Stuttgart, IUGS Serie A, n° 6, p. 223-272.
K. (1985).- Mesozoic calcareous nannofossils.- In: H.M., J.B & K. (eds.), Plankton Stratigraphy.- Cambridge University Press, p. 329-426.
P. (1988).- Die Ammoniten im Valangin-Hauterive Grenzbereich vom Mittlelandkanal bei Pollhagen.- Berliner geowissenschaftliche Abhandlungen, Reihe A, Band 94, p. 15-71.
P.F. (1973).- Lower Cretaceous (Ryazanian-Barremian) marine connections and cephalopod migrations between the Tethyan and Boreal Realms. In: R. & P.F. (eds.), The Boreal Lower Cretaceous.- Geological Journal, Liverpool, Special Issue, vol. 5, p. 131-144.
P.F. (1983).- The Valanginian to Aptian stages current definitions and outstanding problems.- Zitteliana, München, Heft 10, p. 493-500.
P.F. (1993).- The influence of sea level changes on the migration and evolution of early Cretaceous (pre-Aptian) ammonites. In: M.R. (ed.), The ammonoidea: Environment, Ecology, and Evolutionary change.- Systematics Association Special Volume, n° 47, p. 227-242.
P.F. (2004).- Report of the IUGS-ICS Subcommission on Cretaceous Stratigraphy.- Stratigraphy Newsletter, n° 6, 12 p.
P.F & P.J. (reporters), M.B, E., M., S.R.A., J., E., H.G., P., M.R.A., M. & Z. (1999).- Report on the 4th International Workshop of the Lower Cretaceous Cephalopod team (IGPC-Project 362).- Scripta Geologica, Leiden, Special Issue 3, p. 313.
S. (1996).- L'évolution des ammonites du Valanginien-Hauterivien inférieur du bassin vocontien et de la plate-forme provençale (Sud-Est de la France): relations avec la stratigraphie séquentielle et implications biostratigraphiques.- Documents des Laboratoires de Géologie de Lyon, n° 137, 371 p.
S. & F. (1995).- Rôle du climat sur les migrations et la composition des peuplements d'ammonites du Valanginien supérieur du bassin vocontien (S-E de la France).- Geobios, vol. 28, Supplement 1, p. 357-365.
S. & F. (1997).- Quantitative variations of the valanginian ammonite fauna of the Vocontian Basin (south-eastern France) between limestone-marls and within parasequences.- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, vol. 135, n° 1-4, p. 145-155.
S. & F. (1999).- Comments and proposals about the Valanginian-Lower Hauterivian ammonite zonation of south-eastern France.- Eclogae Geologicae Helvetiae, Basel, vol. 92, n° 2, p. 183-197.
S., F., S. & A. (1992).- Renouvellement des ammonites en fosse vocontienne à la limite Valanginien-Hauterivien.- Geobios, n° 25, fasc. 4, p. 469-476.
S., F. & O. (2005).- Ammonoid abundance variations related to changes in trophic conditions across the Oceanic Anoxic Event 1d (Latest Albian, SE France).- Palaios, Tulsa, vol. 20, n° 2, p. 121-141.
S. & P.J. (reporters), M.B., P., F., E.Y., M., G., Y., J. J.L., A., V., F.A., I., N., P., J., J.M., Z., J., H., B. & I. (2006).- Report on the second international meeting of the IUGS Lower Cretaceous Ammonite Working Group, the " Group" (Neuchâtel, Switzerland, 8 September 2005).- Cretaceous Research, London, vol. 27, n° 5, p. 712-715.
S., E., B., F., D. & O. (2003).- Ammonoid and nannoplankton abundance in Valanginian (early Cretaceous) limestone-marl successions from the southeast France Basin: carbonate dilution or productivity?- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, vol. 201, n° 1-2, p. 113-139.
E. (1874).- Tableau des terrains sédimentaires.- Bulletin de la Société Vaudoise des Sciences Naturelles, Lausanne, n° 13, p. 218-52.
