![]()
![]()
![]() Trois serpents munis de pattes sont connus : Pachyrhachis problematicus, Haasiophis terrasanctus et Eupodophis descouensi. Ils possèdent des membres et une ceinture postérieurs mais n'ont ni ceinture ni membres antérieurs, c'est-à-dire qu'ils sont bipèdes. De plus, Pachyophis woodwardi, Mesophis nopcsai et Simoliophis ssp. semblent être étroitement apparentés aux serpents bipèdes ; par conséquent, bien que des membres postérieurs n'aient pas été mis en évidence chez
eux, il est supposé qu'ils étaient aussi bipèdes. Tous les serpents bipèdes et ceux qui sont supposés l'avoir été ont été trouvés dans le Cénomanien seulement. De plus, ces six genres proviennent d'une aire géographique réduite (de l'Europe occidentale et Afrique du nord-ouest au Moyen-Orient). Cette répartition géographique limitée suggère que les serpents sont nés dans la partie "méditerranéenne" de la Téthys, mais la distribution stratigraphique restreinte reste inexpliquée.
Trois serpents munis de pattes sont connus : Pachyrhachis problematicus, Haasiophis terrasanctus et Eupodophis descouensi. Ils possèdent des membres et une ceinture postérieurs mais n'ont ni ceinture ni membres antérieurs, c'est-à-dire qu'ils sont bipèdes. De plus, Pachyophis woodwardi, Mesophis nopcsai et Simoliophis ssp. semblent être étroitement apparentés aux serpents bipèdes ; par conséquent, bien que des membres postérieurs n'aient pas été mis en évidence chez
eux, il est supposé qu'ils étaient aussi bipèdes. Tous les serpents bipèdes et ceux qui sont supposés l'avoir été ont été trouvés dans le Cénomanien seulement. De plus, ces six genres proviennent d'une aire géographique réduite (de l'Europe occidentale et Afrique du nord-ouest au Moyen-Orient). Cette répartition géographique limitée suggère que les serpents sont nés dans la partie "méditerranéenne" de la Téthys, mais la distribution stratigraphique restreinte reste inexpliquée.
Cénomanien ; Crétacé ; Paléogéographie ; Reptiles ; Serpents bipèdes.
Rage J.-C., Escuillié F. (2003).- Le Cénomanien : étage des serpents bipèdes.- Carnets de Géologie / Notebooks on Geology, Maintenon, Article 2003/01 (CG2003_A01)
![]() The Cenomanian: stage of hindlimbed snakes.- Three "snakes with legs" are known: Pachyrhachis problematicus, Haasiophis terrasanctus and Eupodophis descouensi. They have short posterior limbs but lack an anterior girdle and forelimbs. Moreover, Pachyophis woodwardi, Mesophis nopcsai and Simoliophis ssp. appear to be closely related to the hindlimbed taxa; consequently, although the presence of posterior limbs has not been demonstrated for these genera, it is presumed that they too were hindlimbed. All these snakes have been recovered only from the Cenomanian. Moreover, these six genera come from a restricted area (western Europe and
northwesternmost Africa to the Middle East). This limited geographic range suggests that snakes might have originated in the "Mediterranean" part of the Tethys, but the restricted stratigraphical range remains unexplained.
The Cenomanian: stage of hindlimbed snakes.- Three "snakes with legs" are known: Pachyrhachis problematicus, Haasiophis terrasanctus and Eupodophis descouensi. They have short posterior limbs but lack an anterior girdle and forelimbs. Moreover, Pachyophis woodwardi, Mesophis nopcsai and Simoliophis ssp. appear to be closely related to the hindlimbed taxa; consequently, although the presence of posterior limbs has not been demonstrated for these genera, it is presumed that they too were hindlimbed. All these snakes have been recovered only from the Cenomanian. Moreover, these six genera come from a restricted area (western Europe and
northwesternmost Africa to the Middle East). This limited geographic range suggests that snakes might have originated in the "Mediterranean" part of the Tethys, but the restricted stratigraphical range remains unexplained.
![]() Cenomanian ; Cretaceous ; Pachyostosis ; Paleogeography ; Reptilia ; Hindlimbed Snakes.
Cenomanian ; Cretaceous ; Pachyostosis ; Paleogeography ; Reptilia ; Hindlimbed Snakes.
