Court traité de Foraminiférologie
(L'essentiel sur les foraminifères actuels et fossiles)
_
Jean-Pierre Bellier
Robert Mathieu
Bruno Granier
Avant-propos
Après une quarantaine d'années dans le milieu universitaire, et avant d' abandonner
la qualité de salarié de l'Éducation nationale pour entrer dans le nouveau monde des "pensionnés", il est apparu judicieux, au premier auteur
(J.-P.B.) de ce court traité, de rédiger un court document rassemblant de façon synthétique les connaissances acquises, enseignées et pratiquées pendant plus de quatre décennies. Ce fascicule est consacré exclusivement aux foraminifères, lesquels sont, avec les nannofossiles calcaires, les outils fondamentaux de la biostratigraphie moderne, utilisés lors des campagnes océanographiques mondiales et pour les corrélations des forages pétroliers. Il y est question essentiellement des petits foraminifères, c'est-à-dire des formes à tests de petites dimensions sans architecture interne complexe. Ceux-ci comprennent les foraminifères planctoniques dont l'évolution rapide et la grande dispersion permettent de dresser des échelles biochronologiques de référence performantes.
Ce petit cours s'adresse aux étudiants de la préparation au CAPES
(Certificat d'Aptitude au Professorat de l'Enseignement du Second degré) et à l'Agrégation de Sciences
Naturelles, aux étudiants se destinant aux carrières scientifiques dans l'Université ou au
CNRS (Centre National de la Recherche Scientifique), aux étudiants souhaitant intégrer les cadres de l'Industrie pétrolière, là où une spécialité micropaléontologique jugée d'utilité est encore l'objet d'une considération méritée. Il intéressera aussi les naturalistes amateurs et le public profane attirés par les beautés
d'un microcosme vivant ou disparu et fossile.
Introduction
Les foraminifères sont des organismes unicellulaires du monde animal qui vivent sur le fond des lagunes et des mers, ou bien parmi le plancton océanique. Le cytoplasme de la cellule de ces protozoaires est largement inclus dans une coquille (test) diversement composée de matière organique, de substances minérales ou de particules agglutinées. Le test comporte une ou plusieurs loges présentant (une ou) plusieurs ouvertures (foramens, terme dont dérive le nom du groupe) faisant communiquer les loges entre elles et avec l'extérieur. Le cytoplasme sort par (cet ou) ces orifices extérieurs, recouvre plus ou moins le test et émet des pseudopodes fins et réticulés avec lesquels le microorganisme se fixe sur son substrat, se déplace et capture ses proies. Les foraminifères sont connus depuis le début de l'ère primaire jusqu'à l'époque actuelle. Certaines de leurs lignées ont évolué rapidement, ce qui est très utile pour la biochronologie et les corrélations interrégionales précises des strates géologiques. Leur sensibilité écologique les rend aussi particulièrement intéressants pour l'étude des environnements actuels et anciens.
Le texte et les figures qui suivent ne sont qu'un aperçu sur cet embranchement fondamental en micropaléontologie. L'accent sera mis essentiellement sur les "petits foraminifères", c'est-à-dire ceux dont les plus grandes dimensions des tests sont voisines du demi-millimètre, et ceux dont les loges ne présentent pas de structures ou d'architectures internes complexes. La systématique et les relations évolutives ne seront abordées que dans leurs grands traits. Les foraminifères sont des enregistreurs des variations isotopiques des océans en 13C et 18O, mais cet aspect géochimique de leur utilisation n'est pas du domaine de la micropaléontologie fondamentale et ne sera pas développé. Les foraminifères planctoniques feront l'objet d'une attention particulière, étant donné leur grande utilité dans le suivi des sondages scientifiques et des forages pétroliers.
I. Caractères généraux
Les foraminifères sont des Protozoaires pourvus d'une coquille (= test) constituée de loges successives communiquant entre elles par des orifices appelés
foramens. Les loges sont séparées les unes des autres par des cloisons. La dernière loge communique avec l'extérieur par une ou plusieurs ouvertures. Le cytoplasme, qui emplit entièrement toutes les loges, sort par les ouvertures et recouvre extérieurement le test d'où il émet des pseudopodes filamenteux fins granuleux et réticulés ; ceux-ci incluent souvent des grains ou de fines particules de natures variées et jouent un rôle important dans les activités vitales : déplacement, nutrition, construction de nouvelles
loges, etc. Des algues symbiotiques (dinoflagellés) sont souvent présentes dans le cytoplasme.
Le cycle de vie des Foraminifères comporte une alternance de générations. Leur taille est généralement comprise entre 0,1 et 1 mm ; certains peuvent atteindre 10 cm ou plus.
Ce sont donc pour certains des unicellulaires géants !
Ils se trouvent dans tous les milieux aquatiques marins. Certains sont adaptés à des eaux saumâtres. Ils sont absents des eaux douces. On distingue :
- les formes benthiques (Figs. 1
 - 2 ) qui vivent sur le fond, soit à la surface du sédiment, soit enfouies dans celui-ci
(endofaune), soit sur des supports végétaux, rocheux ou des particules (épifaune) ;
- 2 ) qui vivent sur le fond, soit à la surface du sédiment, soit enfouies dans celui-ci
(endofaune), soit sur des supports végétaux, rocheux ou des particules (épifaune) ;
- les formes planctoniques (Fig. 3 ) qui sont passivement
entraînées par les courants, mais susceptibles d'effectuer des migrations verticales.
Les foraminifères à tests organiques sont mal connus [0rdre des Allogromiida]. Il s'agit essentiellement de formes uniloculaires qui ont été très peu étudiées par les micropaléontologues, du fait de leur
piètre potentiel de préservation dans les assemblages fossiles. La plupart des foraminifères que nous
étudions ont des tests minéralisés.
Trois types structuraux de la paroi des tests calcitiques ont été reconnus chez les Foraminifères vivants :
- agglutiné : la paroi d'un Foraminifère agglutinant a un aspect composite, avec un agglutinat
(grains de quartz, spicules, Nannoconus, ...) emprunté au milieu et un ciment secrété de nature "chitinoïde" ou
calcitique [ordres des Astrorhizida, Lituolida, Trochamminida et Textulariida].
Remarque : Depuis le Crétacé, on rencontre quelques rares formes, parmi lesquelles le genre Rzehakina Cushman,
1927, dont le ciment serait (?) siliceux [Ordre des Silicoloculinida] ;
- porcelané : la paroi calcaire, entièrement secrétée par l'animal, a un aspect
uniforme. Elle est imperforée, lisse et homogène, blanche à ambrée, brillante,
avec un aspect de porcelaine [Ordre des Miliolida] ;
- hyalin : la paroi, également secrétée, est perforée, lisse ou non (dans ce dernier cas, les irrégularités sont de même nature que le reste de la paroi), d'aspect vitreux, transparent ou
translucide [ordres des Carterinida (test constitué de
macrocristaux), Spirillinida (à comportement
monocristallin), Lagenida, Buliminida, Rotaliida et Globigerinida].
Remarque : Depuis le Permien, on rencontre des formes affines à tests (originellement) aragonitiques
et non calcitiques [ordres des Involutinida et Robertinida].
Ces trois types de tests calcitiques existent chez les formes fossiles où on
rencontre en outre un quatrième type de test calcitique. En effet, certains foraminifères du
Paléozoïque présentent un type de paroi calcaire dite microgranulaire [0rdre des
Fusulinida], composée comme chez les porcelanés et les hyalins de microcristaux de calcite, mais souvent agencés de façon à donner un aspect strié aux sections
on parle alors de paroi pseudofibreuse.
Le test uniloculaire, la forme la plus simple, est composé d'une seule loge sphérique ou tubulaire. Le plus souvent, le test comporte plusieurs
loges. Une loge est l'unité ajoutée au test pré-existant au cours d'une phase
de croissance. Elle peut être subdivisée en logettes qui résultent toutes de
la même phase de croissance. Les nouvelles loges, séparées par des
cloisons, sont de taille croissante. La jonction entre 2 loges successives, parfois visible à l'extérieur
du test, est appelée suture. Le nombre de loges, leur forme, leur agencement, et partant l'aspect général des tests multiloculaires, sont extrêmement variés.
1) Dans le mode rectiligne ou arqué, les loges sont agencées selon un axe droit ou courbe. S'il n'y a qu'une série de loges, le test est unisérié. S'il y en a plusieurs séries, il est bi-, tri- ou
multi- sérié.
2) Dans le mode enroulé, les loges sont disposées selon une spire. Si elle est plane, le test planispiralé possède deux faces identiques. L'enroulement est évolute quand tous les tours de spire sont visibles latéralement. Il est involute quand le dernier tour recouvre tous les précédents.
L'ombilic est situé dans l'axe de la
spire au point de convergence des sutures.
Si la spire est trochoïde, le test trochospiralé a ses deux faces différentes. L'une, évolute, est dite spirale, l'autre, involute, est dite ombilicale. Remarques :
- La direction de l'enroulement n'est pas toujours uniforme pour tous les individus d'une même espèce (elle peut être dextre ou sénestre), et dépendrait des variations climatiques du milieu.
- Dans la plupart des cas, les loges enroulées sont unisériées. Elles sont exceptionnellement bisériées.
3) Les tests annulaires ou cycliques ont des logettes disposées en anneaux concentriques.
4) Les Miliolidés présentent une disposition particulière, dite "pelotonnée". Les loges arquées, tangentes à leurs deux extrémités à l'axe d'allongement, sont arrangées en cycle
quinque-, tri- ou
bi- loculin.
5) À côté de ces modèles simples, de nombreux tests montrent successivement plusieurs modes d'agencement ; on parle alors de disposition mixte ou composite.
Très importante pour les processus physiologiques du
foraminifère, la disposition des ouvertures va jouer un rôle
significatif dans la classification taxonomique.
Chez les formes benthiques, la face aperturale désigne la partie de la paroi de
la dernière loge où s'insèrent une ou des ouvertures permettant la
communication avec l'extérieur. Elles montrent une très grande
variabilité dans leur forme et leur disposition.
L'ouverture principale put être simple et alors ronde, en fente, en croissant, radiée ou dendritique, quelquefois au sommet d'un col, bordée d'une lèvre lisse ou crénelée, ou partiellement recouverte par des productions calcaires telles que : dent, bulle, etc. Elle peut être au contraire multiple, avec plusieurs petits orifices disposés en file ou sans ordre.
Sa position est variable : elle peut être basale, terminale, suturale ou
périphérique. Dans le cas d'un test trochospiralé, elle peut
être médiane, ombilicale ou spirale.
Chez les formes qui vivent dans un
environnement isotrope, chez les planctoniques en particulier, il peut exister
des ouvertures supplémentaires réparties le long des sutures des loges.
Certaines petites formes benthiques présentent une
structure particulière rattachée au côté interne de l'ouverture, appelée la
"plaque dentale". Si sa fonction exacte n'est pas encore définitivement
élucidée son intérêt taxonomique est néanmoins significatif.
L'ornementation intéresse :
- la surface du test, lisse ou recouverte de stries, de côtes, d'un réticule, de tubercules, d'épines, etc. ;
- les sutures, simples ou limbées, c'est-à-dire soulignées par un bandeau plus ou moins large ;
- la périphérie du test, soulignée ou non par des épines ou une (ou deux) carène(s) ;
- l'ombilic, libre ou occupé par un ou plusieurs boutons.
Tous ces caractères, en se combinant, conduisent à l'extrême diversité morphologique des foraminifères.
Stratigraphiquement, les foraminifères apparaissent dans le Cambrien inférieur (les tous premiers, non
ambigus, proviennent du Cambrien basal de l'Afrique de l'Ouest), à peu près en même temps que les métazoaires à éléments squelettiques. D'après les données moléculaires, cette apparition aurait été précédée par une large radiation d'espèces uniloculaires et nues, non fossilisées, qui auraient divergé d'un ancêtre Cercozoaire, plusieurs millions d'années avant le début du Phanérozoïque (Pawlowski et alii,
2003). Le microfossile tubulaire
Platysolenites, décrit dans le Précambrien terminal de plusieurs régions, n'a pas été définitivement placé ou dans les vers ou dans les foraminifères (Langer,
1999). Les foraminifères planctoniques sont apparus dans le Toarcien (Jurassique inférieur). La question de l'existence possible de taxons "planctoniques" dans le Trias/Jurassique "offshore" du Nord-Ouest de l'Australie n'est pas définitivement résolu (Hart et alii,
2007).
