(Tadeusz
Click on thumbnail to enlarge the image.
This publication is a contribution to the PETRALGA Project http://paleopolis.rediris.es/petralga/ |
Anisian Dasycladales from Upper Silesia
and adjacent regions
_
Zbigniew
Abstract: Anisian Dasycladales (calcareous algae) from the Diplopora Dolomite of the Upper Silesia and adjacent regions of S Poland are revised. All previously reported taxa are critically reviewed and illustrated. New paleontological samples were collected from 74 outcrops and from 45 boreholes. The abundant material includes both specimens visible on fractured rock surfaces and thin-sectioned ones; 24 species of Dasycladales are identified, including three new species: Oligoporella chrzanowensis n.sp., Physoporella polonoandalusica n.sp., and Salpingoporella krupkaensis n.sp. Best-preserved specimens are illustrated in 39 plates. The identified species were compared with Alpine and Carpathian forms of stratigraphic importance. Six Dasycladalean local horizons are defined. The Pelsonian-Illyrian boundary occurs in the middle part of the Diplopora Dolomite. Its uppermost part, despite the presence of Diplopora annulata, belongs to the Illyrian, not to the Fassanian, as also corroborated by conodont correlations. The peculiar state of preservation (internal moulds and double tubes) is discussed; it is due to early syngenetic dolomitisation. The palaeoenvironment of the algae is determined as sublittoral. Dasycladales flourished upon a peri-Tethyan carbonate platform, widely connected with the Alpine-Carpathian seas. Five palaeoecological assemblages are recognized, differing in their bathymetric and turbulence conditions, and living in marine shoals and shallow basins separating them. The Upper Silesian platform was separated from the hypersaline Germanic Basin of the Middle Muschelkalk by banks and oolite and bioclastic barriers, blocking dispersal of marine biota. A wide connection existed with the Alpine-Carpathian seas, allowing immigration of High-Tatric, Križna and even South Alpine flora to the Upper Silesian Carbonate Platform.
Key words: Dasycladales, Anisian, Diplopora Dolomite, Upper Silesia, algal zonation, palaeogeography
Abstrakt: Anizyjskie Dasycladales z Górnego Śląska i obszarów przyległych.- Dokonano rewizji oznaczeń Dasycladales z anizyjskiego dolomitu diploporowego Górnego Śląska i regionów sąsiednich. Zostały krytycznie przedyskutowane i zilustrowane wszystkie dotychczas opisane i wzmiankowane gatunki tamtejszych glonów wapiennych. Zebrano nowe materiały paleontologiczne z 74 stanowisk powierzchniowych ze wszystkich regionów występowania Dasycladales oraz z 45 wierceń, w których zostały znalezione te zielenice. Zebrany obszerny materiał składa się zarówno z okazów widocznych na ułamkach skalnych, jak i z okazów w płytkach cienkich. Oznaczono 24 gatunki Dasycladales, w tym 3 nowe: Oligoporella chrzanowensis, Physoporella polonoandalusica i Salpingoporella krupkaensis. Najlepiej zachowane okazy są zilustrowane na 39 planszach. Oznaczone gatunki porównano z gatunkami alpejskimi i karpackimi - z rejonów, gdzie mają one znaczenie stratygraficzne. Wydzielono 6 lokalnych poziomów diploporowych. Granica pelsonu z ilirem przebiega w połowie dolomitu diploporowego, a jego najwyższa część, mimo obecności gatunku Diplopora annulata, należy do iliru, a nie do fassanu, co wynika także z korelacji konodontowej. Omówiono charakterystyczny stan zachowania glonów (ośródki, podwójne rurki), który jest związany z wczesną, syngenetyczną dolomityzacją. Środowisko życia glonów określone zostało jako sublitoralne. Dasycladales rozwijały się na platformie węglanowej Perytetydy, mającej rozległe połączenie z morzem karpacko-alpejskim. Wyróżniono 5 zespołów paleoekologicznych żyjących w nieco odmiennych warunkach batymetrycznych i turbulencyjnych na mieliznach podmorskich i w dzielących je niegłębokich basenach. Górnośląska platforma była odgraniczona od salinarnego morza germańskiego środkowego wapienia muszlowego przez nasypy i łachy oolitowe i organodetrytyczne, tworzące bariery biologiczne. Istniało natomiast szerokie połączenia z morzem karpacko-alpejskim, przez które flora wierchowa i kriżniańska, a nawet południowoalpejska przenikała na górnośląską platformę węglanową.
Słowa kluczowe: Dasycladales, anizyk, dolomit diploporowy, Górny Śląsk, poziomy glonowe, paleogeografia
(Tadeusz
Click on thumbnail to enlarge the image.
This publication is a contribution to the PETRALGA Project http://paleopolis.rediris.es/petralga/ |
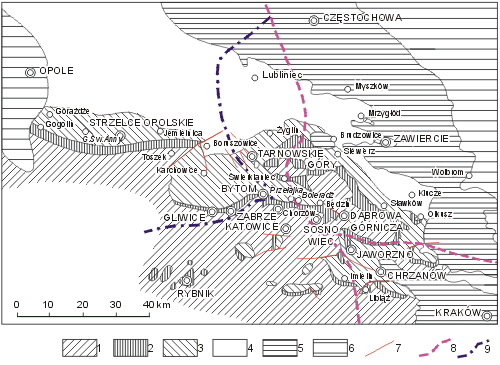
The Upper Silesia, together with adjacent regions of southern Poland, is the only place in Europe where Triassic Dasycladales appear in great abundance and diversity, besides the Alpine orogen, in the peri-Tethyan epicratonic basin. This basin, known since the 19th century as the Germanic Basin, embraced a large territory of the present-day Germany and Poland, reaching through France and Spain as far as to North Africa. The Upper Silesian Triassic yields abundant brachiopods, bivalves, gastropods, and especially Dasyclad algae with Alpine character, allows facial and stratigraphic comparisons of the specific tripartite Germanic Triassic with the classical marine Alpine strata from this period. Thus it is not surprising that the Upper Silesian Triassic attracted attention of geologists working on this system. Their work was greatly facilitated after the discovery of zinc and lead deposits in the Upper Silesia created numerous outcrops where fossil animal remains and Dasycladales were found (Fig. 1 ![]() ).
).
 |
Click on thumbnail to enlarge the image.
Figure 1: Schematic geological map of the Upper Silesian Triassic (modified from , 1973). 1 - Palaeozoic; 2 - Buntsandstein; 3 - Muschelkalk; 4 - Keuper and Rhaetian; 5 - Jurassic; 6 - Cretaceous; 7 - faults; 8 - state borders before World War One; 9 - state borders between the two World Wars.
In the 19th century the area where the fossil Dasycladales occurred belonged to three states: Germany (Prussia), Russia and Austro-Hungary, hampering their comprehensive studies. The earliest research into Triassic algae began in the Prussian Silesia near Strzelce Opolskie, Tarnowskie Góry and Bytom, where pioneering work was conducted by H. (1863), F. (1870), C.W. (1872a) and J. (1906). Soon the Dasycladales were discovered also in the Austrian partition, near Chrzanów ( & , 1866; , 1892). Presence of the Diplopora Dolomite was also noted in the Russian partition near Będzin and in a belt of outcrops from Olkusz to Siewierz (, 1907).
The first palaeontological identifications of the Upper Silesian Dasycladales (then called "Nulliporae", later collectively known as "Diploporae") were done by F. (1870), and especially by C.W. (1872a), who devoted a special monograph to their comparisons with the Alpine forms.
Serious difficulties arose from the different mode of preservation of the Upper Silesian and Alpine Dasyclads. While the Alpine specimens are preserved as calcareous tubes perforated with numerous pore whorls, the Upper Silesian ones are usually preserved as natural moulds and imprints, with tubercles instead of pores. Dasycladales fossilized in such a way were studied by C.W. , who was able to use observations of fractured specimens to recognize many species, some of them still valid. Many 's species were, however, questioned by the eminent specialist on Dasyclads, J. (1912, 1920, in & , 2013), who accepted only species established on thin-sectioned material (, 1931a).
The Upper Silesian Triassic was studied since pre-World War One through the interwar period and during the Second World War by P. (1913, 1926a, 1926b, 1944). He identified the Dasyclads found and consulted J. , by sending him specimens for determination, but lack of coordination in their studies led to many uncertainties and misunderstandings, especially concerning the stratigraphic importance of the Dasycladales.
After part of the Upper Silesia was incorporated to Poland as a result of poll after the World War One, the Dasyclads were studied by F. (1924), C. (1932), and mainly by S. (1935), who continued his research also after the World War Two ( & , 1968). At the same time, the Triassic of the Chrzanów Region was studied by S. (1949, 1952), who extended the lithostratigraphic scheme of P. onto the whole Upper Silesian Muschelkalk. The Muschelkalk of the Siewierz Area was studied by S. (1964). A single Polish paper concerns Dasycladales of the Chrzanów Region ( & , 1960). In Kąty Chrzanowskie (Rosowa Góra) they have found algae preserved as internal moulds and thus based their study on the monograph by (1872a), recognizing and redescribing several species established by him. The unification of the whole Upper Silesia within the territory of Poland after World War Two created opportunities to study all the Dasyclad localities by Polish geologists.
The present author's interest in calcareous algae dates back to the discovery of the first specimens in the Tatra Mountains in the Križna Triassic (, 1963), and then in the High-Tatric and Choč Triassic (, 1965a, 1965b, 1967). The author found his first Dasyclads in the Upper Silesian Muschelkalk in 1966 during work on an assessment of a dam on Biała Przemsza River in Przeczyce. New finds, in 1967, happened during the prospecting at "Kamień Wielki" Quarry within a study of the geological structure of the vicinity of Kamień Śląski to analyse the feasibility of locating there a waste dump for "Blachownia" chemical plant ( et al., 1967). The specimens found were so interesting that the author decided to study them in more detail, aiming at revising the Upper Silesian Triassic Dasycladales (, 1973). Working on Triassic Dasyclads from Bulgaria ( & , 1973), the author saw the necessity of revision of the calcareous algae described by Cz. and S. (1960). It became obvious that the form designated by them as Diplopora annulata var. physoporelloidea is, in fact, conspecific with Physoporella praealpina (, 1979, Pl. IV, 15).
In 1980, the author encouraged by H. , contacted geologists studying Muschelkalk deposits encountered in boreholes in the Zawiercie area, where zinc and lead deposits were intensely prospected by the Polish Geological Institute (a project led by L. ). The best preserved algae were collected by R. in 1980. In the same year, the author identified Acicularia sp. in thin sections from Zawiercie area collected by J. . In 1981, the author personally sampled numerous cores stored in Żarki. Initial results are based on well-preserved specimens of Physoporella praealpina , Physoporella dissita (), Oligoporella pilosa , Macroporella sp., Diplopora annulatissima and Diplopora cf. annulata (, 1981). Identification of the Dasyclads from cores provides critical data for establishing stratigraphic ranges of particular species. In the following years, the author obtained further specimens from R. , H. and W. . The specimens identified from 1982-1986 were published in a monograph "Geology of Poland" (, 1986, Pls. CV-CVI). In 1983, K. provided photographs of thin-sectioned specimens from her study on conodont stratigraphy of the Upper Silesian Muschelkalk (, 1975).
A comprehensive revision of the Dasyclad flora required extensive fieldwork. First, the author compiled a list of all localities where previous researchers found Dasyclads. Most details were gathered from papers by H. (1863), F. (1870), C.W. (1872a) and J. (1906), as well as by P. (1913, 1926a, 1926b, 1944), L. and C. (1866), K. (1907), F. (1924), S. (1935), and S. (1964). Precise location of sites was possible thanks to detailed maps, both German (, 1914, & , 1932, , 1938, et al., 1938; et al., 1914, 1915; et al., 1915) and Polish (, 1955; & , 1956; , 1978; , 1973a, 1973b, 1977; & , 1976a, 1976b; , 1957, 1968; , 1968a, 1968b) with explanations, as well as maps by S. (1935), P. & (1943) and S. (1964).
The author began his fieldwork in the autumn of 1988, accompanied by W. , J. , and W. . Locating individual sites in the field was aided by guidance of S. , then charting Upper Silesian Triassic sediments. We managed to collect samples from such important localities as Kamysz, Nowe Koszyce, Jemielnica, Czeladź, Przełajka, Wojkowice Komorne, Granice, Balin, Rosowa Góra and Stare Gliny. By 1989, first specimens were identified under binocular microscope and photographed.
In summer of 1989, the author, together with W. , J. and W. , located several more sites in the field and collected additional samples from localities visited in the previous year. We visited Libiąż, Imielin, Granice, Krasowy, Rosowa Góra, Stare Gliny, Luszowskie Góry (Krupka), Cezarówka, Będzin, Czeladź, Boleradź, Grodziec, Przełajka, Bytom and Repty. Noteworthy was relocating the important 's and 's locality in Las Segiecki (Segiet Forest). In 1989, the author identified and photographed selected specimens.
In summer of 1990, the author revisited many localities with W. , expanding the collections by sampling the Olkusz-Siewierz belt of Diplopora Dolomite outcrops at localities Bolesław, Krążek, Trzebiesławice, Warpie, Siewierz ("Wiktor Emanuel" Mine), Żelisławice, Brudzowice and Nowa Wioska. During the autumn of 1990, the author continued to determine and photograph specimens of Dasyclads from the collected samples and began to describe the localities with Dasycladalean flora, formulating conclusions about the stratigraphic importance of Dasycladales for the Muschelkalk . A report was written summarizing previous research on Middle Triassic "Diploporae" from Silesia-Cracow area (, 1990).
In 1991-1993, the author completed the photographic documentation and began describing already identified species of Dasycladales from the Upper Silesia and the Tatras. He communicated with B. , sending him materials necessary for a bibliographic inventory of Permian and Triassic Dasycladales he was compiling.
In summer 1994, during preparations to the Third International Meeting on Peri-Tethyan Epicratonic Basins in Cracow, the author and W. met with J. and H. to show them the localities in Przełajka and Boleradź. The author then wrote a report on Middle Triassic Dasycladaceae of the Upper Silesia-Cracow region and their stratigraphical and palaeontological significance (, 1994b) and led a field excursion to Przełajka, Boleradź and Stare Gliny for the Meeting participants.
In the autumn of 1994, W. informed the author about finding abundant and beautifully preserved Dasyclads in a quarry of the "Siewierz" mine in Brudzowice. The author visited the site with W. and H. , collecting many valuable specimens, which were later photographed in colour.
In 1995, the author joined the PETRALGA group, led by B. , and continued taxonomic determinations of the Dasycladales using recent literature. The systematic and comparative work continued in 1996-1998.
In autumn 1998 the author decided to expand the collection from previously studied localities. We revisited some sites with W. and J. , and identified further localities. The research covered all areas where the Diplopora Dolomite is present: the Upper Silesia, Cracow-Chrzanów region and Olkusz-Siewierz Monocline. Exceptional specimens were collected in the Segiet Forest, a renowned 's and 's locality, and in Suchodaniec, where a block of Diplopora Dolomite with well visible layering is protected as a nature monument. During the late autumn the same year, W. worked in the Tarnowskie Góry area. He collected samples from an old mine shaft Głęboka-Fryderyk and from Srebrna Góra. Thus, the list of the Diplopora Dolomite localities has been completed and a map could have been compiled of their distribution. The map with Polish sites and other materials have been sent to B. , who included them in the final version of the bibliographic inventory of Permian and Triassic Dasycladales ( & , 2000).
In 1999, Diplopora Dolomite sites were described from the Triassic of Opole Silesia, Tarnowice Syncline, Bytom Syncline, Cracow-Chrzanów region, Olkusz-Siewierz outcrops belt, and from boreholes in the Zawiercie area. The descriptions of localities and the results of all previous works are discussed as well as listing currently determined Dasycladales are included.
Thus almost all the Silesian Dasyclad localities mentioned in the literature have been reviewed, together with newly discovered sites. Special attention was paid to the old localities, from where originated the first "Diploporae" described by German authors. This is especially important, because the holotypes of Upper Silesian species have been mostly lost, while their illustrations are of imperfect quality or are interpretive drawings. Collecting new material from the type localities created an opportunity to find specimens of the described species and revising them.
In 2000, a collection of Dasyclads on matrix fragments was assembled and transferred to the Geological Museum of the Polish Geological Institute. The specimens were sectioned and the thin sections are also housed in the Geological Museum, PGI. In 2000 and 2001, the author began the systematic descriptions of formerly established species using the new abundant data, as well as descriptions of new species from Chrzanów region and the Opole Triassic (from Kamień Śląski). Systematic arrangement of the described material was easier due to the new important study by & (2000) and the book on Dasycladales by S. and M. (1992).
In 2002, photographic plates were assembled, together with extensive captions, as well as the table of relative abundance of Dasyclads from the localities studied. The algal horizons were recognized and a Dasyclad-based stratigraphic scheme was compiled. The same year, W. conducted additional fieldwork near Bytom and Szarlej, where most old German localities are inaccessible due to mining and metallurgical waste dumps.
In 2003 and 2004, the final editing took place and the text was translated to English. The author worked also on a paper on Diplopora Dolomite localities to be published soon after printing this monograph, thus summarizing four decades of the author's research on Dasycladales of the Upper Silesia and adjacent regions.
[The author died on June 18th, 2005 and the editing has been continued mostly by Karol (who translated the paper) and Witold , coauthor of a parallel paper in Polish.]
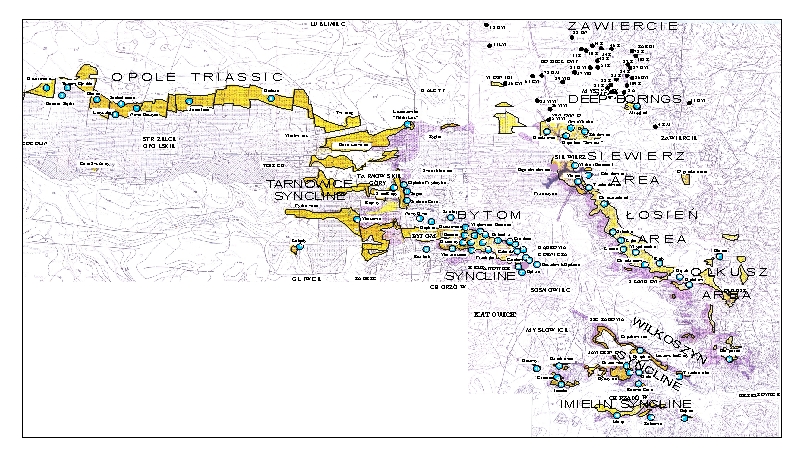
The studied Dasyclad material comes from 75 natural and man-made outcrops, as well as 35 borehole cores scattered throughout the Upper Silesia and adjacent regions. The localities (Fig. 2 ![]() )
should be described in more detail elsewhere. Relative abundance of the Triassic algal fossils in localities studied is summarized in Fig. 5
)
should be described in more detail elsewhere. Relative abundance of the Triassic algal fossils in localities studied is summarized in Fig. 5 ![]() .
.
 |
Click on thumbnail to enlarge the image.
Figure 2: Map of surface and subsurface localities with Anisian Dasycladales (background map from serial geological maps in 1:200,000 scale, sheet Gliwice (, 1977) and sheet Kraków (, 1978). 1 (yellow) - outcrops of Diplopora Dolomite (Jemielnica Formation); 2 (pink) - outcrops of Karchowice Formation; 3 (blue dots) - outcrops of Ore-bearing Dolomite: 1-75, numbering of surface localities; 4 (black dots) - 27 boreholes with Dasycladales: 14-Ż, for instance.
The specimens are housed in the Geological Museum of the Polish Geological Institute in Warsaw (acronym: MuzPIG), as two collections: MuzPIG 1653.II (consisting of 563 rock samples with more than 3000 identifiable specimens, assuming 3-10 Dasyclads per rock fragment) and MuzPIG 1682.II (210 thin sections with about individual 800 algal specimens; 3-4 identifiable specimens per one section). Thus, both collections comprise about four thousands fossil Dasyclads.
About 1500 specimens were photographed; about half of them are illustrated here in 50 black-and white and colour photographic plates. In addition, photocopies were made of all previously published illustrations of Dasycladales from the Upper Silesia and adjacent regions. The figures were scaled to facilitate comparisons and taxonomic determinations.
Dasyclads of the Upper Silesia and adjacent regions are preserved in a peculiar way. Usually fossil calcareous algae retain their calcareous sleeves, but the studied specimens, are preserved as internal moulds (steinkerns) or hollows ("ghosts"); without their calcitic skeletons. Moreover, besides dissolution, during diagenesis, dolomitization occurred. The latter process, responsible for the formation of the Diplopora Dolomite, preserved the general shape of the algae, but obscured some details of their morphology.
Based on these dolomitized steinkerns, the first students of Upper Silesian calcareous algae (, 1862; , 1870; , 1872a; , 1906), regarded the "Diploporae" as foraminifers. They, were able to properly interpret the three-dimensional structure and even some details. W. recognized several species, of which some are still valid. His very accurate drawings are reproduced in some text-figures herein.
A prominent researcher of Alpine and Balkanian Triassic Dasyclads, J. (1912, 1920, in & , 2013) studied calcareous algae in thin sections of limestone. The calcitic skeletons are perfectly preserved in these rocks, allowing to develop new standards of detailed morphological descriptions of fossil Dasycladales and their taxonomy. Study of thin-sectioned Dasyclads has many advantages and became a basic procedure applied by most researchers. The disadvantage, however, was that the sections were randomly oriented - transverse, longitudinal, and oblique at various angles - resulting in dubious restorations and questionable taxonomic attributions (, 1964, p. 83). Because of these problems, the scientists working on Alpine Dasyclads looked for specimens on weathered rock surfaces, revealing the general shape of calcareous sheaths, their segmentation and other morphological details (see , 1965, Pls. 46 & 54; , 1965, tab. XII, 7).
Rarely, for example, in Monte Popera (Eastern Dolomites), the Dasyclads were silicified (E. , 1979, Pls. 4-6), allowing one to observe in detail variously oriented algal fragments, especially the arrangement and shape of branches or the pattern of pores remaining. Silicification could have affected both the algal walls (E. , 1979, Pls. 4-6) and steinkerns (E. , 1979, Pl. 5). Such preservation permits studying the general habitus of the algae and their internal morphology in various aspects.
Some scientists use acetic acid to etch the dolomitized Dasyclad thalli from the matrix. This method was efficiently applied by F. (1958) in his studies on calcareous algae and fauna of the Briançon Triassic and of Préalpes Médianes in Switzerland. He successfully prepared and identified the Dasycladales from three Anisian and Ladinian zones (, 1958, Pl. 4, 5-15; Pl. 6, 12-21; Pl. 7, 17-26), entirely without thin-sectioning the material. F. (1958) and M. (1954) did not share the opinion of J. , that thin sections provide the only reliable method of precise taxonomic determination of Dasyclad algae. They believed that in some cases observation of fractured and etched surfaces is sufficient (see also , 1961, fig. 6). If the pores are large enough, the delicate structure of slightly secondarily recrystallised specimens can be easier observed this way rather than in the sections., The human eye visualizes three dimensional surfaces rather than flat outlines in cross sections (, 1958, p. 177). In some cases, even minute canals (pores remaining after decay of branches) can be seen in chemically treated specimens. Given the general scarcity of Briançon fossils, using the acetic acid etching enabled F. to retrieve and identify gastropods and Dasyclads - "Exploiter un gisement, un nid fossilifère n'est plus dès lors qu'une affaire de patience, de loisirs et d'acide" (, 1958, p. 173).
Chemical preparation method was employed in studies on Carboniferous Dasyclad morphology by S. (1984, 1986, 1987). He studied the algae mostly in thin sections, but performed additional observations on specimens etched with 10% solution of acetic acid (, 1984, figs. 3, 4; 1986, Pl. 9; 1987, Pls. 3-6). Observations of chemically prepared pyritized steinkerns provided the basis for graphic restorations of the actual external appearance of the algae (, 1987, Pl. 3, 3b; Pl. 4, 1b, 5b; text-figs. 1, 2), making it difficult to reconstruct using only randomly oriented cross-sections in thin-sectioned material. In his opinion (, 1986, p. 27), "the taxonomy of any group, based only on the features recognizable in thin sections, will always cause doubts, because the differentiation of sections is usually greater than the variety of real forms in particular fossils. A relation between thin sections and prepared specimens is also unidirectional, i.e., it is possible to find the shape of sections of the calcareous sleeve on basis of internal moulds (, 1986, Pl. 3, fig. 3 and 4 and Pl. 4, fig, 1, 5), but inversely it is difficult."
The mode of preservation of the Upper Silesian Dasyclads is such, that there is no need to apply chemical preparation methods. The algal fossils are well visible on naturally weathered surfaces and on surfaces of freshly fractured dolomite.
The Upper Silesian Diplopora Dolomite is a porous rock. The porosity is due to hollows left after dissolution of gastropod, bivalve, coral and Dasyclad skeletons during dolomitization. Very frequently these are mere "ghosts" whose shape allows recognizing their Dasyclad origin, but their identification of species or even at genus level using thin sections is hardly possible (Pl. XXI ![]() ). Thin sections are useful in rare case of less porous dolomite or preservation in limestone
(Pls. V
). Thin sections are useful in rare case of less porous dolomite or preservation in limestone
(Pls. V ![]() - VI
- VI ![]() ,
XXII
,
XXII ![]() ,
XXV
,
XXV ![]() & XXIX
& XXIX ![]() ). Generally, however, fractured specimens of Dasycladales are much better material for taxonomic studies.
). Generally, however, fractured specimens of Dasycladales are much better material for taxonomic studies.
Several modes of preservation of the Dasyclads visible in fractures can be recognized (, 1981).
Most common are steinkerns, the internal moulds of algae, with the inner axial cavity partly filled with dolomitized sediment or dolomitic spar and covered with tubercles being moulds of pores left after primary branches. The original calcareous sleeve was completely dissolved. The tubercles may sometimes be hollow inside. Such preservation was noted by C.W.
(1872a) and C. and S. (1960), see Figs. 12 ![]() - 13
- 13 ![]() - 14
- 14 ![]() - 15
- 15 ![]() - 16
- 16 ![]() - 17
- 17 ![]() - 18
- 18 ![]() . Dasycladales preserved in such a way can be seen in many photographic plates herein.
. Dasycladales preserved in such a way can be seen in many photographic plates herein.
Fairly rarely the original calcareous sheath can be observed, sometimes dolomitized. It is this mode of fossilisation that allows species identification based on thin sections. Using the latter method, J. (1931a) identified only three Dasycladalean species (Diplopora annulata, Diplopora annulatissima and Diplopora elegans) from the Upper Silesian Triassic, rejecting all taxonomic attributions by C.W. (1872a) based on specimens observed on fractures.
Such a dolomitized sleeve reveals both the inner and outer shape of the alga. Inside the calcareous tube, the pores (inlets of primary branches) are visible; sometimes their shape is also recognizable. Such preservation is most often observed among Physoporellae
(Pls. XVIII, 6 ![]() &
XIX, 3, 7
&
XIX, 3, 7 ![]() ). It is similar to that of 's (1958) acid-etched specimens from the Briançon Triassic (Fig. 18a, p, r
). It is similar to that of 's (1958) acid-etched specimens from the Briançon Triassic (Fig. 18a, p, r ![]() ), to similarly prepared specimens from Swiss Alps (,
1961, fig. 6), and to silicified specimens of Kantia comelicana () from Eastern Dolomites (Fig. 22d, e, f
), to similarly prepared specimens from Swiss Alps (,
1961, fig. 6), and to silicified specimens of Kantia comelicana () from Eastern Dolomites (Fig. 22d, e, f ![]() ).
).
Another common mode of preservation involves formation of a dolomite crust both inside and outside the original calcareous sheath, which later underwent dissolution. Thus a double dolomitic tube was formed, joined by tubercles resulting from partial or complete dolomitization of pores (sometimes hollow - Pl. XIII, 13-16 ![]() ), preserving the original shape of primary branches
(Pls. XVIII, 1-2
), preserving the original shape of primary branches
(Pls. XVIII, 1-2 ![]() &
XXXV, 1
&
XXXV, 1 ![]() ). Such preservation is very advantageous, because it allows one to observe both the inner tube with intusannulation
(Pls. XIV, 5, 10, 14
). Such preservation is very advantageous, because it allows one to observe both the inner tube with intusannulation
(Pls. XIV, 5, 10, 14 ![]() &
XVI, 31
&
XVI, 31 ![]() ) and pores (branch inlets), and the outer surface of the sleeve, with its annulation (Pl. XXVI
) and pores (branch inlets), and the outer surface of the sleeve, with its annulation (Pl. XXVI ![]() ), fissuration or lack thereof. Most important is the fact that in such a double tube the shape of tubercles can be precisely determined
(Pls. XIV
), fissuration or lack thereof. Most important is the fact that in such a double tube the shape of tubercles can be precisely determined
(Pls. XIV ![]() ,
XXIII
,
XXIII ![]() -
XXIV
-
XXIV ![]() &
XXVIII
&
XXVIII ![]() ), and in case of the outer tube breaking away - also the arrangement of tubercles on the inner tube. Even though the shape of an alga underwent some distortions during dolomitization and the tubercles do not exactly reproduce the shape of branches, such preservation allows species identification. Moreover, the three-dimensional view of the algae enhances our understanding of their inner and outer morphology, shape of the apices, their bending, details of tubercle patterns inside the sleeve (in some species, like Kantia comelicana () the branches formed tufts;
Pls. XXIII, 7-8, 10
), and in case of the outer tube breaking away - also the arrangement of tubercles on the inner tube. Even though the shape of an alga underwent some distortions during dolomitization and the tubercles do not exactly reproduce the shape of branches, such preservation allows species identification. Moreover, the three-dimensional view of the algae enhances our understanding of their inner and outer morphology, shape of the apices, their bending, details of tubercle patterns inside the sleeve (in some species, like Kantia comelicana () the branches formed tufts;
Pls. XXIII, 7-8, 10 ![]() &
XXIV, 1, 8, 12, 14-15
&
XXIV, 1, 8, 12, 14-15 ![]() ), and of the structural details of the furrows and interannular lists, depending on the position and shape of branches in whorls (see, e.g., for Diplopora annulatissima -
Pl. XXIV, 1-2, 5-6, 13, 16
), and of the structural details of the furrows and interannular lists, depending on the position and shape of branches in whorls (see, e.g., for Diplopora annulatissima -
Pl. XXIV, 1-2, 5-6, 13, 16 ![]() ).
).
The presence of double tubes in some preservational variants of Upper Silesian Dasycladales was first noted by C.W.
(1872a, p. 92-93) in his description of Gyroporella cylindrica (=Diplopora annulatissima ). His detailed description, appended with numerous drawings (see Fig. 23t ![]() ) allowed to discern such morphological details that are not noticeable in thin sections.
) allowed to discern such morphological details that are not noticeable in thin sections.
The above overview of various modes of preservation of the Upper Silesian Dasycladaleans shows that their species can be identified not only in thin-sectioned specimens, as claimed by J. (1920, 1931a), but also in commonly occurring fractures. If we do not want to neglect huge numbers of Dasyclad specimens, constituting a valuable palaeontological material, we should return to taxonomic designations of C.W. (1872a) and C. and S. (1960), based on algal steinkerns. We should accept the arguments of F. (1958) and M. (1954) who valued the possibility of three dimensional observations of the algae enabled by chemical preparation methods. Nature itself gave us precious palaeontological material due to unique preservation which the present author strived to exploit to its fullest potential.
The monograph by S. and M.J. (1992, p. 17-19; see also , 2012) provides important information on calcification pattern in Recent Dasycladalean algae: "Recent Dasycladales calcify in their natural habitats, although to a different extent. Some Dasycladales develop a strong calcareous skeleton covering nearly the whole thallus, like Cymopolia barbata. Other Dasycladales only develop an extremely fine calcareous coat, like Dasycladus vermicularis. Depending on environmental conditions this fine coat may even be missing. However, in any case, it is a biogenic formation of calcium carbonate which justifies the classification of Dasycladales into the true calcareous algae" ( & , 1992, p. 17).
The skeleton may consist of magnesium calcite or aragonite; the amount of magnesium is temperature-dependent, which allows palaeontological statements to be made. The calcite originates in the diagenesis of unstable aragonite. Early diagenesis happens rather frequently in sediments. The conversion of aragonite into calcite may already occur in the older parts of the skeleton while the thallus is still alive. The percentage of calcite preservation increases in correlation to increasing geologic age. Aragonite is found extremely rarely in skeletons of older geologic formations. The oldest skeletons of Dasycladales that have retained a small amount of the original aragonite mineralogy are found in Triassic strata.
The deposit of biogenic calcium carbonate in calcareous algae may be observed in different places. For example, the mineral may be found mostly or exclusively in cell walls. Other algae may deposit the carbonate mostly or solely in the mucilage. Both cases involve mineralization in crystalline form. However, the lumen of the cell may also be partially filled. The formation of intracellular CaCO3 crystals is initiated in the vacuoles ( & , 1992, fig. 2.8). From here crystals are partly transported and deposited in the cell wall or mucilage cover. Either the complete mucilage may calcify, or only a more or less thick layer around the main axis, laterals and gametophores or gametangia. In Acetabularia, mineralization only occurs in the thin, cortex-like mucilage of the cap rays. Intracellular mineralization of Acicularia has a high value for systematic classification of the algae. The protective function of calcification of productive organs in Dasycladales is evident.
A continuous calcareous coat of fossil specimens is occasionally termed cortex. This coat is formed by incrustation where touching laterals are not, or at least not completely, incrusted.
& (1992, p. 19) supposed that in the fossil Dasycladales only the mucilage and not both the mucilage and cell wall calcified.
Observations on dolomitized Upper Silesian Triassic algae suggest, however, that also the cell wall was calcified. Especially revealing is the preservation as double tube (tube-within-tube), with dolomite coat formed both on the inside and outside of the original calcareous sleeve which was later dissolved. The inner tube allows seeing the pores (branch inlets) and often also intusannulation. Such observations are best made on Diplopora annulatissima
(Pls. XXVI ![]() -
XXVII
-
XXVII ![]() -
XXVIII
-
XXVIII ![]() ) and Clavapora clavaeformis
(Pls. XXX
) and Clavapora clavaeformis
(Pls. XXX ![]() -
XXXI
-
XXXI ![]() ). Obviously, the inner dolomitic coat preserves the inverse relief of the calcified cell wall from the side of the cell lumen. Such preservation shows also the dolomitized casts of the branches connecting the inner and outer tubes, because the actual calcareous sleeve has been dissolved. Observations of specimens with preserved original calcareous wall demonstrate that the primary branches were embedded precisely in that wall and not in the mucilage. Thus, there is convincing evidence that the cell wall was indeed calcified. It is true also for other species of the Upper Silesian Triassic Dasycladaleans, and especially well visible in the Physoporellae and in Kantia comelicana
(Pls. XXIII
). Obviously, the inner dolomitic coat preserves the inverse relief of the calcified cell wall from the side of the cell lumen. Such preservation shows also the dolomitized casts of the branches connecting the inner and outer tubes, because the actual calcareous sleeve has been dissolved. Observations of specimens with preserved original calcareous wall demonstrate that the primary branches were embedded precisely in that wall and not in the mucilage. Thus, there is convincing evidence that the cell wall was indeed calcified. It is true also for other species of the Upper Silesian Triassic Dasycladaleans, and especially well visible in the Physoporellae and in Kantia comelicana
(Pls. XXIII ![]() -
XXIV
-
XXIV ![]() ).
).
Calcification and subsequent dolomitization of mucilage is visible in
phloiophorous Salpingoporella krupkaensis (Pls. III ![]() -
IV
-
IV ![]() ), with occasionally preserved cortex, and in some modes of preservation of Diplopora annulatissima with calcified secondary and tertiary branches within the cortex (Favoporites).
), with occasionally preserved cortex, and in some modes of preservation of Diplopora annulatissima with calcified secondary and tertiary branches within the cortex (Favoporites).
Observation of the Upper Silesian Dasycladales is helpful in resolving the question whether the calcification started from the outer zone of the cell, as envisaged by J.
(1927a, p. 105), or rather from the inside of the cell, as assumed by most modern phycologists. In some specimens of Diplopora annulatissima (Pl. XXVIII ![]() ) it is evident that the shape of interannular furrows depends on arrangement of branches from which the calcification started.
) it is evident that the shape of interannular furrows depends on arrangement of branches from which the calcification started.
J. (1920, p. 77) wondered whether there were branches in the interannular furrows. Of course, it is impossible, because the calcareous coat began its formation only around the branches, in depressions turning into rings due to their calcification. The dolomite crust filled mainly the interannular furrows and surrounded the calcified branches, forming hollow tubercles. saw the process similarly (, 1920, p. 164), stating that the formation of calcareous skeleton started from branches arranged in whorls, and then their calcareous tubes merge into a continuous crust. It is clearly visible that the calcareous sleeve of the thallus consists of merged tubes surrounding branches. Later dolomitization, resulting in formation of the double tube and hollow tubercles, then proceeded from outside of the already dead alga, from interannular furrows, and from inside, from the interior chamber.
According to M. (1972, p. 94), the pores, as negatives ("ghosts") of the calcareous skeletons, formed in the final phase of dolomitization. His conclusion pertained to negatives of Dasycladales preserved in Triassic Dolomites of the Central Carpathians. It is relevant also for the Upper Silesian dolomite, where the algal calcareous sleeves were dissolved already after the double dolomitic tubes encrusted them. Both processes resulted from early diagenetic dolomitization.
This paper aims at redescribing known species and describing new ones, focused on peculiar preservation of Dasyclad algae in the Upper Silesian Triassic. The description should be as detailed as possible and enabling comparisons with other taxa. Thus, diagnostic features need to be selected and illustrated.
The fullest and most objective documentation used by all phycologists is the photographic one. It is an indispensable part of formal description of a new fossil species. This paper is supplemented with numerous photographs, mostly black-and-white, but also in colour. All photographs of specimens visible in fractures are accompanied with explanations much more detailed than simple captions to the pictures of thin-sectioned specimens. Spatial preservation seen on photographs reveals both the general and the detailed morphology; the figure captions point to the most important diagnostic characters visible in particular pictures.
A simple way to reduce the need for wordy description is making a diagrammatic sketch, accentuating the most characteristic features of a given alga and downplaying unimportant ones. The older students of Upper Silesian Dasycladales, such as C.W. (1872a) published many drawings in their papers. The drawings were later reproduced in simplified versions (e.g., , 1928, 1938). Reproductions of these illustrations are shown in this paper as text-figures. It should be noted that also J. (1912, 1920, 1935a, in & , 2013) provided illustrations of species he described based on thin sections, as hand-shaded drawings of the actual observed sections supplemented with restorations. The method of C.W. (1872a) was employed also by C. and S. (1960), including drawings and restorations besides photographs of the algae. Also S. (1984, 1986, 1987) provided restorations of the Dasyclads, more reliable than that based on fractured specimens and not just on cross sections, like those by J. (1920, 1935a).
Some researchers (, 1967; , 1977) applied statistical approach to Dasyclad studies, with very interesting results. However, employing quantitative methods in the case of the Upper Silesian species described herein is very difficult and mostly futile, because only steinkerns or dolomitized remnants of calcareous sleeve dissolution are preserved. Thus, even the most detailed measurements advocated by J. and applied by other researchers on thin sections (e.g., J. , 1964, p. 85), cannot be fully used. Only general division into small (less than 1 mm diameter) and larger (more than 1 mm diameter) forms is practical. Sometimes also the D:d ratio (outer diameter of an alga to the axial cavity diameter) can be calculated. The choice of measured parameters and their precision is limited by the fossils' state of preservation.
The terminology used in descriptions of Dasycladales has been established by J. (1912, 1920, in & , 2013) and is applied by all students of fossil Dasyclads (e.g., , 1963; , 1964; , 1972b, 1972c) and by phycologists working on Recent material (e.g., & , 1992).
Almost all of the terms used for describing thin-sectioned material can be applied to the Upper Silesian Dasycladaleans. The described steinkerns allow one to discern, e.g., annulation, intusannulation, as well as aspondyl, euspondyl and metaspondyl morphology of Dasycladalean thalli, etc. Instead of laterals (branches) there are pores or tubercles, whose shape can be described as phloiophore, vesicular, pyriform, trichophore and acrophore, which is helpful in genera and species identification. There are also additional terms related to dolomitization, such as double tube (tube-within-tube) with inner and outer crust (internal and external wall).
The proportions of certain measurements to the whole thallus, e.g., the diameter of the main axis to the diameter of the thallus, or height of the thallus even the diameter of the laterals to the length of the laterals, are frequently species-specific.
Given the intraspecific variability, a species diagnosis cannot be based on description of a single type specimen. Beside the holotype, paratypes are designated, taking into account different orientation of specimen or different mode of preservation. The large number of described and illustrated specimens allows covering a wide spectrum of variability. A group of conspecific individuals constitute a hypodigm ( & , 1978). All specimens within the hypodigm, including previously designated type specimens, are of equal importance in the species description. Of course, it is not practically possible to describe and illustrate all hypodigm specimens for most species. This problem is partly solved by opening a species description with synonymy. It rarely lists the whole hypodigm but offers its brief overview, useful for comparative purposes. The text-figures present illustrations of all the Upper Silesian Triassic Dasycladales previously pictured in scattered and hardly accessible publications. Together with synonymy and numerous original photographs they are representative for the whole hypodigm.
The species descriptions, necessarily, include such information as derivation of the name, holotype, paratypes, neotypes, syntypes, their repository and collection numbers, diagnosis and nomenclature, material and localities; descriptions, comparisons and discussion, stratigraphical range and geographical distribution.
The principles of Dasycladalean (originally - Dasycladacean) classification are based on the proposals by J. (1920), amended by later researchers, such as M. (1957), J. (1964) and T. (1979). Great advances were made by French scientists ( et al., 1975, 1977, 1979; , 1988; & , 1995) who compiled critical inventories of previously described species and proposed new systematic divisions of Dasycladaceae, elevated by (1988) to the rank of Order Dasycladales. The latest critical bibliographical compilation is that of Permian and Triassic Dasycladales by & (2000). The extensive synonymy compiled by the latter authors is basically accepted here, except for some expansion or adaptation to the needs of describing Upper Silesian Triassic specimens. The taxonomic divisions are largely following those presented in the "Dasycladales" monograph by S. and M.J. (1992), including the convenient subdivision into families, tribes and subtribes, together with the authorities of taxa (see List of described species).
J. (1920, p. 172) noted the second order metamerization: the whorls are paired, and each double whorl is separated from the next pair by a substantial distance. Such metamerization occurs in the tribe Oligoporellinae, where it provided the basis for establishing new species. Within the genus Oligoporella, three groups of species can be discerned: Oligoporella prisca group (with single whorls; O. prisca, O. elegans, O. chia), Oligoporella pilosa group (double whorls; O. silesiaca, O. balinensis) and Oligoporella chrzanowensis group (indistinct whorls or aspondyl arrangement of branches; only the nominal species). Similar groups can be recognized among species of the genus Physoporella: Physoporella pauciforata group (Ph. pauciforata with single, densely spaced whorls and Ph. lotharingica with widely spaced single whorls) and Physoporella praealpina group (double whorl species: Ph. praealpina, Ph. dissita, Ph. minutula and a new species Ph. polonoandalusica with widely spaced double whorls).
Possibly the above outlined groups could be regarded as subgenera within Oligoporella and Physoporella, respectively.
The significance of the Dasycladalean algae for stratigraphic subdivision of the Middle and Upper Triassic is known from numerous publications of (1927a, 1930a, 1931a, 1931b, 1936, 1937a, 1942), and students of geology of the Alpine region (, 1958, 1963; , 1963; & , 1963; , 1972a, 1972b, 1972c, 1974), Balkans (, 1958, 1965; & , 1973) and the Carpathians (, 1964, 1986; & , 1964; & , 1974; , 1970; & , 1973; , 1981; , 1967, 1977, 1979, 1986). They are particularly useful for stratigraphic zonation of calcareous and dolomitic rocks of great thickness, devoid of other fossils. Dasycladales made possible the stratigraphic subdivision of the Triassic carbonate massifs in all tectonic-facial units of the West Carpathians (, 1986). Also in the Tatra Mountains they provided basis for Triassic subdivision and enabled the deciphering of extremely complicated tectonics of napped and scaled carbonaceous rocks (, 1959, 1963, 1965b, 1973).
Nevertheless, despite those successes of Diplopora-based stratigraphy, in recent years their stratigraphic significance was questioned. Several species were demonstrated to be closely associated with facies; this facies-fidelity influenced their regional stratigraphic ranges (, 1967, 1972c; & , 1970). There even appeared an opinion that the Dasycladales were not applicable for the division of the Triassic at all (, 1971). J. (1999) had not even mentioned Dasycladalean algae among the Triassic index fossils. He also does not believe in the reliability of correlation between the Upper Silesian Muschelkalk with the Alpine Middle Triassic based upon them. Even though the correlation was traditionally based on Dasyclads, he rejects it even within the Diplopora Dolomite, containing the Pelsonian-Illyrian boundary. Basing the correlation only on conodonts and echinoderms led to erroneous conclusions, corrected by J. and J. (2000, fig. 8) according to the data of (1994a).
Such situation was the reason, that J. (1986) devoted his last, posthumous publication to the importance of Dasycladalean algae for Triassic stratigraphy.
The importance of particular species for identifying Triassic zones and subzones was noted long ago; e.g., (1920, 1936) assumed that Physoporella pauciforata is characteristic for the Anisian and Diplopora annulata for the Ladinian. However, (1972a, 1972b) noticed that the latter species occurs already in the Upper Illyrian (ammonite Aplococeras avisianus Zone) and survives up to Longobardian. Diplopora annulatissima, regarded by (1930a, 1931a) as index fossil for the Illyrian, was observed by (1965) to co-occur with Diplopora annulata in the Ladinian. In the Briançon Triassic of the Western Alps, (1958, 1963) discerned three diploporan zones - the lowermost with Anisoporella, the middle one with Physoporella praealpina and Diplopora annulatissima (Pelsonian-Illyrian), and the uppermost with Diplopora uniserialis (Ladinian). Teutloporella herculea was regarded the index fossil for the Upper Ladinian (, 1936, 1942), but (1967) extended its range up to the Carnian or even Norian. Clypeina besici is the index species for the Carnian (, 1975), similarly as the Andrusoporella duplicata, while Chinianella and Diplopora phanerospora are index taxa for the Norian (, 1967).
Scattered information about the importance of particular Dasycladales for the stratigraphy of the Eastern Alps was systematised by E. in his stratigraphic table (, 1972a, tab. 2). He recognised four distinct floral assemblages.
1. Physoporella-Oligoporella assemblages - Anisian ("Hydasp"-Lower Illyrian, extinction in the Trinodosus zone).
2. Diplopora annulata group and Teutloporella nodosa, beginning with the Avisianus Zone in the uppermost Anisian and ending in Ladinian, below the Cordevolian.
3. A Carnian flora, containing especially Poikiloporella duplicata (= Andrusoporella duplicata ) and Clypeina besici.
4. A flora of Upper Norian and Rhaetian age, containing endospore species of Diplopora (D. phanerospora, D. tubispora) and species of the genus Chinianella, showing whorls of sterile and fertile branches.
As for Diplopora annulatissima, (1972b) believed it appeared in the Middle Anisian, reached the maximum abundance in the Upper Illyrian and survived up to the Ladinian. E. (1972a, 1972b), like M. and O. (1969) and later O. et al. (1980), followed the distinction between one-branched and two-branched verticil [= single-whorl and double-whorl] species among the Physoporellae and Oligoporellae allowing to discern assemblage zones, and pointed to their importance for the Anisian stratigraphy. This approach was then developed by J. (1982) for the West Carpathians. He correlated the Dasycladalean algal zones with the ammonite zone stages and substages (, 1983) and with the conodont ( & , 1980; , 1974a, 1974b, 1980) and foraminiferal zones ( et al., 1983). (1982, 1986) recognised the following Dasycladalean algal zones in the West Carpathian Triassic:
1. The Physophorella pauciforata - Oligoporella pilosa assemblage zone characterised by the following Dasycladalean algae assemblage;
a) The species of Physoporella with single whorls: Physoporella pauciforata with varieties.
b) The species of Physoporella with double whorls: Physoporella dissita, Physoporella praealpina, Physoporella minutula, and Physoporella varicans.
c) The species of Oligoporella with single whorls: Oligoporella prisca.
d) The species of Oligoporella with double whorls: Oligoporella pilosa with varieties.
e) The species Diplopora proba, Diplopora subtilis, Diplopora hexaster, var. hexaster and var. helvetica, Macroporella alpina, Teutloporella tabulata, Teutloporella peniculiformis, and Diplopora annulatissima.
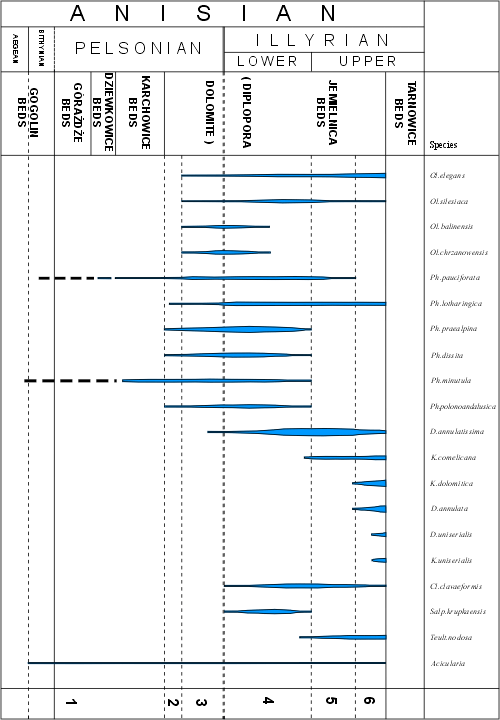
After correlation with ammonite, conodont, and foraminiferal zones, the age of this algal assemblage zones may be established as uppermost Bithynian (?) - Pelsonian - Lower Illyrian (Fig. 5 ![]() )
)
2. The Diplopora annulatissima partial range zone (=Illyrian).
It is represented by a single-species assemblage of Diplopora annulatissima. Its lower border is indicated by disappearance of all species of the Physoporella pauciforata - Oligoporella pilosa Zone. The upper border is indicated by appearance of Diplopora annulata. It corresponds to the ammonite Aplococeras avisianus Zone of the (Upper Illyrian).
3. The Diplopora annulata taxon range zone
It is defined by the range of occurrence of Diplopora annulata zones sensu lato (including Kantia dolomitica). In the lowermost part, it occurs together with Diplopora annulatissima, Diplopora clavaeformis, Diplopora comelicana, and Teutloporella peniculiformis. Diplopora annulatissima does not reach the top of the Diplopora annulata zone. The upper boundary of the zone is indicated by the disappearance of the nominal species and appearance of Teutloporella herculea. It was considered by as characteristic for the Fassanian and Lower Longobardian. The same conception was put forward by E. (1979).
J. (1986) discusses also higher algal zones (Teutloporella herculea interval zone, Andrusoporella duplicata taxon range zone, and Chinianella taxon range zone). These zones are not substantial for our considerations, as they are not present in the Upper Silesian Triassic.
Fig. 5 ![]() presents the stratigraphic ranges and zonation of Dasycladales in the Triassic of the Upper Silesia and adjacent regions. The general conclusion is the same as it follows from
(1994b: tab. 4), with some changes resulting of the more up-to-date species determination. The stratigraphical range of Dasycladalean algae is there limited to the Anisian. Abundant occurrence is characteristic for the Upper Pelsonian and Illyrian (Diplopora Dolomite).
presents the stratigraphic ranges and zonation of Dasycladales in the Triassic of the Upper Silesia and adjacent regions. The general conclusion is the same as it follows from
(1994b: tab. 4), with some changes resulting of the more up-to-date species determination. The stratigraphical range of Dasycladalean algae is there limited to the Anisian. Abundant occurrence is characteristic for the Upper Pelsonian and Illyrian (Diplopora Dolomite).
 |
Click on thumbnail to enlarge the image.
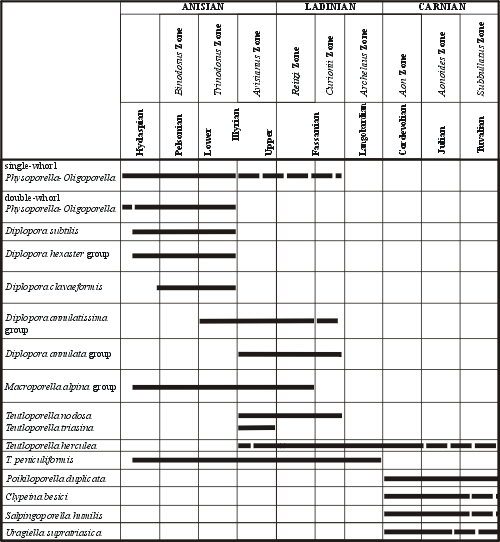
Figure 3: Stratigraphic distribution of the most important Dasycladales in the Alpine Middle Triassic (after , 1972a, tab. 1).
 |
Click on thumbnail to enlarge the image.
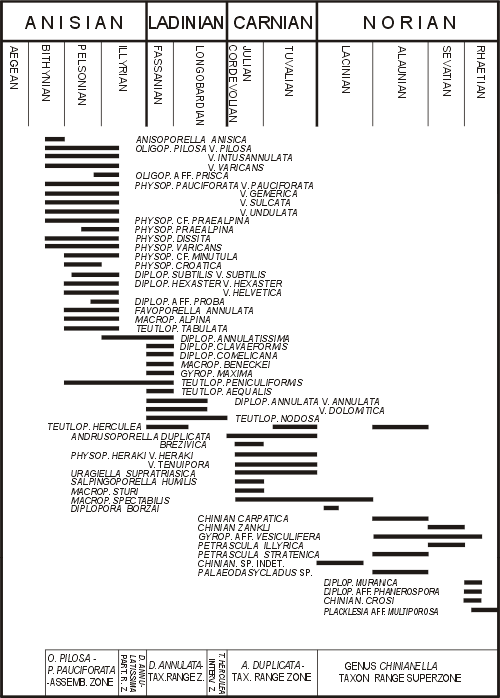
Figure 4: Stratigraphic distribution of the most important Dasycladales in the Triassic of the Western Carpathians (after , 1986, 315).
Occurrence of several algal assemblages may be observed, similarly as in the Alps (Fig. 3 ![]() ) and Western Carpathians (Fig. 4
) and Western Carpathians (Fig. 4 ![]() ):
):
1. The Physoporella-Oligoporella assemblage zone characterised by following Dasycladalean assemblages:
a) Oligoporella prisca group characterised by single whorls, with Oligoporella elegans (uppermost Pelsonian-Upper Illyrian).
b) Oligoporella pilosa group characterised by double whorls, with Oligoporella silesiaca and Oligoporella balinensis including also Oligoporella chrzanowensis (uppermost Pelsonian-lowermost Illyrian).
c) Physoporella pauciforata group, characterised by single whorls, with Physoporella pauciforata (embracing the whole Pelsonian and Illyrian, most abundantly occurring in the uppermost Pelsonian and Lower Illyrian) and with Physoporella lotharingica (Upper Pelsonian and whole Illyrian, up to the bottom of Tarnowice Formation).
d) Physoporella praealpina group, characterised by double whorls, with Physoporella praealpina (Upper Pelsonian-Lower Illyrian), with Physoporella dissita (Upper Pelsonian-Lower Illyrian), Physoporella minutula (Upper Pelsonian-Lower Illyrian), and Physoporella polonoandalusica (Upper Pelsonian-Lower Illyrian).
2. Diplopora annulatissima taxon range zone.
It is characterised by the occurrence of the species Diplopora annulatissima. It is the index fossil for the Illyrian. Frequent in the Lower Illyrian, occurring together with Physoporella praealpina and Physoporella pauciforata groups, and abundant in the Upper Illyrian, reaching up to the bottom of Tarnowice Formation.