![]()
![]() Il est établi depuis longtemps que les serpents sont des tétrapodes. D'ailleurs, les représentants de la plupart des familles actuelles possèdent des vestiges de ceinture pelvienne et de membres postérieurs. Toutefois, chez les actuels, la ceinture pelvienne a perdu tout contact osseux avec la colonne vertébrale et le membre, lorsqu'il existe, consiste en un seul os très réduit, logiquement considéré comme le fémur. Ainsi, les Leptotyphlopidae, plusieurs anilioïdes (Anilius, Cylindrophis, Anomochilus), le booïde Loxocemus et les Boidae possèdent des vestiges de bassin et de fémurs. Ces éléments sont aussi présents chez les Tropidophiidae s.l., mais seulement chez les mâles. Quant aux Typhlopidae, ils conservent un bassin mais ont perdu les fémurs. Notons, qu'il n'existe aucune trace de membres et de ceinture antérieurs chez les serpents, qu'ils soient actuels ou
fossiles.
Il est établi depuis longtemps que les serpents sont des tétrapodes. D'ailleurs, les représentants de la plupart des familles actuelles possèdent des vestiges de ceinture pelvienne et de membres postérieurs. Toutefois, chez les actuels, la ceinture pelvienne a perdu tout contact osseux avec la colonne vertébrale et le membre, lorsqu'il existe, consiste en un seul os très réduit, logiquement considéré comme le fémur. Ainsi, les Leptotyphlopidae, plusieurs anilioïdes (Anilius, Cylindrophis, Anomochilus), le booïde Loxocemus et les Boidae possèdent des vestiges de bassin et de fémurs. Ces éléments sont aussi présents chez les Tropidophiidae s.l., mais seulement chez les mâles. Quant aux Typhlopidae, ils conservent un bassin mais ont perdu les fémurs. Notons, qu'il n'existe aucune trace de membres et de ceinture antérieurs chez les serpents, qu'ils soient actuels ou
fossiles.
![]() On pouvait s'attendre à trouver des serpents fossiles munis de membres, mais ce n'est que récemment que de tels fossiles ont été reconnus ou découverts. Trois taxons sont d'incontestables serpents à membres postérieurs, donc bipèdes ; connus par des squelettes en connexion assez complets, ils correspondent à trois genres distincts. Ces serpents sont très semblables ; ils possèdent tous une ceinture pelvienne et des membres postérieurs courts mais bien formés. Trois autres genres chez qui on n'a pas observé de membres sont maintenant soupçonnés d'avoir possédé des pattes postérieures. Curieusement, tous ont été trouvés dans le Cénomanien. Ils comptent parmi les plus anciens serpents, mais quelques restes difficiles à étudier leur sont antérieurs (voir ci-dessous et
Fig. 1
On pouvait s'attendre à trouver des serpents fossiles munis de membres, mais ce n'est que récemment que de tels fossiles ont été reconnus ou découverts. Trois taxons sont d'incontestables serpents à membres postérieurs, donc bipèdes ; connus par des squelettes en connexion assez complets, ils correspondent à trois genres distincts. Ces serpents sont très semblables ; ils possèdent tous une ceinture pelvienne et des membres postérieurs courts mais bien formés. Trois autres genres chez qui on n'a pas observé de membres sont maintenant soupçonnés d'avoir possédé des pattes postérieures. Curieusement, tous ont été trouvés dans le Cénomanien. Ils comptent parmi les plus anciens serpents, mais quelques restes difficiles à étudier leur sont antérieurs (voir ci-dessous et
Fig. 1
![]() ).
).
![]()
![]() Les plus anciens serpents viennent de la partie moyenne du Crétacé. Un serpent avait été signalé, mais non nommé, dans le Barrémien d'Espagne (Rage & Richter,
1994). Il avait été identifié sur la base de vertèbres isolées. Depuis, l'étude de lézards du Crétacé a montré que les caractères qui étaient supposés démontrer l'appartenance du fossile espagnol aux serpents existent aussi chez des lézards (présence d'un toit zygosphénien, lequel est cependant indenté ; bords subcentraux joignant la partie ventrale des paradiapophyses). Il convient donc de ne plus considérer le fossile du Barrémien comme un serpent.
Les plus anciens serpents viennent de la partie moyenne du Crétacé. Un serpent avait été signalé, mais non nommé, dans le Barrémien d'Espagne (Rage & Richter,
1994). Il avait été identifié sur la base de vertèbres isolées. Depuis, l'étude de lézards du Crétacé a montré que les caractères qui étaient supposés démontrer l'appartenance du fossile espagnol aux serpents existent aussi chez des lézards (présence d'un toit zygosphénien, lequel est cependant indenté ; bords subcentraux joignant la partie ventrale des paradiapophyses). Il convient donc de ne plus considérer le fossile du Barrémien comme un serpent.
