Last modified Aug. 11, 2000
FAMILY DIPLOPORACEAE
TRIBE ?
GENUS Spinaporella FLÜGEL et FLÜGEL-KAHLER in E. FLÜGEL et alii 1984
SPECIES Spinaporella granadaensis FLÜGEL et FLÜGEL-KAHLER in E. FLÜGEL et alii 1984
(by J.C. Braga)
1. Synonymy list
1984 Spinaporella granadaensis n. sp.- E. Flügel & Flügel-Kahler in E. Flügel, Flügel-Kahler et alii, Pl. 21, fig. 7; Pl. 23, fig. 1-4 ; Pl. 30, fig. 2
1987 (?) Spinaporella ? granadaensis.- Braga & Martin, Pl. I, fig. 8
1999 Spinaporella ? granadaensis.- Parente & Climaco, Pl. 40, fig. 3 & 5
2. Types
Holotype: Pl. 23, fig. 3, sample TCH-10a, thin section N° SM.B. 13500, Paläobotanische Abteilung, Forschungsinstitut Senckenberg, Frankfurt am Main (Germany)
Type locality: Pico de la Carne, SSE Monachil - SE La Zubia, Granada (Spain)
Stratum typicum: Norian. The dolostones of Pico de la Carne section were attributed by Flügel et alii (1984) to the Middle Triassic. Braga & Martin (1987) demontrated the Late Triassic (Norian) age of these materials.
3. Diagnosis
Original diagnosis (Flügel & Flügel-Kahler in Flügel et alii, 1984): « Cylindrical, metaspondyle thalli. Only primary branches, arranged in clusters of two or more thick branches. Branches are oriented obliquely to the axis of the stem cell, at an angle of about 30°. Some branches are strongly tilted at the periphery. Distinctly large branches. »
4. Description
Original
description (Flügel & Flügel-Kahler in Flügel et alii,
1984): « The conspicuously long thalli are characterized by
numerous, unbranched trichophore branches, arranged in distinct,
closely-spaced whorls (Pl. 23/2).
Starting at the stem cell the branches are tilted obliquely upwards from
the main axis. Long and shorter branches occur within the same thallus,
also exhibiting different curvatures (Pl. 23/3). Larger branches exhibit a sigmoidal
flexure near the periphery (Pl. 23/1). Some branches are wider near the stem
cell (Pl. 23/1).
Differences in the calcification are caused by a concentration of larger
interspaces between more deeply curved branches within the peripheral
zone (Pl. 23/1, 23/3).
The calcification of the thallus is generally poor, the preservation is
partially due to the strong micritization. »
Most recent description (Parente & Climaco, 1999): « We have found only two specimens of this species along with few very poorly preserved fragments. The calcareous skeleton is cylindrical and very thick (e = 1360-1600 μm) with a relatively narrow central cavity (d = 800-880 μm; D = 3600-4000 μm; d/D = 0.2-0.24). The pores are very crowded so that it is very difficult to be precise about their arrangement. We can only suppose that this species was characterised by short acrophore primary laterals (p1 = 150 μm; l1 = 200-250 μm) bearing tufts of long and segmented trichophore secondary laterals distincly inclined upwards (p2max = 150-200 μm; p2min = 75 μm). However we cannot exclude the possibility that a further branching, giving rise to third order laterals, is present where the laterals become markedly thinner. »
New description: Cylindrical thalli up to 27 mm in length and 1.7 to 5.5 mm in diameter (mean 3.25) Primary branches only arranged in whorls and forming clusters of two-three. This metaspondyle arrangement is masked by the close packing of the branches. Long (1275 to 1910 μm, mean 1610), trichophorous branches with a diameter ranging from 90 to 250 μm (mean 150) at their proximal end. They spread upwards at an angle of 25 to 60 degrees (mainly about 30) to the stem. They curve upwards in their outer half and then downwards in their distal end. Calcification is weak around the branches but it is commonly enhanced by a micritic cement in the studied localities. The resulting wall is very thick (600 to 2200 μm, mean 1200) and its thickness is clearly proportional to the thallus diameter. The outer surface contours the branches giving to the plant a distinct spiny aspect.
Measurements:
Original measurements (Flügel & Flügel-Kahler in Flügel et alii, 1984): « The dimensions are summarized in Table 2. The species is characterized by a rather low d/D ratio, thick "walls" and very large pores. »
|
|
L |
D |
d |
d/D% |
s |
w |
p |
|
Holotype |
27000 |
3500 |
1000 |
28 |
1300-1500 |
|
220-260 |
|
Pl. 23/1 |
|
4100 |
1200 |
30 |
1000-1400 |
400 |
170-240 |
|
Pl. 23/2 |
|
7000 |
700 |
10 |
|
240-340 |
170-290 |
5. Remarks
(none)
6. Stratigraphic range
MIDDLE TRIASSIC, ? Ladinian (E. Flügel & Flügel-Kahler in E. Flügel, Flügel-Kahler et alii, 1984) , Norian (Braga & Martin, 1987
The dolostones of Pico de la Carne section were attributed by Flügel et alii (1984) to the Middle Triassic. Braga & Martin (1987) demonstrated the Late Triassic (Norian) age of these materials.
7. Real distribution
Spain (E. Flügel & Flügel-Kahler in E. Flügel, Flügel-Kahler et alii, 1984; ? Braga & Martin, 1987), Italy (Parente & Climaco, 1999)
8. Paleoecology
Up to recently Spinaporella granadaensis was only known from the Upper Triassic dolostones of the Alpujarride Complex in Sierra Nevada (Betic Cordillera, southern Spain) - Cahorros de Monachil, Pico de la Carne, Picos de las Dehesillas and Picacho Alto sections. There it occurs as monocultures, monospecific accumulations, or associated to Physoporella leptotheca Kochansky-Devide, Gyroporella plumosa (Zanin-Buri) Braga, Griphoporella curvata (Gümbel) Pia and Spinaporella granadaensis Flügel et Flügel-Kahler in lagoon (inner platform) deposits.
|
9. Figures |
More photos | |
|
|
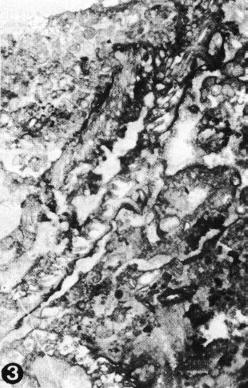 |
from E. Flügel, Flügel-Kahler et alii, 1984: Pl. 23, fig. 3 (holotype) |
10. Reference
FLÜGEL E., FLÜGEL-KAHLER E., MARTIN J.M., MARTIN-ALGARRA A. (1984) -
Middle Triassic reefs
from southern Spain. Facies, Erlangen, 11, p. 173-218, 11 pl. (20-30).