3. Diplopora annulata - Kantia dolomitica taxon range zone.
In the uppermost Illyrian these two taxa appear and disappear abruptly at the bottom of Tarnowice Beds. Accompanying species is Kantia comelicana, appearing in the Upper Illyrian. Near the bottom of Tarnowice Formation one can also observe the first appearance of Diplopora uniserialis and Kantia uniserialis, species characteristic for the Lower Ladinian of Western Alps and Lower Sub-Tatric series.
J. (1920), even though attributing great importance to "Diploporae" in refining the Triassic stratigraphy, noted the limitation of applying this method. He warned against simple extrapolation of algal zones from one area to another. Local subdivisions should be worked out independently for each region, using determinations of actual fossil finds. The zonation may be identical as in the adjacent areas or partly differ from that in the neighbouring regions. This principle was quoted by F. (1958, p. 177) and successfully applied in the Western Alps. He discerned three algal zones in the Vanoise Alps that proved important for the whole Briançon Triassic in the French and Italian Alps, as well as in the Swiss Prealps. The same principle of local stratigraphy has been applied for the Triassic of the Upper Silesia and adjacent regions.
Taking into consideration the described above three assemblage and taxon zones, and also some overlap of particular species, the following Dasycladalean local horizons can be recognised in the Triassic of the Upper Silesia and adjacent regions:
I. First horizon. In the Lower Muschelkalk one can find rare Dasycladalean algae: Physoporella minutula, Physoporella pauciforata, and Acicularia sp. (rare appearance in the Upper Gogolin Formation, Górażdże Formation and Karchowice Formation).
II. Second horizon. At the bottom of the Diplopora Beds (Jemielnica Formation) in some places one can observe rare Physoporellae characterised by single whorls (Physoporella pauciforata group) and by double whorls (Physoporella praealpina group). Very rare is Oligoporella elegans.
III. Third horizon. This horizon is characterised by abundant frequency of Physoporellae and Oligoporellae with both single and double whorls. It is the common assemblage in the lower part of Diplopora Dolomite, testifying of its Upper Pelsonian age. In the vicinity of Chrzanów, besides the aforementioned species there occur also numerous Oligoporella silesiaca (), O. chrzanowensis n.sp., O. balinensis (), Salpingoporella krupkaensis n.sp. and Kantia comelicana (). Acicularia sp. occurs sporadically.
IV. Fourth horizon. The middle part of the Diplopora Dolomite is the richest horizon with very diversified and abundant Dasycladalean algae. The most frequent are single- and double-whorl Physoporellae. Widely distributed are Oligoporellae typical for the Chrzanów region, such as Oligoporella silesiaca and O. chrzanowensis. The Illyrian index species Diplopora annulatissima is very abundant. Clavapora clavaeformis, Kantia comelicana and Salpingoporella krupkaensis are of lesser importance. The flora of the IV Dasyclad horizon is characteristic for the Lower Illyrian.
V. Fifth horizon. The upper part of Diplopora Dolomite is characterised by the predominance of Diplopora annulatissima, indicating its Illyrian age. Species of Physoporella praealpina group are of subordinate importance. Still fairly frequent are Physoporellae of the Physoporella pauciforata group. Oligoporella elegans and Oligoporella silesiaca also range up to this horizon. Kantia comelicana and Salpingoporella krupkaensis are quite numerous, while Teutloporella nodosa appears occasionally.
VI. Sixth horizon. In the uppermost part of the Diplopora Dolomite there appear Diplopora annulata and Kantia dolomitica, accompanied by Kantia comelicana. Below the bottom of the Tarnowice Formation there occur also Diplopora uniserialis and Kantia uniserialis.
 |
Click on thumbnail to enlarge the image.
Figure 5: Stratigraphic ranges and frequency of Anisian Dasycladales in the Upper Silesia and adjacent regions; 1-6 - local Dasyclad horizons.
Diplopora annulata sensu lato was widely considered as an index form for the Ladinian (more precisely for the Fassanian and Lower Longobardian). This created a problem for the algal biostratigraphy of the Muschelkalk, because in Lorraine and Schwarzwald the upper part of the Muschelkalk belonged to the Anisian, with Physoporella lotharingica, while in the Upper Silesia, the upper part of the Diplopora Dolomite seemed to be of Ladinian age due to the presence of Diplopora annulata (,
1930a, 1931a). Therefore,
(1974a) doubted the existence of this species in the Upper Silesia. He stated that directly in the bottom of the lower Tarnowice Formation Diplopora annulatissima occurs, an index species for Illyrian, while Diplopora annulata is absent. However, the determination of this species by
(1920) is based on well preserved specimens and should not be questioned. Diplopora annulata has been found also in the new material from boreholes near Zawiercie (,
1981, 1986, Pl. CV, 3-7), together with Diplopora annulatissima (,
1986, Pl. CV, 10-13); its occurrence was also confirmed in the Opole Triassic and in Tarnowice and Bytom synclines (Pl. XXV ![]() ).
).
Some authors (, 1972a; , 1977; et al., 1980; , 1986) were of the opinion that Diplopora annulata appeared already in the uppermost Illyrian. This is also the case of the Upper Silesian Triassic. K. (1975) found the Upper Illyrian conodonts already in the uppermost Tarnowice Formation and in the lower members of the Wilkowice Formation, where actually lies the boundary between Anisian (Illyrian) and Ladinian (Fassanian). Thus, the results of conodont stratigraphy support the conclusion that Diplopora annulata, Kantia dolomitica, Kantia comelicana, as well as Diplopora uniserialis and Kantia uniserialis first appeared already in the uppermost Illyrian.
It can be concluded that the whole Middle Muschelkalk in the Upper Silesia, i.e., the Diplopora Dolomite (Jemielnica and Tarnowice Formation) belong to the Anisian. Similar conclusion was reached by E. and H. (1993) about the German Muschelkalk.
The classical lithostratigraphic subdivision of the Muschelkalk into beds or formations was done for the Opole Triassic (, 1944). The Diplopora Dolomite (Jemielnica Beds) lies here on limestone Karchowice Beds, and is overlain by marly Tarnowice Beds. In other regions only some units can be recognized, while in the Chrzanów area (, 1952) and in the Olkusz-Siewierz region the Terebratula Beds, Karchowice Beds and the Diplopora Dolomite are united into the Olkusz Formation (, 1961, 1964).
| Traditional lithostratigraphic units (beds) | Formal lithostratigraphic units (formations) | ||
| Upper Muschelkalk | Boruszowice Beds | Boruszowice Formation | |
| Wilkowice Beds | Wilkowice Formation | ||
| Tarnowice Beds | Tarnowice Formation | ||
| Middle Muschelkalk | Jemielnica Beds (Diplopora Dolomite) |
Jemielnica Formation | Olkusz Formation |
| Karchowice Beds | Karchowice Formation | ||
| Lower Muschelkalk | Terebratula Beds | Dziewkowice Formation | |
| Górażdże Beds | Górażdże Formation | ||
| Gogolin Beds | Gogolin Formation | ||
The formalisation of the Silesian Muschelkalk subdivision was possible due to efforts of recent authors (, 1973, 1979, 1998, 2000; , 1989, 1990, 1997; , 2000).
Present studies allow recognizing five algal horizons (II-VI) within the
Diplopora Dolomite. The Dasyclads of the lowest horizon are found in the Karchowice and Górażdże Beds (Fig. 3 ![]() ).
).
The thickness of the Diplopora Dolomite ranges from 20 to 50 m, depending on several factors:
1. Uneven subsidence due to different Triassic substrate and tectonic movements during the Triassic (, 1991, 1999), shown in thickness maps (, 1990).
2. In the eastern part of the study area (Stare Gliny - Brudzowice - Zawiercie zone), the Lower Muschelkalk transgresses over Devonian substrate and not all algal horizons are developed there (Fig. 4 ![]() ).
).
3. Incomplete syngenetic dolomitization. In the western part of the Opole Triassic the lower Diplopora strata belong to the Karchowice limestones. Also in other areas there occur limestone beds within the Diplopora Dolomite.
4. Epigenetic dolomitization (, 1944; et al.,
1975; , 1985) that affected a large part of the Diplopora Dolomite. In the Bytom and Tarnowice synclines, east of Mikulczyce - Wieszowa - Tarnowskie Góry tectonic discontinuity, the bottom of the syngenetic Diplopora Dolomite consists of epigenetic Ore-bearing Dolomite, within which vanished the lower Diplopora horizons as well as some Lower Muschelkalk members (Figs. 4 ![]() - 5
- 5 ![]() ).
).
5. The top of the Diplopora Dolomite has often eroded away. There are several epigenetic erosional surfaces: Upper Keuper, Rhaetian and Cainozoic (Fig. 5 ![]() ). Most often, the erosion removed the upper Diplopora horizons. Thus the uppermost strata of the Diplopora Dolomite are usually preserved under marls of the Tarnowice Beds, where there is an uninterrupted transition of the Jemielnica Beds into the overlying Tarnowice Beds.
). Most often, the erosion removed the upper Diplopora horizons. Thus the uppermost strata of the Diplopora Dolomite are usually preserved under marls of the Tarnowice Beds, where there is an uninterrupted transition of the Jemielnica Beds into the overlying Tarnowice Beds.
The Recent Dasycladales have particular preferences regarding salinity, bathymetry, light, temperature and substrate ( & , 1992). They mainly settle in the shallow marine waters of subtropical regions where they are protected from strong storms. They form "meadows" on the rocks or coral reefs in the tropical seas ( & , 1963, p. 517). Dasycladalean meadows may also grow on deeper, better protected marine bottom covered with micritic calcareous mud ( & , 1979).
Nearly all extant Dasycladalean species need euhaline seawater and therefore they prefer the infralittoral zone, like open lagoons, protected but open bays, and the shelves of submarine ridges. Dasycladales are rarely found in closed lagoons with extremely high salinity due to water evaporating or with extremely low salinity due to freshwater supply, leading to brackish conditions. Bathophora oerstedii, an alga typical for mangroves is even found in freshwater lakes in Florida. Bathophora is therefore the exceptional euryhaline plant, which can live in the sea of normal salinity, in the brackish waters and in freshwater (, 1979).
Bioclasts, lithoclasts and hard ground as well as stabilised, partly sandy or muddy soft ground serve as substrates for Dasycladales also in other habitats, e.g., on the Bahamas carbonate platform.
The temperature tolerance of the Recent Dasycladales is also low. Nearly all extant genera are restricted to tropical and subtropical regions of the oceans. Only a few species have advanced into marginal subtropical waters. Their northernmost distribution is the Mediterranean Sea, where large populations of Acetabularia acetabulum may occasionally be found in the shallow waters within 0-10 m depth, depending also on the water clarity ( & , 1992). According to J. (1967), Dasycladus clavaeformis () lives in separated bays on more on less consolidated substrate or hard ground. They are not found in stormy seas. In calm waters, the aggregations of algae are partly covered with silt already in vivo. They can cover large patches at 3 to 6 m depth. Acetabularia mediterranea grows in bays on hard ground at 1.5-3 m depth, forming belts of algal aggregations stretching parallel to the shoreline. Patchy occurrences are noted throughout the sublittoral zone. Best developed algal covers are found in clear and well lit sublittoral waters on southern and southwestern slopes. The caps are shed in summer and are accumulated by currents at 25-30 m depth. Between islands, where strong current occur, there can be no sediment accumulation at all (, 1988). According to (1988), the surface water temperature in the Adriatic Sea reaches 23-24 °C, while at 100-200 m it is 10-13 °C. The salinity is 34-35 ‰, more than the average for oceans. The aeration is 100 % in spring, and 70-80 % in summer and autumn.
According to (1979), Recent Dasycladales may attach to rocks and stones (epilithic mode of life), to mangrove roots (epiphytic), may grow in the mud (pelophytic), in the sands (psammophytic) or on broken shells and fragmented corals (zoophytic).
Extrapolating the ecological data on Recent Dasycladales to their relatives from earlier geologic periods in order to answer palaeoecological questions requires an assumption that the environmental preferences of these algae have not changed, or, at least, changed only slightly during their evolution. Investigations concerning this problem have shown that the extinct Dasycladales did not settle in essentially different habitats than the Recent algae of this group.
The major factor limiting their development is the water temperature. Both Recent and fossil species have tropical distribution. Several modern and fossil species managed to adapt to fairly varied salinity conditions. There are forms living in normally saline seawater, as well as in lagoonal environments. To establish palaeoenvironmental conditions for particular facies, whole assemblages of algae and marine fauna have to be considered, as some taxa may be more reliable indicators of salinity conditions and other environmental parameters (, 1979).
& (1969) observed that the assemblage found in the Middle Triassic of the Romanian Carpathians represents a reef biocoenosis, containing Dasycladales, Codiaceae and Solenoporaceae, accompanied by detrital remains of corals, gastropods, crinoid trochites and echinoid plates.
According to E. (1972b, p. 254), the formation of reefs began in the Late Anisian; the incipient reefs on ridges exist already in the Lower Muschelkalk. The growing reefs surrounding basins increased the area of lagoonal sedimentation. The lagoonal deposits show a typical succession of algal stromatolites and Dasyclad banks. The Dasyclads are variably distributed within a lagoon. For example, Teutloporella nodosa and T. herculea occupy a zone along the reef margin, while Diplopora annulata occurs only in the central part of the lagoon.
E. (1979), describing a Dasyclad assemblage from the Anisian of Italian Alps, noted that in some cases, the thalli occur in very fine-grained micritic clasts, probably representing the original sediment where the algae grew. It was a low-energy environment, such as a lagoon with muddy-sandy bottom. The water circulation was limited and thus the plant remains could have fossilized. More often, however, the Dasyclad thalli are crushed, densely packed, preferentially oriented, partly deteriorated and concentrated in layers enriched in remains of these algae, alternating with layers without the Dasyclads (e.g., interbedded with stromatolite strata). This indicates that the algal remains were transported by water currents in high-energy conditions, e.g., within tidal channels.
Some algae co-occur in high-energy environments with organisms ecologically so varied as bryozoans, brachiopods and charophytes. However, they are redeposited into such context. They are allodapic algae (, 1979). Most Dasyclads of the Pyrenean Jurassic lived in calm infralittoral habitats protected from turbulence and with normal salinity; some could have been able to survive episodic increases in salinity. There were also eurytopic algae, inhabiting marine waters of elevated salinity. This is the case of Acicularia, present both in the Diplopora Dolomite in the Upper Silesia, as well as much farther north in the hyperhaline Muschelkalk sea of the Holy Cross Mountains ( & , 1974). A similar case was described from the Triassic of Iran (, 1979).
One of the most fascinating and difficult to explain phenomena is the presence of typically Alpine Dasyclads, known from inner sedimentary zones, on the Upper Silesian carbonate platform, located at the distant periphery of the Alpine-Carpathian orogen. Such Alpine species are: Oligoporella pilosa, O. prisca, Physoporella pauciforata, P. praealpina, P. dissita, Diplopora annulatissima, D. annulata, D. uniserialis, Kantia dolomitica, K. uniserialis, K. comelicana and Clavapora clavaeformis. The presence of these species allowed correlating the stratigraphy of the Diplopora Dolomite with the more universal Alpine stratigraphic scheme. However, besides these far-ranging species there are also taxa that can be currently regarded as endemic for the Upper Silesia: Oligoporella elegans, O. silesiaca, O. balinensis, O. chrzanowensis and Salpingoporella krupkaensis. There are also forms typical for the Upper Silesia but with wider palaeogeographic connections: Physoporella lotharingica, P. minutula and P. polonoandalusica. Presence of so numerous species, both more cosmopolitan ones and more endemic, allowed to discern local Dasyclad horizons.
Occurrence of so abundant and diverse Dasyclad flora indicates favourable palaeoecological conditions on the Upper Silesian carbonate platform. Also the development of a diverse fauna (gastropods, bivalves, crinoids, echinoids, corals) suggests normal salinity in the Middle Muschelkalk sea. F. (1958) regarded the Triassic Vanoise fauna, lacking cephalopods, as a depleted assemblage, closer to those of Germany and Upper Silesia than to the Triassic fauna of the Eastern Alps. This was due to the fact that the salinity of the Briançon Triassic sea was high enough to prevent development of the cephalopod fauna. A Briançon - Pre-Alpine - High-Tatric - Upper Silesian palaeogeographic province (, 1967, 1994a, 1994b) was then formed, transitional between thr Alpine and the Germanic provinces. But what separated the latter two provinces?
Some researchers believe that there was a Vindelitic-Beskidy Wall, including the Bohemian Massif (, 1938, 1959; , 1982; , 1985, 1991; , 1991, 1999). The Germanic Sea could communicate with the Alpine Sea only through gates, such as the Eastern Carpathian Gate ( & , 1975; , 1974a), Silesian-Moravian Gate ( in & , 1953; , 1985, 1991; , 1991, 1999), and Burgundian Gate (, 1974b; , 1991). Some scientist regarded the Vindelitic-Beskidy Wall as a chain of islands (, 1991, 1999). If so, the islands should have supplemented the neighbouring basins with detritic material, as was the case, e.g., with the margins of the Massif Central (Mont d'Or, Crussol) and the outer massifs of the Alps, where transgressive detritic sediments had accumulated during formation of the Muschelkalk. Such Hercynian massifs as the Vosges, Schwarzwald, Harz, and the Bohemian Massif were not emerged at that time, however, and were not recorded in the Muschelkalk palaeogeography (, 1963; , 1964; , 1972; et al., 1990, 1993; , 1998).
Also the Beskidy Wall did not exist in the Middle Triassic ( & , 1964), even though during the Early and Late Triassic it supplied detritic material to the Tatra Werfen ( & , 1964; & , 1960). The Germanic Sea communicated with the Alpine Sea through a passage wider than the Moravian Gate (, 1928; & , 1964; , 1972). This way the Upper Silesian Anisian assemblage was enriched, e.g., by Kantia comelicana, known only from the Italian Alps and the Gemer Series in southern Slovakia (, 1979; , 1986).
(1963) believed that during the Middle Muschelkalk times there was a submarine step between the Briançon area and the outer areas, which separated the hypersaline waters from the open sea. However, in the Tatra Mountains, the Middle Triassic strata, both of the High-Tatric and Križna Series, contain numerous dolomitic intercalations, formed in highly saline sea ("pea dolomites" of , 1963). Only occasionally there were deposited dolomites with rich Dasycladacean flora - Illyrian in the High-Tatric Series and Fassanian in the Križna Series (, 1967, 1986). According to (1963), in the Ladinian no emerged land barrier separated the Germanic Muschelkalk sea from the marine areas of French Alps inhabited by calcareous algae. Only an environmental barrier due to salinity or temperature gradient or currents (turbulences) could separate the two provinces. Accepting this explanation and extrapolating it onto the Polish Triassic realm, it should be noted that such an environmental dispersal barrier was not always tight (Illyrian and Fassanian biotic interchanges) and allowed the Dasycladacean flora of the open Alpine Sea to enter the Upper Silesian carbonate platform.
An important question is identifying the factors preventing the Upper Silesian biota from invading the extensive Germanic Basin, and, on the other hand, what blocked the hypersaline waters of the Germanic Basin from entering the Upper Silesian Sea, given the fact that they sometimes reached the High-Tatric and Križna areas. envisaged the hypersaline areas separated from the open sea by submarine steps, allowing precipitation of primary dolomites, calcium sulphate, sodium chloride, and sometimes potassium chloride from the salines. Fairly arid climate facilitated evaporation. Large thickness of the Middle Triassic sediments in the Germanic Basin accumulated due to strong subsidence of large areas of Central Europe, varying because of the movements of the Permian salt masses in their substratum (, 1990; , 1998).
It is, however, unclear what could constitute the submarine step separating highly saline Germanic sea from the normally saline Upper Silesian Sea. It could not have been a coral reef, because corals, fairly common in the Diplopora Dolomite, did not form a barrier reef but instead occur as patchy bioherms (, 1974, 1988). According to (1972), such protection from turbulent open-sea waters was often offered by oolitic mounds. Oolitic, oncolitic and stromatolite intercalations are indeed quite common in the Diplopora Dolomite sections, originally forming belts of submarine mounds and banks (, 1992). Also (1999, 2000) considers such explanation plausible. Protected by these barriers, the carbonate platform formed under conditions of euhaline or slightly hyperhaline sea and low water turbulence, where lagoon-like environmental conditions allowed the prolonged development of Dasycladacean assemblages during the Pelsonian and Illyrian.
The Upper Silesian carbonate platform probably extended much farther eastwards, but is now hidden under Flysch Carpathian nappes (Fig. 6 ![]() ). Middle Muschelkalk are usually missing there due to erosion. They are quite frequently documented by flysch exotics. K.
(1914); quoted also in
et al. (1988), found a "Nullipora dolomite" among exotics in Kruhel near Przemyśl. In the Pieniny Klippen Belt, even farther south, the Middle Triassic resembles that of the High-Tatric Series (,
1963; & , 1964) without any marked influence of the hypothetical Beskidy Wall.
). Middle Muschelkalk are usually missing there due to erosion. They are quite frequently documented by flysch exotics. K.
(1914); quoted also in
et al. (1988), found a "Nullipora dolomite" among exotics in Kruhel near Przemyśl. In the Pieniny Klippen Belt, even farther south, the Middle Triassic resembles that of the High-Tatric Series (,
1963; & , 1964) without any marked influence of the hypothetical Beskidy Wall.
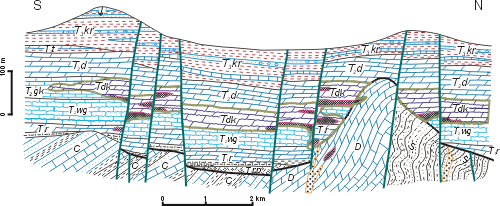 |
Click on thumbnail to enlarge the image.
Figure 6: Schematic geological section through a fragment of Triassic deposits in the north-eastern part of the Silesia-Cracow area (after , 1976). S - Silurian; D - Devonian; C - Carboniferous; T1pp - Lower Buntsandstein; T1r - Upper Buntsandstein (Röt); T2wg - Gogolin Beds; T2gk - Górażdże, Terebratula and the Karchowice Beds; Tdk - Ore-bearing Dolomite; T2d - Diplopora Dolomite; T2t - Tarnowice Beds; T3kr - Keuper-Rhaetian; J - Jurassic.
The lithofacies, environmental restorations and sedimentation regime of the Diplopora Dolomite of the eastern part of the Silesia-Cracow area (Bytom Syncline, Cracow-Chrzanów region, and Olkusz-Siewierz Monocline) were described by J. (1992). She compiled lithological profiles showing the sequence of ten lithofacies recognized using macro- and microscopic observation (microfacial analysis). Three rock complexes were discerned within the Diplopora Dolomite (Lower, Middle and Upper), differing in their sedimentary structure and conditions of origin. Chrzanów, Olkusz, Siewierz and Bytom areas were characterized. For each region, the environmental energy was estimated typical for the rock complexes.
According to J. , whose conclusions were based on a very keen microfacial analysis, the Diplopora Dolomite formed in sublittoral environment, at depths typical for continental shelf. Locally, deposition was occurring in littoral shallows, associated with laminites and stromatolites. These sediments contain various early diagenetic structures (birdseyes, or ocellar pores) indicating periodic emergence of the sea bottom and formation of islands. Also the presence of microstalactite cement within birdseyes indicates littoral conditions and is regarded as indicative of subaerial exposure of the sediments and their subjection to factors of marine vadose diagenesis (, 1993).
After the freshwater supply (from rainfall) ceased, the saline environment regained dominance. The above processes occurred in the carbonate sediments prior to dolomitization; the latter process was probably due to alternating influx of freshwater (meteoric) and saltwater. Magnesium-rich salines caused early synsedimentary dolomitization of the carbonate sediments together with their fossilized fauna and algal flora. Such dolomitization did not substantially deform the Dasyclads. Their calcareous skeletons were usually dissolved but the moulds preserved the shape of branches and segmentation of thalli well enough to enable taxonomic identification of the Dasycladalean remains (, 1981).
Similar conclusions about the origin of the Diplopora Dolomite were reached by J. (1994), who regarded the sedimentary conditions as subtidal to intertidal. The syngenetic dolomitization caused by mixing meteoric-saline waters also speaks in favour of the interpretation of sedimentary environment as a shallow marine one.
All researchers working on Dasycladales noted that the algae are excellent markers of facies and environmental conditions. Many detailed observations were made by H. (1972b, 1976, 1977), who worked on well-preserved Anisian and Ladinian Dasyclads from the South Alps.
(1976, 1977) noted a correlation between the annulation of the algae and the fine-grained fraction of the sediment. The finer is the matrix, the more annulated are the calcareous skeletons. In Diplopora annulata and other segmented species, the spacing between the annuli depends on water movements. In especially calm, silty environments the annulation is denser than in more turbulent habitats. The species forms dense "meadows" in suitable habitats. Also in other species in calm and mud-rich environments, the furrows are closer spaced, while the distances between them are greater in more turbulent biotopes. This correlation provides an explanation as to why small specimens (and species) of Dasyclads show denser segmentation. Small plants, growing under partial protection of larger ones were less exposed to water movements and could have developed finely annulated skeleton, more prone to breaking, but allowing greater mobility.
The spacing of interannular furrows depends, however, not only on environmental conditions. The spacing between annuli varies. If the distance is large enough, a furrow appears. With denser spacing, it is possible to estimate the turbulence of the environment looking at the width of the furrow. In Ladinian Diplopora exuberans, very similar to Diplopora annulata, (1977) observed that in deeper waters almost exclusively single-whorl segments occurred, while double-whorl segments predominate in shallower facies, where even triple-whorl specimens can be found. Fine-grained matrix is typical for samples yielding single-whorl segments. Samples with double or triple whorls lack fine-grained sediment, suggesting stronger water agitation. Segmentation of the skeleton is thus clearly correlated with environmental conditions.
T. and R. (1970) studied Anisian (Pelsonian) strata in the Dolomites of Southern Tyrol, noted the following facial correlations: In the western part of the study area there occur sandstones and conglomerates formed near shore. They gradually pass into marly limestones with numerous Dasyclad fossils. The algae are even more abundant in shallow marine limestones deposited on carbonate platform far from the shore. The Dasyclads are missing from the center of the carbonate platform, where marls were deposited in a fairly deep basin. Thus, the near-shore and basin sediments are devoid of Dasyclads. Too close to the shore, the sediment supply and water turbulence were too intense, while in the middle of the basin it was too deep for the algae to thrive.
& (1970) studied the Dasyclad species involved, and their assemblages. The authors identified 14 species and recognized four ecological groups.
Closest to the land Group A (Diplopora hexaster) predominate, consisting of species less sensitive to sediment influx and turbulence. These are Dasyclads with cup-shaped branch tips, tightly arranged. They were adapted to poorly lit, turbid water. The cup-like instead of hair-like shape of the laterals was an adaptation reducing the area of turbulent water action and making it more difficult for sediment particles to stick to the alga.
Group B (double-whorl Physoporella-Oligoporella assemblage) developed on muddy bottom farther from shore. The water was calmer, with only fine silt reaching this zone from the land. The Physoporella and Oligoporella species belonging to this assemblage had in fact apically pointed branches (, 1969).
Group C is heterogeneous as far as branch shape is concerned; it includes Macroporella with funnel-shaped branches (widening distally) and Teutloporella with hair-like branches, giving it a "bottle-cleaner" appearance. These algae inhabited well lit and turbulent shallows, without much suspended sediment in the water.
Group D (single-whorl Physoporella-Oligoporella assemblage) developed in the interior part of the carbonate platform, protected by shallow banks. The optimum photic conditions stimulated growth of species with widely spaced whorls.
The above examples not only demonstrate the correlation of the distribution of particular Dasycladalean species with their sedimentary environment, but also reveal the morphological adaptations to various environmental conditions (, 1976).
The aforementioned observations and inferred correlations allow an attempt to estimate the living conditions of the ecological assemblages of Dasycladales from the Upper Silesian carbonate platform during the Pelsonian and Illyrian (Fig. 7 ![]() ).
).
1. Single-whorl Physoporella-Oligoporella assemblage. This assemblage includes Oligoporella prisca, O. elegans, Physoporella pauciforata and P. lotharingica. The algae developed in the interior part of the platform, protected by shallow banks (localities Laryszka, Segiet, Przełajka). Thanks to optimum lighting conditions, even P. lotharingica could thrive there, despite its widely spaced whorls. Minute O. elegans could grow there protected by larger P. pauciforata, with which it often co-occurs. This is an assemblage typical for the Upper Pelsonian.
2. Double-whorl Physoporella-Oligoporella assemblage. The assemblage includes Oligoporella pilosa, O. silesiaca, O. balinensis, O. chrzanowensis (Rosowa Góra, Balin), Physoporella praealpina, P. dissita, P. minutula and P. polonoandalusica (Boleradź). The algae developed on muddy bottom away from the shore. The water movement was limited, with fine silt reaching this zone from the land, forming the micritic rock matrix. P. praealpina was the predominant species, locally forming "diplopore meadows" (Przełajka). Good lighting conditions encouraged growth of P. polonoandalusica, with large distances between its double whorls. Small Oligoporellae O. silesiaca, O. balinensis and O. chrzanowensis formed an endemic assemblage, typical for the Chrzanów area, but extending also onto adjacent regions. Also this assemblage is typical for the Upper Pelsonian.
3. Salpingoporella krupkaensis defines a separate assemblage, typical for Luszowskie Góry (locality Krupka) and common also in the Chrzanów area (Rosowa Góra and Mały Balin) and in Olkusz area (Stare Gliny). This species thrived on muddy bottom. The water was so calm, that even the most external, sticky layer (the cortex) was preserved around the dead algae resting on the seafloor. This assemblage is typical for the beds around the Pelsonian-Illyrian boundary.
4. Diplopora annulatissima also forms a distinct assemblage. The species had regularly arranged double whorls, indicating steady low turbulence. On shallow banks, "diplopore meadows" developed extensively, consisting mostly of this single species (Jemielnica, Segiet), with minor admixture of a related species Clavapora clavaeformis (Przełajka). Under their cover grew small Oligoporellae O. silesiaca, O. balinensis and O. chrzanowensis. Nearby there were meadows with Physoporella praealpina and P. pauciforata, as well as muddy basins with Salpingoporella krupkaensis. On shallower banks with more turbulent water, there were unsegmented algae Kantia comelicana (Mały Balin, Klucze). This assemblage is typical for the Lower Illyrian.
5. Diplopora annulata assemblage with related taxa Kantia dolomitica, Diplopora uniserialis and Kantia uniserialis. All the species are segmented. Diplopora annulata and Kantia dolomitica form annuli composed usually of two whorls, indicating low turbulence. Concentrations of Diplopora annulatissima formed under similar conditions. Under the cover provided by those large diplopores, there grew small, densely segmented Diplopora uniserialis and Kantia uniserialis (Tarnów Opolski, Kamień Śląski, Segiet, Libiąż, Wojkowice Komorne, some boreholes near Zawiercie). In shallower, more turbulent habitats, there were patches of unsegmented Kantia comelicana. This assemblage is typical for the Upper Illyrian.
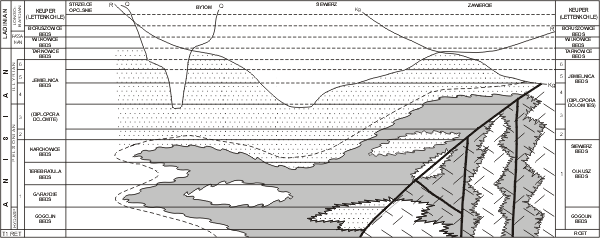 |
Click on thumbnail to enlarge the image.
Figure 7: Schematic section through the Middle Triassic strata of the Upper Silesian and adjacent regions (after Z. , 1994a, fig. 4. 8). 1-6 - local Diplopora horizons; dotted areas represent early diagenetic dolomites (Diplopora, Siewierz dolomites, as well as dolomites in the Lower Muschelkalk); shaded surfaces - epigenetic Ore-bearing Dolomite; differently oriented dashes - Devonian basement of transgressive Triassic deposits; surfaces of epigenetic erosion cutting Middle Triassic deposits: Kg - Upper Keuper; R - Rhaetian; Q - Quaternary.
The Dasycladalean assemblages described above are often accompanied by a fauna of minute gastropods, feeding on the algae (, 1958; , 1986). Much rarer are small bivalves.
The palaeoecological cross-sections (Fig. 7 ![]() ) show a theoretical distribution of Dasycladalean assemblages living on the Upper Silesian carbonate platform during the Pelsonian and Illyrian. It is, however, impossible to precisely locate the described assemblages for several reasons. First, the diplopores are rarely found in situ. This happens when we recover very long (up to 10 cm) individuals, consisting of many segments (10 to 15 annuli), rarely broken, belonging to a single species or to few species living together in ecological interdependence. They sank to the seafloor for biological reasons (diplopores die after releasing gametes; & ,
1971), or due to the destruction of algal meadows by storm events and covered with coarser sediment. Also pelophytic assemblages with preserved delicate details, like the cortex, may be regarded as occurring in situ.
) show a theoretical distribution of Dasycladalean assemblages living on the Upper Silesian carbonate platform during the Pelsonian and Illyrian. It is, however, impossible to precisely locate the described assemblages for several reasons. First, the diplopores are rarely found in situ. This happens when we recover very long (up to 10 cm) individuals, consisting of many segments (10 to 15 annuli), rarely broken, belonging to a single species or to few species living together in ecological interdependence. They sank to the seafloor for biological reasons (diplopores die after releasing gametes; & ,
1971), or due to the destruction of algal meadows by storm events and covered with coarser sediment. Also pelophytic assemblages with preserved delicate details, like the cortex, may be regarded as occurring in situ.
However, most of the diplopores found come from multispecies accumulations, comprising forms with different environmental preferences, forming only few centimeter thick beds, (e.g., Suchodaniec, MuzPIG 1653.II.21, 22). The thalli are often broken, crushed, damaged and oriented along a common direction. Such deposits indicate high-energy environments and probably formed in tidal channels. Several such intercalations may occur in a single geological section, and in each such bed, the fossils may have originated from areas inhabited by different assemblages.
In diplopore-bearing intercalations there commonly occur faunal remains belonging to animals living outside the algal concentrations. The diplopore meadows were frequented only by small gastropods and bivalves feeding on the algae; these molluscs can be regarded as co-occurring with the Dasyclads in situ. But the diplopore-bearing intercalations contain also shells of large snails and bivalves, mostly crushed, as well as corals and echinoids. Only rarely are crinoid columnals found. Obviously, the crinoid and Dasyclad habitats were widely separated due to different environmental requirements. Already the first Silesian diplopore researchers noticed that inverse correlation, and did not look for the algae in rocks with abundant crinoid fossils.
 |
Click on thumbnail to enlarge the image.
Figure 8: Palaeogeographical sketch map showing situation of the Upper Silesian Carbonate Platform in relation to the German evaporitic Middle Muschelkalk and the Carpathian-Alpine Middle Triassic (after , 1982), RM - Rhenish Massif, ECG - East Carpathian Gate, RFH - Ringkobing-Fyn High, USCP - Upper Silesian Carbonate Platform, SMG - Silesian-Moravian Gate, A in Pelsonian (3rd Diplopora horizon), B in Lower Illyrian (4th Diplopora horizon).
 |
Click on thumbnail to enlarge the image.
Figure 9: Palaeoecological sections showing environments where Anisian Dasycladales flourished on the Upper Silesian Carbonate Platform.
Taking all the above into account, we can realize how difficult would be an attempt to reconstruct palaeogeographic-facies maps in a real geographic context.
Such difficulties were encountered by J. (1992; Fig. 9), who tried to map the extent of Diplopora Dolomite sediments representing various facies, using microfacies analysis. Her restoration of palaeoenvironments and mode of sedimentation is correct and generally fits the restoration constructed using the palaeoecological analysis of various diplopore assemblages. However, the lower, middle and upper complexes she discerned are not always comparable, because the age of each complex may vary between profiles. In most cases, the complexes do not include the whole depositional sequence of the Diplopora Dolomite, but only the Pelsonian and its boundary with the lowermost Illyrian. Both the microfacies studies and the palaeoecological observations on the Dasyclad assemblages indicate a large palaeogeographic and facies variability within the Diplopora Dolomite sediments of the Upper Silesian carbonate platform, and demonstrate the difficulty to indicate it on maps.
 |
Click on thumbnail to enlarge the image.
Figure 10: Relative abundance of the identified Dasycladacean species in the Diplopora Dolomite localities of the Upper Silesia, Cracow-Chrzanów Region, Olkusz-Siewierz Monocline and in the deep boreholes of the Zawiercie Region. 1 - very abundant; 2 - abundant; 3 - numerous; 4 - rare; 5 - single.
| ORDER Dasycladales , 1931 | |||||
| FAMILY Seletonellaceae (, 1950) et al., 1975 | |||||
| TRIBE Seletonelleae (, 1950) et al., 1979 | |||||
| SUBTRIBE Teutloporellinae , 1920 | |||||
| GENUS Teutloporella , 1920 | |||||
| Teutloporella nodosa (, 1863) ex , 1920 | |||||
| FAMILY Triploporellaceae (, 1920) | |||||
| TRIBE Salpingoporellae et al., 1979 | |||||
| SUBTRIBE Salpingoporellinae et al., 1979 | |||||
| GENUS Salpingoporella ( in , 1917) et al., 1973 | |||||
| Salpingoporella krupkaensis n.sp. | |||||
| SUBTRIBE Oligoporellinae et al., 1979 | |||||
| GENUS Oligoporella , 1912 | |||||
| Oligoporella prisca group: | |||||
| • Oligoporella prisca , 1912 | |||||
| • Oligoporella elegans ex , 1931a | |||||
| Oligoporella pilosa group: | |||||
| • Oligoporella pilosa , 1912 | |||||
| • Oligoporella silesiaca (, 1872a) | |||||
| • Oligoporella balinensis (, 1892) | |||||
| Oligoporella chrzanowensis group: | |||||
| • Oligoporella chrzanowensis n.sp. | |||||
| GENUS Physoporella (, 1903) , 1912 | |||||
| Physoporella pauciforata group: | |||||
| • Physoporella pauciforata (, 1872a) , 1912 | |||||
| • Physoporella lotharingica (, 1898) | |||||
| Physoporella praealpina group: | |||||
| • Physoporella praealpina , 1920 | |||||
| • Physoporella dissita (, 1872a) | |||||
| • Physoporella minutula (, 1872a) , 1912 | |||||
| • Physoporella polonoandalusica n.sp. | |||||
| FAMILY Diploporaceae (, 1920) , 1988 | |||||
| TRIBE Diploporae (, 1920) , 1979 | |||||
| GENUS Kantia (, 1912) , 1979 | |||||
| Kantia dolomitica (, 1912) , 1979 | |||||
| Kantia uniserialis (, 1912) , 1979 | |||||
| Kantia comelicana (, 1979) | |||||
| GENUS Diplopora (, 1863) , 1979 | |||||
| Diplopora annulata (, 1863) | |||||
| Diplopora uniserialis (, 1920) , 1979 | |||||
| Diplopora annulatissima , 1920 | |||||
| GENUS Clavapora , 1979 | |||||
| Clavapora clavaeformis (, 1920) , 1979 | |||||
| FAMILY Acetabulariaceae () , 1885 [Polyphysaceae (, 1841)] | |||||
| TRIBE Acetabulariae , 1842 | |||||
| GENUS Acicularia d', 1843 | |||||
Order Dasycladales , 1931
Family Seletonellaceae (, 1950) et al., 1975
Tribe Seletonelleae (, 1950) et al., 1979
Subtribe Teutloporellinae , 1920
Teutloporella (, 1912) et al., 1978
Teutloporella nodosa (, 1863) ex , 1920
1863 Diplopora nodosa n.sp. - Pl. LXVe, 19-20.
1920 Teutloporella nodosa n.comb.- , Pl. II, 11-13.
1965 Teutloporella nodosa.- , Pl. II, 1, Pl. III, 1-4; Pl. IV, 1.
1969 Teutloporella nodosa.- & Pl. I, 4.
1969 Teutloporella sp. 2.- & Pl. IV, 9; Pl. V, 3.
1973 Teutloporella aff. nodosa.- & Pl. III, 12.
1979 Diplopora nodosa.- , Pls. I-XVI.
1986 Teutloporella nodosa.- , p. 305.
1994 Teutloporella nodosa.- & , p. 55, 73.
1995 Teutloporella nodosa.- -, Pl. VI, 8.
2000 Teutloporella nodosa.- & , p. 42-44.
Material and locality.- A single specimen in thin section from Laryszka (locality no. 5), Opole Triassic. Z. 's coll. No. 1682.II.5.1.
Descriptions and discussion.- Massive, well calcified thallus, divided into movable, discrete funnel-shaped segments, separated by deep furrows, reaching up to the central cavity. The segments are concrescent near the axial cell and diminishing very fast outside, thus having appearance of little baskets. The arrangement of pores is not regularly euverticillate, and owing to very close connection of verticils and intense calcification, the boundaries between verticils are obliterated. By inference, when observing the external margins of the thallus, which is often shredded, one can distinguish 4-5 verticillated branches. The branches are trichophore, swollen proximally and thinner outside. Dimensions (D= 2.5 mm, H= 0.9-1.25 mm) are comparable with the data of (1920, p. 45; , 1965, p. 9, 10) and of & (1969, p. 71).
Stratigraphical range and geographical distribution.- Teutloporella nodosa is typical for the Ladinian limestones and dolomites of Marmolata and was also found in northern Tirol, and in the Appenines (, 1920). It is frequent in the Wettersteinkalk of the Austrian Northern Alps (Zugspitze, Karwendel, Kaisergebirge - , 1974), where its age is uppermost Anisian and Ladinian. It is also known from Slovenia, Dalmatia, Apuseni Mts (Romania) and Southern Slovakia (, 1986). The age is late Anisian to early Ladinian; the accompanying flora contains Diplopora annulata ( & , 1994).
Teutloporella aff. nodosa is found in the Upper Silesia in locality no. 5, Laryszka, together with Diplopora annulatissima, Physoporella pauciforata, Physoporella lotharingica and Oligoporella elegans. It is of Illyrian age.
Remarks and discussion.- The shape of two preserved segments is almost identical as restored by (1920, fig. 9). The upper segment is larger than the lower one, because of the diminishing number of verticals. The exact shape of branches is not possible to determine, but they are evidently larger proximally. Some parts of verticals are preserved, but branches are not grouping in clusters. This is the reason, why the species nodosa is classified among Teutloporella (see & , 1994) and not Diplopora ( & , 2000). The general shape of segments is also similar to Teutloporella triasina (, 1912, in & , 2013, see Pl. III (IV), 14), but in our material no series of pores are observable. C.W. (1872a) remarked that Gyroporella triasina, known from Recoaro in Southern Alps, was absent from the Upper Silesia.
 |
|
|
Click on thumbnail to enlarge the image.
Figure 11: Teutloporella aff. nodosa (, 1863) ex , 1920. Photograph of a thin section x 12; longitudinal section. Locality no. 5 - Laryszka, Opole Triassic, Upper Silesia. MuzPIG 1682.II, thin section no. 5.1.
Family Triploporellaceae (, 1920)
Tribe Salpingoporelleae et al., 1978
Subtribe Salpingoporellinae et al., 1979
Salpingoporella ( in , 1917) et al., 1973
Salpingoporella krupkaensis n.sp.
Material and localities.- Several dozens of specimens on the fractures from the Kraków-Chrzanów region (localities: Rosowa Góra, no. 48; Balin, no. 50; Krupka, no. 52), Olkusz-Siewierz Monocline (Stare Gliny, no. 57; Łosień, no. 62; Trzebiesławice, no. 65; Warpie, no. 66; Brudzowice, no. 70; Nowa Wioska, no. 71), deep borings of the Zawiercie region (borehole 72 KM Koziegłowy), Bytom Syncline (localites: Przełajka, no. 25; Kamyce, no. 26; Grodziec, no. 33).
Holotype.- Pl. I, 6 ![]() .
.
Paratypes.- Pl. I, 2, 7 ![]() ; Pl. IV, 1, 5
; Pl. IV, 1, 5 ![]() .
.
Repository.- Polish Geological Institute, Warsaw, Geological Institute, coll. of Z. no. 1653.II.
Type locality.- Krupka (locality no. 52) in Luszowskie Góry area, Cracow-Chrzanów region.
Derivation of the name.- After the type locality Krupka.
Type horizon.- Upper part of the Diplopora Dolomite, Upper Anisian.
Diagnosis.- Straight thallus without annulation or undulation, with simple, closely spaced perpendicular phloiophorous branches arranged in clearly developed whorls, thus constituting the euspondyl arrangement of branches.
Description.- Branches (tubercles) weakly attached to the wall of central cavity, rapidly expanding distally, and forming funnel-like tubercles (Pl. I, 7 ![]() ), merging with the adjacent rock, where they begin to touch, forming probably a rarely and poorly preserved cortex (Pl. II, 4
), merging with the adjacent rock, where they begin to touch, forming probably a rarely and poorly preserved cortex (Pl. II, 4 ![]() ), absent from Jurassic and Cretaceous species (,
1958, p. 110). Tubercles are perpendicular to the central cavity and arranged in single rows. Normally, they are observable from inside, because the tubercles are weakly attached to the wall of central cavity and strongly adhering to the adjacent rock. Central cavity is fairly large, with a smooth wall. It is hollow or filled with sediment, forming the central stem. Whorls are straight (Pl. I, 2
), absent from Jurassic and Cretaceous species (,
1958, p. 110). Tubercles are perpendicular to the central cavity and arranged in single rows. Normally, they are observable from inside, because the tubercles are weakly attached to the wall of central cavity and strongly adhering to the adjacent rock. Central cavity is fairly large, with a smooth wall. It is hollow or filled with sediment, forming the central stem. Whorls are straight (Pl. I, 2 ![]() ), or slightly wavy, if the tubercles are a little alternating (Pl. IV, 3, 9
), or slightly wavy, if the tubercles are a little alternating (Pl. IV, 3, 9 ![]() ). Tubercles are rarely hollow (Pl. IV, 12
). Tubercles are rarely hollow (Pl. IV, 12 ![]() ).
).
Comparisons.- Salpingoporella krupkaensis n.sp. belongs to the genus Salpingoporella with euspondyl whorls of phloiophorous branches, thus differing from the genus Macroporella with generally aspondyl branches. R. (1962) proposed a name Pianella for euspondyl Macroporella, but I. (in & , 1971) noted that this is a later synonym of Salpingoporella in , 1917. This view was adopted by E. (1974) and by B. and T. (2000), who rejected the name Pianella and used the name Salpingoporella, following the rule of priority.
Genus Salpingoporella is common in the Upper Jurassic and Lower Cretaceous (, 1974), but was also found in the Carnian (Julian) in Slovakia (, 1967). Salpingoporella krupkaensis n.sp. found in Silesian Illyrian, is the oldest species of the genus Salpingoporella. A species belonging to the tribe Salpingoporelleae, Nanopora anglica has been reported from the Carboniferous Upper Visean of Lublin region (, 1986).
Salpingoporella krupkaensis n.sp. is the only Triassic species of Macroporella sensu lato with consequently euspondyl whorls and branches perpendicular to the long axis of the thallus. Therefore it differs from all other Triassic species. There are some superficial similarities with Kantia comelicana (), because if observed from inside on fractures, its branches are also weakly attached to the wall of central cavity. However, they differ in the shape of their branches (phloiophorous in Salpingoporella krupkaensis and vesiculiferous in Kantia comelicana). Branches are never grouped in clusters in Salpingoporella krupkaensis and are always arranged in clusters of three or four in Kantia comelicana.
Stratigraphic range and geographic distribution.- Salpingoporella krupkaensis
occurs together with Physoporellae and frequently with Diplopora annulatissima (see Fig. 5 ![]() ). Therefore, the age of this part of the Diplopora Dolomite is Illyrian. Salpingoporella krupkaensis is the most frequent in Chrzanów region (Wilkoszyn Syncline) and in the Olkusz-Siewierz Monocline, but it occurs also in the Zawiercie boreholes (Pl. XXXVI, 2-3
). Therefore, the age of this part of the Diplopora Dolomite is Illyrian. Salpingoporella krupkaensis is the most frequent in Chrzanów region (Wilkoszyn Syncline) and in the Olkusz-Siewierz Monocline, but it occurs also in the Zawiercie boreholes (Pl. XXXVI, 2-3 ![]() ) and in the Upper Silesia, in the Bytom Syncline. This species has not yet been found in the Opole Triassic.
) and in the Upper Silesia, in the Bytom Syncline. This species has not yet been found in the Opole Triassic.
Subtribe Oligoporellinae et al., 1979
Oligoporella , 1912
Oligoporella prisca group
Oligoporella prisca , 1912
1912 Oligoporella prisca .- , Pl. V (IV), 1-2.
1912 Physoporella pauciforata emend.- , Pl. V (IV), 18
1964 (?) Oligoporella cf. prisca ? - , Pl. IX, 4, 5
1973 Oligoporella cf. prisca (= Physoporella pauciforata var. pauciforata).- & , Pl. VIII, 31
1986 (?) Oligoporella cf. prisca - , Pl. I, 4, excerpt from (1964, Pl. IX, 6)
2013 Oligoporella prisca .- in & , Pl. V (IV), 1-2.
Material.- This species is relatively frequent in several localities of the Diplopora Dolomite. It is however difficult to distinguish from Oligoporella elegans ex . The best specimens were collected from locality Żelisławice (no. 72).
Description and discussion - The cylindrical tube is straight or rarely slightly banded. The average diameter is more than 1 mm. The outer wall is flat, without undulation. Axial cavity is narrower than O. elegans. The common feature of this two compared species is the presence of single row whorls, wedge-like, mainly trichophorous, but rarely phloiophorous. Nevertheless, the whorls of O. prisca are very dense and perpendicular to the outer wall, rarely oblique. The tubercles of neighbouring whorls are alternating. The number of whorls per 1 mm of tube length is 4. Thus the average whorl spacing is 0.2 mm. Numbers of branches (tubercles) in one whorl varies from 14 to 18.
Stratigraphical range.- Oligoporella prisca is known from Middle Anisian Reifling limestone in the Northern Alps. J (1964) described Oligoporella cf. prisca from Southern Slovakia in Pelsonian limestones. In the Upper Silesia and adjacent regions this species occurs in Pelsonian and Lower Illyrian Diplopora Dolomite.
Oligoporella elegans ex , 1931a
1872a Cylindrella silesiaca. - Pl. D.IV, 4a-d.
1926a (nom. nud.) Diplopora elegans n.sp.- , p. 55-56, without figure.
1930a Diplopora elegans.- , p. 172.
1931a Oligoporella elegans n.comb.- , Pl. XXI, 3-6.
1931a Oligoporella silesiaca.- , p. 272.
1938 Oligoporella elegans.- , fig. 8.b., excerpt from (1931a, Pl. XXI, 3).
1944 Diplopora elegans.- , p. 63.
1960 Diplopora elegans.- & , p. 690-691.
1986 Oligoporella elegans.- , Pl. CV, 22, excerpt from (1931a, Pl. XXI, 4); Pl. CV, 23, excerpt from (1931a, Pl. XXI, 3); Pl. CV, 24-25, excerpt from (1931a, Pl. XXI, 5-6).
2000 Oligoporella elegans.- & , p. 111.
Syntypes.- , 1931a, Pl. XXI, 3-6.
Type locality.- Nowe Koszyce no. 7, NW Strzelce Opolskie, Upper Silesia.
Derivation of name.- Probably after distinct and elegant arrangement of pores.
Diagnosis.- Long and narrow central stem with thin calcareous wall. Pores are perpendicular or slightly oblique to the wall and are open from outside. They are arranged in single whorls. Distance between whorls is rather large (0.2 mm). The number of pores in one whorl is about 12-20. The shape of pores is generally trichophore (wedge-like), but one can observe the enlargement of pores in some thin sections. Measurements: D=0.58-1 mm, d=0.27-0.64 mm, D:d=46%-63%. This species belongs to the Oligoporella prisca group (one row of pores per one whorl).
Material.- Description of weathered specimen offered to by and 4 figures of thin sections in (1931a, figs. 3-6) from J. collection, Naturhistorisches Museum in Vienna (Austria). Beside this are several weathered specimens in the Z. 's collection MuzPIG 1653.II at the Polish Geological Institute in Warsaw (Poland), as well as in the collections of thin sections MuzPIG 1682.II. It is one of the most frequent Dasycladales species in the Upper Silesia and adjacent regions.
Descriptions and discussion.- The species name elegans has been proposed by P. (1926a) as Diplopora elegans n.sp. (p. 55-56). However, it was a nomen nudum, because the form in question has not been illustrated. The specimens he described came from an area south of Tarnowskie Góry (Segiecki Las, Segiet hunting cottage, site no. 11). noted that besides large specimens of Diplopora annulata () [D. annulatissima and Physoporella praealpina ] the locality yields visibly different small specimens 1 mm in diameter. Cylindrical, thick walled tube is slightly bent. It is covered with minute pores being the outlets of branches. The annular segments are barely visible on the outer surface of the tube. They are 0.4-0.5 mm high, and 0.8-1.0 mm wide. There are 4 rows of pores per segment. He believed that this resembles Alpine species, like Gyroporella triasina or G. multiserialis .
J. (1930a, p. 172) originally accepted 's name Diplopora elegans, but later (, 1931a, p. 273-274) changed it to Oligoporella elegans. He received from a quite weathered specimen, being one of those described by . The steinkern revealed that the pores are filled from the axial cavity up the surrounding rock. Thus the pores were open from outside, and not closed as in Physoporella lotharingica. The tube was about 0.8 mm across. There are about 4 whorls of pores per 0.7 mm of tube length. Thus the average whorl spacing is 0.175 mm.
This description of the weathered specimen is consistent with the description of thin sections (,
1931a, Pl. XXI, 3-6, reproduced herein as Fig. 12e-h ![]() ). They clearly show the single row whorls, wedge-like. Number of branches in one whorl varies from 12 to 20. The pores are usually slightly obliquely directed, but not so steep as in Oligoporella silesiaca (). The tube is often slightly undulating. In some sections, the pores widen outwards (,
1931a, Pl. XXI, 6). The dimensions of the four illustrated specimens are as follows:
). They clearly show the single row whorls, wedge-like. Number of branches in one whorl varies from 12 to 20. The pores are usually slightly obliquely directed, but not so steep as in Oligoporella silesiaca (). The tube is often slightly undulating. In some sections, the pores widen outwards (,
1931a, Pl. XXI, 6). The dimensions of the four illustrated specimens are as follows:
| I | II | III | IV | |
| D | 1 mm | 0.74 mm | 0.76 mm | 0.58 mm |
| d | 0.64 mm | 0.38 mm | 0.37 mm | 0.27 mm |
| D/d | 63 % | 52 % | 49 % | 46 % |
According to , the Oligoporella elegans is very close to Oligoporella prisca. Despite several differences (the Silesian specimens are a little smaller than the Alpine ones, have wider axial cavity and lesser pore density), they exhibit important common features - single-row whorls and thinning pores, occasionally slightly widening outwardly. He suggested, however, that both species should be retained, until intermediate forms will be found. This was also the opinion of J. (1964, p. 116), although he remarked that the relationship of this species to Oligoporella prisca is unclear. Besides the type locality at Nowe Koszyce (no. 7), identified Ol. elegans from the following localities: Stare Repty (no. 10), Fryderyk Deep Shaft (no. 12) and Leśna borehole. Its co-occurrence with D. annulatissima indicates its late Anisian age.
Oligoporella elegans () was schematically illustrated by M.
(1938, fig. 8b). This
slightly generalized drawing of (1931a, Pl. XXI, 3) is reproduced herein (Fig. 12i ![]() ). Also Z. (1986, Pl. CV, 22-25) reproduced all four original figures by .
). Also Z. (1986, Pl. CV, 22-25) reproduced all four original figures by .
