Last modified Mar. 30, 2000
FAMILY TRIPLOPORELLACEAE
TRIBE SELLIPORELLEAE
GENUS Neoteutloporella BASSOULLET et alii 1978
SPECIES Neoteutloporella rajkae PARENTE et CLIMACO 1999
(by M. Parente)
1. Synonymy list
1999 Neoteutloporella rajkae n. sp.- Parente & Climaco, Pl. 34, fig. 1-8; Pl. 35, fig. 1-8
2. Types
Holotype: Pl. 34, fig.
1 &
3, thin section N° AC572.7;
Isotypes : Pl. 34, fig.
2 &
4, thin section N° AC554.23; Pl. 34, fig.
5, thin section N° AC354.21; Pl. 34, fig.
6, thin section N° AC561.2; Pl. 34, fig.
7, thin section N° AC561.6; Pl. 34, fig.
8, thin section N° AC560; Pl. 35, fig.
1, thin section N° AC559.1; Pl. 35, fig.
2, thin section N° AC554.22; Pl. 35, fig.
3, thin section N° AC561.1; Pl. 35, fig.
4, thin section N° AC554.23; Pl. 35, fig.
5, thin section N° AC562.2; Pl. 35, fig.
6, thin section N° AC565.12; Pl. 35, fig.
7, thin section N° AC554.20; Pl. 35, fig.
8, thin section N° AC565.1, V.
Zamparelli Collection, Dipartimento di
Scienze della Terra, Università di Napoli "Federico II" (Italy)
Type locality: Mte Rotonda, E Maratea, Basilicata (Italy)
Stratum typicum: Upper Triassic, Norian
3. Diagnosis
Original diagnosis (Parente & Climaco, 1999): « Calcareous skeleton simple, articulated (undulated) with well spaced swellings consisting of thin hook-like apophyses. Primary laterals inclined upward, short and acrophore, arranged in well spaced whorls. Each primary lateral bears a tuft of 4-6 elongated secondary laterals each consisting of 3-4 segments separated by constrictions. First segment of secondary laterals acrophore; second segment acrophore to slightly trichophore, third and fourth segment slender and trichophore. Secondary laterals keep at first the same inclination as the primary laterals, then bend upward setting subvertical and finally bend downward paralleling the hook-like apophyses at the outer surface of the skeleton. Reproductive organs unknown, seemingly cladospore and located in the first or second segment of secondary laterals. »
4. Description
Original description (Parente & Climaco, 1999):
« General characters of the
calcareous skeleton. The calcareous skeleton of Neoteutloporella rajkae n. sp.
is articulated. Its outer surface is characterised by well spaced
swellings whose maximum height, taken in the proximal portion, is
about 250-350 μm. The long (800-1440 μm) portions of
skeleton intervening between two subsequent swellings are bound
outward by slightly concave surfaces.
The type of articulation present in Neoteutloporella rajkae n. sp. can be described as undulation,
following the nomenclature of PIA
(1912) and DE CASTRO
(1997), because even in the most constricted parts the skeleton is
rather thick. As in other species showing this type of articulation
the skeleton was massive, undulation did not affect its rigidity and
specimens are found as whole skeletons and not as isolated articles.
The swellings are made of thinning
outward apophyses of the calcareous skeleton. These thin and fragile
apophyses can be partly to almost completely obliterated by subsequent
abrasion and breakage. Where completely preserved they show an
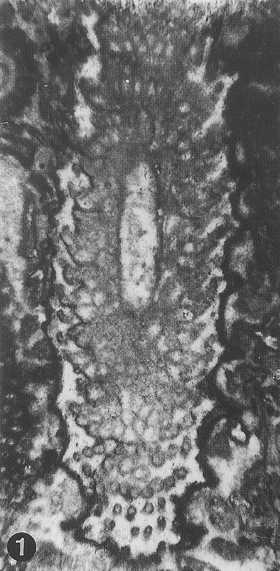
hook-like shape in longitudinal and oblique sections (Pl. 34/2, 35/7) with a short and thicker proximal part directed upward, a longer
and thinner median part perpendicular to the central stem axis and a
very short and thin distal part bending downward. Most frequently in
our material subsequent abrasion truncated the hook-like distal part
so that only the proximal and median ones are preserved (Pl. 35/3, 35/4).
In a few specimens abrasion went as deep as to leave only short stumps
at the outer surface of the skeleton (Pl. 34/1, 35/1). The
variable morphology of the apophyses is certainly a preservation
artefact since different parts of the same specimen often show the
whole range of shapes (Pl. 35/1,
35/4, 35/7).
