![]()
Contents
[Introduction]
[Main taxonomic characters]
[The Obolus example]
[Recommendations]
[References]
and ...
[Figures]
CNRS UMR 6540, Centre d'Océanologie, Rue de la
Batterie-des-Lions, 13007 Marseille (France)
Manuscript online since November 27, 2002
![]()
This study points out some basic problems of linguloid systematics and proposes solutions for them. A taxonomic examination of the unique species of the genus Obolus found in the Upper Cambrian of Estonia and Russia, O. apollinis (= O. ruchini, O.
transversus, O. rebrovi and Ungula convexa) is used as an example of a methodology employing all of the characters valid for distinguishing species of both extant and fossil Lingulidae. These characters are:
- umbonal region;
- body musculature;
- septa or ridges;
- main mantle canals - as established and figured by (1982,
1983) and and
(1993). All of them have been determined to be taxonomically stable and have been studied and compared to take into account intraspecific variability; they should be used to describe or to redescribe any taxon of the superfamily Linguloidea. Characters of the shell and valves, such as shape, size, and dimensional ratios have no taxonomic value.
Taxonomy; Obolus; Brachiopoda; Cambrian - Ordovician; Estonia.
C. (2002).- Tools for linguloid taxonomy: the genus Obolus (Brachiopoda) as an example.- Carnets de Géologie / Notebooks on Geology, Maintenon, Article 2002/01 (CG2002_A01)
Des outils pour la taxinomie des
Lingulidoida : le genre Obolus (Brachiopoda) pris comme exemple.- Cette étude met en exergue des
problèmes fondamentaux de la systématique des Linguloïdes et propose des solutions méthodologiques basées sur les caractères utilisés pour identifier les espèces de Lingulides actuelles et fossiles. L'unique espèce du genre
Obolus, O. apollinis (= O. ruchini, O. transversus, O. rebrovi
et Ungula convexa), récoltée dans divers gisements du Cambrien
moyen et supérieur en Estonie et Russie, sert ici d'exemple. Les caractères sont :
- la région umbonale ;
- la musculature du corps ;
- les septums et crêtes internes ;
- les canaux du manteau ; selon la description et la représentation faites par (1982,
1983) et
et
(1993). Tous ces caractères se sont révélés taxinomiquement stables tout en présentant une relative variabilité intraspécifique ; ils sont donc à être utilisés pour décrire ou redécrire tous les taxons de la superfamille des Linguloidea. Il convient de souligner que les caractères de la coquille ou des valves, tels que formes, tailles, rapports dimensionnels n'ont aucune valeur taxinomique.
Taxinomie ; Obolus ; Brachiopoda ; Cambrien - Ordovicien ; Estonie.
As stated by (1977) diagnoses of the extant species of the genera Lingula , 1797, and Glottidia , 1870, are too poor to permit discrimination between species. Consequently, all species referred to these genera have been redescribed by (1982, 1983) using a new taxonomy based on morpho-anatomical characters. Recently, these characters have been used successfully by and (1993), et alii (1996) and - et alii (1999) to revise the taxonomy of Palaeozoic and Mesozoic brachiopod genera and species.
During recent field trips to Estonia hundreds of well preserved specimens were collected from the Upper Cambrian (,
1996) near Tallinn. Previously, brachiopods from these strata were classified as belonging to the genera Ungula , 1830, Oepikites
et , 1984, and Schmidtites
et , 1929, all representatives of the family Obolidae (Lingulida: Linguloidea). Other specimens studied are in the collections of the Museum of Geology in the Institute of Geology of the University of Tartu, Estonia; they were collected by L. from:
(1) localities along the Sarya and Volkhov rivers (Ingra, Russia: Middle Cambrian) and identified as Obolus apollinis , 1829,
(2) the locality Sarya River (Ingria Russia, Middle Cambrian) identified as O. ruchini
et , 1984,
(3) the Sayas River locality identified as O.
transversus (, 1830),
(4) from Ladoga and Suma River localities (Ingria Russia: Upper Cambrian) identified as Ungula convexa , 1830.
