![]()
![]()
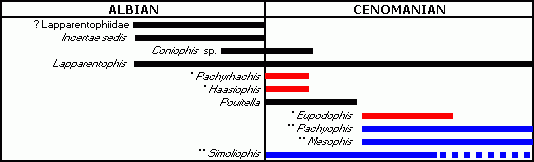
![]() Three "snakes with legs" are known: Pachyrhachis problematicus, Haasiophis terrasanctus and Eupodophis descouensi. They have short posterior limbs but lack an anterior girdle and forelimbs. Moreover, Pachyophis woodwardi, Mesophis nopcsai and Simoliophis ssp. appear to be closely related to the hindlimbed taxa; consequently, although the presence of posterior limbs has not been demonstrated for these genera, it is presumed that they too were hindlimbed. All these snakes have been recovered only from the Cenomanian. Moreover, these six genera come from a restricted area (western Europe and
northwesternmost Africa to the Middle East). This limited geographic range suggests that snakes might have originated in the "Mediterranean" part of the Tethys, but the restricted stratigraphical range remains unexplained.
Three "snakes with legs" are known: Pachyrhachis problematicus, Haasiophis terrasanctus and Eupodophis descouensi. They have short posterior limbs but lack an anterior girdle and forelimbs. Moreover, Pachyophis woodwardi, Mesophis nopcsai and Simoliophis ssp. appear to be closely related to the hindlimbed taxa; consequently, although the presence of posterior limbs has not been demonstrated for these genera, it is presumed that they too were hindlimbed. All these snakes have been recovered only from the Cenomanian. Moreover, these six genera come from a restricted area (western Europe and
northwesternmost Africa to the Middle East). This limited geographic range suggests that snakes might have originated in the "Mediterranean" part of the Tethys, but the restricted stratigraphical range remains unexplained.
Cenomanian; Cretaceous; Pachyostosis; Paleogeography; Reptilia; Hindlimbed Snakes.
Rage J.-C., Escuillié F. (2003).- The Cenomanian: stage of hindlimbed snakes.- Carnets de Géologie / Notebooks on Geology, Maintenon, Article 2003/01 (CG2003_A01)
![]() Le Cénomanien: étage des serpents bipèdes.-
Trois serpents munis de pattes sont connus: Pachyrhachis problematicus, Haasiophis terrasanctus et Eupodophis descouensi. Ils possèdent des membres et une ceinture postérieurs mais n'ont ni ceinture ni membres
antérieurs, c'est-à-dire qu'ils sont bipèdes. De plus, Pachyophis woodwardi, Mesophis nopcsai et Simoliophis ssp. semblent être étroitement apparentés aux serpents bipèdes; par
conséquent, bien que des membres postérieurs n'aient pas été mis en évidence chez
eux, il est supposé qu'ils étaient aussi bipèdes. Tous les serpents bipèdes et ceux qui sont supposés l'avoir été ont été trouvés dans le Cénomanien
seulement. De plus, ces six genres proviennent d'une aire géographique réduite (de l'Europe occidentale et Afrique du nord-ouest au
Moyen-Orient). Cette répartition géographique limitée suggère que les serpents sont nés dans la partie
"méditerranéenne" de la Téthys, mais la distribution stratigraphique restreinte reste
inexpliquée.
Le Cénomanien: étage des serpents bipèdes.-
Trois serpents munis de pattes sont connus: Pachyrhachis problematicus, Haasiophis terrasanctus et Eupodophis descouensi. Ils possèdent des membres et une ceinture postérieurs mais n'ont ni ceinture ni membres
antérieurs, c'est-à-dire qu'ils sont bipèdes. De plus, Pachyophis woodwardi, Mesophis nopcsai et Simoliophis ssp. semblent être étroitement apparentés aux serpents bipèdes; par
conséquent, bien que des membres postérieurs n'aient pas été mis en évidence chez
eux, il est supposé qu'ils étaient aussi bipèdes. Tous les serpents bipèdes et ceux qui sont supposés l'avoir été ont été trouvés dans le Cénomanien
seulement. De plus, ces six genres proviennent d'une aire géographique réduite (de l'Europe occidentale et Afrique du nord-ouest au
Moyen-Orient). Cette répartition géographique limitée suggère que les serpents sont nés dans la partie
"méditerranéenne" de la Téthys, mais la distribution stratigraphique restreinte reste
inexpliquée.
![]() Cénomanien; Crétacé; Paléogéographie; Reptiles; Serpents bipèdes.
Cénomanien; Crétacé; Paléogéographie; Reptiles; Serpents bipèdes.
![]()
![]() That snakes are tetrapods was established long ago.
Furthermore, the representatives of most existing families have vestiges of a pelvic girdle and posterior limbs. However, in all modern snakes the pelvic girdle, if indeed present, has lost all bone-to-bone contact with the vertebral column. The limb is but a single very small bone that logically is considered to represent the femur. Thus the Leptotyphlopidae, several
anilioids (Anilius, Cylindrophis, Anomochilus), the booid Loxocemus and the Boidae have vestiges of a pelvis and a femur. These elements are also present in the Tropidophiidae s.l., but only in the male. As for the Typhlopidae, they still have a pelvis but no femur. Note that there is no trace of forelimbs or an anterior girdle in snakes, living or fossil.
That snakes are tetrapods was established long ago.
Furthermore, the representatives of most existing families have vestiges of a pelvic girdle and posterior limbs. However, in all modern snakes the pelvic girdle, if indeed present, has lost all bone-to-bone contact with the vertebral column. The limb is but a single very small bone that logically is considered to represent the femur. Thus the Leptotyphlopidae, several
anilioids (Anilius, Cylindrophis, Anomochilus), the booid Loxocemus and the Boidae have vestiges of a pelvis and a femur. These elements are also present in the Tropidophiidae s.l., but only in the male. As for the Typhlopidae, they still have a pelvis but no femur. Note that there is no trace of forelimbs or an anterior girdle in snakes, living or fossil.
![]() One might expect to find fossil snakes with limbs, but it is only recently that such fossils have been recognized or discovered. Three taxa are unquestionably snakes with hind legs; they are known from articulated and largely complete skeletons. They represent three distinct genera. These snakes are very similar: they all have a pelvic girdle and short but well developed hind legs. Curiously, all were found in strata of Cenomanian age. So they are among the oldest snakes yet found, although some older and problematic remains are considered to be snakes (see below and
Fig. 1
One might expect to find fossil snakes with limbs, but it is only recently that such fossils have been recognized or discovered. Three taxa are unquestionably snakes with hind legs; they are known from articulated and largely complete skeletons. They represent three distinct genera. These snakes are very similar: they all have a pelvic girdle and short but well developed hind legs. Curiously, all were found in strata of Cenomanian age. So they are among the oldest snakes yet found, although some older and problematic remains are considered to be snakes (see below and
Fig. 1
![]() ).
).
![]()
![]() The oldest snakes come from the middle Cretaceous. One had been reported but not named from the Barremian of Spain
(Rage & Richter,
1994). It was identified on the basis of isolated vertebrae. Afterward, the study of Cretaceous lizards showed that the characters supposedly demonstrating the Spanish fossil to be a snake also exist in some lizards (the presence of a zygosphenal roof, although it is notched; the existence of subcentral ridges reaching the ventral part of the paradiapophyses). Consequently, the Barremian fossil can no longer be regarded as a snake.
The oldest snakes come from the middle Cretaceous. One had been reported but not named from the Barremian of Spain
(Rage & Richter,
1994). It was identified on the basis of isolated vertebrae. Afterward, the study of Cretaceous lizards showed that the characters supposedly demonstrating the Spanish fossil to be a snake also exist in some lizards (the presence of a zygosphenal roof, although it is notched; the existence of subcentral ridges reaching the ventral part of the paradiapophyses). Consequently, the Barremian fossil can no longer be regarded as a snake.