P.H. (1978).- Calcareous nannoplankton biostratigraphy and oceanography of the Northwestern Atlantic Ocean.- Initial Reports of the Deep Sea Drillling Project, College Station, vol. 44, p. 731-760.
P.H. (1983).- Jurassic and Lower Cretaceous Calcareous nannofossils in the Western North Atlantic (Site 534): biostratigraphy, preservation, and some observations on biogeography and paleoceanography.- Initial Reports of Deep Sea Drilling Project, College Station, vol. 76, p. 587-621.
B. van de, O., T., P.& K. (2003).- Decoupling of P- and Corg-burial following Early Cretaceous (Valanginian-Hauterivian) platform drowing along the NW Tethyan margin.- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, vol. 199, n° 3-4, p. 315-331.
W. (1977).- Biostratigraphy of Cretaceous calcareous nannoplankton.- Geologie en Mijnbouw, Amsterdam, vol. 56, n° 1, p. 37-65.
C. (1967).- Ammonites and stratigraphy of the Bathonian in the Digne-Barrême area (south-eastern France, dept. Basses-Alpes).- Bollettino della Società Paleontologica Italiana, Modena, vol. 5, n° 1, p. 3-57.
H.R. (1973).- Lower Cretaceous calcareous nannoplankton biostratigraphy.- Abhandlungen der Geologischen Bundesanstalt, Wien, Band 29, 52 p.
H.R. (1976).- Mesozoic calcareous nannoplankton biostratigraphy of marine sediments.- Marine Micropaleontology, Amsterdam, vol. 1, p. 325-362.
J.-P. (1973).- The occurrence and distribution of boreal ammonites from the Neocomian of southeast France (Tethyan Province). In: R. & P.F. (eds.), The Boreal Lower Cretaceous.- Geological Journal, Liverpool, Special Issue, vol. 5, p. 289-302.
J.-P. (1977a).- La zone à Callidiscus du Valanginien supérieur vocontien (Sud-Est de la France). Lithostratigraphie, ammonitofaune, limite Valanginien-Hauterivien, corrélations.- Géologie Alpine, Grenoble, t. 53, p. 83-143.
J.-P. (1977b).- Les ammonites boréales des formations néocomiennes du Sud-Est français (province subméditerranéenne).- Geobios, n° 10, fasc. 3, p. 395-461.
J.-P., M. & L.G. (1990).- Biostratigraphie du Crétacé inférieur de l'Arc de Castellane (S.E. de la France). 1 : Faunes d'ammonites du Valanginien supérieur et âge de l'horizon de "La Grande Lumachelle".- Geologie Méditerranéenne, Marseille, vol. XVII, n° 1, p. 55-99.
M.G.E. (2001).- Die tiefe Unter-Kreide (Berrias bis Unter-Hauterive) im südwestmarokkanischen Becken: Ammonitenfauna, Bio-und Sequenzstratigraphie.- Thesis Universität Bochum, 142 p.
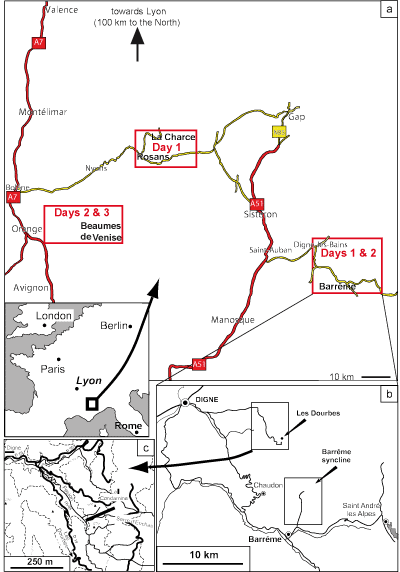
Click on the thumbnail to enlarge it.
Figure 1.1. (a) Location of the stops to be made during the fieldtrip in SE France. (b) Location of the sections to be visited during day 1 near the towns of Digne and Barrême. (c) Topographic map showing the location of the Feston section near Les Dourbes (SE of Digne; day 1, stop 4).