Les foraminifères ont été particulièrement affectés par les grandes crises : extinction du
Permo-Trias, remaniements majeurs lors du passage Crétacé-Paléogène, par exemple. C'est par leur étude qu'a été souligné le caractère catastrophique de certaines transitions majeures pour de nombreux groupes d'organismes, transitions correspondant à des coupures importantes des temps géologiques. À plus petite échelle, la connaissance de la succession des espèces dans des familles bien documentées permet de disposer d'un outil de datation qui
peut être des plus précis. Leur sensibilité aux conditions de milieu en font, d'autre part, des indicateurs de conditions
(paléo-) environnementales très performants.
Historiquement, les foraminifères sont connus depuis l'Antiquité : Hérodote (vers 484-425 av. J.C.), Strabon (63 av. J.C.
- 20 ap. J.C.), puis Pline l'Ancien (23-79 ap. J.C.) avaient mentionné une accumulation d'objets en forme de lentilles dans les calcaires des pyramides d'Égypte : ils avaient détecté la présence des Nummulites (grands foraminifères benthiques).
La Classe des Foraminifera a été créée, au début du
19ème siècle, par Alcide d'Orbigny (1802-1857), dans son ouvrage intitulé
"Tableau méthodique de la classe des Céphalopodes" (1826). Fondateur de la Micropaléontologie, cet auteur décrivit plus de 1000 espèces et fut le premier à étudier le mode de vie et l'écologie de ces petits organismes qu'il classait avec les ammonites. La nature unicellulaire des foraminifères a été découverte en 1835 par Félix Dujardin (1801-1860). Les échantillons des fonds océaniques récoltés lors de la première campagne océanographique scientifique du "Challenger" (1873-1876), étudiés jusque dans les années 1880 par des spécialistes comme Brady, ont apporté une somme de données fondamentales sur le vivant et le fossile. Les travaux poursuivis pendant le
20ème siècle ont développé l'utilisation des foraminifères comme outils
biostratigraphiques. Y sont
associés des noms prestigieux tels que Cushman, Loeblich et Tappan aux États-Unis, Subbotina en Union Soviétique, Bolli et Sigal en
Europe, pour n'en citer que quelques-uns.
II. Quelques données du Vivant
1. Nourriture
Les foraminifères benthiques herbivores actifs broutent des algues (et des bactéries) en se déplaçant sur leur substrat. Les herbivores passifs sont des formes épifaunes sessiles qui capturent la nourriture (diatomées) au voisinage du site de fixation. Les proies des carnivores sont de petits arthropodes et d'autres foraminifères. La digestion dure environ un jour. Certains sont
suspensivores. La majorité des foraminifères vivant dans les sédiments fins situés sous la zone photique sont détritivores et bactérivores. Ils y prédominent dans la partie superficielle (1-2
cm) immédiatement sous l'interface eau-sédiment. Beaucoup de foraminifères benthiques sont d'opportunistes
omnivores et beaucoup de grands benthiques hébergent des algues symbiontes qui
leur fournissent un appoint énergétique.
Les espèces planctoniques à épines mangent essentiellement des organismes zooplanctoniques comme les copépodes. D'autres crustacés,
tintinnoïdiens, radiolaires, polychètes et larves de gastéropodes, ptéropodes et tuniciers sont digérés aussi. Les copépodes calanoïdes sont digérés en 7 à 9 heures, les copépodes cyclopoïdes plus longuement, en 9 à 20 heures. Les copépodes harpacticoïdes ne sont digérés qu'exceptionnellement. Les besoins sont évalués à une capture puis une digestion toutes les 24 heures. Les espèces sans épines mangent essentiellement du phytoplancton : surtout des diatomées, ainsi que des
coccolithophoridés et des dinoflagellés. Ces foraminifères peuvent ingérer des copépodes, mais seulement quand ceux-ci sont immobilisés ou morts. Les temps de digestion sont beaucoup plus longs que ceux observés chez les espèces à épines (suivant Hembelen et alii,
1989).
Lorsque des foraminifères ont été ingérés par d'autres organismes ( vers, gastéropodes, crustacés, échinodermes, poissons,
...) ils parviennent le plus souvent à traverser le système digestif sans
être particulièrement affectés.
2. Fonctions du test
Les six fonctions possibles du test sont (d'après Murray,
1991) :
- protéger le foraminifère contre la prédation ;
- servir de barrière à un environnement défavorable ;
- constituer un réceptacle pour de la matière excrétée ;
- aider dans le processus de reproduction ;
- contrôler les mouvements de l'organisme ;
- assister la croissance de la cellule.
Les foraminifères hyalins construisent des tests plus poreux dans les eaux chaudes et les moins salées (Hembelen et alii,
1989). Ces eaux sont les moins denses. La porosité des foraminifères planctoniques peut donc être utilisée comme un outil paléocéanographique pour évaluer la température, la salinité et la densité relatives de l'eau de mer. Les dimensions et la densité des "perforations" sont aussi des critères de distinction systématique chez les planctoniques actuels et fossiles : on oppose classiquement, par exemple, les formes microperforées et les formes macroperforées dans la classification des genres du Crétacé inférieur (Moullade et alii,
2002).
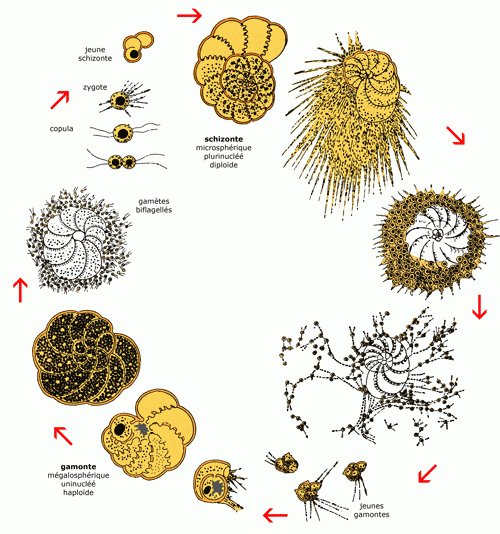
3. Reproduction
Sur approximativement 4.000 espèces vivantes, le cycle de vie d'une
trentaine d'entre elles est bien connu. Étudié en laboratoire chez les benthiques, ce cycle est
haplo-diplophasique, avec alternance d'une génération haploïde et d'une génération diploïde
(Fig. 4 ). Les formes haploïdes sont appelées
gamontes. Ceux-ci donnent, par division du noyau, des gamètes (cellules
sexuelles indifférenciées). La fusion de deux gamètes donne un individu diploïde, le
schizonte, plurinucléé, qui se reproduit par mitose. Après
méiose et division du cytoplasme autour de chaque noyau, les schizontes donnent de nouveaux
gamontes. Gamontes et schizontes se distinguent par leur taille et surtout par celle de leur loge initiale. Les gamontes sont aussi appelés formes mégalosphériques et les schizontes formes microsphériques.
Chez les foraminifères planctoniques, le dimorphisme est inconnu et il n'y a
probablement pas de reproduction asexuée
en accord avec leur stratégie de vie. Chez de nombreuses espèces, une couche de calcite est
sécrétée sur les loges du dernier tour pendant la gamétogénèse. La forme ultime de cette calcification constitue la "croûte
calcitique" ; sa signification est énigmatique
(résistance à la dissolution ?).
Beaucoup de formes fossiles ont été décrites bien avant que les
modalités de reproduction des foraminifères n'aient été connues, si bien que de nombreux morphotypes ne différant que par la taille et la dimension de la première loge ont été placés dans des espèces différentes. Des réajustements postérieurs à la découverte de l'existence des deux phases du cycle ont amené les micropaléontologues à conserver un double nom spécifique pour dénommer certains taxons, ce qui est contraire aux règles de la nomenclature linnéenne. Les exemples sont multiples chez les Nummulites.
4. Productivité
La longévité des foraminifères, difficile à évaluer, est comprise entre 15 jours et 16 mois. Les tests de nombreux foraminifères sont partiellement dissous ou partiellement désintégrés pendant la phase reproductive. Sur le substratum, la production carbonatée, évaluée chez les grands foraminifères benthiques tropicaux ou subtropicaux, varie de 40 à
2.800
g/m2.an. Les comptages indiquent des densités de populations variant de 0 à 8.600 Foraminifères par dizaine de
cm2 de surface de fond. Les tests de petite taille et à paroi fine sont souvent transportés en suspension. Il s'agit d'adultes de petites espèces et de juvéniles d'espèces plus grandes qui ont généralement moins de 200 µm de diamètre ou de longueur (selon Murray,
1991). Dans de nombreux de cas, ils sont transportés
post mortem loin de leur source, souvent sur des distances de plus de 100 km.
Le grand intérêt géologique des foraminifères planctoniques tient à leur immense productivité. Ils vivent dans toute la colonne d'eau. Les plus fortes concentrations se trouvent à des profondeurs comprises entre 10 et 50 m. Les populations vivantes comptent 1 à 200 individus par
m2 en surface. Dans la nature actuelle, les individus et les espèces sont plus abondants dans les eaux chaudes. Il a pu être compté jusqu'à 10.000 Foraminifères par
m3 d'eau, mais jusqu'à moins de 1 individu dans 100 m3 à des profondeurs supérieures à 1000 m. Actuellement, on estime que 47% des fonds océaniques,
soit 126.000.000
km2, sont couverts par leurs coquilles qui tombent en pluie constante, à la vitesse moyenne de 2 cm/s, sur le sol marin. Plusieurs milliers de
spécimens peuvent être contenus dans un gramme de sédiment.
III. Méthodes d'étude
Pour rechercher les foraminifères les sédiments meubles (sables, argiles, marnes, etc.) sont lavés à l'eau du robinet sur des tamis à mailles très fines (communément de 63 µm). L'opération peut être facilitée en remuant l'échantillon avec une brosse souple. Elle est terminée lorsque l'eau qui traverse le (ou les) tamis est aussi claire qu'à la sortie du robinet. Les résidus de lavage sont séchés, puis
éventuellement fractionnés à l'aide de tamis secs. Ils sont triés à la loupe binoculaire, à un grossissement compris entre x 20 et x 100, avec une aiguille frottée sur de la
pâte à modeler ou, mieux encore, un pinceau très fin (000) humidifié. Le tri a pour but de prélever et de regrouper dans des cellules les tests, en respectant, autant que possible, leur fréquence dans les assemblages. Après chaque lavage, les tamis sont précautionneusement brossés, passés à l'air comprimé, plongés dans une solution de bleu de méthylène et séchés. Le passage au bleu à pour effet de marquer les objets susceptibles d'avoir échappé au nettoyage et, en les repérant facilement dans les préparations ultérieures, d'éviter les contaminations.
Les roches dures (calcaires, grès) nécessitent la confection de lames minces. Les foraminifères sont alors observés en sections, au microscope optique ou à la loupe binoculaire. Dans la plupart des cas, les lames sont taillées à
environ trois centièmes de
millimètres (30 µm) d'épaisseur, afin de préserver la netteté des contours et la lisibilité des structures internes des tests. Les méthodes d'extraction à l'acide acétique donnent rarement de bons résultats et sont peu utilisées. Les roches semi-indurées
(marnes, craie) peuvent être lavées avec succès après concassage, trempage puis éventuel traitement à chaud à l'eau oxygénée.
Le microscope électronique à balayage (MEB) traditionnel est utilisé en routine pour les illustrations et les observations à fort grossissement. Les foraminifères, simplement disposés à l'aide de
adhésif collant double face sur un plot métallique, sont recouverts d'une fine pellicule d'or et/ou de carbone dans un évaporateur. Le microscope électronique environnemental ne nécessite pas de métallisation, mais il n'équipe pas encore, à ce jour, tous les centres de recherche. Les prises de vues numériques facilitent largement la réalisation des planches photographiques qui illustrent nécessairement les articles publiés dans les revues de micropaléontologie.