![]() The oldest unequivocal snakes known come from the Upper Albian (level C of the "série de Tiout") of Algeria. They are represented only by poorly preserved
vertebrae (Cuny et alii,
1990) but incontestably they are snakes. The vertebrae represent a possible Lapparentophiidae and a snake
incertae sedis
(Fig. 1
The oldest unequivocal snakes known come from the Upper Albian (level C of the "série de Tiout") of Algeria. They are represented only by poorly preserved
vertebrae (Cuny et alii,
1990) but incontestably they are snakes. The vertebrae represent a possible Lapparentophiidae and a snake
incertae sedis
(Fig. 1
![]() ). The first was certainly terrestrial, but the second shows possible adaptations to an aquatic life.
Lapparentophis defrennei (Lapparentophiidae), a terrestrial snake also found in Algeria
(Hoffstetter,
1959), comes from the "série d'In Akhamil" that may correspond to the Upper Albian or Cenomanian
(Cuny et alii,
1990). In North America (USA), Coniophis sp. has been reported from the Upper Albian or Lower Cenomanian
(Gardner & Cifelli,
1999); this terrestrial snake, probably a burrower, is attributed to the Aniliidae
s.l., a family that at least when its fossil representatives are included is probably paraphyletic and is comprised of generally primitive forms. These snakes are known only from isolated vertebrae so it is impossible to determine if they had legs.
). The first was certainly terrestrial, but the second shows possible adaptations to an aquatic life.
Lapparentophis defrennei (Lapparentophiidae), a terrestrial snake also found in Algeria
(Hoffstetter,
1959), comes from the "série d'In Akhamil" that may correspond to the Upper Albian or Cenomanian
(Cuny et alii,
1990). In North America (USA), Coniophis sp. has been reported from the Upper Albian or Lower Cenomanian
(Gardner & Cifelli,
1999); this terrestrial snake, probably a burrower, is attributed to the Aniliidae
s.l., a family that at least when its fossil representatives are included is probably paraphyletic and is comprised of generally primitive forms. These snakes are known only from isolated vertebrae so it is impossible to determine if they had legs.
![]() The Cenomanian has provided several taxa. Only one of them was terrestrial. This is Pouitella, from the Lower Cretaceous of France
(Rage,
1988); represented by only one vertebra, it is impossible to know if it had legs. The others were completely adapted to an aquatic life and all were found in marine sediments. Pachyrhachis, Eupodophis and Haasiophis are three snakes from the Cenomanian of the Middle East with hind legs.
Pachyophis and Mesophis come from the Middle or more probably from the Upper Cenomanian of Bosnia.
Lee et alii
(1999) supposed that Pachyophis may have had hind legs (see below). Mesophis is very poorly known; the only specimen seems to be lost. Finally, Simoliophis, represented by vertebrae and isolated ribs is known from the Lower, Middle, and perhaps Upper Cenomanian of Western Europe and North Africa.
The Cenomanian has provided several taxa. Only one of them was terrestrial. This is Pouitella, from the Lower Cretaceous of France
(Rage,
1988); represented by only one vertebra, it is impossible to know if it had legs. The others were completely adapted to an aquatic life and all were found in marine sediments. Pachyrhachis, Eupodophis and Haasiophis are three snakes from the Cenomanian of the Middle East with hind legs.
Pachyophis and Mesophis come from the Middle or more probably from the Upper Cenomanian of Bosnia.
Lee et alii
(1999) supposed that Pachyophis may have had hind legs (see below). Mesophis is very poorly known; the only specimen seems to be lost. Finally, Simoliophis, represented by vertebrae and isolated ribs is known from the Lower, Middle, and perhaps Upper Cenomanian of Western Europe and North Africa.
![]()
Pachyrhachis problematicus Haas, 1979
![]() In
1979 Haas described a fossil under the name Pachyrhachis problematicus. This fossil, found in the house of a quarrier, comes from the Cenomanian limestone of Ein Jabrud near Ramallah in
Palestine. Its age, earliest Cenomanian (Chalifa & Tchernov,
1982), is unquestioned. The fossil comprises the skull and a large portion of the vertebral column, but not the posterior part. The skull proved indisputably that the animal was a squamate and the vertebral column showed that it was snake-like. But
Haas, although indicating that the skull had characteristics of a snake, considered the fossil to be a varanoid lizard (Platynota). He was concerned particularly with the primitive characteristics of the skull (that obviously suggested affinities with lizards) and the pachyostosis
(i.e. thickening) of a part of the vertebrae and ribs. This pachyostosis resembled that of Simoliophis, a Cenomanian fossil known only from vertebrae and ribs, and generally considered to be a snake. Based on vertebral morphology,
Haas put Pachyrhachis in the same family as Simoliophis (Simoliophiidae) and considered the two genera to be snake-like varanoid lizards.
In
1979 Haas described a fossil under the name Pachyrhachis problematicus. This fossil, found in the house of a quarrier, comes from the Cenomanian limestone of Ein Jabrud near Ramallah in
Palestine. Its age, earliest Cenomanian (Chalifa & Tchernov,
1982), is unquestioned. The fossil comprises the skull and a large portion of the vertebral column, but not the posterior part. The skull proved indisputably that the animal was a squamate and the vertebral column showed that it was snake-like. But
Haas, although indicating that the skull had characteristics of a snake, considered the fossil to be a varanoid lizard (Platynota). He was concerned particularly with the primitive characteristics of the skull (that obviously suggested affinities with lizards) and the pachyostosis
(i.e. thickening) of a part of the vertebrae and ribs. This pachyostosis resembled that of Simoliophis, a Cenomanian fossil known only from vertebrae and ribs, and generally considered to be a snake. Based on vertebral morphology,
Haas put Pachyrhachis in the same family as Simoliophis (Simoliophiidae) and considered the two genera to be snake-like varanoid lizards.
![]() Haas remarked on a particularly striking aspect of the skull, its macrostomate (large-mouth) structure (see below: Phyletic Considerations), but did not take it into account in establishing the affinities of Pachyrhachis.
Haas remarked on a particularly striking aspect of the skull, its macrostomate (large-mouth) structure (see below: Phyletic Considerations), but did not take it into account in establishing the affinities of Pachyrhachis.
![]() In 1980, Haas described (as
Ophiomorphus colberti) a second serpentiform squamate acquired under the same conditions and from the same locality as
Pachyrhachis. The fossil consists of a vertebral column (a part of its anterior region and the posterior part of the tail are missing), a pelvic girdle and short hind legs. In addition there is a poorly preserved skull under the vertebral column. The vertebrae and ribs are pachyostotic, but less so than in
Pachyrhachis. Haas interpreted this fossil as an intermediate between varanoid lizards and snakes.
In 1980, Haas described (as
Ophiomorphus colberti) a second serpentiform squamate acquired under the same conditions and from the same locality as
Pachyrhachis. The fossil consists of a vertebral column (a part of its anterior region and the posterior part of the tail are missing), a pelvic girdle and short hind legs. In addition there is a poorly preserved skull under the vertebral column. The vertebrae and ribs are pachyostotic, but less so than in
Pachyrhachis. Haas interpreted this fossil as an intermediate between varanoid lizards and snakes.
![]() Wallach
(1984) thereafter proposed the genus name Estesius to replace Ophiomorphus, which was preoccupied.
Wallach
(1984) thereafter proposed the genus name Estesius to replace Ophiomorphus, which was preoccupied.
![]() McDowell
(1987) was the first to consider forthrightly that Pachyrhachis is a snake; on the other hand he removed Ophiomorphus
(i.e. Estesius) from this group.
McDowell
(1987) was the first to consider forthrightly that Pachyrhachis is a snake; on the other hand he removed Ophiomorphus
(i.e. Estesius) from this group.
![]() In 1997, Caldwell
& Lee showed that
In 1997, Caldwell
& Lee showed that
![]() 1) the specimen described as Pachyrhachis problematicus is indeed a snake, and
1) the specimen described as Pachyrhachis problematicus is indeed a snake, and
![]() 2) that the specimen corresponding to Estesius colberti belongs to Pachyrhachis problematicus
, the difference in pachyostosis being ontogenetic. Therefore, Pachyrhachis problematicus was a snake with hind legs! Some doubt might still exist because the skull that demonstrates its serpenthood is on one specimen while the legs are on the other specimen (the skull of the latter specimen is present but it is crushed under the ribs). But subsequently the discovery of Eupodophis descouensi
and Haasiophis terrasanctus proved that Caldwell & Lee
(1997) were right to put the two specimens in one taxon.