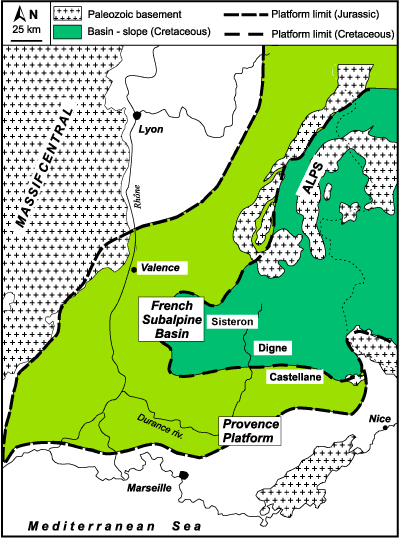
Click on the thumbnail to enlarge it.
Figure 1.2. The French Subalpine Basin or Vocontian Basin was bounded by the crystalline basement of the Massif Central to the West, by the Provençal Platform to the South and it was open to the East towards the Tethys Ocean. The basin-platform boundaries for Jurassic and Cretaceous times are also shown.

Click on the thumbnail to enlarge it.
Figure 1.3. Picture (taken from the road D19) showing the different formations encountered in the Feston Les Dourbes section, in particular the transition from the Calcaires à Cancellophycus to the Terres Noires formations is clearly visible. Ammonoid zones are also indicated.
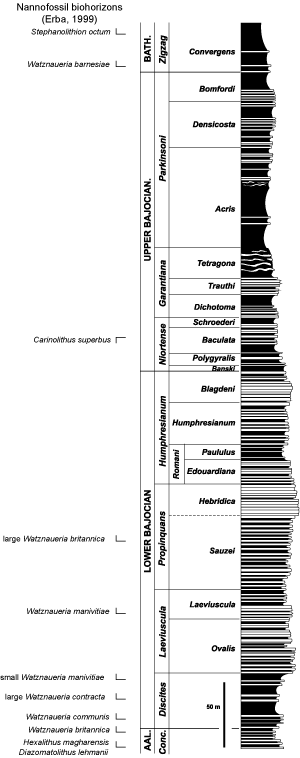
Click on the thumbnail to enlarge it.
Figure 1.4. Stratigraphical log of the Feston Les Dourbes section (modified after & , 1995). Biostratigraphical limits are sited in accordance with (1973, 1983). The relationships of nannofossil biohorizons to ammonoid biozones are those of (1990), who has studied three sections in the Digne area. Note that large Watznaueria contracta and small W. manivitiae sensu & (1999) correspond respectively to Watznaueria sp. 4 and Watznaueria sp. 5 of (1990).
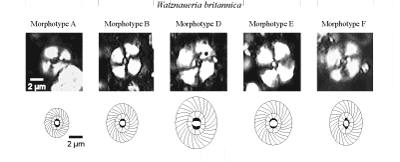
Click on the thumbnail to enlarge it.
Figure 1.5. Five of the six morphotypes of Watznaueria britannica described by et alii (2006) were found in the Feston Les Dourbes section. Note that preservation in the samples ranges from moderately good to poor.
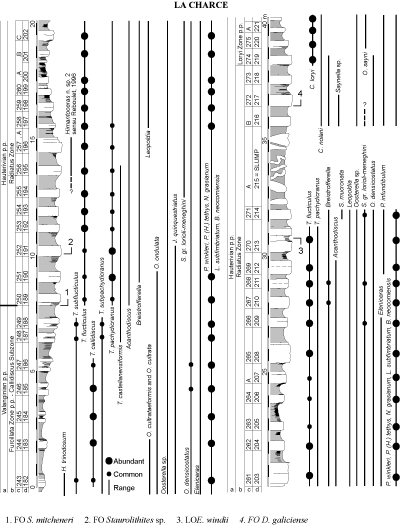
Click on the thumbnail to enlarge it.
Figure 2.1. Ranges of the ammonoids in and zonation of the La Charce section based on data from et alii (1992), et alii (1993, 1996), (1995), (1996), & (1997, 1999). This figure is the fruit of a collaboration with Luc . (a) Stages; (b) ammonoid zones; (c) bed numbers according to L. ; (d) bed numbers according to S. . The main nannofossil events recorded in the La Charce section are also shown (data from Silvia , unpublished).