IV. Éléments de Systématique
Dans la classification des êtres vivants selon Margulis et alii
(1998), les foraminifères (= Granuloreticulata) correspondent à un embranchement parmi les trente que comptent les
"Protoctista". Sur la base de caractères moléculaires, ils sont aujourd'hui placés dans un ensemble plus vaste qui rassemblent plusieurs taxons unicellulaires sans caractère morphologique commun évident (Lecointre & Le Guyader,
2001).
Une taxonomie moins moderne ("non cladistique" et se trouvant dans toute bonne encyclopédie) propose une hiérarchie des emboîtements systématiques ordonnée de la sorte :
l'Ordre des Foraminifères (plus savamment des Foraminiferida) appartient au Royaume des Protistes
(Protista), Sous-royaume des Protozoaires (Protozoa), Phylum des Sarcomastigophores
(Sarcomastigophora), Sous-phylum des Sarcodines (Sarcodina),
Super-classe des Rhizopodes (Rhizopoda), Classe des Granuloreticulosés (Granuloreticulosa).
La plupart des classifications des composants de l'Ordre - élevé ici au
rang de Classe - des Foraminifères, comme celle de Loeblich & Tappan
(1987), révisée par Sen Gupta
(1999), restent artificielles et sont fondées sur la nature de la paroi, puis l'agencement des loges et les types d'ouverture.
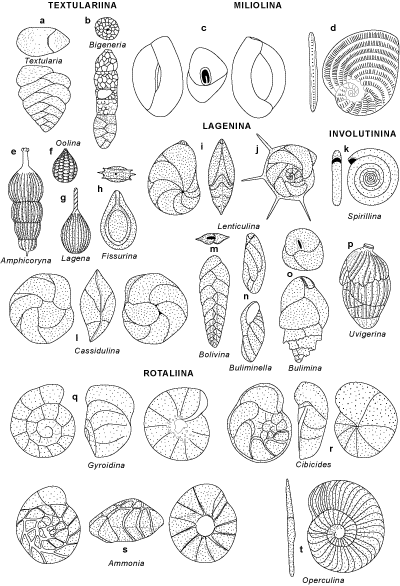
Ainsi l'Ordre des ALLOGROMIIDA rassemble les foraminifères à tests organiques
alors que ceux à tests minéralisés sont classés en
15 ordres dont les plus importants sont les suivants :
- ASTRORHIZIDA, LITUOLIDA (Orbitolina), TROCHAMMINIDA et TEXTULARIIDA
(Textularia, Spiroplectammina, Bigenerina) : 4 ordres qui regroupent
tous les Foraminifères agglutinants ;
- INVOLUTINIDA (Spirillina) et ROBERTINIDA : 2 ordres réduits qui regroupent les Foraminifères à test
aragonitique ;
- FUSULINIDA (Fusulina) qui regroupe tous les Foraminifères à test microgranulaire
(exclusivement paléozoïques) ;
- MILIOLIDA (Quinqueloculina, Alveolina, Borelis,
Orbitolites, Parasorites, Peneroplis, Spirolina)
qui regroupe tous les Foraminifères à test porcelané ;
- LAGENIDA (Lenticulina, Dentalina, Amphicoryna,
Lagena, Fissurina, Oolina) : formes hyalines présentant une structure de test originale
(monolamellaire) et une ouverture de type radiée ;
- BULIMINIDA et ROTALIIDA qui rassemblent la plupart des espèces benthiques à test
hyalin (bilamellaire). Les représentants de ces ordres sont extrêmement variés et représentent l'immense majorité des formes benthiques.
Quelques exemples de Super-familles :
- S.f. Buliminacea (B) (Buliminella, Bulimina) : formes à trochospire haute ;
- S.f. Bolivinacea (B) (Bolivina, Uvigerina) : formes bisériées ;
- S.f. Cassidulinacea (B) (Cassidulina) et Nonionacea
(B) (Melonis, Astrononion) : formes bisériées ou planispiralées ;
- S.f. Discorbacea (R) (Gyroidina, Cibicides, Heterolepa,
Planorbulina) : formes à trochospire basse ;
- S.f. Rotaliacea (R) (Ammonia, Cribroelphidium,
Elphidium, Operculina, Nummulites, Amphistegina,
Orbitoides) : formes trochospiralées ou planispiralées à structure interne complexe.
- GLOBIGERINIDA qui regroupe tous les foraminifères planctoniques (tous de
type hyalin bilamellaire).
Quelques exemples de Super-familles :
- S.f. Globotruncanacea, uniquement fossile, renferment les morphotypes bicarénés ;
- S.f. Globororataliacea, à test trochospiralé à périphérie souvent carénée et loges peu nombreuses ;
- S.f. Globigerinacea qui regroupe des formes à loges globuleuses.
V. Foraminifères et biochronologie
1. Intérêt biostratigraphique des foraminifères
Les foraminifères ont une extension chronostratigraphique allant du Cambrien inférieur à l'époque actuelle (Fig.
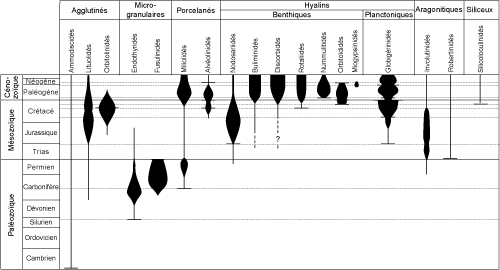
5 ). Les premières formes apparues ont des tests organiques ou de simples tubes agglutinés. Les espèces sont rares jusqu'au Dévonien. Les tests calcitiques
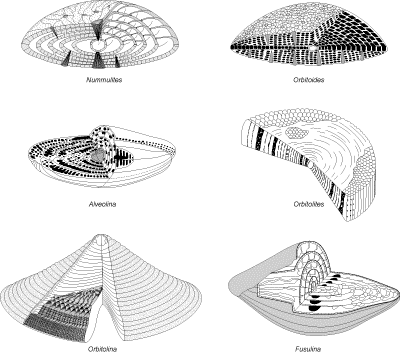
microgranulaires culminent au Carbonifère et au Permien avec le développement des Fusulinidés qui disparaissent à la fin du Paléozoïque. Les tests porcelanés
apparaissent au Carbonifère avec les Cornuspiridés. Ils sont suivis au cours du Mésozoïque par la multiplication des tests agglutinés (les Lituolidés au Jurassique et les Orbitolinidés au Crétacé) et par la radiation des tests hyalins (dont celle des Nodosariidés au Jurassique). Les premières formes sont toutes benthiques, les formes planctoniques n'existent pas avant le Jurassique. Les niveaux marins élevés et les climats chauds du Crétacé favorisent la diversification des planctoniques dont de nombreuses formes sont affectées par les extinctions majeures de la fin du Mésozoïque. Une rapide explosion évolutive eu lieu dès la base du Cénozoïque avec l'apparition de morphotypes rappelant les formes planctoniques actuelles. Les grands foraminifères benthiques se diversifient également dans le Paléogène avec les Alvéolinidés, les Nummulitidés et les
"orbitoïdiformes" ; ces derniers disparaissent à la base du Néogène. La diversité des planctoniques a aussi décliné depuis la fin du Crétacé avec quelques brèves recrudescences pendant les périodes chaudes du Paléogène et du Néogène. La distribution des types de tests et des familles au cours des temps ne peut être utilisée pour construire une échelle stratigraphique fine. Une bonne précision ne peut être espérée qu'en travaillant au niveau spécifique en se basant sur une connaissance approfondie des relations phylétiques des espèces (microévolution), afin de calibrer au mieux les extensions des taxons.
L'étude des strates sédimentaires basée sur le contenu fossilifère, plutôt que sur la lithologie ou d'autres paramètres géologiques, est la biostratigraphie. Les
(micro-)paléontologues organisent les couches sédimentaires en biozones fondées sur les premières et les dernières occurrences d'espèces sélectionnées. Ces niveaux d'apparition et d'extinction sont appelés biohorizons ("datums" en anglais). De tels taxons sont connus sous une variété de noms, comprenant "marqueur", "index", "guide" ou
"indicateur" (McGowran, 2005).
Les caractéristiques d'une espèce biostratigraphiquement utile sont les suivantes :
- extension stratigraphique courte ;
- dispersion géographique rapide et large ;
- bonne préservation ;
- reconnaissance relativement facile.
La reconnaissance des zones permet d'établir l'âge relatif, ou chronostratigraphie, d'une section sédimentaire basée sur la succession connue d'espèces fossiles au cours des temps géologiques, suivant les modalités de l'évolution biologique. Au cours du demi-siècle dernier, les occurrences des fossiles et les biozones successives ont été intégrées avec les données paléomagnétiques et les âges radiométriques partout dans le Monde de façon à construire
et calibrer une robuste échelle des temps géologiques. La géochronologie est basée sur l'âge absolu plutôt que sur l'âge relatif. Des biozones bien datées peuvent être corrélées avec des sections très distantes dans lesquelles les données paléomagnétiques ou autres outils de datation font défaut, étendant l'application de l'échelle des temps au Globe entier. Ces principes ont été appliqués à tout le Phanérozoïque
(quelques 540 millions d'années), dans des sections continentales et océaniques.
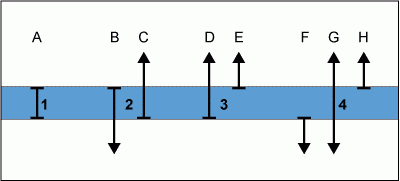
Les paléontologues ont reconnu différents types de biozones basées sur l'extension stratigraphique d'une ou plusieurs espèces. Le but de l'utilisation de différents types de zones est d'établir des intervalles stratigraphiques distincts représentant de courtes fractions de l'enregistrement du temps géologique. La Figure
6
présente quatre types de zones différents (A-H représentent les extensions stratigraphiques de 8 espèces différentes). Leurs définitions sont les suivantes :
- Zone d'extension totale de l'espèce A : depuis l'apparition de l'espèce A, jusqu'à la disparition de l'espèce A.
- Zone d'extension concomitante des espèces B et C : partie commune des extensions des espèces B et C, depuis l'apparition de l'espèce C et jusqu'à la disparition de l'espèce B.
- Zone d'extension partielle de l'espèce D : depuis l'apparition de l'espèce D, jusqu'à l'apparition de l'espèce E.
- Zone d'intervalle de l'espèce G : occurrence de l'espèce G entre la disparition de l'espèce F et l'apparition de l'espèce H.
2. Esquisse phylogénétique des planctoniques
Les foraminifères planctoniques sont d'excellents fossiles stratigraphiques du Crétacé jusqu'à l'époque actuelle, en raison de leur grande dispersion et de la rapidité de leur évolution. C'est pourquoi ils sont un outil biostratigraphique de premier
choix (Bolli
et alii, 1985).
Lesdits planctoniques sont vraisemblablement issus de foraminifères benthiques dont une phase planctonique du cycle se serait complètement séparée au début des temps mésozoïques. Ce scénario est parfaitement en accord avec le fait que les premières espèces du Jurassique et du Crétacé basal sont inféodées à des environnements de plate-forme externe (Caron,
1983).