![]() Les plus anciens serpents incontestables viennent de l'Albien supérieur d'Algérie (niveau C de la série de Tiout). Ils sont représentés uniquement par des vertèbres mal conservées (Cuny et alii,
1990), mais, leur appartenance aux serpents est incontestable. Il s'agit d'un possible Lapparentophiidae et d'un serpent incertae sedis
(Fig. 1
Les plus anciens serpents incontestables viennent de l'Albien supérieur d'Algérie (niveau C de la série de Tiout). Ils sont représentés uniquement par des vertèbres mal conservées (Cuny et alii,
1990), mais, leur appartenance aux serpents est incontestable. Il s'agit d'un possible Lapparentophiidae et d'un serpent incertae sedis
(Fig. 1
![]() ). Le premier était certainement terrestre mais le second montre des adaptations possibles à la vie aquatique. Lapparentophis defrennei (Lapparentophiidae), serpent terrestre lui aussi trouvé en Algérie (Hoffstetter,
1959), provient de niveaux (série d'In Akhamil) qui pourraient correspondre à l'Albien supérieur ou au Cénomanien (Cuny et alii,
1990). En Amérique du Nord (USA), Coniophis sp. a été signalé dans l'Albien supérieur ou Cénomanien inférieur (Gardner & Cifelli,
1999) ; ce serpent terrestre, sans doute fouisseur, est attribué aux Aniliidae s.l., famille probablement paraphylétique (au moins quand les fossiles sont inclus) qui comprend des formes généralement très primitives. Ces serpents ont été trouvés sous forme de vertèbres isolées et il n'est pas possible d'établir s'il possédaient des
membres.
). Le premier était certainement terrestre mais le second montre des adaptations possibles à la vie aquatique. Lapparentophis defrennei (Lapparentophiidae), serpent terrestre lui aussi trouvé en Algérie (Hoffstetter,
1959), provient de niveaux (série d'In Akhamil) qui pourraient correspondre à l'Albien supérieur ou au Cénomanien (Cuny et alii,
1990). En Amérique du Nord (USA), Coniophis sp. a été signalé dans l'Albien supérieur ou Cénomanien inférieur (Gardner & Cifelli,
1999) ; ce serpent terrestre, sans doute fouisseur, est attribué aux Aniliidae s.l., famille probablement paraphylétique (au moins quand les fossiles sont inclus) qui comprend des formes généralement très primitives. Ces serpents ont été trouvés sous forme de vertèbres isolées et il n'est pas possible d'établir s'il possédaient des
membres.
![]() Le Cénomanien a fourni plusieurs taxons. Un seul d'entre eux était terrestre. Il s'agit de Pouitella, du Cénomanien inférieur français (Rage,
1988) ; représenté par une unique vertèbre, il est impossible de savoir s'il possédait des membres. Les autres étaient hautement adaptés à la vie aquatique et ont tous été trouvés dans des sédiments marins. Pachyrhachis, Eupodophis et Haasiophis sont trois serpents du Cénomanien du Moyen-Orient, munis de membres postérieurs. Pachyophis et Mesophis proviennent du Cénomanien moyen ou plus probablement supérieur de Bosnie. Lee et alii
(1999) ont supposé que Pachyophis avait peut-être des membres postérieurs (voir ci-dessous). Mesophis est très mal connu, l'unique spécimen semble perdu. Enfin, Simoliophis, représenté par des vertèbres et des côtes isolées, est connu dans le Cénomanien inférieur, moyen et peut-être supérieur d'Europe occidentale et d'Afrique du Nord.
Le Cénomanien a fourni plusieurs taxons. Un seul d'entre eux était terrestre. Il s'agit de Pouitella, du Cénomanien inférieur français (Rage,
1988) ; représenté par une unique vertèbre, il est impossible de savoir s'il possédait des membres. Les autres étaient hautement adaptés à la vie aquatique et ont tous été trouvés dans des sédiments marins. Pachyrhachis, Eupodophis et Haasiophis sont trois serpents du Cénomanien du Moyen-Orient, munis de membres postérieurs. Pachyophis et Mesophis proviennent du Cénomanien moyen ou plus probablement supérieur de Bosnie. Lee et alii
(1999) ont supposé que Pachyophis avait peut-être des membres postérieurs (voir ci-dessous). Mesophis est très mal connu, l'unique spécimen semble perdu. Enfin, Simoliophis, représenté par des vertèbres et des côtes isolées, est connu dans le Cénomanien inférieur, moyen et peut-être supérieur d'Europe occidentale et d'Afrique du Nord.