It is of fundamental importance that the taxonomic criteria used here to define the Cambrian genus Obolus are applied in the revision of the systematics of and within the superfamily Linguloidea.
The main taxonomic criteria used to discriminate between linguloid taxa were established and figured by
(1982, 1983) and
and
(1993). They are listed below, illustrated for the genus Obolus on
Fig. 1
![]() and described in Tables 1
and described in Tables 1 ![]() and 2
and 2 ![]() .
.
Most of these features are used to define species, but some appear to have generic value also, for example the septa in the extant genus Glottidia. Some serve as criteria to define higher taxa among the Linguloidea, for example the ridge in the dorsal valve between the anterior oblique muscles, a characteristic perhaps of the entire superfamily. Symmetrical musculature is a defining characteristic of the family Obolidae and asymmetrical musculature serves to distinguish the Lingulidae. All of these criteria exhibit great variability, so when the erection of a new linguloid genus or species is contemplated its characteristics must be measured and compared not only with a single related taxon but also with all possible relatives. The description of fossil taxa should be based on no less than 20 to 30 well-preserved specimens, none fragmentary. To attain this requisite number, several hundred specimens may have to be collected, prepared and studied in detail. Furthermore a species should be defined only on the basis of its occurrence in at least three discrete geographic populations, as suggested by (1981).
In addition, it is highly desirable that the full range and limits of variability of diagnostic characters be figured, and that the figures be integrated into the definition of the species as has been done here in
Fig. 1
![]() for Obolus apollinis.
for Obolus apollinis.
External features, such as the shape, size, and dimensional ratios of the valves, have been demonstrated to have no taxonomic value (, 1982, 1983; and , 1993). Consequently, they cannot be used to define either genus or species but may be given additional to the diagnosis. Only exceptionally is shell form diagnostic of a species, for example the quadrangular shape of the shell of Lingula adamsi when compared to those of the other species of Lingula (, 1982).
Among the lingulid taxa of the Middle Cambrian to Ordovician (Tremadocian) in Estonia and Russia, the genus Obolus (Table
1 ![]() )
has been the subject of controversy regarding its systematics (see et alii,
1989; , 1996 for reviews), because many species have been assigned to this genus without regard to its taxonomic characteristics stated above;
)
has been the subject of controversy regarding its systematics (see et alii,
1989; , 1996 for reviews), because many species have been assigned to this genus without regard to its taxonomic characteristics stated above;
The genus Obolus has been redefined based on a study of four species: i.e. O. apollinis, O. ruchini, O. transversus (= O. rebrovi
et , 1984), and Ungula convexa (Table 2 ![]() ). Consequently, three of these species must be considered junior synonyms of O. apollinis, the type-species, originally described by
(1829).
). Consequently, three of these species must be considered junior synonyms of O. apollinis, the type-species, originally described by
(1829).
All of the specimens of the above cited species show so many similarities in their characters that they are described and discussed herein under the name Obolus apollinis. Furthermore their original descriptions (, 1829; , 1830; and , 1984; et alii, 1989) do not establish valid distinctions between them.
The shell ranges from subcircular to a subtriangular in outline, the former being most common
(Fig. 1
![]() ). Externally, the valves are smooth with fine growth lines, some of which are stronger. Sometimes weak radial marks are present. The insides of both valves shows a thickened visceral area, ranging from a very slight elevation to a very well developed thickened area, common in large specimens. In the ventral valve there is a heart-like depression between the anterior and posterior adductor muscles. et alii
(1989) stated that Obolus differs from Ungula in having a subcircular, thinner and flatter shell and lacking a heart-like depression in the ventral visceral area: However these characteristics have no taxonomic value because they occur in both Obolus and Ungula.
). Externally, the valves are smooth with fine growth lines, some of which are stronger. Sometimes weak radial marks are present. The insides of both valves shows a thickened visceral area, ranging from a very slight elevation to a very well developed thickened area, common in large specimens. In the ventral valve there is a heart-like depression between the anterior and posterior adductor muscles. et alii
(1989) stated that Obolus differs from Ungula in having a subcircular, thinner and flatter shell and lacking a heart-like depression in the ventral visceral area: However these characteristics have no taxonomic value because they occur in both Obolus and Ungula.