2) that the specimen corresponding to Estesius colberti belongs to Pachyrhachis problematicus
, the difference in pachyostosis being ontogenetic. Therefore, Pachyrhachis problematicus was a snake with hind legs! Some doubt might still exist because the skull that demonstrates its serpenthood is on one specimen while the legs are on the other specimen (the skull of the latter specimen is present but it is crushed under the ribs). But subsequently the discovery of Eupodophis descouensi
and Haasiophis terrasanctus proved that Caldwell & Lee
(1997) were right to put the two specimens in one taxon.
![]() The anatomy of
Pachyrhachis problematicus was revised in detail by Lee & Caldwell
(1998). In particular it shows that Pachyrhachis was truly macrostomate. Pachyrhachis is the largest of the hindlimbed snakes, but its length is difficult to estimate (more than 1.5 meters).
The anatomy of
Pachyrhachis problematicus was revised in detail by Lee & Caldwell
(1998). In particular it shows that Pachyrhachis was truly macrostomate. Pachyrhachis is the largest of the hindlimbed snakes, but its length is difficult to estimate (more than 1.5 meters).
Eupodophis descouensi (Rage & Escuillié, 2000)
![]() This taxon is represented by a specimen found in the Middle Cenomanian of Al Nammoura in Lebanon
(Fig. 2
This taxon is represented by a specimen found in the Middle Cenomanian of Al Nammoura in Lebanon
(Fig. 2 ![]() , 3
, 3
![]() ).
The specimen includes the skull, the vertebral column (a part is missing in the posterior third), the pelvic girdle and the hind limb. The skull is less well-preserved than that of
Pachyrhachis but it confirms the macrostomate condition of hindlimbed snakes. Most presacral vertebrae and ribs are thickened and pachyostotic. One of the most interesting points is the excellent preservation of the posterior portion of the vertebral column which shows that the tail was astonishingly short (Fig.
4
).
The specimen includes the skull, the vertebral column (a part is missing in the posterior third), the pelvic girdle and the hind limb. The skull is less well-preserved than that of
Pachyrhachis but it confirms the macrostomate condition of hindlimbed snakes. Most presacral vertebrae and ribs are thickened and pachyostotic. One of the most interesting points is the excellent preservation of the posterior portion of the vertebral column which shows that the tail was astonishingly short (Fig.
4
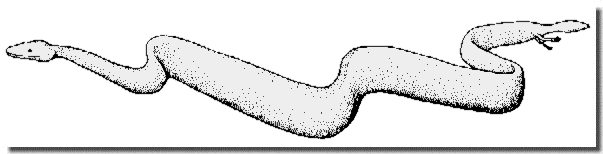
![]() ). While the total length of the animal is estimated at 85 cm, the leg is located only 5 cm from the end of the tail. The leg is about 2 cm long. In the tail the chevron bones are articulated with the vertebrae, not united to them; this is a plesiomorphic (i.e. primitive) character important in phylogenetic analysis.
). While the total length of the animal is estimated at 85 cm, the leg is located only 5 cm from the end of the tail. The leg is about 2 cm long. In the tail the chevron bones are articulated with the vertebrae, not united to them; this is a plesiomorphic (i.e. primitive) character important in phylogenetic analysis.
![]() Eupodophis
differs from Pachyrhachis in the more pointed form of the skull, by its quadrate bone that is much less drawn out anteroposteriorly and by the presence of special dorsal tubercles on the vertebrae (Rage & Escuillié,
2000). In addition, the body seems to be more flattened laterally than in
Pachyrhachis.
Eupodophis
differs from Pachyrhachis in the more pointed form of the skull, by its quadrate bone that is much less drawn out anteroposteriorly and by the presence of special dorsal tubercles on the vertebrae (Rage & Escuillié,
2000). In addition, the body seems to be more flattened laterally than in
Pachyrhachis.
Haasiophis terrasanctus Tchernov et alii, 2000
![]() This taxon too is represented by only one specimen; it is almost complete and undoubtedly the best preserved of all the specimens of hindlimbed snakes. It comes from the same site as
P. problematicus and therefore is dated earliest Cenomanian. Its size is close to that of Eupodophis. The published description is rather brief and concerns mainly the skull; in particular the caudal region is poorly described and the description must be completed in its entirety before its characteristics can be fully known. It was flattened laterally but the degree of flattening has not been determined. Judged by its principal characteristics it is very similar to the two other taxa. It is distinct in its very much reduced coronoid and a much less marked macrostomate condition.
This taxon too is represented by only one specimen; it is almost complete and undoubtedly the best preserved of all the specimens of hindlimbed snakes. It comes from the same site as
P. problematicus and therefore is dated earliest Cenomanian. Its size is close to that of Eupodophis. The published description is rather brief and concerns mainly the skull; in particular the caudal region is poorly described and the description must be completed in its entirety before its characteristics can be fully known. It was flattened laterally but the degree of flattening has not been determined. Judged by its principal characteristics it is very similar to the two other taxa. It is distinct in its very much reduced coronoid and a much less marked macrostomate condition.
![]() In addition it differs from
Pachyrhachis in its anteroposteriorly narrow quadrate and from Eupodophis in the absence of dorsal tubercles on the vertebrae. The position of the leg in relation to the caudal extremity is unknown. Furthermore, the presence of haemapophyses in the tail reported by
Tchernov et alii
(2000), poses a problem: does it concern haemapophyses in a strict sense (united to the centrum) or are they chevrons (articulated to the centrum)? The presence of true haemapophyses in Haasiophis, while Eupodophis has articulated chevrons, would be surprising.
In addition it differs from
Pachyrhachis in its anteroposteriorly narrow quadrate and from Eupodophis in the absence of dorsal tubercles on the vertebrae. The position of the leg in relation to the caudal extremity is unknown. Furthermore, the presence of haemapophyses in the tail reported by
Tchernov et alii
(2000), poses a problem: does it concern haemapophyses in a strict sense (united to the centrum) or are they chevrons (articulated to the centrum)? The presence of true haemapophyses in Haasiophis, while Eupodophis has articulated chevrons, would be surprising.
Pachyophis woodwardi Nopcsa, 1923
![]() This snake too is represented by only one specimen. It comes from a quarry near Bileca in Bosnia-Herzegovina. At first considered as
early Cretaceous (Neocomian) it is now dated as middle or probably late Cretaceous
(Sliskovic,
1970).
This snake too is represented by only one specimen. It comes from a quarry near Bileca in Bosnia-Herzegovina. At first considered as
early Cretaceous (Neocomian) it is now dated as middle or probably late Cretaceous
(Sliskovic,
1970).![]()
![]() A little smaller than Eupodophis, it must have reached some sixty centimeters. The specimen includes the dentition, some fragments of skull bones and the vertebral column (vertebrae and ribs) but the caudal portion is hidden under another part of the column (Lee et alii,
1999). The vertebrae and ribs are strongly pachyostotic; the ribs, very thickened, form an almost continuous "wall". The lateral flattening of the body must have been comparable to that in Eupodophis. It is difficult to compare
Pachyophis to the other taxa because it is known only from its dorsal side and the cranial bones are very incomplete. However it can be noted that pachyostosis is stronger than that in the other taxa, although the animal is smaller. So the difference in the degree of pachyostosis cannot be attributed to ontogeny, but represents a valid characteristic of this genus.
A little smaller than Eupodophis, it must have reached some sixty centimeters. The specimen includes the dentition, some fragments of skull bones and the vertebral column (vertebrae and ribs) but the caudal portion is hidden under another part of the column (Lee et alii,
1999). The vertebrae and ribs are strongly pachyostotic; the ribs, very thickened, form an almost continuous "wall". The lateral flattening of the body must have been comparable to that in Eupodophis. It is difficult to compare
Pachyophis to the other taxa because it is known only from its dorsal side and the cranial bones are very incomplete. However it can be noted that pachyostosis is stronger than that in the other taxa, although the animal is smaller. So the difference in the degree of pachyostosis cannot be attributed to ontogeny, but represents a valid characteristic of this genus.