The maximum outer diameter of the calcareous
skeleton (Dmax), taken at the distal end of the apophyses
is 2080-5120 μm (3495 ± 791 μm). The corresponding
maximum thickness of the skeleton (emax) is 700-2060 μm
(1315 ± 344 μm). The minimum outer
diameter, taken at the constrictions of the skeleton (Dmin),
is 1520-4720 μm (2623 ± 818 μm). The corresponding
minimum thickness of the skeleton (emin) is 460-1860 μm
(889 ± 365 μm). These values have
been measured from longitudinal and oblique sections of specimens
(Tab. 1). The biometric values of specimens in transversal sections
vary because the section may run through a swelling or through a
constriction (Tab. 2).
The longest specimens in our material
are up to 2 cm long. They show that the outer diameter of the skeleton
increases slightly and regularly upward (Pl. 35/4). Unfortunately the
inner surface of the skeleton is not preserved in these very long
specimens so that we cannot control if the diameter of the central
stem is increasing too.
In well-preserved specimens of Neoteutloporella rajkae n. sp.
the calcareous skeleton envelopes the laterals from their insertion
point on the central stem up to their distal ends. Therefore in these
specimens the inner diameter of the skeleton (d) corresponds to the
diameter of the central stem. Its value is 560-1120 μm (835 ± 158 μm) (Tab. 3).
We observed many specimens and
fragments of Neoteutloporella rajkae n. sp. where only the outer part of the
skeleton is preserved. Their specific identification is based on the
very peculiar morphology of the hook-like apophyses of the skeleton.
In some specimens the inner part of the skeleton is totally missing
(Pl. 35/4). In others the central stem wall is preserved and lined by
a thick micritic rim but large and irregular cavities occur in the
proximal and median part of the skeleton (Pl. 35/2).
The
laterals. In fossil Dasycladales,
shape, size and arrangement of laterals are deduced from the pores of
the calcareous skeleton as observed in random sections (De Castro, 1997). In specimens
of Neoteutloporella rajkae
n. sp. the preservation of the pores of the skeleton is far from
perfect. In many specimens the pores have been almost completely
obliterated by recrystallization (Pl. 34/2, 34/3, 34/7). Very often
the inner part of the calcareous skeleton is completely missing or
largely destroyed by dissolution. Our conclusions on shape and
arrangement of laterals are therefore largely based on a few
"lucky" specimens where the pores are preserved as cement
filled cavities lined by a thick micritic rim.
Neoteutloporella
rajkae n. sp. is
characterised by short acrophore primary laterals bearing a tuft of
long and segmented trichophore secondary laterals.
The primary laterals are arranged in well spaced
whorls (Pl. 34/1, 35/3 ). The distance between whorls (h) is
800-1440 μm (1123 ± 165 μm). Each whorl is made
of 15-26 (21 ± 4) laterals (Tab. 3).
The primary laterals are short and acrophore (Pl. 34/1 , 34/3, 35/3)
and are always inclined upward (a1=35°-65°).
Their length (l1) is 250-350 μm (296 ± 40 μm). The vertical
diameter (p1r) is 125-150 μm (148 ± 8 μm); the verticillar
diameter (p1v) is 150-200 μm (180 ± 21 μm).
Each primary lateral bears a tuft of
secondary laterals. Their number per whorl is very difficult to count
the number of laterals per tuft. Some longitudinal and oblique
sections show that each primary bears 3-4 secondary laterals in a
longitudinal plane (Pl. 34/1,
34/3,
35/5). Transversal sections show
1-2 secondary per primary lateral (Pl. 34/7). In tangential sections
(or tangential parts of oblique sections), cutting through the
proximal part of a tuft, the pores are so crowded that is impossibile
to correlate each pore to its tuft (Pl. 35/1). Our best guess is that
each primary lateral bears a tuft of 4-6 secondary laterals.
The secondary laterals of Neoteutloporella rajkae n. sp.
are segmented (Pl. 4/3,
34/4, 34/6, 35/2, 35/6, 35/8). Each lateral consists
of at least 3-4 segments separated by constrictions. The first segment
is acrophore (Pl. 34/1, 34/2, 34/4).
Its length (l2') is 450-600 μm (525 ± 65 μm), its vertical
diameter (p2'r) is 150-175 μm (153 ± 9 μm). The second segment
is more elongated, acrophore to slightly trichophore (Pl. 34/1, 34/2, 34/4,34/6, 35/6). Its length (l2") is 400-900 μm (650 ± 250 μm); the vertical
diameter (p2"r) is 100-150 μm (113 ± 25 μm). The remaining part
of the secondary lateral is long, slender and distinctly trichophore (Pl. 34/1, 34/2, 34/6, 35/3, 35/4, 35/6). Because of the poor preservation of
most specimens it is difficult to state if the trichophore part of
secondary laterals comprises just one very long or two shorter
segments. The length of the secondary laterals from the apex of their
second segment to the outer surface of the skeleton (not including the
hook-like end) is 850-1500 μm (1158 ± 227 μm). The diameter of the
distal end of secondary laterals (p2min) is 50-100 μm (75 ± 19 μm).