Oligoporella elegans has been found in very numerous localities within the Upper Silesia and adjacent regions (see Fig. 2 ![]() ). It occurs both in Pelsonian, and in Illyrian. It is very common in the thin sections from the Opole Triassic: from Tarnów Opolski (locality no. 2) and Kamień Śląski (no. 3) - see Pl. V
). It occurs both in Pelsonian, and in Illyrian. It is very common in the thin sections from the Opole Triassic: from Tarnów Opolski (locality no. 2) and Kamień Śląski (no. 3) - see Pl. V ![]() . It is also abundant in thin sections from Laryszka (no. 5), Suchodaniec (no. 6), Nowe Koszyce (no. 7) and Jemielnica (no. 8), as well as from the Tarnowice syncline: Segiet (no. 11), Fryderyk Deep Shaft (no. 12), Srebrna Góra (no. 13), Łabędy (no. 14), and Wieszowa (no. 15). It is fairly numerous in the Bytom syncline: in Bytom (no. 23), Dąbrówka Wielka (no. 24), Przełajka (no. 25), Kamyce (no. 26), Wojkowice (no. 27-29), Grodziec (no. 33), and Czeladź (no. 34, 35). In the Cracow-Chrzanów area, Oligoporella elegans () is most abundant in thin sections from Balin (no. 50), and Cezarówka (no. 51). It is also common in Krupka (no. 52) and Klucze (no. 57). In the Siewierz region, the species is most common in Żelisławice (no. 72). In the boreholes from the Zawiercie area, it is most abundant in the borehole 51-Ż (Myszków), and in borehole 34 Ż (Żarki Letnisko). This species ranges farthest north, as it has been found in the borehole Odra IG 1. Oligoporella elegans () from the thin sections is illustrated in
Pls. V
. It is also abundant in thin sections from Laryszka (no. 5), Suchodaniec (no. 6), Nowe Koszyce (no. 7) and Jemielnica (no. 8), as well as from the Tarnowice syncline: Segiet (no. 11), Fryderyk Deep Shaft (no. 12), Srebrna Góra (no. 13), Łabędy (no. 14), and Wieszowa (no. 15). It is fairly numerous in the Bytom syncline: in Bytom (no. 23), Dąbrówka Wielka (no. 24), Przełajka (no. 25), Kamyce (no. 26), Wojkowice (no. 27-29), Grodziec (no. 33), and Czeladź (no. 34, 35). In the Cracow-Chrzanów area, Oligoporella elegans () is most abundant in thin sections from Balin (no. 50), and Cezarówka (no. 51). It is also common in Krupka (no. 52) and Klucze (no. 57). In the Siewierz region, the species is most common in Żelisławice (no. 72). In the boreholes from the Zawiercie area, it is most abundant in the borehole 51-Ż (Myszków), and in borehole 34 Ż (Żarki Letnisko). This species ranges farthest north, as it has been found in the borehole Odra IG 1. Oligoporella elegans () from the thin sections is illustrated in
Pls. V ![]() and
VI
and
VI ![]() , and from fractures in Pl. VII
, and from fractures in Pl. VII ![]() .
.
Strangely, such an abundant species has not been recorded before 's studies and 's identification. However, noted (1931a, p. 272), that Cylindrella silesiaca (1872a, p. 281-282, Pl. D.IV, 4a-d) might be a peculiar form of preservation of Oligoporella elegans.
(1872a, p. 281-282) wrote that Cylindrella silesiaca is an abundant species found in Upper Silesian Muschelkalk, locally known as the Jemielnica Dolomite. Below is 's description of this species, important for identifying isolated, weathered specimens of Oligoporella elegans (Fig. 12a-d ![]() ).
).
"Minute cylindrical, slightly curving tubes do not have annuli on their surface, but instead are covered with delicate points of pores, so that they are smooth or moderately rough. The tube has two walls: outer and inner one. The outer wall is thin and fragile; if broken, it reveals the underlying inner tube, containing a wide inner chamber. The inner tube is connected with the outer tube via numerous tubules (tubercles) arranged in rings, and extending from an annular swelling. The tubules are hollow and contain canals leading from the inner chamber towards the outer surface. Thus, between the inner and outer tubes of the cylinder there is a fairly large space, crossed only by the delicate tubules".
Independently from , the present author has noticed such preservation in different material (, 1981, 1986, Pl. CVI, 3-4, 6). There is no doubt that the specimens described by are conspecific with Oligoporella elegans , as evident from their single row rearrangement of branches (canals), oriented slightly obliquely and minute size of double tubes. Thus, one may wonder, if 's name should not be given priority over that by . However, the species epithet silesiaca has been used by to another species (Gyroporella silesiaca), belonging to the same genus Oligoporella, according to 's synonymy. Thus it seems reasonable to give up the name Cylindrella silesiaca and retain instead the well established Oligoporella elegans () , widely used since 75 years.
F. (1958) noted that his new species Oligoporella (?) n.sp. somewhat resembles Oligoporella elegans. In his opinion, the slender thalli, prepared with acetic acid etching (op. cit., Pl. 4, 10-15), are close to those of the Upper Silesian species. Later, however, the 's new species has been synonymised by G. (1961) with Anisoporella occidentalis (see & , 2000, p. 11), an index species for the Lower Anisian.
M. (1967) described a Ladinian species from the Chios Island on the Aegean Sea, Oligoporella chiae. It has single rows of thin trichophore branches (pores), arranged slightly obliquely to the axial cavity. He noted the similarity of this species to the Anisian species Oligoporella pilosa , from which it differs in having widely spaced single-row whorls and in pronounced undulation. The presence of single-row whorls indicates, however, that Oligoporella chiae belongs to the Oligoporella prisca group, together with Oligoporella elegans . O. chiae differs from the latter two species in its filamentous oblique trichophorous pores (occasionally the pores may be sausage-like; , 1972b), wide spacing of the whorls, and strong undulation.
The following sentence by P. (1944, p. 63) on occurrence of calcareous algae in dolomitic and limestone strata of Middle Muschelkalk has generated substantial controversy: "Die Formen gehören nach J. nicht wie er früher annahm (1927, S. 197) zu der im mediterranen Triasgebiet vorkommenden Diplopora annulata, sondern zu D. elegans, die auf das oberschlesische Gebiet in weiterem Sinne beschränkt ist". This sentence was quoted by C. and S. (1960, p. 690), who rightly remark that does not identify the paper by supposedly claiming that in the Upper Silesia Diplopora annulata would be replaced by Diplopora elegans. S. (1952, p. 106, 108), following , and not knowing important papers by , wrote that the alga Diplopora annulata would be replaced by Diplopora elegans characteristic for the Silesian region. J. (1930a, p. 171-173; 1931a, p. 273-274) stated clearly that the Upper Silesian forms include only three well recognized species: Oligoporella elegans (probably limited in its range to the Upper Silesia), undisputable Diplopora annulata with beautiful specimens, and the most abundant Diplopora annulatissima, an index fossil for the Illyrian.
C. and S. (1960) suggested instead that Oligoporella elegans occurs in lower strata than the Diplopora annulata and D. annulatissima, namely in Pelsonian Karchowice Beds, even though they did not find it in those strata. Actually, Oligoporella elegans occurs in numerous localities within the Diplopora Dolomite, both in Pelsonian (Physoporella pauciforata - Oligoporella pilosa assemblage Zone), and in Illyrian, together with Diplopora annulatissima. The controversial and unsubstantiated 's and 's opinion, resulting from ignoring 's work, is thus of only historical interest.
 |
|
|
Click on thumbnail to enlarge the image.
Figure 12: Previously published illustrations of Oligoporella elegans ex from the Upper Silesia: a-d pencil drawings by (1872a: Pl. D.IV, 4a-d) showing broken or whole specimens assigned by him to Cylindrella silesiaca: a - partly fractured specimen enabling to see the interior, x 15; b - the same specimen, actual size; c - vertical fracture of the specimen, x 15; d - specimen with preserved surface, actual size; e-h - pencil drawings of polished sections of newly erected Oligoporella elegans by (1931a, figs. 3-6): e - slightly oblique longitudinal section x 15; f - tangential section, x 16; g - oblique section, x 16, h - oblique section, x 16; i - schematic drawing by (1938, fig. 8b), redrawn from (1931a, fig. 3) and reduced (compare i and e herein), x 7.5.
Oligoporella pilosa group
Oligoporella pilosa , 1912
1912 Oligoporella pilosa .- , Pl. IV (III), 1-4, 6-7.
1920 Oligoporella pilosa .- , p. 48.
1935a Oligoporella pilosa typica .- , text-fig. 5-15.
1957 Oligoporella pilosa typica .- , Pl. IV, fig. 1-2.
1964 Oligoporella pilosa var. pilosa , , Pl. X, 2, 4, 6
1965 Oligoporella pilosa .- , Pl. XI, 4-9.
1986 Oligoporella pilosa var. pilosa .- , Pl. I, 9.
1994 Oligoporella pilosa var. pilosa .- E. et al., Pl. 2, 3.
1997 Oligoporella pilosa var. pilosa ex .- I. , Pl. I, 1-17.
2013 Oligoporella pilosa .- in & , Pl. IV (III), 1-4, 6-7.
Material. - This species is probably largely distributed in the outcrops of Diplopora Dolomite, but the state of preservation makes difficult to determine it with certitude. Several good preserved specimens derive from locality Żelisławice (no. 72) - see Pl. V ![]() .
.
Description and discussion.- The thallus is cylindrical, without distinct undulation. Tubercles (branches) of trichophorous type are rather very thick at the base, thinning outward. They are arranged in double rows, separated by narrow spaces. Tubercles are perpendicular to the external wall, rarely slightly oblique. The diameter of the thallus is 1.5-2.5 mm. Axial cavity is large (1.3-1.8 mm). The number of whorls per 1 mm of tube length is 2. The average whorl spacing is 0.5 mm. Number of branches (tubercles) per one whorl varies from 16 to 20.
Several varieties were distinguished by J. (1912, in & , 2013) and J. (1964). They are described in this section only. M. (1965) is of the opinion, that they all represent intraspecific variability.
Stratigraphical range.- Oligoporella pilosa is largely distributed in Dinarides (Bosnia, Dalmatia) and Southern Alps in the Anisian limestones (Bithynian, Pelsonian) and also in Southern Slovakia (Slovakian Karst, Plešivecka Planina) where it is of Pelsonian and Lower Illyrian age (, 1964). In the Upper Silesia and adjacent regions it occurs together with Physoporellae (Pelsonian) and Diplopora annulatissima (Lower Illyrian).
Oligoporella silesiaca (, 1872a)
1863 pars Diplopora nodosa n.sp.- , Pl. LXV. e, 19-20.
1872a non Cylindrella silesiaca n.sp.- , Pl. D.IV, 4a-d.
1872a Gyroporella silesiaca n.sp.- , Pl. D.III, 6, 7a-b?, p. 276.
1872a Gyroporella infundibuliformis.- , Pl. D III, 8a-c, 10, p. 277.
1920 Diplopora annulata or Teutloporella nodosa.- , p. 30.
1926a Diplopora silesiaca.- , p. 505, not figured.
1928 Diplopora silesiaca.- , fig. 12, excerpt from (1872a, Pl. D.III, 6).
1931a Oligoporella elegans.- , p. 272.
1960 Diplopora cf. silesiaca.- & , fig. 7; Pl. III, 13 [= fig. 7].
1979 Diplopora nodosa.- .
2000 (?) Diplopora nodosa.- & .- p. 78.
2000 Diplopora minutula.- & .- p. 127.
Holotype.- , 1872a, Pl. D III, 6 (Fig. 13a ![]() herein)
herein)
Neotypes.-
& , 1960, Pl. III, 13; Fig.
7 (from , 1995); Pl. VIII, 8 (Fig. 13h-j ![]() herein).
herein).
Type localities.- Jemielnica (no. 8) in the Upper Silesia, Opole Triassic Region; Kąty Chrzanowskie (Rosowa Góra; no. 48) - Cracow-Chrzanów Region.
Derivation of name.- After distribution in the Diplopora Dolomite of Upper Silesia.
Diagnosis.- Cylindrical thallus of small diameter (ca. 2 mm) with double whorls of trichophorous tubercles. The elongated tubercles are arranged very obliquely to the thallus, forming cones merging down in such a whorl. Tubercles are alternating in the double whorl. Their number per one whorl is about 20. This species belongs to the Oligoporella pilosa group (two rows of pores per one whorl).
Material.- Two type specimens - holotype of and neotype of & . Beside this, there is another neotype by and several specimens in his collection MuzPIG no. 1653.II, from localities Jemielnica (no. 8), Przełajka (no. 25), Grodziec (no. 33), Czeladź (no. 34), Będzin (no. 36, 37), Libiąż (no. 45), Klucze (no. 57) and Brudzowice (no. 71). Among them is a third neotype, from the type locality Kąty Chrzanowskie (Rosowa Góra, locality no. 48; see Pl. VIII, 8 ![]() ), where it is very abundant.
), where it is very abundant.
Nomenclature and discussion.- The species Oligoporella silesiaca has been described and illustrated by
(1872a) as Gyroporella silesiaca, and the specimen figured in Pl. D.III, 6 is regarded as the holotype. This specimen from Jemielnica has very steeply oriented long, pointed tubercles in double whorls, merging downwards into sort of cone-in-cone structure. This specimen is a breakage with partly preserved shell, slightly undulating. The holotype of is illustrated in this paper (Fig. 13a ![]() ). A similar conical arrangement of tubercle whorls is visible in specimens from the Mendola Dolomite in Italian Alps (Pl. D III, 7a-b; Fig. 13b-c
). A similar conical arrangement of tubercle whorls is visible in specimens from the Mendola Dolomite in Italian Alps (Pl. D III, 7a-b; Fig. 13b-c ![]() herein). Perhaps this species includes also specimens described as Gyroporella infundibuliformis in Pl. D III, 8a-c, 10; this Hungarian form (Fig. 13d
herein). Perhaps this species includes also specimens described as Gyroporella infundibuliformis in Pl. D III, 8a-c, 10; this Hungarian form (Fig. 13d ![]() ) is similar to the Upper Silesian specimens, according to . The above specimens differ markedly from specimens that classified as Cylindrella silesiaca, and that were included by J. (1931a) into his species Oligoporella elegans. Such a position is accepted herein.
) is similar to the Upper Silesian specimens, according to . The above specimens differ markedly from specimens that classified as Cylindrella silesiaca, and that were included by J. (1931a) into his species Oligoporella elegans. Such a position is accepted herein.
J. (1920) included 's species into Diplopora annulata sensu lato. However, P.
(1926a) regarded 's species as valid, repeated its diagnosis and named Diplopora silesiaca (). But neither he, nor have subsequently found this species in the Upper Silesian material. M. (1928) also accepted validity of Diplopora silesiaca and reproduced a simplified version of the figure by
(1872a, Pl. D.III, 6) of the Jemielnica specimen, reproduced also herein (Fig. 13g ![]() ). (1931a) doubted the existence of this species, because he did not observe it and thus believed it could belong to Oligoporella elegans. Besides, he speculated (
1920, p. 27) that the original calcareous tube could have been replaced with a secondary carbonate incrustation; thus it would represent not a separate species, but a particular mode of preservation.
). (1931a) doubted the existence of this species, because he did not observe it and thus believed it could belong to Oligoporella elegans. Besides, he speculated (
1920, p. 27) that the original calcareous tube could have been replaced with a secondary carbonate incrustation; thus it would represent not a separate species, but a particular mode of preservation.
It should be noted, that when referring to that species, various authors use different generic names: Gyroporella (), Diplopora (, , ), and Oligoporella (). The elongated shape of its rarely spaced pointed branches (tubercles) suggests its inclusion to the genus Oligoporella. It cannot belong to Diplopora, because there are no branches spreading from one point and forming metaverticillate tufts.
E. and S. (1960) studying material collected in Kąty Chrzanowskie (Rosowa Góra - locality no. 48) identified a form
labelled by them Diplopora cf. silesiaca, which could be treated as a neotype of this species (
& , 1960, Pl. III, 7, and Fig. 13h-j ![]() herein). In the preserved fragment of a cylinder there are three double whorls of tubercles. The tubercles of trichophore type are situated obliquely to the cylindrical surface and are elongated, and taper distally. The tubercles alternate, and there are 20 per whorl. The double whorls resemble cones; all tubercles of each whorl converge downwards. Double whorls are separated by wide spaces (Fig. 13h-j
herein). In the preserved fragment of a cylinder there are three double whorls of tubercles. The tubercles of trichophore type are situated obliquely to the cylindrical surface and are elongated, and taper distally. The tubercles alternate, and there are 20 per whorl. The double whorls resemble cones; all tubercles of each whorl converge downwards. Double whorls are separated by wide spaces (Fig. 13h-j ![]() ).
).
Such a description mostly fits the original diagnosis by . However, the species silesiaca should not be classified within the genus Diplopora, because the branches do not form tufts. Evidently, the authors followed 's generic assignment, not knowing the paper by J. (1931a), who suggested inclusion of the 's species to Oligoporella elegans. Inclusion of the species silesiaca in Oligoporella seems justified, but it could not be Oligoporella elegans, because the latter species resembles Oligoporella prisca with a single row of trichophore branches per whorl. The double-row whorls suggest a close relationship between Oligoporella silesiaca and Oligoporella pilosa instead.
Some similarity of the discussed species to Diplopora nodosa () has been noted; and that the latter names would have priority (, 1979; & , 2000). It has not been proven however, that in D. nodosa the branches form tufts. J. (1920) thoroughly discussed the inadequacy of assigning this species to Diplopora, because of the lack of tufts. That is why he assigned it to the genus Teutloporella. The segments typical to Teutloporella nodosa are indeed somewhat similar to the tubercles forming whorls in the species silesiaca, but the branches are differently arranged within segments. Besides, the branches in Teutloporella are much smaller and not so obliquely directed as in the species silesiaca. This is another reason for a separate status of Oligoporella silesiaca. It is, however fully appropriate to use the name Teutloporella nodosa, instead of Diplopora nodosa, as proposed by & (2000). This is supported by morphological features, especially by the presence of tufts. The purely formal priority issues should be overruled by the nomina conservanda principle, because the name Teutloporella nodosa is widely used for 80 years.
 |
|
|
Click on thumbnail to enlarge the image.
Figure 13: Previously published illustrations of Oligoporella silesiaca () from the Upper Silesia and similarly preserved specimens from the Alps: a - pencil drawing of the holotype specimen (from Jemielnica, Opole Silesia) of his new species Gyroporella silesiaca by (1872a, Pl. D.III, 6), x 15; b-c - pencil drawings of a conspecific specimen from Mendola Dolomite (Italian Alps) by (1872a, Pl. D.III, 7a, b): b - actual size, c - x 5; d-f - specimens from Hungary similar to the Silesian ones according to (1872a, Pl. D.III, 8a-c): d - actual size, e and f - x 5; g - schematic drawing by (1928, fig. 12), redrawn from (1872a, Pl. D.III, 6) and reduced (compare g and a herein), x 8; h-j - photographs (h, i) and drawing (j) of the neotype illustrated by & (1960, Pl. III, 7; text-fig. 13) and designated as Diplopora cf. silesiaca: h - tangential fracture x 46, i - the same x 23, j - x 30.
Oligoporella balinensis (, 1892)
1892 (nom. nud.) Gyroporella balinensis n.sp.- , p. 7-8, without figure.
1960 Oligoporella pilosa forma balinensis n.stat.- & , fig. 9; Pl. III, 14 [=fig. 9].
1960 Oligoporella balinensis.- & , p. 699 (English summary).
1964 ? Oligoporella pilosa.- , p. 111.
2000 Oligoporella pilosa var. balinensis.- & , p. 112.
Neotype.- & , fig. 9; Pl. III, 14 [= fig. 9]
Type locality.- Balin, near Chrzanów, locality no. 50 (lost 's holotype), Kąty Chrzanowskie (Rosowa Góra, no. 48) near Chrzanów (neotype of & ).
Derivation of name.- After first discovery by (1892) in Balin.
Diagnosis.- Very small cylindrical thallus of 1.2 mm diameter. Tubercles are arranged in double whorls, distinctly separated. Tubercles alternating in one whorl have shape similar to trichophorous. The height of tubercles does not exceed 0.3 mm. They are arranged obliquely to the central stem. This species belongs to the Oligoporella pilosa group (two rows of pores per one whorl).
Material.- The photograph of neotype in & (1960, Pl. III, 14 = fig. 9). Collection preserved in Chair (Department?) of Ore Deposits, AGH University of Science and Technology (Katedra Złóż Rud AGH), Cracow (Poland). Several specimens (photo V) from Balin are preserved in MuzPIG collection no. 1653.II, locality no. 50, of Z. .
Nomenclature and discussion.- M. (1892, p. 532-533) noted, that in Balin there are tiny Dasycladales, much smaller than those known to him. He did not give a full diagnosis of this minute form. He wrote only that "on each very low ring there are two rows of tubules" (p. 532-533). By then the monograph by (1872a) was already known, with descriptions of several Upper Silesian taxa, e.g., Gyroporella silesiaca, G. cylindrica, and G. minutula. There was also a paper by A. (1878), where he reproduced specimens of 's (1872a) Cylindrum annulatum (, 1878, Pl. VI, 10-11). And it was to those species that compared the form he discovered and regarded as a new species Gyroporella balinensis. It is, however, a nomen nudum, because the species was not illustrated by this author. 's collection with this new species has probably been lost, because it could not be located in any of the Cracow's museums.
The specific epithet balinensis has been revived only after 68 years by C. and S. (1960). Exploring for ore deposits near Chrzanów, they found in a
borehole from Kąty Chrzanowskie and in Rosowa Góra outcrop (locality no. 48) numerous tiny Dasycladales and the smallest of these they regarded as the equivalent of the species described by ( & , 1960, Pl. III, 14). If we should regard it as a neotype of this species, than it fits the above description. The diagnosis of the species is given after the neotype designated by & (Fig. 14a-c ![]() herein). They regarded it as a variety of the species Oligoporella pilosa differing only by its smaller size - in Ol. pilosa the tube diameter ranges from 1.38 to 3.00 mm, while in Ol. balinensis reaches merely 1.2 mm. J. (1964) was of opinion, that the size difference is not sufficient to establish a new variety, especially that it is based on the weathered filling of the axial cavity and pores, while the calcareous sleeve is missing. Because the general shape of the pores and their passage through the calcareous shell are unknown, he regards the comparisons with Oligoporella pilosa as problematic. It should be noted, however, that the presence of double row of branches in the whorl and their trichophore shape suffice to compare the discussed form with Ol. pilosa. It is also noteworthy, that & (1960) in their description use a name Oligoporella pilosa f. balinensis , but in the English abstract (p. 699) they restored Oligoporella balinensis (). Also the similarity between Oligoporella balinensis and Oligoporella silesiaca should be noted. Both species are generally similar to Oligoporella pilosa (double rows of trichophore pores), but differ from the latter in having more steeply arranged pores, being a feature common to both these species.
herein). They regarded it as a variety of the species Oligoporella pilosa differing only by its smaller size - in Ol. pilosa the tube diameter ranges from 1.38 to 3.00 mm, while in Ol. balinensis reaches merely 1.2 mm. J. (1964) was of opinion, that the size difference is not sufficient to establish a new variety, especially that it is based on the weathered filling of the axial cavity and pores, while the calcareous sleeve is missing. Because the general shape of the pores and their passage through the calcareous shell are unknown, he regards the comparisons with Oligoporella pilosa as problematic. It should be noted, however, that the presence of double row of branches in the whorl and their trichophore shape suffice to compare the discussed form with Ol. pilosa. It is also noteworthy, that & (1960) in their description use a name Oligoporella pilosa f. balinensis , but in the English abstract (p. 699) they restored Oligoporella balinensis (). Also the similarity between Oligoporella balinensis and Oligoporella silesiaca should be noted. Both species are generally similar to Oligoporella pilosa (double rows of trichophore pores), but differ from the latter in having more steeply arranged pores, being a feature common to both these species.
Oligoporella balinensis is fairly rare in the Diplopora Dolomite. It has been recorded from several localities in the vicinity of Chrzanów (Balin, Rosowa Góra, Krupka), but it was also found in the Opole Triassic, Tarnowice Syncline and Bytom Syncline. It accompanies the Physoporella pauciforata - Oligoporella pilosa assemblage zone from the Pelsonian, and continues into the Illyrian together with Diplopora annulatissima.
 |
|
|
Click on thumbnail to enlarge the image.
Figure 14: Neotype of Oligoporella balinensis () from Kąty Chrzanowskie near Chrzanów (Rosowa Góra - locality no. 48) as illustrated by & (1960):
a - photograph of the neotype of Oligoporella pilosa f. balinensis ( & , 1960, Pl. III, 14), x 45;
b - drawing ( & , 1960, Pl. III, text-fig. 14), x 30;
c - the same as in a, reduced twice (x 22.5) to facilitate comparison with b, and with the newly found specimens (Pl. IX ![]() ).
).
Oligoporella chrzanowensis group
Oligoporella chrzanowensis n.sp.
1960 Gyroporella multiserialis .- & , Pl. II, 11.
Holotype.- Pl. X, 1 ![]() .
.
Paratypes.- Pl. X, 2, 5, 14 ![]() ; Fig. 15
; Fig. 15 ![]() .
.
Repository.- Polish Geological Institute, Warsaw, Geological Institute, coll. of Z. no. MuzPIG 1653.II.
Derivation of the name.- After the type area Chrzanów: locality Kąty Chrzanowskie (Rosowa Góra), no. 48, and Balin, no. 50, in the Cracow-Chrzanów region.
Type horizon.- Upper part of the Diplopora Dolomite, Upper Anisian.
Diagnosis.- Thallus without annulation or undulation, with amassment of short, trichophore oblique branches (tubercles) arranged in indistinct, dense whorls (euspondyl arrangement) or irregularly (aspondyl arrangement).
Description.- In fractures one can observe large (1.5-3 mm in diameter) fragments of thallus, covered with densely arranged tubercles. The tubercles are oblique and form regular cones with wide bases, rapidly thinning
(Pl. X, 1-2, 5, 12, 23-24 ![]() ). Whorls exist, but they are not always observable, frequently the arrangement of tubercles is irregular. In some cases rows were observed in groups of 3 or 4 (Fig. 15
). Whorls exist, but they are not always observable, frequently the arrangement of tubercles is irregular. In some cases rows were observed in groups of 3 or 4 (Fig. 15 ![]() ). Central cavity is moderately large, with a distinct internal wall
(Pl. X, 15, 23
). Central cavity is moderately large, with a distinct internal wall
(Pl. X, 15, 23 ![]() ). External wall is not preserved, and the exact outline of the alga is not observable.
). External wall is not preserved, and the exact outline of the alga is not observable.
Material and localities.- Several specimens on the fractures from the Cracow-Chrzanów region (localities: Rosowa Góra, no. 48; Balin, no. 50; Cezarówka, no. 51; Krupka, no. 52), Olkusz-Siewierz Monocline (Stare Gliny, no. 57; Żelisławice, no. 72), deep borings of the Zawiercie region (boreholes: 10Z Lgota Nadwarcie; 25 WW Winowno), Bytom Syncline (localites: Kamyce, no. 26; Czeladź, no. 35), Opole Triassic (Jemielnica, no. 8).
Discussion and comparisons.- Attribution of Oligoporella chrzanowensis n.sp. to the genus Oligoporella is based on the shape of branches, as they are clearly trichophore, forming short, regular cones. This is the difference from Teutloporella, having thin and long euspondyl or aspondyl branches. Also, Teutloporella has twice as many branches per whorl. The newly described species cannot belong to the Diplopora multiserialis (comp. & , 1960) because the tubercles are not arranged in clusters of vesicular type nor there is any observable multiseriality. And D. (Gyroporella) multiserialis , 1872a, is a junior synonym of D. annulata , 1853, according to (1912, in & , 2013).
Within the genus Oligoporella there are two groups of species (, 1967): the Oligoporella prisca group, with one row of pores per one whorl, and Oligoporella pilosa group, with two rows of branches in one whorl). Oligoporella chrzanowensis does not belong to neither of these groups. It forms a third group, indistinctly euspondyl or aspondyl, with oblique short-conical branches. Oligoporella elegans is typically single-row euspondyl species, belonging to the Oligoporella prisca group. To the contrary, euspondyl species Oligoporella silesiaca and Oligoporella balinensis possess two rows of branches in each whorl and belong to the Oligoporella pilosa group. A feature common to these two species shared with Oligoporella chrzanowensis is the oblique arrangement of their branches.
Stratigraphical range and geographical distribution.- Oligoporella chrzanowensis n.sp. is very frequent in the vicinity of Chrzanów, where it occurs with Upper Pelsonian Physoporellae and other Oligoporellae (locality no. 48, Rosowa Góra), and Illyrian Diplopora annulatissima and Kantia comelicana. (locality no. 50, Balin, and no. 57, Stare Gliny). Occurrence in other regions is sporadic.
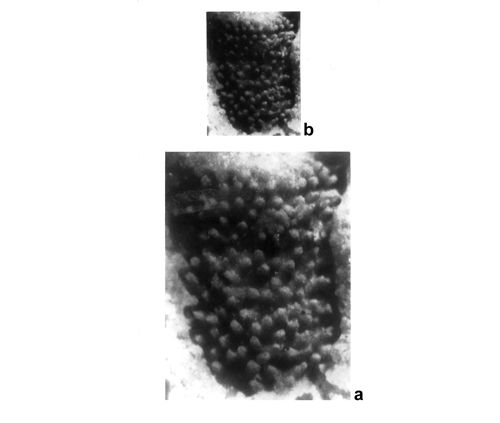 |
|
|
Click on thumbnail to enlarge the image.
Figure 15: Oligoporella chrzanowensis n.sp. from Kąty Chrzanowskie near Chrzanów (Rosowa Góra - locality no. 48) as illustrated by
& (1960): a - photograph of a specimen assigned to Gyroporella multiserialis by & (1960, Pl. II, 11), x 13;
b - the same as in a, reduced twice (x 6) to facilitate comparison with other photographs of Oligoporella chrzanowensis n.sp. (Pl. X ![]() ).
).
Physoporella pauciforata group
Physoporella pauciforata (, 1872a) , 1912
Pls. XI ![]() ,
XVIII
,
XVIII ![]() -
XIX
-
XIX ![]() -
XX
-
XX ![]() -
XXI
-
XXI ![]() &
XXXII
&
XXXII ![]() -
XXXIII
-
XXXIII ![]() -
XXXIV
-
XXXIV ![]() ;
Fig. 16
;
Fig. 16 ![]()
1872a Gyroporella pauciforata n.sp. , Pl. D.III, 2b-c, 3a-b.
1872a Gyroporella minuta [sic] , Pl. D.III, 5a, b.
1872b Gyroporella pauciforata n.sp. .
1903 Physoporella pauciforata n.comb.- , p. 50.
1903 Physoporella pauciforata.- , p. 17.
1912 Physoporella pauciforata.- , Pl. V (IV), 9-19.
1920 Physoporella pauciforata.- , Pl. 3, 10-15.
1927 Physoporella pauciforata.- , Pl. IX, 5; Pl. X, 4a-b; Pl. XIII, 7a-b.
1928 Gyroporella cf. minutula.- , fig. 11b.
1935 Physoporella pauciforata var. simplex n.var.- , p. 223.
1962 Physoporella pauciforata () var. gemerica n.var.- , Pl. 4, 4-5.
1964 Physoporella pauciforata var. pauciforata n.var.- , Pl. 13, 1-6; Pl. 14, 4, 5b, 6.
1964 Physoporella pauciforata () var. gemerica - , Pl. 16, 1-5.
1965 Physoporella pauciforata var. simplex.- , Pl. XIII, 5.
1967 Physoporella pauciforata var. pauciforata.- , Pl. I, 1.
1967 Physoporella pauciforata.- , Pl. 8, 5.
1972 Physoporella pauciforata var. pauciforata.- et al., Pl. I, 1-3.
1973 Physoporella pauciforata var. pauciforata.- & , Pl. X, 1-6.
1973 Physoporella pauciforata.- & , Pl. IV, 14-15, 16 pars; Pl. VII, 25, 26 pars, 27.
1975 Physoporella pauciforata.- & , Pl. IV, 24.
1986 Physoporella pauciforata.- , Pl. CVI, 9b.
1989 Physoporella pauciforata.- & , Pl. 10, 4.
1994 Physoporella pauciforata.- et al., Pl. 11, fig. 8-9, 11.
2000 Physoporella pauciforata.- & , p. 131.
2013 Physoporella pauciforata.- in & , Pl. V (IV), 9-19.
Diagnosis.- Cylindrical thallus without segmentation. Only very little undulation may be observable. The branches (tubercles) are distributed in single whorls. They are pyriform, mostly closed within the calcareous wall (if present), oriented perpendicularly to the longitudinal axis. Whorls are rather closely spaced. It is a model representative of Physoporella pauciforata group (one row of pores per one whorl).
Material.- This species is present in all study areas - frequent in the Upper Silesia: Opole Triassic (Laryszka, no. 5, Suchodaniec, no. 6, Jemielnica, no. 8), Tarnowice Syncline (Segiet, no. 8), Bytom Syncline (Przełajka, no. 25, Wojkowice-Komorne, no. 28, Boleradź, no. 31), quite rare in the Cracow-Chrzanów region, and abundant in the Olkusz-Siewierz region (Stare Gliny, no. 57, Brudzowice, no. 70, Nowa Wioska, no. 71) and in boreholes of the Zawiercie region (31-KW Morsko, 26-WN Winowno, 42-Ż Dzierżążno, 10-Ż Lgota Nadwarcie, 46-Ż Wysoka, 78-Ż Żarki). All specimens are housed in the Geological Museum of the Polish Geological Institute (Warsaw), Z. 's collection no. 1653.II.
Description.- The three-dimensional structure can be studied in fractures, both from outside (tubercles/branches) and from inside (pore inlets in the axial cavity). The tubercles form whorls (always single rows), close to each other, sometimes arranged in vertical rows
(Pl. XI, 2-3, 15 ![]() ). The tubercles are pyriform, closed from outside. Sometimes they are hollow inside
(Pl. VI, 15
). The tubercles are pyriform, closed from outside. Sometimes they are hollow inside
(Pl. VI, 15 ![]() ). Pores, visible from the axial cavity and being the inlets of branches, form regular single-row whorls
(Pls. XI, 4
). Pores, visible from the axial cavity and being the inlets of branches, form regular single-row whorls
(Pls. XI, 4 ![]() &
XVIII, 4
&
XVIII, 4 ![]() ). Pores are situated in furrows separated by ridges
(Pl. XI, 1
). Pores are situated in furrows separated by ridges
(Pl. XI, 1 ![]() ). The ridges undulate because of the alternating arrangement of pores. The axial cavity is fairly wide
(Pl. XI, 3, 10, 23
). The ridges undulate because of the alternating arrangement of pores. The axial cavity is fairly wide
(Pl. XI, 3, 10, 23 ![]() ) and hollow
(Pl. XVIII, 6
) and hollow
(Pl. XVIII, 6 ![]() ). Sometimes the internal wall is thickened
(Pl. XI, 20
). Sometimes the internal wall is thickened
(Pl. XI, 20 ![]() ). In many cases two walls, internal and external, are visible
(Pls. XVIII, 3
). In many cases two walls, internal and external, are visible
(Pls. XVIII, 3 ![]() -
XIX, 1
-
XIX, 1 ![]() ). The outer surface of the alga is smooth, without annulation nor undulation
(Pls. XVIII, 6
). The outer surface of the alga is smooth, without annulation nor undulation
(Pls. XVIII, 6 ![]() &
Pls. XX, 3, 7
&
Pls. XX, 3, 7 ![]() ). The apex is rounded, with slightly wavy external surface
(Pl. XI, 15
). The apex is rounded, with slightly wavy external surface
(Pl. XI, 15 ![]() ).
).
Comprehensive description of specimens from the fractures is given in explanations to Pl. XVIII ![]() .
(1962, 1964, 1967) discerned several varieties. It is not always possible to assign the specimens studied to particular variety. Nevertheless, most specimens shown in Pl. XI
.
(1962, 1964, 1967) discerned several varieties. It is not always possible to assign the specimens studied to particular variety. Nevertheless, most specimens shown in Pl. XI ![]() belong to Physoporella pauciforata () var. gemerica - . The pyriform (pear-shaped) whorls are very densely spaced and often form also vertical rows. The diameter equals the distance of 4 to 8 whorls, being the most characteristic feature of 's concept of the gemerica variety. In the
Pl. XI, 1-3, 12, 14-15, 21-25
belong to Physoporella pauciforata () var. gemerica - . The pyriform (pear-shaped) whorls are very densely spaced and often form also vertical rows. The diameter equals the distance of 4 to 8 whorls, being the most characteristic feature of 's concept of the gemerica variety. In the
Pl. XI, 1-3, 12, 14-15, 21-25 ![]() , show specimens of the var. gemerica, and only in Pl. XI, 13
, show specimens of the var. gemerica, and only in Pl. XI, 13 ![]() , there is a representative of var. pauciforata. Surprisingly, it is the most southern of 's varieties, typical for the Slovakian Karst and northern Hungary, that is most frequent in the Upper Silesia and adjacent regions, situated much farther north.
, there is a representative of var. pauciforata. Surprisingly, it is the most southern of 's varieties, typical for the Slovakian Karst and northern Hungary, that is most frequent in the Upper Silesia and adjacent regions, situated much farther north.
Stratigraphical range and geographical distribution.- Physoporella pauciforata () is considered as an index form for Upper Pelsonian and Lower Illyrian in former Yugoslavia (, 1935a; , 1965) and in the Slovakian Karst (, 1964, 1986). It has a similar stratigraphic range in the Alps (, 1937a), but in the Austrian Alps it can reach up to Ladinian (, 1974). It is one of the fossils, forming Physoporella pauciforata-Oligoporella pilosa assemblage zone (, 1986). Similar stratigraphic position is established in Romania ( & , 1973) and in Bulgaria ( & , 1973). In the High-Tatric series of the Polish Tatra Mountains Ph. pauciforata occurs together with other Physoporellae of pauciforata and praealpina group, as well as with Diplopora annulatissima , which determines the Illyrian age of the Tatric specimens. This species is a representative of the Briançon - Préalpian - High-Tatric - Bulgarian palaeogeographical province of the Middle Triassic ( & , 1973; , 1977, 1994a, 1994b, 1995; et al., 2004). It belongs to the Physoporella pauciforata-Oligoporella pilosa assemblage zone, but it also continues into Illyrian with D. annulatissima and D. annulata.
Nomenclature and discussion.- Surprisingly, J. never acknowledged the
occurrence of Physoporella algae in the Upper Silesian Triassic, although C.W.
(1872a) clearly mentioned species of this genus. It was mainly due to the fact that underestimated the possibility of precise determination of specimens found on breakage and weathered surfaces, and such is the occurrence of Physoporellae from the Upper Silesia and adjacent regions, where they occur abundantly in numerous localities (Fig. 5 ![]() ).
).
The holotype of Physoporella pauciforata () is recognized as the specimen figured by
(1872a, Pl. D.III, 2b), and coming from Reifling Limestone in Northern Calcareous Alps in Austria. It is a drawing of weathered wavy surface with few wide pores in single-row whorls (Fig. 16a ![]() ). The cross section (,
1872a, Pl. D.III; Fig. 16c
). The cross section (,
1872a, Pl. D.III; Fig. 16c ![]() herein) should be regarded as a syntype. included to this species also Upper Silesian specimens (,
1872a, Pl. D.III, 3b), although he was unsure as to their conspecific nature (fig. 5b). In the next paper (,
1872b) confirmed his doubt about presence of this species in the Upper Silesia. J. (1931a, p. 271) confirmed this doubt. Also from the Upper Silesia come specimens that
(1872a, Pl. D.III, 5a, b), classified as Gyroporella minutula. They cannot belong to G. minutula, because the weathered surface shows wide tubercles arranged in dense single-row whorls, a typical feature of Physoporella pauciforata (Physoporella minutula has double-row whorls; see
Fig. 15d-e
herein) should be regarded as a syntype. included to this species also Upper Silesian specimens (,
1872a, Pl. D.III, 3b), although he was unsure as to their conspecific nature (fig. 5b). In the next paper (,
1872b) confirmed his doubt about presence of this species in the Upper Silesia. J. (1931a, p. 271) confirmed this doubt. Also from the Upper Silesia come specimens that
(1872a, Pl. D.III, 5a, b), classified as Gyroporella minutula. They cannot belong to G. minutula, because the weathered surface shows wide tubercles arranged in dense single-row whorls, a typical feature of Physoporella pauciforata (Physoporella minutula has double-row whorls; see
Fig. 15d-e ![]() herein). Schematic drawings of these specimens were labelled by as Ph. cf. minutula
(1928, fig. 11b; Fig. 16f-g
herein). Schematic drawings of these specimens were labelled by as Ph. cf. minutula
(1928, fig. 11b; Fig. 16f-g ![]() herein).
herein).
A photograph of Physoporella pauciforata () from the borehole Krusin 23WW near Zawiercie was published by Z. (1986, Pl. CVI, 9b), and its assignment to this species was confirmed by & (2000, p. 131). It is reproduced in
Pl. XVIII, 8b ![]() .
.
 |
|
|
Click on thumbnail to enlarge the image.
Figure 16: Previously published illustrations of Physoporella pauciforata () from the Upper Silesia and similarly preserved specimens from the Alps: a-e - pencil drawings by (1872a): a tangential longitudinal fracture of a specimen from Reiflinger Kalk (Reissalpe, Niederösterreich), that can be regarded a lectotype specimen of his new species Gyroporella pauciforata (, 1872a, Pl. D.III, 2b), x 12; b - fractured conspecific specimen from the Upper Silesia (, 1872a, Pl. D.III, 3b), x 15; c - transverse fracture of a similar specimen from the Austrian Alps (, 1872a, Pl. D.III, 2e) that can be regarded a paralectotype, x 10; d-e - Upper Silesian specimens attributed to Gyroporella minutula by , 1872a, Pl. D.III, 5): d - longitudinal fracture along the outer surface, x 15; e - transverse fracture, x 15; f-g - schematic drawings by (1928, fig. 11b, x 6), redrawn from (1872a, Pl. D.III, 5a, b) and reduced (compare f and d, and g and e herein).
Physoporella lotharingica (, 1898)
1898 Diplopora lotharingica n.sp.- , figs. 1-2.
1913 Diplopora lotharingica.- , Pl. I, 1.
1920 Physoporella pauciforata var. ? lotharingica n.stat. n.comb.- , Pl. III, 11, 13.
1920 Diplopora uniserialis.- , Pl. V, 6.
1928 Physoporella var. lothringiaca [sic!].- , fig. 10, excerpt from (1913, Pl. I, 1).
1931a Physoporella lotharingica.- , Pl. XXI, 8.
1935b non Physoporella aff. lotharingica.- in , Pl. II, 2.
1958 Physoporella lotharingica.- , Pl. 7, 17, 19.
1958 non Physoporella lotharingica [= Physoporella leptotheca].- , fig. 12.
1965 Physoporella lotharingica.- , Pl. XIV, 4-6.
1969 (?) Physoporella sp. or Diplopora uniserialis.- in & , Pl. XIX, 7-9.
1974a Physoporella pauciforata subsp. lotharingica.- , p. 25.
1979 pars Physoporella leptotheca.- , Pl. 5, 1-2, 4.
1997 non Physoporella lotharingica.- , Pl. 1, 10-13.
2000 Physoporella lotharingica.- & , p. 126.
Diagnosis: The thallus is cylindrical of very small diameter (1-2 mm). The calcareous wall is rather tiny. Pores (tubercles) are pyriform (closed) and distributed in single whorls. The spaces between whorls are so large, that particular whorls of tubercles are distinctly separated. This species belongs to the Physoporella pauciforata group (one row of pores per one whorl).
Material and localities.- Physoporella lotharingica is abundantly distributed in the Upper Silesia and adjacent territories. It is a common species in many localities of the Upper Silesia (Opole Triassic: Nowe Koszyce, no. 7, Jemielnica, no. 8; Tarnowice Syncline: Segiet, no. 11, Srebrna Góra, no. 13, Łabędy, no. 14; Bytom Syncline: Wojkowice Komorne, no. 29, Boleradź, no. 31, Czeladź, no. 34, 35, 36, Będzin, no. 37) and adjacent regions (Cracow-Chrzanów region - Balin, no. 50, Krupka, no. 52; and Olkusz-Siewierz region: Stare Gliny, no. 57, Brudzowice, no. 70, Nowa Wioska, no. 71, Żelisławice, no. 72). It is also found in few boreholes in the Zawiercie region (10-Ż Lgota Nadwarcie, 14-Ż Żarki).
Nomenclature and discussion; geographical distribution and stratigraphical range.- The species described as Diplopora lotharingica, was erected by E.W. (1898) for material from Middle Muschelkalk of Faulquement (Falkenberg) in Lorraine (Lothringen), near the border of the limestone facies of the Muschelkalk. It is important to note the mode of preservation of the diplopores from the Upper Silesia. The calcareous sheath precipitated by the algae has been dissolved and all that remains is a cylindrical infilling of the internal chamber or an impression of the calcareous sleeve. 's (1898) fig. 1 shows in its left part the outer imprint (a - top), and to the right, the "tentaculite-like" infilling (b - bottom). To present the internal structure of these minute algae (ca. 1 mm diameter), made a composite drawing (ibid., fig. 2) of the two specimens seen in a. In the bottom part of the drawing in each row there is only one whorl of tubercles (branches), and there is only about ten of these. In the upper part of the drawing, the preserved, slightly undulating outer
calcareous sleeve shows swellings with pores, being the outlets for thin branches (b). Such simply built forms were previously known from the Muschelkalk of the Upper Silesia, described as Gyroporella pauciforata and G. minutula . Similar specimens were reported from Weilderstadt in eastern Schwarzwald by
(1913). His illustration (Pl. I, 1) shows single rows of swellings in the outer sheath. Each isolated row consists of 8-10 swellings. 's figure was reproduced by M. (1928, fig. 10), labelled as Physoporella lothringiaca [sic!] - Fig. 17 ![]() .
.
J. (1920) introduced the name Physoporella pauciforata var. lotharingica to describe small (1.7-2 mm) forms from the Eastern Alps (Reifling) and from the vicinity of Sarajevo in Bosnia; these fossils resembled Physoporella pauciforata (), and were very similar to 's specimens from Lorraine. Their calcareous skeleton is poorly developed and forms a thin cover, replicating the contours of soft parts so closely, that a swelling occurs above each branch. The whorls of these swellings, quite widely spaced, are always arranged in one row (, 1920, Pl. III, 11, 13). Probably this species also encompasses the specimen figured in his Pl. V, 6, classified by as Diplopora uniserialis . In his subsequent works (1926, 1930a) consequently used the name Physoporella lotharingica (), regarding it as a separate species. (1931a) received new specimens from the 's locality in Lorraine, and confirmed his description. The spacing between whorls equals approximately half the outer diameter. photographed one of these specimens (, 1931a, Pl. XXI, 8), which in his opinion reveals a steinkern in the bottom part and an imprint of the outer surface in the upper part, as in 's drawing (his fig. 2). Actually, there is an outer view of the tube with visible swellings in the lower part of the photograph, while the upper part shows single rows of pores, representing branch inlets from the tube's lumen. The axial cavity is neither calcified nor sediment-filled. Such a mode of preservation is common among the Upper Silesian specimens.
(1935a, fig.
5g-h) described from Prades near Barcelona (Catalonia, Spain) small forms (about 1 mm in diameter), that he classified as Physoporella aff. lotharingica . However, his description reveals that they show paired rows of tubercles, as in Ph. praealpina . Wide spacing between the rows indicates, they represent the new species Ph. polonoandalusica (Fig. 21c ![]() ).
).
(1935b) attributed to Physoporella lotharingica only the forms from Lorraine, Schwarzwald and Catalonia. He delegated all Alpine and Bosnian material to Ph. pauciforata var. undulata. The main criterion was their different diameter: below 1 mm in German forms, and slightly over 1.5 mm in the Alpine and Bosnian ones.
F. (1958) found in his Third Diplopore Zone of Briançon Triassic (Ladinian) forms similar to Ph. lotharingica () (, 1958, Pl. 7, 17-19), in association with Diplopora uniserialis , and he regarded them as dwarf, atrophic variants of the latter species (Fig. 17g-h ![]() ). According to him, both variants are linked through a series of transitional morphs, but the most abundant are specimens with single whorls of tubercles, separated by wide spaces (, 1958, Pl. 7, 17). Only such forms can be attributed to Ph. lotharingica (). They are identical to many very well preserved Upper Silesian specimens. Very numerous specimens of Ph. lotharingica are present in the collection assembled by the author from the Briançonian Triassic (Col d'Izoard), where they also co-occur with Diplopora uniserialis. Thus, the forms in question cannot be regarded as atrophied variety of D. uniserialis , but as a separate species Ph. lotharingica (). Such a solution was adopted by & (2000, p. 126). Another specimen of (1958, fig. 11), that he believed should be assigned to the latter species or to D. uniserialis, they attribute to Physoporella leptotheca ( & , 2000, p. 125). E.
(1958), who studied diplopores of the Berne Alps, tended to accept 's (1958, fig. 11, p. 194) view that Ph. lotharingica is just a variety and peculiar mode of preservation of D. uniserialis. However, he believed that there may be present real forms belonging to Physoporella lotharingica (). J.-P. (in & , 1969), who determined diplopores from the Maritime Alps, in the Ladinian of the Briançon Series extending up to there, found forms with sharp triangular annuli, which in his opinion are not transitional to Diplopora uniserialis , but instead belong to the genus Physoporella. Perhaps the forms he illustrated (Pl. 1, 7-9) belong to Ph. lotharingica (), as assumed by & (2000, p. 126).
). According to him, both variants are linked through a series of transitional morphs, but the most abundant are specimens with single whorls of tubercles, separated by wide spaces (, 1958, Pl. 7, 17). Only such forms can be attributed to Ph. lotharingica (). They are identical to many very well preserved Upper Silesian specimens. Very numerous specimens of Ph. lotharingica are present in the collection assembled by the author from the Briançonian Triassic (Col d'Izoard), where they also co-occur with Diplopora uniserialis. Thus, the forms in question cannot be regarded as atrophied variety of D. uniserialis , but as a separate species Ph. lotharingica (). Such a solution was adopted by & (2000, p. 126). Another specimen of (1958, fig. 11), that he believed should be assigned to the latter species or to D. uniserialis, they attribute to Physoporella leptotheca ( & , 2000, p. 125). E.
(1958), who studied diplopores of the Berne Alps, tended to accept 's (1958, fig. 11, p. 194) view that Ph. lotharingica is just a variety and peculiar mode of preservation of D. uniserialis. However, he believed that there may be present real forms belonging to Physoporella lotharingica (). J.-P. (in & , 1969), who determined diplopores from the Maritime Alps, in the Ladinian of the Briançon Series extending up to there, found forms with sharp triangular annuli, which in his opinion are not transitional to Diplopora uniserialis , but instead belong to the genus Physoporella. Perhaps the forms he illustrated (Pl. 1, 7-9) belong to Ph. lotharingica (), as assumed by & (2000, p. 126).
E. (1979) described and illustrated numerous specimens of Physoporella leptotheca from an Anisian locality at Monte Poper (Belluno) in the Eastern Dolomites. She stated that particular whorls of pyriform branches are very variable. However, some specimens preserved as internal moulds (, 1979, Pl. 5, 1-2, 4) seem to have fairly stable, wide distances between the whorls of tubercles (Fig. 17i-k ![]() ), and thus resemble the Upper Silesian representatives of Physoporella lotharingica (). On the other hand, the Ladinian Physoporellae from Brenta Dolomites, attributed to Ph. lotharingica by A.
(1997), though minuscule (about 0.5 mm), have densely spaced pore whorls and belong probably to Physoporella pauciforata var. undulata .
), and thus resemble the Upper Silesian representatives of Physoporella lotharingica (). On the other hand, the Ladinian Physoporellae from Brenta Dolomites, attributed to Ph. lotharingica by A.
(1997), though minuscule (about 0.5 mm), have densely spaced pore whorls and belong probably to Physoporella pauciforata var. undulata .
Physoporella lotharingica () in Lorraine and Germany is typical for the Upper Anisian ( & , 1993), as it does in the Upper Silesia where it occurs in the assemblage zone Physoporella pauciforata - Oligoporella pilosa (Pelsonian - , 1986) and within the Diplopora annulatissima zone (Illyrian). In the Dinaride Mountains D. lotharingica () is known from the Anisian and Ladinian (, 1965). In the Briançon Series of the Western Alps it occurs in the Ladinian, together with Diplopora uniserialis (, 1958, in & , 1969), as confirmed also by my unpublished observations.
M. (1967) described a new species from the Chios Island on the Aegean Sea, Physoporella minutuloidea, resembling Ph. minutula () in general appearance and pore shape. It differs from the latter species in having quite widely spaced single whorls. Thus it belongs to the Physoporella pauciforata () group, together with Physoporella lotharingica. The latter species is, however, thinner, with even more distant pores, and shows no prominent undulation. Both species discussed above occur in the Upper Anisian (Illyrian).
 |
|
|
Click on thumbnail to enlarge the image.
Figure 17: Previously published illustrations of Physoporella lotharingica () from Lorraine, Schwarzwald and Alps, similarly preserved as those from the Upper Silesia: a - fragment of a drawing in (1898, fig. 1), with imprints of the outer surface of two minute specimens of his new species Diplopora lotharingica, x 1; b - composite drawing of two specimens from Faulquemont (Falkenberg) in Lorraine (Lotharingien) shown in a, treated as a holotype of Diplopora lotharingica (, 1898, fig. 2), widely spaced single whorls of tubercles are visible, x 10; c - photograph by (1931a, fig. 8) of a specimen from 's type locality in Lotharingien, labelled Physoporella lotharingica; in the lower part this is an outside view of the thallus with swellings, and in the upper part, single rows of pores - branch inlets seen from inside of the central cavity; this mode of preservation is common in Upper Silesia, x 12; d - drawing (enlarged more than twice) by , of a specimen from Schwarzwald (scale bar = 1 cm); e - pencil drawing by (1913, Pl. 1, 1) of a specimen from Schwarzwald, labelled "Diplopora lotharingica" and showing an outer view of a slightly bent alga with widely spaced rows of swellings (scale bar = 1 cm); f - slightly enlarged drawing by (1928, fig. 10) of 's specimen named Physoporella pauciforata var. lothringiaca [sic] (scale bar = 1 cm); g-h - acetic acid-etched specimens from the Vanoise Massif in the French Alps labelled "Physoporella lotharingica" by (1958, Pl. 7, 17, 19), with widely spaced single whorls of tubercles, both x 13; i - pencil drawing by (1920, Pl. V, 6); attributed to Diplopora uniserialis, but considered by (1958) as "Physoporella lotharingica" (); j-l - silicified specimens from M. Popera (Belluno Province, Eastern Dolomites), etched by weathering and showing widely spaced whorls of tubercles, assigned by (1979, Pl. 5, 1-2, 4) to Physoporella leptotheca , j-k x 100, l x 75.
Physoporella praealpina group
Physoporella praealpina , 1920
Pls. XIII ![]() -
XIV
-
XIV ![]() -
XV
-
XV ![]() ,
XVIII
,
XVIII ![]() -
XIX
-
XIX ![]() -
XX
-
XX ![]() -
XXI
-
XXI ![]() &
XXXII
&
XXXII ![]() -
XXXIII
-
XXXIII ![]() -
XXXIV
-
XXXIV ![]() -
XXXV
-
XXXV ![]() -
XXXVI
-
XXXVI ![]() -
XXXVII
-
XXXVII ![]() -
XXXVIII
-
XXXVIII ![]() ;
Fig. 18
;
Fig. 18 ![]()
1870 Cylindrum annulatum (Nullipora annulata ).- , Pl. 11, 3, 4.
1872a Gyroporella pauciforata.- , Pl. D.III, 2a.
1872a Gyroporella macrostoma.- , Pl. D.II, 4a, 4b.
1920 Physoporella praealpina n.sp.- , Pl. III, 1-9.
1928 Diplopora annulata.- , fig. 7.
1935a Physoporella aff. praealpina.- , figs. 37-39.