![]() Lee et alii
(1999), noting the similarities between Pachyophis and Pachyrhachis (the only hindlimbed taxon known when this work was published) concluded that Pachyophis might also have hind legs. If they are present, which is quite possible or even probable, they must be hidden under the medial portion of the vertebral column.
Lee et alii
(1999), noting the similarities between Pachyophis and Pachyrhachis (the only hindlimbed taxon known when this work was published) concluded that Pachyophis might also have hind legs. If they are present, which is quite possible or even probable, they must be hidden under the medial portion of the vertebral column.
![]() Lee et alii
(1999) considered that Pachyophis and Pachyrhachis form a clade and put them in the same family, the Pachyophiidae. Based on the available material, it does not seem possible to confirm this point of view; the possibility that hindlimbed snakes form a paraphyletic assemblage cannot be set aside definitively (Rage & Escuillié,
2000).
Lee et alii
(1999) considered that Pachyophis and Pachyrhachis form a clade and put them in the same family, the Pachyophiidae. Based on the available material, it does not seem possible to confirm this point of view; the possibility that hindlimbed snakes form a paraphyletic assemblage cannot be set aside definitively (Rage & Escuillié,
2000).
Mesophis nopcsai Bolkay, 1925
![]() This taxon is represented by an almost complete specimen in which the caudal portion is poorly preserved and perhaps incomplete. Apparently the specimen is lost so this snake is known only from Bolkay's
(1925) description which is not complete enough to establish its relations to the other taxa. It was found in the same site as
Pachyophis. It is flattened laterally but the degree of flattening could not be determined. A part of the vertebrae are pachyostotic while the ribs are not at all or but weakly affected by pachyostotis. Because it is clearly smaller than
Pachyophis (approximately half its size) it is possible that the difference is ontogenetic and as the two taxa come from the same site it is possible to suppose, as did
Nopcsa
(1931: 30), that Mesophis is a young Pachyophis.
This taxon is represented by an almost complete specimen in which the caudal portion is poorly preserved and perhaps incomplete. Apparently the specimen is lost so this snake is known only from Bolkay's
(1925) description which is not complete enough to establish its relations to the other taxa. It was found in the same site as
Pachyophis. It is flattened laterally but the degree of flattening could not be determined. A part of the vertebrae are pachyostotic while the ribs are not at all or but weakly affected by pachyostotis. Because it is clearly smaller than
Pachyophis (approximately half its size) it is possible that the difference is ontogenetic and as the two taxa come from the same site it is possible to suppose, as did
Nopcsa
(1931: 30), that Mesophis is a young Pachyophis.
![]() Whatever the case, one can only conjecture as long as the specimen is not found again. It might well be another hindlimbed snake.
Whatever the case, one can only conjecture as long as the specimen is not found again. It might well be another hindlimbed snake.
Simoliophis Sauvage, 1880
![]() In contrast with the genera discussed previously, Simoliophis comprises several species. On the other hand it is represented by disarticulated specimens consisting only of vertebrae and ribs.
In contrast with the genera discussed previously, Simoliophis comprises several species. On the other hand it is represented by disarticulated specimens consisting only of vertebrae and ribs.
![]() Pachyostosis affects the vertebrae of the middle and posterior trunk portions as well as the ribs that probably came from the same regions. Vertebral morphology is very
similar to that of Eupodophis, the only unquestionable hindlimbed snake of which a disarticulated vertebra is known. The vertebrae of Simoliophis are distinguished from those of
Eupodophis by the absence of the dorsal tubercles that characterize the neural arch of that genus. On the other hand, what is known about
Pachyrhachis and Haasiophis does not allow a distinction between the vertebrae of these two genera and those of Simoliophis, unless the reconstructions of Pachyrhachis vertebrae as proposed by Lee & Caldwell
(1998) are exact. If that be the case, Pachyrhachis's vertebrae would be quite different from those of
Eupodophis and Simoliophis. As reconstructed, the vertebrae of Pachyrhachis differ from those of the two other genera mainly in having by far less pachyostotic and more cylindrical centra, more elongated prezygapophyses, less protruding paradiapophyses, and larger condyles and cotyles. Whatever may be determined, the vertebrae of Eupodophis and Simoliophis (and probably those of Pachyrhachis
and Haasiophis) are very similar and are markedly different from those of non-hindlimbed snakes. For this reason one may postulate that these taxa are either closely related or at least at a similar grade, and therefore the existence of hind legs in Simoliophis can be inferred with some confidence.
Pachyostosis affects the vertebrae of the middle and posterior trunk portions as well as the ribs that probably came from the same regions. Vertebral morphology is very
similar to that of Eupodophis, the only unquestionable hindlimbed snake of which a disarticulated vertebra is known. The vertebrae of Simoliophis are distinguished from those of
Eupodophis by the absence of the dorsal tubercles that characterize the neural arch of that genus. On the other hand, what is known about
Pachyrhachis and Haasiophis does not allow a distinction between the vertebrae of these two genera and those of Simoliophis, unless the reconstructions of Pachyrhachis vertebrae as proposed by Lee & Caldwell
(1998) are exact. If that be the case, Pachyrhachis's vertebrae would be quite different from those of
Eupodophis and Simoliophis. As reconstructed, the vertebrae of Pachyrhachis differ from those of the two other genera mainly in having by far less pachyostotic and more cylindrical centra, more elongated prezygapophyses, less protruding paradiapophyses, and larger condyles and cotyles. Whatever may be determined, the vertebrae of Eupodophis and Simoliophis (and probably those of Pachyrhachis
and Haasiophis) are very similar and are markedly different from those of non-hindlimbed snakes. For this reason one may postulate that these taxa are either closely related or at least at a similar grade, and therefore the existence of hind legs in Simoliophis can be inferred with some confidence.
![]() Simoliophis has been found only in strata of Cenomanian age (see below).
Simoliophis has been found only in strata of Cenomanian age (see below).
![]() Two valid species have been described,
S. rochebrunei and S. libycus. In addition, specimens of Simoliophis from the Cenomanian of Baharija (Egypt), referred to
S. rochebrunei by Nopcsa
(1925), represent a third, unnamed species.
Two valid species have been described,
S. rochebrunei and S. libycus. In addition, specimens of Simoliophis from the Cenomanian of Baharija (Egypt), referred to
S. rochebrunei by Nopcsa
(1925), represent a third, unnamed species.
Simoliophis rochebrunei Sauvage, 1880
![]() S. rochebrunei, the type species of the genus, has been found in the Cenomanian of the central west and southwest of France as well as in Portugal, near Lisbon (Jonet,
1981; Rage, 1984). The species has been recovered from a dozen localities that with two exceptions are dated either
Early or Middle Cenomanian. The exact level of the Cenomanian present at Sillac and Basseau, two localities in the Charente now lost, is not known. Consequently it may be that S. rochebrunei occurs in the
Late Cenomanian.
S. rochebrunei, the type species of the genus, has been found in the Cenomanian of the central west and southwest of France as well as in Portugal, near Lisbon (Jonet,
1981; Rage, 1984). The species has been recovered from a dozen localities that with two exceptions are dated either
Early or Middle Cenomanian. The exact level of the Cenomanian present at Sillac and Basseau, two localities in the Charente now lost, is not known. Consequently it may be that S. rochebrunei occurs in the
Late Cenomanian.
Simoliophis libycus Nessov, Zhegallo & Averianov, 1998
![]() This species was found in Libya at Draa Ubari. According to Nessov et alii
(1998), it is distinguishable from S. rochebrunei by a narrower centrum, a lower neural spine on the anterior vertebrae, the smaller anteroposterior length of the neural spine, and less well developed paradiapophyses. As the species was based on a small amount of material and is smaller than
S. rochebrunei, it remains to be determined whether or not these differences are only ontogenetic and/or are related to the position of the vertebrae in the vertebral column.