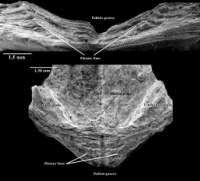
The pseudointerarea always overhangs the internal valve surface, except at the level of the pedicle groove, which is generally continuous with the internal side of the valve. Laterally it is limited by well-marked and elevated flexure lines, which are aligned with the Anterior (or Median) Internal Oblique - Anterior Oblique muscles. The flexure line is sometimes accentuated by a narrow groove. Furthermore, in the ventral valve the pseudointerarea, triangular and slightly concave, extends laterally over about 30 to 60 per cent of the umbonal region of which the beak is rather rounded. A rather narrow subparallel pedicle groove medially divides the ventral pseudointerarea (Fig.
1A
![]() ,
2
,
2 ![]() ). The length of the pedicle groove ranges between 1.3 and 3.8 mm (mean = 2.2 mm; n = 31). In larger specimens the groove sometimes is less prominent or unimpressed anteriorly over half to one-third of the length of the pseudointerarea
). The length of the pedicle groove ranges between 1.3 and 3.8 mm (mean = 2.2 mm; n = 31). In larger specimens the groove sometimes is less prominent or unimpressed anteriorly over half to one-third of the length of the pseudointerarea
On the dorsal valve, the subtriangular pseudointerarea is slightly concave and rounded; it has an extent similar to that of the ventral pseudointerarea, i.e. it occupies from 30 to 60 per cent of the umbonal region. The median length of the pseudointerarea ranges from 0.6 and 3.4 mm (mean = 1.6 mm; n = 34). The flexure lines are well-marked and elevated: they are more or less aligned with the three postero-lateral muscles, i.e. the Median Lateral Oblique muscle, the Anterior (or Median) Internal Oblique muscle and the Anterior Lateral Oblique muscle. Anteriorly, the flexure lines on either side extend towards the Anterior Lateral Oblique muscle (Fig.
1B
![]() , 3
, 3
![]() ).
).
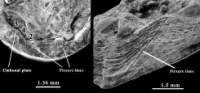
The clearly established existence of flexure lines on the dorsal valve
(Fig. 3
![]() )
is cited here for the first time as regards the genus Obolus, because their lack was used by et alii
(1989) to distinguish the genus Obolus from the genus Ungula. Consequently, this character cannot be used to separate the genera.
)
is cited here for the first time as regards the genus Obolus, because their lack was used by et alii
(1989) to distinguish the genus Obolus from the genus Ungula. Consequently, this character cannot be used to separate the genera.
On each side of the pseudointerarea is a flat, broad lateral umbonal plate, which largely overhangs the Oblique muscle scars. These plates are commonly as wide as the length of the pedicle groove. Nevertheless, such plates appear rather fragile and are partly broken in many specimens. Pseudointerarea and lateral umbonal plates overhang the scars of the Anterior and Internal Oblique muscles sometimes so far that they are hidden completely. These features may vary in their expression in relation to the degree of internal thickening of the valve.
The muscle system of Obolus was first described by (1939). The absence in the obolids of the Internal Posterior Oblique muscle, considered by this author as an undivided Median Internal Oblique muscle, is an important feature because the muscle arrangement is bi-symmetrical in the obolids and asymmetrical in the lingulides. This phylogenetic character was overlooked in the diagnosis of the family Obolidae by and (2000). However, it is a character which should perhaps be applied at superfamily level, thus implying a revision of the higher taxa in the order Lingulida.