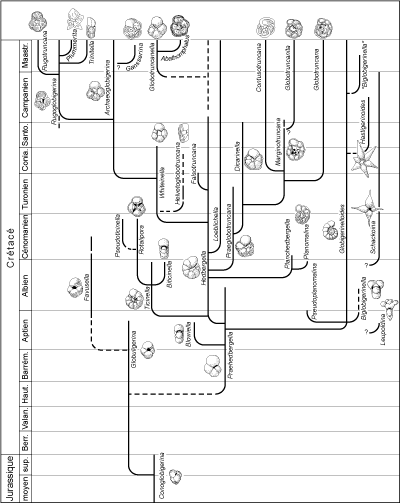
Le tableau phylogénétique des genres mésozoïques (Fig. 7 ) tient compte des principaux réajustements apportés à la taxonomie et la classification des morphotypes du Crétacé inférieur (Moullade et alii,
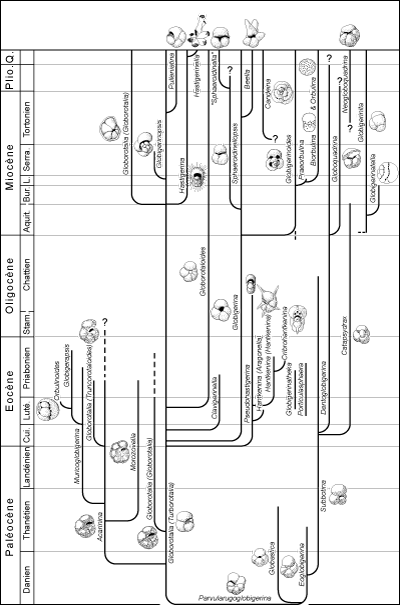
2002). Le tableau des genres cénozoïques (Fig. 8 ) est moins moderne en raison de la simplification recherchée à laquelle ne souscrira pas un spécialiste de métier. Il dresse cependant un panorama assez complet des événements évolutifs pour comprendre "ce qui s'est passé" au cours des temps, depuis la limite Crétacé-Paléocène jusqu'à l'époque actuelle. Dans les deux modèles représentés ne figurent que les morphotypes principaux trochospiralés ou planispiralés. Les morphotypes à loges sériées, souvent de moindre intérêt stratigraphique, ne sont pas pris en considération.
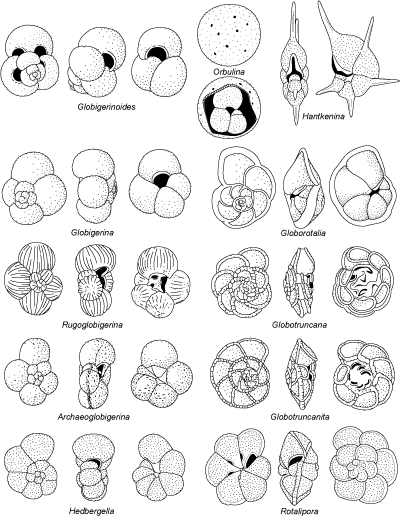
a - Les "Globigérinomorphes" du Jurassique
Des homéomorphes des Globigérines tertiaires et actuelles apparaissent au Lias et forment un groupe nettement différencié au Dogger. Les tests sont de petite taille, faiblement perforés, avec une ouverture
virguline, non parfaitement ombilicale, pourvue d'une petite lèvre. Les racines phylogénétiques de ces formes sont obscures. Elles ne constituent pas un groupe très homogène. Elles ont porté le nom de
"Protoglobigérines" ou de genre
"Globigerina". Protoglobigerina étant un nomen nudum et les liaisons avec les morphotypes ressemblants du Tertiaire n'existant pas, les termes génériques pour les désigner sont maintenant différents :
Conoglobigerina pour les formes à spire haute et Globuligerina pour les formes à spire basse. Les représentants de ces dernières au Crétacé inférieur seront la souche des
Praehedbergella puis des Favusella à paroi épaissie et ornementation en nids d'abeilles, encore inféodées à des environnements de plate-forme.
b - Les genres du Crétacé
En se basant sur l'histoire des foraminifères planctoniques, le Crétacé peut être divisé en 3 périodes : le temps des Hedbergelles et des Rotalipores, celui des Dicarinelles et des
Marginotruncana puis celui des Globotruncana et des Rugoglobigérines.
Au Crétacé inférieur, les foraminifères planctoniques sont principalement représentés par des formes peu spécialisées à test globigériniforme
(Praehedbergella microperforées, Hedbergella
macroperforées, etc.). Ces morphotypes à test trochospiralé et loges globuleuses constituent le tronc principal de l'arborescence évolutive. Les
Hedbergella, dont l'ouverture ombilicale-extraombilicale est bordée par un revers, constituent un stock durable très plastique dont les diverses tendances sont mises en évidence par les
différents taxons vers lesquels elles évoluent : enroulement devenant planispiralé
(Globigerinelloides), augmentation du nombre de loges par tour et ouverture de l'ombilic, puis remplacement des revers par des portiques
("portici") avec ouvertures infralaminales
(Ticinella), extension de l'ouverture vers la face spirale et apparition d'ouvertures
supplémentaires de ce côté (Loeblichella, exclusivement américaine), déplacement de l'ouverture vers une position plus ombilicale
(Whiteinella).
L'adjonction d'une carène est un phénomène important qui marque différents
rameaux en même temps (notons que la première carène était apparue
auparavant chez Pseudoplanomalina). Ainsi s'individualisent successivement
Planomalina, puis Rotalipora (ce dernier est en outre
caractérisé par l'adjonction de vraies ouvertures supplémentaires),
Praeglobotruncana et Helvetoglobotruncana. De Rotalipora se rapproche
Pseudoticinella dont la carène s'estompe au dernier tour. Les représentants de ces deux derniers genres disparaissent brusquement dans la zone à "Grandes
Whiteinelles" située à la transition Cénomanien-Turonien.
Peu avant et peu après cet événement, d'importantes mutations se produisent au sein des lignées et font apparaître isolément des caractères qui se retrouvent presque en totalité chez les
Globotruncana du Crétacé supérieur. Les principales étapes de cette évolution, à partir du
genre Praeglobotruncana, se résument de la façon suivante : apparition de 2 carènes
(Dicarinella), étirement des loges dans le sens de l'enroulement et courbure des sutures septales
de la face ombilicale (Marginotruncana), migration de l'ouverture principale vers l'ombilic et développement de plaques ombilicales,
appelées "tegilla" (Globotruncana).
C'est par le même processus qui a mené à Globotruncana que s'individualise le genre
Archaeoglobigerina. Chez Rugoglobigerina les loges sont ornées de côtes radiales typiquement alignées vers leur pôle périphérique. Le stock de ces dernières est affecté de variations tendant vers l'acquisition d'une carène
(Rugotruncana), la compression (Trinitella), l'allongement radial
(Plummerita). Si les affinités des Contusotruncana à spire élevée sont détectables, celles des
Globotruncanita monocarénées ainsi que l'origine des Globotruncanella et des
Abathomphalus, chez lesquels l'ouverture glisse en position
extraombilicale, sont difficiles à élucider.
D'un point de vue écologique, les représentants les moins spécialisés
(Hedbergelliformes) ont pu s'adapter à des milieux très divers, bien
qu'exclusivement marins. Leur habitat se situait aussi bien en haute mer que sur la plate-forme continentale ;
ils sont connus autant dans la zone intertropicale que dans les régions boréales. Les formes ornées ou carénées étaient certainement moins tolérantes et montrent majoritairement une répartition latitudinale assez stricte (zones intertropicales et tempérées). L'extinction massive de la fin du Crétacé a affecté principalement les formes spécialisées très ornementées et fortement carénées.
Un spécialiste raisonnable compte une centaine d'espèces de l'Albien au
Maastrichtien, permettant de délimiter 25 zones d'une durée moyenne de 1,75 Ma.
c - Les genres du Cénozoïque
En suivant l'histoire des foraminifères planctoniques, au cours du Cénozoïque se succèdent le temps du regain ou des formes naines, faisant suite à la catastrophe fini-crétacée, le temps des Acarinines et des Morozovelles puis celui des Globigérines et des
Globigerinoides.
Les espèces du Tertiaire seraient issues d'une souche résistante, probablement peu spécialisée, seule capable de survivre à un changement brutal des conditions de milieu dû à une conjonction de catastrophes. Une faune de petits morphotypes globigériniformes
(Parvularugoglobigerina, Eoglobigerina, Globastica), à structure très simple, commence à se développer à l'aurore du Paléocène. Le développement rapide et la diversification des espèces commence avec des globigérinomorphes
(Subbotina) et les Globorotalia non ornées (Turborotalia), suivies au Paléocène moyen et supérieur par les
Globorotalia carénées (Globorotalia sensu stricto) et des formes à surface épineuse
(Acarinina et Morozovella). Cette tendance s'affirme à l'Éocène inférieur (apparition de
Globigerina, Truncorotaloides, Catapsydrax, etc.) et s'accélère à l'Éocène moyen et supérieur où apparaissent plusieurs nouveaux genres
(Hantkenina, Cribrohantkenina, Globigerinatheca, Globorotaloides,
Orbulinoides, etc.).
Le développement de cette faune caractéristique s'interrompt brutalement à la fin de l'Éocène. Ce sont principalement les formes très évoluées et de courte longévité qui s'éteignent alors, tandis que plusieurs espèces de Globigérines persistent à l'Oligocène. C'est aussi à un soudain changement écologique, probablement une chute de température, qu'est due la disparition d'espèces hautement spécialisées.
Débutant à l'Oligocène inférieur, pour la troisième fois (comme au Crétacé inférieur et au Paléocène inférieur), se développe une faune, à partir de formes globigériniformes simples, de
Globorotalia non carénées, et plus tard , au Miocène, de Globorotalia carénées. Au Miocène inférieur apparaissent les
Globigerinoides, les Globoquadrina et les Globigerinita. Au cours du Néogène apparaissent encore de nombreuses formes spécialisées (par exemple
Globigerinatella, Sphaeroidinellopsis, Hastigerina, Candeina,
Orbulina, etc.). Cette tendance à la diversification, qui a commencé à l'Oligocène inférieur, continue encore aujourd'hui.
À l'échelle mondiale, au Paléocène, la distribution des taxons est uniforme à des latitudes avoisinant 50°. Les changements climatiques influencent cette large distribution géographique, d'abord à la fin du Paléogène, puis d'une façon beaucoup plus prononcée au cours du Néogène, avec comme conséquence des associations différentes en fonction des zones climatiques : populations des zones tropicales-subtropicales, tempérées et froides. Bien qu'il y ait des chevauchements de ces zones de températures, les corrélations à l'échelle interrégionale deviennent difficiles. C'est vrai en particulier pour le Miocène.
D'après la répartition des foraminifères planctoniques, le Cénozoïque est aujourd'hui divisé en une quarantaine de zones d'une durée moyenne de 1,75 Ma, comme dans le Crétacé.
VI. Foraminifères et (paléo-) environnements
1. Foraminifères et dépôts océaniques
Les océans couvrent 70% de la surface de la Planète. Ils sont une grande fosse dans laquelle se retrouvent des matériaux qui ont été arrachés aux continents, mais ils produisent aussi leurs propres matériaux sédimentaires, principalement par la vie qu'ils renferment.
a - Environnements marins
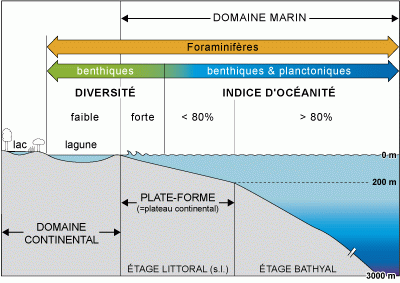
Le domaine margino-littoral (ou péritidal) correspond à une zone de bordure maritime baignée par des eaux présentant des variations de concentration saline (apports d'eaux douces et/ou isolement avec la mer). Le plateau ou plate-forme continentale, de 0 à 200 m en moyenne, est le domaine de pénétration de la lumière (zone photique ou zone des algues
photophiles). Il correspond au domaine littoral
(s.l.) qui est subdivisé en étages infralittoral et circalittoral (où se développent des algues supportant un faible éclairement). Le domaine infralittoral est celui des eaux agitées ; il est fortement influencé par des variations saisonnières (thermoclines saisonnières). L'étage bathyal, englobe la pente continentale (ou talus) et le glacis jusqu'à une profondeur d'environ 3.000 m. L'étage abyssal est situé au delà de 3.000 m (grandes plaines abyssales).
b - Dépôts océaniques terrigènes
La sédimentation de la marge continentale est principalement terrigène, c'est-à-dire que les matériaux proviennent de l'érosion des continents. Les débris de roches, plus ou moins altérés, arrachés au continent, sont transportés, s'accumulent et sont plus ou moins cimentés pour donner des sédiments ou roches détritiques. La nature des roches détritiques terrigènes dépend de la nature des roches soumises à l'érosion, du régime climatique de l'aire d'alimentation, des conditions
hydro- et/ou aérodynamiques de transport et de dépôt, ainsi que des transformations post-dépôt (que l'on appelle la diagenèse).