This species was found in Libya at Draa Ubari. According to Nessov et alii
(1998), it is distinguishable from S. rochebrunei by a narrower centrum, a lower neural spine on the anterior vertebrae, the smaller anteroposterior length of the neural spine, and less well developed paradiapophyses. As the species was based on a small amount of material and is smaller than
S. rochebrunei, it remains to be determined whether or not these differences are only ontogenetic and/or are related to the position of the vertebrae in the vertebral column.
![]() Nessov et alii
(1998) considered the age of the outcrops at Draa Ubari as Santonian-Campanian, but all of the stratigraphically significant taxa found at the site are of Cenomanian age, and some may be older (Rage & Cappetta,
2002); the exact age within the Cenomanian cannot be established.
Nessov et alii
(1998) considered the age of the outcrops at Draa Ubari as Santonian-Campanian, but all of the stratigraphically significant taxa found at the site are of Cenomanian age, and some may be older (Rage & Cappetta,
2002); the exact age within the Cenomanian cannot be established.
The other Simoliophis
![]() Nopcsa
(1925) described a series of vertebrae and ribs from the Cenomanian of Baharija, Egypt. He attributed these fossils to S. rochebrunei, the only species then known. In fact, in reconstructing the vertebral column,
Nopcsa probably mixed the vertebrae of two snakes. Some of them, middle and posterior trunk vertebrae, are truly Simoliophis but those considered anterior trunk may belong to another distinct genus (opinion based on as yet unpublished specimens of Simoliophis). In addition, the vertebrae of Simoliophis belong neither to S. rochebrunei
nor to S. libycus, but to another, unnamed species as shown by the marked groove on the base of the anterior edge of the neural spine.
Nopcsa
(1925) described a series of vertebrae and ribs from the Cenomanian of Baharija, Egypt. He attributed these fossils to S. rochebrunei, the only species then known. In fact, in reconstructing the vertebral column,
Nopcsa probably mixed the vertebrae of two snakes. Some of them, middle and posterior trunk vertebrae, are truly Simoliophis but those considered anterior trunk may belong to another distinct genus (opinion based on as yet unpublished specimens of Simoliophis). In addition, the vertebrae of Simoliophis belong neither to S. rochebrunei
nor to S. libycus, but to another, unnamed species as shown by the marked groove on the base of the anterior edge of the neural spine.
![]() There is no agreement on the precise age of the Baharija locality within the Cenomanian: For some,
Early Cenomanian (Lapparent de Broin,
2000), for others Late Cenomanian (Werner,
1989). However, on the basis of ammonites Dominik (1985)
proposed a Late Cenomanian age for at least a part of the Baharija Formation.
There is no agreement on the precise age of the Baharija locality within the Cenomanian: For some,
Early Cenomanian (Lapparent de Broin,
2000), for others Late Cenomanian (Werner,
1989). However, on the basis of ammonites Dominik (1985)
proposed a Late Cenomanian age for at least a part of the Baharija Formation.
![]() Several localities in the Kem Kem (Morocco) have also furnished Simoliophis vertebrae that resemble those of
S. libycus (work in progress). The Kem Kem beds are mainly of Cenomanian age but it is not impossible that the lowermost levels are of
Late Albian age. However, all of the levels that furnished specimens of Simoliophis are dated Cenomanian (Sereno et alii,
1996).
Several localities in the Kem Kem (Morocco) have also furnished Simoliophis vertebrae that resemble those of
S. libycus (work in progress). The Kem Kem beds are mainly of Cenomanian age but it is not impossible that the lowermost levels are of
Late Albian age. However, all of the levels that furnished specimens of Simoliophis are dated Cenomanian (Sereno et alii,
1996).
![]() It is incontestable that three snakes have hind limbs:
Pachyrhachis problematicus and Haasiophis terrasanctus of the
earliest Cenomanian, along with Eupodophis descouensi of the Middle Cenomanian. All three were found in the Middle East. Three other genera may possess such limbs: Pachyophis (unique species: P. woodwardi),
Mesophis (unique species: M. nopcsai), both from the Middle, or more probably the
Late Cenomanian, and Simoliophis ( three species:
S. rochebrunei, S. libycus and an unnamed species) from the
Early, Middle and perhaps Late, Cenomanian.
It is incontestable that three snakes have hind limbs:
Pachyrhachis problematicus and Haasiophis terrasanctus of the
earliest Cenomanian, along with Eupodophis descouensi of the Middle Cenomanian. All three were found in the Middle East. Three other genera may possess such limbs: Pachyophis (unique species: P. woodwardi),
Mesophis (unique species: M. nopcsai), both from the Middle, or more probably the
Late Cenomanian, and Simoliophis ( three species:
S. rochebrunei, S. libycus and an unnamed species) from the
Early, Middle and perhaps Late, Cenomanian.
![]() It is not impossible that either Pachyrhachis
or Haasiophis is a synonym of Simoliophis, for the latter is known only from vertebrae and isolated ribs. The dorsal tubercles on the vertebrae of
Eupodophis make it impossible for that genus to be a synonym of Simoliophis.
It is not impossible that either Pachyrhachis
or Haasiophis is a synonym of Simoliophis, for the latter is known only from vertebrae and isolated ribs. The dorsal tubercles on the vertebrae of
Eupodophis make it impossible for that genus to be a synonym of Simoliophis.
![]()
![]() It is difficult here not to touch on the question of the relationships of "snakes with legs" even though their phylogenetic aspects are not the main topics of this article. We shall discuss only the three species that are unquestionably hindlimbed.
It is difficult here not to touch on the question of the relationships of "snakes with legs" even though their phylogenetic aspects are not the main topics of this article. We shall discuss only the three species that are unquestionably hindlimbed.
![]() The phylogenetic position of the hindlimbed taxa in snake classification has led to much debate. Two main facts must be kept in mind:
The phylogenetic position of the hindlimbed taxa in snake classification has led to much debate. Two main facts must be kept in mind:
![]() 1) these three taxa have hind legs, small but perfectly formed; logically, in snakes this is a plesiomorphic characteristic;
1) these three taxa have hind legs, small but perfectly formed; logically, in snakes this is a plesiomorphic characteristic;
![]() 2) the three hindlimbed snakes have a macrostomate skull;
2) the three hindlimbed snakes have a macrostomate skull;
but in existing snakes this character appears only in forms considered to be the most "advanced", the Macrostomata;
a priori, this structure should be derived.
![]() The name macrostomate is given to a structural plan that allows the buccal opening to enlarge. This enlargement is due mainly to a posterior elongation of the supratemporal bone that shifts the quadrate posteriorly, and the dorsoventral elongation of the quadrate. A macrostomate structure permits the ingestion of very large prey, sometimes greater in diameter than the predatory snake itself.
The name macrostomate is given to a structural plan that allows the buccal opening to enlarge. This enlargement is due mainly to a posterior elongation of the supratemporal bone that shifts the quadrate posteriorly, and the dorsoventral elongation of the quadrate. A macrostomate structure permits the ingestion of very large prey, sometimes greater in diameter than the predatory snake itself.
![]() The association of these two characters, the presence of hind legs and a macrostomate structure, poses a serious problem.
The association of these two characters, the presence of hind legs and a macrostomate structure, poses a serious problem.
![]() Several phyletic analyses have led to diametrically opposite results: snakes with legs may be either
Several phyletic analyses have led to diametrically opposite results: snakes with legs may be either
![]() 1) primitive snakes, sister group to all the other snakes
(Caldwell & Lee,
1997; Lee & Caldwell,
1998; Rage & Escuillié,
2000; Scanlon & Lee,
2000; Lee & Scanlon, 2002), or
1) primitive snakes, sister group to all the other snakes
(Caldwell & Lee,
1997; Lee & Caldwell,
1998; Rage & Escuillié,
2000; Scanlon & Lee,
2000; Lee & Scanlon, 2002), or
![]() 2) "advanced" snakes (in spite of the presence of legs), sister group to modern Macrostomata
(Zaher & Rieppel,
1999; Tchernov et alii, 2000; Rieppel & Zaher,
2000; Zaher & Rieppel, 2000).