New characteristics of musculature in Obolus are established: muscle scars "deeply" impressed into the inner surface of the valves, in particular the postero-lateral scars; in the ventral valve, the muscle succession Anterior Oblique - Internal Oblique is more or less aligned with the flexure lines of the pseudointerarea, covered by the lateral umbonal plates until about the middle of the Anterior Oblique muscle, and less extended antero-laterally than in Ungula; in the composite muscle formed by the Anterior Adductor and the Anterior Lateral Oblique, the latter is located in the posterior part of the scar; in the inner side of the dorsal valve the lateral umbonal plates overhang the alignment of muscles: Median Lateral Oblique - Anterior Lateral Oblique - Anterior Internal Oblique (or Median Internal Oblique) until about the middle of the Anterior Lateral Oblique muscle; the Anterior Adductor muscles are elongate and slightly convergent posteriorly as observed also in the obolid Schmidtites celatus (unpublished
data) (Fig. 1
![]() ,
2
,
2
![]() , 3
, 3
![]() ).
).
On the ventral valve a narrow to well-developed median ridge extends over several millimetres at the level of and between the posterior adductor muscles (Fig.
1
![]() ,
2
,
2
![]() ). This ridge was first cited as the "Seitensepta" by
(1896). On the dorsal valve, a median ridge at the level of the anterior oblique muscle scars is narrow and poorly developed. One may infer that, as in extant lingulides, the importance of this ridge generally increases with the size of the specimen.
). This ridge was first cited as the "Seitensepta" by
(1896). On the dorsal valve, a median ridge at the level of the anterior oblique muscle scars is narrow and poorly developed. One may infer that, as in extant lingulides, the importance of this ridge generally increases with the size of the specimen.
The vascula lateralia are described as arcuate, submarginal by et alii (1989). No observation is given on the vascula media.
As a result of this study, a new diagnosis for Obolus , 1829, is provided (Table
1 ![]() )
as well as for the unique species Obolus apollinis , 1829, remaining in the genus (Table
2
)
as well as for the unique species Obolus apollinis , 1829, remaining in the genus (Table
2 ![]() ). Furthermore, Ungula convexa described by
(1830) was considered by subsequent designation (,
1965: see and ,
2000) as the type species of Ungula but this species is now assigned to Obolus and considered a synonym of Obolus apollinis. Thus, a new type species for Ungula must be proposed (paper in preparation): that is Ungula ingrica (, 1829), originally described under Obolus ingricus, and new diagnoses will be provided for the species and for Ungula.
). Furthermore, Ungula convexa described by
(1830) was considered by subsequent designation (,
1965: see and ,
2000) as the type species of Ungula but this species is now assigned to Obolus and considered a synonym of Obolus apollinis. Thus, a new type species for Ungula must be proposed (paper in preparation): that is Ungula ingrica (, 1829), originally described under Obolus ingricus, and new diagnoses will be provided for the species and for Ungula.
The genus Obolus now represented by only one species is an example of what occurs when the taxonomic characters discussed above are applied. Furthermore, this genus along with the genus Lingula, both of which have provided a name for the family, share an occurrence unique among the inarticulated brachiopods: that is many more or less complete fossils specimens have been assigned to one or the other genus based solely on the shape of the shell.
Because Obolus was selected as the genotype at a family level (see and , 2000) the taxonomic criteria used here should be applied to the genera and their species in all the Obolidae. The same remark is valid for all the Lingulidae (see Lingulidae in http://paleopolis.rediris.es/BrachNet/).
Taxonomy is a tool based primarily on a diagnosis of each taxon. When comparing the diagnoses in Tables
1 ![]() and 2
and 2 ![]() , remember that the definition of the term diagnosis as given in ICZN
(1999), differs slightly in the French and English versions: "Énoncé écrit établissant l'ensemble des caractères d'un taxon qui suffisent à le distinguer des autres taxons auxquels il peut être utilement comparé" and "A statement in words that purposes to give those characters which differentiate the taxon from other taxa with which it is likely to be confused". A diagnosis should also comply with Recommendation 13A (ICZN,
1999) as well as recommendation 13B that concerns the languages in which a diagnosis should be given.
, remember that the definition of the term diagnosis as given in ICZN
(1999), differs slightly in the French and English versions: "Énoncé écrit établissant l'ensemble des caractères d'un taxon qui suffisent à le distinguer des autres taxons auxquels il peut être utilement comparé" and "A statement in words that purposes to give those characters which differentiate the taxon from other taxa with which it is likely to be confused". A diagnosis should also comply with Recommendation 13A (ICZN,
1999) as well as recommendation 13B that concerns the languages in which a diagnosis should be given.