On note une répartition spatiale en fonction de la granulométrie : les plus grossières sur les plages (cordons de galets, sables grossiers) jusqu'à des boues et argilites lorsque la profondeur d'action des vagues de tempête (une centaine de mètres) est
dépassée (si on ne tient pas compte des turbidites, tsunamites et
countourites).
c - Dépôts océaniques biogènes
Le plancton est un des éléments essentiels des océans. Il est constitué de l'ensemble des micro-organismes qui vivent à la surface des océans, dans une couche qui fait jusqu'à plusieurs dizaines de mètres d'épaisseur et qui dépasse même les 100 mètres par endroits ; c'est une véritable soupe organique. Une grande proportion des micro-organismes planctoniques possèdent un squelette minéralisé, soit en carbonate de calcium
(CaCO3, calcite ou aragonite), comme par exemple les foraminifères ou certaines micro-algues du nannoplancton, soit en silice
(SiO2), comme les diatomées et les radiolaires. Après la mort d'un individu, son squelette devient une particule sédimentaire. Il s'ensuit que la surface des océans produit une pluie continuelle de très fines particules. Cette pluie, composée de matières organiques non encore oxydées, de calcaire et de silice produit une couche sédimentaire sur le plancher océanique. Il existe une limite naturelle en milieu océanique qu'on appelle la CCD
(calcite compensation depth = niveau de compensation de la calcite), profondeur à laquelle tout l'apport de calcaire est compensé par la dissolution. En somme, le gros des sédiments au large des marges continentales est produit par l'océan lui-même, biologiquement ; ces sédiments forment, à la surface des plaines abyssales et des zones de dorsales, une couche composée d'un mélange de matières organiques, de silice et possiblement de carbonates, avec des proportions variables d'argiles et de poussières atmosphériques.
Les composants majeurs des vases (ou boues) carbonatées pélagiques, c'est-à-dire de milieux marins francs et ouverts, sont les foraminifères et les coccolithophoridés dont le test est en calcite et les ptéropodes dont le test en aragonite n'est conservé que dans les domaines de faible bathymétrie. Dans les milieux plus côtiers, de faible profondeur, tout un cortège de macro-organismes (mollusques, gastéropodes, échinodermes, etc.), d'algues encroûtantes et d'ooïdes (dans les mers chaudes) viennent accompagner les carbonates d'origine planctonique.
Les foraminifères benthiques préfèrent les zones relativement peu profondes des plates-formes continentales où ils prolifèrent en fonction de différents facteurs (remontée de
nutriments ou éclairement important, etc.).
Il existe une liaison étroite entre la morphologie du test et le mode de vie. Les formes qui vivent dans les milieux agités possèdent une architecture solide, des coquilles bien calcifiées.
Les foraminifères vivent parfois en symbiose avec des algues. Il existe ainsi une liaison entre la nature du test et l'éclairement : chez les porcelanés, la lumière est
filtrée car trop importante (ces foraminifères s'adaptent à de très faibles profondeurs d'eau dans les régions très ensoleillées) ; chez les hyalins, elle peut être canalisée par les
cristaux du test (adaptation à une plus faible luminosité).
D'une façon générale, les formes sphériques ou lenticulaires (biconvexes, symétriques) se tiennent à la surface du substratum ; les foraminifères fusiformes sont fouisseurs ; les formes trochospiralées dissymétriques, avec une face plane, sont fixées ; l'aplatissement du test
s'accroît, en général, avec la profondeur. On observe, aussi, une liaison entre la profondeur et l'ornementation : dans les milieux peu profonds, pour résister à l'abrasion, le test présente souvent de nombreux nodules et épines.
Les foraminifères planctoniques exigent des conditions physico-chimiques stables, et ne supportent pas, en particulier, les nuisances liées aux eaux continentales (turbidité, baisse de salinité, etc.). Ils sont bien adaptés à la flottaison : test mince, inclusions protoplasmiques graisseuses, capsules gazeuses, etc.
Les formes actuelles à test épineux (Globigerina, etc.) prospèrent dans les eaux superficielles ; celles à test lisse
(Globorotalia, etc.) commencent leur cycle vital près de la surface, avant de s'enfoncer ensuite à plusieurs dizaines, ou centaines, de mètres de profondeur.
Comme cela nous a été rappelé par L. Hottinger
(communication personnelle, 26 Juin 2009), le rapport benthique-planctonique
dans les sédiments est lié au cycle de vie des foraminifères planctoniques
et en particulier à la profondeur à laquelle ils se reproduisent. La perte des épines lors de
la descente dans la colonne d'eau est un facteur connexe important dans le
processus. Un grand nombre de tests planctoniques
apparaissent sur un gradient profondeur seulement là où les tests
"maternels" vides vont sédimenter après la phase de reproduction.
Ils ne signalent pas où l'espèce vivait mais plus exactement où elle se
reproduisait.
Certaines espèces sont ubiquistes, d'autres ont une distribution localisée en fonction de la latitude.
2. Caractérisation des milieux par les foraminifères
La plupart des taxons ont des conditions d'existence bien définies et peuvent être utilisés comme indicateurs de milieu (bathymétrie, essentiellement), mais la connaissance taxonomique suffisante des foraminifères ne peut être acquise en peu de temps. Plus simplement, les caractéristiques générales suivantes sont importantes
(Sartorio & Venturini,
1988) :
- en milieu lagunaire, les benthiques sont seuls présents, les espèces en nombre restreint; genres typiques:
Alveolinella, Ammobaculites, Peneroplis, Trochammina ;
- dans les mers épicontinentales, ce sont les benthiques qui dominent, avec un nombre relativement important d'espèces; genres typiques à proximité du rivage:
Ammonia, Elphidium, Quinqueloculina ; en plate-forme interne:
Discorbinella, Eponides, Lenticulina, Textularia ; en plate-forme moyenne:
Amphicorina, Bigenerina, Lenticulina ; en plate-forme externe:
Bolivina, Bulimina, Discorbis, Nodosaria, Uvigerina,
Globigerina ;
- en domaine bathyal, les assemblages comportent en majeure partie des planctoniques
(Globigerina, Globigerinoides, Globorotalia, etc.) ; taxons benthiques typiques:
Epistominella, Nodosariidae, Pyrgo ;
- en domaine abyssal, les formes agglutinées simples prédominent.
a - Grands foraminifères et paléoenvironnements
Dans la nature actuelle, la distribution des grands foraminifères le long d'un profil continent – océan fournit des modèles de partition écologique qui sont transférables, avec beaucoup de précautions, dans certains domaines et à certaines périodes géologiques.
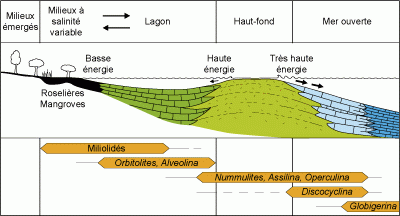
Le modèle d'Arni (1965, voir Fig.
9 ) est le plus classique d'entre eux pour l'Éocène
téthysien. Il met en jeu, outre les algues, les grands porcelanés et les grands benthiques auxquels sont adjoints les Miliolidés et les planctoniques. En utilisant uniquement des caractères supra-spécifiques "relativement simples", il permet de positionner approximativement n'importe quel assemblage dans la
succession des paléoenvironnements, depuis la côte jusqu'à la mer ouverte. Il est applicable, dans ses grandes lignes et avec prudence, à toutes les plates-formes carbonatées du Cénozoïque. C'est un outil pratique pour évaluer rapidement la profondeur, la salinité, l'oxygénation et le dynamisme des eaux lagunaires et/ou océaniques.
b - Petits foraminifères et paléoenvironnements
Indice d'océanité
L'indice d'océanité d'une population de foraminifères (Gibson, 1989) est le rapport quantitatif entre le nombre de planctoniques et le nombre total d'individus (benthiques + planctoniques) présents dans le sédiment
i = P / P + B
P est le nombre de foraminifères planctoniques comptés dans un volume défini de sédiment ; B est le nombre de foraminifères benthiques comptés dans le même volume. P + B devrait être compris entre 100 et 300.
La productivité optimale des foraminifères planctoniques se situe dans les eaux marines non côtières. Les foraminifères benthiques, quant à eux, prolifèrent sur les plate-formes continentales. L'indice
d'océanité permet donc d'évaluer la bathymétrie, et, par voie de conséquence, les différents milieux allant de la plate-forme continentale au domaine abyssal.
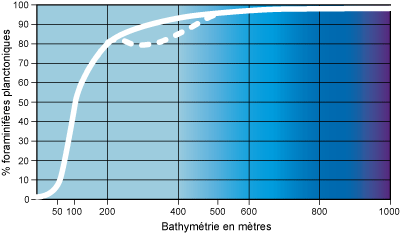
D'après des études réalisées dans différentes régions du Globe, la proportion de 50% de foraminifères planctoniques est atteinte entre
100 m et 200 m de profondeur et correspond à la zone externe de la plate-forme continentale (étage
circalittoral). La proportion augmente très rapidement sur le rebord de la plate-forme et au delà de
200 m de profondeur l'indice
d'océanité dépasse 80% (Figs. 10 -
11 ).
Ce premier indice permet de distinguer l'environnement marin franc, des environnements sous influence continentale (estuaires, lagunes, lagons, etc.).
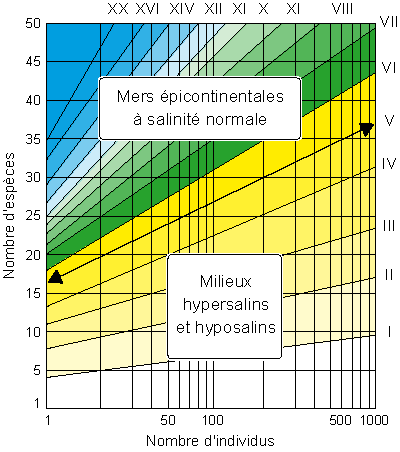
Indice de diversité et stratégies de vie
Cet autre indice permet de distinguer les différents environnements sous influence
continentale (Murray,
1991) ; les foraminifères benthiques sont alors seuls pris en considération.
La meilleure manière d'exprimer la diversité spécifique est de mettre en relation, dans une population de foraminifères, le nombre d'individus et le nombre d'espèces auxquelles ils appartiennent. Dans sa forme la plus simple, la diversité spécifique est le nombre d'espèces présentes.
Le calcul de l'indice de diversité peut être effectué mathématiquement, mais, d'une façon pratique, les valeurs de l'indice sont lues sur une abaque où sont portés en abscisse le nombre d'individus, et, en ordonnée, le nombre d'espèces
(Fig. 12 , indices en chiffres romains). Se distinguent assez clairement les mers épicontinentales à salinité normale des lagunes et marais hypo- et/ou
hyper- salins, la séparation étant la ligne de diversité d'indice V.
L'habitat d'une espèce conditionne sa stratégie démographique. Les habitats stables, permanents, ont des caractéristiques prévisibles, par opposition aux habitats instables, tels que les
marais littoraux ou les lagunes. Les espèces à stratégie r des habitats instables sont qualifiées d'opportunistes et les espèces à stratégie K des habitats stables sont des espèces d'équilibre. Les espèces opportunistes ont une durée de vie restreinte ; ce sont de petites formes à croissance limitée, à progéniture nombreuse, à diversité faible. Les espèces d'équilibre ont une croissance plus longue, une progéniture restreinte, une diversité forte.