2) "advanced" snakes (in spite of the presence of legs), sister group to modern Macrostomata
(Zaher & Rieppel,
1999; Tchernov et alii, 2000; Rieppel & Zaher,
2000; Zaher & Rieppel, 2000).
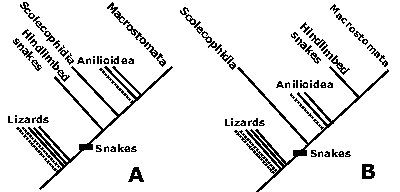
![]() The first hypothesis (hindlimbed snakes are "primitive";
Fig.
5A
The first hypothesis (hindlimbed snakes are "primitive";
Fig.
5A
![]() )
proposes either that the macrostomate structure appeared twice by convergence (in hindlimbed snakes first and in the Macrostomata again) or, more probably, that the macrostomate structure is plesiomorphic (i.e. primitive) among snakes (contrary to what was believed) and that in non-macrostomate snakes it was lost because of the adoption of a burrowing habitat
(Rage & Escuillié,
2000); in fact, all non-macrostomate snakes are burrowers, a mode of life that entails a reduction in the size of the mouth in vertebrates.
)
proposes either that the macrostomate structure appeared twice by convergence (in hindlimbed snakes first and in the Macrostomata again) or, more probably, that the macrostomate structure is plesiomorphic (i.e. primitive) among snakes (contrary to what was believed) and that in non-macrostomate snakes it was lost because of the adoption of a burrowing habitat
(Rage & Escuillié,
2000); in fact, all non-macrostomate snakes are burrowers, a mode of life that entails a reduction in the size of the mouth in vertebrates.
![]() The second hypothesis (hindlimbed snakes are "advanced";
Fig.
5B
The second hypothesis (hindlimbed snakes are "advanced";
Fig.
5B
![]() )
leads to the supposition either that several groups of snakes (five?) independently lost their hind legs or that hind legs
reappeared in hindlimbed snakes. Although theoretically not entirely impossible, this last possibility seems most unlikely.
)
leads to the supposition either that several groups of snakes (five?) independently lost their hind legs or that hind legs
reappeared in hindlimbed snakes. Although theoretically not entirely impossible, this last possibility seems most unlikely.
![]() These divergences of opinion concerning the phyletic position of hindlimbed fossils are not without influence on our conceptions of the origin of snakes. It being understood that the lizards are the stem group from which snakes evolved, it was thought for a long time that snakes evolved after a burrowing or semi-fossorial phase
(Rage,
1987). But as hindlimbed snakes are marine, if they are really the sister of or stem group to other taxa, the snakes may have originated in a marine environment. This hypothesis is clearly supported by the fact that the Mosasauroidea, Cretaceous lizards also markedly adapted to a marine life, are widely recognized as the group most closely related to the snakes
(e.g. Lee,
1997; Caldwell, 1999; Lee & Caldwell,
2000; see, however, the divergent views of Rieppel & Zaher,
2000). On the other hand, if hindlimbed snakes are "advanced", which seems most unlikely, the origin of snakes subsequent to a burrowing or sub-burrowing phase remains plausible.
These divergences of opinion concerning the phyletic position of hindlimbed fossils are not without influence on our conceptions of the origin of snakes. It being understood that the lizards are the stem group from which snakes evolved, it was thought for a long time that snakes evolved after a burrowing or semi-fossorial phase
(Rage,
1987). But as hindlimbed snakes are marine, if they are really the sister of or stem group to other taxa, the snakes may have originated in a marine environment. This hypothesis is clearly supported by the fact that the Mosasauroidea, Cretaceous lizards also markedly adapted to a marine life, are widely recognized as the group most closely related to the snakes
(e.g. Lee,
1997; Caldwell, 1999; Lee & Caldwell,
2000; see, however, the divergent views of Rieppel & Zaher,
2000). On the other hand, if hindlimbed snakes are "advanced", which seems most unlikely, the origin of snakes subsequent to a burrowing or sub-burrowing phase remains plausible.
![]() A remaining question is whether the hindlimbed snakes form a clade or are a paraphyletic group. This question, raised by
Rage & Escuillié
(2000), has not yet been answered.
A remaining question is whether the hindlimbed snakes form a clade or are a paraphyletic group. This question, raised by
Rage & Escuillié
(2000), has not yet been answered.
![]()
![]() The fact that all snakes confirmed or inferred to be limbed are of Cenomanian age is striking. It is true, too, that the rare fossils found in older beds (Upper Albian), are all in the form of poorly preserved isolated vertebrae so that it is not possible to determine whether or not they had legs. Whatever may be the case, it must be stated that legged snakes and those supposed to have had them have vertebrae of a very special morphology that distinguishes them from all other snakes. But this type of vertebra has been found only in the Cenomanian. Consequently, one may assume that legged snakes occur only in strata assigned this stage.
The fact that all snakes confirmed or inferred to be limbed are of Cenomanian age is striking. It is true, too, that the rare fossils found in older beds (Upper Albian), are all in the form of poorly preserved isolated vertebrae so that it is not possible to determine whether or not they had legs. Whatever may be the case, it must be stated that legged snakes and those supposed to have had them have vertebrae of a very special morphology that distinguishes them from all other snakes. But this type of vertebra has been found only in the Cenomanian. Consequently, one may assume that legged snakes occur only in strata assigned this stage.
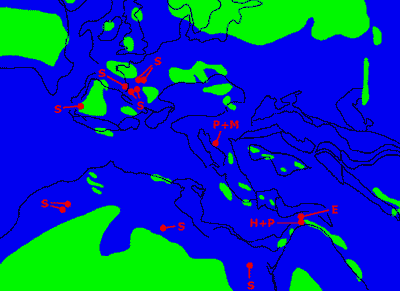
![]() In addition, these snakes have a very restricted geographic distribution. All occur in the "Mediterranean" area of the Tethys or in its immediate vicinity: the north, east and south margins of the existing Mediterranean and its extension as far as the transitional area between the Aquitaine and Paris basins
(Fig.
6
In addition, these snakes have a very restricted geographic distribution. All occur in the "Mediterranean" area of the Tethys or in its immediate vicinity: the north, east and south margins of the existing Mediterranean and its extension as far as the transitional area between the Aquitaine and Paris basins
(Fig.
6
![]() ).
).
![]() If hindlimbed snakes really belong to a basal group (sister group to all other snakes) as we uphold, and as did
Caldwell & Lee
(1997) and Scanlon & Lee (2000), then their range that is very restricted in both time and space deserves attention. As a consequence, snakes would have originated in an aquatic, marine environment
(Nopcsa,
1923; Caldwell, 1999; Rage,
2000), although Lee et alii (1999) did not reject the possibility that legged snakes were only secondarily aquatic. If snakes really originated in a marine environment, the ''Mediterranean'' part of the Tethys might be regarded as the cradle of the whole group. As far as their very limited stratigraphic range (Cenomanian only) is concerned, no explanation can be brought forward, although a taphonomic bias cannot be definitely ruled
out.
If hindlimbed snakes really belong to a basal group (sister group to all other snakes) as we uphold, and as did
Caldwell & Lee
(1997) and Scanlon & Lee (2000), then their range that is very restricted in both time and space deserves attention. As a consequence, snakes would have originated in an aquatic, marine environment
(Nopcsa,
1923; Caldwell, 1999; Rage,
2000), although Lee et alii (1999) did not reject the possibility that legged snakes were only secondarily aquatic. If snakes really originated in a marine environment, the ''Mediterranean'' part of the Tethys might be regarded as the cradle of the whole group. As far as their very limited stratigraphic range (Cenomanian only) is concerned, no explanation can be brought forward, although a taphonomic bias cannot be definitely ruled
out.
