Phoronida: Fósiles Último cambio el 13 de Febrero de 2023 |
||
|
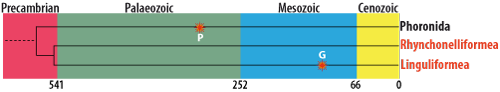
El origen de los Phoronida, ciertamente monofilético con los otros Lophophorata, se remonta hace unos 1000 MA. Aunque como fósiles, los foronídeos no se conocen nada más que a partir del Devónico (los braquiópodos, en cambio desde el Cámbrico y los briozoos desde el Ordovícico), son precisamente los Phoronida quienes han conservado una forma y un modo de vida ancestral, próximo al del antecesor Protolofoforado (Emig, 1984). Varios autores (Fenton & Fenton, 1934; Avnimelech, 1955; Josey, 1959; Voigt, 1972; Mackinnon & Biernat, 1970; Sokoloski, 2005; Wisshak et al., 2017) sugirieron que algunos tubos o madrigueras cilíndricas del registro fósil (por otra parte, descritos bajo distintos icnogeneros, como Skolithos, Talpina, Diorygma...) pertenecen a Phoronida. Recientemente, Voigt (1975) demostró la identidad del icnogénero Talpina von Hagenow, 1840, con el género Phoronis y lo confirmó en 1978. Voigt (1972) también probó que Conchotrema Teichert 1945 es un sinónimo de Talpina, lo que fue ratificado por Plewes (1996). Talpina es abundante y conocido del Cretáceo (Voigt, 1972) y Jurásico (Fürsich et al., 1994). El icnogénero se extiende hacia atrás hasta el Devónico superior (Thomas, 1911; Rodriguez & Gutschick, 1970; véase Bromley, 2004). Tubos fósiles de foronídeos se conocen desde el Devónico (véase Emig, 1982). Talpina es perforante en varios substratos como en algas calcáreas, en equínidos, conchas de moluscos o en el rostro de belemnites. Voigt da criterios para discriminar los tubos de foronídeos de otros similares como talofitas, esponjas, briozoos o "gusanos". Los tubos del icnogénero Talpina ramosa que ocurre con frecuencia dentro de los rostros de Belemnella y Belemnitella (cf. Voigt, 1972; Wisshak et al., 2017) han sido descritos dentro de las cavidades que provienen de la destrucción diagenética de corales aragoníticos de la creta del Maastrichtiano (Voigt, 1978). No ha sido confirmada en la actualidad la presencia frecuente de foraminíferos aglutinados alrededor de la abertura del tubo de un animal fósil parecido a Phoronopsis ni la sugerencia de un comensalismo entre ambos organismos fósiles en Maastrichtiano Superior (Voigt, 1970). Towe (1978) y Larsson (1979) mencionaron la posibilidad que Tentaculites fueron relacionados con los braquiópodos o quizás más de cerca de los foronídeos. Sin embargo, el ultimo autor señaló que el sistema de alimentación de los foronídeos es incompatible con el modo de vida planctónico propuesto para algunos tentaculites. Vinn (2005) y Vinn & Mutvei (2005) formularon la hipótesis que tal vez los foronídeos estaban los parientes vivos más cercanos de Cornulites y de Tentaculites, pero tambien de los braquiópodos o bryozoos (Vinn & Isakar, 2007). Mientras que reconstruir los Cornulites y otros tentaculitidos sobre la base del “esqueleto foronídeo” es especulativo. Los dos únicos grupos, con los cuales Cornulites puede ser comparados directamente, son los briozoos y los cnidarios, con estos últimos morfologías y estructuras de la concha comparables fueron descritas (Herringshaw et al., 2007). Iotuba chengjiangensis fue descrita por Chen & Zhou (1997) en la fauna del Chengjiang (Cámbrico Inferior) a partir de tres ejemplares: se trataría de un foronídeo con un tubo digestivo en U y un penacho de tentáculos. Encontrado en la misma fauna, otro fósil, de cuerpo blando, ha sido afirmado como foronídeo y llamado Eophoronis por Chen (2004). Pero, las afinidades de estos fósiles han sido cuestionadas por Cohen & Weydmann (2005), y de acuerdo con Conway Morris (2006) y Huang (2006), pueden ser priapúlidos. Zhang et al. (2023) consideran estas descripciones como nomina nuda según el art. 13.1 del Código porque no fueron acompañadas de diagnosis [*]; en consecuencia, definen un nuevo género y especie bajo Iotuba chengjiangensis cuyas características son las de los anélidos flabelligeroides, pues resulta que no tienen afinidades con Lophophorata y Phoronida en particular.
Skolithos linearis Haldeman, 1840, la especie tipo del icnogénero, ha sido redescrito por Knaust et al. (2018) del Cámbrico: el posible constructor del tubo sería un poliqueto anélido o un foronídeo. Pero los tubos de las fig. 11 y 12 de estos autores no pueden atribuirse a un foronídeo, porque su parte posterior está redondeada y cerrada. En los foronídeos, esta parte es cónica y siempre tiene una pequeña abertura terminal: este detalle a menudo se ignora y, sin embargo, es primordial en el tubo (Pourreau, 1979). En los trabajos recientes existe la tendencia a atribuir tubos o perforaciones fósiles al grupo de los foronídeos, sin realizar su comparación con la estructura y la composición química de los tubos de ese grupo, que son muy característicos. Además difícilmente puede entenderse que se construya un árbolo filogenético apoyándose en un tubo hipotéticamente atribuido a un foronídeo como hacen Skovsted et al. (2008). Una corta revisión fue propuesta por Emig (2010), y veasei "fósiles de rastro mandados a foronídeos.".
Según estos autores, la edad promedio de la divergencia en los braquiópodos entre Linguliformea y Rhynchonelliformea es 547 MA, mientras que entre braquiópodos y foronídeos es 578 MA. Los tubos de gusanos generalmente identificados como "Spirorbis" se extienden desde el Ordovícico hasta la actualidad. Generalmente, incrustan conchas y otros sustratos. Si los Spirorbis actuales son anélidos poliquetos, Vinn & Taylor (2006) consideran los "Spirorbis" antes del Cretácico, segun los detalles de la estructura del tubo, como Microconchida. Es un orden extinto de lofoforados muy hipotéticos, caracterizados por un tubo spirorbiforme microchonchide. Ese tubo no se puede confundir con los de Phoronida, que tienen una estructura muy específica y una forma rectilínea (véase adelante; y Referencias). Recuerdo : El tubo de fornido es tan característico que no puede ser confundido con otro tubo. Esto implica haberlo visto ya y tocado en realidad y no en foto. Cuando a su fosilización, la cuestión queda puesta, porque parece posible sólo en condiciones muy particulares - como en el caso de las valvas de Lingulidos (Brachiopoda) (véase Emig, 1990).
Más... |
||
|
|