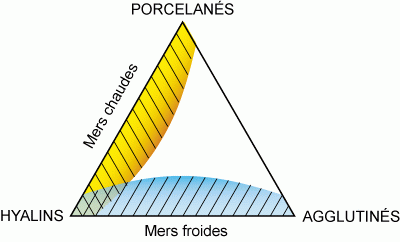
Diagramme triangulaire et conditions physico-chimiques
Dans le cas des domaines infra- et circa- littoraux, la proportion relative des tests hyalins, porcelanés et agglutinés permet d'évaluer la température de
l'eau (Murray,
1991). Ainsi une analyse simple de la composition d'une biocénose, fondée sur la distinction des trois types de natures des tests, informe-t-elle sur les conditions physico-chimiques ambiantes
(Fig. 13 : un point situé à chaque sommet du triangle signifie que la population comporte 100% du type concerné, sur les bases du triangle 0% du type indiqué au sommet opposé). Il ressort nettement de ces diagrammes que les biocénoses dans lesquelles dominent les hyalins et les porcelanés correspondent aux mers chaudes, alors que les hyalins et les agglutinés prédominent dans les mers froides.
c - Foraminifères planctoniques, paléogéographies et paléoclimats
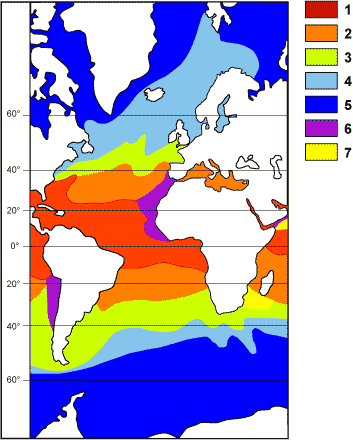
La biodiversité des foraminifères planctoniques actuels permet de délimiter cinq provinces biogéographiques latitudinales par hémisphère correspondant à autant d'intervalles thermiques, caractérisés par des associations spécifiques index (Bé & Tolderlund,
1971 ; compléments dans Hembelen et alii,
1989). Il s'agit des provinces (Fig.
14 ) :
- tropicale (1 : 0 à 25°N et 0 à 10°S ; températures de 24 à 30°C) ;
- subtropicale (2 : 25 à 40°N et 10 à 30°S ; températures de 18 à 24°C) ;
- de transition (3 : 40 à 45°N et 30 à 40°S ; températures de 10 à 18°C) ;
- subpolaire (4 : 45 à 60°N et 40 à 50°S ; températures de 5 à 10°C) ;
- polaire (5 : 60 à 90°N et 50 à 90°S ; températures de 0 à 5°C).
Cette configuration en cinq bandes s'estompe progressivement quand on remonte dans le
Cénozoïque. Au Mésozoïque les écarts thermiques sont moins tranchés mais les foraminifères planctoniques sont encore discriminants au Crétacé. Paradoxalement, les reconstitutions paléobioprovinciales concernant ce système et publiées à la fin du siècle dernier (synthèse dans Hart,
2000), ne tiennent pas compte de la composition spécifique précise des associations. Elles prennent en considération, sur la base de distinctions morphologiques générales, la prépondérance relative des formes à loges globuleuses et des formes carénées.
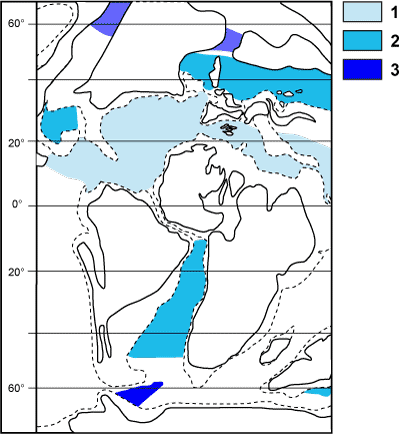
Un exemple peut être pris à l'Albien terminal, période clé de l'évolution des planctoniques au cours de laquelle un dispositif carénal robuste apparaît sur les tests. Trois biogéoprovinces par hémisphère sont ainsi distinguées (Fig.
15 ) :
- province téthysienne indivisée (1 = tropicale : 0 à 40°N) ;
- province transitionnelle (2 : 40 à 50°N et 0 à 45°S) ;
- province boréale (ou australe, 3 : 50 à 90°N et 45 à 90°S).
Pour cette même période, la prise en considération des extensions des espèces fournies par les données révisées des forages "DSDP"
(Deep Sea Drilling Project) et
"ODP" (Ocean Drilling Program) permet d'envisager une subdivision de la province téthysienne en trois bandes latitudinales dont les frontières se dessinent vers 25 et 30°N (Bellier & Vrielynck,
2007). Dans les trois sous-ensembles de cette province tropicale, les associations sont composées de morphotypes carénés et de morphotypes non carénés, sans que l'un ou l'autre des deux types prédomine. Dans les eaux atlantiques actuelles, une telle cohabitation caractérise les domaines tropical, subtropical et de transition, soit un créneau latitudinal compris entre l'équateur et 45°N ou 45°S. D'après les données paléobotaniques, une zonation paléoclimatique est affirmée à
l'Albien (Chumakov, 1995,
in Skelton et alii,
2003), avec trois ceintures de l'équateur au pôle : une ceinture aride équatoriale, une ceinture chaude de moyennes latitudes et une ceinture tempérée de hautes latitudes. Si les distributions des foraminifères crétacés sont en relation avec les variations de températures, comme c'est le cas actuellement, les planctoniques de
l'Albien supérieur paraissent indiquer des paléoprovinces océaniques plus restreintes que celles des plantes terrestres. Il semble donc que le microplancton recèle des indicateurs paléoclimatiques des plus sensibles.
Le passage Albien-Cénomanien serait caractérisé par des fluctuations importantes des températures, d'après des mesures du δ18O effectuées sur des
bélemnites et des mollusques (Frakes, 1999) :
l'Albien correspondrait à un des premiers épisodes chauds qui annoncerait l'effet de serre qui s'installera au Crétacé supérieur, après un épisode froid de très courte durée affectant le passage Albien terminal-Cénomanien basal. Les foraminifères planctoniques sont bien un matériel de choix pour des analyses isotopiques, mais les données mesurées dans l'intervalle considéré sont rares et inutilisables, car les valeurs proposées sont contradictoires. De plus, les habitats indiqués par les mesures ne sont qu'exceptionnellement en accord avec ce qui est connu des paléoprofondeurs vitales attribuées aux différents taxons, d'après des critères purement paléoenvironnementaux (Hart,
2000). Quand on observe les
variations de la distribution de toutes les espèces dans l'Atlantique Nord à la transition
Albien-Cénomanien, il apparaît une augmentation de la biodiversité qui pourrait aller de paire avec un
réchauffement. De même, quand on regarde les seules (18) espèces qui perdurent tout au long de cette transition, on ne constate pas un déplacement sensible de l'extension maximale des taxons vers le Sud (Bellier & Vrielynck,
2007). Les foraminifères planctoniques n'ont donc pas enregistré le refroidissement indiqué par les mesures isotopiques pratiquées sur les invertébrés.
Il est intéressant de constater que les biogéoprovinces de dinoflagellés présentent, à
l'Albien supérieur, un front de séparation, localisé entre 25 et 30°N, entre un domaine équatorial et tropical peuplé de taxons endémiques et cosmopolites et un domaine subtropical dans lequel seuls les taxons cosmopolites subsistent (Masure et alii,
2003). Ce front de transition, qui est perceptible dans deux groupes micropaléontologiques distincts, botanique et zoologique, semble donc bien correspondre à une réalité
paléoclimatique.
Conclusion
Les foraminifères, fraction importante de la productivité océanique, sont des indicateurs environnementaux et des fournisseurs d'âges souvent disponibles en assez grande quantité. Ces qualités sont particulièrement appréciées dans les travaux de recherches fondamentaux, la cartographie géologique, les explorations pétrolières et les forages océaniques.
Dans les campagnes scientifiques en mer, l'utilité des foraminifères est exemplaire : ce sont les foraminiféristes embarqués, associés aux
nannofossilistes, qui fournissent les déterminations des âges relatifs initiaux des séquences sédimentaires traversées en suivant la progression des sondages. Tous les 9,5 m
dans le cas de la plate-forme de forage du JOIDES Resolution, par exemple, le carottier est remonté du fond marin et un échantillon provenant du "core catcher", le "bouchon" qui empêche la carotte de glisser hors du tube, est fractionné et réparti parmi les deux équipes de spécialistes. Des montages entre lames et lamelles sont préparés pour l'étude des nannofossiles calcaires, et une plus grosse partie est lavée pour extraire des foraminifères, en routine. La détermination de l'âge peut être obtenue en moins d'une heure, quelquefois en moins de 15 minutes. L'établissement d'une chronostratigraphie "à grande vitesse" est fondamental pour toutes les analyses effectuées à bord, particulièrement pendant les expéditions visant des événements critiques dans l'histoire de la Terre, comme la limite Crétacé-Paléogène ou le maximum thermique du Paléocène-Éocène. Ainsi, les stratégies de forage peuvent être ajustées, si nécessaire, pour réduire le risque que l'intervalle intéressant soit disloqué ou perdu entre deux prélèvements.
La reconnaissance des zones biostratigraphiques aide beaucoup à l'identification des inversions magnétiques qui sont alors corrélables avec les chrons de l'échelle de polarité géomagnétique. La biostratigraphie joue aussi un rôle clé en développant un modèle d'âge pour chacune des séquences sédimentaires forées ; des diagrammes âge versus profondeur dans le puits permettent de quantifier les variations des taux de sédimentation. Le contrôle de l'âge relatif et une approche de l'âge absolu sont des nécessités pour caler dans le temps tous les types de données et comprendre comment les strates traversées s'insèrent dans le grand schéma temporel de référence.
Remerciements
Nous gardons (J.-P.B. et R.M.) le meilleur souvenir de notre cohabitation, tout au long de ces longues années, avec nos anciens collègues et néanmoins amis Françoise Dépêche, Claude Guernet et Gérard Bignot (†), de nos confrontations avec les occupants permanents ou épisodiques, enseignants-chercheurs, chercheurs et thésards du Laboratoire de Micropaléontologie de Paris, ayant subi les pérégrinations dudit laboratoire depuis les préfabriqués de la rue Saint-Jacques jusqu'aux tours 15-25 puis 46-56 du campus Jussieu. Nous
exprimons aussi notre reconnaissance à L. Hottinger
et à M. Moullade pour les suggestions qui ont
contribué à améliorer la version préliminaire de notre manuscrit.
Initialement prévu pour venir en complément d'un Manuel de
Micropaléontologie, la publication de ce fascicule précède finalement
celle du susdit ouvrage co-écrit par le même groupe d'auteurs (auxquels se
sont adjoints pour l'occasion d'autres collègues). La raison de ce décalage
est d'abord juridique, ensuite technique. Pour certaines illustrations que nous
souhaitions utiliser, n'étant pas détenteurs des droits, nous avons dans un
premier temps sollicité des autorisations de reproduction auprès des auteurs
et des éditeurs. Nous remercions vivement M. Hart, ainsi que
le B.R.G.M.
(Orléans), Elsevier (Oxford), la Geological Society Publishing House (Londres),
Koninklijke Brill (Leiden), Zitteliana (Munich) et leurs représentants
respectifs : F. Trifigny, C. Truter, A. Hills, G. van Rietschoten
et M. Krings pour
nous avoir accordé ces permissions ou assisté dans nos démarches. Toutefois
il est vite apparu que la traçabilité des droits de toutes les figures
n'était pas garantie. Pour cette raison, nous avons opté pour reprendre
l'ensemble de l'iconographie et présenter, en lieu et place, de nouvelles
illustrations, certes inspirées de celles citées en référence, mais
significativement modifiées. Nos remerciements vont à Alexandre Lethiers et Claude Abrial, de l'Atelier d'Infographie du Département de Géologie sédimentaire de l'Université Pierre et Marie
Curie, dessinateurs talentueux et respectifs des figures 1-3 &
5-13 et des figures 14-15.