Click on the thumbnail to enlarge it.
Figure 2.2. La Charce section. (a) Picture showing Upper Valanginian marl-limestone alternations (Verrucosum ammonoid Zone). (b) Marl-limestone alternations at the base of the Hauterivian Stage. (c) Panorama of the La Charce section.
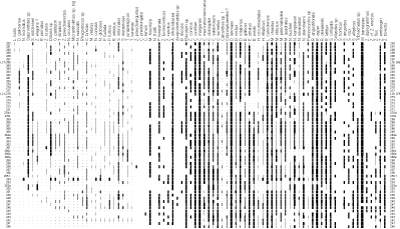
Click on the thumbnail to enlarge it.
Figure 3.1. Distribution chart of nannofossil taxa in the La Charce section across the Valanginian/Hauterivian boundary.
Click on the thumbnail to enlarge it.
Figure 3.2. Micrographs showing some nannofossil taxa recorded in the La Charce section. Scale bar is 5 µm. 1. Eiffellithus windii, sample LCH 251; 2-3. Eiffellithus striatus, LCH 255. Same specimen rotated; 4. Cruciellipsis cuvillierii, LCH 255; 5-6. Staurolithites mitcheneri, LCH 255. Same specimen rotated; 7. Tubodiscus verenae, LCH 272; 8. Rhagodiscus asper, LCH 270; 9. Nannoconus bucheri, LCH 260; 10. Nannoconus cornuta, LCH 260; 11. Nannoconus circularis, LCH 266; 12. Tribrachiatus sp., LCH 251; 13. Diloma galiciense, LCH 278; 14. Corollithion silvaradion, LCH 272; 15. Staurolithites sp., LCH 272; 16. Calcicalathina oblongata, LCH 272.

Click on the thumbnail to enlarge it.
Figure 4.1. Picture showing the interval in the Blieux section.

Click on the thumbnail to enlarge it.
Figure 4.2. Total abundance (specimens per view), diversity, and percentage of selected calcareous nannofossil taxa across the interval of the Blieux section (after et alii, 2003). The colour variations from pale-grey to dark-grey reflect change from oligotrophic to mesotrophic conditions.
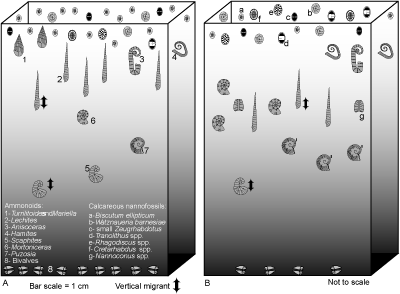
Click on the thumbnail to enlarge it.
Figure 4.3. Sketches showing the presence/absence of some Late Albian ammonoids, their varying abundance and their position in the water column in relation to trophic conditions during the interval of the Blieux section (after et alii, 2003). (A) Lower part of the interval: mesotrophic conditions in surface waters and episodic density stratification of the water column; (B) Upper part of the interval: oligotrophic conditions in surface waters and more stable paleoenvironment.
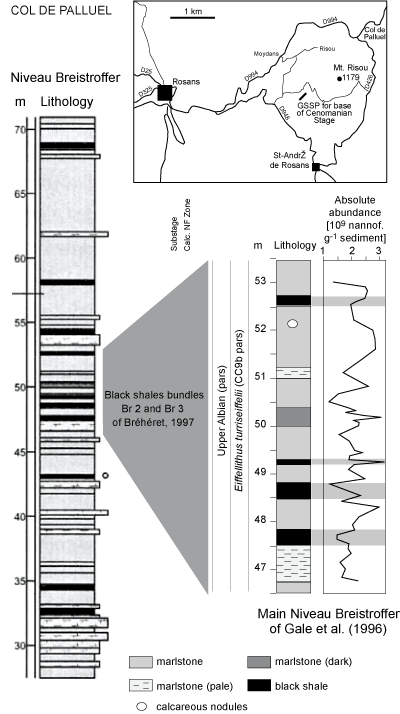
Click on the thumbnail to enlarge it.