![]()
Pachyrhachis problematicus Haas, 1979
![]() Haas, en
1979, a décrit un fossile sous le nom de Pachyrhachis problematicus. Ce fossile, trouvé chez un carrier, provenait de l'exploitation de calcaire cénomanien de Ein Jabrud près de Ramallah, en Palestine. L'âge cénomanien basal (Chalifa & Tchernov,
1982) ne semble pas discuté. Le fossile comprend le crâne et une partie importante de la colonne vertébrale, mais pas la partie postérieure. Le crâne prouvait indiscutablement qu'il s'agit d'un Squamate et la colonne vertébrale montrait que l'animal était serpentiforme. Mais Haas, tout en montrant que le crâne présente des caractères de serpent, a interprété ce fossile comme un lézard varanoïde (platynote). Il avait surtout relevé des caractères primitifs du crâne (lesquels suggéraient évidemment des affinités avec les lézards) et la pachyostose d'une partie des vertèbres et des côtes. Cette pachyostose évoquait Simoliophis, fossile du Cénomanien connu uniquement par des vertèbres et des côtes isolées, généralement considéré comme un serpent. Sur la base de la morphologie vertébrale, Haas plaçait Pachyrhachis dans la même famille que Simoliophis (Simoliophiidae) et considérait ces deux genres comme des lézards varanoïdes serpentiformes.
Haas, en
1979, a décrit un fossile sous le nom de Pachyrhachis problematicus. Ce fossile, trouvé chez un carrier, provenait de l'exploitation de calcaire cénomanien de Ein Jabrud près de Ramallah, en Palestine. L'âge cénomanien basal (Chalifa & Tchernov,
1982) ne semble pas discuté. Le fossile comprend le crâne et une partie importante de la colonne vertébrale, mais pas la partie postérieure. Le crâne prouvait indiscutablement qu'il s'agit d'un Squamate et la colonne vertébrale montrait que l'animal était serpentiforme. Mais Haas, tout en montrant que le crâne présente des caractères de serpent, a interprété ce fossile comme un lézard varanoïde (platynote). Il avait surtout relevé des caractères primitifs du crâne (lesquels suggéraient évidemment des affinités avec les lézards) et la pachyostose d'une partie des vertèbres et des côtes. Cette pachyostose évoquait Simoliophis, fossile du Cénomanien connu uniquement par des vertèbres et des côtes isolées, généralement considéré comme un serpent. Sur la base de la morphologie vertébrale, Haas plaçait Pachyrhachis dans la même famille que Simoliophis (Simoliophiidae) et considérait ces deux genres comme des lézards varanoïdes serpentiformes.
![]() Un caractère du crâne particulièrement frappant, la structure macrostomate (voir ci-dessous : Aspects phylétiques), était partiellement relevé par Haas mais il n'en avait pas tenu compte pour établir les affinités de Pachyrhachis.
Un caractère du crâne particulièrement frappant, la structure macrostomate (voir ci-dessous : Aspects phylétiques), était partiellement relevé par Haas mais il n'en avait pas tenu compte pour établir les affinités de Pachyrhachis.
![]() En 1980, Haas a décrit, sous le nom de Ophiomorphus colberti, un second squamate serpentiforme provenant du même gisement et acquis dans les mêmes conditions que Pachyrhachis. Le fossile consiste en une colonne vertébrale (dont il manque une partie de la région antérieure et la partie postérieure de la queue), un bassin et de courts membres postérieurs. En outre, un crâne mal préservé, apparaît sous la colonne vertébrale. Les vertèbres et les côtes sont pachyostotiques, mais moins que celles de Pachyrhachis. Haas a interprété ce fossile comme un intermédiaire entre les lézards varanoïdes et les serpents.
En 1980, Haas a décrit, sous le nom de Ophiomorphus colberti, un second squamate serpentiforme provenant du même gisement et acquis dans les mêmes conditions que Pachyrhachis. Le fossile consiste en une colonne vertébrale (dont il manque une partie de la région antérieure et la partie postérieure de la queue), un bassin et de courts membres postérieurs. En outre, un crâne mal préservé, apparaît sous la colonne vertébrale. Les vertèbres et les côtes sont pachyostotiques, mais moins que celles de Pachyrhachis. Haas a interprété ce fossile comme un intermédiaire entre les lézards varanoïdes et les serpents.
![]() Wallach
(1984) a ensuite proposé le nom de genre Estesius pour remplacer Ophiomorphus, ce dernier étant
préoccupé.