1958 Physoporella praealpina.- , Pl. 6, 12-14, 16; Pl. 22, 2.
1960 Diplopora annulata forma physoporelloidea, n.form.- & , fig. 6; Pl. IV, 15 [= fig. 6].
1960 Diplopora sp.- & , fig. 8; Pl. II, 12.
1961 Physoporella praealpina.- , fig. 6; Pl. IV, 1-3, 4 pars.
1964 Physoporella cf. praealpina.- , Pls. XIX, 1-9; Pl. XX, 1-3.
1965 Physoporella cf. praealpina.- , Pl. 12, 5.
1965 Physoporella praealpina.- & , Pls. III, 1-2, 4, 6; Pl. IV, 1-2, 5-6.
1966 Physoporella praealpina.- , Pl. VII, 2a, 3-4.
1967 Physoporella praealpina.- , p. 282.
1969 Physoporella praealpina.- in , figs. 14-15.
1970 Physoporella praealpina.- , Pl. V, 4-5.
1972c Physoporella praealpina.- , fig. 1.1-21.
1973 Physoporella praealpina.- & , Pls. VI, 1-12; Pl. VII, 1-17.
1975 Physoporella praealpina.- & , Pl. IV, 21-23.
1979 Physoporella praealpina.- , Pl. 79, 1.a-b, 2.a-b, 2.f, 3, 10.a-c, 16, excerpt from & (1960: Pl. IV, 15 = fig. 6).
1981 Physoporella praealpina.- , p. 75, 78.
1982 Physoporella praealpina.- , Pl. II, fig. 4.
1986 Physoporella praealpina.- , Pl. CIII, 1.a-b, 2.a-b, 2.f, 3, 10.a-c; Pl. CIII, 1b, excerpt from (1979; Pl. 79, 1.a-b, 2.a-b, 2.f, 3, 10.a-c; Pl. CIII, 16, excerpt from & (1960: Pl. IV, 15 = fig. 6); Pl. CV, 14-16; Pl. CVI, 1-2, 4-5, 7, 9.a.
2000 Physoporella praealpina.- & , p. 134.
Material.- Ph. praealpina is the most frequent and widely distributed species. It is common in the Diplopora Dolomite in the Upper Silesia and adjacent regions (see Fig. 2 ![]() with localities and Fig. 5
with localities and Fig. 5 ![]() ). The species is relatively infrequent in the Opole Triassic (Suchodaniec no. 6) and in the Tarnowice Syncline (Segiet no. 11). It does occur very frequently in the Bytom Syncline, where it is present in almost all the localities. The most numerous and best preserved specimens come from Przełajka (no. 25), Wojkowice-Komorne (no. 27-30), Boleradź (no. 31), and Czeladź (no. 34, 35); see Pl. XIII, 7-8
). The species is relatively infrequent in the Opole Triassic (Suchodaniec no. 6) and in the Tarnowice Syncline (Segiet no. 11). It does occur very frequently in the Bytom Syncline, where it is present in almost all the localities. The most numerous and best preserved specimens come from Przełajka (no. 25), Wojkowice-Komorne (no. 27-30), Boleradź (no. 31), and Czeladź (no. 34, 35); see Pl. XIII, 7-8 ![]() . The species is also abundant in Cracow-Chrzanów area, (Krasowy no. 41, Granice no. 43, Imielin no. 44, Libiąż no. 45, Rosowa Góra no. 48, and Balin no. 50, see Pl. XVIII, 8a
. The species is also abundant in Cracow-Chrzanów area, (Krasowy no. 41, Granice no. 43, Imielin no. 44, Libiąż no. 45, Rosowa Góra no. 48, and Balin no. 50, see Pl. XVIII, 8a ![]() ), as well as in the Olkusz-Siewierz Monocline (Krążek no. 56, Klucze no. 57, Łosień no. 62, Warpie no. 66). It is the most abundant, however, in the Siewierz region (Brudzowice no. 70, Nowa Wioska no. 71, Żelisławice no. 72, near Mrzygłód, Winowno, Myszków, and Żarki. The best preserved specimens are shown in Pl. XVIII, 1-3
), as well as in the Olkusz-Siewierz Monocline (Krążek no. 56, Klucze no. 57, Łosień no. 62, Warpie no. 66). It is the most abundant, however, in the Siewierz region (Brudzowice no. 70, Nowa Wioska no. 71, Żelisławice no. 72, near Mrzygłód, Winowno, Myszków, and Żarki. The best preserved specimens are shown in Pl. XVIII, 1-3 ![]() (e.g., from boreholes 25-WW Winowno and 23-WW Krusin) Physoporella praealpina is difficult to detect in thin sections (Z. 's collection MuzPIG no. 1682.II), where only its "ghosts" are visible (see Pl. XXI, 1-17
(e.g., from boreholes 25-WW Winowno and 23-WW Krusin) Physoporella praealpina is difficult to detect in thin sections (Z. 's collection MuzPIG no. 1682.II), where only its "ghosts" are visible (see Pl. XXI, 1-17 ![]() ).
).
Diagnosis.- This species is characterized by a cylindrical, generally undivided thallus and by alternating vertical branches (pores, tubercles), forming two rows belonging to one whorl. Whorls are closely spaced and tubercles are very regularly arranged. If calcareous (or dolomitic) wall is preserved, initial undulation or fissuration can be observed. The shape of pores (tubercles) is pyriform. Generally, they are perpendicular to the main axis, but in some cases they are slightly oblique to the wall.
Description.- Detailed description of Ph. praealpina is given in explanation of
Pls. XIII ![]() -
XIV
-
XIV ![]() -
XV
-
XV ![]() , where the photographs of fractures are presented. Many specimens are preserved three-dimensionally, permitting to study many details, not observable in thin sections.
, where the photographs of fractures are presented. Many specimens are preserved three-dimensionally, permitting to study many details, not observable in thin sections.
Stratigraphical range and geographical distribution.- Physoporella praealpina is a typical West Alpine species, described by J. (1920) from the material collected by F. from the Préalpes Medianes (, 1913). The name "praealpina" had been proposed by (fide , 1920, p. 55). This species is also typical for the Briançonian Series, particularly in the Vanoise Massif (, 1958). In these two places, this species is considered as the index fossil for Upper Anisian, occurring here together with Diplopora annulatissima , an index Illyrian fossil. Ph. praealpina appears here with Ph. dissita () and Ph. minutula () of the same group, but without the Ph. pauciforata group. Physoporella aff. praealpina together with Ph. pauciforata were described by (1935a) from Bosnia, by (1965) from Dalmatia, by (1970) from Eastern Serbia, by (1972c) from the Austrian Alps and by (1964) from Southern Slovakia (Western Carpathians). In all these descriptions Ph. praealpina is considered as index fossil of Pelsonian, but it reaches up to the Illyrian, where it appears together with Diplopora annulatissima . Bulgarian algae ( & , 1973) are Pelsonian in age, as D. annulatissima is absent.
In the Tatra Mts (High-Tatric series, Polish and Slovakian Western Carpathians) Physoporella praealpina , determined by & (1965) and by (in & , 1965; , 1965), appears together with Diplopora annulatissima (, 1967, 1979, 1986).
Physoporella praealpina must have been found in the Upper Silesia by the pioneer German researchers, as it is the most common species among the Dasycladales. The specimen illustrated by F. (1870, Pl. 11, 3, 4), and labelled as Cylindrum annulatum (Nullipora anullata ) does not belong to Diplopora annulata, but to Physoporella praealpina (fig. 7a-d). J. (1931a, p. 271) believed, that the field geologists understood Cylindrum annulatum as equivalent to the species Diplopora annulata in later usage, though it is a collective name and it is difficult to establish which species was meant. This may be demonstrated by the fact that M. (1928, fig. 7) reproduces 's illustration and uses the name Diplopora annulata . to label it, misquoting it after . Perhaps it was because of that later geologists used the name Diplopora annulata (.) for all large specimens of Dasycladales they found in the field.
But the form illustrated by (Fig. 18a-d ![]() ) and (Fig. 18e-f
) and (Fig. 18e-f ![]() ) has all features of the Physoporella praealpina (Fig. 18j
) has all features of the Physoporella praealpina (Fig. 18j ![]() ). It is a simple non-segmented tube, covered with pyriform tubercles arranged in double whorls. Tubercles alternate, and particular double rows lay fairly close to each other, separated only with narrow spaces.
). It is a simple non-segmented tube, covered with pyriform tubercles arranged in double whorls. Tubercles alternate, and particular double rows lay fairly close to each other, separated only with narrow spaces.
Described specimen has been found by in the Segiet Forest, south of Tarnowskie Góry (locality no. 11), that yielded also numerous well preserved specimens of Physoporella praealpina and Diplopora annulatissima .
Undoubtedly, Physoporella praealpina is represented by a specimen from the Austrian Alps, illustrated by
(1872a, Pl. D.III, 2a) and attributed by him to then more widely interpreted species Physoporella pauciforata (Fig. 18g ![]() ). His illustration shows large tubercles in double whorls, separated by small spaces with fissuration lists. Characters typical for Ph. praealpina are also visible in the specimen from Mendola Dolomite in Italian Alps, classified by
(1872a, Pl. D.II, 4a, b), to Gyroporella macrostoma (Fig. 18h-i
). His illustration shows large tubercles in double whorls, separated by small spaces with fissuration lists. Characters typical for Ph. praealpina are also visible in the specimen from Mendola Dolomite in Italian Alps, classified by
(1872a, Pl. D.II, 4a, b), to Gyroporella macrostoma (Fig. 18h-i ![]() ).
).
The presumption that large Dasycladales from the Upper Silesia and adjacent regions belong to Diplopora annulata (.) is also evident in the paper by C. and S. (1960). They determined a large specimen (2.8 mm in diameter) found in Żelisławice (locality no. 72) as Diplopora annulata . f. physoporelloidea n.f. (op. cit., Pl. IV, 15; text-figs. 6, 10). In their description, they state, that pyriform tubercles are arranged in double whorls, separated by non-tuberculate bands. They do not present any evidence of tubercles forming tufts, being a major diagnostic character of Diplopora annulata sensu lato (, 1920 - metaspondyl arrangement of pores or tubercles). The photograph does not show evidence for annulate morphology of the specimen. Tubercles are pyriform, unusual for the genus Diplopora. The described specimen has no characters typical for Diplopora, and all diagnostic characters of Physoporella. The authors must have noticed that, when establishing a new form, and naming it physoporelloidea. Actually, all the above-mentioned characters indicate that their specimen belongs to Physoporella praealpina , as noted by Z. , 1967 (also in & , 1973). Illustrated with reproduced photograph by & (op. cit., Pl. IV, 15 = fig. 6), the specimen has been relabelled as Physoporella praealpina (, 1979, Pl. 79, 16; , 1986, Pl. CIII, 16).
Also the specimen regarded by & (1960, Pl. II, 12, text-fig. 8) as Diplopora sp. undoubtedly belongs to Physoporella praealpina . It is a Ph. praealpina with condensed whorls (Fig. 18n ![]() ).
).
F. (1958), determining Middle Triassic Dasycladales from the Vanoise Massif
(Briançonian Series of the French Alps), etched the specimens with acetic acid, thus obtaining a three-dimensional appearance (Fig. 18o-r ![]() ), similar to naturally occurring preservation in Upper Silesian fractures. The chemical etching has been also successfully applied to study of Dasycladales from Berne Alps in Switzerland (,
1961, text-fig. 6; Fig. 18k
), similar to naturally occurring preservation in Upper Silesian fractures. The chemical etching has been also successfully applied to study of Dasycladales from Berne Alps in Switzerland (,
1961, text-fig. 6; Fig. 18k ![]() herein), where the Briançon Series from French Alps continues in the Pre-Alpine nappes (, 1958).
herein), where the Briançon Series from French Alps continues in the Pre-Alpine nappes (, 1958).
 |
|
|
Click on thumbnail to enlarge the image.
Figure 18: Previously published illustrations of Physoporella praealpina from the Upper Silesia and similarly preserved specimens from the Alps: a-d - pencil drawings by (1870) of specimens from Segiet Forest near Tarnowskie Góry, assigned by him to Cylindrum annulatum (Nullipora annulata ): a - enlarged view of a fracture with double whorls of alternating fairly large tubercles separated by small gaps (not forming annuli, as explained); b - enlarged upper view upon a whorl; the elongated branches do not belong to the latter specimen, but to Diplopora annulatissima (compare Fig. 13b); c-d - the same specimens as in a and b, but reproduced almost in the same scale as 's original figures; e-f - reproduction of drawings by (1928, fig. 7), who attributed the material to Diplopora annulata ., x 7.5; g - pencil drawing by (1872a, Pl. D.III, 2a) of a specimen from Austrian Alps, assigned by him to Gyroporella pauciforata, even though there are visible whorls of double rows of alternating tubercles with pronounced fissuration (typical for Ph. praealpina); h-i - pencil drawings by (1872a, Pl. D.III, 4a, 4b) of a specimen from Italian Alps, assigned by him to Gyroporella macrostoma: h - actual size; i - the same specimen enlarged x 8, with whorls of double rows of alternating tubercles separated with fissures; j - pencil drawing by (1920, fig. 5) of a polished section of a syntype of Physoporella praealpina from Horboden in Berne Alps; k - photograph by of an acetic acid-etched specimen of Physoporella praealpina (, 1961: text-fig. 6), x 25; l-m - illustrations of a specimen from Żelisławice near Siewierz (locality no. 72), attributed by & (1960) to Diplopora annulata . f. physoporelloidea: l - drawing, x 16; m - reduced photograph (x 11) from & (1960, Pl. IV, 15; originally x 22), according to (1967), & (1973), (1979, Pl. 79, 16), and (1986, Pl. CIII, 16) this is a typical Physoporella praealpina ; n - photograph of a specimen from Kąty Chrzanowskie (Rosowa Góra - locality no. 48) labelled as Diplopora sp. by & (1960, Pl. II, 12); this is a Physoporella praealpina with dense double whorls of tubercles; o-r - drawings of specimens from Berne Alps assigned to Physoporella praealpina by (1958, Pl. 6, 12-14); etching with acetic acid revealed their three-dimensional structure, similar to that on fractures of the specimens from the Upper Silesia.
Physoporella dissita (, 1872a)
Pls. XVI ![]() ,
XX
,
XX ![]() -
XXI
-
XXI ![]() ,
XXXIV
,
XXXIV ![]() -
XXXV
-
XXXV ![]() &
XXXVII
&
XXXVII ![]() -
XXXVIII
-
XXXVIII ![]() ;
Fig. 19
;
Fig. 19 ![]()
1872a Gyroporella dissita n.sp.- , Pl. D.III, 1.
1912 Physoporella dissita n.comb.- , Pl. VI (V), 1-4.
1958 Physoporella dissita.- , Pl. 6, 17.
1964 Physoporella dissita.- , Pl. XX, 6; Pl. XXI, 1-7.
1965 Physoporella cf. dissita.- & , Pl. 4, 3.
1966 Physoporella dissita.- , Pl. VI, 5; Pl. VII, 1-2b.
1967 Physoporella dissita.- , Pls. I, 5; Pl. II, 2-4.
1967 Physoporella dissita.- , Pl. 2, 2-4; Pl. 3, 2-3.
1973 Physoporella dissita.- & , Pl. VIII, 1-23.
1979 Physoporella dissita.- , Pl. 79, 2. c, 5-6.
1981 Physoporella dissita.- , p. 75, 78.
1983 Physoporella dissita.- , Pl. III, 19.
1986 Physoporella dissita.- , Pl. III, 1, from (1966, Pl. VII, 1); Pl. III, 2.
1986 Physoporella dissita.- , Pl. CIII, 2. c, 5-6, from (1979: Pl. 79, fig. 2. c, 5-6); Pl. CV, 17-19.
1994 Physoporella dissita.- et al., Pl. 11, fig. 3.
1997 Physoporella lotharingica.- , Pl. I, fig. 10-13.
2000 Physoporella dissita.- & , p. 122-123.
2013 Physoporella dissita.- in & , Pl. VI (V), 1-4.
Diagnosis.- Markedly segmented or even annulated forms. Each segment is connected with one whorl, composed of two rows with alternating pores. This segmentation is very well developed and is an indispensable feature of this species. The tubercles are pyriform, but not so remarkable pear-shaped, as in the Ph. praealpina. Nevertheless, the pores never, or only very rarely, transect the wall of the cylinder, which is decisive in assigning these forms to the genus Physoporella (, 1967, 1969). Dimensions are comparable to those of Ph. praealpina ( & , 1973). If the wall is not preserved, the only feature permitting to distinguish these two species is the distances between whorls of double-row tubercles, which are distinctly larger in Ph. dissita. This feature, noted by (1958), is clearly observable in our material. The largest distances between the double rows are a characteristic feature of Ph. polonoandalusica.
Material and localities.- Physoporella dissita is distributed in all regions of the Upper Silesia and adjacent areas (see Fig. 2 ![]() ), but is rather infrequent and difficult to determine. In the Opole Triassic it was found in Nowe Koszyce, no. 7, and Jemielnica, no. 8. In the Tarnowice Syncline it is present in Segiet, no. 11, and Srebrna Góra, no. 13. Its occurrences are more abundant in the Bytom Syncline: Przełajka, no. 25, Wojkowice Komorne, nos. 27, 28, and particularly Czeladź, nos. 34, 35. This species is also present in the Cracow-Chrzanów area: Granice, no. 43, Rosowa Góra, no. 48. It appears more frequently in the Olkusz-Siewierz Monocline (Łosień, no. 62, Trzebiesławice, no. 53, Siewierz, no. 68, and particularly Brudzowice, no. 70, and Nowa Wioska, no. 71. In the deep borings of the Zawiercie region, it was found in boreholes 51-Ż Myszków and 10 Ż Lgota Nadwarcie, where it was determined from relatively good thin sections (,
1986, Pl. CV, 17-19). Attribution of the above specimens figured by to Ph. dissita has been accepted by & (2000, p. 122-123). Ph. dissita was also found in many thin-sections (see Pl. XXI
), but is rather infrequent and difficult to determine. In the Opole Triassic it was found in Nowe Koszyce, no. 7, and Jemielnica, no. 8. In the Tarnowice Syncline it is present in Segiet, no. 11, and Srebrna Góra, no. 13. Its occurrences are more abundant in the Bytom Syncline: Przełajka, no. 25, Wojkowice Komorne, nos. 27, 28, and particularly Czeladź, nos. 34, 35. This species is also present in the Cracow-Chrzanów area: Granice, no. 43, Rosowa Góra, no. 48. It appears more frequently in the Olkusz-Siewierz Monocline (Łosień, no. 62, Trzebiesławice, no. 53, Siewierz, no. 68, and particularly Brudzowice, no. 70, and Nowa Wioska, no. 71. In the deep borings of the Zawiercie region, it was found in boreholes 51-Ż Myszków and 10 Ż Lgota Nadwarcie, where it was determined from relatively good thin sections (,
1986, Pl. CV, 17-19). Attribution of the above specimens figured by to Ph. dissita has been accepted by & (2000, p. 122-123). Ph. dissita was also found in many thin-sections (see Pl. XXI ![]() ), but it is hardly determinable, because of its poor state of preservation - as "ghosts", with few recognizable features.
), but it is hardly determinable, because of its poor state of preservation - as "ghosts", with few recognizable features.
Description.- In many specimens shown in Pl. XVI ![]() there are observable features allowing to their attribution to Physoporella dissita.
Pl. XVI, 1, 4-8, 13, 17, 20-23, 32
there are observable features allowing to their attribution to Physoporella dissita.
Pl. XVI, 1, 4-8, 13, 17, 20-23, 32 ![]() show pronounced segmentation. In many other cases, when the outer wall has not been preserved, the segmentation is indicated by fairly large distances between the double rows of alternating tubercles
(Pl. XVI, 1, 10-12, 16, 32
show pronounced segmentation. In many other cases, when the outer wall has not been preserved, the segmentation is indicated by fairly large distances between the double rows of alternating tubercles
(Pl. XVI, 1, 10-12, 16, 32 ![]() ) and by undulating surface of surrounding rock. Some specimens exhibit intusannulation
(Pl. XVI, 6, 32
) and by undulating surface of surrounding rock. Some specimens exhibit intusannulation
(Pl. XVI, 6, 32 ![]() ), while the ridges separating furrows with pores are wide and flat
(Pl. XVI, 21a, 32
), while the ridges separating furrows with pores are wide and flat
(Pl. XVI, 21a, 32 ![]() ). Pyriform shape of branches (tubercles) is well visible. In transverse fractures, the number of tubercles ranges from 23
(Pl. XVI, 24
). Pyriform shape of branches (tubercles) is well visible. In transverse fractures, the number of tubercles ranges from 23
(Pl. XVI, 24 ![]() ) to 25 (Pl. XVI, 19
) to 25 (Pl. XVI, 19 ![]() ). The axial cavity is usually hollow
(Pl. XVI, 3, 5-7, 21b, 23-24, 31
). The axial cavity is usually hollow
(Pl. XVI, 3, 5-7, 21b, 23-24, 31 ![]() ), rarely filled with sediment completely
(Pl. XVI, 19, 26
), rarely filled with sediment completely
(Pl. XVI, 19, 26 ![]() ). In some specimens, two walls - outer and inner one - are visible, connected by branches
(Pl. XVI, 1, 17, 21a, 23-24, 26, 31-32
). In some specimens, two walls - outer and inner one - are visible, connected by branches
(Pl. XVI, 1, 17, 21a, 23-24, 26, 31-32 ![]() ); both walls are undulating and the undulation coincides with intusannulation.
); both walls are undulating and the undulation coincides with intusannulation.
Stratigraphical range and geographical distribution.- Physoporella dissita is a typical West Alpine species (, 1912, 1920, in & , 2013) and is characteristic for the Prealpian Upper Anisian (Illyrian) because of its appearance together with Diplopora annulatissima. It occupies the same stratigraphical position in the Briançon Series, where it occurs together with Ph. praealpina, Ph. minutula and Diplopora annulatissima (, 1958; , 1960). Of the same age is Ph. dissita in the High-Tatric series, where it occurs together with Ph. praealpina, Ph. minutula, Ph. pauciforata and Diplopora annulatissima (, 1967, 1979, 1986). Ph. dissita from Slovakian Karst is of Pelsonian age (, 1957, 1964, 1966). Pelsonian age is also ascribed to this species in Bulgaria, where it occurs together with other species of genus Physoporella, but without Diplopora annulatissima ( & , 1973).
In the the Upper Silesia and adjacent regions, Physoporella dissita is of Pelsonian and Illyrian age.
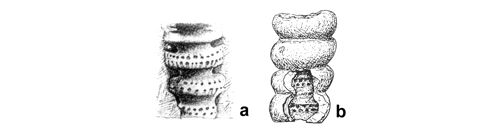 |
|
|
Click on thumbnail to enlarge the image.
Figure 19: Previously published illustrations of Physoporella dissita () from the Alps, similarly preserved as new specimens from the Upper Silesia: a - drawing of the holotype from Wettersteinkalk, Zugspitze Group, Eastern Alps, fracture with pronounced undulation and widely spaced double whorls (, 1872a, Pl. D.III, 1), x 10; b - drawing of a specimen from the Middle Triassic of the Vanoise Massif in French Alps, etched with acetic acid (, 1958, Pl. 6, 17), x 10, attributed by to Physoporella praealpina , a form transitional to Physoporella minutula (). Widely spaced double whorls and pronounced deep undulation indicate that it belongs to Physoporella dissita ().
Physoporella minutula (, 1872a) , 1912
1872a Gyroporella minutula n.sp.- , Pl. DIII, 4a.
1872a non Gyroporella minutula n.sp.- , Pl. DIII, 5a, 5b.
1912 Physoporella minutula n.comb.- , Pl. VI (V), 5-10, 12.
1920 Physoporella minutula.- , Pl. II, 19-22.
1928 Gyroporella cf. minutula.- , fig. 11a, c.
1928 non Gyroporella cf. minutula.- , fig. 11 b.
1958 Physoporella minutula.- , Pl. 6, 18-19.
1958 Clavaphysoporella minutula n.gen, n.comb.- , p. 260, without figure.
1961 Physoporella minutula.- , Pl. VI, 4 pars.
1964 Physoporella minutula.- , Pl. XX, 4; Pl. XXI, 10.
1965 Physoporella cf. minutula.- & , Pl. III, 5.
1969 Physoporella minutula.- & , Pl. IV, 7-8.
1970 Physoporella minutula.- , Pls. I, 9, II, 3-5; Pl. V, 6.
1973 Physoporella minutula.- & , Pl. IX, 1-35.
1973 Physoporella minutula.- & , Pl. VI, 21 pars; Pl. VII, 28; Pl. VIII, 30 pars; Pl. X, 38-39 pars; Pl. XIV, 55 pars; Pl. XV, 57 pars; Pl. XVI, 63 pars.
1975 Physoporella minutula.- & , Pl. IV, 25; Pl. VI pars.
1979 Clavaphysoporella minutula.- , p. 633, without figure.
1979 Physoporella minutula.- , Pl. 79, 11-12.
1981 Physoporella minutula.- , p. 75, 78.
1986 Physoporella minutula.- , Pl. CIII, 11-12 (from , 1979, Pl. 79, 11-12); Pl. CV, 20-21.
2000 Physoporella minutula.- & , p. 126-127.
2013 Physoporella minutula.- in & , Pl. VI (V), 5-10, 12.
Description.- Most frequently algae with divided (undulated or even annulated) cylinder. Segments, if they exist, are sharper that in Ph. dissita and have a very characteristic shape (comp. 's holotype - Fig. 20a ![]() ). This feature is so characteristic, that it is frequently used for species determination even when pores are not visible. The two rows of pores in one whorl are very well marked and the shape of pores, although still pyriform, is as a rule elongated, having rather a "sausage-like" character. Nevertheless, in some cases it is typically pyriform. The contour of the inner tube of cylinder is straight, regular and very markedly separated from the wall. Dimensions are comparable with Ph. praealpina, or smaller.
). This feature is so characteristic, that it is frequently used for species determination even when pores are not visible. The two rows of pores in one whorl are very well marked and the shape of pores, although still pyriform, is as a rule elongated, having rather a "sausage-like" character. Nevertheless, in some cases it is typically pyriform. The contour of the inner tube of cylinder is straight, regular and very markedly separated from the wall. Dimensions are comparable with Ph. praealpina, or smaller.
Material and discussion.- The 's holotype (Gyroporella minutula) was described from the Reifling Limestone at Reissalpe in the Northern Calcareous Alps (,
1872a, Pl. D.III, 4a - see Fig. 20a ![]() ). He remarked that this species is probably present in Jemielnica (Himmelwitz) in the Upper Silesia (,
1872a, Pl. D.III, 5a 5b). M. (1928, fig. 11c) reproduced schematically 's holotype under the name Gyroporella cf. minutula, assigning him erroneously the Upper Silesian origin (actually, it is of Alpine provenance). On the other hand, & considered the specimen from Jemielnica as belonging to Gyroporella minutula. However, this specimen shows only one row of tubercles in one whorl, and not two rows, as it is typical for Physoporella minutula. This is why the specimen from Jemielnica is here attributed to Ph. pauciforata (Fig. 16d-e
). He remarked that this species is probably present in Jemielnica (Himmelwitz) in the Upper Silesia (,
1872a, Pl. D.III, 5a 5b). M. (1928, fig. 11c) reproduced schematically 's holotype under the name Gyroporella cf. minutula, assigning him erroneously the Upper Silesian origin (actually, it is of Alpine provenance). On the other hand, & considered the specimen from Jemielnica as belonging to Gyroporella minutula. However, this specimen shows only one row of tubercles in one whorl, and not two rows, as it is typical for Physoporella minutula. This is why the specimen from Jemielnica is here attributed to Ph. pauciforata (Fig. 16d-e ![]() ).
(1926a) did not find this species in Jemielnica, but found some similar species (Diplopora cf. minutula) in the Drama Valley (Karchowice Beds). (1931a) considered 's descriptions and illustration, as well as 's description, as insufficient and was of opinion that the specimen from Jemielnica belongs instead to Oligoporella elegans. Physoporella minutula is probably frequent in the Upper Silesia and adjacent regions, but the state of their preservation (lack of cylinder and segments with only the internal cast and tubercles preserved) is not favourable for their recognition and precise determination. The example of determinable specimen with cylinder and pores preserved is described and illustrated from borehole Winowno 25WW (, 1986, Pl. CV,
20-21). It is confirmed by & (2000).
).
(1926a) did not find this species in Jemielnica, but found some similar species (Diplopora cf. minutula) in the Drama Valley (Karchowice Beds). (1931a) considered 's descriptions and illustration, as well as 's description, as insufficient and was of opinion that the specimen from Jemielnica belongs instead to Oligoporella elegans. Physoporella minutula is probably frequent in the Upper Silesia and adjacent regions, but the state of their preservation (lack of cylinder and segments with only the internal cast and tubercles preserved) is not favourable for their recognition and precise determination. The example of determinable specimen with cylinder and pores preserved is described and illustrated from borehole Winowno 25WW (, 1986, Pl. CV,
20-21). It is confirmed by & (2000).
Stratigraphical range and geographical distribution.- Physoporella minutula has in general a rather similar stratigraphical range and distribution as Ph. dissita and Ph. praealpina. It is a typical species of Western Alps (Préalpes Medianes - , 1912, 1920, in & , 2013; , 1958; , 1961), where it was first found by F. . Its age here is Upper Anisian (Illyrian), as it appears together with Diplopora annulatissima. The same applies to this species in the Briançon Series (, 1958) and in the High-Tatric Series in the Tatra Mts. (, 1967, 1979, 1986). D. minutula is also known in Slovakia (, 1964), Romania ( & , 1969; , 1970; & , 1973; & , 1975) and Bulgaria ( & , 1973); in the last locality it is of Pelsonian age. Generally speaking, its age coincides with Physoporella-Oligoporella assemblage zone and Diplopora annulatissima zone.
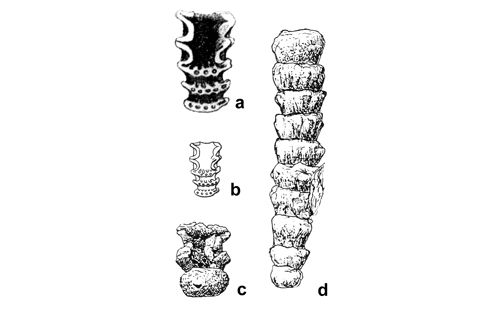 |
|
|
Click on thumbnail to enlarge the image.
Figure 20: Selected previously published illustrations of Physoporella minutula () from the Alps, similarly preserved as new specimens from the Upper Silesia: a - pencil drawing of a specimen from Reiflinger Kalk (Reissalpe, Austrian Alps), the holotype of Gyroporella minutula (, 1872a, Pl. D.III, 4a), fractured specimen with typical undulation and widely spaced double whorls of pores, x 12; b - schematic drawing of 's holotype by (1928, fig. 11b), reduced (x 6) and labelled Gyroporella cf. minutula; c-d - drawings of specimens from Vanoise Massif in French Alps, etched with acetic acid (, 1958, Pl. 6, 18-19), x 8, attributed by to Physoporella minutula. Characteristic outer appearance of the alga is visible, with asymmetric whorls (d), and widely spaced double whorls of pores.
Physoporella polonoandalusica n.sp.
1906 ? Diplopora rauffi.- , Pl. 3, 1; p. 81.
1920 Diplopora annulata.- , p. 28.
1926a ? Diplopora rauffi.- , p. 505.
1931a Diplopora annulata.- , p. 273, 275.
1935a Physoporella aff. lotharingica.- , Pl. II, 2.
1952 Physoporella aff. lotharingica.- , p. 2-3.
1954 Physoporella lotharingica.- et al., p. 58.
1958 Physoporella praealpina.- , Pl. 6, fig. 15.
1979 pars Physoporella leptotheca.- , Pl. 5, fig. 3.
2000 ? Physoporella aff. lotharingica.- & , p. 126.
Holotype.- Pl. XVII, 6 ![]()
Paratypes.- Pl. XVII, 1, 5 ![]() .
.
Localities.- Przełajka, no. 25 (the type locality), Granice, no. 43, Klucze (Stare Gliny), no. 53.
Repository.- Polish Geological Institute, Warsaw, Geological Institute, coll. of Z. no. MuzPIG 1653.II.
Derivation of the name.- After a palaeogeographical province in the Middle Triassic, stretching from Poland to Andalusia.
Diagnosis.- Cylindrical thallus of very small diameter (1-2 mm), very long and frequently bended. The calcareous wall is rather thin and slightly undulated. Pores (tubercles) are pyriform and are distributed in double whorls, very widely spaced. This species belongs to the Physoporella praealpina group (two rows of alternate pores (tubercles) per one whorl).
Material.- Multiple specimens in all described regions of the Upper Silesia (Tarnowice Syncline, Bytom Syncline) and adjacent regions (Cracow-Chrzanów region, Olkusz-Siewierz regions and deep boreholes from Zawiercie region). For distribution in particular localities, see Fig. 5 ![]() )
)
Description.- Extensive description of specimens present on fractures is given in explanations to Pl. XVII ![]() . The specimen from Przełajka (locality no. 25) shown in Pl. XVII, 6
. The specimen from Przełajka (locality no. 25) shown in Pl. XVII, 6 ![]() is proposed as the holotype. The slightly bent thallus is flat, irregularly undulated. Pyrifom tubercles (branches) are arranged in double rows, separated by large surfaces without tubercles. The distance between double rows is distinctly larger than in Physoporella dissita (). Similar features are present in the specimen shown in Pl. XVII, 1
is proposed as the holotype. The slightly bent thallus is flat, irregularly undulated. Pyrifom tubercles (branches) are arranged in double rows, separated by large surfaces without tubercles. The distance between double rows is distinctly larger than in Physoporella dissita (). Similar features are present in the specimen shown in Pl. XVII, 1 ![]() , also from Przełajka, selected as the paratype. Another paratype (Pl. XVII, 4
, also from Przełajka, selected as the paratype. Another paratype (Pl. XVII, 4 ![]() ) comes from Czeladź, locality no. 34. The third paratype
(Pl. XVII, 10
) comes from Czeladź, locality no. 34. The third paratype
(Pl. XVII, 10 ![]() ), from Klucze (Stare Gliny, no. 57) is much larger and shows no distinct undulation. The fourth paratype
(Pl. XVII, 16
), from Klucze (Stare Gliny, no. 57) is much larger and shows no distinct undulation. The fourth paratype
(Pl. XVII, 16 ![]() ), again from the type locality Przełajka, is very similar to the holotype
(Pl. XVII, 6
), again from the type locality Przełajka, is very similar to the holotype
(Pl. XVII, 6 ![]() ), with slight undulation of the outer wall and more double alternating tubercles. In Pl. XVII, 8
), with slight undulation of the outer wall and more double alternating tubercles. In Pl. XVII, 8 ![]() , two walls are visible: undulating outer one, and flat inner one. The specimen shown in Pl. XVII, 11
, two walls are visible: undulating outer one, and flat inner one. The specimen shown in Pl. XVII, 11 ![]() is very thin, similar to Physoporella lotharingica (), but has double rows of alternating tubercles.
is very thin, similar to Physoporella lotharingica (), but has double rows of alternating tubercles.
Nomenclature and geographical distribution.- The new species described above is very abundant in the Diplopora Dolomite of the Upper Silesia and neighbouring regions. Probably the species comprises also the insufficiently described and inadequately illustrated Diplopora rauffi , from Granice (,
1906, Pl. 3; Fig. 21a ![]() herein). It has been listed under the same name by
(1926a, p. 505), who noted that this species differs from Diplopora annulata () in its diameter to annulus height ratio being 3:1, not 5.5:1. (1928, fig. 8a) reproduced the illustration and included his species into Diplopora annulata var. septemtrionalis. According to his description, the branches are filamentous, thickening distally (vesiculiferous?), forming indistinct tufts. The description does not fit the figure by (Fig. 21a
herein). It has been listed under the same name by
(1926a, p. 505), who noted that this species differs from Diplopora annulata () in its diameter to annulus height ratio being 3:1, not 5.5:1. (1928, fig. 8a) reproduced the illustration and included his species into Diplopora annulata var. septemtrionalis. According to his description, the branches are filamentous, thickening distally (vesiculiferous?), forming indistinct tufts. The description does not fit the figure by (Fig. 21a ![]() ) nor its reproduction by (Fig. 21b
) nor its reproduction by (Fig. 21b ![]() ), and seems to be biased to make it fit the results of taphonomical alterations of the genus Diplopora.
(1920, p. 28; 1931a, p. 273, 275) included D. rauffi to D. annulata, but without explicit justification; he merely noted that both species differ in some characters. Indeed, the figure in there are no characters diagnostic for Diplopora - no annuli, no tufts of branches, and there are only rarely spaced whorls of tubercles of undetermined shape. It seems, that these are not single rows of tubercles (a character typical for Physoporella lotharingica; & , 2000, p. 126, placed D. rauffi in the synonymy of Ph. lotharingica, albeit with a question mark), but double, condensed rows of alternating tubercles. Such a pattern allows to tentatively assign this dubious form to the new species, common in Granice, Physoporella polonoandalusica n.sp. (Fig. 21a-b
), and seems to be biased to make it fit the results of taphonomical alterations of the genus Diplopora.
(1920, p. 28; 1931a, p. 273, 275) included D. rauffi to D. annulata, but without explicit justification; he merely noted that both species differ in some characters. Indeed, the figure in there are no characters diagnostic for Diplopora - no annuli, no tufts of branches, and there are only rarely spaced whorls of tubercles of undetermined shape. It seems, that these are not single rows of tubercles (a character typical for Physoporella lotharingica; & , 2000, p. 126, placed D. rauffi in the synonymy of Ph. lotharingica, albeit with a question mark), but double, condensed rows of alternating tubercles. Such a pattern allows to tentatively assign this dubious form to the new species, common in Granice, Physoporella polonoandalusica n.sp. (Fig. 21a-b ![]() ). It is the more probable, that himself regarded his new species as most similar to Gyroporella dissita (), which has double rows of tubercles.
). It is the more probable, that himself regarded his new species as most similar to Gyroporella dissita (), which has double rows of tubercles.
Specimens very similar to those from Poland have been found in Spain. J.
(1935b) described them from Anisian of Prades, Tarragona province of Catalonia, from the collection of S. made in 1931. His so-called minor form is less than 1 mm across. Fairly long tubes are straight or bent. Double row (paired) tubercle whorls are
separated by deep furrows. Pl. II, 2 ![]() shows the inner side of a tube with pores (branches' inlets) in furrows separated by wide flat ridges (Fig. 21c
shows the inner side of a tube with pores (branches' inlets) in furrows separated by wide flat ridges (Fig. 21c ![]() ). These characters prompted
(1935b, p. 16) to attribute these specimens to Physoporella aff. lotharingica (), even though, as he noted himself, they exhibit double whorls, not observed in West German specimens. He believed this difference is
neglectible, because the character is very irregular among Physoporella praealpina. However, this particular character is actually stable and typical for the new species here discussed.
). These characters prompted
(1935b, p. 16) to attribute these specimens to Physoporella aff. lotharingica (), even though, as he noted himself, they exhibit double whorls, not observed in West German specimens. He believed this difference is
neglectible, because the character is very irregular among Physoporella praealpina. However, this particular character is actually stable and typical for the new species here discussed.
M. (1952) attributed the species Physoporella aff. lotharingica described by to the German-Andalusian province extending to the north of Betic Alp, which in Triassic belonged to the Alpine-Dinaride
palaeogeographic province. In his next paper ( et al.,
1954), wrote on Physoporella lotharingica, as a species typical for the German-Andalusian province, situated peripherally from the Briançon province, to the south of which the Alpine-Dinaride province is situated. The new specific name proposed herein alludes to the known extremes of its range - in Poland and Andalusia. Physoporella polonoandalusica n.sp. probably occurs also in Briançon and Alpine provinces. F. (1958, Pl. 6, 15) figured a specimen with double, widely spaced pore rows (seen from inside of the tube) that he regarded as Ph. praealpina , but which can be assigned to the new species (Fig. 21d ![]() ). The same is probable the case with the specimen (, 1979, Pl. 5, 3) assigned by E. (1979) to Physoporella leptotheca , where widely, though irregularly spaced double rows of tubercles are visible (Fig. 21e
). The same is probable the case with the specimen (, 1979, Pl. 5, 3) assigned by E. (1979) to Physoporella leptotheca , where widely, though irregularly spaced double rows of tubercles are visible (Fig. 21e ![]() ).
).
 |
|
|
Click on thumbnail to enlarge the image.
Figure 21: Selected previously published illustrations of specimens from the Upper Silesia, Andalusia, and Alps, similarly preserved as new specimens from the Upper Silesia, and belonging probably to Physoporella polonoandalusica n.sp.: a - pencil drawing of a holotype of Diplopora rauffi (1906, Pl. 3, 1) from Granice (locality no. 43 in the Kraków-Chrzanów Region of the Upper Silesia), a small form with widely spaced double whorls of alternating pores, x 4; b - schematic drawing of the same, redrawn from by (1938, fig. 8a), x 4; c - Physoporella aff. lotharingica from the Anisian of Prades, Tarragona Province, Andalusia (, 1935b, Pl. II, 2) with double whorls, x 5; d - a drawing of a specimen from Vanoise Massif in French Alps, etched with acetic acid, with widely spaced double whorls of pores (, 1958, Pl. 6, 15), x 7, attributed by to Physoporella praealpina; e - naturally weathered silicified specimens from the Anisian of M. Popera, Belluna Province, Eastern Dolomites (, 1979, Pl. 5, 3) with widely, though irregularly, spaced double whorls of tubercles, x 54.2.
Family Diploporaceae (, 1920) , 1988
Tribe Diploporeae (, 1920) , 1979
Kantia dolomitica (, 1912) , 1979
1912 Kantia dolomitica n.sp.- , Pl. VI (V), 14-16.
1920 Diplopora annulata var. dolomitica (forma vesiculifera) n.st. n.comb.- , Pl. V, 18-19, 21, 26, 27.
1963 Diplopora annulata.- , figs. 3-6, 10, 15.
1964 Diplopora annulata var. dolomitica.- , Pls. XXIX, 5; Pl. XXX, 1-3, 6; Pl. XXXI, 2-5; Pl. XXX, 4-6; Pl. XXXI, 1.
1973 Diplopora annulata var. dolomitica.- , Pl. VI, 4 pars.
1973 Diplopora annulata forma vesiculifera.- & , Pls. XVII, 65-68; Pl. XVIII, 72 pars.
1979 Kantia dolomitica emend.- , p. 631.
1979 Diplopora annulata dolomitica.- , Pl. 80, 2a-d; Pl. 80, 10-12, 14.
1986 Diplopora annulata dolomitica.- , Pls. CIV, 2a-d, 10-12, 14; Pl. CV, 2, from (1920, Pl. V, 21).
2000 Kantia dolomitica.- & , p. 83.
2013 Kantia dolomitica.- in & , Pl. VI (V), 14-16.
Material and localities.- This species may include the form from Libiąż (locality 45) illustrated by J. (1920, Pl. V, 20) as Diplopora annulata var. dolomitica (forma vesiculifera), and named by Diplopora annulata annulata (, 1986; Pl. V, 21), and which & (2000) classified recently to Kantia dolomitica. The illustration of this specimen does not show clear pores, thus it is not possible to determine, whether they are of vesiculiferous, or trichophorous type. J. (1931a, p. 276-277) described well preserved large specimens in thin sections. Specimens and sections from this locality are preserved in the 's collection, housed in the Naturhistorisches Museum Wien (Vienna, Austria). In the locality Libiąż, in the uppermost part of the quarry, Dasycladales from the Pelsonian-Illyrian boundary can be currently found. Previously, however, the topmost part of the Diplopora Dolomite was outcropping further towards NW, in the railroad cuts, where the Upper Muschelkalk and Keuper used to be exposed. Probably the specimen described by J. came from this outcrop.
New finds of Kantia dolomitica came from the Opole Triassic (Kamień Śląski, locality no. 3) from the Tarnowice Syncline (Srebrna Góra, no. 13), and from the Zawiercie area (deep borehole 11-Ż Żarki Letnisko).
The best material comes from Kamień Śląski, from Karchowice limestones, where (1975) preliminarily determined "Diplopora sp.". I received from Dr. K. two large photographs full of diplopores. Many
specimens from her photographs are reproduced in Pl. XXII ![]() . The original thin sections from which 's pictures were taken are now unfortunately lost.
. The original thin sections from which 's pictures were taken are now unfortunately lost.
Kantia dolomitica from Kamień Śląski has fairly large size. Its thallus is irregularly segmented. The annuli are wide, separated by deep troughs. The branches are clearly vesiculiferous type of "stalk and cup", grouped in several whorls per annulus. The troughs and annuli with branches are slightly obliquely arranged to the main axis of the alga. The axial cavity is wide, and the calcareous crust fairly thick.
The specimens from Srebrna Góra are poorly preserved, but in cross sections the vesiculiferous branches are visible (Pl. XXII, 14-15 ![]() ).
).
Some specimens (Pl. XXII, 23, 25 ![]() ) from the borehole 11-Ż (Żarki Letnisko) have already been illustrated before (, 1986, Pl. CV, 5, 7) as Diplopora annulata
() in the old, larger sense. Two new specimens are now added (Pl. XXII, 22, 26
) from the borehole 11-Ż (Żarki Letnisko) have already been illustrated before (, 1986, Pl. CV, 5, 7) as Diplopora annulata
() in the old, larger sense. Two new specimens are now added (Pl. XXII, 22, 26 ![]() ).
).
Diagnosis.- Cylindrical thallus with wide axial cavity. Annulation is regular and interannular furrows are large and deep. In every segment, there are several whorls of branches (minimum two). Branches are metaspondyl, vesiculiferous. The uppermost part of plant is frequently unsegmented with thin, trichophorous pores.
Stratigraphical range and geographical distribution.- Kantia dolomitica usually co-occurs with Diplopora annulata and is regarded an index fossil for the Ladinian (, 1912, 1920, 1931a, 1937b, 1942, in & , 2013; , 1957, 1965; , 1964) or the Lower Ladinian (, 1986). However, co-occurrence with Diplopora annulatisima may also indicate presence in the Upper Anisian (Illyrian - see , 1974, , 1964). In the Eastern Dolomites, in Monte Popera (Comelica), the species is regarded by (1979) as Upper Anisian (Avisianus Zone).
originally regarded Kantia dolomitica as a South Alpine and Dinaride species, and Diplopora septemtrionalis (=annulata) as a North Alpine form. Later, however, K. dolomitica has been also found in the Northern Calcareous Alps (, 1942) and in the Western Carpathians (, 1964, 1986). In the Tatras, K. dolomitica occurs abundantly, together with Diplopora annulata, in the Lower Sub-Tatric Series (Križna Nappe), where it belongs to the Lower Ladinian (, 1963, 1979, 1986).
The single specimen of K. dolomitica found in Libiąż, J. (1931a, p. 277), regarded as Ladinian and thus assumed that in the Cracovian Triassic only the top part of the Upper Silesian Diplopora Dolomite is preserved, and that its lower part belongs to the Anisian (ibid., p. 276). Nowadays, it is known that the whole Diplopora Dolomite is of Anisian age, and that Diplopora annulata co-occurs with D. annulatissima, belonging to the Upper Illyrian (Avisianus Zone - , 1986). The Illyrian age is corroborated by the Illyrian conodonts found by K. (1975) above the Tarnowice Beds. Anisian age of the Diplopora Dolomite has been assumed by H. (1974a, 1974b) and by E. and H. (1993). The large facial variability of the Opole Triassic is evidenced by the find of Dasycladales in Karchowice Limestone in Kamień Śląski (, 1975), which were here assigned to Kantia dolomitica. The limestone facies in this section reaches up to the Upper Illyrian.
Nomenclature.- The nomenclatorical issues on Diplopora and Kantia are discussed in more detail when describing Diplopora annulata. T. (1979) restored original 's designations: Diplopora annulata and Kantia dolomitica; this has been accepted by & (2000). Despite some doubts concerning the strict formal nomenclature, their terms have been used herein.
Kantia uniserialis (, 1912) , 1979
1912-1913 Kantia philosophi.- in , p. 217.
1920 Diplopora uniserialis n.sp. (forma vesiculifera).- , Pl. V, 1, 4-5, 7.
1933 Diplopora annulata var. brianconnensis n.var. forma vesiculifera.- , Pls. I, 1; Pl. II, 3-4.
1958 Diplopora cf. brianconnensis = Diplopora uniserialis.- , Pl. 7, 21-22, 23; Pl. 22, 4.
1961 Diplopora uniserialis.- , Pl. 4.
1979 Kantia uniserialis.- , p. 631, Pl. V.
1979 Diplopora uniserialis.- , Pl. 80, 6-9 (pars).
1986 Diplopora uniserialis.- , Pl. CIV, 6-9 (pars).
1994 Kantia, cf. Kantia uniserialis.- et al., Pl. 13, 8.
2000 Kantia uniserialis.- & , p. 89-90.
Discussion.- J. (1920) erected the species Diplopora uniserialis based on material from Berne Alps provided by F. and discerned two forms within the new species: forma trichophora and forma vesiculifera. The two forms were regarded by T. (1979) as separate species: Diplopora uniserialis and Kantia uniserialis. The latter species has vesiculiferous pores (,
1961, Pl. lV, 4, p. 66). More detailed description and stratigraphic-palaeogeographic comments are given in the section describing Diplopora uniserialis. Both species are typical for the Briançon - Križna - High-Tatric zone. They are especially abundant in the lower Ladinian of the Križna Series, with rarer occurrences in the
High-Tatric Illyrian. In the Ladinian strata their co-occur with Diplopora annulata and Kantia dolomitica, and in the Illyrian - with Diplopora annulatissima (, 1986). In the Upper Silesia (Opole Triassic), poorly preserved and rare Kantia cf. uniserialis (Pl. XXV, 15-17 ![]() ) occurs in the uppermost Illyrian with Diplopora annulata and Kantia dolomitica.
) occurs in the uppermost Illyrian with Diplopora annulata and Kantia dolomitica.
Kantia comelicana (, 1979)
Pls. XX ![]() ,
XXIII
,
XXIII ![]() -
XXIV
-
XXIV ![]() &
XXXVII
&
XXXVII ![]() ;
Fig. 22
;
Fig. 22 ![]()
1966 Diplopora annulata var. dolomitica.- , p. 255, text-fig. 2.
1979 Diplopora comelicana n.sp.- , Pl. 1, 1-7; Pl. 2, 1-5; Pl. 3, 10a; Pl. 4, 1-12.
1982 Diplopora comelicana.- , Pl. 2, 2.
1983 Diplopora comelicana.- & , Pl. 1, a (pars), c (pars).
1986 Diplopora comelicana.- , Pl. IV, 2, 4.
2000 Diplopora comelicana.- & , p. 40.
Material and localities.- Several specimens in fractures from the Upper Silesia. In the Cracow-Chrzanów area (localities no. 46, Żelatowa; no. 48, Rosowa Góra; no. 50, Balin; no. 51, Cezarówka; no. 52, Krupka), the Siewierz-Olkusz Monocline (localities no. 57, Stare Gliny, no. 57, Brudzewice; no. 72, Żelisławice), in boreholes from the Zawiercie area (borehole 83-Ż Myszków, 51-Ż Myszków, 10Ż Lgota Nadwarcie, 37-WB Winowno-Bendusz; 72-KM Koziegłowy), Bytom Syncline (locality no. 25, Przełajka).
Description.- Thallus cylindrical, never annulated nor undulated, with sporadic fissuration. Outer wall continuous
(Pls. XXIII, 2 ![]() &
XXIV, 5
&
XXIV, 5 ![]() ). Central cavity large (Pl. XXIII, 2, 4
). Central cavity large (Pl. XXIII, 2, 4 ![]() ), hollow (Pl. XXIII, 2, 4, 16
), hollow (Pl. XXIII, 2, 4, 16 ![]() ) or filled with sediment, forming central stem
(Pls. XXIII, 18-19
) or filled with sediment, forming central stem
(Pls. XXIII, 18-19 ![]() &
XXIV, 3, 8, 14-17
&
XXIV, 3, 8, 14-17 ![]() ). Branches (tubercles) metaspondyl, densely whorled
(Pls. XXIII, 3
). Branches (tubercles) metaspondyl, densely whorled
(Pls. XXIII, 3 ![]() &
XXIV, 8, 12, 14, 19-20
&
XXIV, 8, 12, 14, 19-20 ![]() ),
acroporous to slightly vesiculiferous, very frequently diverging (Pls. XXIII, 2
),
acroporous to slightly vesiculiferous, very frequently diverging (Pls. XXIII, 2 ![]() &
XXIV, 7-8, 12, 14-15, 17
&
XXIV, 7-8, 12, 14-15, 17 ![]() ). Metaspondyl branches grouped in clusters of three or four
(Pls. XXIII, 7-11
). Metaspondyl branches grouped in clusters of three or four
(Pls. XXIII, 7-11 ![]() &
XXIV, 1, 3-4, 9, 14, 16, 18-19
&
XXIV, 1, 3-4, 9, 14, 16, 18-19 ![]() ). Most frequently they are double, kidney-shaped, but with a marked vesicle. In some cases very characteristic arrangement of branches is observable - typical double branches with a third branch of the cluster growing between them
(Pls. XXIII, 7
). Most frequently they are double, kidney-shaped, but with a marked vesicle. In some cases very characteristic arrangement of branches is observable - typical double branches with a third branch of the cluster growing between them
(Pls. XXIII, 7 ![]() &
XXIV, 8
&
XXIV, 8 ![]() ), see Fig. 22c
), see Fig. 22c ![]() (with arrow) of the paratype. The presence of typical vesiculiferous branches is the reason for attribution of this species to the genus Kantia, not to Diplopora¸ according to the scheme of (1979), accepted by & (2000).
(with arrow) of the paratype. The presence of typical vesiculiferous branches is the reason for attribution of this species to the genus Kantia, not to Diplopora¸ according to the scheme of (1979), accepted by & (2000).
Stratigraphical range and geographical distribution.- Kantia comelicana () is typical for the Upper Anisian (Avisianus Zone) at Monte Popera in the Eastern Dolomites (Belluno). J. (1982, 1986) attributes it to the Lower Ladinian (Fassanian) of the uppermost Sub-Tatric Series (Stražov Nappe). The Upper Silesia is the third region where occurrence of Kantia comelicana is found. This species, previously found in the Southern Alps and in the uppermost Sub-Tatric Series in S Slovakia, was considered a typical representative of the southern Alpine zones in the Tethys realm. For that reason, the presence of widely distributed, perfectly preserved and easily determinable Kantia comelicana in so northerly situated region of the Tethyan periphery is very surprising. Apparently, the connections of the remote Germanic Basin in the Upper Silesia with the Tethys Ocean were still sufficiently large and easy.
 |
|
|
Click on thumbnail to enlarge the image.
Figure 22: Selected previously published illustrations of Kantia comelicana (): a-c - microphotographs of sectioned specimens from Upper Anisian of M. Popera, Belluna Province, Eastern Dolomites, attributed by (1979, Pl. 1, 1-3) to her new species Diplopora comelicana: a - longitudinal section (holotype), x 8.3, b - an agglomeration of specimens visible in various sections (paratype), x 8.2, c - longitudinal section (paratype), x 8.5; d-f - naturally weathered silicified specimens: d - inside view of the central cavity with pore clusters, e-f - upper view on typical vesiculiferous branches (pores), e x 78, f x 85; drawings (, 1966, text-fig. 2) based on microphotographs of sectioned specimens from the Lower Ladinian of Drienok Series (uppermost Sub-Tatric Nappe, Central Slovakia), assigned by him to Diplopora annulata () var. dolomitica (), and by (1979) to Diplopora comelicana: g - tangential section, h-f - oblique longitudinal sections, k-n - transverse sections.
Diplopora annulata (, 1863)
1853 Nullipora annulata n.sp.- , Pl. VI, 1a-f.
1862 Nullipora annulata.- , Pl. XVI, p. 246, 309.
1863 Diplopora annulata n.gen. n.comb.- , Pl. LXV. e, 6a-b.
1912 Diplopora annulata.- , Pl. VII (VI), 1-2, 14-15; Pl. VII (VI), 9-11; Pl. VII (VI), 12-13; Pl. VII (VI), 16; Pl. VII (VI), 17; Pl. VIII (VII), 1-2.