The very peculiar shape of the tufts of
secondary laterals, as seen in longitudinal sections, is due to
changes of inclination of the laterals along their course (Pl. 34/1).
As stated above 3-4 laterals are usually seen in a longitudinal plane
cutting through the proximal part of a tuft. The first and second
segment of the central lateral(s) keeps the same inclination as the
primary lateral , the upper and lower laterals at first diverge (first
segment) and then converge (second segment) so to set parallel to the
"central" lateral(s) (Pl. 34/3). Then all the secondary
laterals bend upward setting at an angle of 10°-20° while they
converge (Pl. 34/1). Finally the distal part of the secondary laterals
bends downward following the hook-like course of the skeleton
apophyses. The radial diameter of the tuft of secondary laterals
therefore reaches its maximum at the first segment and thins out
markedly at the third and fourth (?) segment (Pl. 34/1). The radial
diameter of the tuft at its distal end is not larg enough to
accomodate 3-4 laterals in a vertical plane. Some of them are probably
displaced so that only one or two thin secondary laterals per tuft are
seen in longitudinal sections at the outer surface of the skeleton.
Reproductive organs. We
can only suppose that Neoteutloporella
rajkae n. sp. was provided with uncalcified cladospore
reproductive organs seemingly located in the primary laterals or in
the first or second segment of secondary laterals. »
|
d |
Dmin |
Dmax |
d/Dmin |
d/Dmax |
emin |
emax |
p1r |
l1 |
n |
15 |
14 |
16 |
13 |
15 |
13 |
15 |
10 |
8 |
min |
a |
a |
a |
a |
a |
a |
a |
a |
260 |
max |
a |
a |
a |
a |
a |
a |
a |
a |
360 |
mean |
a |
a |
a |
a |
a |
a |
a |
a |
298 |
st. dev. |
139.1 |
818.4 |
791 |
0.063 |
0.033 |
364.7 |
344 |
8 |
40 |
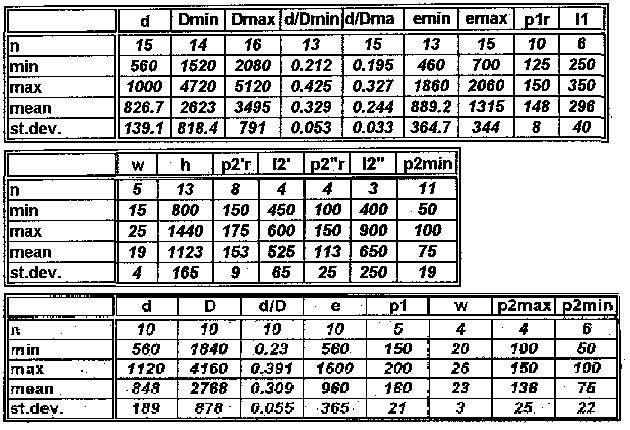
Measurements (see above: blocks 1 & 2, longitudinal and oblique sections; block 3, transversal sections)
5. Remarks
6. Stratigraphic range
Norian (Parente & Climaco, 1999)
7. Real distribution
Italy (Parente & Climaco, 1999)
8. Paleoecology
|
9. Figures |
|||
|
|
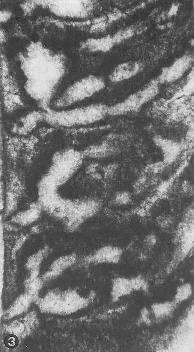
from Parente & Climaco, 1999: Pl. 34, fig. 1 (holotype) |
 |
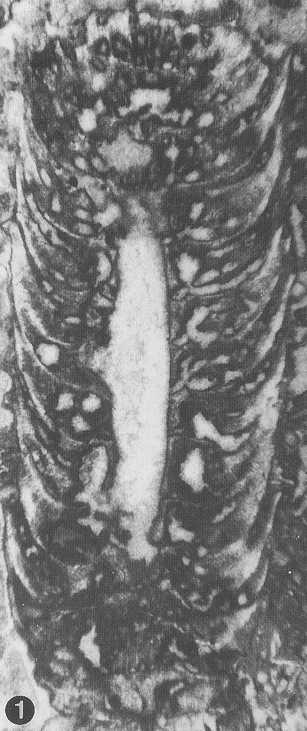
from Parente & Climaco, 1999: Pl. 34, fig. 3 (detail of the holotype) |
{kind=link}
{kind=link}
{kind=link}
10. Reference
PARENTE M., CLIMACO A. (1999).-
Dasycladalean
green algae from the Upper Triassic of Mt. Rotonda (Verbicaro Unit, Calabria-Lucania border, Southern Italy).
Facies, Erlangen, 41, p. 159-182, 7 pl. (34-40).