Wallach
(1984) a ensuite proposé le nom de genre Estesius pour remplacer Ophiomorphus, ce dernier étant
préoccupé.
![]() McDowell
(1987), le premier a clairement considéré Pachyrhachis comme un serpent ; en revanche il rejetait Ophiomorphus (c'est-à-dire Estesius) de ce
groupe.
McDowell
(1987), le premier a clairement considéré Pachyrhachis comme un serpent ; en revanche il rejetait Ophiomorphus (c'est-à-dire Estesius) de ce
groupe.
![]() En 1997, Caldwell & Lee ont montré que :
En 1997, Caldwell & Lee ont montré que :
![]() 1) le spécimen décrit comme Pachyrhachis problematicus représente bien un serpent, et
1) le spécimen décrit comme Pachyrhachis problematicus représente bien un serpent, et
![]() 2) que le spécimen correspondant à Estesius colberti appartient à Pachyrhachis problematicus, la pachyostose moins forte résultant d'une différence ontogénique.
2) que le spécimen correspondant à Estesius colberti appartient à Pachyrhachis problematicus, la pachyostose moins forte résultant d'une différence ontogénique.
Ainsi, Pachyrhachis problematicus était un serpent à membres postérieurs ! Un doute pouvait subsister du fait que le crâne, qui montre l'appartenance aux serpents, est sur un spécimen alors que les membres sont sur l'autre spécimen (toutefois, le crâne de ce dernier spécimen est présent mais il est écrasé sous les côtes). Mais par la suite, la découverte de Eupodophis descouensi et de Haasiophis terrasanctus a prouvé que Caldwell & Lee
(1997) avaient eu raison de réunir ces deux spécimens en un seul taxon.
![]() L'anatomie de Pachyrhachis problematicus a été révisée en détail par Lee
& Caldwell
(1998). Ils montrent en particulier que Pachyrhachis était bien macrostomate. Pachyrhachis est le plus grand des serpents bipèdes, mais sa longueur est difficile à estimer (plus de 1,5 m).
L'anatomie de Pachyrhachis problematicus a été révisée en détail par Lee
& Caldwell
(1998). Ils montrent en particulier que Pachyrhachis était bien macrostomate. Pachyrhachis est le plus grand des serpents bipèdes, mais sa longueur est difficile à estimer (plus de 1,5 m).
Eupodophis descouensi (Rage & Escuillié, 2000)
![]() Ce taxon est représenté par un spécimen trouvé dans le Cénomanien moyen de Al Nammoura, au Liban. Le spécimen comprend le crâne, la colonne vertébrale (dont il manque une partie vers le tiers postérieur), le bassin et le membre postérieur (Fig. 2
Ce taxon est représenté par un spécimen trouvé dans le Cénomanien moyen de Al Nammoura, au Liban. Le spécimen comprend le crâne, la colonne vertébrale (dont il manque une partie vers le tiers postérieur), le bassin et le membre postérieur (Fig. 2 ![]() , 3
, 3
![]() ). Le crâne est moins bien conservé que celui de Pachyrhachis mais il confirme la macrostomatie des serpents bipèdes. Une grande partie des vertèbres et des côtes est pachyostotique. L'un des points les plus intéressants est l'excellente conservation de la partie postérieure de la colonne vertébrale qui montre que la queue était étonnamment courte (Fig.
4
). Le crâne est moins bien conservé que celui de Pachyrhachis mais il confirme la macrostomatie des serpents bipèdes. Une grande partie des vertèbres et des côtes est pachyostotique. L'un des points les plus intéressants est l'excellente conservation de la partie postérieure de la colonne vertébrale qui montre que la queue était étonnamment courte (Fig.
4
![]() ) : alors que la longueur totale de l'animal est estimée à 85 cm, le membre se situe à moins de 5 cm seulement de l'extrémité caudale. Le membre mesure environ 2 cm. Dans la queue, les os chevrons sont articulés aux vertèbres et non soudés ; il s'agit d'un caractère primitif important pour l'analyse phylétique.
) : alors que la longueur totale de l'animal est estimée à 85 cm, le membre se situe à moins de 5 cm seulement de l'extrémité caudale. Le membre mesure environ 2 cm. Dans la queue, les os chevrons sont articulés aux vertèbres et non soudés ; il s'agit d'un caractère primitif important pour l'analyse phylétique.
![]() Eupodophis diffère de Pachyrhachis par la forme plus pointue de son crâne, par son carré beaucoup moins étalé antéropostérieurement et par la présence de tubercules dorsaux particuliers sur les vertèbres (Rage & Escuillié,
2000). De plus, le corps semble plus aplati latéralement que chez Pachyrhachis.