Figure 4.4. Location of the Col de Palluel section. Columns show the lithology of the succession and the absolute abundance of calcareous nannofossils in the Col de Palluel section (modified after et alii, 2005).
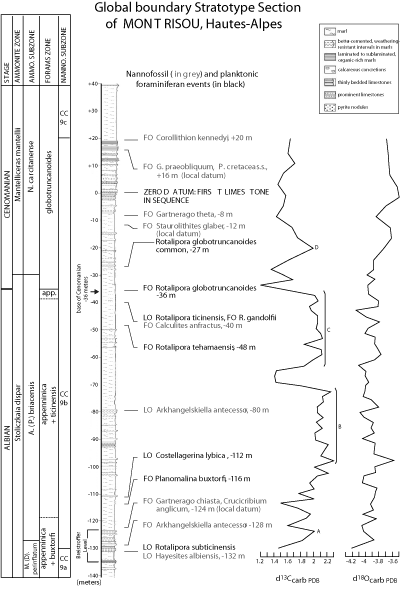
Click on the thumbnail to enlarge it.
Figure 5.1.
See Figure 4.4 ![]() for the location of the Global Stratotype Section and Point of Mont Risou. Integrated litho-, biostratigraphy and stable isotopic data across the Albian-Cenomanian
boundary at Mt Risou (modified after et alii,
2004). Nannofossil data are from J.A. .
for the location of the Global Stratotype Section and Point of Mont Risou. Integrated litho-, biostratigraphy and stable isotopic data across the Albian-Cenomanian
boundary at Mt Risou (modified after et alii,
2004). Nannofossil data are from J.A. .
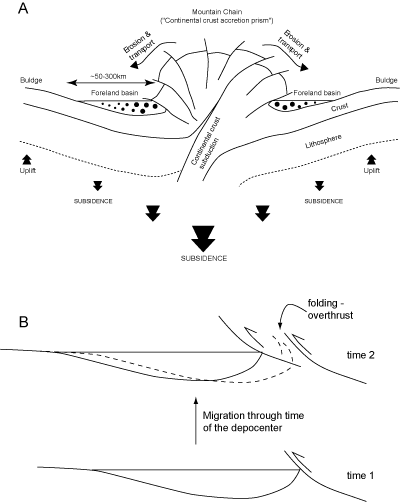
Click on the thumbnail to enlarge it.
Figure 6.1. Diagrams showing the relationships between Alpine mountain-building and the development of foreland basins (see the text for a more detailed explanation).
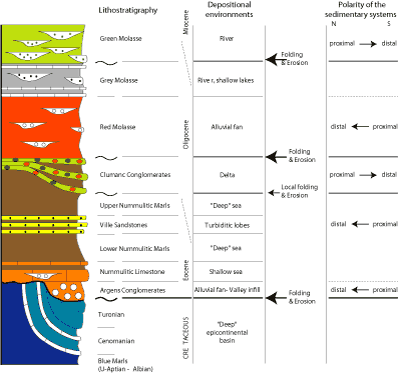
Click on the thumbnail to enlarge it.
Figure 6.2. Lithostratigraphy and facies of the stratigraphic entities encountered in the Barrême syncline. The major unconformities are also shown. An interpretation of the evolution of the different environmental conditions is shown as well as the polarity of the several sedimentary systems.

Click on the thumbnail to enlarge it. [Some rights reserved]
Figure 6.3. Topographic map of the Barrême syncline.
 |
|
|
A |
B |
Click on the thumbnail to enlarge it.
Figure 6.4. (A) Picture showing the unconformity between the Albian Blue Marls and the deposits of the Nummulitic Sea at Les Sauzeries Basses. The Ville sandstones are visible at the top of the hill. (B) Picture illustrating the complex geometrical relationships between the Clumanc Conglomerates and the Nummulitic Limestone and Marls at the Clumanc Castle.