Enfin, Nestor Sander a fourni une aide inestimable
par ses contributions à la version anglaise du texte et des figures.
Références bibliographiques
Arni P. (1965).- L'évolution des Nummulitinae en tant que facteur de modification des dépôts littoraux. In : Colloque International de Micropaléontologie (Dakar, 6-11 mai 1963).- Mémoires du Bureau de Recherches et Minières, Orléans, N° 32, p. 7-20.
Bé A.W.H. & Tolderlund D.S. (1971).- Distribution and ecology of living planktonic foraminifera in surface waters of the Atlantic and Indian Oceans. In : Funnell B.M. & Riedel W.R. (eds.), The micropalaeontology of oceans.- Cambridge University Press, p. 105-149.
Bellier J.-P., Dépêche F. & Mathieu R. (1995).- Introduction à la Micropaléontologie.- Documents pédagogiques du Laboratoire de Micropaléontologie, Université Pierre et Marie Curie, Paris, N° 1, 50 p.
Bellier J.-P. & Vrielynck B. (2007).- Distribution des foraminifères planctoniques au passage Albien-Cénomanien dans l'Atlantique Nord : Indices de l'existence d'une zonation latitudinale dans la province téthysienne.- Revue de Paléobiologie, Genève, vol. 26, n° 1, p. 55-62.
Bignot G., Dépêche F. & Mathieu R. (1975).- Initiation pratique à la Micropaléontologie.- Travaux du Laboratoire de Micropaléontologie, Université Pierre et Marie Curie, Paris, N° 4, 217 p.
Bignot G. (2001).- Introduction à la micropaléontologie.- Gordon and Breach Science Publishers, Paris, 258 p.
Blow W.H. (1979).- The Cainozoic Globigerinida: A study of the morphology, taxonomy, evolutionary relationships and the stratigraphical distribution of some Globigerinida (mainly Globigerinacea).- 3 vols., E.J. Brill, Leiden, 1413 p.
Bolli
H.M., Saunders J.B. & Perch-Nielsen
K. (1985).- Plankton stratigraphy.-
Cambridge University Press, 1032 p.
Caron M. (1983).- Taxonomie et phylogénie de la famille des Globotruncanidae. In : 2. Symposium Kreide. München 1982.- Zitteliana, München, (Reihe B: Abhandlungen der Bayerischen Staatssammlung für Paläontologie und Geologie), 10, p. 677-681.
Chumakov N.M. (1995).- Climatic zones in the middle of the Cretaceous Period.- Stratigraphy and Geological Correlation, Moscow, vol. 3, p. 3-14.
Frakes L.A. (1999).- Estimating the global thermal state from Cretaceous sea surface and continental temperature data. In : Barrera E. & Johnson C.C. (eds.), Evolution of the Cretaceous ocean-climate system.- Geological Society of America, Special Paper, Boulder, vol. 332, p. 49-57.
Gibson T.G. (1989).- Planktonic
benthonic foraminiferal ratios: modern patterns and Tertiary applicability.- Marine Micropaleontology, Amsterdam, vol.
15, n° 1-2, p. 29-52.
Hart M.B. (2000).- Climatic modelling in the Cretaceous using the distribution of planktonic Foraminiferida. In : Hart M.B. (ed.), Climates: Past and
Present.- Geological Society, Special Publication, London, n° 181, p.
33-41.
Hart M., Hudson W. & Smart C.W. (2007).- Palaeobiogeography of early planktonic foraminifera.- 1er Symposium international de Paléobiogéographie (Paris, 10-13 juillet 2007), Résumés, p. 47.
Hembelen C., Spindler M. & Anderson O.R. (1989).- Modern planktonic foraminifera.- Springer-Verlag, New-York, xiv + 363 p.
Hottinger L. (2006).-
Illustrated glossary of terms used in foraminiferal research.- Carnets
de Géologie / Notebooks on Geology, Brest, Memoir 2006/02 (CG2006_M02),
126 p.
Langer M.R. (1999).- Origin of foraminifera: Conflicting molecular and paleontological data?.- Marine Micropaleontology, vol. 38, p. 1-5.
Lecointre G. & Le Guyader H. (2001).- Classification phylogénétique du Vivant.- Ed. Belin, 544 p. + annexes.
Lister J.J. (1895).- VIII. Contributions to the life-history of the foraminifera.- Philosophical Transactions of the Royal Society of London B, vol. 186, p. 401-453, pls. 6-9.
Loeblich A. & Tappan H. (1987).- Foraminiferal genera and their classification.- Van Nostrand Reinhold, New York, vol. 1, 970 p. ; vol. 2, 212 p. + 847 pls.
Margulis L., Gould S.J., Schwartz K.V. & Margulis A.R. (1998).- Five kingdoms: An illustrate guide to the phyla of life on Earth.- Freeman & Co, New York, 448 p.
Masure E., Vrielynck B. & Fiet N. (2003).- Les Dinoflagellés et le gradient de température des eaux océaniques de surface à l'Albien supérieur. In : Bassins crétacés de France et d'Europe occidentale.- Séance spécialisée de la Société Géologique de France, 6-7 novembre 2003, résumé.
Mathieu R. (1986).-
Sédiments et foraminifères actuels de la marge continentale atlantique du Maroc.-
Thèse de Doctorat d'État ès Sciences naturelle, Université Pierre et Marie Curie, Paris 6 ;
Mémoires des Sciences de la Terre, Paris, n° 86-14, 419 p.
Mathieu R. (1988).- Foraminifères actuels et résurgences côtières sur la marge continentale atlantique du Maroc. In : Benthos'86.- Revue de Paléobiologie, Genève, vol. spécial, n° 2, Partie II, p. 845-850.
McGowran B. (2005).-
Biostratigraphy. Microfossils and geological time.- Cambridge University Press, 480 p.
Moullade M., Bellier J.-P. & Tronchetti G. (2002).- Hierarchy of criteria, evolutionary processes and taxonomic simplification in the classification of Lower Cretaceous planktonic Foraminifera.- Cretaceous Research, London, vol. 23, n° 1, p. 111-148.
Murray J.W. (1991).- Ecology and palaeoecology of benthic foraminifera.- Longman Scientific & Technical, 397 p.
Pawlowski J., Holzmann M., Berney C., Fahrni J., Gooday A.J., Cedhagen T., Habura A. & Bowser S.S. (2003).- The evolution of early Foraminifera.- Proceedings of the National Academy of Sciences of the United States of America, Washington, vol. 100, n° 20, p. 11494-11498.
Sartorio D. & Venturini S. (1988).-
Southern Tethys biofacies.- Agip, San Donato Milanese, 235 p.
Sen Gupta B.K. (1999).-
Systematics of modern Foraminifera. In : Sen Gupta B.K.
(ed.), Modern Foraminifera.- Kluwer Academic Publishers, Dordrecht, p. 7-36.
Skelton W., Spicer R.A., Kelley S.P. & Gimour L. (2003).- The Cretaceous world.- The Open University, Cambridge University Press, 360 p.
Figures
Cliquer sur l'image pour l'agrandir.
Figure
1 : Quelques petits foraminifères benthiques (plus grande dimension de
l'ordre du demi-centimètre en moyenne).
Cliquer sur l'image pour l'agrandir.
Figure
2 : Quelques grands foraminifères benthiques (plus grande dimension de
l'ordre du centimètre en moyenne).
Cliquer sur l'image pour l'agrandir.
Figure
3 : Quelques foraminifères planctoniques (plus grande dimension de
l'ordre du demi-millimètre en moyenne).
Cliquer sur l'image pour l'agrandir.
Figure
4 : Cycle de reproduction des Foraminifères benthiques (illustré par Elphidium
crispum (Linné), ex "Polystomella crispa", d'après J.J. Lister, modifié).
Cliquer sur l'image pour l'agrandir.
Figure
5 : Extension stratigraphique de quelques familles de Foraminifères (d'après Bignot,
2001).
Cliquer sur l'image pour l'agrandir.
Figure
6 : Principaux types de zones.
Cliquer sur l'image pour l'agrandir.
Figure
7 : Phylogénie des genres planctoniques du Mésozoïque (d'après Caron,
1983, modifié).
Cliquer sur l'image pour l'agrandir.
Figure
8 : Phylogénie des genres planctoniques du Cénozoïque (d'après Blow,
1979, modifié).
Cliquer sur l'image pour l'agrandir.
Figure
9 : Écozonation des plates-formes mésogéennes à l'Éocène
(il y a entre -40 à -50 Ma environ) avec les Foraminifères (d'après Arni, 1965,
simplifié).
Cliquer sur l'image pour l'agrandir.
Figure
10 : Indice d'océanité, diversité des Foraminifères et (paléo-)
environnements.
Cliquer sur l'image pour l'agrandir.
Figure
11 : Indice d'océanité (d'après Gibson, 1989, simplifié).
Sur les marges continentales soumises aux upwellings, la courbe de pourcentage
des foraminifères planctoniques subit, au sommet de la pente continentale (entre
200 m et 500 m), une inflexion (courbe en pointillés) liée à la productivité
importante des foraminifères benthiques (d'après Mathieu,
1988).
Cliquer sur l'image pour l'agrandir.
Figure
12 : Indice de diversité (d'après Murray,
1991) : hyposalin ‹ 33‰, normal = 33-37‰,
hypersalin › 37‰.
Cliquer sur l'image pour l'agrandir.
Figure
13 : Emplacement sur un diagramme triangulaire des biocénoses des
domaines infra- et circa- littoraux (d'après Murray,
1991).
Cliquer sur l'image pour l'agrandir.
Figure
14 : Provinces biogéographiques délimitées par la biodiversité des
foraminifères planctoniques actuels (modifié d'après Bé
& Tolderlund, 1971 ; Hemleben et alii,
1989). 1) province tropicale ; 2) province subtropicale ;
3) province de transition ; 4) province subpolaire ; 3) province
polaire ; 6) zones de résurgences côtières ("upwelling") ; 7) province subtropicale/tropicale.
Cliquer sur l'image pour l'agrandir.
Figure
15 : Paléobiogéoprovinces reconstituées avec les foraminifères
planctoniques crétacés à l'Albien terminal (d'après Hart,
2000, extrait modifié) : 1) tropicale (Téthysienne); 2)
de transition; 3) boréale (Nord) et australe (Sud).
Planches

Cliquer sur l'image pour l'agrandir.
Planche 1 :
Vues au MEB de morphotypes représentatifs des genres Spiroplectammina
(a), Bigenerina (b-c), Quinqueloculina (d et g), Parasorites
(e : vue oblique ; f : vue latérale), Borelis (h), Peneroplis
(i : vue latérale ; j : vue oblique), Spirolina (k) - Marge atlantique du Maroc. Échelles graphiques = 100 μm.

Cliquer sur l'image pour l'agrandir.
Planche
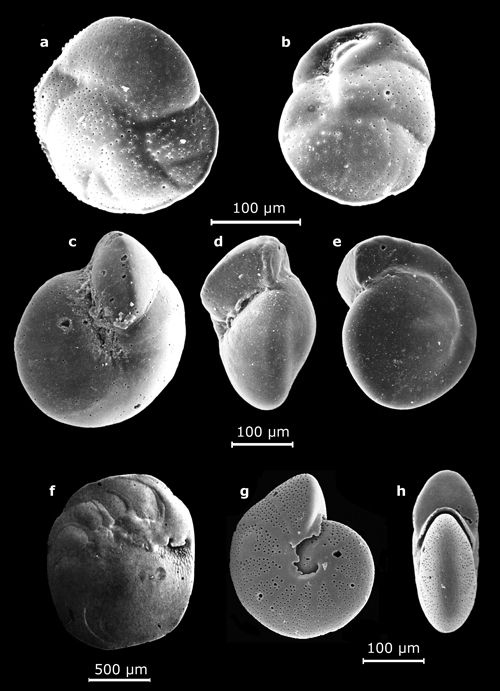
2 : Vues au MEB de morphotypes représentatifs des genres Oolina
(a), Bulimina (b : in
Mathieu, 1986, Pl. 8, fig. 10), Uvigerina (c : vue latérale in Mathieu,
1986,
Pl. 9, fig. 10), Lagena (d), Fissurina (e :
vue oblique ; g : vue apicale), Lenticulina
(f : in Mathieu, 1986, Pl. 6, fig. 10), Dentalina
(h), Amphicoryna (i), Bolivina (j-m) - Marge atlantique du Maroc. Échelles graphiques = 100 μm.