1920 Diplopora annulata var. septemtrionalis (forma trichophora) n.var.- , Pl. V, 14-15, 23; Pl. V, 16, 20.
1920 non Diplopora rauffi.- , p. 28.
1928 non Diplopora annulata.- , fig. 7.
1928 Diplopora annulata, var. septemtrionalis.- , fig. 8b.
1928 non Diplopora annulata, var. septemtrionalis.- , fig. 8a.
1935a non Diplopora rauffi.- , p. 273, 275.
1938 Diplopora annulata.- , Pl. I, fig. 1.
1950 Diplopora annulata.- , Pl. I, 2-3; Pl. II, 1-2; Pl. IV, 1 pars.
1957 Diplopora annulata subsp. annulata, n.subsp. , p. 52.
1963 Diplopora annulata.- , Pls. III, 1-2; IV, 1-2.
1964 Diplopora annulata var. annulata nom.nov.- , Pl. XXIX, fig. 1-3.
1965 Diplopora annulata subsp. annulata.- , Pl. VI, 2-4.
1965 Diplopora annulata var. annulata forma trichophora.- , Pl. 54, 4-6; Pl. 55.
1966 Diplopora annulata var. annulata.- , Pl. VIII, 7.
1967 Diplopora annulata.- , Pl. IV, 5.
1969 Diplopora annulata var. annulata. & , Pl. VI, 1-3; Pl. VII, 3.
1970 Diplopora annulata var. annulata.- , Pl. III, 1.
1972b Diplopora annulata.- , Pl. I, 1-7; Pl. II, 1, 7-8.
1979 Diplopora annulata annulata.- , Pl. 80, 13.
1979 Diplopora annulata.- , p. 630.
1981 Diplopora annulata.- et al., fig. 7, 15.8.
1982 Diplopora annulata var. annulata.- , Pl. 2, 1 pars, 3-4; Pl. 4, 2 pars, 3 pars.
1986 Diplopora annulata var. annulata.- , Pl. IV, 1, 6.
1986 Diplopora annulata.- , Pl. CV, 3-7.
1993 Diplopora annulata.- et al., Pl. 55, 2-5.
2000 Diplopora annulata.- & , p. 32-38.
2013 Diplopora annulata.- in & , Pl. VII (VI) 1-2, 14-15; Pl. VII (VI), 9-11; Pl. VII (VI), 12-13; Pl. VII (VI), 16; Pl. VII (VI), 17; Pl. VIII (VII), 1-2.
Diagnosis.- The thallus is cylindrical, irregularly annulated, with branches generally trichophorous, rarely vesiculiferous (in the upper part of thallus), arranged in tufts and whorls, from one to several whorls in each segment (but never only one, as in D. annulatissima). In the Diplopora Dolomite from the Upper Silesia, with its peculiar state of preservation, annulation is hardly discernible. In this case, D. annulata differs from D. annulatissima by larger spaces between double or triple whorls of tubercles (branches).
Material and localities.- There are very few preserved rock samples and thin sections from the Polish Upper Silesia and adjacent regions. According to (1931a, p. 275), in the collections stored in the Naturhistorisches Museum Wien there are rock samples labelled "Schlesien leg.". This material yielded a beautiful longitudinal section of Diplopora annulata, illustrated by (1920, Pl. V, 16) and reproduced by (1986, Pl. CV, 1). Specimens of from the Upper Silesia, labelled by him Gyroporella annulata and G. cylindrica, and actually possibly belonging to D. annulata (see synonymy in & , 2000, p. 32), are housed in the collection of prof. in Munich. These are, however, probably specimens of D. annulatissima . A. (1878) received from F. from the vicinity of Tarnowskie Góry, that he used in comparative studies on Jurassic Actinoporellae from Niżniów in Podole. His paper contains the first figures of Triassic Dasycladales published in the Polish literature (, 1878, Pl. VI, 9-11), then labelled Gyroporella annulata. These are, however, not illustrations of specimens obtained from , but reproduced figures of (1872a, Pl. D II, 1a, e, 2e, f), showing forms that he assigned to Gyroporella annulata and G. cylindrica, and actually belonging partly to Diplopora annulatissima. Specimens given to by are probably lost; at least they could not be located in any of the Cracow museums. (1926a, p. 505) believed that D. annulata was the most common species in the Diplopora Dolomite, especially in its lower part. This assumption is erroneous, because in fact the species is very rare and occurs only in the topmost part of the Diplopora Dolomite.
Housed in the Geological Museum of the Polish Geological Institute in Warsaw there is a collection of Z. , MuzPIG no. 1653.II and 1682.II. It contains thin sections with Diplopora annulata from the borehole Żarki-Letnisko 11-Ż (, 1986, Pl. CV, 3-7) and rock fragments from the localities Segiet (no. 11) and Fryderyk Deep Shaft (no. 12) with indicated D. annulata, co-occurring with D. annulatissima.
Nomenclature.- 's species, that he originally named Nullipora annulata (1853), and later (1863) called Diplopora annulata, has been accepted by (1912, in & , 2013) under the latter name. Later changed his original diagnosis (, 1920), discerning three varieties: dolomitica, septemtrionalis, and debilis; and two forms: trichophora and vesiculifera, while rejecting the previously established genus Kantia with vesiculiferous pores. Both forms were interpreted by as sexually dimorphic, and regarded as geographical varieties. M. (1957) criticized 's interpretation, rejecting the possibility of sexual dimorphism in Diplopora annulata, and in Dasycladales in general (it is absent among Recent Dasycladales). He also rejected the varieties, and instead erected two subspecies - Diplopora annulata subsp. annulata and Diplopora annulata subsp. dolomitica. J. (1964) did not accept 's arguments and continued to discern 's varieties - Diplopora annulata var. dolomitica and var. debilis, substituting only var. septemtrionalis with var. annulata, to conform the rules of botanical nomenclature. Many subsequent authors used the names Diplopora annulata annulata and D. annulata dolomitica (see , 1970; , 1979, 1986). Applying the strict formal rulings of the International Code of Botanical Nomenclature (1961), T. (1979) restored the original 's names Diplopora annulata, Kantia dolomitica, and they were accepted by & (2000).
Remarks.- The species Diplopora annulata (.) is the most commonly listed by field geologists, both older and modern. Also (1920) believed it is the most common species in the Upper Silesia and in adjacent regions. However, soon it became obvious (, 1931a, p. 272), that, incidentally, the first specimens he obtained from Silesian geologists indeed belonged to this species. Also (1906, p. 81) and (1926a, p. 505) regarded this species as very common. Specimens obtained later from field geologists and from the Preussischen Geologischen Landesanstalt, persuaded that Diplopora annulata is actually not so widely distributed as originally believed. Usually, the collective name was used for large specimens, visible with naked eye, and belonging to Diplopora annulatissima and Physoporella praealpina . (1920, p. 28; 1931a, p. 275) included to this species also Diplopora rauffi , from the locality Granice (no. 43). The research in this locality shown that there are no representatives of the genus Diplopora, and only numerous Physoporellae. 's inadequately described and illustrated Diplopora rauffi show some similarity to Physoporella polonoandalusica n.sp. The figure in (1906, T. 3, 3) does not show clearly, if there are single or double rows of condensed, alternating tubercles. If there are single rare rows of tubercles, than the species in question would be similar to Physoporella lotharingica (). If, however, there are double rare rows of tubercles, the species would be closer to Ph. polonoandalusica n.sp.
The single long wonderfully preserved specimen of Diplopora annulata (),
labelled on section and illustrated by
(1920, Pl. V, 16) comes from the Upper Silesia ("Schlesien, leg."), with no exact locality given (probably from the Tarnowice Syncline). The species occurs also in other localities from the syncline (Segiet, no. 11 and Fryderyk Deep Shaft, no. 12), in Kamień Śląski - no. 3 (Pl. XXV, 18a-b ![]() ) and in boreholes from the vicinity of Zawiercie (, 1986, Pl. CV, 3-7).
) and in boreholes from the vicinity of Zawiercie (, 1986, Pl. CV, 3-7).
Localities.- Diplopora annulata () occurs in the uppermost part of the Diplopora Dolomite, slightly below Tarnowice Beds. This uppermost part of the Diplopora Dolomite has been intensely eroded subsequently and is preserved only in few localities. As for the Opole Triassic, photographs of Karchowice limestone from Kamień Śląski (locality no. 3), published by (1975, Pl. 27) show some Diplopora annulata specimens among numerous Kantia dolomitica specimens.
Diplopora annulata has been found by older German researchers in the Tarnowice Syncline ("Schlesien, leg." - , 1920, Pl. V, 16), but most often Diplopora annulatissima occurs here (, 1931a, p. 270). Resent research led to finding of Diplopora annulata () with rarely spaced interannular furrows in Segiet (locality no. 11) and Fryderyk Deep Shaft (no. 12). These specimens are visible on fractures. Near Zawiercie, in the borehole Żarki Letnisko 11-Ż, a Diplopora annulata () was determined in thin section, luckily preserved below Tarnowice Beds (, 1986, Pl. CV, 3-7).
Stratigraphical range and geographical distribution.- Diplopora annulata () was regarded by (1912, 1920, 1931a, 1937b, 1942, in & , 2013) as a Ladinian species - its stratum typicum is Wetterstein Dolomite from the Northern Calcareous Alps. A similar range and age is shown by the species from numerous localities from other parts of the Alps, Dalmatia, Bosnia, and Velebit (, 1937a; , 1957). In the Western Carpathians the species is usually Lower Ladinian (co-occurrence with Teutloporella herculea in the Upper Sub-Tatric Nappe - , 1973) and Upper Anisian (co-occurrence with D. annulatissima - , 1964, 1986). In the Lower Sub-Tatric Series (Križna Series) in the Tatras, it is the index species for Lower Ladinian (, 1963, 1979, 1986). Many scientists, especially E. (1974) and J. (1964, 1986), believed that co-occurrence of D. annulata with D. annulatissima indicates Upper Illyrian age (Avisianus Zone) of the lowest Wetterstein Dolomite and its equivalents. A similar situation concerns the Upper Silesia and adjacent areas, where D. annulata co-occurs with Kantia dolomitica and D. annulatissima (, 1986) in the upper part of the Diplopora Dolomite, below the Tarnowice Beds. Because K. (1975) found Illyrian conodonts above the Tarnowice Beds, the upper part of the Diplopora Dolomite also belongs to the Illyrian. Thus, the long held opinion, that the Diplopora Dolomite is of Ladinian age (, 1912, 1920, 1931a, in & , 2013; , 1952; , 1979), is of purely historical interest. The lower part of the Diplopora Dolomite belongs to the Pelsonian, and the upper part - to the Illyrian. The Opole Triassic, where the Karchowice limestone facies reaches up to the Upper Illyrian, shows substantial facial variability. Photographs of Karchowice limestone from Kamień Śląski (locality no. 3), published by (1975, Pl. 27) show some Diplopora annulata specimens among numerous Kantia dolomitica specimens.
Diplopora uniserialis (, 1920) , 1979
1912-1913 Kantia philosophi.- in , p. 217.
1920 Diplopora uniserialis n.sp. (forma trichophora).- , Pl. V, 2, 6.
1933 Diplopora annulata var. brianconnensis n.var. forma trichophora.- , Pl. I, 2-3; Pl. II, 1-2, 5-8.
1969 Diplopora uniserialis var. ligurica. & , Pl. I, 4-6, 13-14.
1979 Diplopora uniserialis.- , Pl. 80, 6-9 (pars).
1979 Diplopora uniserialis.- , p. 630.
1986 Diplopora uniserialis.- , Pl. CIV, 6-9 (pars).
2000 Diplopora uniserialis.- & , p. 48-49.
Diagnosis and nomenclature.- J. (1920, p. 89) erected the species Diplopora uniserialis based on material from Berne Alps provided by F. . Earlier, he regarded the species as Diplopora annulata var. debilis. The name Diplopora uniserialis refers to the presence of a single whorl of branches in each segment of this densely segmented species. This alga is much smaller than Diplopora annulata. Annuli are very short and narrowing outwardly, assuming triangular shape (in cross section); the interannular furrows are wide and widening outwardly. The branches are perpendicular to the central axis. There are about 4 branches in each tuft.
discerned two forms within the new species: forma trichophora (with thin pores, narrowing towards the outer surface and opening to the outside) and forma vesiculifera (with thick, club-shaped pores encased in calcareous crust). The two forms were regarded by T. (1979) as separate species and resurrected the old generic name Kantia (, 1912, in & , 2013). Thus, the material now belongs to two species: Diplopora uniserialis and Kantia uniserialis.
Material and localities.- Both species co-occur in the same strata. In the upper part of the Upper Silesian Diplopora Dolomite they are infrequent and difficult to discern in fractures. They can be diagnosed based on thin sections, albeit also with difficulties. Diplopora cf. uniserialis is recognizable from the borecore Żarki Letnisko 11-Ż (Pl. XXV, 3-4 ![]() ), Tarnów Opolski (locality no. 2). Better recognizable Diplopora uniserialis occurs in Srebrna Góra (no. 13), Tarnów Opolski (no. 2) and in Kamień Śląski (no. 3) - Pl. XXV, 11-15
), Tarnów Opolski (locality no. 2). Better recognizable Diplopora uniserialis occurs in Srebrna Góra (no. 13), Tarnów Opolski (no. 2) and in Kamień Śląski (no. 3) - Pl. XXV, 11-15 ![]() . In the Opole Triassic, the species occurs with Kantia uniserialis, Kantia dolomitica and Diplopora annulata in the uppermost Illyrian.
. In the Opole Triassic, the species occurs with Kantia uniserialis, Kantia dolomitica and Diplopora annulata in the uppermost Illyrian.
Stratigraphical range and geographical distribution.- Diplopora uniserialis was first collected by F. , and determined by (1920), in Berne Alps, in Swiss Prealpes, where it was regarded an Upper Anisian taxon. D. (1933), found it in the Briançon Triassic. In the Briançon Series of the French Inner Alps it has been later found by F. (1958). Diplopora uniserialis forms there the third diploporan horizon, belonging to the Lower Ladinian. Also in the Alpes Maritimes, Diplopora uniserialis is Ladinian ( & , 1969). The species is absent from the Eastern Alps (, 1920). It appears, however, in the Tatra Mountains, where it is especially abundant in the Lower Sub-Tatric Series (Križna Nappe) of Lower Ladinian age (, 1979, 1986), but occurs also in the Upper Anisian High-Tatric Series. Križna and Hightatric Series are palaeogeographic equivalents of the Briançon Series in the Western Alps (, 1977, 1994b). F. (1963) found a continuation of the Briançon Series in the Aosta River Valley, at the Mount Blanc foreland. He also believed that its further equivalents are present in the Graubünden in the Eastern Switzerland and in the High Tauern in Austria.
The presence of Diplopora uniserialis and Kantia uniserialis in the Upper Silesia (Opole Triassic, Tarnowice Syncline) and adjacent regions (boreholes in the Zawiercie area) indicates wide contacts between the Briançon-High-Tatric zone with the Upper Silesian zone. Both co-occurring species are accompanied by Diplopora annulata and Kantia dolomitica in the uppermost Illyrian.
Diplopora annulatissima , 1920
Pls. XXVI ![]() -
XXVII
-
XXVII ![]() -
XXVIII
-
XXVIII ![]() -
XXIX
-
XXIX ![]() -
XXX
-
XXX ![]() ;
Fig. 23
;
Fig. 23 ![]()
1870 Cylindrum annulatum.- , Pl. 11, 1-2.
1872a Gyroporella cylindrica n.sp.- , Pl. D.I, 8; Pl. D.II, 2a-o.
1872a pars Gyroporella annulata n.sp.- , Pl. D.II, 1.
1878 Gyroporella annulata.- .
1878 Gyroporella annulata.- , Pl. VI, 9 (from , 1872a, Pl. D.II, 1a), 10 (from , 1872a, Pl. D.II, 1c).
1912 Diplopora cf. cylindrica.- & , p. 739 and 745.
1919 ? Kantia monregalensis n.sp.- , fig. 6, 6.a-c.
1920 Diplopora annulatissima n.sp.- , Pl. IV, 11-16.
1931a Diplopora annulatissima.- , Pl. XXI, 1-2.
1933 Diplopora annulata var. briançonnensis var.nov.- , Pl. II, 13-14.
1957 Diplopora annulatissima.- , Pl. VII, 3.
1957 ?Diplopora monregalensis.- , p. 49.
1958 Diplopora cf. annulatissima.- , Pl. 6, 20-21.
1964 Diplopora annulatissima.- , Pl. XXIV, 4-5; Pl. XXV, 4; Pl. XXVI, 3-4; Pl. XXVII, 1-3; Pl. XXVIII, 1-4.
1968 Favoporella annulata n.gen. n.sp.- , Pl. I, 1-4; Pl. II, 1-4; Pl. III, 1-5; Pl. IV, 1-5.
1979 Diplopora annulatissima.- , Pl. 79, 1c, 2 d-e, 7-9.
1986 Diplopora annulatissima.- , Pl. CIII, 1c, 2d-e, 7-9 (from , 1979, Pl. 79, 1c, 2d-e, 7-9); Pl. CV, 8 (from , 1931a, Pl. XXI, 1), 9, (from , 1931a, Pl. XXI, 2), 10-13.
1986 Diplopora annulatissima.- E. , Pl. 4, 8.
1986 Diplopora annulatissima.- , Pl. III, 8-9.
1993 Diplopora annulatissima.- et al., Pl. 55, 1, 6, 8; Pl. 56, 14.
2000 Kantia monregalensis.- & , p. 85.
Diagnosis.- Perfect annulation with regular thin segments, separated by deep interannular furrows, reaching to the very large central cavity. In each segment there is only one whorl of distinct extremely trichophorous branches arranged in tuffs (metaspondylity) with 3-6 branches in one tuft. Branches, large at the base, are consequently thinning towards the external wall, forming double row of pores (tubercles). Occasionally, not only the branches of 1st order are calcified, but calcification also extends onto the cortex with delicate geometric patterns (comp. Favoporella annulata ).
Material and localities.- Diplopora annulatissima is one of the most frequent species in the Diplopora Dolomite (Figs. 2 ![]() & 5
& 5 ![]() ). It is distributed in all regions - in the Upper Silesia (Opole Triassic, Tarnowice Syncline, Bytom Syncline) and in adjacent areas (Cracow-Chrzanów region, Olkusz-Siewierz region, as well as in boreholes within the Zawiercie area). It was found in the upper part of the Diplopora Dolomite and is the index fossil for the Illyrian substage. The fractures with best preserved specimens come from the locality no. 8, Jemielnica
(Pls. XXVI
). It is distributed in all regions - in the Upper Silesia (Opole Triassic, Tarnowice Syncline, Bytom Syncline) and in adjacent areas (Cracow-Chrzanów region, Olkusz-Siewierz region, as well as in boreholes within the Zawiercie area). It was found in the upper part of the Diplopora Dolomite and is the index fossil for the Illyrian substage. The fractures with best preserved specimens come from the locality no. 8, Jemielnica
(Pls. XXVI ![]() -
XXVII
-
XXVII ![]() -
XXVIII
-
XXVIII ![]() ) and Przełajka (Pl. XXX
) and Przełajka (Pl. XXX ![]() ). The specimens from thin sections are shown in Pl. XXIX
). The specimens from thin sections are shown in Pl. XXIX ![]() .
.
Description: The description is based upon the analysis of specimens visible in fractures and their photographs
(Pls. XXVI ![]() -
XXVII
-
XXVII ![]() -
XXVIII
-
XXVIII ![]() -
XXIX
-
XXIX ![]() ). The axial cavity is fairly wide
(Pls. XXVI, 4-5
). The axial cavity is fairly wide
(Pls. XXVI, 4-5 ![]() -
XXVII, 6, 8-13
-
XXVII, 6, 8-13 ![]() -
XXVIII, 1, 4, 8-18
-
XXVIII, 1, 4, 8-18 ![]() -
XXIX, 3, 5, 9-12, 14-17
-
XXIX, 3, 5, 9-12, 14-17 ![]() ), hollow or filled with sediment. The surface of the chamber is smooth, although a delicate intusannulation may be occasionally visible, consisting of furrows and ridges. The pores (branch outlets) are located in furrows; but their whorls may occasionally overlap ridges
(Pls. XXVI, 3-13
), hollow or filled with sediment. The surface of the chamber is smooth, although a delicate intusannulation may be occasionally visible, consisting of furrows and ridges. The pores (branch outlets) are located in furrows; but their whorls may occasionally overlap ridges
(Pls. XXVI, 3-13 ![]() &
XXIX, 9
&
XXIX, 9 ![]() ). Sometimes the axial cavity wall is preserved
(Pls. XXVII, 4
). Sometimes the axial cavity wall is preserved
(Pls. XXVII, 4 ![]() &
XXVIII, 9, 16
&
XXVIII, 9, 16 ![]() ), reached by the annuli and interannular furrows
(Pls. XXVII, 8, 10
), reached by the annuli and interannular furrows
(Pls. XXVII, 8, 10 ![]() &
XXIX, 9
&
XXIX, 9 ![]() ).
).
The annulation is very dense and regular. The annuli are covered from outside with a thin calcareous crust
(Pls. XXVI, 3-5, 11 ![]() &
XXVII, 2-4, 7, 9
&
XXVII, 2-4, 7, 9 ![]() ). Usually, however, the fracture passes through an annulus, and then the branches (tubercles) are visible. Interannular furrows are much better preserved. They consist of two lists: upper and lower, connected with a streamlined joint near the wall of the axial cavity
(Pls. XXVI, 8
). Usually, however, the fracture passes through an annulus, and then the branches (tubercles) are visible. Interannular furrows are much better preserved. They consist of two lists: upper and lower, connected with a streamlined joint near the wall of the axial cavity
(Pls. XXVI, 8 ![]() ,
XXVII, 7, 10-13
,
XXVII, 7, 10-13 ![]() &
XXVIII, 6
&
XXVIII, 6 ![]() ). The joint is best visible when the interannular furrow is completely filled with sediment
(Pls. XXVII, 10-12
). The joint is best visible when the interannular furrow is completely filled with sediment
(Pls. XXVII, 10-12 ![]() &
XXVIII, 6
&
XXVIII, 6 ![]() ). Usually, however, the furrow is empty, and then there appears a visible fissure separated by furrow lists from the annulus
(Pls. XXVI, 1, 5, 8-9, 12, 14
). Usually, however, the furrow is empty, and then there appears a visible fissure separated by furrow lists from the annulus
(Pls. XXVI, 1, 5, 8-9, 12, 14 ![]() &
XXVII, 1, 12
&
XXVII, 1, 12 ![]() ). The filled interannular furrows look like ribs
(Pl. XXVIII, 6
). The filled interannular furrows look like ribs
(Pl. XXVIII, 6 ![]() ), slightly convex in the middle
(Pl. XXVIII, 6, 8, 13
), slightly convex in the middle
(Pl. XXVIII, 6, 8, 13 ![]() ). The annuli and furrows are usually perpendicular to the axis of an alga, but sometimes they are slightly oblique
(Pls. XXVI, 4-5
). The annuli and furrows are usually perpendicular to the axis of an alga, but sometimes they are slightly oblique
(Pls. XXVI, 4-5 ![]() &
XXVII, 4-5, 11-12
&
XXVII, 4-5, 11-12 ![]() ).
).
On tangential-longitudinal fractures (Pls. XXVI ![]() -
XXVII
-
XXVII ![]() ) the annuli are evidently filled with branches (tubercles). In fractures closer to the axial cavity, a double row of alternating tubercles is visible. In more peripheral fractures, triple rows of densely packed tubercles are visible. In longitudinal fractures, the shape of tubercles is difficult to ascertain, abut in internal fractures they are noticeably thicker than in the external ones. Some tubercles are hollow
(Pl. XXVI, 1, 5
) the annuli are evidently filled with branches (tubercles). In fractures closer to the axial cavity, a double row of alternating tubercles is visible. In more peripheral fractures, triple rows of densely packed tubercles are visible. In longitudinal fractures, the shape of tubercles is difficult to ascertain, abut in internal fractures they are noticeably thicker than in the external ones. Some tubercles are hollow
(Pl. XXVI, 1, 5 ![]() ). At the base, near the wall of the axial cavity, the branches (tubercles) form tufts of 3-6 branches of metaspondyl type
(Pls. XXVII, 5-6
). At the base, near the wall of the axial cavity, the branches (tubercles) form tufts of 3-6 branches of metaspondyl type
(Pls. XXVII, 5-6 ![]() ,
XXVIII, 1, 10, 15
,
XXVIII, 1, 10, 15 ![]() &
XXIX, 4
&
XXIX, 4 ![]() ). The shape of branches is best visible in transverse fractures
(Pl. XXVIII
). The shape of branches is best visible in transverse fractures
(Pl. XXVIII ![]() ). The branches are long and thin, slightly thinning towards the
outside (thus of trichophore type). It is shown in many photographs (see
Pls. XXVII, 12a
). The branches are long and thin, slightly thinning towards the
outside (thus of trichophore type). It is shown in many photographs (see
Pls. XXVII, 12a ![]() ,
XXVIII, 1-3, 5, 7-10, 12-16
,
XXVIII, 1-3, 5, 7-10, 12-16 ![]() &
XXIX, 1-4
&
XXIX, 1-4 ![]() ). Quite rarely the branches expand slightly externally
(Pl. XXVIII, 4
). Quite rarely the branches expand slightly externally
(Pl. XXVIII, 4 ![]() ), but never form a vesicle, characteristic of the vesiculiferous type. Thus, it is well established that this species belongs to the genus Diplopora and not to Kantia.
), but never form a vesicle, characteristic of the vesiculiferous type. Thus, it is well established that this species belongs to the genus Diplopora and not to Kantia.
Nomenclature.- This very typical species was described as Diplopora annulatissima by (1920). The name alludes to the unusually regular fine annulation, different from irregular annulation of D. annulata. In each annulus, there is a single row of branches (pores), thick at the base, and gradually thinning distally (trichophorous type). The pores form tufts of 3-4 branches, extending from a very large axial cavity. The name has been widely accepted and has been described and illustrated from many countries (see synonymy), especially that it was regarded an index species for the Illyrian (, 1930a, 1931a, 1936, 1937a; , 1964, 1986; , 1965). However, M. (1957) noted that Kantia monregalensis (, 1919, fig. 6) is actually identical with a species described by a year later (1920, Pl. IV, 12-14); thus the priority issue arise. If we assume that Kantia monregalensis was erected through legitimate description, the species should be officially named Diplopora monregalensis, as the generic name Kantia has been rejected by himself (1920). However, (1957, p. 49) regards this impossible, because under the name Kantia monregalensis heterogeneous specimens were described, and the description by does not fit the types, currently regarded as the true representatives of the species. Thus, the name Diplopora annulatissima should be retained. It is to be noted here, however, that A. (1919) describing her species explicitly stated that the pores widened distally, and it is because of that feature she classified her species monregalensis to the genus Kantia (with vesiculiferous pores) and not to Diplopora (with trichophorous pores). Thus, 's species clearly differs from 's D. annulatissima, which according to diagnoses by , & , had distinctly trichophorous branches. M. (1957, p. 50) tried to explain this by comparing Diplopora annulatissima with Diplopora annulata (in the wide sense of , 1920) and with Diplopora uniserialis. In both species, there are both vesiculiferous and trichophorous forms (varieties, subspecies). thus believes that the same approach can be applied to Diplopora annulatissima.
B. and T. (2000), despite the above mentioned differences in pore shapes and applying strictly the priority principle, assigned all specimens described till then to Kantia monregalensis , 1919.
However, if the priority principle were to be strictly applied, the priority would surely belong to C.W.
(1872a), whose Gyroporella cylindricum (cylindrica) and some specimens of Gyroporella annulata show remarkable similarity to Diplopora annulatissima . It is particularly interesting, as it concerns Upper Silesian specimens. J. (1931a, p. 271, 274), noted that, citing the above mentioned species by and stressing that in Silesian thin sections there are no segments with more than one whorl of pores and they never widen outwardly. To the contrary, it is visible that the pores are wide in the inner part and are thinning distally, exactly conforming to the characters of Diplopora annulatissima from Swiss Alps. This is even more pronounced in the fractured specimens from the Diplopora Dolomite
(Pls. XXVI ![]() -
XXVII
-
XXVII ![]() -
XXVIII
-
XXVIII ![]() -
XXIX
-
XXIX ![]() -
XXX
-
XXX ![]() ), and is also seen on 's specimens. Thus it seems obvious that the species in question should bear the name Diplopora cylindrica (). This is even more feasible, since the specimens of had not been lost, as presumed by (1931a, p. 271), but are housed in Munich, in Prof. 's collection
(fide & , 2000). It should be possible to choose from this collection a syntype most resembling the holotype of
(1920). It should be noted, that & in their synonymy of Kantia monregalensis omitted the work of
(1872a), and their listing begins with the paper by
(1919).
(1872a, p. 92-93) gave, however, a detailed and accurate description of Gyroporella cylindrica, illustrated with numerous drawings of Upper Silesian specimens (see Fig. 23
), and is also seen on 's specimens. Thus it seems obvious that the species in question should bear the name Diplopora cylindrica (). This is even more feasible, since the specimens of had not been lost, as presumed by (1931a, p. 271), but are housed in Munich, in Prof. 's collection
(fide & , 2000). It should be possible to choose from this collection a syntype most resembling the holotype of
(1920). It should be noted, that & in their synonymy of Kantia monregalensis omitted the work of
(1872a), and their listing begins with the paper by
(1919).
(1872a, p. 92-93) gave, however, a detailed and accurate description of Gyroporella cylindrica, illustrated with numerous drawings of Upper Silesian specimens (see Fig. 23 ![]() ). These comprehensive descriptions of
three-dimensionally preserved specimens seen in fractures are much more informative than descriptions of accidentally chosen sections. They provide insight into previously unknown structural details. Thus, 's priority is based not only upon formal issues, but also on the fact that his descriptions are more in-depth and better illustrated than many later works, and so are still a relevant source of data.
). These comprehensive descriptions of
three-dimensionally preserved specimens seen in fractures are much more informative than descriptions of accidentally chosen sections. They provide insight into previously unknown structural details. Thus, 's priority is based not only upon formal issues, but also on the fact that his descriptions are more in-depth and better illustrated than many later works, and so are still a relevant source of data.
However, according to the rules of the International Code of Botanical Nomenclature (2000, St Louis Edition), another solution is possible. The article 14 of the ICBN provides for conserving widely used names in the interest of nomenclatorical stability. The name Diplopora annulatissima has gained wide circulation since its establishing in 1920, and thus would be a good candidate for retention. The proposal of entering Diplopora annulatissima to the list of nomina conservanda should be approved by the International Committee on Botanical Nomenclature. Such a proposal could be submitted by the phycologists grouped in the specialist committee PETRALGA.
In the above state of affairs and, taking into account the serious formal and factual obstacles against strict application of the priority principle, I opt for conserving the name Diplopora annulatissima and its inclusion to the list of nomina conservanda.
Stratigraphical range and geographical distribution.- Diplopora annulatissima was described by (1920) upon the material collected by A. and F. (1912). In the Swiss Préalpes correctly labelled the specimens after 's descriptions as Diplopora cf. cylindrica. Locus et stratum typicum is Zweckenalpe bei Mythen, belonging to the uppermost Anisian. It is very common in the Roman Préalpes (Mont d'Or, St. Triphon), where it co-occurs with Physoporella praealpina, Ph. dissita, and Ph. minutula in the Upper Anisian (, 1961). Together with the above mentioned Physoporellae, D. annulatissima occurs also in Préalpes Médianes and in the whole Briançon Series of the French Alps. In the Vanoise Massif it defines, together with the Physoporellae the so-called "second diplopore zone" of the Upper Anisian (, 1958; , 1960). D. (1933), who discovered the diplopores in the Briançon Triassic, correctly suspected that some forms he classified as Diplopora annulata (, 1933, Pl. II, 13, 14), might actually belong to D. annulatissima . J. (1937a) listed numerous occurrences of D. annulatissima in localities of Western and Central Alps where it belongs, together with Ph. praealpina, to Illyrian. In the Eastern Alps, D. annulatissima occurs in the bottom part of the Wetterstein Dolomite, still in the Anisian.
Diplopora annulatissima has been reported from many localities within the Western Carpathians, both in southern (Gemerian) units of the Slovak Kras (, 1959, 1964), and in the Upper Sub-Tatric Nappe (Stražov Nappe) - , 1964, 1966), Middle Sub-Tatric Nappe (Choč - , 1967) and in the High-Tatric Series ( & , 1965; in & , 1965; in , 1965; , 1967, 1979, 1986). All these occurrences belong to the Upper Anisian. The species has been also found in Transylvania and many other places of the Romanian Carpathians (, 1969), where it also constitutes a separate zone in the uppermost Anisian. The species is however lacking from some areas of the Romanian Carpathians ( & , 1969) and Bulgaria ( & , 1973).
Diplopora annulatissima is also common in Southern Alps (,
1919; , 1937a; , 1965; ,
1972a; , 1997) and Dinaride Mountains (, 1957, 1965). According to (1965) and
(1974), the species belongs to the Upper Anisian and Lower Ladinian. even published a photograph of partially weathered specimen, where D. annulatissima
occurs with D. annulata and with ammonites (Fig. 23z ![]() ). If D. annulata is treated as a Ladinian index fossil, then D. annulatissima would be also of Ladinian age. However, (1965) agrees that D. annulatissima is present both in the Upper Anisian and in Ladinian, together with D. annulata.
(1926, 1930a, fig. 3) regarded D. annulatissima as an index fossil for the Upper Illyrian. J. (1982, 1986) assumed that both species could co-occur in the Illyrian and lowermost Fassanian. In the Upper Silesia, Diplopora annulatissima is an index fossil for Illyrian, and in the uppermost Illyrian it
occurs together with D. annulata, Kantia dolomitica, Kantia uniserialis and Diplopora uniserialis.
Illyrian age of both species is documented by the presence of upper Illyrian conodonts above the Diplopora Dolomite and the Tarnowice Beds (, 1975). Anisian age for the Diplopora Dolomite was assumed also by H.
(1974a, 1974b) and by E. and H.
(1993).
). If D. annulata is treated as a Ladinian index fossil, then D. annulatissima would be also of Ladinian age. However, (1965) agrees that D. annulatissima is present both in the Upper Anisian and in Ladinian, together with D. annulata.
(1926, 1930a, fig. 3) regarded D. annulatissima as an index fossil for the Upper Illyrian. J. (1982, 1986) assumed that both species could co-occur in the Illyrian and lowermost Fassanian. In the Upper Silesia, Diplopora annulatissima is an index fossil for Illyrian, and in the uppermost Illyrian it
occurs together with D. annulata, Kantia dolomitica, Kantia uniserialis and Diplopora uniserialis.
Illyrian age of both species is documented by the presence of upper Illyrian conodonts above the Diplopora Dolomite and the Tarnowice Beds (, 1975). Anisian age for the Diplopora Dolomite was assumed also by H.
(1974a, 1974b) and by E. and H.
(1993).
 |
|
|
Click on thumbnail to enlarge the image.
Figure 23: Selected previously published illustrations of specimens of Diplopora annulatissima from the Upper Silesia, Alps and Dinaride Mountains, similarly preserved as in the Upper Silesia: a-c - drawings of specimens from the Upper Silesian Muschelkalk (Segiet Forest near Tarnowskie Góry, locality no. 14) assigned by (1870, Pl. 11, 1, 2, 4) to Cylindrum annulatum (Nullipora annulata ): a - a piece of dolomite with numerous scattered specimens; besides Diplopora annulatissima , there are also representatives of Physoporella pauciforata (), Ph. prealpina , and Oligoporella elegans () , actual size; b - enlarged fractured specimen of Diplopora annulatissima with visible interior of the central cavity with single whorls of pores (branch inlets) and dense annulation, becoming more oblique apically; c - enlarged upper view upon a single annulus with numerous (24) branches; d-e - drawings of specimens from the Zugspitze Group in Bavarian Alps, assigned by (1872a) to Gyroporella annulata: d - finely annulated tube with protruding core of a central cavity (, 1872a, Pl. D.II, 1b), actual size; e - thallus with dense double whorls of small elongated tubercles and non-segmented apex (, 1872a, Pl. D.II, 1e), x 5; f-s - drawings of specimens from the Upper Silesian Jemielnica Dolomite assigned by (1872a) to Gyroporella cylindrica (=Diplopora annulatissima): f - algal accumulation in the rock (, 1872a, Pl. D.II, 2a), actual size; g - a single finely annulated thallus (, 1872a, Pl. D.II, 2aa), actual size; h - outer surface of a thalus with dense annulation (12 annuli separated with interannular furrows) covered with tubercles and pits, being possibly pore (branch) outlets (, 1872a, Pl. D.II, 2b), x 10; i - fractured thallus with outer annulation, with intusannulated wall of the central cavity; in vertical section wall visible are annuli with canals and interannular furrows; fissuration lists are slightly convex, with smooth surface (, 1872a, Pl. D.II, 2c), x 10; j - upper view upon a surface of a single annulus with rare long branches (22 canals) belonging to one row of a double whorl (, 1872a, Pl. D.II, 2d), x 10; k - upper view upon a surface of a single annulus with long thin branches (36 canals) belonging to both rows of a double whorl (, 1872a, Pl. D.II, 2e), x 10; l - smooth, slightly convex surface of a fissuration list (, 1872a, Pl. D.II, 2f), x 12; m - oblique view upon a fractured annulus with sections of branches and an interannular furrow; n - oblique-longitudinal fracture with segments containing double whorls of minute branches, separated with interannular furrows with locally preserved fissuration lists (, 1872a, Pl. D.II, 2h), x 10; o - longitudinal fracture of a specimen with preserved apex and eight segments consisting of double whorls of thin branches, perpendicular to the outer wall, and oblique at the apex; in some places the branches aggregate in tufts (, 1872a, Pl. D.II, 2i), x 10; p - accidental arrangement of one tube within another (, 1872a, Pl. D.II, 2m), x 2; r - apex of an alga with granulated outer surface (cortex?); the fracture reveals interior of the central cavity with rows of pores situated in furrows separated by flattened ridges (intusannulation) (, 1872a, Pl. D.II, 2k), x 10; s - slightly oblique longitudinal section of an alga; visible are annuli with two slightly oblique canals (branches) in each segment, separated with fissures, locally forming oval structures filled with sediment; in the lower part the fissures are separated from the segments with double whorls of tubercles by fissuration lists; annuli and fissures are connected by slightly undulating outer wall; a wide central cavity shows intusannulation (, 1872a, Pl. D.II, 2o), x 10; t - a diagrammatic representation of the structure of Gyroporella cylindrica (=Diplopora annulatissima) based upon studies of the Upper Silesian specimens (, 1872a, Pl. D.I, 8), x 30: c - canals (branches) passing through the algal wall; t - fissure of the inner wall of a central cavity; m - interannular suture on the inner wall of a central cavity; m' - pores (inlets of canals); x - probably encrustation swelling; z - interannular furrows with fissuration lists; u-w - drawings by (1878: Pl. VI, 10, 11a, 11b) being copies of figures by (1872a): u =k; v = l; w = e; x - schematic drawings by (1919, figs. 6, 6a, 6b, 6c) depicting her new species Kantia monregalensis from Villanova-Mondovi in Piemont Alps; numerous dense oblique annuli are separated by fissures; locally visible are double whorls of alternating pores; the branches supposedly widen outwardly, but the oversimplified drawing do not allow to see this well; thus the attribution of the specimens to the genus Kantia is poorly substantiated. Trichophorous branches have been observed in hundreds of specimens of Diplopora annulatissima , thus confirming the placement of this species in the genus Diplopora; y - specimens from Vanoise Massif in French Alps, etched with acetic acid (, 1958, Pl. 6, 20, 21), x 10, attributed by him to Diplopora cf. annulatissima ; in the lower specimen one can see dense annuli separated with outwardly thinning fissures, widened in the middle of the wall; pores are not well visible, but their traces are observable in the furrows of the intusannulated central cavity wall; the upper specimen is a fragmentary annulus with smooth fissuration list; z - weathered rock surface (from the Middle Triassic of Dinaride Mountains) with an ammonite and Diplopora annulatissima , showing dense annulation (from , 1965, Pl. VII, 1), actual size.
Clavapora , 1979
Clavapora clavaeformis (, 1920) , 1979
Pls. XXX ![]() -
XXXI
-
XXXI ![]() ;
Figs. 24
;
Figs. 24 ![]() - 25
- 25 ![]()
1920 Diplopora clavaeformis n.sp.- , Pl. IV, 17.
1935a Diplopora clavaeformis.- , p. 246, text-fig. 55.
1974 Diplopora clavaeformis.- , Pl. I, 1-4; Pl. II, 1-3; Pl. III, 1-5; Pl. IV, 1-3; Pl. V, 1-4.
1979 Clavapora clavaeformis n.gen. n.comb.- , p. 631, without figure.
1982 Diplopora clavaeformis.- , Pl. 1, 1-4; Pl. 4, 2 pars.
1986 Diplopora clavaeformis.- , Pl. IV, 3 (from , 1982; Pl. 1, 1).
2000 Clavapora clavaeformis.- & , p. 24-25.
Diagnosis.- Large, club-shaped and clearly annulated calcareous wall. Segments are oriented slightly upward and situated obliquely. The central cavity is very broad, occupying more than 2/3 of total diameter in the upper part of the plant. The trichophorous branches are arranged in whorls, each segment bearing one whorl. The whorls consist of a row of tufts formed of 4, exceptionally 5 branches, which are linked together by a common base. This species is similar to Diplopora annulatissima , from which it differs by larger dimensions, by the club-shaped top of the thallus and by larger furrows between two neighbouring segments.
Material and description.- The specimen from locality no. 50 Balin (Fig. 25 ![]() ) is preserved in similar manner as the specimen of B.
(1974, Pl. 1; Fig. 24
) is preserved in similar manner as the specimen of B.
(1974, Pl. 1; Fig. 24 ![]() herein) from the Mt Velebit (Dinaride Mts), preserved on the bottom bedding planes. The most characteristic is the club-shaped and clearly annulated skeleton. Dense annulation is observable in the lower part, where the segments with branches are separated by interannular furrows. The shape of branches is not clearly pronounced, as only the top of branches is observable, forming thin dense tubercles in two rows. The upper part of described specimen is enlarged, club-shaped. The central cavity forms main, very large stem, surrounded by widely spaced, slightly oblique segments separated by large interannular furrows. In the upper-right segments some branches are preserved, having trichophorous, slightly vesiculiferous shape. In this section tufts are not observable. In the uppermost part of the plant no segmentation is marked, but the top segment has the shape of calotte, as observed by
(1974).
herein) from the Mt Velebit (Dinaride Mts), preserved on the bottom bedding planes. The most characteristic is the club-shaped and clearly annulated skeleton. Dense annulation is observable in the lower part, where the segments with branches are separated by interannular furrows. The shape of branches is not clearly pronounced, as only the top of branches is observable, forming thin dense tubercles in two rows. The upper part of described specimen is enlarged, club-shaped. The central cavity forms main, very large stem, surrounded by widely spaced, slightly oblique segments separated by large interannular furrows. In the upper-right segments some branches are preserved, having trichophorous, slightly vesiculiferous shape. In this section tufts are not observable. In the uppermost part of the plant no segmentation is marked, but the top segment has the shape of calotte, as observed by
(1974).
Completely different is the state of preservation of the specimens from locality no 25 Przełajka
(Pls. XXX, 5-17 ![]() &
XXXI, 1-4
&
XXXI, 1-4 ![]() ). The fractures are passing through the central cavity, showing the internal wall from inside. The internal wall shows frequently delicate intusannulation, composed of shallow furrows separated by low ridges. Furrows are covered by numerous alternating pores, forming two or more rows. Pores reach up to the sides of ridges, which in effect of
alternation possess wavy course (Pls. XXX, 9, 14-15
). The fractures are passing through the central cavity, showing the internal wall from inside. The internal wall shows frequently delicate intusannulation, composed of shallow furrows separated by low ridges. Furrows are covered by numerous alternating pores, forming two or more rows. Pores reach up to the sides of ridges, which in effect of
alternation possess wavy course (Pls. XXX, 9, 14-15 ![]() &
XXXI, 1-2, 4
&
XXXI, 1-2, 4 ![]() ). Intusannulation is not always present and in this cases pores are rather irregular
(Pl. XXXI, 3
). Intusannulation is not always present and in this cases pores are rather irregular
(Pl. XXXI, 3 ![]() ). This feature may be observed on some fractures of 's (1974, Pl. I,
1-2, text-figure 14a, d) specimens. The large central cavity is surrounded by dense segmentation, reaching to the internal wall
(Pls. XXX, 9-13, 16-17
). This feature may be observed on some fractures of 's (1974, Pl. I,
1-2, text-figure 14a, d) specimens. The large central cavity is surrounded by dense segmentation, reaching to the internal wall
(Pls. XXX, 9-13, 16-17 ![]() &
XXXI, 1-3
&
XXXI, 1-3 ![]() ). In some segments trichophorous branches are preserved, grouped in tufts
(Pls. XXX, 14
). In some segments trichophorous branches are preserved, grouped in tufts
(Pls. XXX, 14 ![]() &
XXXI, 4
&
XXXI, 4 ![]() ). Interannular furrows are limited by wavy lists
(Pls. XXX, 7, 12-13
). Interannular furrows are limited by wavy lists
(Pls. XXX, 7, 12-13 ![]() &
XXXI, 1-2
&
XXXI, 1-2 ![]() ). Some specimens are club-shaped
(Pls. XXX, 6, 12
). Some specimens are club-shaped
(Pls. XXX, 6, 12 ![]() &
XXXI, 3
&
XXXI, 3 ![]() ). No segmentation is present on the top of the plant, where according to
's (1974) observation, a large segment forming calotte may be seen.
). No segmentation is present on the top of the plant, where according to
's (1974) observation, a large segment forming calotte may be seen.
Stratigraphical range and geographical distribution.- The holotype of Diplopora clavaeformis was described by (1920) from the vicinity of Han Bulog in Bosnia from the uppermost Anisian Trinodosus Limestone (, 1942). His description was based on a single specimen and was not very detailed. The second locality is area of Gračac, on the northeastern slopes of Mt. Velebit. The full description is based on many perfectly preserved thin sections (, 1974), belonging to Lower Ladinian (co-occurence with Diplopora annulata). Diplopora clavaeformis was also found by (1982, Pl. 1, 1-4; Pl. 4, 2 pars) in the Stražov nappe (Slovakia) in the lower part of Wetterstein Limestone. It occurs together with D. annulatissima , D. annulata and Kantia comelicana (), belonging to the Diplopora annulatissima Zone (, 1986, p. 303-306, Pl. IV, 3).
T. (1979) proposed erecting a new genus Clavapora sp., based on the Diplopora clavaeformis , for club-shaped strongly annulated specimens with trichophorous branches arranged in whorls, and grouped in tufts. This concept was accepted by S. and M.J. (1992) and by B. and T. (2000). In the Upper Silesia, Clavapora clavaeformis () was found in numerous fractures in the Diplopora Dolomite (Illyrian) from the locality Przełajka (no. 25), together with Physoporellae, Oligoporellae, and Diplopora annulatissima (see Fig. 5 ![]() ). The second locality, also of Illyrian age, is Mały Balin (no. 50), where Clavapora clavaeformis was found in a thin section. The accompanying Dasycladales are: Diplopora annulatissima, Kantia comelicana, Physoporella polonoandalusica, Physoporella praealpina, Physoporella lotharingica, Physoporella pauciforata, Oligoporella chrzanowensis, Oligoporella balinensis, Oligoporella silesiaca and Oligoporella elegans.
). The second locality, also of Illyrian age, is Mały Balin (no. 50), where Clavapora clavaeformis was found in a thin section. The accompanying Dasycladales are: Diplopora annulatissima, Kantia comelicana, Physoporella polonoandalusica, Physoporella praealpina, Physoporella lotharingica, Physoporella pauciforata, Oligoporella chrzanowensis, Oligoporella balinensis, Oligoporella silesiaca and Oligoporella elegans.
Family Acetabulariaceae () , 1885
[Polyphysaceae (, 1841)]
Tribe Acetabularieae , 1842
Acicularia d', 1843
General remarks.- In thin sections of the Triassic sediments from the Upper Silesia, there are tiny round or oval cross-sections (0.3-0.5 mm across), with regular pores usually distributed along the edge of the cross section. Such cross sections were also found in Triassic strata of the Alps and Carpathians. They were named Acicularia sp. (, 1955; , 1964; , 1967, 1973, 1979, 1981, 1994a; & , 1969; & , 1971; -, 1975; et al., 1998) or Aciculella (, 1930b; & , 1974, 1975; , 1975; et al., 1980; et al., 1994; -, 1975). Perhaps using two generic designations has sometimes resulted from inadvertent confusion of the two similar names (, 1974, p. 43). However, the main reason is the diametrally different understanding and interpretation of the cross sections visible in thin sections. The authors using the name Acicularia refer to the Cainozoic and Recent genus of the same name, represented mostly by its reproductive organs (an organo-species), and to Recent species Acetabularia mediterranea. In this case, the observed cross sections would represent lime spicules of the cap rays with gametangia ( & , 1971, p. 85, fig. 3; & , 1992, p. 127, 157, fig. 2.9e-f; , 1994, Pl. 4, 3). On the other hand, the authors preferring the name Aciculella regard the observed structures as cross-sections of the whole algal stem with sporangia (, 1930b, Pl. 4; , 1971; & , 1974, fig. 3; , 1975, Pl. I-III; & , 1992, fig. 2.9g). In this case, it would be a form-species (, 1971) or paragenus ("Sammelgattung" - , 1958, p. 103, 105) representing in fact several biological species (, 1975). It is difficult to find characters differing Aciculella from Acicularia. (1930a, p. 160) stated that Aciculella differs from the spicules of Acicularia by having one end sharply pointed. However, all Recent Acetabulariaceae have pointed one (inner) end of their spicules, while the other one is rounded.
If the observed structures are sections of the whole algal stem, the central cavity should be visible. It is in fact so in several sections described by (1975, Pl. III, 6-9) and assigned by him to Aciculella. In most cases, however, there is no central cavity in the sections (see, e.g., & , 1974, Pl. I), thus indicating that they pertain to the spicules, not to the whole algae. Also the small size of the structures (within fraction of a millimeter range) supports this interpretation. The Recent Acicularia is morphologically identical to Acetabularia: "The striking difference lays in the fact that the gametangia in Acetabularia are completely free and not calcified, while the gametangia in the genus Acicularia are embedded into a coherent calcareous structure. This structure including the gametangia might be freed of the gametophore as one unit. The cysts can be easily detached from their bed, leaving cavities" ( & , 1992, p. 157). That is the reason why Recent Acicularia specimens are seldom found intact, but rather occur as separate lime spicules (gametophores) of the cap rays. Whole fossil algae are never found; only the spicules called Acicularia are present in the fossil record.
Assuming such interpretation, the structures seen in thin sections would belong to choristophore Dasycladales, with special reproductive organs (ray-shaped gametophores), such as in the Recent umbrellophore Acetabulariaceae. If, however, the sections would represent whole stems of Aciculella, they would belong to endospore Dasycladales (with sporangia inside their thallus). The endospore species are believed to be more primitive than the choristophore Dasycladales. The endospore species are known in the Triassic (Diplopora phanerospora , D. muranica , D. praecursor , and Teutloporella herculea () .- unpublished data by the author). However, umbrellospore species are known in the Permian (Clypeina; Permian-Paleogene), Carboniferous (Atractyliopsis, Coelosporella.- , 1986), and even in the Devonian (Paradella.- & , 1971). Therefore, there are no reasons to preclude the existence of umbrellospore Dasycladales, such as Acicularia, also in the Triassic, thus not appearing only in the Jurassic ( & , 1992, p. 42).
Stratigraphical range and geographical distribution.- Acicularia has been up to now known since Jurassic till Recent ( & , 1992). However, paragenus Aciculella (, 1930a; , 1958; & , 1974, 1975; , 1975), is virtually indistinguishable from the Triassic Acicularia (, 1964; , 1967, 1973, 1979, 1981, 1994a). The genus Acicularia has no importance for stratigraphy. In the Upper Silesia it is know from the Górażdże Beds ( & , 1974) and Karchowice Beds (, 1967, 1973, 1979, 1981, 1994a), of Pelsonian age (boreholes 94 WB, 91 WB, and Odra IG1). It occurs also in the Diplopora Dolomite in the following localities: Srebrna Góra, no. 13; Wojkowice Komorne, no. 30; and Trzebionka, no. 53 (Pelsonian-Illyrian). It is probably a fairly common genus, but is not visible on fractured surfaces, and recognizable only in thin sections. In Poland, besides the Upper Silesia and the Zawiercie area, it is also known from the Holy Cross Mountains (Zajączków, Łukowa Beds, Pelsonian - & , 1974).
 |
|
|
Click on thumbnail to enlarge the image.
Figure 24: Selected previously published (, 1974, Pl. I) illustrations of specimens of Diplopora clavaeformis from the Dinaride Mountains (Mt Velebit), similarly preserved as new specimens from the Upper Silesia, x 2.5.
 |
|
|
Click on thumbnail to enlarge the image.
Figure 25: Fractured specimen of Clavapora clavaeformis () from Balin (locality no. 51), x 2.5.
Special thanks are due to Witold , M.Sc. (PGI), who for years participated in my field research. The study was done in the Polish Geological Institute, Warsaw; I am grateful to the Director of the Geological Museum, PGI, Dr. Włodzimierz , for enabling me to produce numerous photographs and thin sections in the museum labs, and for his help in the transfer of the Dasyclad collection consisting of rock samples and thin sections to the museum archival storage. Mr. Karol , M.Sc. (PGI), kindly translated the paper to English and offered many valuable editorial comments. I am obliged to the late Prof. E. for access to the collection of J. von at the Naturhistorisches Museum Wien, and for sending me copies of rare publications. I would like to express my thanks also to Prof. Bruno , leader of the PETRALGA Project, for inviting me to participate in this joint effort.
This publication is a contribution to the PETRALGA Project http://paleopolis.rediris.es/petralga/
J. (1906).- Die Trias im südlichen Oberschlesien.- Königl. Preuß. Geol. Landesanst. u. Bergakad., Abh., Berlin, 50: 1-163.
A. von (1878).- O galicyjskich gatunkach skamieniałych otwornic rodzaju Gyroporella Rozprawy i Sprawozdania z posiedzeń Wydz. mat.-przyr., III Akad. Umiejętności w Krakowie, V: 71-112.
D. (1938).- Sur quelques fossiles triasiques des Carpathes occidentales.- Vestník král. česk. společnosti nauk, Tř. matemat.-přirodovęd., Praha, R. 1937, 20 p.
D. (1959).- Geologia československých Karpát.- Vyd. Slov. Akad. Vied, Bratislava, II.
E.J. d' (1843).- Description géologique du département de l'Aisne.- Mém. de la Soc. géol. de France, Paris, 1 sér., V, 3: 129-418.
P. (1913).- Beitrag zur Kenntniss der Stratigraphie der oberschlesischen Muschelkalks.- Jb. der Preuß. Geol. Landesanst., Berlin, 34, 1: 268-340.
P. (1914).- Geologischen Karte von Preussen und banachbarten Bundesstaaten landern - Blatt Broslawitz [Zbrosławice] - Geologish-agronomisch bearbeitet durch P. ; Erläutert durch P. ; mit einem agronomischen Teil von W. (, 1914).- Königlich Preussischen Geologischen Landesanstalt; A. W. Schade Buchdruckerei, Berlin. 62 pp.
P. (1926a).- Die Fauna der Wirbellosen und die Diploporen der oberschlesischen Trias mit Ausnahme der Brachiopoden, Lamellibranchiaten, Gastropoden und Korallen.- Jb. der Preuß. Geol. Landesanst., Berlin, 46 (1925): 504-527.