Eupodophis diffère de Pachyrhachis par la forme plus pointue de son crâne, par son carré beaucoup moins étalé antéropostérieurement et par la présence de tubercules dorsaux particuliers sur les vertèbres (Rage & Escuillié,
2000). De plus, le corps semble plus aplati latéralement que chez Pachyrhachis.
Haasiophis terrasanctus Tchernov et alii, 2000
![]() Ce taxon est lui aussi représenté par un seul spécimen ; ce dernier, presque complet, est sans doute le mieux conservé des exemplaires de serpents bipèdes. Il provient du même gisement que P. problematicus et est donc daté du Cénomanien basal. Sa taille est voisine de celle de Eupodophis. Sa description publiée est assez succincte et porte essentiellement sur le crâne ; en particulier, la région caudale reste mal connue et il faudra attendre la description complète pour connaître ses caractéristiques. Il était aplati latéralement, mais le degré d'aplatissement n'est pas établi. Par ses caractères principaux, il est très semblable aux deux autres taxons. Il s'en distingue par un coronoïde beaucoup plus réduit et une macrostomatie beaucoup moins
marquée.
Ce taxon est lui aussi représenté par un seul spécimen ; ce dernier, presque complet, est sans doute le mieux conservé des exemplaires de serpents bipèdes. Il provient du même gisement que P. problematicus et est donc daté du Cénomanien basal. Sa taille est voisine de celle de Eupodophis. Sa description publiée est assez succincte et porte essentiellement sur le crâne ; en particulier, la région caudale reste mal connue et il faudra attendre la description complète pour connaître ses caractéristiques. Il était aplati latéralement, mais le degré d'aplatissement n'est pas établi. Par ses caractères principaux, il est très semblable aux deux autres taxons. Il s'en distingue par un coronoïde beaucoup plus réduit et une macrostomatie beaucoup moins
marquée.
![]() En outre, il diffère de Pachyrhachis par son carré beaucoup plus étroit antéropostérieurement et de Eupodophis par l'absence de tubercules dorsaux sur les vertèbres. La position du membre par rapport à l'extrémité caudale n'est pas connue. De plus, la présence d'hémapophyses dans la queue, citée par Tchernov et alii
(2000), pose un problème : s'agit-il d'hémapophyses au sens strict (soudées au centrum) ou de chevrons ? La présence de vraies hémapophyses chez Haasiophis, alors que Eupodophis porte des chevrons articulés, serait
surprenante.
En outre, il diffère de Pachyrhachis par son carré beaucoup plus étroit antéropostérieurement et de Eupodophis par l'absence de tubercules dorsaux sur les vertèbres. La position du membre par rapport à l'extrémité caudale n'est pas connue. De plus, la présence d'hémapophyses dans la queue, citée par Tchernov et alii
(2000), pose un problème : s'agit-il d'hémapophyses au sens strict (soudées au centrum) ou de chevrons ? La présence de vraies hémapophyses chez Haasiophis, alors que Eupodophis porte des chevrons articulés, serait
surprenante.
Pachyophis woodwardi Nopcsa, 1923
![]() Ce serpent est lui aussi représenté par un spécimen unique. Il provient d'une carrière proche de Bileca, en Bosnie-Herzégovine. D'abord considéré comme Crétacé inférieur (Néocomien), il est maintenant daté du Cénomanien moyen ou plus probablement supérieur (Sliskovic,
1970).
Ce serpent est lui aussi représenté par un spécimen unique. Il provient d'une carrière proche de Bileca, en Bosnie-Herzégovine. D'abord considéré comme Crétacé inférieur (Néocomien), il est maintenant daté du Cénomanien moyen ou plus probablement supérieur (Sliskovic,
1970).
![]() Un peu plus petit que Eupodophis, il devait atteindre une soixantaine de centimètres. Le spécimen comprend un dentaire, quelques fragments d'os crâniens et la colonne vertébrale (vertèbres et côtes), mais la partie caudale est masquée sous une autre partie de la colonne (Lee et alii,
1999). Vertèbres et côtes sont fortement pachyostotiques ; les côtes, très épaissies, forment un revêtement presque continu. L'aplatissement latéral du corps devait être comparable à celui de Eupodophis. Il est difficile de comparer Pachyophis aux autres taxons car il n'est connu que par sa face dorsale et ses os crâniens sont très incomplets. Toutefois, il faut remarquer que sa pachyostose est plus forte que chez les autres taxons alors qu'il est plus petit. La différence dans le degré de pachyostose n'est donc pas liée à l'ontogénie et elle représente une réelle caractéristique de ce genre.