Cliquer sur l'image pour l'agrandir.
Planche
3 : Vues au MEB de morphotypes représentatifs des genres Cassidulina
(a : face spirale in Mathieu, 1986,
Pl. 10, fig. 7 ; b : face ombilicale), Gyroidina (c : face ombilicale ;
d : profil in Mathieu,
1986, Pl. 17, fig. 5 ; e :
face spirale in Mathieu, 1986,
Pl. 17, fig. 4), Amphistegina (f :
face ombilicale), Melonis (g : vue
latérale in Mathieu, 1986,
Pl. 14, fig. 10 ; h : profil in Mathieu,
1986, Pl. 14, fig. 12) - Marge atlantique du Maroc. Échelles graphiques = 100 μm,
sauf f = 500 μm.

Cliquer sur l'image pour l'agrandir.
Planche
4 : Vues au MEB de morphotypes représentatifs des genres Heterolepa
(a : face ombilicale ; b : profil in Mathieu, 1986, Pl.
15, fig. 3 ; c : face spirale), Ammonia (d : face
ombilicale ; e : face spirale), Cribroelphidium (f : vue oblique), Elphidium (g : vue
latérale in Mathieu, 1986, Pl. 16, fig.
10 ; h :vue oblique in
Mathieu, 1986, Pl. 16, fig.
11 ; i : détail de h) - Marge atlantique du Maroc. Échelles graphiques = 100 μm.
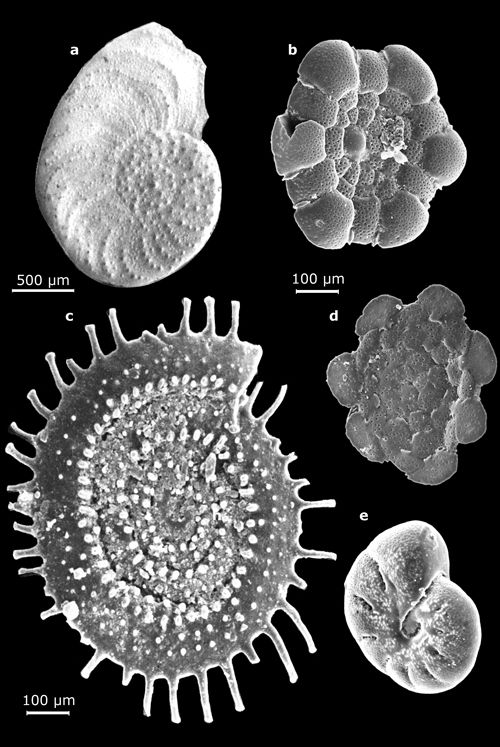
Cliquer sur l'image pour l'agrandir.
Planche
5 : Vues au MEB de morphotypes représentatifs des genres Operculina
(a :
vue latérale), Planorbulina
(b : face ombilicale in Mathieu, 1986, Pl.
14, fig. 2 ; d : face spirale in
Mathieu, 1986, Pl. 14, fig. 1), Spirillina
(c : vue latérale), Astrononion (e : vue oblique in Mathieu,
1986, Pl. 14, fig. 7) - Marge atlantique du Maroc. Échelles graphiques = 100 μm,
sauf a = 500 μm.
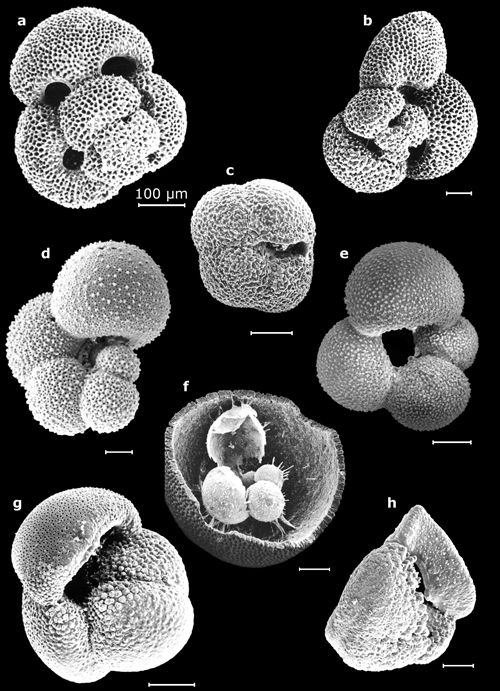
Cliquer sur l'image pour l'agrandir.
Planche
6 : Vues au MEB de morphotypes représentatifs des genres Globigerinoides
(a : vue spirale ; b: vue spirale in Mathieu, 1986, Pl. 18, fig. 3), Neogloboquadrina
(c : vue ombilicale), Globigerinella (d : vue
latérale in Mathieu,
1986, Pl. 18, fig. 5), Globigerina (e :
vue ombilicale in Mathieu,
1986, Pl. 18, fig. 7), Orbulina (f : vue
éclatée in Mathieu,
1986, Pl. 18, fig. 1), Globorotalia (g :
vue ombilicale ; h : profil in Mathieu, 1986, Pl.
19, fig. 11) - Marge atlantique du Maroc. Échelles graphiques = 100 μm.
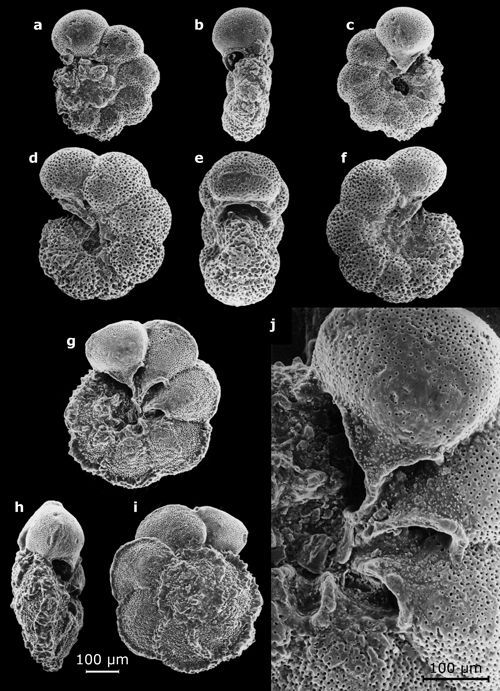
Cliquer sur l'image pour l'agrandir.
Planche
7 : Vues au MEB de morphotypes représentatifs des genres Ticinella
(a : face spirale ; b : profil ; c : face ombilicale), Biticinella (d et f : faces latérales ; e : profil), Rotalipora (g :
face spirale ; h : profil ; i : face ombilicale ;
j : détail de g, ouverture principale et ouvertures supplémentaires) - ODP Leg 171B, Site
1050. Échelles graphiques = 100 μm.
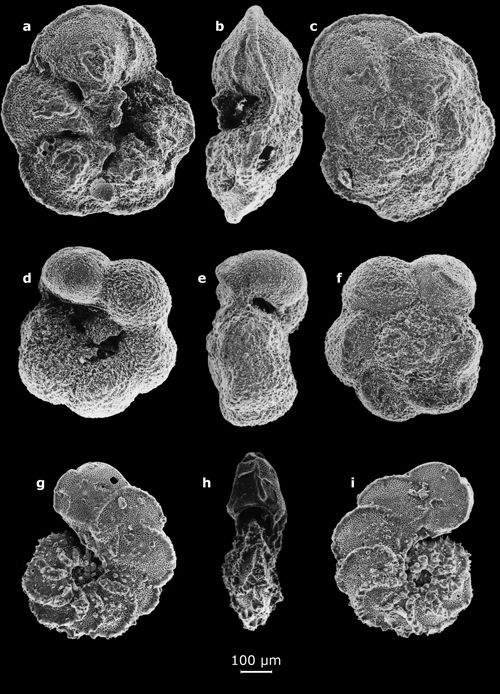
Cliquer sur l'image pour l'agrandir.
Planche
8 : Vues au MEB de morphotypes représentatifs des genres Rotalipora (a : face ombilicale ; b : profil ; c : face
spirale), Helvetoglobotruncana (d : face ombilicale ; e :
profil ; f : face spirale), Planomalina (g et i :
faces latérales ; h : profil) - ODP Leg 171B, Site 1050. Échelles graphiques = 100 μm.
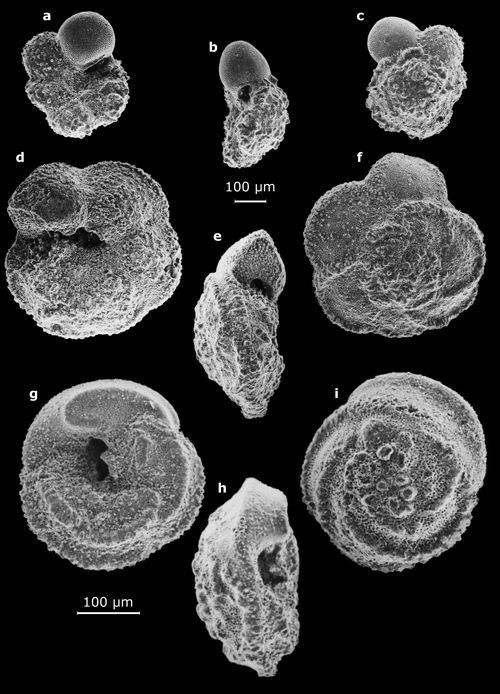
Cliquer sur l'image pour l'agrandir.
Planche
9 : Vues au MEB de morphotypes représentatifs des genres Praeglobotruncana
(a : face ombilicale ; b : profil ; c : face
spirale), Dicarinella (d : face ombilicale ; e :
profil ; f : face spirale), Contusotruncana (g : face
ombilicale ; h : profil ; i : face spirale) - ODP Leg 171B, Site
1050. Échelles graphiques = 100 μm.
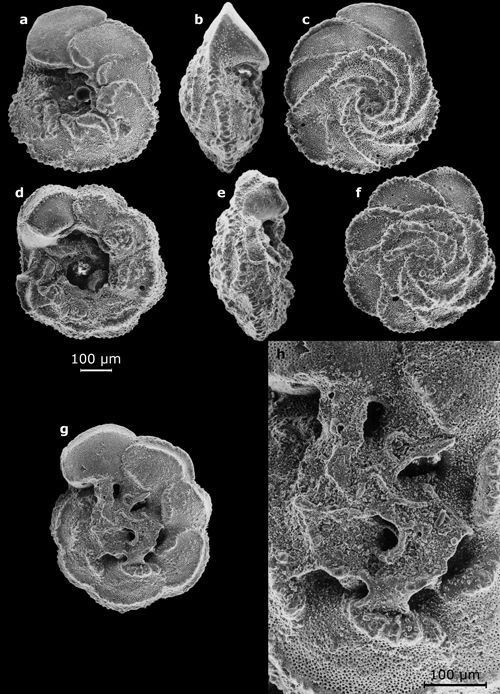
Cliquer sur l'image pour l'agrandir.
Planche
10 : Vues au MEB de morphotypes représentatifs des genres Globotruncanita
(a : face ombilicale ; b : profil ; c : face
spirale), Globotruncana (d et g : faces ombilicales ; e :
profil ; f : face spirale ; h : détail de g, plaques ombilicales) -
ODP Leg 171B, Site 1050. Échelles graphiques = 100 μm.
Citation
Bellier J.-P., Mathieu R. & Granier B. (2010).- Court traité de
foraminiférologie (L'essentiel sur les foraminifères actuels et fossiles) [Short Treatise on Foraminiferology (Essential on modern and fossil
Foraminifera)].- Carnets
de Géologie - Notebooks on Geology, Brest, Livre 2010/02 (CG2010_B02),
104 p., 15
figs, 10 pls.