P. (1926b).- Die Tiefbohrung "Oppeln".- Jb. der Preuß. Geol. Landesanst., Berlin, 46: 373 - 395.
P. (1944).- Die Stratigraphie der oberschlesischen Trias. Teil 2: Der Muschelkalk.- Abh. d. Reichsamt fűr Bodenforschung (Berlin), N.F. 208: 1-124.
P. & R. (1932).- Geologische Karte von Preussen und banachbarten deutschen landern - Blatt Leschnitz [Leśnica] Geologish-agronomisch aufgenommen von P. & R. ; erlautert von P. .- Preussischen Geologischen Landesanstalt, Berlin, 52 pp.
P. & J. (1943).- Ubersichtskarte des vortertiären Untergrundes im Oberschlesischen Triasgebiet.- Reichsamt fur Bodenforschung. Berlin
P., W. & R. (1914).- Geologischen Karte von Preussen und banachbarten Bundesstaaten landern - Blatt Zabrze. Aufgenommen und erlautert dur , & (, & , 1914).- Koniglich Preussischen Geologischen Landesanstalt, A. W. Schade Buchdruckerei, Berlin,103 pp.
M. & K. (1965).- Budowa fałdu Czerwonych Wierchów na wschodnim zboczu Doliny Kościeliskiej w Tatrach.- Acta Geol. Pol., 15, 3: 350-352.
A. (1919).- Contributio allo studio delle Siphoneae verticillatae del calcare di Villanova-Mondovi.- Atti Soc. Ital. Sc. Natur., 58: 216-236, Pavia.
J.-P., P., R., P., M., A.F. & G. (1975).- Réflexions sur la systématique des Dasycladales fossiles.- Géobios, 8: 259-290.
J.-P., P., R., P., M., A.F. & G. (1977).- Classification criteria of fossil Dasycladales. In: E. (ed.), Fossil Algae.- Springer, Berlin: 154-166.
J.-P., P., R. & D. (1979).- Essai de classification des Dasycladales en tribus. In: 2čme Symposium international sur les Algues fossiles (Paris, 23-26 avril 1979).- Bull. des Centres de Rech. Expl.-Prod. Elf-Aquitaine, Pau, 3, 2: 429-442.
T. & R. (1970).- Das Anis. zwischen St. Vireil und dem Hölensteintal (Pragser und Olanger Dolomite, Sűdtirol).- Festband d. Geol. Inst., 300-Jahr-Feier Univ. Innsbruck: 9-109.
E.W. (1898).- Diplopora und einige andere Versteinerungen im elsass-lothringischen Muschelkalk.- Mitt. Geol. Landesanst. V. Elsass-Lothringen, Strassburg, 4: 277-285.
S. & M.J. (1992).- Dasycladales. An illustrated monograph of a fascinating algal order.- Thieme, Stuttgart, New York: 247 pp.
A. & J. (1964).- Die Dasycladazeen in der Trias der Westkarpaten.- Geol. Sbor., 15, 2: 173-188.
G. (1955).- Szczegółowa Mapa Geologiczna Polski w skali 1:50,000, ark. Pyskowice.- IG, Warszawa
G. & M. (1956).- Szczegółowa Mapa Geologiczna Polski w skali 1:50,000, ark. Katowice.- IG, Warszawa
M., C. & Ş. (1972).- Contribuţii la biostratigrafia depozitelor triasice din Platoul Vaşcău (Munţii Apuseni).- Dări de seamă ale şedinţelor, Bucureşti, 58 (1971), 3. Paleontologie: 5-25.
J.-P. & R. (1969).- Présence de Diplopora annulata dans le Ladinien de la zone briançonnaise des Alpes cottiennes méridionales (Alpes occidentales franco-italiennes).- C.R. Acad. Sci., Paris, (sér. D), 269: 1166-1168.
A. (1989).- Biostratinomy and sedimentary environment of the echinoderm-sponge biostromes in the Karchowice Beds, Middle Triassic of Upper Silesia.- Ann. Soc. Geol. Pol., 59, 3-4: 331-350.
A. (1990).- Unit IV (Karchowice Beds) Regression stage. In: International Workshop - Field Seminar the Muschelkalk - Sedimentary Environments, Facies and Diagenesis.- Excursion Guidebook and Abstracts.- Kraków-Opole.
A. (1997).- Formacja karchowicka: definicja i stratygrafia.- Geologos, 2: 165-199.
K., S., C. & P. (1975).- Origin of the ore-bearing dolomite in the Triassic of the Cracow-Silesian Pb-Zn ore district.- Rocz. Pol. Tow. Geol. 45: 139-155.
K. (1907).- Materialy dla izuchenia rakovinnogo izvestnaka Dombrowskogo bassieyna.- Trudy Geol. Komiteta. N.S. Vyp. 35: 1-99, Petersburg.
G. (1961).- Étude géologique de la région du Mont d'Or (Préalpes romandes).- Eclog. geol. Helvet., 54, 1: 29-106.
I.I. (1997).- Révision de la variété Pilosa ex , 1964, de l'espèce Oligoporella pilosa , 1912, une algue Dasycladacée de l'Anisien.- Rev. Paléobiol., 16, 1: 181-186.
I.I., C. & D. (1994).- Middle Triassic carbonate deposits and calcareous algae from the Sasca zone (Southern Carpathians, Romania).- Facies, 30: 85-100.
J. (1957).- Príspevok ku poznaniu diplopór triasu gemeríd [Beitrag zur Kenntnis der Diploporen der Gemeriden-Trias].- Geol. Sbor., 8, 2: 226-241.
J. (1959).- Výskyt riasy Diplopora annulatissima v Slovenskom krase [Vorkommen der Alge Diplopora annulatissima im Gebirge Slovenský kras].- Geol. pr., Bratislava, 16: 177-182.
J. (1962).- Nové Dasycladaceae triasu Slovenského krasu [New Dasycladaceae of the Triassic in the Slovak Karst].- Geol. Sbor., 13, 2: 227-240.
J. (1964).- Slovenský kras. Stratigrafia a Dasycladaceae mezozoika Slovenského krasu [Stratigraphie und Dasycladaceen des Gebirges Slovensky kras].- Ústredný ústav Geologický, Bratislava. 204 pp.
J. (1966).- La stratigraphie et les Dasycladacées du Trias moyen de la série du Drienok.- Geol. Sbor.- Geol. carpath., 17, 2: 241-257.
J. (1967).- Die obertriadischen Dasycladazeen der Westkarpaten. Geol. Sbor.- Geol. Carpath., 18, 2: 285-309.
J. (1975).- Genus Aciculella , 1930 (Kalkalgen) in der Trias der Westkarpaten.- Geol. Sbor.- Geol. Carpath., 36, 1: 1-20.
J. (1982).- Attribution to the Strážovská hornatina Mts. Triassic reef complex stratigraphy on the basis of Dasycladaceae (the West Carpathians Mts., Slovakia).- Geol. Sbor.- Geol. Carpath., 33, 1: 79-88.
J. (1983).- Physoporella croatica , 1958, of the Slovak Karst Anisian (Slovakia, the West Carpathians Mts.).- Geol. Sbor.- Geol. carpath., Bratislava, 34, 5: 507-533.
J. (1986).- Stratigraphic ranging and zonation of Dasycladal algae in the West Carpathians Mts., Triassic.- Mineral. Slov., Bratislava, 18, 4: 289-321.
J. & J. (1965).- Triadische Dasycladaceen in der hochtatrischen Hüllenserie der Hohen Tatra.- Geol. Sbor., 16, 1: 11-18.
A. (1955).- Une nouvelle espèce de "Clypeina" (Dasycladacée) dans le Berriasien du Salève et du Jura central.- Archiv. Sci. (Genève) 9, 4: 471-477.
R. (1938).- Geologischen Karte von Preussen und banachbarten deutschen landern - Blatt Jakobswalde [Kotlarnia]. Aufgenommen von R. ; mit einem beitrag von K. Ihnen (R. , 1938).- Preussischen Geologischen Landesanstalt, Otto Meusel Buchdruckerei und Verlag, Berlin, 30 pp.
R., P. & W. (1938).- Geologischen Karte von Preussen und banachbarten deutschen landern - Blatt Peiskretscham [Pyskowice]. Aufgenommen von R. , P. und W. ; erläutert von R. ; mit einem beitrag von K. (R. , 1938).- Preussischen Geologischen Landesanstalt, Otto Meusel Buchdruckerei und Verlag, Berlin, 53 pp.
P. (1979).- Osservazioni su Diplopora nodosa , 1863 (alga verde Dasicladacea) delle scogliere triassiche della Lucania (Appennino meridionale).- Boll. Soc. Nat. Napoli, 88: 1-91.
J. (1960).- Comparaison du Trias haut-tatrique avec celui des Alpes occidentales (Zone intra-alpine). Acta Geol. Pol. 10, 2: 107-114.
J. & M. (1963).- État actuel de nos connaissances sur la stratigraphie du Trias dans le Briançonnais sensu stricto.- Mém. Bureau Rech. Géol. Min., 15: 232-242.
R. (1988).- Nouvelle taxonomie des Algues Dasycladales.- Bull. Centr. Rech. Expl.-Prod. Elf-Aquitaine, 12, 1: 165-217.
G. & J. (1963).- Flores triasiques de France.- Mém. Bureau Rech. Géol. Min., 15: 507-523.
J., L.E., S., G., H., J., M., M., A. & P. (1990).- Anisian to Oligocene paleogeography of the European margin of Tethys (Geneva to Baku).- Mém. Soc. Géol. France, N.S. 154, 3: 159-190.
J., L.E. & B. (1993).- Atlas Tethys palaeoenvironmental maps.- Atlas and Explanatory Notes, Gauthier Villars Ed., CGMW Paris, 307 p.
M. & O. (1969).- Triassic calcareous algae from the Apuseni Mountains (Romania).- Rev. Palaeobot. Palynol., Amsterdam, 9, 1-2: 63-101.
S. (1935).- Arkusz Grodziec.- PIG, Warszawa
S. & B. (1968).- Mapa geologiczna Górnośląskiego Zagłębia Węglowego w skali 1:100,000, bez utworów młodszych od triasu.- Warszawa.
O. (1969).- Algues calcaires du Jurassique supérieur et du Crétacé inférieur de Roumanie.- Rev. Micropal. 12: 53-62.
O. (1981).- Mesozoic Dasycladaceae from Romania: Distribution and biostratigraphical importance.- Facies, 4: 165-196.
O., M., E. & R. (1980).- Biostratigraphy of the Triassic formations in the east of the Pădurea Craialui Mountains.- D. S. Inst. geol. geofiz. 64, 4: 29-61.
O. & E. (1975).- Asupra unor alge, foraminifere, sphinctozoare şi microproblematice din Triasicul din Carpatüii Orientali şi Dobrogea de nord [Some Triassic Algae, Foraminiferae, Sphinctozoa and Microproblematicae from the Eastern Carpathians and North Dobrogea ].- Studii Cercet. Geol. Geofiz., Geogr., ser. Geol., Bucureşti, 20, 2: 247-254.
W. (1990).- Ocena zasięgu genezy dolomitu kruszconośnego w rejonie Żarki - Zachód. CPBR nr 1. 8 "Budowa Geologiczna Polski i Poszukiwania Złóż Surowców Mineralnych" Cel nr 11, Wyjaśnienie zasięgu i genezy dolomityzacji formacji mezozoicznych i młodopaleozoicznych w rejonie Żarki Zachód.- CAG Warszawa nr 503/91, Archiwum PIG.
W. (1995).- Geologiczne przesłanki poszukiwań czystych odmian dolomitów w dewonie i triasie.- CAG Warszawa nr 530, Archiwum PIG.
S. & R. (1960).- Source of the Lower Triassic clastics in the Tatra Mts.- Bull. Acad. Pol. Sc. ser. geol.-geogr., 8, 1.
H. (1862).- Nullipora annulata im Muschelkalk Oberschlesiens.- Zeitschr. d. Deutsch. Geol. Gesellsch. Bd 14. Berlin.
H. (1863).- Über den Opatowitzer Kalkstein des Oberschlesischen Muschelkalkes.- Zeitschr. d. Deutsch. Geol. Gesellsch. Bd 15. Berlin.
F. (1958).- Étude géologique du pays de Vanoise.- Mémoires pour servir à l'explication de la Carte géologique détaillée de la France, Paris. 561 pp.
F. (1963).- Trias à faciès briançonnais de la Vanoise et des Alpes occidentales.- Mém. Bur. Rech. Géol. Min., 15: 215-231.
G.F. (1971).- The nature of Aciculella (calcareous algae).- Palaeontology, London, 14, 4: 629-636.
P., L. & M. (1954).- Observations sur le Trias bétique et ses Algues calcaires.- Mem. Com. Inst. Geol. prov., Barcelona, 11.
E. (1986).- Zur Mikrofazies oberanisischer Riffkalke in den östlichen Nordkarawanken, Kärnten.- Carinthia II, Klagenfurt, 96, 176: 463-478.
E. & H. (1993).- Dasycladaceen aus dem Oberen Muschelkalk (Mitteltrias) des Hohenloher Landes, Süddeutschland.- Zitteliana (München), 20: 93-103.
E., A. & I.I. (1994).- Middle Triassic (Anisian) limestones from Bled, Northwestern Slovenia: microfacies and microfossils.- Geologija, Ljubljana, 36 (1993): 157-181.
E. (1979).- A new Dasycladacean (calcareous algae) assemblage from Triassic of M. Popera (Belluno, Italy).- Riv. Ital. Paleont. Strat., 85, 1: 57-84.
E. & F. (1983).- La dorsale paleocarnica anisica di Pontebba.- Riv. Ital. Paleont. Stratigr., 89, 1: 3-30.
M. & M. (1989).- The Anisian (Middle Triassic) carbonate bank of Camorelli (Lombardy, southern Alps).- Facies. 21: 41-56.
A. & W.R. (1974).- Green algae Aciculella from the Muschelkalk of the Holy Cross Mts. (Poland).- Bull. de l'Acad. Pol. des Sci, Ser. Sciences de la Terre, 22, 1: 27-32.
A. & W.R. (1975).- Revision of fossil sporangia Aciculella and Acicularia (Dasycladaceae) from the Mesozoic of Poland.- First International Symposium on Fossil Algae (Erlangen, 6-10 Oct 1975), Program and Abstracts: 12-13.
E. jr (1958).- Ein Beitrag zur Stratigraphie der südlichen Klippendecke im Gebiet Spillgerten-Seehorn (Berner Oberland).- Eclog. Geol. Helvet., 51, 1: 151-211.
S., S. & J. (1988).- Pozycja "skałek" jurajskich Kruhela Wielkiego w profilu formacji z Ropianki (fm). In: J. & K.(eds.), Przewodnik 59. Zjazdu PTG.- Karpaty Przemyskie Kraków: 266-273.
A. (1979).- The stratigraphic distribution of fossil calcareous algae in southern Iran.- Bull. Centr. Rech. Explor.-Prod. Elf-Aquitaine, 3: 619-624.
B. (2012).- The contribution of calcareous green algae to the production of limestones: a review. In: D. & B. (eds.), Calcareous algae and the global change: from identification to quantification.- Geodiversitas, 34, 1: 35-60.
B. & R. (1994).- Inventaire critique des Algues Dasycladales fossiles. II partie - Les Algues Dasycladales du Jurassique et du Crétacé.- Rev. Paléobiol., 12, 1 (1993): 19-65.
B. & R. (1995).- Inventaire critique des Algues Dasycladales fossiles. 3. Les Algues Dasycladales du Permien et du Trias.- Rev. Paléobiol., 14, 1: 49-84.
B. & T. (2000).- Les Algues Dasycladales du Permien et du Trias. Nouvelle tentative d'inventaire bibliographique, géographique et stratigraphique [Permian and Triassic Dasyclad Algae Bibliographic, Geographic, and Stratigraphic Reappraisal].- Geol. Croat., 53, 1: 1-197.
B. & N.J. (2013).- The XXIst Century (the 100th Anniversary) Edition of the "New studies on Triassic Siphoneae verticillatae" by Julius von .- Carnets de Géologie [Notebooks on Geology], Brest, Book 2013/01 (CG2013_B01), 108 pp.
C.W. (1872a).- Die sogenannten Nulliporen (Lithothamnium und Dactylopora) und ihre Betheiligung an der Zusammensetzung der Kalkgesteine. Zweiter Theil: Die Nulliporen des Thierreichs (Dactyloporideae) nebst Nachtrag zum ersten Theile.- Abh. II. Cl. kön. bayer. Akad. Wiss., Mathem. phys. Cl., München, 11, 1: 231-290.
C.W. (1872b).- Ueber die Dactyloporen-ähnlichen Fossilien der Trias.- Vehr. k. k. geol. Reichsanstalt. Sitzung am 5 März 1872.
C.W. (1873).- Ueber Conodictyum bursiforme , einer Foraminifere aus der Gruppe der Dactyloporideen.- Sitzungsberichten kön. bayer. Akad. Wiss., Sitzung der mathem.-phys. Classe, München, 3: 282-294.
C.W. (1874).- Ueber neue Gyroporellen aus den Gailthaler Gebirge.- Verh. k.-k. geol. Reichsanstalt, Wien, 1874, 4: 79-80.
T. (1979).- Dasycladacées métaspondyles du Paléozoďque supérieur et du Trias. In: 2me Symposium international sur les Algues fossiles (Paris, 23-26 avril 1979).- Bull. Centr. Rech. Explor.-Prod. Elf-Aquitaine, Pau, 3, 2: 625-637.
H. (1985).- Immigration of crinoids into the German Muschelkalk Basin. In: U. & A. (eds.), Sedimentary and Evolutionary Cycles.- Lect. Not. Earth Sc., 1: 237-254.
H. (ed., in cooperation with T. & J.) (1991).- Muschelkalk. A Field Guide.- Korb Verlag (Goldschneck), Stuttgart, 80 pp.
H. & E. (1993).- Palaeobiogeography and stratigraphy of Muschelkalk echinoderms (Crinoidea, Echinoidea) in Upper Silesia. In: H. & A. (eds.), Muschelkalk. Schöntaler Symp. 1991: 165-176. Goldschneck, Stuttgart.
M. (1957).- Some problems in the Dasyclad genus Diplopora-. Micropaleontology, 3, 1: 49-52.
M. (1958).- The Dasyclad genus Physoporella in the Anisian of Yugoslavia.- J. Palaeont. Soc. India, Lucknow, Birbal Sahni Mem. No., 3: 59-63.
M. (1963).- Trias de Yugoslavie.- Mém. Bureau Rech. Géol. Min. 15: 477-494.
M. (1965).- Comparative study of some Triassic Dasycladaceae in Yugoslavia [Komparativni studij nekih trijaskih dasikladaceja u Jugoslaviji].- Geol. vjesnik, Zagreb, 18, 1 (1964): 3-34.
M. (1967).- Mesozoische Kalkalgen der Insel Chios (Griechenland).- N. Jb. Geol. Paläont, Abh., Stuttgart, 129, 1: 97-104.
V. (1913).- Beiträge zur Kenntnis des mittleren Muschelkalks und des unteren Trochitenkalks am östlichen Schwarzwaldrand.- Geol. Palaeont. Abh., Jena, N.F., 12, 2, 1-100.
L. & C. (1866).- Geognostische Karte des ehemaliges Gebietes von Krakau mit den südlich angrenzenden Theile von Galizien.- Erläuterungen: Denkschr. math.-naturwiss. Cl. Wiener Akad. Wiss. 26.
H. (1967).- Variationsstatistische Untersuchungen an anisischen Dasycladaceen aus den Pragser Dolomiten in Südtirol.- N. Jb. Geol. Paläont, Abh., 128, 1: 41-100.
H. (1969).- Umbildungstendenzen der Astformen in Physoporella-Oligoporella-Populationen (Dasycladaceen) aus dem Anis der Pragser Dolomiten (Italien).- N. Jb. Geol. Paläont., Mh., 1969, 2: 104-120.
H. & R. (1971).- Diplopora praecursor, a Mesozoic calcareous alga (Dasycladaceae) with endospore cysts.- Nova Hedwigia, 21: 505-519. Verlag von J. .
A., R. & V. (1998).- A new Dasycladalean alga from the Middle Triassic carbonates of northern Calabria (southern Italy).- Facies, 39: 67-74.
ICBN (1961).- International Code of Botanical Nomenclature, adopted by the Ninth International Botanical Congress, Montreal, August 1959. In: J. et al. (eds.), Regnum Vegetabile 23. Koeltz Scientific Books, Königstein.
ICBN (2000).- International Code of Botanical Nomenclature (St-Louis Code) adopted by the Sixteenth International Botanical Congress St Louis, Missouri, July-August 1999. In: W. et al. (eds.), Regnum Vegetabile 138. Koeltz Scientific Books, Königstein.
A. (1998).- Middle Muschelkalk palaegeography, In: R., S. & J. (eds.), Atlas paleogeograficzny epikontynentalnego permu i mezozoiku w Polsce 1:2,500,000 [Palaeogeographical Atlas of the Epicontinental Permian and Mesozoic in Poland 1:2,500,000].- Państwowy Instytut Geologiczny, Warszawa.
A. (1912-1913).- Monographie géologique des Tours d'Aď et des régions avoisinantes (Préalpes vaudoises).- Mat. Carte géol. Suisse, 34: 466 pp.
A. & F. (1912).- Le Trias du bord radical des Préalpes médianes entre le Rhône et l'Aar.- Ecl. Geol. Helvet., 11: 739-747.
E. (1958).- Über das System und die Stammesgeschichte der Dasycladacean (Siphoneae verticillatae).- Ann. Naturh. Mus. Wien, 62: 95-122.
H. (1978).- Mapa geologiczna Polski w skali 1:200,000 ark. B bez utworów czwartorzędowych, ark. Kraków.- Wyd. Geol. Warszawa.
M. (1969).- Lithostratigraphische und sedimentologische Untersuchungen in der kalkalpinen Mitteltrias (Anisian und Ladinian) des Rätikon (Österreich und Fürstentum Liechtenstein).- Mitteil. Geol. Inst. Eidgenöss. Tech. Hochschule Univ. Zürich, N.F., 118, 1-149.
V. & I. (1971).- Evolutions-Tendenzen der Dasycladaceen mit besonderer Berücksichtigung neuer Funde in Jugoslawien.- Palaont. Z., 45, 1-2: 82-91.
V. & J. (1974).- Übersicht über den gegenwärtigen Stand der Biostratigraphie der Trias der Westkarpaten. In: H. (ed.), Die Stratigraphie der alpin-mediterranen Trias [The stratigraphy of the Alpine-Mediterranean Triassic ] Symposium (Wien, 1973).- Öster. Akad. Wiss., Schrift. Erdwiss. Kom., 2: 125-136.
K. B. (1950).- K morfologii mutovchatykh sifonej karbona severnogo Urala.- Doklady AN SSSR, 73, 3: 569-571.
Z. (1959).- Stratigraphy, sedimentology and palaeogeography of the high-Tatric Triassic in the Tatra Mts.- Acta Geol. Pol. 9, 2: 113-143.
Z. (1963).- Stratygrafia i litologia triasu regli zakopiańskich [Stratigraphie et lithologie du Trias subtatrique de la région de Zakopane].- Acta Geol. Pol., 13, 3-4: 317-385.
Z. (1965a).- Analogie litologiczne triasu tatrzańskiego z triasem wschodnioalpejskim.- Rocznik PTG, 35, 2: 143-162.
Z. (1965b).- Budowa geologiczna pasma reglowego między Doliną Małej Łąki i Doliną Kościeliską.- Acta Geol. Pol. 15, 3: 257-320.
Z. (1967).- Palaeontological basis of the Triassic stratigraphy in the Tatra Mts.- Geol. Sb. Slov. Akad. Věd. Geol. Carpath., 18, 2: 277-288.
Z. (1973).- Upper and Middle Subtatric Nappes in the Tatra Mts.- Bull. de l'Acad. Pol. des Sci., Sér. Sci. de la Terre, 21, 1: 75-83.
Z. (1977).- Trias. In: H. (ed.), Geologia historyczna.- Wyd. Geol. Warszawa: 477-558.
Z. (1979).- Gromada Chlorophyta, rząd Siphonales, rodzina Dasycladaceae. In: A. et al. (eds.), Budowa geologiczna Polski. Tom III. Atlas skamienialosci przewodnich i charakterystycznych. Część 2a. Mezozoik. Trias.- Instytut Geologiczny, Wyd. Geol., Warszawa: 218-222.
Z. (1981).- Nowe obserwacje nad Dasycladaceami z dolomitu diploporowego z wierceń okolic Zawiercia (Wyżyna Śląsko-Krakowska). In: Fauna i flora triasu obrzeżenia Gór Świętokrzyskich i Wyżyny Śląsko-Krakowskiej. Materiały V Krajowej Konferencji Paleontologów. Kielce-Sosnowiec: 74-78, Kielce.
Z. (1986).- Macroflora. Class Chlorophyta. In: A. et al. (eds.), Geology of Poland. Vol. III. Atlas of guide and characteristic fossils. Part 2a. Mesozoic. Triassic.- Instytut Geologiczny, Wyd. Geol., Warszawa: 176-184, 218-220.
Z. (1990).- Znaczenie diplopor dla badań dolomitów środkowotriasowych regionu śląsko-krakowskiego. CPBR cel 71.00.024. pkt. kontr. 4.- Archiwum PIG, Warszawa.
Z. (1994a).- Środkowotriasowe Dasycladaceae z Polski i innych brzegów Perytetydy. Abstr. II Kongr. Międzynar. 343 Projektu IGCP "Perytetyda". Bukareszt.- Przegl. Geol. 42, 5: 390-394.
Z. (1994b).- Middle Triassic Dasycladaceae of Upper Silesian-Cracow Region and their stratigraphical and palaeoecological significance.- Third Intern. Meeting of Peri-Tethyan Epicratonic Basins. Cracow. (Poland). Excursion Guidebook. Pol. Geol. Instit. Warszawa, 59-66.
Z. (1995).- Middle Triassic Dasycladaceae of Algeria.- Second International Symposium on the Geology of the Eastern Mediterranean Region (Abstracts), Jerusalem, Israel: 12.
Z. & G.A. (1973).- Triassic Dasycladaceae from central Balkan and fore-Balkan, Bulgaria.- Acta Pal. Pol., 18, 2: 187-210.
Z., G. & T. (2004).- Reptile tracks (Rotodactylus) from the Middle Triassic of Djurdjura Mountains in Algeria.- Geol. Quart., 48, 1: 89-96.
Z., J., S. & K. (1967).- Budowa geologiczna kamieniołomów "Kamień Wielki" na tle budowy okolic Kamienia Śląskiego i Śląska Opolskiego. In: Studium geologiczno-inżynierskie nad możliwością składowania odpadów z ZakładówChemicznych "Blachownia". Problem składowania w kamieniołomach "Kamień Wielki".- Archiwum Zakładu Prac Inżyniersko-Geologicznych Uniwersytetu Warszawskiego: 1-64.
S. (1973a).- Szczegółowa Mapa Geologiczna Polski 1:50,000, ark. Strzelce Opolskie.- Wyd. Geol. Warszawa.
(1973b).- Objaśnienia do Szczegółowej Mapy Geologicznej Polski 1:50,000, ark. Strzelce Opolskie.- Wyd. Geol. Warszawa.
S. (1977).- Mapa geologiczna Polski w skali 1:200,000, ark. B bez utworów czwartorzędowych, ark. Gliwice.- Wyd. Geol. Warszawa.
S. & M. (1976a).- Szczegółowa Mapa Geologiczna Polski 1:50,000, ark. Tworóg.- Wyd. Geol. Warszawa.
S. & M. (1976b).- Objaśnienia do Szczegółowej Mapy Geologicznej Polski 1:50,000, ark. Tworóg.- Wyd. Geol. Warszawa.
S. & H. (1980).- Stratigraphische Reichweite der wichtigsten conodonten (ohne Zohnreihen-conodonten) der Mittel und Obertrias.- Geol.-Paläont. Mitt., Innsbruck, 10, 2: 47-78.
H. (1974a).- Biostratigraphie der germanischen Mitteltrias.- Freiberger Forsch.-H.C. 280,1: 1-56, 2: 1-71, 3: 12.
H. (1974b).- Probleme der Triasgliederung und Parallelisierung der germanischen und tethyalen Trias. Teil I - Abgrenzung und Gliederung der Trias.- Freiberger Forsch.-H.C. 298: 139-197.
H. (1980).- Revision der Conodontenzonierung der Mittel und Obertrias des tethyalen Faunenreichs.- Geol. Paläont. Mitt., Innsbruck, 10, 3-4: 79-172.
L. (1983).- Das Epidaurus-Profil (Griechenland) - ein Beitrag zur Conodonten-Standartzonierung des tethyalen Ladin und Unterkarn.- Österr. Akad. Wiss. Schriftenreihe Erdwiss. Komm., Wien, 5: 231-258.
M. & J. (1953).- Geologia Polski.- Wyd. Geol. Warszawa.
C. (1932).- Złoża rud ołowiu w okolicy Siewierza.- Spraw. PIG 7, 1: 3-74. Warszawa.
M. (1952).- Sur la présence de Dasycladacées dans le trias de la chaine bétique (Espagne).- C.R. Acad. Sci., 234: 1382-1384.
M. (1954).- Étude des Algues calcaires et de leur répartition. In: P. et al. (eds.), Observations sur le Trias bétique et ses Algues calcaires.- Memorias y Communicaciones del Instituto geologico provincial, Barcelona, 11: 23-60.
M. (1963).- Algues calcaires du Trias de France et des régions limitrophes.- Mém. Bureau Rech. Géol. Min., 15: 564-566.
R. (1914).- Geologischen Karte von Preussen und banachbarten Bundesstaaten landern - Blatt Tarnowitz-Brinitz [Tarnowskie Góry, Brynica]. Geologisch aufgenommen durch R. , F. und P. ; Erläutert durch R. .- Königlich Preussischen Geologischen Landesanstalt; A.W. Schade Buchdruckerei, Berlin, 108 pp.
R., P. & W. (1915).- Geologische Karte von Preussen und banachbarten deutschen landern. Blatt Beuthen - Laurahütte [Bytom-Siemianowice] Geologisch und bodenkundlich bearbeitet durch P. , R. , W. und F. , 1907-1910; Erläutert durch R. , P. und W. , 1915; Mit 18 Abbildungen im Text.- Königlich Preussischen Geologischen Landesanstalt, Hansa Buchdruckerei, Berlin, 131 pp.
M. (1972).- Lithologische und fazielle Analyse der Mittleren Trias der Kerngebirge der Westkarpaten.- Acta geol. geogr. Univ. Comen., Geologica, 22: 5-154.
E. (1974).- Koralowce z warstw karchowickich z Kamienia Śląskiego k. Opola.- Przewodnik 46 Zjazdu Pol. Tow. Geol. IG, Warszawa.
E. (1988).- Middle Triassic Scleractinia from the Cracow-Silesia region, Poland.- Acta Paleont. Pol., 33: 91-121.
J. (1992).- Litofacje i sedymentacja dolomitów diploporowych (środkowy wapień muszlowy) wschodniej części obszaru śląsko-krakowskiego.- Ann. Soc. Geol. Poloniae, 62: 19-62.
J. (1993).- Diagenesis of Diplopora Dolomite (Middle Muschelkalk) in the East Part of the Cracovian-Silesian Region.- Geologia. AGH, Kraków 19, 4: 171-202.
M. (1990).- Ocena bazy zasobowej surowców węglanowych dla potrzeb przemysłu materiałów ogniotrwałych.- CPBR 1. 8, cel 24. Archiwum CPBR - CAG, PIG Warszawa.
K. & J. (2004).- Controls of migrations of conodont fauna in peripheral oceanic areas - on example from Middle Triassic of the Northern Peri-Tethys.- Géobios 37: 425-436.
J. & J. (2000).- The middle Triassic magnetostratigraphy from the Peri-Tethys basin in Poland.- Earth, Planet. Sc. Letters, 182: 77-92.
R. (2000).- Litostratygrafia formacji górażdżańskiej i formacji dziewkowickiej na Śląsku Opolskim.- Pr. Geol.-Mineral. U. Wr., 71: 1-72.
M.M. (1927).- Das Grödener-, Fassa- und Enneberggebiet in den Südtiroler Dolomiten. Geologische Beschreibung mit besonderer Berücksichtigung der Überschiebungserscheinungen.- Abh. Geol. Bundesanst., Wien, 24, 2, 1-89.
E. (1967).- Dasycladazeen (Kalkalgen) aus der nordalpinen Obertrias.- Mitteil. der Bayer. Staatssamml. f. Paläont. u. hist. Geol., München, 7: 205-226.
E. (1972a).- Die Kalkalgen-Chronologie der alpinen Mitteltrias in Angleichung an die Ammoniten-Chronologie.- N. Jb. Geol. Paläont, Abh., 141, 1: 81-115.
E. (1972b).- Mitteltriadische Riffe der Nördlichen Kalkalpen und altersgleiche Bildungen auf Karaburun und Chios (Ägäis). In: Mikrofazies und Mikrofauna der Alpinen Trias und deren Nachbargebiete (Symposium Innsbruck, 20.-23. März 1972).- Mitt. Ges. Geol.- und Bergbaustud. Öst., Innsbruck, 21, 1: 251-276.
E. (1972c).- Neufunde anisischer Dasycladaceen (Kalkalgen) in Österreich.- Anzeiger der math.-naturwiss. Kl. Öster. Akad. Wiss., Wien, 1972, 9: 187-200.
E. (1974).- Algae (Dasycladaceae). In: H. (ed.), Catalogus Fossilium Austriae. Eine systematisches Verzeichnis aller auf österreichischen Gebiet festgestellten Fossilien.- Öster. Akad. Wiss., Wien (Springer-Verlag, Wien), 17b: 1-64.
S. (1967).- Mikropaleontološke karakteristike srednjeg i gornjeg trijasa planine Tare (zapadna Srbija) [Les caractéristiques micropaléontologiques du Trias moyen et supérieur de la montagne Tara (Serbie occidentale)].- Vesnik, Ser. A (Geologija), Beograd, Knjiga, 24/25 (1966-1967): 245-254.
S. (1970).- Mikropaleontološke karakteristike trijaskog stuba antiklinale Ždrela (istočna Srbija) [Caractéristiques micropaléontologiques de la colonne triasique de l'anticlinal de Ždrelo (Serbie oriental)].- Vesnik, Ser. A (Geologija), Beograd, Knjiga, 28: 377-386.
- S. (1975).- Trijaske mikrofacije Dinarida. Crna Gora, istočna Bosna i Hercegovina i zapadna Srbija [Les microfaciès triasiques des Dinarides. Le Monténégro, la Bosnie orientale et l'Hérzegovine et Serbie occidentale].- Društvo za nauku i umjetnost Crne Gore, Titograd (Podgorica), Posebna izd., 4, Od. Prir. nauka, 4: 1-56.
- S. (1995).- Mikrofosili donjeg i srednjeg trijasa u jugozapadnom delu bosanskog Podrinja [Lower and Middle Triassic microfossils in southwestern areas of Bosnian Podrinje].- Geološki anali Balkanskoga poluostrva, Beograd, 59, 2: 155-178.
C. & S. (1960).- Występowanie glonów (Dasycladaceae) w dolomitach kruszconośnych okolic Chrzanowa [Algae (Dasycladaceae) from ore-bearing dolomites in the vicinity of Chrzanów].- Kwart. Geol., 4, 3: 679-699.
D. (1970).- Inventar sumar al algelor Dasycladaceae triasice din Carpatii românesti.- Dări de seamă ale şedinţelor, Bucureşti, 55 (1967-1968), 3, Pal: 187-196.
J. (1985).- Rozwój facjalny morskich osadów triasu na obszarze śląsko-krakowskim.- Biul. Inst. Geol. 349: 41-92.
B. (1979).- Les algues du Jurassique et du Crétacé inférieur des Pyrénées franco-espagnoles. Intéręt biostratigraphique et paléoécologique.- Bull. Centr. Rech. Explor.-Prod. Elf-Aquitaine, 3: 733-741.
J. von (1912).- Neue Studien über die triadischen Siphoneae verticillatae.- Beiträge zur Paläontologie und Geologie Österreich-Ungarns und des Orients, 25: 25-81.
J. von (1918).- Zur Alterbestimmung des Chocz-dolomites.- Jahresber. kön. Ung. Geol. R.-A., (Budapest) 1916, 1: 256 - 258.
J. von (1920).- Die Siphoneae verticillatae vom Karbon bis zur Kreide.- Abh. zool.-bot. Gesellschaft, Wien, 11, 2, 1-263 [Les Siphonées verticillées du Carbonifère au Crétacé. Éditions Technip, Rueil-Malmaison, 1961].
J. von (1926).- Die Diploporen der deutschen Trias und die Frage der Gleichsetzung der deutschen und alpinen Triasstufen.- Z. deutsch. geol. Ges., 78: 192-201.
J. von (1927a).- Die Diploporen der deutschen und alpinen Triasstufen.- Z. deutsch. geol. Ges., 23: 192-201.
J. von (1927b).- 1. Abteilung: Thallophyta. In: M. (ed.), Handbuch der Paläobotanik.- Oldenbourg, München, Berlin, I: 31-136.
J. von (1930a).- Grundbergiffe der Stratigraphie mit ausfürlicher Anwendung auf die europäische Mitteltrias.- Deudicke, Leipzig, Wien, 255 pp.
J. von (1930b).- Upper Triassic fossils from the Burmo-Siamese frontier. A new Dasycladacea, Holosporella siamensis nov.gen., nov.sp., with a description of the allied genus Aciculella .- Rec. Geol. Surv. India, 63, 1: 177-181.
J. von (1931a).- Die Dasycladaceen der germanischen Trias.- Ann. Naturhist. Mus. Wien, 45: 265-279.
J. von (1931b).- Grundsätzliches zum Vergleiche germanischer und alpiner Trias.- Mitt. Geol. Ges. Wien, 23: 160-163
J. von (1935a).- Die Diploporen der anisischen Stufe Bosniens.- Geološki anali Balkanskog poluostrva, Beograd, 12, 2: 190-246.
J. von (1935b).- Algen und Pseudoalgen aus der spanischen Trias. In: M. (ed.), Fossilien der spanischen Trias.- Abh. Heidelberger Akad. Wiss., M.-n. Kl., 22: 9-17.
J. von (1936).- Description of the algae. In: L.R. & J. von (ed.), Fossil algae from the Uppermost Cretaceous beds (the Niniyur Group) of the Trichinopoly District, S. India.- Mem. Geol. Surv. India, Palaeont. Indica, N.S. 21, 4: 13-44.
J. von (1937a).- Die wichtigsten Kalkalgen des Jungpaläozoikums und ihre geologische Bedeutung.- Deuxième Congrès pour l'Avancement des Études de Stratigraphie Carbonifère (1935), Compte-Rendu, Heerlen, 2: 765-856.
J. von (1937b).- Sammelberichte über fossile Algen: Dasycladaceae 1928-1936 mit nachträgen aus früheren Jahre.- Neues Jahrb. Min. Geol. Palaeont. 3: 985-1027.
J. von (1942).- Uebersicht über fossile Kalkalgen und die geologischen Ergebnisse ihrer Untersuchung (Vortrag).- Mitteilungen des alpenländischen geologischen Vereines (Mitteil. Geol. Gesellschaft Wien), 33 (1940): 11-34.
S. (1976).- An outline of the metallogeny of zinc and lead ores in the Silesian-Cracovian region. In: J. (ed.), The current metallogenic problems of central Europe.- Wydawnictwa Geologiczne, Warsaw: 367–384.
W., R., W. & W. (1915).- Geologischen Karte von Preussen und banachbarten Bundesstaaten - Blatt Gleiwitz [Gliwice]. Geologisch-agronomich bearbeitet durch W. , R. , W. und F. ; Erlautert durch W. .- Koniglich Preussischen Geologischen Landesanstalt, A.W. Schade Buchdruckerei, Berlin, 84 pp.
J. (1965).- Trias środkowy masywu Kominów Tylkowych i jego tektonika. Le Trias moyen et la tectonique de massif de Kominy Tylkowe.- Acta Geol. Pol. 15, 3: 355-386.
E. & O. (1973).- Alge şi foraminifere triasice (anisian-ladinian) din estul Pădurii Craiului (Munţii Apuseni). Studii şi Cercetări de Geologie, Geofizică, Geografie, seria Geologie, Bucureşti, 18, 2: 425-442.
A. (1997).- Middle-Late Triassic Dasycladales (green algae) from Brenta Dolomites (Giudicarie Alps, Italy). Riv. Ital. Paleont. Strat. Milano, 103, 1: 71-80.
F. (1913).- Les Préalpes entre le Simmental et Dierntigal.- Fasc. I. Mat. Carte Geol. Suisse, 35: 1-130.
M. (1892).- Zapiski paleobotaniczne. Kosmos, Lwów, 11-12, 8 p.
R. (1962).- O jednoj novoj gornjojurskoj dazikladacei Pianella grudii nov.gen., nov.spec. [Sur une Dasycladacée nouvelle du Jurassique supérieur Pianella grudii nov.gen., nov.sp.].- Vjesnik, Ser. A (Geologija) Beograd, 20: 190-199, 201-210.
D.M. & S.M. (1978).- Principles of paleontology (2nd ed.).- W.H. Freeman, New York.
J. (1963).- Esquisse paléogéographique de la France aux temps triasiques. Colloque sur le Trias de la France et des régions limitrophes. Montpellier 1961.- Mém. Bur. Rech. Géol. Min., 15: 715-734.
F. (1870).- Geologie von Oberschlesien.- Jb. Schles. Ges. Vaterl. Kultur., 1-587, Breslau.
F. (1924).- Stratygrafja wapienia muszlowego w północnej części Zagłębia Dąbrowskiego. Spraw. PIG 2, 3-4: 431-495.
J., K. & O. (1983).- Triassic Foraminifers of the West Carpathians.- Geol. Úst. D. Štura, 213 pp.
J. (1967).- Répartition bathymétrique des Algues fossilisables en Méditerranée.- Geol. Sbor. Geol. Carpath., 18, 1: 141-150.
J. (1988).- Principles of study of Adriatic shelf ecosystems from the viewpoint of applications in geology.- Geol. Sbor.- Geol. Carpath., 39, 3: 285-300.
H. (1973).- Trias. Wyżyna Śląsko-Krakowska. In: Budowa geologiczna Polski. T. 1. Stratygrafia. Cz. 2. Mezozoik.- IG, Warszawa: 31-40.
H. (1979).- Możliwości sformalizowania podziału litostratygraficznego środkowego i górnego triasu epikontynentalnego w Polsce.- Kwart. Geol., 23: 583-600.
H. (1998).- Trias północno-wschodniego obrzeżenia Górnośląskiego Zagłębia Węglowego.- Biul. Państw. Inst. Geol., 378: 5-66.
H. (2000).- Stratygrafia retu i wapienia muszlowego z południowo-zachodniej Polski.- Biul. Państw. Inst. Geol., 390: 97-166.
H. & A. (1975).- Stratygrafia i paleogeografia triasu.- Biul. Inst. Geol., 252: 135-151.
K.E. (1853).- Beiträge zur näheren Kenntnis der Bayern'schen Voralpen.- N. Jb. Geol. Paläont, 1853: 299-319.
K.E. (1863).- Süd-Bayerns Lethaea Geognostica. Geognostisch Betrachtet in ihren Petrefacten.- L. Voss, Leipzig.
M. (1928).- Die Lebewelt unserer Trias.- Hohenlohe'sche Buchhandlung Ferdinand Rau, Oehringen.
M. (1938).- Die Lebewelt unserer Trias.- Hohenlohe'sche Buchhandlung Ferdinand Rau, Oehringen, Nachtrag 1938.
D. (1933).- Sur la découverte de nouveaux gisements de Diplopores (Algues calcaires) dans le Trias de la zone du Briançonnais.- Travaux Lab. Géol. Fac. Sci. l'Univ. Grenoble, 17, 1: 59-75.
B. & P. (1979).- Distributional patterns of calcareous algae within Upper Triassic patch reef structures of the Northern Calcareous Alps (Salzburg). 2me Symposium international sur les Algues fossiles (Paris, 23-26 avril 1979).- Bull. Centr. Rech. Explor.-Prod. Elf-Aquitaine, 3, 2: 811-820.
S. (1949).- Zagadnienia stratygrafii morskich osadów triasu krakowskiego.- Rocznik PTG 18: 191-272.
S. (1952).- Utwory geologiczne obszaru pomiędzy Chrzanowem a Kwaczałą.- Biul. Inst. Geol. 60: 1-230.
S. (1984).- The functional morphology of the Carboniferous Dasycladacean genus Kulikia.- N. Jb. Geol. Paläont. Mh., 7: 427-436.
S. (1986).- Upper Viséan calcareous algae from the Lublin Coal Basin.- Acta Geol. Pol., 36: 1-3: 251-280.
S. (1987).- The Dasycladacean nature of the Late Paleozoic palaeoberesellid algae.- Acta Geol. Pol., 37, 1-2: 21-31.
B. (1968).- A new genus of calcareous algae from the Middle Triassic of Velebit [Novi rod vapnenačke alge iz srednjeg trijasa Velebita].- Geol. vjesnik, 21 (1967): 207-212.
B. (1974).- Diplopora clavaeformis (Chlorophyta, Dasycladaceae) from the Lower Ladinian of Mt. Velebit [O vrsti Diplopora clavaeformis (Chlorophyta, Dasycladaceae) iz donjeg ladinika Velebita].- Geol. vjesnik, Zagreb, 27: 133-141.
G. (1903).- "Tetraporella Remeši", eine neue Dasycladacea aus dem Tithon von Stramberg.- Beiträge zur Paläontologie und Geologie Österreich-Ungarns und des Orients, Wien, 15, 2-3: 45-54.
J. (1991).- The Muschelkalk in Poland. In: H. (ed.), Muschelkalk. A Field Guide.- Korb (Goldschneck): 58-78.
J. (1994).- Sedimentary environment of the Diplopora Beds. In: Z. (ed.), 3rd Intern. Meeting of IGCP 343, Excursions Guidebook.- Cracow: 6.
J. (1999).- Anisian-Carnian evolution of the Germanic basin and its eustatic, tectonic and climatic controls.- Zbl. Geol. Paläont., (1998), 1, 7-8: 813-852.
J. (2000).- Middle Triassic evolution of the northern Peritethys area as influenced by early opening of the Tethys Ocean.- Ann. Soc. Geol. Pol., 70, 1: 1-48.
S. (1961).- Warstwy olkuskie.- Rudy i Met. Nież., 6, 12: 526-529.
S. (1964).- Geologia obszaru siewierskiego.- Prace Geol. Komisji Nauk Geol. PAN, 25: 1-74.
G. (1979).- Approche paléoécologique du monde des Dasycladales à partir de l'écologie des formes actuelles.- Bull. Centr. Rech. Explor.-Prod. Elf-Aquitaine, 3: 859-866.
K. (1914).- Jura Kruhela Wielkiego pod Przemyślem.- Rozprawy Pol. Akad. Umiej. (seria B), 53, 1-2: 409-490; 543-629; 54, 3-4: 13-69, 141-182; Kraków.
J. (1957).- Szczegółowa Mapa Geologiczna Polski w skali 1:50,000, ark. Zabrze.
J. (1968).- Objaśnienia do Szczegółowej Mapy Geologicznej Polski - 1:50,000, ark.- Kalety. Wyd. Geol., Warszawa.
C. (1965).- Il Trias in Lombardia (Studi geologici e paleontologici). 13. Le Alghe calcaree delle Prealpi Lombarde.- Riv. Ital. Paleont. Strat., 71, 2: 449-544.
K. (1974).- Stratygrafia triasu opolskiego na podstawie konodontów.- Przewodnik 46. Zjazdu PTG Opole: 27 - 29.
K. (1975).- Conodont stratigraphy and sedimentary environment of the Muschelkalk in Upper Silesia. Acta Geol. Pol., 25, 2: 217-256.
P.A. (1990).- Geological Atlas of Western and Central Europe.- Shell Intern. Petr. Maatsch. B. V., Hague.
H. (1971).- Paläontologische, stratigraphische und sedimentologische Untersuchungen des Salvatoredolomits (Mitteltrias) der Tessiner Kalkalpen. Unter besonderer Berücksichtigung der Mikrofazies, Diagenese und Taxionomie der Lamellibranchiata.- In: E. & B. (eds.), Der Triasfauna der Tessiner Kalkalpen.- Schweiz. Paläont. Abh., Basel, 91: 1-90.
H. (1972a).- Mikrofazielle Analyse eines mitteltriassischen Riffkomplexes in den Tessiner Kalkalpen. In: Mikrofazies und Mikrofauna der Alpinen Trias und deren Nachbargebiete (Symposium Innsbruck, 20.-23. März 1972).- Mitt. Ges. Geol.- und Bergbaustud. Öster., Innsbruck, 21, 1: 123-142.
H. (1972b).- Bau und Funktion der Wirteläste einiger triassischer Dasycladaceen (Kalkalgen).- N. Jb. Geol. Paläont, Mh., 12: 727-740.
H. (1976).- Über den Lebensraum fossiler Wirtelalgen in der Trias der Alpen.- Naturwiss., 63: 426-429.
H. (1977).- Der Einfluß der Umwelt auf die Variation fossiler Arten.- Natur u. Museum, Frankfurt/Main, 107, 1: 1-5.
E. (1968a).- Szczegółowa Mapa Geologiczna Polski w skali 1:50,000 arkusz Bytom.- Wyd. Geol., Warszawa.
E. (1968b).- Objaśnienia do Szczegółowej Mapy Geologicznej Polski - 1:50,000, ark. Bytom.- Wyd. Geol., Warszawa.
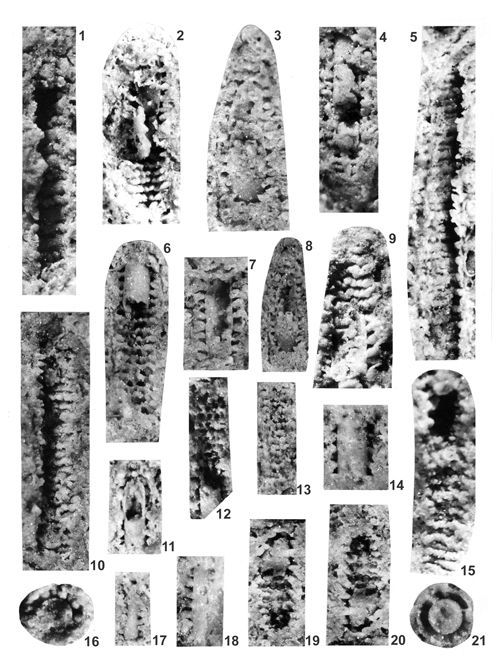 |
|
|
Click on thumbnail to enlarge the image.
Plate I: Salpingoporella krupkaensis n.sp. 1-21 Krupka.
1. Longitudinal fracture of the thallus with single rows of phloiophorous tubercles seen from inside. Tubercles are strictly connected with surrounding rock; x 20; 2. Longitudinal fracture of the thallus with partly preserved internal tube connected with surrounding rock by single rows of phloiophorous tubercles; x 20, paratype; 3. Longitudinal fracture of the thallus with phloiophorous tubercles set on internal tube and enlarging towards the surrounding rock; x 20; 4. Longitudinal fracture of the thallus with partly preserved internal tube connected with surrounding rock by phloiophorous tubercles; x 20; 5. Longitudinal fracture of the thallus with dense single rows of phloiophorous tubercles seen from inside. Tubercles are strictly connected with surrounding rock; x 20; 6. Longitudinal fracture of the thallus with partly preserved smooth internal tube connected with surrounding rock by single rows of phloiophorous tubercles; x 20, holotype; 7. Longitudinal fracture of the thallus with perfectly preserved phloiophorous tubercles, set on the internal tube and rapidly enlarging towards the surrounding rock, forming funnel-like features, x 20, paratype; 8. Longitudinal fracture of the thallus with perfectly preserved internal tube connected with external tube by phloiophorous tubercles; x 20; 9. Longitudinal fracture of the thallus with phloiophorous tubercles set on partly preserved internal tube and rapidly enlarging towards the surrounding rock with relics of the cortex (?); x 20; 10. Longitudinal fracture of the thallus with single rows of phloiophorous tubercles from inside; x 20; 11. Longitudinal fracture of the thallus with two tubes: internal and external one, connected by phloiophorous tubercles; x 20; 12. Longitudinal fracture of the thallus with single rows of phloiophorous tubercles and remnant of internal tube; x 20; 13. Longitudinal fracture of the thallus through dense single rows of phloiophorous tubercles; x 20; 14. Longitudinal fracture of the thallus showing smooth surface of internal tube with phloiophorous tubercles set on and rapidly enlarging and merging with surrounding rock; x 20; 15. Longitudinal fracture of the thallus through dense single rows of tubercles, merging with surrounding rock; x 20; 16. Transverse fracture with phloiophorous tubercles initiating from internal tube filled with sediment and merging with surrounding rock; x 20; 17. Longitudinal fracture of the thallus showing smooth internal tube, with phloiophorous tubercles set on it and merging with surrounding rock; x 13; 18. Longitudinal fracture of the thallus showing smooth internal tube connected by phloiophorous tubercles with surrounding rock; x 20; 19. Longitudinal fracture of the thallus showing singular rows of phloiophorous tubercles, merging with surrounding rock; x 20; 20. Longitudinal fracture of the thallus showing singular rows of phloiophorous tubercles, merging with surrounding rock; x 20; 21. Transverse fracture with internal tube filled by sediment and dark hollow space between internal and external tube, with some preserved phloiophorous tubercles; x 20.
 |
|
|
Click on thumbnail to enlarge the image.
Plate II: Salpingoporella krupkaensis n.sp. 1-11, 16 Kamyce, 12-15 Krupka.
1. Longitudinal fracture of the thallus with single rows of tubercles setting on the smooth surface of internal tube, phloiophorous tubercles merge with surrounding rock; x 20; 2. Longitudinal fracture through central cavity, partly filled and through the wall connected with surrounding rock by phloiophorous tubercles, rapidly enlarging and merging with adjacent rock; x 20; 3. Longitudinal fracture through central cavity, partly filled and through internal tube, connected with surrounding rock by remnants of tubercles; x 20; 4. Longitudinal fracture of the thallus, partly filled central cavity and internal tube with setting phloiophorous tubercles, rapidly enlarging and merging with surrounding rock and remnants of the cortex; x 20; 5. Longitudinal fracture of the thallus with phloiophorous tubercles; x 20; 6. Longitudinal fracture of the thallus with singular rows of phloiophorous tubercles and partly uncovered smooth internal tube; x 20; 7. Longitudinal fracture of the thallus with partly uncovered internal tube, connected by phloiophorous tubercles with external tube; x 20; 8. Longitudinal fracture of the thallus with singular rows of tubercles; x 20; 9. Transverse fracture of the thallus with large, filled central cavity, connected with adjacent rock by very large phloiophorous tubercles; x 32; 10. Transverse fracture of the thallus with central cavity filled by sediment; phloiophorous tubercles are disconnected from central cavity and merging with adjacent rock; x 32; 11. Longitudinal oblique fracture of the thallus viewed from inside, with single rows of phloiophorous tubercles connecting with surrounding rock; x 20; 12. Oblique fracture of the thallus with large central cavity and phloiophorous tubercles merging with adjacent rock; x 32; 13. Longitudinal fracture of thin thallus with hollow central cavity, thin internal wall and tubercles connecting it with external wall; x 20; 14. Longitudinal fracture of the thallus viewed from inside, with single rows of phloiophorous tubercles merging with adjacent rock; x 20; 15. Longitudinal fracture of the thallus with perfectly preserved smooth internal tube, connected with adjacent rock by single rows of phloiophorous tubercles, in some places thin external wall is partly preserved; x 20; 16. Longitudinal fracture of the thallus with partly hollow central cavity and fragmentarily preserved internal tube connected with adjacent rock by singular rows of phloiophorous tubercles; x 20.
 |
|
|
Click on thumbnail to enlarge the image.