![]()
![]() One of the traits that most clearly distinguish the legged snakes is pachyostosis, that is a non-pathologic "thickening" of the bones of a large portion of the vertebral column. Hoffstetter
(1955) presented two hypotheses to explain the advantages to the animal of
pachyostosis:
One of the traits that most clearly distinguish the legged snakes is pachyostosis, that is a non-pathologic "thickening" of the bones of a large portion of the vertebral column. Hoffstetter
(1955) presented two hypotheses to explain the advantages to the animal of
pachyostosis:
![]() a) an increase in the amount of tissues that make erythrocytes, thus an augmentation of respiratory capacity;
a) an increase in the amount of tissues that make erythrocytes, thus an augmentation of respiratory capacity;
![]() b) as ballast to improve the animal's swimming capabilities.
b) as ballast to improve the animal's swimming capabilities.
No one else has followed up on Hoffstetter's hypothesis concerning erythrocytes (which does not mean that it is erroneous), but
Buffrénil & Rage
(1993) and Scanlon et alii (1999) support the idea that pachyostosis played a hydrostatic role. Indeed, the area in which the bones are pachyostotic and could serve as ballast is situated in the region of the lungs; consequently pachyostosis may have helped in buoyancy control. More precisely, according to Scanlon et alii
(1999), thanks to pachyostosis, the mean density of the animal was probably close to that of water and its center of gravity must have been near that of buoyancy. From a
study of Pachyrhachis, they concluded that the animal was not a surface dweller and that pachyostosis probably allowed it to remain at depth.
![]() Buffrénil & Rage
(1993), based on a study of Simoliophis (the existence of legs in some snakes was not known at the time) and
Scanlon et alii
(1999) from Pachyrhachis, reached very similar conclusions concerning the habitat of these snakes. These animals lived in shallow water among reefs, in lagoons, and could not have been deep divers. They were slow, not very mobile, and probably ate slow-moving prey, or cornered faster prey in burrows or crevices.
According to Scanlon et alii
(1999), they were able to hunt through ambush, as do modern sea snakes.
Buffrénil & Rage
(1993), based on a study of Simoliophis (the existence of legs in some snakes was not known at the time) and
Scanlon et alii
(1999) from Pachyrhachis, reached very similar conclusions concerning the habitat of these snakes. These animals lived in shallow water among reefs, in lagoons, and could not have been deep divers. They were slow, not very mobile, and probably ate slow-moving prey, or cornered faster prey in burrows or crevices.
According to Scanlon et alii
(1999), they were able to hunt through ambush, as do modern sea snakes.
![]() The major characteristics of the anatomy of all the legged snakes is so homogenous that the major points regarding their way of life derived from Pachyrhachis and Simoliophis should be applicable to all of them.
The major characteristics of the anatomy of all the legged snakes is so homogenous that the major points regarding their way of life derived from Pachyrhachis and Simoliophis should be applicable to all of them.
![]() Finally and inevitably is the question of the function of the back legs in these snakes. Very much reduced, they could not have really aided locomotion. It has been suggested that they may have served as a way to anchor the body in fissures, or as aids to mating
(Scanlon et alii,
1999). But as they were simply limbs in the process of total regression perhaps they had no function.
Finally and inevitably is the question of the function of the back legs in these snakes. Very much reduced, they could not have really aided locomotion. It has been suggested that they may have served as a way to anchor the body in fissures, or as aids to mating
(Scanlon et alii,
1999). But as they were simply limbs in the process of total regression perhaps they had no function.
![]() The content of this article was presented originally at a colloquium on the Cenomanian organised by B. Ferré,
M. Fouray and J. Tabouelle at Rouen, France (October, 2001). We are indebted to N.J. Sander who translated the text in a hurry. E. Buffetaut,
M.W. Caldwell, and M.S.Y. Lee made helpful suggestions and we also thank B. Granier for much editorial work.
The content of this article was presented originally at a colloquium on the Cenomanian organised by B. Ferré,
M. Fouray and J. Tabouelle at Rouen, France (October, 2001). We are indebted to N.J. Sander who translated the text in a hurry. E. Buffetaut,
M.W. Caldwell, and M.S.Y. Lee made helpful suggestions and we also thank B. Granier for much editorial work.
![]()
Bolkay S.J. (1925).- Mesophis nopcsai n.g. n.sp. ein neues, schlangenähnliches reptil aus der unteren Kreide (Neocom) von Bilek-Selista (Ost-Hercegovina).- Glasnik zemaljskog Muzeja u Bosni i Hercegovini, Sarajevo, t. 37, pp. 125-135.
Buffrénil V. de & Rage J.C. (1993).- La "pachyostose" vertébrale de Simoliophis (Reptilia, Squamata): données comparatives et considérations fonctionnelles.- Annales de Paléontologie, Paris, vol. 79, pp. 315-335.
Caldwell M.W. (1999).- Squamate phylogeny and the relationships of snakes and mosasauroids.- Zoological Journal of the Linnean Society, Oxford, vol. 125, pp. 155-147.
Caldwell M.W. & Lee M.S.Y. (1997).- A snake with legs from the marine Cretaceous of the Middle East.- Nature, London, t. 386, pp. 705-709.
Chalifa Y. & Tchernov E. (1982).- Pachyamia latimaxillaris, new genus and species (Actinopterygii: Amiidae), from the Cenomanian of Jerusalem.- Journal of Vertebrate Paleontology, Los Angeles, vol. 2, pp. 269-285.
Cuny G., Jaeger J.J., Mahboubi M. & Rage J.C. (1990).- Les plus anciens serpents (Reptilia, Squamata) connus. Mise au point sur l'âge géologique des serpents de la partie moyenne du Crétacé.- Comptes rendus des séances de l'Académie des Sciences, Paris, II, t. 311, pp. 1267-1272.
Dominik W. (1985).- Stratigraphie und Sedimentologie (Geochemie, Schwermineralanalyse) der Oberkreide von Bahariya und ihre Korrelation zum Dakhla-Becken (Western-Desert, Ägypten). -Berliner Geowissenschaftliche Abhandlungen, Berlin A, vol. 62, pp. 1-172.
Gardner J.D. & Cifelli R.L. (1999).- A primitive snake from the Cretaceous of Utah.- Special Papers in Palaeontology, London, nº 60, pp. 87-100.
Haas G. (1979).- On a new Snakelike Reptile from the Lower Cenomanian of Ein Jabrud, near Jerusalem.- Bulletin du Muséum national d'Histoire Naturelle de Paris, Paris, t. 1, C, pp. 51-64.
Haas G. (1980).- Remarks on a new ophiomorph reptile from the lower Cenomanian of Ein Jabrud, Israel.- In: Aspects of Vertebrate History (Jacobs L.L., Ed.), Museum of Northern Arizona Press, Flagstaff, pp. 177-192.
Hoffstetter R. (1955).- Squamates de type moderne.- Traité de Paléontologie, 5. Masson, Paris, pp. 606-662.
Hoffstetter R. (1959).- Un serpent terrestre dans le Crétacé du Sahara.- Bulletin de la Société géologique de France, Paris, (7), t. I, pp. 897-902.
Jonet S. (1981).- Contribution à l'étude des vertébrés du Crétacé portugais et spécialement du Cénomanien de l'Estremadure.- Comunicações dos Serviços Geológicos de Portugal, Lisboa, t. 67, pp. 191-300.
Lapparent de Broin F. de (2000).- The oldest pre-podocnemidid turtle (Chelonii, Pleurodira), from the early Cretaceous, Ceará State, Brasil, and its environment.- Treballs del Museu de Geologia de Barcelona, Barcelona, nº 9, pp. 43-95.
Lee M.S.Y. (1997).- The phylogeny of varanoid lizards and the affinities of snakes. -Philosophical Transactions of the Royal Society of London, London, B, vol. 352, pp. 53-91.
Lee M.S.Y. & Caldwell M.W. (1998).- Anatomy and relationships of Pachyrhachis problematicus, a primitive snake with hindlimbs.- Philosophical Transactions of the Royal Society of London, London, B, vol. 353, pp. 1521-1552.
Lee M.S.Y., Caldwell M.W. & Scanlon J.D. (1999).- A second primitive marine snake: Pachyophis woodwardi from the Cretaceous of Bosnia-Herzegovina.- Journal of Zoology, London, vol. 248, pp. 509-520.