Un peu plus petit que Eupodophis, il devait atteindre une soixantaine de centimètres. Le spécimen comprend un dentaire, quelques fragments d'os crâniens et la colonne vertébrale (vertèbres et côtes), mais la partie caudale est masquée sous une autre partie de la colonne (Lee et alii,
1999). Vertèbres et côtes sont fortement pachyostotiques ; les côtes, très épaissies, forment un revêtement presque continu. L'aplatissement latéral du corps devait être comparable à celui de Eupodophis. Il est difficile de comparer Pachyophis aux autres taxons car il n'est connu que par sa face dorsale et ses os crâniens sont très incomplets. Toutefois, il faut remarquer que sa pachyostose est plus forte que chez les autres taxons alors qu'il est plus petit. La différence dans le degré de pachyostose n'est donc pas liée à l'ontogénie et elle représente une réelle caractéristique de ce genre.
![]() Lee et alii
(1999), notant les similitudes existant entre Pachyophis et Pachyrhachis (le seul taxon bipède connu au moment de leur travail), ont conclu que Pachyophis pourrait avoir lui aussi des membres postérieurs. Si un membre est présent, ce qui est fort possible voire probable, il est inévitablement masqué sous la partie moyenne de la colonne vertébrale.
Lee et alii
(1999), notant les similitudes existant entre Pachyophis et Pachyrhachis (le seul taxon bipède connu au moment de leur travail), ont conclu que Pachyophis pourrait avoir lui aussi des membres postérieurs. Si un membre est présent, ce qui est fort possible voire probable, il est inévitablement masqué sous la partie moyenne de la colonne vertébrale.
![]() Lee et alii
(1999) ont considéré que Pachyophis et Pachyrhachis forment un clade et les ont placés dans la même famille, les Pachyophiidae. Sur la base du matériel disponible, il ne semble pas possible de confirmer ce point de vue ; la possibilité de la paraphylie des serpents bipèdes ne peut pas être définitivement écartée (Rage & Escuillié,
2000).
Lee et alii
(1999) ont considéré que Pachyophis et Pachyrhachis forment un clade et les ont placés dans la même famille, les Pachyophiidae. Sur la base du matériel disponible, il ne semble pas possible de confirmer ce point de vue ; la possibilité de la paraphylie des serpents bipèdes ne peut pas être définitivement écartée (Rage & Escuillié,
2000).
Mesophis nopcsai Bolkay, 1925
![]() Ce taxon est représenté par un spécimen presque complet mais dont la partie caudale est mal conservée et peut-être incomplète. Le spécimen est apparemment perdu et ce serpent n'est donc connu que par la description de Bolkay
(1925), laquelle n'est pas suffisante pour établir les relations avec les autres taxons. Il a été trouvé dans le même gisement que Pachyophis. Il est aplati latéralement mais le degré d'aplatissement ne peut pas être évalué. Une partie des vertèbres est pachyostotique alors que les côtes ne sont pas, ou ne sont que faiblement affectées par la pachyostose. Comme il est nettement plus petit que Pachyophis (approximativement la moitié) il est possible que cette différence ne résulte que de l'ontogénie et comme les deux taxons viennent du même gisement on peut supposer, comme l'a fait Nopcsa
(1931 : 30), que Mesophis est un juvénile de Pachyophis.
Ce taxon est représenté par un spécimen presque complet mais dont la partie caudale est mal conservée et peut-être incomplète. Le spécimen est apparemment perdu et ce serpent n'est donc connu que par la description de Bolkay
(1925), laquelle n'est pas suffisante pour établir les relations avec les autres taxons. Il a été trouvé dans le même gisement que Pachyophis. Il est aplati latéralement mais le degré d'aplatissement ne peut pas être évalué. Une partie des vertèbres est pachyostotique alors que les côtes ne sont pas, ou ne sont que faiblement affectées par la pachyostose. Comme il est nettement plus petit que Pachyophis (approximativement la moitié) il est possible que cette différence ne résulte que de l'ontogénie et comme les deux taxons viennent du même gisement on peut supposer, comme l'a fait Nopcsa
(1931 : 30), que Mesophis est un juvénile de Pachyophis.
![]() Quoi qu'il en soit, on ne pourra que conjecturer tant que le spécimen ne sera pas retrouvé. Toujours est-il qu'il pourrait s'agir d'un autre serpent bipède.