Plate III: Salpingoporella krupkaensis n.sp. 1-4, 8, 9, 13-18, 20, 23-24 Krupka, 5, 11 Rosowa Góra, 6-7, 10, 12, 21 Balin, 19 Kamyce, 25-26 Stare Gliny.
1 . Longitudinal fracture of the thallus with hollow central cavity, partly preserved internal tube and phloiophorous tubercles merging with adjacent rock; x 20; 2. Longitudinal fracture of the thallus showing internal tube with phloiophorous tubercles attached to it and merging with adjacent rock; x 20; 3. Longitudinal fracture of the thallus with single rows of phloiophorous tubercles attached to partly preserved internal tube and merging with adjacent rock; x 20; 4. Longitudinal fracture of the thallus viewed from inside, with singular dense rows of phloiophorous tubercles, merging with adjacent rock; x 20; 5. Longitudinal fracture of the thallus showing central stem with phloiophorous tubercles attached to it; x 20; 6. Longitudinal fracture of the thallus, showing slightly bent central stem with phloiophorous tubercles attached to it and merging with adjacent rock; x 20; 7. Longitudinal fracture of the thallus, showing hollow central cavity and internal tube with phloiophorous tubercles partly attached to it and merging with adjacent rock; x 20; 8. Longitudinal fracture of the thallus with central stem, internal wall and single rows of phloiophorous tubercles, reaching to thin external wall; x 20; 9. Longitudinal fracture of the thallus with partly preserved internal tube and phloiophorous tubercles attached to it and reaching to thin external wall; x 20; 10. Longitudinal fracture of the thallus showing partly hollow smooth internal tube with rounded uppermost part; partly preserved phloiophorous tubercles; x 20; 11. Longitudinal fracture of very long the thallus viewed from inside, with several (more than 30) single rows of phloiophorous tubercles merging with adjacent rock; x 20; 12. Longitudinal oblique fracture of the thallus showing almost hollow central cavity and internal wall with attached phloiophorous tubercles, merging with adjacent rock; x 20; 13. Longitudinal fracture of the thallus with single rows of phloiophorous tubercles, attached to remnant of the inner tube and merging with adjacent rock; x 20; 14. Longitudinal fracture of the thallus showing hollow central cavity and internal wall with phloiophorous tubercles attached to it and merging with adjacent rock; x 20; 15. Transverse fracture with central stem and phloiophorous tubercles closely joined with adjacent rock; x 32; 16. Transverse fracture with central stem and phloiophorous tubercles closely joined with adjacent rock; x 20; 17. Transverse fracture with central stem and phloiophorous tubercles closely joined with adjacent rock; x 20; 18. Transverse fracture of the thallus with hollow central cavity, internal wall and phloiophorous tubercles closely joined with adjacent rock; x 20; 19. Transverse fracture with central stem and partly preserved phloiophorous tubercles, closely joined with adjacent rock; x 32; 20. Longitudinal fracture of the thallus with phloiophorous tubercles on the inner tube, closely joined with adjacent rock; x 20; 21. Transverse fracture with central stem and phloiophorous tubercles closely joined with adjacent rock; x 20; 22. Transverse fracture with central stem and phloiophorous tubercles, reaching to the outer wall; x 20; 23. Longitudinal fracture of the thallus with partly preserved internal tube and attached to it phloiophorous tubercles, forming single rows and merging with adjacent rock; x 13; 24. Longitudinal fracture of the thallus with central stern, external wall and partly preserved tubercles; x 13; 25. Transverse fracture with central stem and phloiophorous tubercles, closely joined with adjacent rock; x 32; 26. Transverse fracture of the thallus with very large central stem and phloiophorous tubercles, closely joined with adjacent rock; x 32.
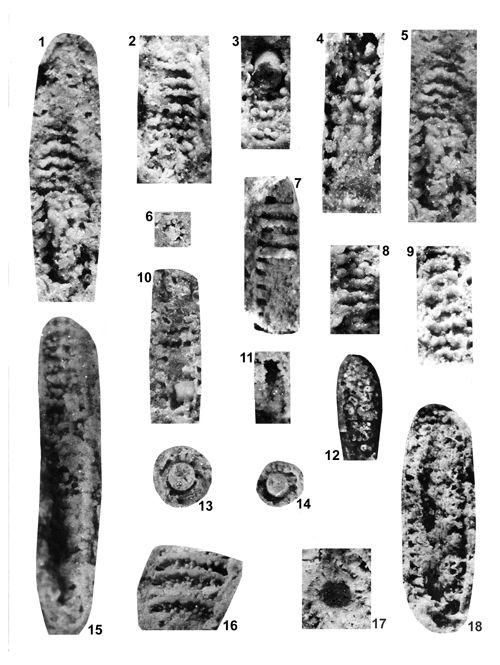 |
|
|
Click on thumbnail to enlarge the image.
Plate IV: Salpingoporella krupkaensis n.sp. 1-3, 5-6, 8-9, 11, 13, 17 Krupka, 4 Stare Gliny, 7, 16 Balin, 10, 14 Kamyce, 12, 18 Przełajka.
1. Longitudinal fracture of the thallus viewed from inside, showing single rows of phloiophorous tubercles and relics of the internal tube; x 20; 2. Longitudinal fracture of the thallus viewed from inside, with single rows of phloiophorous tubercles; x 20; 3. Longitudinal fracture of the thallus viewed from inside, showing single rows of phloiophorous tubercles, connecting two tubes - internal and external; x 20; 4. Longitudinal fracture of the thallus with single rows of bulky tubercles; x 32; 5. Longitudinal fracture of the thallus with single rows of phloiophorous tubercles, merging with adjacent rock; x 32, paratype; 6. Transverse fracture showing central stem surrounding by round phloiophorous tubercles; x 13; 7. Longitudinal fracture of the thallus with double rows of round tubercles separated by interannular furrows; x 20; 8. Longitudinal fracture with single rows of bulky tubercles; x 20; 9. Longitudinal fracture of the thallus showing single, bent rows of round tubercles; x 20; 10. Longitudinal fracture of the thallus with tubercles in single rows; x 30; 11. Longitudinal fracture showing internal view with tubercles in single rows; x 13; 12. Longitudinal fracture showing internal view with hollow tubercles in single rows; x 20; 13. Transverse fracture showing the central stem with phloiophorous tubercles merging with surrounding rock; x 20; 14. Transverse fracture showing the central stem with phloiophorous tubercles merging with surrounding rock; x 20; 15. Longitudinal fracture of the thallus showing the single rows of phloiophorous tubercles; x 20; 16. Longitudinal fracture showing from inside the annular rings filled by thin tubercles, separated by interannular furrows; x 20; 17. Transverse fracture showing the hollow central cavity surrounded by tubercles merging with adjacent rock; x 30; 18. Longitudinal fracture of the thallus showing from inside the rows of partly hollow tubercles reaching to the external wall; x 20.
 |
|
|
Click on thumbnail to enlarge the image.
Plate V: Oligoporella elegans () Photographs of thin sections from locality no. 2 - Tarnów Opolski (1-23) and locality no. 3 - Kamień Śląski (24-37).
1. Longitudinal and oblique sections; x 9.9; 2. Transverse and oblique sections; x 10.2; 3. Oblique section; x 9.9; 4. Longitudinal oblique section; x 10.3; 5. Longitudinal oblique section; x 10.2; 6. Longitudinal section; x 9.9; 7. Longitudinal oblique section; x 10.2; 8. Transverse oblique section; x 10.3; 9. Oblique section; x 10.2; 10. Longitudinal oblique section; x 10.3; 11. Differently oriented section; x 10.3; 12. Oblique section; x 10.3; 13. Transverse and longitudinal oblique sections; x 10.2; 14. Transverse oblique section; x 10.3; 15. Transverse section; x 10.3; 16. Longitudinal section; x 10.3; 17. Transverse section; x 9.9; 18. Oblique section; x 9.9; 19. Transverse section; x 10.2; 20. Transverse section; x 10.2; 21. Transverse section; x 10.2; 22. Transverse section; x 9.9; 23. Transverse section; x 10.2; 24. Longitudinal oblique section; x 12.2; 25. Longitudinal oblique section; x 12.2; 26. Longitudinal oblique section; x 12.2; 27. Longitudinal oblique section; x 12.2; 28. Longitudinal section; x 14; 29. Longitudinal oblique sections; a - x 9, b - x 14; 30. Longitudinal section; x 9; 31. Transverse oblique section; x 9; 32. Transverse section; x 9; 33. Oblique section; x 9; 34. Transverse section; x 9; 35. Transverse section; x 9; 36. Oblique section; x 12.2.
 |
|
|
|
|
Click on thumbnail to enlarge the image.
Plate VI: Oligoporella elegans () Photographs of thin sections from several localities.
1. Longitudinal oblique section, x 11.3; locality no. 5, Laryszka; 2. Longitudinal oblique section, x 7; locality no. 6, Suchodaniec; 3. Transverse oblique section, x 7; locality no. 6, Suchodaniec; 4. Transverse oblique section, x 7; locality no. 6, Suchodaniec; 5. Transverse section, x 7; locality no. 6, Suchodaniec; 6. Longitudinal oblique section, x 8.8; locality no. 6, Suchodaniec; 7. Longitudinal oblique section, x 8.7; locality no. 7, Nowe Koszyce; 8. Longitudinal oblique section, x 10; locality no. 7, Nowe Koszyce; 9. Transverse oblique section, x 10; locality no. 7, Nowe Koszyce; 10. Longitudinal oblique section, x 10; locality no. 7, Nowe Koszyce; 11. Transverse oblique section, x 10; locality no. 7, Nowe Koszyce; 12. Longitudinal oblique section, x 10; locality no. 7, Nowe Koszyce; 13. Transverse section, x 10; locality no. 7, Nowe Koszyce; 14. Transverse oblique section, x 10; locality no. 7, Nowe Koszyce; 15. Transverse oblique section, x 10; locality no. 7, Nowe Koszyce; 16. Transverse oblique section, x 10; locality no. 7, Nowe Koszyce; 17. Two transverse section, x 10; locality no. 7, Nowe Koszyce; 18. Two transverse oblique section, x 10; locality no. 7, Nowe Koszyce; 19. Transverse oblique section, x 10; locality no. 7, Nowe Koszyce; 20. Transverse section, x 10; locality no. 7, Nowe Koszyce; 21. Transverse oblique section, x 10; locality no. 7, Nowe Koszyce; 22. Transverse section, x 7.7; locality no. 8, Jemielnica; 23. Transverse oblique section, x 9.3; locality no. 8, Jemielnica; 24. Longitudinal oblique section, x 9.3 ; locality no. 8, Jemielnica; 25. Transverse oblique section, x 9.3; locality no. 8, Jemielnica; 26. Longitudinal oblique section, x 7.5; locality no. 11, Segiet; 27. Longitudinal oblique section, x 10; locality no. 13, Srebrna Góra; 28. Oblique and transverse section, x 10.3; locality no. 13, Srebrna Góra; 29. Transverse oblique section, x 10.3; locality no. 13, Srebrna Góra; 30. Transverse section, x 10.3; locality no. 13, Srebrna Góra; 31. Longitudinal oblique section, x 10.3; locality no. 13, Srebrna Góra; 32. Longitudinal section, x 10.5; locality no. 44, Imielin; 33. Longitudinal and transverse sections, x 7.2; locality no. 50, Balin; 34. Three oblique sections, x 7.2; locality no. 50, Balin; 35. Oblique section, x 10; locality no. 52, Krupka; 36. Longitudinal oblique section, x 10; locality no. 52, Krupka; 37. Longitudinal oblique section, x 10; locality no. 62, Łosień; 38. Oblique section, x 10; locality no. 70, Brudzowice; 39. Longitudinal section, x 10.4; locality no. 68, Siewierz ("Wiktor Emanuel" mine); 40. Two transverse sections, x 9; locality no. 70, Brudzowice; 41. Transverse section, x 9; locality no. 70, Brudzowice; 42. Transverse section, x 9; locality no. 70, Brudzowice; 43. Transverse section, x 9; locality no. 70, Brudzowice; 44. Transverse section, x 9; locality no. 70, Brudzowice; 45. Longitudinal section, x 9; locality no. 70, Brudzowice; 46. Longitudinal section, x 9; locality no. 70, Brudzowice; 47. Longitudinal section, x 10; locality no. 71, Dziewki (Nowa Wioska); 48. Oblique section, x 10; locality no. 71, Dziewki (Nowa Wioska); 49. Oblique section, x 10; locality no. 71, Dziewki (Nowa Wioska); 50. Longitudinal section, x 10; locality no. 71, Dziewki (Nowa Wioska).
 |
|
|
Click on thumbnail to enlarge the image.
Plate VII: Oligoporella elegans () Photographs of fractures from several localities.
1. Very long longitudinal fracture of thick central stem with regular whorls (more than 20) of single trichophorous tubercles; x 16; locality no 57, Klucze (Stare Gliny); 2. Longitudinal fracture - central stem with regular whorls of single tubercles; x 16; locality no. 8, Jemielnica; 3. Longitudinal fracture of partly hollow central stem with rarely preserved tubercles; x 12; locality no. 48, Rosowa Góra; 4. Longitudinal fracture with the wall of hollow central stern, tubercles are partly preserved; x 12; locality no. 26, Kamyce; 5. Longitudinal fracture showing the interior of hollow central cavity with internal wall and partly preserved tubercles; x 12; locality no. 52, Krupka; 6. Very long longitudinal fracture with partly preserved central stern; x 12; locality 31KW, Morsko; 7. Longitudinal fracture with central stem and tubercles; x 10; locality no. 50, Balin; 8. Longitudinal fracture showing regular whorls of single trichophorous tubercles and partly preserved internal wall viewed from interior of hollow central cavity; x 20; locality no. 26, Kamyce; 9. Longitudinal fracture showing the central stem with regular whorls of single tubercles; x 12; locality no. 26, Kamyce; 10. Longitudinal fracture showing the interior of central cavity with slightly bent internal wall covered by single whorls of tubercles; x 10; locality no. 50, Balin; 11. Longitudinal oblique fracture with central stem and single whorls of tubercles; x 10; locality no. 25, Przełajka; 12. Transverse fracture showing filled central cavity and hollow places of former trichophorous branches (more than 10 in one whorl); x 10; locality no. 26, Kamyce; 13. Transverse fracture showing filled central cavity and hollow places of former trichophorous branches, x 10; locality no. 52, Krupka; 14. Transverse oblique fracture showing hollow central cavity and internal wall with trichophorous tubercles; x 12; locality no. 34, Czeladź; 15. Transverse fracture of filled central cavity and hollow places of former trichophorous branches (more than 12 in one whorl); x 10; locality no. 26, Kamyce; 16. Transverse fracture of filled central cavity and hollow places of former trichophorous branches (more than 12 in one whorl); x 12; locality no. 26, Kamyce; 17. Transverse fracture of filled central stem and hollow places of former trichophorous branches (more than 12 in one whorl); x 10; locality no. 26, Kamyce; 18. Transverse fracture of filled central stem and internal wall covered with thin trichophorous branches; the former calcareous wall is dissolved and only the round fissure is a trace of it; x 12; locality no. 34, Czeladź; 19. Longitudinal fracture showing partly preserved central stem covered with regular single whorls of trichophorous tubercles; x 10; locality no. 52, Krupka; 20. Oblique fractures showing filled central stem and rare tubercles; x 10; locality no. 50, Balin; 21. Longitudinal fracture of central stem and rare tubercles; x 10; locality no. 50, Balin; 22. Longitudinal fracture showing filled central stern, rare thin trichophorous branches and two parallel fissures - the place of former calcareous wall, completely dissolved; x 10; locality no. 52, Krupka; 23. Longitudinal fracture of thick central stem and partly preserved internal wall with rows of irregular tubercles; x 16; locality no. 57, Stare Gliny; 24. Longitudinal fracture of thick central stem covered with rare, partly deformed trichophorous branches; x 16; locality no. 48, Rosowa Góra; 25. Longitudinal fracture of hollow central stem and slightly bent internal wall with partly preserved thin tubercles in single rows; x 10; locality no. 48, Rosowa Góra; 26. Longitudinal fracture of central stem covered by tubercles; x 10; locality no. 52, Krupka; 27. Longitudinal fracture of central stem covered by single rows of tubercles; x 16; locality no. 50, Balin; 28. Longitudinal fracture of central stem; x 10; locality no. 50, Balin; 29. Longitudinal fracture of very long central stem with regular rows of trichophorous tubercles (branches); x 12; locality no. 48, Rosowa Góra.
 |
|
|
Click on thumbnail to enlarge the image.
Plate VIII: Oligoporella silesiaca (). 1-11, 14, 17-27 Rosowa Góra, 12-13 Jemielnica, 15 Przełajka, 16 Stare Gliny.
1. Longitudinal fracture of the thallus with dense double whorls of alternating trichophorous tubercles, oblique to the main axes; x 16; 2. Longitudinal fracture of the thallus with relatively largely spaced double whorls of alternating trichophorous tubercles; x 16; 3. Longitudinal fracture of the thallus with dense double whorls of alternating trichophorous oblique tubercles; x 16; 4. Longitudinal fracture of the thallus filled up by dense double whorls of alternating trichophorous thick oblique tubercles; x 23; 5. Longitudinal fracture of the thallus with apex, filled up by dense double whorls of alternating trichophorous thick oblique tubercles; x 20; 6. Longitudinal fracture of the thallus with double whorls separated by fissures and composed of alternating trichophorous oblique tubercles, slightly obliterated; x 16; 7. Longitudinal fracture of the thallus showing double whorls of alternating trichophorous oblique tubercles; x 20; 8. Longitudinal fracture of the thallus showing dense double whorls of alternating trichophorous oblique tubercles; x 20; 9. Longitudinal fracture of the thallus with the view to inside of central cavity and remnants of trichophorous oblique tubercles; x 16; 10. Longitudinal fracture of the thallus with relatively largely spaced double whorls of alternating trichophorous thick oblique tubercles; x 20; 11. Longitudinal fracture of the fragment of the thallus with double whorls of alternating trichophorous oblique tubercles; x 23; 12. Longitudinal fracture of the thallus showing several dense double whorls of irregularly arranged alternating trichophorous oblique tubercles; x 16; 13. Longitudinal fracture of the thallus with fragmentarily preserved central cavity and double whorls of alternating trichophorous oblique tubercles; the oblique whorls with tubercles form cones merging down, entering one into another; external wall is partly preserved and is slightly undulated; x 10; 14. Longitudinal fracture of the thallus with dense double whorls of trichophorous oblique tubercles, reaching from external wall to the central cavity; x 12; 15. Longitudinal fracture of the thallus with dense double rows of alternating trichophorous oblique tubercles; in the lower part of specimen the basal parts of branches are set on the internal tube with pores and score ridges; x 16; 16. Longitudinal fracture of the thallus filled up by tubercles, some of them oblique; x 16; 17. Fragment of the thallus with dense double whorls of the trichophorous oblique tubercles; x 16; 18. Fragment of the thallus with dense double of the trichophorous oblique tubercles; x 16; 19. Longitudinal fracture of the thallus with trichophorous oblique tubercles; x 12; 20. Fragment of the thallus with trichophorous oblique tubercles; x 12; 21. Transverse fracture of the thallus with trichophorous tubercles, connecting the internal and external wall; x 20; 22. Longitudinal fracture of the thallus showing dense whorls of trichophorous oblique tubercles; x 16; 23. Longitudinal fracture of the thallus showing dense whorls of trichophorous oblique tubercles; x 20; 24. Fragment of the thallus with double whorls of trichophorous alternating oblique tubercles; x 20; 25. Longitudinal fracture of the thallus showing whorls with alternating flattened trichophorous oblique tubercles; x 20; 26. Accumulation of the fragments with oblique tubercles; x 20; 27. Longitudinal fracture of the thallus filled up by the dense double whorls with trichophorous alternating oblique tubercles; x 20.
 |
|
|
Click on thumbnail to enlarge the image.
Plate IX: Oligoporella balinensis (). 1, 9 Tarnów Opolski, 2, 7 Balin, 3-4 Nowe Koszyce, 5, 8 Segiet, 6 Czeladź, 10 Srebrna Góra, 11 Krupka-10.
1. Longitudinal section showing double rows of alternating oblique tubercles; 1a x 10, 1b x 25; 2. Longitudinal tangential fracture with double rows of alternating oblique tubercles; 2a x 10, 2b x 30; 3. Longitudinal tangential fracture with double rows of alternating oblique tubercles; 3a x 12, 3b x 30; 4. Longitudinal fracture showing double rows of alternating oblique trichophorous tubercles; 4a x 19, 4b x 47; 5. Longitudinal section with hollow central cavity and double rows of alternating oblique trichophorous tubercles; 5a x 19, 5b x 47; 6. Longitudinal section with hollow central cavity and horizontal rows of oblique trichophorous tubercles; 6a x 20, 6b x 50; 7. Longitudinal fracture with horizontal rows of alternating oblique trichophorous tubercles; 7a x 20, 7b x 50; 8. Transverse fracture with hollow central cavity and trichophorous tubercles; 8a x 12, 8b x 30; 9. Transverse fracture with filled up central cavity and trichophorous tubercles; 9a x 12, 9b x 30; 10. Longitudinal fracture showing double rows of alternating oblique tubercles; 10a x 20, 10b x 50; 11. Accumulation of differently oriented specimens; x 20.
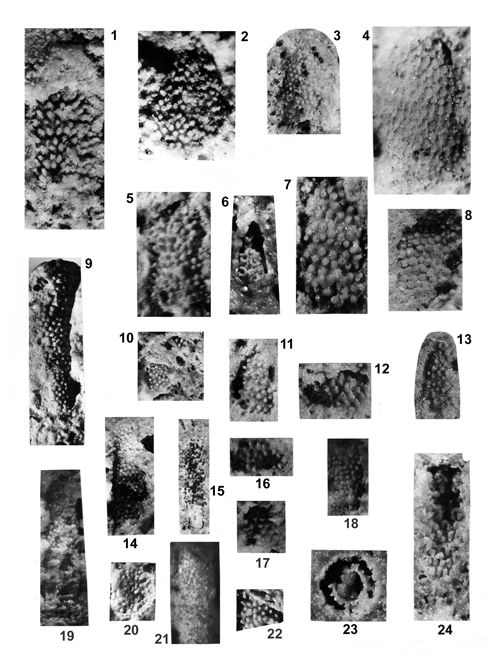 |
|
|
Click on thumbnail to enlarge the image.
Plate X: Oligoporella chrzanowensis n.sp. 1-5, 7-11, 13, 16-17, 19-20 Rosowa Góra, 6, 21 Stare Gliny, 12, 14 Balin, 15 Czeladź, 18 Jemielnica, 22 25 KW Winowo, 23-24 Krupka.
1. Longitudinal fracture of the large thallus showing probably the apex of the alga with flat external wall; in the middle part of the specimen one can observe the dense amassment of trichophorous oblique tubercles, arranged in hardly discernible horizontal whorls; nevertheless some horizontal plan is present; x 10, holotype; 2. Longitudinal fracture of the large thallus showing dense amassment of the oblique trichophorous tubercles, not arranged in the distinct whorls; x 10, paratype; 3. Longitudinal fracture of the thallus showing dense amassment of the trichophorous oblique tubercles not arranged in the distinct whorls and partly covered by the transparent external wall; x 6; 4. Longitudinal fracture of the thallus showing it from outside, traces of tubercles are present on the transparent external wall; the general shape suggests the apex in the upper part of specimen; x 10; 5. Longitudinal fracture of the thallus with trichophorous oblique tubercles in indistinct whorls; x 10; 6. Longitudinal fracture with the fragment of the thallus showing some part of the central stem with trichophorous tubercles, connecting the internal wall with external one; x 6; 7. Longitudinal fracture of the thallus, filled up by relatively thick trichophorous oblique tubercles, arranged in indistinct whorls; x 10; 8. Longitudinal fracture of the large thallus, filled up by relatively thick tubercles arranged in dense whorls; x 10; 9. Longitudinal fracture of long and narrow thallus filled up by trichophorous oblique tubercles arranged in indistinct whorls; x 6; 10. Two fragments of the thallus with irregular oblique trichophorous tubercles; x 6; 11. Longitudinal fracture of the thallus with trichophorous oblique tubercles in indistinct whorls; x 6; 12. Fragment of the thallus with amassment of trichophorous oblique tubercles; x 10; 13. Longitudinal fracture of the thallus with amassment of trichophorous oblique tubercles; x 6; 14. Longitudinal fracture of the long thallus with dense whorls of trichophorous oblique tubercles; x 6; 15. Longitudinal fracture of the long and narrow thallus with irregularly arranged thin trichophorous tubercles, viewed from inside; internal wall is partly preserved; x 6; 16. Transverse oblique fracture of the thallus with irregularly arranged dense trichophorous oblique tubercles; x 10; 17. Small fragment of the thallus with dense whorls of trichophorous oblique tubercles; x 10; 18. Longitudinal fracture of the thallus with dense whorls of trichophorous oblique tubercles; x 10; 19. Longitudinal fracture of the thallus with dense whorls of trichophorous oblique tubercles; x 6; 20. Longitudinal fracture of the thallus with dense whorls of trichophorous oblique tubercles; x 6; 21. Longitudinal fracture of the thallus with thin oblique trichophorous tubercles; x 6; 22. Small fragment of the thallus with dense whorls of trichophorous oblique tubercles; x 10; 23. Transverse fracture of the thallus showing partly filled central cavity and internal wall with trichophorous oblique tubercles (about 20) reaching in some places to the external wall; x 10; 24. Longitudinal fracture of the thallus showing the internal view with dense whorls of trichophorous oblique tubercles; x 10.
 |
|
|
Click on thumbnail to enlarge the image.
Plate XI: Physoporella pauciforata () 1-5, 7-15, 22-25 Przełajka, 20 10Ż Lgota Nadwarcie, 21 Jemielnica.
1. Longitudinal fracture of the thallus, viewed from inside; pores are arranged in horizontal single whorls separated by curled fillets; on the sides of specimen pores penetrate inside of the tubercles, reaching up to the external wall; x 8; 2. Longitudinal fracture of the thallus filled up by singular dense whorls of pyriform tubercles, forming also vertical rows; tubercles reach to the external wall, which forms regular tube; x 8; 3. Longitudinal fracture of the thallus with smooth central stem (in the middle), covered by single dense whorls of pyriform tubercles, arranged also in vertical rows (upper part) and reaching to the external wall; x 8; 4. Longitudinal fracture of the thallus filled up by pyriform tubercles; x 5; 5. Longitudinal fracture of two specimens; a) the left one is observed from outside with no fissuration or annulation; b) the right one shows the central stem and pyriform slightly deformed tubercles; x 5; 6. Longitudinal fracture of the thallus filled up by horizontally arranged single whorls of pyriform tubercles; x 5; 7. Transverse oblique fracture showing pyriform tubercles; x 8; 8. Transverse fracture showing small central stem and pyriform tubercles (6 in the half of section); x 8; 9. Transverse fracture with thick central stem and pyriform tubercles (10 in the half of section); x 8; 10. Longitudinal oblique fracture with large central cavity and pyriform tubercles arranged in single whorls; x 8; 11. Longitudinal fracture of the thallus with inner tube covered by single whorls of pyriform tubercles; x 8; 12. Longitudinal oblique fracture of the thallus, showing the central cavity with pores, penetrating in the pyriform tubercles arranged in single whorls, reaching up to the external wall; x 8; 13. Longitudinal fracture with pyriform tubercles arranged in rare whorls; x 8; 14. Longitudinal fracture with pyriform tubercles arranged in dense whorls; x 8; 15. Longitudinal fracture of the apex of specimen with pyriform tubercles arranged in dense horizontal whorls and forming vertical rows; x 15; 16. Longitudinal fracture of the thallus showing the central cavity, internal wall with pores in dense single whorls and penetrating into tubercles reaching to the external wall; x 5; 17. Longitudinal fracture of the thallus, showing pyriform tubercles arranged in dense single whorls; x 8; 18. Longitudinal fracture of the thallus, showing the surface of the central stem with pyriform tubercles arranged in single whorls; x 8; 19. Longitudinal fracture of the thallus showing the internal wall of central cavity with pores arranged in single whorls and penetrating into tubercles reaching to the external wall; x 5; 20. Longitudinal fracture of the thallus with densely arranged pyriform tubercles in single whorls; x 15; 21. Longitudinal fracture of the thallus with pyriform tubercles arranged in dense single whorls; x 8; 22. Longitudinal fracture with pyriform branches; x 8; 23. Longitudinal fracture of the thallus showing hollow central cavity and pyriform tubercles contacting on the internal wall, arranged in dense single whorls; x 8; 24. Longitudinal fracture of the thallus showing partly preserved pyriform tubercles arranged in dense single whorls; x 5; 25. Longitudinal fracture of the thallus with pyriform branches arranged in dense single whorls; tubercles are small in the lower part of specimen, where they are sitting on the internal wall, and much greater in the upper part, where they reach tu the external wall; x 5.
 |
|
|
Click on thumbnail to enlarge the image.
Plate XII: Physoporella lotharingica (). 1 10Ż Lgota-Nadwarcie, 2, 7, 11, 13, 17, 19 Krupka, 3, 5, 12 Kamyce, 4 Nowa Wioska (Dziewki), 6 37WB Winowno-Będusz (depth 118 m), 8, 11, 15, 18, 20 Czeladź, 9 Balin, 10 Przełajka, 14 25WW Winowno, depth 90.56 m, 16 Stare Gliny.
1. Longitudinal section with largely spaced single rows of pyriform tubercles; x 6.5; 2. Longitudinal fracture of the hollow central cavity with widely spaced single rows of pyriform tubercles; x 10; 3. Longitudinal fracture showing external part with widely spaced single rows of pyriform tubercles; x 6; 4. Longitudinal fracture showing central cavity partly filled up by sediment and with widely spaced single rows of tubercles; x 10; 5. Longitudinal fracture with widely spaced single rows of tubercles; x 12; 6. Oblique fracture showing widely distributed single rows of tubercles; x 6; 7. Longitudinal fracture with widely spaced single rows of tubercles; x 6; 8. Oblique fracture with widely spaced single rows of pyriform tubercles; x 6; 9. Longitudinal fracture with widely spaced single rows of pyriform tubercles; x 10; 10. Longitudinal fracture with partly hollow central cavity and single rows of pyriform tubercles; x 6; 11. Longitudinal fracture showing partly hollow central cavity and single rows of pyriform tubercles; x 10; 12. Transverse fracture showing pyriform tubercles stemming from central cavity filled up by sediment; x 6; 13. Longitudinal fracture showing external part with widely spaced single rows of pyriform tubercles; x 6; 14. Longitudinal fracture showing partly preserved single rows of pyriform tubercles; x 6; 15. Longitudinal fracture showing partly preserved single rows of pyriform tubercles; x 6; 16. Longitudinal fracture with partly obliterated single rows of pyriform tubercles; x 10; 17. Longitudinal fracture showing widely spaced single rows of pyriform tubercles; x 6; 18. Longitudinal fracture with widely spaced single rows of pyriform tubercles; x 6; 19. Oblique fracture with widely spaced single rows of pyriform tubercles; x 10; 20. Four differently oriented specimens with widely spaced single rows of pyriform tubercles; x 6.
 |
|
|
Click on thumbnail to enlarge the image.
Plate XIII: Physoporella praealpina 1, 6 Wojkowice Komorne, 2-3, 7-8 Przełajka, 4-5, 9-17 Czeladź.
1. Tangential-longitudinal fracture of the outer part of the thallus showing 15 double whorls of alternating tubercles; their pyriform shape is visible on sides; some whorls separated by narrow furrows. Wojkowice Komorne, coll. S. ; x 10; 2. Tangential-longitudinal fracture near the outer part of the thallus with faintly visible double whorls of alternating pyriform tubercles; the whorls are closely spaced, without separating furrows; in the lower part of the image, the whorls are covered with a uniform crust of obliterated tubercles; x 10; 3. Tangential-longitudinal fracture along the outer part of thallus, with preserved large parts of the crust; from below, there appear double whorls of closely spaced alternating pyriform tubercles; x 10; 4. Tangential-longitudinal fracture of the apical part of the thallus. In the lower part, three closely spaced double whorls of alternating pyriform tubercles are visible; in the upper part, there are 4-5 closely spaced whorls consisting of single rows. The apical rows are separated from the double rows by a fissuration ridge, above which there are preserved remnants of the first, double row of the apical part; x 12; 5. Tangential-longitudinal fracture of the apical part of the thallus with preserved apex, inclined towards the viewer, 8- 9 closely spaced whorls are visible, of which 7 consist of double rows of alternating pyriform tubercles, and the two apical ones are single. To the right, some tubercles end with openings on top, leading to hollow interior; x 12; 6. Tangential-longitudinal fracture along the outer part of the thallus. Ten double whorls of alternating pyriform tubercles are visible; the whorls are separated by furrows. Specimen from Wojkowice Komorne, coll. S. ; x 10; 7. Tangential-longitudinal fracture close to the outer part of the thallus. Five whorls of alternating pyriform tubercles visible; the three upper rows are double, the other two are quadruple. The distances between whorls are greater in the upper part. Some tubercles open with pores, and are hollow. Traces of undulation are visible on sides, but it is not evident due to the lack of preserved outer wall; x 12; 8. Tangential-longitudinal fracture close to the outer part of the thallus. Under partially preserved crust, there appear 4 densely spaced double whorls of alternating pyriform tubercles; x 12; 9. Tangential-longitudinal fracture of the fragment of the thallus with three whorls consisting of 3, 5, and 2 rows, respectively (from the top down), of alternating pyriform tubercles; x 12; 10. Transverse fracture of the thallus showing almost hollow central cavity, with partly preserved inner wall, with two rows of pores (deeper and shallower one). The tubercles are pyriform (wider inside and thinning conically outwards, towards the partially preserved outer wall; x 12; 11. Transverse fracture of the thallus showing almost completely filled central cavity in the middle, forming a continuous stem, to which ca. 22 pyriform tubercles are attached; the branches are fairly thick, slightly thinning or thickening outwards. They form a double row (some are higher, some are lower); x 12; 12. Tangential-longitudinal fracture of he thallus with three double whorls of alternating pyriform tubercles; x 12; 13. Tangential-longitudinal fracture close to the outer part of the thallus. At least five loosely spaced double whorls of alternating pyriform tubercles are visible. Tubercles end with pores and are hollow. Only below the third whorl from the top there is an empty fissure, bordered with fissuration ridges; x 12; 14. Transverse fracture of the thallus showing hollow, partially filled central cavity around which there are preserved more than 14 tubercles. The tubercles are hollow, and are covered with crust, usually sealing them completely (in the upper part of the picture). Some tubercles (in the right part of the specimen) are cracked in the apical part; x 12; 15. Tangential-longitudinal fracture through hollow central cavity. In the lower part, there are whorls of alternating pores, inlets to tubercles. In the upper part, pyriform tubercles are visible; some of them open with pores on top, and are hollow; x 12; 16. Longitudinal fracture of the thallus slightly above the wall of central cavity. Intusannulation of the upper (outer) surface of the wall is visible. In the furrows, there are double whorls of alternating hollow pyriform tubercles; x 12; 17. Longitudinal fracture of the thallus slightly above the wall of central cavity. Deep intusannulation of its wall is clearly visible. In the furrows, there are double whorls of alternating hollow pyriform tubercles, occasionally encroaching on ridges, thus making them denticulate. In the upper part of the specimen, there is a preserved fragment of its outer side, with double whorls of alternating pyriform tubercles. Sideways, visible are elongated pyriform branches reaching the outer wall which is smooth, without undulation; x 12.
 |
|
|
Click on thumbnail to enlarge the image.
Plate XIV: Physoporella praealpina 1, 7 Czeladź, 2-6, 8-23 Przełajka.
1. Tangential-longitudinal fracture along the outer part of the thallus showing double whorls of alternating pyriform tubercles; some whorls separated by fissuration ridges; x 12; 2. Tangential-longitudinal fracture of the outer part of the thallus. About 10 double whorls of alternating pyriform tubercles are visible, separated with narrow spaces. A hole in the middle of the specimen reveals hollow central cavity. Fragments of the internal wall are visible, overlapping the whorls; x 10; 3. Tangential-longitudinal fracture of the outer part of the thallus showing about 14 double whorls of alternating partly obliterated pyriform tubercles; fragments of the outer wall are preserved; x 10; 4. Tangential-longitudinal fracture of the outer part of the thallus. Intusannulation of the inner surface of the wall facing the central cavity is visible. In furrows, there are 6 double whorls of alternating fairly large pores separated by ridges. Some ridges reveal fissuration cracks; the outer wall is slightly undulating; x 10; 5. Tangential-longitudinal fracture through hollow central cavity. On the inner side of the wall, there are fairly widely spaced double whorls of pores (inlets to tubercles). In the upper part of the specimen, fragment of its outer side is a preserved. In the lower part, there are double whorls of alternating pyriform tubercles. Between the outer and the inner walls, there is a hollow fissure crossed by poorly preserved pyriform tubercles. The outer wall which is smooth, slightly undulating; x 10; 6. Tangential-longitudinal fracture of the outer part of the thallus with densely packed double whorls of alternating pyriform tubercles. Some tubercles open with pores, and are hollow. A hole in the middle of the specimen reveals hollow central cavity; x 12; 7. Tangential-longitudinal fracture of the outer part of the thallus showing widely spaced double whorls of alternating pyriform tubercles; tubercles of the lowest row open with pores, and are hollow; x 12; 8. Tangential-longitudinal fracture of the outer part of the thallus showing widely spaced double whorls of alternating pyriform tubercles; fragments of the encrusted outer wall are preserved; x 12; 9. Oblique fracture of the thallus showing filled central cavity. In its upper part a double whorl of deformed hollow pyriform tubercles is preserved; x 12; 10. Longitudinal fracture of the fragment of the thallus showing its structure on various levels. In the lower part: 8 double whorls of alternating pores, enters the inner wall from the central cavity. The whorls are set in furrows separated by ridges (intusannulation). In the upper part of the thallus, a pattern of denser arrangement of double whorls of alternating tubercles is visible. From the top and left side, the whorls are partially obscured by fragmentarily preserved outer wall. The outer and the inner walls are separated by a hollow fissure crossed by poorly preserved tubercles. The walls are slightly undulating; x 10; 11. Tangential-longitudinal fracture of the outer part of the thallus showing 8 double whorls of alternating pyriform tubercles; x 12; 12. Oblique tangential-longitudinal fracture of the outer part of the thallus with densely spaced double whorls of alternating pyriform tubercles; x 12; 13. Tangential-longitudinal fracture of the outer part of the thallus. In the apical part, the outer wall is partly preserved. Below are exposed 3 densely spaced double whorls of alternating pyriform tubercles; x 10; 14. Longitudinal fracture of the fragment of the thallus showing its structure on various levels. A hole in the middle of the specimen reveals the internal wall of hollow central cavity with whorls of alternating pores previously leading to branches. Below there are double whorls of alternating tubercles. In the lowest part, fragmentarily preserved outer surface is visible. On both sides, there are fissures between the outer and the inner walls, situated against the whorls of pores on the inner surface. These are hollow tubercles; x 10; 15. Transverse fracture of the thallus. The hollow, partially filled central cavity is encircled by fairly thick pyriform tubercles. There are at least 16 of them, arranged in alternating double whorls (higher and lower); x 12; 16. Transverse fracture of the thallus. The almost hollow, central cavity surrounded by thickened wall of the inner tube, with about 22 small pyriform tubercles. The tubercles are separated from the outer wall by a narrow fissure; x 10; 17. Transverse fracture of the thallus. Both inner and outer tubes are partly preserved. Between them are pyriform tubercles (in the upper part of the specimen). In the lower part the tubercles are attached to the outer wall and separated from the inner wall by a hollow fissure. Large central cavity is almost filled with sediment; x 12; 18. Tangential-longitudinal fracture of the thallus with 4 tightly arranged double whorls of alternating tubercles; x 10; 19. Oblique transverse fracture of the thallus with both walls (tubes) preserved, with about 20 fairly thick pyriform tubercles between them. Large central cavity is almost filled with sediment in the upper part and hollow in the lower part; x 12; 20. Transverse fracture of the thallus with filled central cavity surrounded by about 20 pyriform tubercles, usually tapering outwardly; x 12; 21. Oblique transverse fracture of the thallus with hollow central cavity showing intusannulated interior surface of the inner tube. In furrows, there are poorly visible pores (branch inlets). Between the inner and the outer tubercles there are pyriform tubercles, pointed to outside. In the lower part, there are hollow tubercles with pores; x 12; 22. Fracture of the thallus with large hollow tubercles; x 12; 23. Tangential-longitudinal fracture of the thallus with two densely packed double whorls of tubercles. Some have pores, indicating they are hollow inside. On sides of the specimen there are fragmentarily preserved outer walls of the specimen; x 10.
 |
|
|
Click on thumbnail to enlarge the image.
Plate XV: Physoporella praealpina 1-9, 11-19 Rosowa Góra, 10 Przełajka, 21-22 Stare Gliny.
1. Longitudinal fracture of the thallus showing dense double whorls of alternating pyriform tubercles; x 12; 2. Longitudinal fracture of the thallus with partly exposed central cavity, thickened internal wall and relatively largely spaced double whorls of alternating pyriform tubercles; x 12; 3. Longitudinal fracture of the thallus showing 8 double whorls of alternating pyriform tubercles; x 10; 4. Longitudinal fracture of the thallus showing from outside dense double whorls of alternating pyriform tubercles; x 12; 5. Longitudinal fracture of the thallus with alternating pyriform tubercles arranged in double whorls; x 12; 6. Longitudinal fracture of the thallus with alternating pyriform tubercles arranged in double whorls; x 10; 7. Longitudinal fracture of the thallus showing dense double whorls of alternating pyriform tubercles; every two whorls are separated by thin fissure; x 10; 8. Longitudinal fracture of the thallus showing double whorls of alternating pyriform tubercles; x 10; 9. Longitudinal fracture of the fragment of the thallus with alternating pyriform tubercles arranged in double whorls; x 10; 10. Longitudinal fracture of the fragment of the thallus showing alternating pyriform tubercles arranged in double whorls; x 10; 11. Longitudinal fracture of the thallus showing partly hollow central cavity, remnants of internal tube and pyriform tubercles, arranged in the lower part of specimen in relatively largely spaced double whorls with alternating tubercles, reaching (middle part, left) to the external wall; x 10; 12. Longitudinal fracture of the thallus with double whorls of alternating pyriform tubercles; x 10; 13. Longitudinal fracture of the thallus showing relatively largely spaced double whorls of alternating pyriform tubercles; x 12; 14. Longitudinal fracture showing double whorls of alternating pyriform tubercles; x 10; 15. Longitudinal fracture of the thallus with double whorls of alternating pyriform tubercles; x 10; 16. Longitudinal fracture of the thallus with double whorls of alternating pyriform tubercles; x 10; 17. Longitudinal fracture of the thallus showing slightly spaced double whorls of alternating pyriform tubercles; x 10; 18. Longitudinal fracture of the thallus showing relatively largely spaced double whorls of alternating pyriform tubercles; x 12; 19. Longitudinal fracture of the thallus showing slightly spaced double whorls of alternating pyriform tubercles; x 10; 20. Longitudinal fracture of the thallus relatively largely spaced double whorls of alternating pyriform tubercles; x 10; 21. Longitudinal fracture of the thallus showing from inside of the specimen alternating pyriform tubercles; x 10.
22-23. Physoporella pauciforata () Balin.
22. Longitudinal fracture of the thallus with preserved apex, showing dense single whorls of pyriform tubercles, forming also vertical rows; x 12; 23. Longitudinal fracture of the thallus with preserved apex, showing dense single whorls of pyriform tubercles; x 12.
 |
|
|
Click on thumbnail to enlarge the image.
Plate XVI: Physoporella dissita () 1, 5, 10-12, 18, 20, 25-28, 31 Czeladź, 2, 14 Wojkowice Komorne, 3 Jemielnica, 4, 6-9, 13, 15-17, 19, 21-24, 29-30 Przełajka.
1. Longitudinal fracture of the thallus showing the inner tube with rudimentarily preserved pyriform tubercles arranged in double whorls; tubercles reach to the external tube, which is markedly undulated; x 12; 2. Longitudinal fracture of the thallus subdivided in two parts; in the lower part the interior of the central cavity is visible, with fragmentary double whorls of alternating tubercles, exterior wall is undulated; in the upper part of the specimen the central stem is preserved, covered with widely spaced double whorls of pyriform tubercles; x 12; 3. Longitudinal-oblique fracture of the thallus; thick tube with hollow central cavity is composed of double whorls of pyriform tubercles, rudimentarily preserved external wall is slightly undulated; x 12; 4. Longitudinal fracture of the thallus with evidently undulated external tube; x 10; 5. Longitudinal fracture of the thallus showing hollow central cavity, internal tube with rudimentarily preserved pyriform tubercles of double whorls and fragments of external undulated wall; x 10; 6. Longitudinal fracture of the thallus showing the interior of central cavity with alternating pores in double whorls, separated by ridges, forming intusannulation; pyriform tubercles in double whorls are preserved in the upper part of the specimen and on its side, where external slightly undulated wall is partly preserved; x 12; 7. Longitudinal fracture of the thallus with distinct undulation of widely spaced double whorls of alternating pyriform tubercles, fissuration (carbonaceous infilling) is preserved beneath the uppermost whorl; x 12; 8. Longitudinal fracture of the thallus showing hollow central cavity and thickened internal tube with moderately spaced double whorls of alternating pyriform tubercles; undulation is clearly marked; x 10; 9. Longitudinal fracture of the thallus with alternating pyriform tubercles; double rows are spaced; x 10; 10. Longitudinal fracture of the thallus showing regular spaced double whorls of alternating pyriform tubercles; undulation of dissolved external wall is observable; x 12; 11. Longitudinal fracture of the thallus with regularly spaced double whorls of alternating pyriform tubercles; undulation of dissolved external wall is clearly seen; x 12; 12. Longitudinal fracture of the thallus showing double whorls separated by large distances; alternating tubercles are pyriform and form swellings of contact with adjacent rock; x 12; 13. Longitudinal fracture of the thallus with moderately spaced double whorls of pyriform obliterated tubercles; undulation of the external wall is present; x 10; 14. Longitudinal fracture of the fragment of the thallus showing moderately spaced double whorls of alternating pyriform tubercles; x 10; 15. Longitudinal fracture of the fragment of the thallus with double whorls of alternating pyriform tubercles; whorl are separated by relatively large distances; x 10; 16. Longitudinal fracture of the slightly bent thallus, subdivides in two separated parts; in the lower part the internal tube is covered by double whorls of alternating pyriform tubercles, in the upper part of specimen the interior of the central cavity is seen, with preserved interior wall penetrated by the pores arranged in moderately spaced double whorls of pores, separated by the ridges forming intusannulation; external wall is partly undulated; x 12; 17. Transverse fracture of two specimens with central stem, internal wall and pyriform tubercles; x 10; 18. Transverse fracture of the thallus with central stem and pyriform tubercles; x 12; 19. Longitudinal fracture of the slightly bent the thallus showing interior of the central cavity, with relatively widely spaced double whorls of alternating tubercles, separated by the ridges forming intusannulation; external wall is undulated (upper left side); x 10; 20. a) Longitudinal fracture of the thallus showing the interior with double whorls separated by relatively large and flat ridges (intusannulation), undulation of external wall is observable; x 10; b) Transverse section of the thallus with hollow central cavity and internal and external wall joined by pyriform tubercles; x 10; 21. Longitudinal fracture of the thallus with double whorls of pyriform tubercles; x 12; 22. Longitudinal fracture of the thallus with partly uncovered hollow central cavity and double whorls of pyriform tubercles between two undulated walls - internal and external; x 10; 23. Transverse fracture of the thallus with hollow central cavity and two walls joined by pyriform tubercles; x 12; 24. Longitudinal fracture of the thallus filled up by moderately spaced double whorls of alternating pyriform tubercles; x 10; 25. Transverse fracture of the thallus with double whorls of pyriform tubercles; x 12; 26. Transverse fracture of two specimens with central stem and pyriform tubercles; x 12; 27. Longitudinal fracture of the thallus with double whorls of alternating pyriform tubercles; x 10; 28. Longitudinal fracture of the thallus filled up by double whorls of alternating pyriform tubercles; x 10; 29. Longitudinal fracture of two specimens filled up by relatively dense rows of alternating pyriform tubercles and pores; x 10; 30. Transverse fracture of the thallus with central cavity, thickened internal wall and pyriform tubercles; x 12; 31. Longitudinal fracture of the thallus observable from interior central cavity; internal wall with intusannulation composed of double whorls of alternating pores, separated by large flat ridges; tubercles begin from the pores and reach to the undulated external wall; x 12.
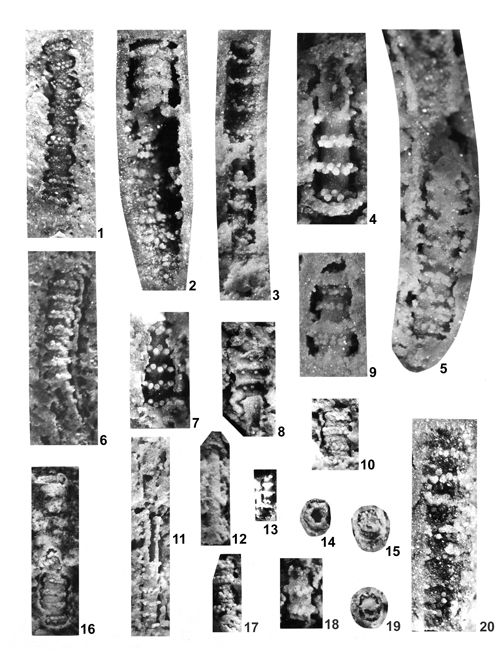 |
|
|
Click on thumbnail to enlarge the image.
Plate XVII: Physoporella polonoandalusica n.sp. 1, 6, 10, 16, 18 Przełajka, 2-3, 5, 9, 17 Stare Gliny, 4, 7-8, 11, 13-15, 19-20 Czeladź, 12 Jemielnica.
1. Longitudinal fracture of the slightly bent the thallus showing double whorls separated by large distances; tubercles are pyriform, alternating; the fragmentary preserved external wall is slightly undulated; x 10; paratype; 2. Longitudinal fracture of the thallus showing in the upper part double whorls of pyriform tubercles separated by large distances set on the central stem; lower part of the specimen shows pyriform tubercles sticking up to the straight external wall; x 12; 3. Longitudinal fracture of the slightly bent the thallus showing in the upper part of specimen double whorls of pyriform alternating tubercles, sticking up to the external wall; in the lower part of specimen the thin central stem is preserved with double whorls of alternating pyriform tubercles; in both parts the whorls are separated by large distances, external wall is straight without undulation; x 10; 4. Longitudinal fracture of the thallus showing rare double whorls of alternating pyriform tubercles; x 12; 5. Longitudinal fracture of the slightly bent the thallus showing rare double whorls of alternating pyriform tubercles; external wall is thick and strait without undulation; x 12, paratype; 6. Longitudinal fracture of the slightly bent the thallus showing rare double whorls of alternating pyriform tubercles, set on the central tube; external wall is flat, but in some places slightly undulated; x 10, holotype; 7. Longitudinal fracture of the thallus showing rare double whorls of alternating pyriform tubercles; external wall is slightly undulated; x 12; 8. Longitudinal fracture of the thallus showing the undulated external wall with rare double whorls of alternating pyriform tubercles (in some whorls tubercles are missing); in the lower part of the specimen the fragment of central tube is preserved; x 10; 9. Longitudinal fracture of the thallus with rare double whorls of alternating pyriform tubercles; x 12; 10. Longitudinal fracture of the thallus showing rare double whorls with alternating pyriform tubercles; x 10, paratype; 11. Longitudinal fracture of the long and narrow the thallus with pyriform tubercles in rare double whorls; x 10; 12. Longitudinal fracture of the thallus showing the central stem with fragmentary preserved rare double whorls of alternating pyriform tubercles; x 10; 13. Longitudinal fracture of the thallus showing the central stem with rare double whorls of alternating pyriform tubercles; x 8; 14. Transverse fracture with hollow central cavity and pyriform tubercles connecting the internal and external walls; x 10; 15. Transverse oblique fracture showing central stem and pyriform tubercles connecting it with thickened external wall; x 10; 16. Longitudinal fracture of the bent the thallus subdivided in two parts; in the lower part one can observe the internal tube with rare double whorls of alternating pyriform tubercles, connecting the central tube with undulated external wall; the upper part shows the external undulated wall with small fragments of the tubercles; in the uppermost part the entrance to the central cavity is preserved; x 12, paratype; 17. Longitudinal fracture of the thallus with rare double whorls of alternating pyriform tubercles; x 10; 18. Longitudinal fracture with pyriform tubercles arranged in double whorls; x 10; 19. Transverse fracture showing the pyriform tubercles connecting the central cavity with external wall; x 10; 20. Longitudinal fracture of the thallus showing the rare double whorls of alternating pyriform tubercles; x 12.
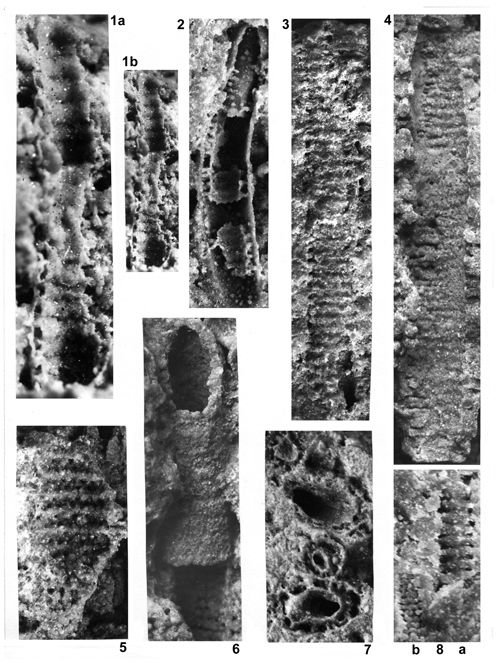 |
|
|
Click on thumbnail to enlarge the image.
Plate XVIII: Physoporella praealpina and Physoporella pauciforata () Boreholes of the Zawiercie region.
1. a and b Physoporella praealpina , borehole 11Ż Żarki Letnisko, depth 128.4-129.2; upper part of Diplopora Dolomite (Illyrian); longitudinal fracture of two thin tubes (inner and outer), connected by pyriform tubercles; outer tube slightly undulated; on the inner tube inlets of double rows of pores are observable; 1a x 10, fig. 1b x 5; from , 1986, Pl. CVI, f. 3; 2. Physoporella praealpina , borehole 23WW Krusin, depth 143.4 m; lower part of Diplopora Dolomite (Pelsonian); longitudinal fracture of outer tube connected with fragments of the inner tube by pyriform tubercles arranged in double rows; x 9; from , 1986, Pl. CVI, 4; 3. Physoporella praealpina , borehole 25WW, Winowno, depth 90.56 m; longitudinal fracture of double rows of pyriform tubercles on the smooth surface of tube; x 9; 4. Physoporella pauciforata () , borehole 25WW, Winowno, depth 90.30-90.35 m; lower part of Diplopora Dolomite (Pelsonian); longitudinal fracture with dense single rows of tubercles; x 8; 5. Physoporella praealpina , borehole 25WW, Winowno, depth 90.35-90.40 m; lower part of Diplopora Dolomite (Pelsonian); longitudinal fracture with double rows of pores on the inner tube; from , 1986, Pl. CVI, 5, 7; x 10; 6. Physoporella pauciforata () , borehole 25WW, Winowno, depth 90.30-90.35 m; x 8; lower part of Diplopora Dolomite (Pelsonian); outer view of single, partly split tube with double rows of alternating pores in the inner surface; x 8; from , 1986, Pl. CVI, 8a; x 10; 7. Physoporella sp., borehole 23WW Krusin, depth 143.4 m; lower part of Diplopora Dolomite (Pelsonian); transverse fractures through double tubes, both parts connected by tubercles; x 9; from , 1986, Pl. CVI, 6; 8. Physoporella praealpina (a) and Physoporella pauciforata () (b); borehole 23WW Krusin, depth 143.4 m; lower part of Diplopora dolomite (Pelsonian); longitudinal fracture with double (a) and single (b) rows of tubercles; x 9; from , 1986, Pl. CVI, 9a, b.