A diagnosis cannot be based on features without taxonomic value, or even more significantly on chacters without phylogenetic value, or include only a part of the characters that define a taxon. A species should not be identified by any of the characters that define the genus and vice versa, so that the diagnosis for each hierarchical taxon is clearly differentiated (Recommendation 13A).
Information on the sites where types are deposited and their accessibility for study or revision is sometimes rather difficult to obtain for linguloid taxa, contrary to Recommendation 72F of the ICZN (1999). None of the types of Obolus species are available from the Geological Museum in St-Petersburg (Russia) and the only specimens of Obolus that can be studied currently are in Tartu (Estonia).
Finally, a scientific approach to systematics should be based on a good knowledge of the taxa, comparative morphology and anatomy. Ecological requirements may also be criteria in systematics ( and , 1987). Systematics and taxonomy must propose new ideas and test hypotheses. Systematics as a tool to identify a taxon is but a technical consequence. This paper is a first step in better understanding in linguloid taxonomy, as well as in suggesting new hypotheses and cladistic analyses. In these latter, a plesiomorphous or apomorphous condition has to be proved and discussed; that such a state exists cannot be based only on a simple computer calculation.
This study was supported by a CNRS-Estonia grant for cooperation between France and Estonia and through a grant by the French Embassy in Tallinn. I thank Ivar and the Institute of Geology staff (University of Tartu, Estonia) for laboratory facilities and for help in collecting specimens, and Chantal (Centre d'Océanologie, Marseille) for SEM preparations. I am most grateful to Nestor J. (USA) for comments and English improvement of the earlier draft, as well as to the two referees, Art (OSU, Corvallis, USA) and Fernando (Universidad de Oviedo, Spain).
P.M. and C.C. (1987).- La population, unité fonctionnelle de la biocoenose. In: Biologie des Populations.- Actes du Colloque National du Centre National de la Recherche Scientifique, Lyon 1986, pp. 69-72.
G. and C.C. (1993).- Anatomical distinctions of the Mesozoic lingulide brachiopods.- Acta Palaeontologica Polonica, Warsaw, Vol. 38, N° 1/2, pp. 1-20.
J.G. (1797).- Vers, coquilles, mollusques et polypiers. Tableau encyclopédique et méthodique des trois règnes de la nature, vol. 2. Agasse, Paris, Pl. 96-314.
O.M. (1939).- Muscle systems of some inarticulate brachiopods.- Geological Magazine, Cambridge, Vol. 76, pp. 434-444.
W.H. (1870).- A revision of the terebratulidae and lingulidae.- American Journal of Conchology, Vol. 6, pp. 88-168.
E. (1829).- Zoologia specialis, quam expositis animalibus tum vivis, tum fossilibus potissimum Rossiae in universum et Poloniae in specie, in usum lectionum publicarum in Universitate Caesarea Vilnensi. Volume 1.- Josephi Zawadski, Vilnae, 314 p.
C.C. (1977).- Réflexions sur la taxonomie des espèces du genre Lingula (Brachiopodes, Inarticulés).- Comptes Rendus de l'Académie des Sciences, Paris, Vol. 285, pp. 523-525
C.C. (1982).- Taxonomie du genre Lingula (Brachiopodes, Inarticulés).- Bulletin du Muséum National d'Histoire Naturelle, Paris, Sér. 4, Vol. 4, N° 3/4, pp. 337-367
C.C. (1983).- Taxonomie du genre Glottidia (Brachiopodes Inarticulés).- Bulletin du Muséum National d'Histoire Naturelle, Paris, Sér. 4, Vol. 5 (Sect. 4), N° 2, pp. 469-489.
P.Y., A., C.C., T., D. and M. (1996).- New paleontological and geological data on the Ordovician and Silurian of Bolivia.- Journal of South America Earth Sciences, Vol. 9, N° 5/6, pp. 329-347.