Lee M.S.Y. & Caldwell M.W. (2000).- Adriosaurus and the affinities of mosasaurs, dolichosaurs, and snakes.- Journal of Paleontology, Lawrence, vol. 74, pp. 915-937.
Lee M.S.Y. & Scanlon J.D. (2002).- Snake phylogeny based on osteology, soft anatomy and ecology.- Biological Review, Cambridge, vol. 77, pp. 333-401.
McDowell S.B. (1987).- Systematics.- In: Snakes. Ecology, and evolutionary Biology (Seigel R.A., Collins J.T. & Novak S.S., Eds.), McMillan, New York, pp. 3-50.
Nessov L.A., Zhegallo V.I. & Averianov A.O. (1998).- A new locality of Late Cretaceous snakes, mammals and other vertebrates in Africa (western Libya).- Annales de Paléontologie, Paris, vol. 84, pp. 265-274.
Nopcsa F. (1923).- Vorläufige Notiz über die Pachyostose und Osteosklerose einiger mariner Wirbeltiere.- Anatomischer Anzeiger, Jena, t. 56, pp. 353-359.
Nopcsa F. (1925).- Die Symoliophis-Reste.-In: Ergbenisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens, II.- Abhandlungen der Bayerischen Akademie der Wissenschaften, Mathematisch-naturwissenschaftliche Abteilung, München, vol. 30, pp. 1-27.
Nopcsa F. (1931).- Osteologia Reptilium fossilium et recentium. II: Appendix.- Fossilium Catalogus I. Animalia, Berlin, Pars 50, pp. 1-62.
Rage J.C. (1984).- Serpentes.- In: Handbuch der Paläoherpetologie (Wellnhofer P., Ed.), part 11. Gustav Fischer Verlag, Stuttgart/New York, xii + 80 p.
Rage J.C. (1987).- Fossil History.- In: Snakes. Ecology, and evolutionary Biology (Seigel R.A., Collins J.T. & Novak S.S., Eds.), McMillan, New York, pp. 51-76.
Rage J.C. (1988).- Un serpent primitif (Reptilia, Serpentes) dans le Cénomanien (base du Crétacé supérieur).- Comptes rendus des séances de l'Académie des Sciences, Paris, II, t. 307, pp. 1027-1032.
Rage J.C. (2000).- Phylogénie et origine des serpents.- Bulletin de la Société herpétologique de France, Paris, nº 96, pp. 57-69.
Rage J.C. & Cappetta H. (2002).- Vertebrates from the Cenomanian, and the geological age of the Draa Ubari fauna (Libya).- Annales de Paléontologie, Paris, vol. 88, pp. 79-84.
Rage J.C. & Escuillié F. (2000).- Un nouveau serpent bipède du Cénomanien (Crétacé). Implications phylétiques.- Comptes rendus des séances de l'Académie des Sciences, Paris, IIa, t. 330, pp. 513-520.
Rage J.C. & Escuillié F. (2002).- Eupodophis, new name for the genus Podophis Rage and Escuillié, 2000, an extinct bipedal snake, preoccupied by Podophis Wiegmann, 1843 (Lacertilia, Scincidae).- Amphibia-Reptilia, Leyden, vol. 23, pp. 232-233.
Rage J.C. & Richter A. (1994).- A snake from the Lower Cretaceous (Barremian) of Spain: The oldest known snake.- Neues Jarbuch für Geologie und Paläontologie, Monatshefte, Stuttgart, fasc. 9, pp. 561-565.
Rieppel O. & Zaher H. (2000).- The intramandibular joint in squamates, and the phylogenetic relationships of the fossil snake Pachyrhachis problematicus Haas.- Fieldiana, Chicago, nº 3, pp. 1-69.
Sauvage H.E. (1880).- Sur l'existence d'un reptile du type ophidien dans les couches à Ostrea columba des Charentes.- Comptes rendus hebdomadaires des séances de l'Académie des Sciences, Paris, t. 91, pp. 671-672.
Sauvage H.E. (1898).- Vertébrés fossiles du Portugal. Contribution à l'étude des poissons et des reptiles du Jurassique et du Crétacique.- Memorias Commissão do Serviço Geológico de Portugal, Lisboa, 1897-1898, pp. 1-46.
Scanlon J.D., Lee M.S.Y., Caldwell M.W. & Shine R. (1999).- The palaeocology of the primitive snake Pachyrhachis.- Historical Biology, Chur, vol. 13, pp. 127-152.
Scanlon J.D. & Lee M.S.Y. (2000).- The Pleistocene snake Wonambi and the early evolution of snakes.- Nature, London, vol. 403, pp. 416-420.
Sereno P.C., Dutheil D.B., Iarochene M., Larsson H.C.E., Lyon G.H., Magwene P.M., Sidor C.A., Varrichio D.J. & Wilson J.A. (1996).- Predatory dinosaurs from the Sahara and late Cretaceous faunal differentiation.- Science, Washington, vol. 272, pp. 986-991.
Sliskovic T. (1970).- Die stratigraphische Lage der Schichten mit Pachyophiidae aus Seliste bei Bileca (Ostherzegowina).- Bulletin scientifique, Conseil des Académies des Sciences et des Arts de la RSF de Yougoslavie, Zagreb, t. 15, pp. 389-390.
Tchernov E., Rieppel O., Zaher H., Polcyn M.J. & Jacobs L.L. (2000).- A fossil snake with limbs.- Science, Washington, vol. 287, pp. 2010-2012.
Voigt S. (1996).- Paläobiogeographie oberkretazicher Inoceramen und Rudisten - Ozeanographische und Klimatologische Konsequenzen einer neuen Paläogeographie.- Münchner Geowissenschaftliche Abhandlungen, München, A, vol. 31, pp. 1-101.
Wallach V. (1984).- A new name for Ophiomorphus colberti Haas, 1980.- Journal of Herpetology, Riverside, vol. 18, p. 239.
Werner C. (1989).- Die Elasmobranchier-Fauna des Gebel Dist Member der Bahariya Formation (OberCenoman) der Oase Bahariya, Ägypten.- Paleo Ichthyologica, München, vol. 5, pp. 1-112.
Zaher H. & Rieppel O. (1999).- The phylogenetic relationships of Pachyrhachis problematicus, and the evolution of limblessness in snakes (Lepidosauria, Squamata).- Comptes rendus des séances de l'Académie des Sciences, Paris, IIa, t. 329, pp. 831-837.
Zaher H. & Rieppel O. (2000).- A brief history of snakes.- Herpetological Review, Saint-Louis, vol. 31, pp. 73-76.
![]()
Click on thumbnail to enlarge the image.
Figure 1: Stratigraphic distribution of the oldest snakes (*unquestionable hindlimbed snakes; **probable hindlimbed snakes). Note: with the exception of Simoliophis, each taxon comes from only one locality; so each bar indicates the maximum possible range.

Click on thumbnail to enlarge the image.
Figure 2: Eupodophis descouensi. Al Nammoura, Lebanon, Cenomanian. Ventral view. The animal has been broken and the posterior portion is fossilized near the head. The total length is about 85 cm.

Click on thumbnail to enlarge the image.
Figure 3: Eupodophis descouensi. Al Nammoura, Lebanon, Cenomanian. Sacral and caudal regions and the back leg; the leg includes the femur, the tibia and the fibula, plus some more distal bones.
Click on thumbnail to enlarge the image.
Figure 4: Reconstruction of Eupodophis descouensi (note the size and position of the limbs).
Click on thumbnail to enlarge the image.
Figure 5: Relationships of the hindlimbed snakes. They may be "primitive" (A) or "advanced" (B).
Click on thumbnail to enlarge the image.
Figure 6: Geographic distribution of hindlimbed and presumed hindlimbed snakes. Paleogeography of the Mediterranean region during the Cenomanian (from Voigt, 1996, simplified). E: Eupodophis; H+P: Haasiophis and Pachyrhachis; P+M: Pachyophis and Mesophis; S: Simoliophis (because of their proximity, all the localities with Simoliophis in France, Portugal and Morocco are not shown).
![]()