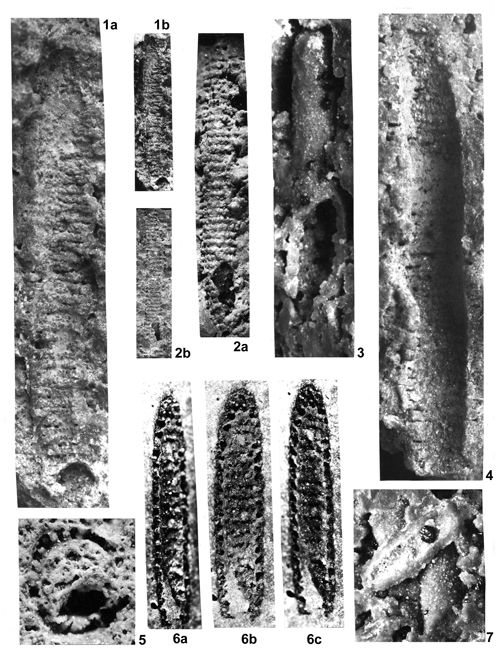 |
|
|
Click on thumbnail to enlarge the image.
Plate XIX: Physoporella praealpina and Physoporella pauciforata () Boreholes of the Zawiercie region.
1. Physoporella praealpina ; borehole 25WW Winowno, depth 90.05-90.15 m; lower part of Diplopora Dolomite (Pelsonian); longitudinal fracture of the external part of tube with dense double rows of pyriform tubercles; 1a x 6.5; 1b x 2; 2. Physoporella praealpina ; borehole 25WW Winowno, depth 90.56 m; lower part of Diplopora Dolomite (Pelsonian); longitudinal fracture of the external surface part of tube with dense double rows of pyriform tubercles; 2a x 9, 2b x 3; from , 1986, Pl. CVI, 2; 3. Physoporella praealpina ; borehole 25WW Winowno, depth 90.56 m; lower part of Diplopora Dolomite (Pelsonian); thick tube with smooth external surface; x 8; from , 1986, Pl. CVI, 1; 4. Physoporella praealpina ; borehole 15WW Winowno, depth 90.30-90.35 m; lower part of Diplopora Dolomite; longitudinal fracture of internal part of tube; x 8; 5. Physoporella praealpina ; borehole 37WB Winowno-Będusz, depth 118.0 m; transverse fracture with pyriform tubercles; x 10; 6. Physoporella pauciforata () ; borehole 37WB Winowno-Będusz, depth 118.0 m; 6a - left side of tube, 6b - tube in horizontal position, 6c - right side of tube; longitudinal fracture with single rows of pores on the internal surface of inner tube; x 6; 7. Physoporella sp. borehole 25WW Winowno, depth 90.56 m; lower part of Diplopora Dolomite; accumulation of thick smooth tubes; x 8; from , 1986, Pl. CVI, 1.
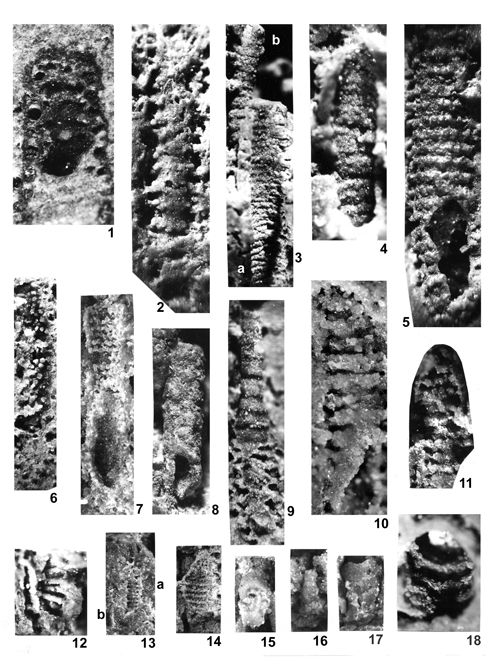 |
|
|
Click on thumbnail to enlarge the image.
Plate XX: Various Dasycladales from boreholes of the Zawiercie region.
1. Kantia comelicana (); borehole 37 WB Winowno-Będusz, depth 118.0 m; upper part of Diplopora Dolomite (Illyrian); longitudinal fracture with rows of vesiculiferous tubercles with clear ramification; x 8; 2. Kantia comelicana (); borehole 37 WB Winowno-Będusz, depth 118.0 m; upper part of Diplopora Dolomite (Illyrian); longitudinal fracture with rows of tubercles in some places showing arrangement in tufts; x 8; 3. a) Physoporella dissita () ; b) Physoporella praealpina ; borehole 31 KW Morsko, depth ca 293.0 m; upper part of Diplopora Dolomite (Illyrian); longitudinal fracture with dense double rows of tubercles (b) and spaced single rows (a); x 8; 4. Physoporella praealpina ; borehole 31 KW Morsko, depth ca 293.0 m; upper part of Diplopora Dolomite (Illyrian); longitudinal fracture of the surface of outer tube with double rows of pyriform tubercles; x 8; 5. Physoporella pauciforata () ; borehole 51 Z Myszków (Nowa Wieś); depth 59.0 m; middle part of Diplopora Dolomite (Pelsonian-Illyrian boundary); longitudinal fracture of the surface of outer tube with single rows of pyriform tubercles; x 8; 6. Physoporella praealpina ; borehole 23 WW Krusin, depth 143.4 m; lower part of Diplopora Dolomite; longitudinal fracture with pyriform tubercles attached from inside to the external tube, arranged in double rows; x 6; 7. Physoporella pauciforata () ; borehole 37 WB Gliniana Góra (Winowno-Będusz), depth 118.0 m; upper part of Diplopora Dolomite; longitudinal fracture with pyriform tubercles attached from inside to the external tube, arranged in single rows; x 6; 8. Physoporella sp. ; borehole 31 KW Morsko, depth ca 293.0 m; upper part of Diplopora Dolomite (Illyrian); external view of a thick tube; x 6; 9. Physoporella dissita () , borehole 51 Z Myszków (Nowa Wieś), depth 59.0 m; middle part of Diplopora Dolomite (Pelsonian-Illyrian boundary); longitudinal fracture with segmentation; x 8; 10. Physoporella praealpina ; borehole 51 Z Myszków (Nowa Wieś), depth 59.0 m; middle part of Diplopora Dolomite (Pelsonian-Illyrian boundary); longitudinal fracture with double rows of pyriform tubercles distinctly spaced; x 10; 11. Physoporella pauciforata () ; borehole 37 WB Gliniana Góra (Winowno-Będusz), depth 118.0 m; upper part of Diplopora Dolomite (Illyrian); longitudinal fracture with pyriform tubercles arranged in single rows; x 8; 12. Diplopora annulatissima , borehole 37 WB Gliniana Góra (Winowno-Będusz), depth 118.0 m; upper part of Diplopora Dolomite (Illyrian); transverse oblique fracture with long and narrow trichophorous tubercles, attached to the internal tube; x 10; 13. Physoporella praealpina (a) and Physoporella pauciforata () ; borehole 23 WW Krusin, depth 1434 m; lower part of Diplopora Dolomite (Pelsonian); longitudinal fracture with double (a) and single (b) rows of tubercles; x 4; 14. Physoporella praealpina borehole 25 WW Winowno, depth 90.35-90.40 m, lower part of Diplopora Dolomite (Pelsonian); longitudinal fracture with double rows of pores on the inner tube; from , 1986, Pl. CVI, 7; x 9; 15. Physoporella minutula () , borehole 25 WW Winowno; depth 90.56 m, lower part of Diplopora Dolomite (Pelsonian); external view of segmented tube; x ca 10; 16. Physoporella minutula () , borehole 25 WW Winowno; depth 90.56 m, lower part of Diplopora Dolomite (Pelsonian); external view of segmented tube; x ca 8; 17. Physoporella minutula () , borehole 25 WW Winowno; depth 90.56 m, lower part of Diplopora Dolomite (Pelsonian); external view of segmented tube; x ca 10; 18. Diplopora annulatissima , borehole 31 KW Morsko, depth ca 293.0 m; upper part of Diplopora Dolomite (Illyrian); oblique fracture with segments and furrows viewed from inside; x ca 10.
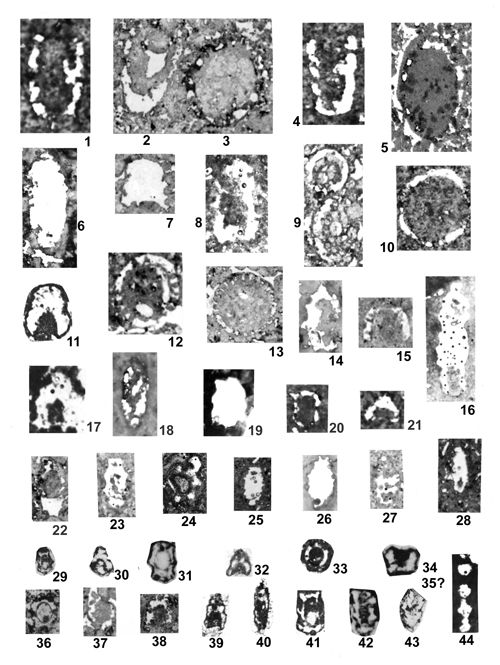 |
|
|
|
|
Click on thumbnail to enlarge the image.
Plate XXI: Various Physoporellae in thin sections. 1-17 Physoporella cf. pauciforata or Physoporella cf. praealpina, 18-28 Physoporella cf. dissita, 29-44 Physoporella cf. minutula.
Localities and numbers of thin sections: 1. 31.1 Boleradź x 8; 2-3. 71.12 Brudzowice x 11; 4. 31.1 Boleradź x 8; 5. 71.14 Brudzowice x 11; 6. 14.4 Łabędy x 14.4; 7. 71.5 Brudzowice x 11; 8. 43.6 Granice x 9; 9. 71.10 Brudzowice x 11; 10. 44.4 Imielin x 10.5; 11. 3 Kamień Śląski x 14; 12. 71.9 Brudzowice x 11; 13. 71.5 Brudzowice x 11; 14. 5.5 Laryszka x 11.3; 15. 73.1 Żelisławice x 9; 16. 62.6 Łosień x 10; 17. 61.3 Łęka x 10; 18. 61.3 Łęka x 9.5; 19. 5.5 Laryszka x 11.3; 20. 62.4 Łosień x 10; 21. 31.1 Boleradź x 8; 22. 71.1 Brudzowice x 9; 23. 62.6 Łosień x 10; 24. 11.3 Segiet 7.8; 25. 52.15 Krupka x 10; 26. 5.5 Laryszka x 11.3; 27. 62.6 Łosień x 10; 28. 52.15 Krupka x 10; 29. 3 Kamień Śląski x 9; 30. 3 Kamień Śląski x 9; 31. 3 Kamień Śląski x 14; 32. 7.6 Nowe Koszyce x 10; 33. 6.1 Suchodaniec x 12; 34. 3 Kamień Śląski x 14; 35. 3 Kamień Śląski x 14; 36. 7.6 Nowe Koszyce x 10; 37. 71.1 Brudzowice x 9; 38. 71.4 Brudzowice x 9; 39. 7.6 Nowe Koszyce x 10; 40. 7.6 Nowe Koszyce x 10; 41. 3 Kamień Śląski x 14; 42. 3 Kamień Śląski x 14; 43. 3 Kamień Śląski x 14; 44. 62.4 Łosień x 10.
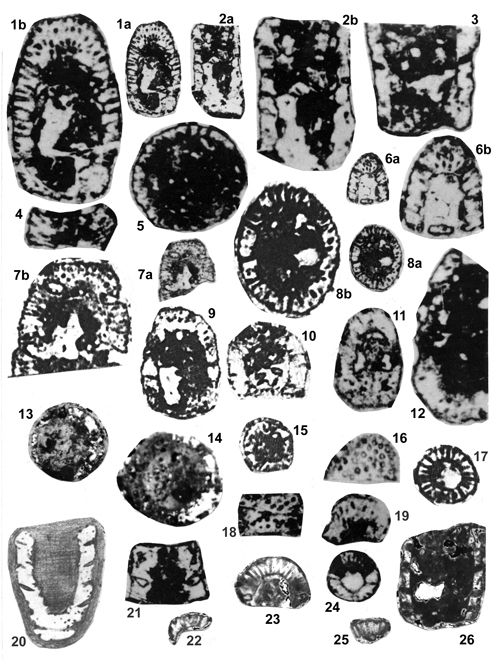 |
|
|
Click on thumbnail to enlarge the image.
Plate XXII: Kantia dolomitica () . Thin sections. 1-12, 16-19, 21, 24 Kamień Śląski, 13-15 Srebrna Góra, 20 Libiąż, 22-23, 25-26 Żarki Letnisko 11Ż.
1. Oblique longitudinal section of a thick crust with vesiculiferous branches and tufts; a x 6; b x 12; 2. Longitudinal section showing distinct dense segmentation; a x 6; b x 12; 3. Longitudinal section showing rare segmentation; x 12; 4. Longitudinal section of a segment with vesiculiferous branches; x 12; 5. Longitudinal section of a thin crust with endings of vesiculiferous branches; x 12; 6. Oblique longitudinal section of a thick segmented crust; a x 6; b x 12; 7. Oblique longitudinal section of a thick segmented crust with dense pores, locally diverging and forming clusters; a x 6; b x 12; 8. Oblique longitudinal section with thick vesiculiferous pores; a x 6; b x 12; 9. Oblique longitudinal section of a segmented crust with pore clusters (branch tufts); x 12; 10. Oblique longitudinal section; x 12; 11. Oblique longitudinal section with pore clusters; x 12; 12. Oblique longitudinal section of a thick segmented crust; x 13; 13. Oblique transverse section of a segmented crust of variable thickness; x 10; 14. Transverse section with vesiculiferous pores; x 10; 15. Transverse section with vesiculiferous pores; x 10; 16. Oblique section with thick pores; x 10; 17. Transverse section with vesiculiferous pores; x 10; 18. Longitudinal section with clusters of thick pores; x 10; 19. Oblique longitudinal section with vesiculiferous pores; x 10; 20. Oblique longitudinal section of segmented thallus with indistinct vesiculiferous pores; Specimen of (1920, Pl. 5, 20) from Libiąż; x 10, reproduced by (1986, Pl. CV, 2); 21. Longitudinal section showing segmentation; x 10; 22. A fragment of transverse section with vesiculiferous pores, borehole 11 Ż Żarki Letnisko, depth 128.4-129.2 m; x 12.6; 23. A fragment of transverse section with vesiculiferous pores, borehole 11 Ż Żarki Letnisko, depth 128.4-129.2 m; x 12.6; 24. Transverse section with vesiculiferous pores; x 6; 25. A fragment of longitudinal section with vesiculiferous pores, borehole 11 Ż Żarki Letnisko, depth 128.4-129.2 m; x 12.6; 26. A fragment of longitudinal section of a segmented thallus with indistinct vesiculiferous pores, borehole 11 Ż Żarki Letnisko, depth 128.4-129.2 m; x 12.6.
|
|
|
|
Click on thumbnail to enlarge the image.
Plate XXIII: Kantia comelicana (). 1, 3-8, 11-12, 14-16, 20 Balin, 2 Przełajka, 9-10, 13, 17-19 Rosowa Góra.
1. Longitudinal fracture of the thallus with partly filled central cavity; internal wall very thin, tubercles are double, diverging, reniform (kidney-shaped), arranged in whorls; x 8; 2. Longitudinal fracture of the thallus showing hollow central cavity and two walls - internal and external, connected with vesiculiferous tubercles; frequently diverging; x 8; 3. Longitudinal fracture of the thallus filled up by vesiculiferous tubercles arranged in dense single whorls; x 4; 4. Longitudinal fracture of the thallus showing partly filled central cavity and internal wall with started from it vesiculiferous reniform tubercles, in some places evidently double, and diverging; x 8; 5. Longitudinal fracture of the thallus filled up by vesiculiferous tubercles arranged in dense single whorls; x 8; 6. Longitudinal fracture of the thallus filled up by vesiculiferous tubercles, arranged in single whorls, sometimes with cross-setting; some tubercles are double, diverging, hollow with pore on the top; x 8; 7. Transverse fracture of two specimens with central stem, surrounding by vesiculiferous tubercles; on the top of upper specimen one can observe triple diverging branch; x 8; 8. Longitudinal oblique fracture with perfectly visible double diverging vesiculiferous tubercles, very quickly expanding distally; x 8; 9. Longitudinal fracture of the external part of the thallus, passing close to the proximal part of tubercles, grouped in clusters of four and three; x 8; 10. Longitudinal fracture of the thallus, passing through the clusters of tubercles of four or rarely three; x 8; 11. Longitudinal fracture of the thallus filled up by single whorls of double diverging vesiculiferous tubercles, grouped in some places on the inner wall in clusters of four and three; x 10; 12. Oblique fracture showing the hollow central cavity and internal wall with vesiculiferous reniform tubercles, in some places double and diverging; x 8; 13. Transverse oblique fracture with vesiculiferous reniform double diverging tubercles; x 8; 14. Oblique fracture with vesiculiferous reniform tubercles, set on the internal tube; x 10; 15. Longitudinal oblique fracture with vesiculiferous, slightly oblique tubercles; x 4; 16. Longitudinal fracture of the thallus with single whorls of vesiculiferous tubercles, in upper right part one can observe reniform double diverging tubercles; x 8; 17. Longitudinal oblique fracture of the thallus with partly hollow central cavity and inner tube with single whorls of vesiculiferous tubercles, merging with adjacent rock; x 8; 18. Longitudinal oblique fracture with central stem and vesiculiferous reniform tubercles; x 8; 19. Transverse oblique fracture showing central stem and vesiculiferous reniform double diverging tubercles; x 10; 20. Longitudinal oblique fracture with partly preserved internal tube and vesiculiferous double diverging tubercles; x 8.
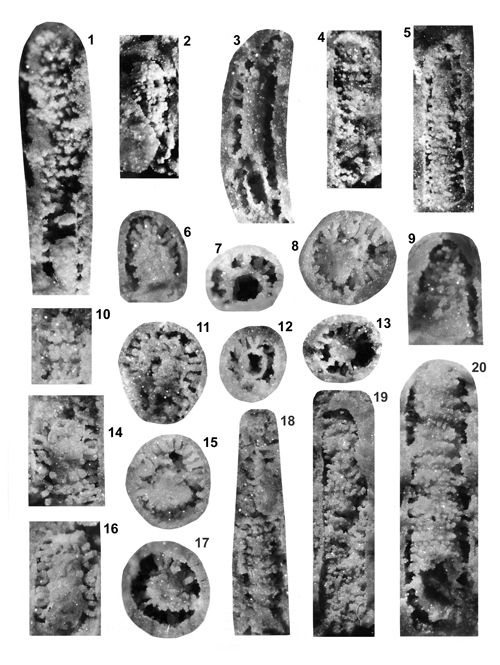 |
|
|
Click on thumbnail to enlarge the image.
Plate XXIV: Kantia comelicana (). 1 Balin, 2-20 Stare Gliny.
1. Longitudinal fracture of the thallus showing single whorls regularly spaced composed of tubercles grouped in clusters by four and three (upper and middle part of specimen where they are cut near their proximal part, close to the internal wall); in the lower part, the internal tube is partly preserved with vesiculiferous tubercles starting from this surface, where the alternating arrangement of proximal parts of tubercles may be observable; x 10; 2. Longitudinal fracture of the thallus filled up by whorls of clusters with bifurcating tubercles; x 10; 3. Longitudinal fracture of the thallus with partly broken internal tube; from the surface of the tube grow the vesiculiferous tubercles, reaching up to the wall of external tube; in the upper left side of the specimen trichotomous branch are visible; x 8; 4. Longitudinal fracture of the thallus filled up by vesiculiferous tubercles, arranged in single whorls (lower part of the specimen) and forming clusters of three and four (upper part) and typical alternating arrangement; x 8; 5. Longitudinal fracture of the thallus filled up by vesiculiferous tubercles arranged in single dense whorls; tubercles reach up the wall of external tube and in some places stick to it; x 8; 6. Oblique fracture with vesiculiferous reniform tubercles, starting from the surface of inner tube and reaching up to the wall of external tube; x 10; 7. Transverse fracture showing hollow central cavity, the thickened inner tube and vesiculiferous bifurcating tubercles, reaching to the external tube; x 12; 8. Transverse fracture showing the central stem and perfectly preserved tubercles with triple and double divergences; tubercles reach up to the external tube and are partly adhering to it in some places; x 12; 9. Longitudinal fracture of the thallus filled up by tubercles grouped in clusters of four and rarely three; x 10; 10. Longitudinal fracture of the thallus filled up with tubercles, grouped in clusters of five, four and three, showing also typical alternating arrangement; x 10; 11. Oblique fracture with long vesiculiferous tubercles, starting from the internal tube and reaching up to the external wall; x 12; 12. Transverse fracture showing partly filled central cavity, internal tube and vesiculiferous reniform bifurcating tubercles, reaching up to the external wall; x 10; 13. Transverse fracture showing the central stem and vesiculiferous reniform tubercles reaching up to the external wall; x 12; 14. Longitudinal fracture of the thallus with the central stem and vesiculiferous reniform tubercles, bifurcating in the upper part of specimen, where they reach up to the external wall, and grouped in clusters of four and three in the lower part; x 10; 15. Transverse fracture showing the filled central stem surrounded by vesiculiferous reniform tubercles, in some places bifurcating and reaching to the external wall; x 12; 16. Longitudinal fracture of the thallus showing the internal wall with traces of clusters of four, attached to it and vesiculiferous reniform double and triple diverging tubercles, reaching up to the external wall; x 12; 17. Transverse fracture showing the filled central stem and attached to it vesiculiferous reniform double diverging tubercles, reaching up to the external wall; x 12; 18. Longitudinal fracture of the thallus, filled up by tubercles arranged in single whorls and grouped in clusters of four and three; x 8; 19. Longitudinal fracture of the thallus, filled up by vesiculiferous tubercles, arranged in dense single whorls and grouped in clusters of four and three; x 8; 20. Longitudinal fracture of the thallus, filled up by dense single whorls of vesiculiferous tubercles, reaching up to the external wall; x 10.
 |
|
|
Click on thumbnail to enlarge the image.
Plate XXV: Photographs of thin sections (1-18) and fracture (19) Diplopora annulata (), Diplopora uniserialis , Kantia dolomitica () and Kantia uniserialis ().
1. Diplopora annulata () - longitudinal oblique section of regularly annulated specimen with 15 segments; trichophorous branches arranged in two or more rows in each segment and grouped in whorls; x 8; locality probably no. 11, Segiet, , 1920: Pl. 5, 16; , 1986, Pl. CV, 1; 2. Diplopora annulata () - longitudinal oblique section of regularly annulated specimen with 11 segments; trichophorous branches arranged in two rows in each segment and grouped in whorls; x 12.6; locality 11 Ż, Żarki Letnisko; , 1986, Pl. CV, 3; 3. Diplopora uniserialis - longitudinal oblique section, densely annulated; x 12.6; locality no. 11 Ż, Żarki Letnisko; 4. Diplopora cf. annulata - longitudinal section, rarely annulated with trichophorous branches x 12.6; locality 11 Ż, Żarki Letnisko; 5. Diplopora cf. uniserialis - longitudinal oblique section, densely annulated; x 10.2; locality no. 5, Tarnów Opolski; 6. Diplopora annulata () - oblique section; x 12.6; locality 11 Ż, Żarki Letnisko; , 1986, Pl. CV, 6; 7. Kantia cf. dolomitica - longitudinal oblique section, rarely annulated, with trichophorous branches; x 12.6; locality 11 Ż, Żarki Letnisko; 8. Diplopora cf. annulata - transverse section; x 12.6; locality 11 Ż, Żarki Letnisko; 9. Diplopora annulata () - longitudinal oblique section with several rows of branches in each segment; x 12.6; locality 11 Ż, Żarki Letnisko, , 1986, Pl. CV, 4; 10. Diplopora annulata () - oblique section of one large segment with trichophorous branches; x 12. 6; locality 11 Ż, Żarki Letnisko; 11. Diplopora uniserialis () - longitudinal oblique section of characteristic segments; x 10, locality no. 13, Srebrna Góra; 12. Diplopora uniserialis () - longitudinal oblique section of regular characteristic segments; x 10, locality no. 13, Srebrna Góra; 13. Diplopora uniserialis () - three sections differently oriented, x 10.2; locality no. 2, Tarnów Opolski; 14. Diplopora uniserialis () - transverse section with trichophorous branches; x 9; locality no. 3, Kamień Śląski; 15. Kantia uniserialis () - transverse oblique section with trichophorous branches; x 9; locality no. 3, Kamień Śląski; 16. Kantia uniserialis () - transverse section with trichophorous branches; x 9; locality no. 3, Kamień Śląski; 17. Kantia uniserialis () - longitudinal tangential section densely annulated; x 9, locality no. 5 Laryszka; 18. Diplopora annulata () - longitudinal tangential section of five segments with two or more rows of vesiculiferous branches, grouped in whorls; a x 9, b x 14; locality no. 3, Kamień Śląski; 19. Diplopora cf. annulata () - longitudinal fracture of external part of specimen with large segments separated by intusannulated fissures (left center of photograph); close to it small fragment of Physoporella polonoandalusica n.sp. with widely spaced double alternating rows of pyriform tubercles; in the right part of the photograph several fragments of Physoporella praealpina and Physoporella polonoandalusica n.sp. are visible; x 9, locality no. 30, Wojkowice Komorne, collected by S.
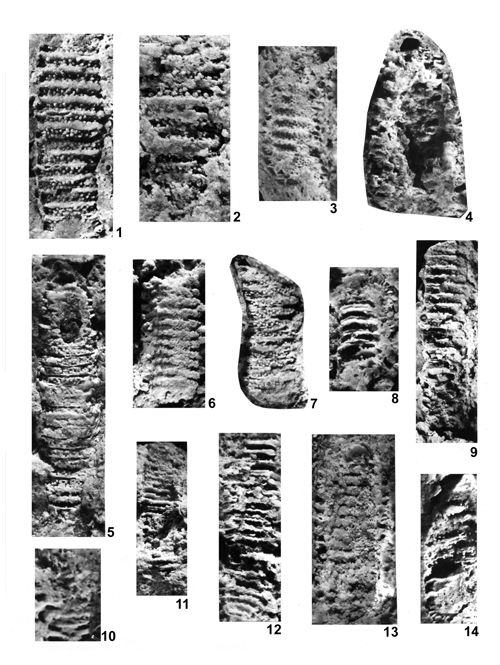 |
|
|
Click on thumbnail to enlarge the image.
Plate XXVI: Diplopora annulatissima . 1-14 Jemielnica.
1. Longitudinal tangential fracture containing 15 annuli, separated with fissuration lists rounded and closed externally. Four lists are broken showing the interior of the interannular furrow. In the annuli usually two rows of alternating small tubercles are visible. Some of them (in the lower part) are hollow. Partly preserved is the outer crust of the alga, connecting annuli with furrows; x 8; 2. Longitudinal tangential fracture. Annuli of alternating tubercles separated with fissuration lists; x 8; 3. Longitudinal fracture near the central cavity. Visible are branch inlets (canals) in transverse depressions, forming whorls consisting of 2-3 rows, separated with ridges without pores (intusannulations) In the upper part of the specimen there are clusters of pores (4-5-7). On both sides of the thallus there are interannular furrows with partly preserved lists; x 8; 4. Oblique-longitudinal fracture with visible central cavity. On the left side there are annuli separated with furrows; x 8; 5. Longitudinal, slightly oblique fracture, consisting of 26 annuli, separated with narrow furrows. The annuli are partly covered with calcareous crust from outside. The crust is mostly lacking, revealing densely spaced tiny alternating tubercles ranged in two or three rows. In the upper part of the specimen there are elongated tubercles thinning distally. In the lower part of the specimen the tubercles are thicker, with an opening (hollow). The interannular furrows are empty and separated by lists from annuli. In the upper part of the specimen a central cavity is visible with poorly pronounce intusannulation; x 8; 6. Longitudinal tangential fracture with 12 annuli filled with slightly obliterated tubercles. At places one can see clusters of 4-5 tubercles. On the upper side of the specimen preserved are a few fissuration lists; x 8; 7. Slightly oblique longitudinal tangential fracture, consisting of 12 annuli with tiny tubercles forming 2-3 alternating roves. Annuli separated with fissures mostly covered by fused lists; x 8; 8. Tangential fracture with well visible six slightly oblique interannular furrows bordered from below and from above by slightly bent lists. The fracture passes closely to their closure from the central cavity side. In the lower part there are slightly oblique tubercles in open annuli; x 8; 9. Tangential fracture consisting of more than 20 annuli separated with furrows. In some places the furrows are closed near the central cavity. Usually, however, the fissuration lists are broken and their slightly undulating surfaces are visible. In the lower part of the specimen the annuli are slightly oblique; x 8; 10. Longitudinal tangential fracture with annuli partly closed from outside. Interannular furrows enter deeply into the thallus and are slightly oblique; x 8; 11. Longitudinal tangential fracture passing near the central cavity, thus showing rounded closures of the interannular furrows; x 8; 12. Longitudinal tangential fracture slightly oblique with about 20 annuli. The fracture passes in the upper part through the rounded closures of the interannular furrows. In the middle part the fracture crosses the furrows and the lists surrounding annuli are visible. They have undulating surface connected with the arrangement of branches. In the lower part of the specimen the surface of the crust is visible from the side of the central cavity, slightly undulated. In transverse depressions there are inlets of alternating canals; x 8; 13. Longitudinal fracture through more than 15 annuli passing through the inner surface of the crust with visible inlets of alternating pores (branches). On both sides of the specimen there are slightly oblique annuli separated with furrows. The apex is visible in the upper part of the thallus; x 8; 14. Oblique longitudinal fracture with annuli and furrows. On the left side of the specimen it is visible that the crust surrounds both the annuli and some furrows; x 8.
 |
|
|
Click on thumbnail to enlarge the image.
Plate XXVII: Diplopora annulatissima . 1-13 Jemielnica.
1. Longitudinal fracture tangential to the outer surface. From below the crust, densely spaced annuli emerge with double whorls of tubercles and lists bordering the interannular furrows; x 8; 2. Longitudinal fracture tangential to the outer surface. There are annuli with double whorls of small pointed tubercles separated by full interannular furrows. Some annuli in the lower part of the specimen are closed from outside; x 8; 3. Longitudinal fracture, tangential to the inner surface. Pores (branch inlets) ranged in whorls. On both sides of the thallus there are infilled interannular furrows. Some of them are hollow or partly filled; x 8; 4. Longitudinal fracture with concave surface of the wall of central cavity with pores (branch inlets) in transverse furrows arranged in whorls. On both sides of the thallus there are evenly spaced slightly oblique interannular furrows, usually completely filled. On the left side of the thallus the outer crust connects annuli with interannular furrows; x 8; 5. Longitudinal fracture passing through the middle of annuli with densely spaced tubercles arranged in clusters of 3-4 branches, usually hollow. On the right side of the specimen there are visible infillings of interannular furrows; x 10; 6. Oblique fracture showing annuli from the side and from below. In the annuli the tubercles are densely packed in several rows. Whorls consisting of 4-6 tubercles are visible; x 10; 7. Longitudinal tangential fracture close to the outer surface of the thallus. From below the outer crust there appear several annuli consisting of 2-3 rows of alternating tubercles. The annuli are separated with thin rows, partly hollow, bordered from both sides by interannular lists to with the tubercles adhere; x 10; 8. Oblique fracture with central cavity completely filled and surrounded by wall reached by annuli and interannular furrows, possessing also an outer wall; x 8; 9. Oblique fracture showing the annuli from the side and below separated with fissuration lists; x 8; 10. Oblique fracture showing empty central cavity reached by rounded ribbed ends of semicircular fissuration lists. The tubercles are clustered at the base; x 10; 11. Longitudinal fracture near the contact of central cavity with annuli and ribbed interannular furrows. In the upper part of the specimen infilling of the central cavity is visible. Also the slightly oblique interannular furrows are filled. Between them there are scattered tubercles in the annuli. Both the annuli and furrows in the upper left part of the specimen are connected from outside by the outer wall; x 8; 12. Group of the specimen in different positions. a) Oblique fracture tangential to the rounded surface of the specimen near the apex. The rare annuli consisting of elongated thin slightly oblique tubercles arranged in double rows. The annuli are separated with furrows surrounded by rounded lists, between them the rare partially empty spaces. b) Fragment of specimen with ribbed rounded closures of interannular furrows from inside of the thallus and with annuli full of tubercles. c) Fragment of the specimen seen obliquely from inside. Rounded closures of interannular furrows are visible; between them the rare scattered tubercles. Interannular lists are thin in the middle and they thicken toward the outside. d) Longitudinal fracture from the center of the alga. Visible are rounded, ribbed closures of the interannular furrows and thin branches in the annuli; x 8; 13. Group of the specimens in different positions. a) Transverse fracture through the thallus with filled central cavity and thin tubercles ranged in alternating rows. b) Longitudinal fracture from the middle of the thallus. Intusannulation is visible, with pores in transverse furrows. c) Oblique longitudinal fracture; visible are ribbed infillings of interannular furrows. In open annuli there are scattered tubercles tightly adhering to fissuration lists. d) Transverse fracture through the thallus with infilled central cavity surrounded by annulus with numerous thin tubercles; x 8.
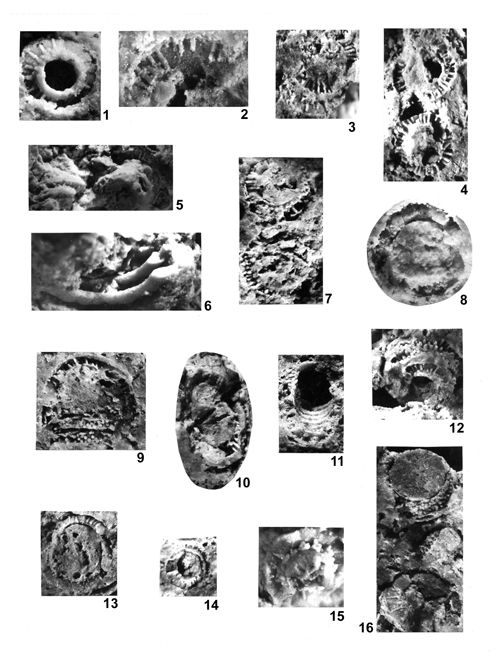 |
|
|
Click on thumbnail to enlarge the image.
Plate XXVIII: Diplopora annulatissima . 1-16 Jemielnica.
1. Transverse fracture through the alga with hollow central cavity. The surface of the list bordering the interannular furrow is visible, with long and thin trichophorous tubercles. Only one row of a double whorl is preserved. Traces of tubercles belonging to the second whorl are partly preserved only near the central cavity opening. A few tufts made of 4-5 branches are visible; x 10; 2. Fragment of the transverse fracture with trichophorous tubercles laying on the surface of the interannular list. The annulus is bordered from outside by a partly preserved wall; x 12; 3. Oblique-transverse with two whorls of thin trichophorous tubercles; x 8; 4. Transverse fracture through two specimens. Two walls are visible, the inner and the outer one, and between them there is an annulus filled with thin trichophorous tubercles, locally slightly swelling outwards, but a closed before the outer wall. Central cavity only partially filled; x 8; 5. Oblique-transverse through two specimens with filled central cavity, forming a thick column, and ribbed interannular furrows. Thin trichophorous tubercles are visible in open annuli; x 10; 6. Oblique fracture with smooth closures, shaped as convex ribs, fully filled interannular furrows, seen from below and from inside. In the open annuli tubercles are locally preserved; x 12; 7. Transverse fracture through two specimens. In both specimens the central cavity is partly filled. Both walls, inner and outer, are visible; between them there is an annulus consisting of a double whorl of thin trichophorous tubercles. Some of them are slightly swollen at the ends; x 8; 8. Transverse fracture of the thallus with large, filled central cavity. Around the cavity there is the annulus with few preserved tubercles lying on a slightly convex surface of the fissuration list; x 12; 9. Oblique transverse fracture. In the upper part there is a semicircular filled central cavity, surrounded by an annulus bordered by two walls - inner and outer ones. Between them there are few thin tubercles. In the lower part there are two cross-cut annuli densely filled with alternating tubercles, arranged in two or three rows, separated by empty furrows with visible fissuration lists; x 10; 10. Oblique fracture through the specimen with filled central cavity. To the right, there are two annuli with long thin tubercles. Some of them form tufts (three to four tubercles) and some fork dichotomously. The annuli are separated with the filled furrow sharply closed from inside; x 8; 11. Oblique fracture with a view into the empty central cavity. In the lower part there are semicircular ribbed closures of interannular furrows, and small tubercles lay between them. In the outer part of the specimen there are ribbed fillings of interannular furrows, and between them there are open annuli filled with long, thin tubercles; x 8; 12. Oblique view from below on a specimen with hollow central cavity and annuli separated with filled furrows. In the annuli there are densely spaced, slightly oblique trichophorous tubercles. The specimen is partly covered with an outer crust, covering both the furrows and the annuli; x 8; 13. Transverse fracture with a view on to a filled large central cavity, surrounded by an annulus with a long, outwardly thinning tubercles, lying on a convex surface of fissuration lists; x 10; 14. Oblique transverse fracture of a specimen with a filled central cavity surrounded by an annulus with minute thin tubercles; x 8; 15. Oblique transverse fracture with a view on filled stem of a central cavity surrounded by an annulus of thin and long tubercles grouped by four to six. They lie on a wide, smooth fissuration list; x 10; 16. Oblique transverse fractures through two specimens. In the upper specimen a large, filled central cavity is visible, with partly preserved inner wall. From below it is adhered by annuli with tiny densely spaced alternating tubercles in double whorls. A broken hollow furrow is visible, consisting of two lists with a rounded closure. In the lower specimen, a moderately wide central cavity is surrounded by a flat surface of a interannular list upon which lie long and thin, outwardly thinning trichophorous tubercles; x 10.
 |
|
|
Click on thumbnail to enlarge the image.
Plate XXIX: Diplopora annulatissima . Photographs of thin sections from various localities.
1. Longitudinal section of annulated specimen with pores separated by fissures, (, 1920, Pl. XXI, 2, description p. 275), also reproduction , 1986, Pl. CV, 9; x 16; locality no. 16, Deep Friedrichstolle (Fryderyk Deep Shaft); 2. Oblique longitudinal section with regular annulation (9 segments) and pores grouped in whorls; , 1920, Pl. XXI, 1, reproduced in , 1986, Pl. CV, 8; x 12; locality no. 7, Nowe Koszyce; 3. Longitudinal section with regular annulation; x 12; locality no. 6, Suchodaniec; 4. Longitudinal-oblique section with regular annulation; x 12; locality no. 6, Suchodaniec; 5. Longitudinal section with regular annulation (more than 10 segments); x 12; locality no. 7, Nowe Koszyce; 6. Longitudinal section with regular annulation (10 segments); x 12; locality no. 7, Nowe Koszyce; 7. Longitudinal section with regular annulation; x 12; locality no. 6, Suchodaniec; 8. Oblique section, x 10, locality no. 7, Nowe Koszyce; 9. Transverse section with trichophorous branches, x 10, locality no. 7, Nowe Koszyce; 10. Longitudinal section with regular annulation, x 10, locality no. 7, Nowe Koszyce; 11. Longitudinal section of three good preserved segments; x 12; locality no. 5, Laryszka; 12. Longitudinal oblique section with regular annulation; x 9; locality no. 6, Suchodaniec; 13. Longitudinal oblique section with traces of annulation; x 9. 5; locality no. 8, Jemielnica; 14. Longitudinal oblique section, x 10, locality no. 7, Nowe Koszyce; 15. Longitudinal tangential section, x 9, locality no. 7, Nowe Koszyce; 16. Longitudinal oblique section with regular annulation; x 7; locality no. 6, Suchodaniec; 17. Longitudinal section of three segments, x 12, locality no. 7, Nowe Koszyce; 18. Longitudinal section with regular annulation, x 9, locality no. 7, Nowe Koszyce; 19. Longitudinal tangential section, ; x 12; locality no. 5, Laryszka; 20. Longitudinal tangential section with pores; x 12.6; locality 11 Ż, Żarki Letnisko, depth 128.4 -192.2 m; from , 1986, Pl. CV, 10; 21. Longitudinal section of one segment with pores; x 12.6; locality 11 Ż, Żarki Letnisko, depth 128.4 - 192.2 m; from , 1986, Pl. CV, 11; 22. Longitudinal section of one segment with pores; x 12.6; locality 11Ż, Żarki Letnisko, depth 128.4 - 192.2 m; from , 1986, Pl. CV, 12; 23. Longitudinal section with regular annulation; x 12.6; locality 11 Ż, Żarki Letnisko, depth 128.4 - 192.2 m; 24. Longitudinal section of one segment with pores; x 14; locality no. 3, Kamień Śląski; 25. Longitudinal section of one segment with pores; x 9; locality no. 3, Kamień Śląski; 26. Longitudinal section of one segment with pores; x 9; locality no. 3, Kamień Śląski; 27. Longitudinal section of one segment with pores, in the upper part transverse section of Oligoporella elegans () ; x 9; locality no. 3, Kamień Śląski; 28. Longitudinal section of two relatively large segments, belonging probably to Diplopora annulata (), x 9, locality no. 7, Nowe Koszyce; 29. Longitudinal section with regular annulation; x 9.8; locality no. 3, Kamień Śląski; 30. Oblique section with segmentation; locality no. 50, Balin; x 9; reduced reproduction of figure by (1992, Pl. VII, 2).
 |
|
|
Click on thumbnail to enlarge the image.
Plate XXX: 1-4. Diplopora annulatissima Czeladź Przełajka.
1. Oblique-transverse fracture. In the upper part, on the fissuration lists there are long outwardly thinning tubercles. Below there are variably positioned tubercles. Specimen of S. (MuzPIG 135.II.53 from Czeladź); x 12; 2. Lateral view of long thin tubercles branching at the base; x 12; 3. Transverse fracture. From partly filled central stem there spread thin trichophorous tubercles; x 10; 4. Oblique-transverse section. Long tubercles spread from tufts of 4-6, in the lower part of the specimen, and are thinning distally, reaching the outer wall; x 12.
5-17. Clavapora clavaeformis () Przełajka.
5. Longitudinal fracture. The hollow central cavity is reached by annuli and interannular furrows. They are often hollow, separated by fissuration lists; x 10; 6. Longitudinal fracture. Interior of the central cavity is visible, with dense transverse furrows and ridges (intusannulation). In the furrows there are inlets of branches (pores). The central cavity, rounded in the upper part (apex) is bordered by dense annuli and interannular furrows, partly hollow; x 10; 7. Longitudinal tangential fracture through annuli and interannular furrows. In the annuli there are locally visible tubercles, grouped in tufts. Fissuration lists are locally with dented edges, resulting from the arrangement of tubercles; x 10; 8. Longitudinal tangential fracture passing through the annuli and interannular furrows and cutting into a fissure-like central cavity. Locally there are tufts consisting of 3-5 branches (tubercles); x 10; 9. Longitudinal fracture through the interior of the central cavity with densely spaced transverse furrows and ridges. Seen from above, the wall of the central cavity is covered with dense rows of pores. Indented fissuration lists are connected with the arrangement of pores (branch inlets). To the right of the specimen there are densely spaced annuli and interannular furrows, partly hollow, separated by fissuration lists. The whole system of furrows and annuli (annulation), reaches the well visible inner wall. From outside the alga edge is frayed; x 12; 10. Longitudinal fracture through the narrow central cavity, bordered from a sides and from above by dense annuli and interannular furrows; x 8; 11. Longitudinal section through the right edge of a thallus with a annuli and interannular furrows. Locally, long thin outwardly pointed trichophorous tubercles are visible. The furrows are usually hollow, oval; x 10; 12. Longitudinal section through partly hollow central cavity, bordered from sides and from the top (apex), with a system of annuli and interannular furrows. Long thin pointed trichophorous tubercles are locally forking dichotomously. The edges are frayed; x 12; 13. Longitudinal tangential section through a set of annuli and interannular furrows. Indented edges of fissuration list are visible, related to the arrangement of adjacent tubercles. In several places tubercles are visible, sometimes doubled; x 12; 14. View on the interior of the central cavity. Intusannulation of the inner wall is visible - in the transverse furrows there are pores (branch inlets), locally grouped in tufts. The left edge of the thallus is frayed; x 10; 15. View on the interior of the central cavity with intusannulated inner wall. Pores are situated in the transverse furrows, but some occur also on ridges separating them. Locally the pores (tubercles) are grouped in tufts of 3-5 tubercles; x 12; 16. Longitudinal fracture through the hollow central cavity and surrounding it set of annuli and interannular furrows. The furrows are hollow and the annuli have denticulate fissuration lists with tubercles; x 10; 17. Longitudinal fracture passing through the central cavity and the set of annuli and interannular furrows surrounding it from the sides and from below. The central cavity is partly filled with preserved convex inner wall seen from outside. The annulation is very dense. In the annuli there are tiny tubercles situated on fissuration lists and making them denticulate; x 10.
 |
|
|
Click on thumbnail to enlarge the image.
Plate XXXI: Clavapora clavaeformis () Przełajka.
1. Longitudinal fracture, passing through the central cavity and surrounding set of annuli and interannular furrows. Visible intusannulation of the internal wall, consisting of transverse furrows and ridges. In furrows there are rows of alternating pores, locally entering slopes of the ridges. Note wide central cavity. Annulation is very dense, on the sites perpendicular to the central cavity, and oblique in the upper part forming apex. In the annuli there are dense spaced small alternating tubercles. The sides of the thallus are frayed and no distinct outer wall is visible; x 12; 2. Enlargement of the middle part of photo 1. Clearly visible is intusannulation of the wall of the central cavity from inside. Alternating pores form rows not only in transverse furrows but also entering slopes of the ridges. In the annuli of the sites of the thallus there are clearly visible rows of tubercles tightly adjacent to the fissuration lists, making them indented. Long thin trichophorous tubercles are visible on the left side of the specimen; x 20; 3. Longitudinal fracture of two specimens. In the upper one, lying transversely internal wall of the central cavity with intusannulation is visible. In furrows there are 2-3 rows of alternating pores (branch inlets). Ridges are thin and sharp. The lower specimen has slightly widening club-like enlargement of the central cavity forming apex in the upper part. The inner side of the wall of the central cavity numerous densely spaced pores are irregularly dispersed in the club-like part of the thallus and forming rows below. Intusannulation is not pronounced. In the club-like part of the thallus there is clearly visible inner wall reach by a set of annuli and interannular furrows separated by lists; x 12; 4. Longitudinal fracture showing two long specimens. The left one has intusannulated walls of the central cavity, seen from inside in the upper part of the specimen. Rows of alternating pores overlap ridges making them indented. In the middle part of the specimen central cavity wall is preserved visible from outside. Intusannulation encompasses the whole thickness of the wall. Two rows of alternating pores occur in furrows partly overlapping ridges. In the lower part of the specimen some tufts are visible. In the lower part of the right specimen intusannulation of the central cavity wall is visible from inside, with two or three rows of alternating pores overlapping the slopes of the ridges resulting in their indentations. On the sides of the thallus there is a poorly preserved set of annuli and interannular furrows. The upper, widening club-like part of the thallus is poorly preserved. Only locally a set of annuli and interannular furrows is visible; x 10.
 |
|
|
Click on thumbnail to enlarge the image.
Plate XXXII: Accumulations of Physoporellae. Przełajka.
1. Accumulation of Physoporella dissita () , (a) and Physoporella praealpina (b); x 8; 2. Accumulation of Physoporella praealpina , (a), Physoporella pauciforata () (b) and Physoporella dissita () (c); x 8; 3. Accumulation of Physoporella pauciforata () (a), Physoporella praealpina (b) and Physoporella dissita () (c); x 8; 4. Accumulation of Physoporella pauciforata () (a) and Physoporella praealpina (b); x 10.
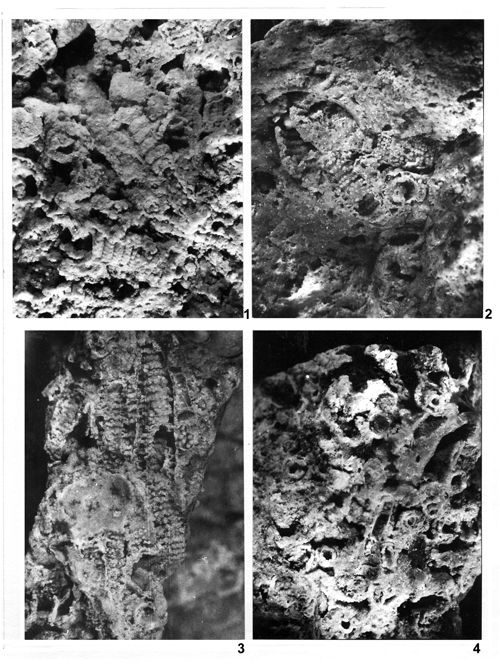 |
|
|
Click on thumbnail to enlarge the image.
Plate XXXIII: Accumulations of Physoporellae. Przełajka.
1. Accumulation of Physoporella praealpina (a), Physoporella pauciforata () (b) and Physoporella dissita ) (c); x 8; 2. Accumulation of Physoporella praealpina (a) and Physoporella pauciforata () (b); x 8; 3. Accumulation of Physoporella pauciforata () and Physoporella praealpina ; x 8; 4. Accumulation of Physoporella pauciforata () (a), Physoporella praealpina (b) and Oligoporella elegans () (c); x 8.
 |
|
|
Click on thumbnail to enlarge the image.
Plate XXXIV: Accumulations of Physoporellae. 1, 3-4 Przełajka, 2 borehole Winowno 25 WW.
1. Accumulation of Physoporella pauciforata () (a) and Physoporella praealpina (b); x 8; 2. Accumulation of Physoporella praealpina and Physoporella pauciforata () ; x 6; 3. Accumulation of Physoporella praealpina (a), Physoporella pauciforata () (b) and Physoporella dissita () (c); x 4; 4. Accumulation of Physoporella praealpina (a), Physoporella pauciforata () (b) and Physoporella dissita () (c); x 4.
 |
|
|
Click on thumbnail to enlarge the image.
Plate XXXV: Accumulations of Physoporellae. Przełajka.
1. Accumulation of Physoporella praealpina (a), Physoporella pauciforata () (b) and Physoporella dissita () (c); x 6; 2. Accumulation of Physoporella praealpina (a), Physoporella pauciforata () (b) and Physoporella dissita () (c); x 6.
 |
|
|
Click on thumbnail to enlarge the image.
Plate XXXVI: Accumulations of Dasycladales from various localities.
1. Accumulation of Oligoporella chrzanowensis n.sp. (a) with subordinate Physoporella praealpina (b); Rosowa Góra; x 8; 2. Accumulation of Salpingoporella krupkaensis n.sp. (a), Physoporella pauciforata () (b) and Oligoporella elegans () (c); Krupka; x 6; 3. Accumulation of Physoporella pauciforata () (a) and Physoporella praealpina (b); borehole 10Ż Lgota Nadwarcie; x 6; 4. Accumulation of Physoporella praealpina (a), Physoporella pauciforata () (b) and Oligoporella elegans () (c); borehole 10Ż Lgota Nadwarcie; x 8.
 |
|
|
Click on thumbnail to enlarge the image.
Plate XXXVII: Accumulations of various Dasycladales from boreholes of the Zawiercie region.
1. Accumulation of Kantia comelicana () (a), Oligoporella chrzanowensis n.sp. (b), Physoporella praealpina (c) and Oligoporella elegans () (d); borehole 10 Ż Lgota Nadwarcie; x 8; 2. Accumulation of Kantia comelicana () (a), Salpingoporella krupkaensis n.sp. (b), Physoporella dissita () (c), Physoporella praealpina (d) and Oligoporella elegans () (e); borehole 10 Ż Lgota Nadwarcie; x 8; 3. Accumulation of Physoporella dissita () (a), Physoporella praealpina () (b) and Salpingoporella krupkaensis n.sp. (c); borehole 10 Ż Lgota Nadwarcie; x 6; 4. Accumulation of Physoporella praealpina (a), Physoporella pauciforata () (b) and Oligoporella elegans () (c); borehole 10 Ż Lgota Nadwarcie; x 10.
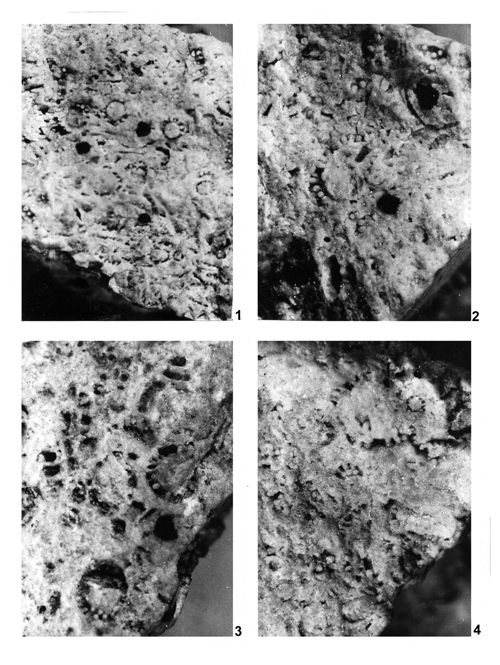 |
|
|
Click on thumbnail to enlarge the image.
Plate XXXVIII: Accumulations of various Dasycladales from boreholes of the Zawiercie region.
1. Accumulation of Physoporella praealpina (a), Physoporella pauciforata () (b) and Oligoporella elegans () (c); borehole 10 Ż Lgota Nadwarcie; x 8; 2. Accumulation of Physoporella praealpina (a) and Physoporella dissita () (b); borehole 10 Ż Lgota Nadwarcie; x 10; 3. Accumulation of Physoporella praealpina (a) and Diplopora annulatissima (b); borehole 10 Ż Lgota Nadwarcie; x 10; 4. Accumulation of Physoporella praealpina (a) and Oligoporella elegans () (b); borehole 10 Ż Lgota Nadwarcie; x 12.
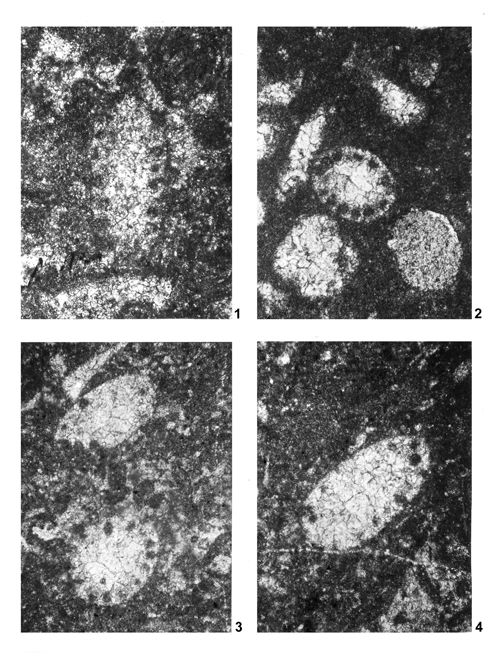 |
|
|
Click on thumbnail to enlarge the image.
Plate XXXIX: Acicularia sp. from boreholes of the Zawiercie Region.
1. Winowno-Będusz 94-WB, depth 95.65-102.60 m; Karchowice Beds; coll. J. ; x 4.5; 2. Winowno-Będusz 94-WB, depth 103.8-115.50 m; Karchowice Beds; coll. J. ; x 4.5; 3. Winowno-Będusz 71-WB, depth 187.0 m; Karchowice Beds or the top of Gorażdże Beds; coll. J. ; x 4.5; 4. Winowno-Będusz 71-WB, depth 187.0 m; Karchowice Beds or the top of Gorażdże Beds; coll. J. ; x 4.5.