L.E. and L.E. (2000).- Lingulida. In: R.L. (Ed.), Revised Brachiopoda.- Treatise on Invertebrate Paleontology, Geological Society of America, New York, and University of Kansas, Lawrence, Part H, Vol. 2, pp. 32-95.
INTERNATIONAL COMMISSION ON ZOOLOGICAL NOMENCLATURE (1999).- International code of zoological nomenclature, Fourth Edition, adopted by the International Union of Biological Sciences, The International Trust for Zoological Nomenclature / The Natural History Museum, London, xxix + 306 pp.
K.K. and L.E., In: K.K., L.E. and L.M. (1984).- Inarticulate brachiopods, ostracodes (bradoriids) and hyolithelminths from the Sablinka Formation of the Leningrad District (in Russian).- Paleontologicheskiy Zhurnal, Moscow, Vol. 1984, N° 4, pp. 33-47.
- A., C.C. and J.M. (1999).- Triassic lingulide brachiopods from the Iberian Range (Spain).- Géobios, Lyon, Vol. 32, pp. 815-821.
A. (1896).- Über die Brachiopodengattung Obolus .- Mémoires de l'Académie Impériale des Sciences de St Pétersbourg, Classe physico-mathématique, St-Péterbourg, Vol. 4, N° 2, pp. 1-216.
C.H. (1830).- Beiträge zur Geognosie des Russischen Reiches, St. Petersburg, 165 p.
L., K.K., N.G., S.P. and R.F. (1989).- The Key sections and stratigraphy of the phosphate bearing Obolus beds of the North-East of the Russian platform (in Russian). Nauka Trudy, Leningrad, Vol. 18, pp. 1-222.
I. (1996).- Lingulate brachiopods and biostratigraphy of the Cambrian-Ordovician boundary beds in Baltoscandia.- Dr. Thesis, Univ. Uppsala, 136 pp.
A.J. (1965).- Inarticulata. In: R.C. (Ed.), Brachiopoda.- Treatise on Invertebrate Paleontology, Geological Society of America, New York, and University of Kansas, Lawrence, Part H, p. 260-296.
C. and C.M. (1929).- Brachiopoda (generum et genotyporum index et bibliographia). In: Fossilium Catalogues, Vol. 1, Animalia, Pars 42. Junk, Berlin 140 p.
E. O. (1981).- Phylogenetics. The theory and practice of phylogenetic systematics.- Wiley-Interscience, New-York, 439 p.
Click on thumbnail to enlarge the image.
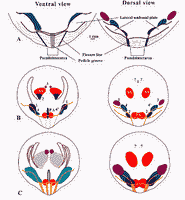
Figure 1: Diagrams of the main taxonomic characters of Obolus apollinis.
A. Umbonal regions (internal view), see also Fig.
2
![]() ,
3
,
3
![]() ;
;
B. Average disposition of muscles and main mantle canals (drawn from a real specimen);
C. Variability in musculature (established from more than 20 specimens) and main mantle canals (on 3 specimens).
Muscle terminology as established by
(1982):
1 Anterior Oblique;
2 Anterior Lateral Oblique;
3 Median Lateral Oblique;
4 Anterior Internal Oblique;
4' Median Internal Oblique;
AA Anterior Adductor;
PA Posterior Adductor.
Click on thumbnail to enlarge the image.
Figure 2: Obolus apollinis: Internal umbonal region of the ventral valve: profile view from the beak and a frontal view.
Click on thumbnail to enlarge the image.
Figure 3: Obolus apollinis: Internal umbonal region of the dorsal valve: frontal view and postero-lateral view with well-marked flexure lines.

Click on thumbnail to enlarge the image.
Table 1: Obolus , 1829: the previous diagnosis and the proposed one emended (in italics the non-taxonomic characters, without generic significance).

Click on thumbnail to enlarge the image.
Table 2: The new diagnosis of Obolus apollinis , 1829, is compared to the previous diagnosis as translated and modified by (1996) from the paper of et alii (1989), concerning O. apollinis, O. ruchini, O. transversus, and Ungula convexa. Non-taxonomic characters are in italics.