Quoi qu'il en soit, on ne pourra que conjecturer tant que le spécimen ne sera pas retrouvé. Toujours est-il qu'il pourrait s'agir d'un autre serpent bipède.
Simoliophis Sauvage, 1880
![]() A la différence des genres discutés auparavant, Simoliophis comprend plusieurs espèces. D'autre part, il n'est représenté que par des spécimens désarticulés, des vertèbres et côtes uniquement.
A la différence des genres discutés auparavant, Simoliophis comprend plusieurs espèces. D'autre part, il n'est représenté que par des spécimens désarticulés, des vertèbres et côtes uniquement.
![]() La pachyostose affecte les vertèbres des régions dorsales moyenne et postérieure ainsi que les côtes provenant sans doute des mêmes régions. La morphologie vertébrale est très semblable à celle de Eupodophis, seul serpent bipède avéré dont une vertèbre désarticulée est connue. Les vertèbres de Simoliophis se distinguent de celles de Eupodophis par l'absence des tubercules dorsaux qui caractérisent l'arc neural de ce dernier genre. En revanche, ce que l'on connaît de Pachyrhachis et Haasiophis ne permet pas de distinguer les vertèbres de ces deux genres de celles de Simoliophis, à moins que les reconstitutions de vertèbres de Pachyrhachis proposées par Lee & Caldwell
(1998) ne soient exactes. Dans ce dernier cas, les vertèbres de Pachyrhachis seraient sensiblement différentes de celles de Eupodophis et Simoliophis. Telles qu'elles ont été reconstituées, les vertèbres de Pachyrhachis diffèrent de celles des deux autres genres par leur centrum nettement moins pachyostotique et plus cylindrique, leurs prézygapophyses plus allongées, leurs paradiapophyses moins saillantes et leurs plus grands cotyle et condyle. Quoi qu'il en soit, les vertèbres de Eupodophis et Simoliophis (ainsi que, probablement, celles de Pachyrhachis et Haasiophis) sont très semblables et elles se distinguent très nettement de celles des serpents non bipèdes. Pour cette raison, on peut supposer que ces genres sont étroitement apparentés, ou au moins au même grade, et donc que Simoliophis possédait lui aussi des membres postérieurs.
La pachyostose affecte les vertèbres des régions dorsales moyenne et postérieure ainsi que les côtes provenant sans doute des mêmes régions. La morphologie vertébrale est très semblable à celle de Eupodophis, seul serpent bipède avéré dont une vertèbre désarticulée est connue. Les vertèbres de Simoliophis se distinguent de celles de Eupodophis par l'absence des tubercules dorsaux qui caractérisent l'arc neural de ce dernier genre. En revanche, ce que l'on connaît de Pachyrhachis et Haasiophis ne permet pas de distinguer les vertèbres de ces deux genres de celles de Simoliophis, à moins que les reconstitutions de vertèbres de Pachyrhachis proposées par Lee & Caldwell
(1998) ne soient exactes. Dans ce dernier cas, les vertèbres de Pachyrhachis seraient sensiblement différentes de celles de Eupodophis et Simoliophis. Telles qu'elles ont été reconstituées, les vertèbres de Pachyrhachis diffèrent de celles des deux autres genres par leur centrum nettement moins pachyostotique et plus cylindrique, leurs prézygapophyses plus allongées, leurs paradiapophyses moins saillantes et leurs plus grands cotyle et condyle. Quoi qu'il en soit, les vertèbres de Eupodophis et Simoliophis (ainsi que, probablement, celles de Pachyrhachis et Haasiophis) sont très semblables et elles se distinguent très nettement de celles des serpents non bipèdes. Pour cette raison, on peut supposer que ces genres sont étroitement apparentés, ou au moins au même grade, et donc que Simoliophis possédait lui aussi des membres postérieurs.
![]() Simoliophis n'a été trouvé que dans le Cénomanien (voir ci-dessous).
Simoliophis n'a été trouvé que dans le Cénomanien (voir ci-dessous).
![]() Deux espèces valides ont été décrites, S. rochebrunei et S. libycus. En outre, un Simoliophis du Cénomanien de Baharija (Égypte), rapporté à S. rochebrunei par Nopcsa
(1925), représente une troisième espèce, non nommée.
Deux espèces valides ont été décrites, S. rochebrunei et S. libycus. En outre, un Simoliophis du Cénomanien de Baharija (Égypte), rapporté à S. rochebrunei par Nopcsa
(1925), représente une troisième espèce, non nommée.
Simoliophis rochebrunei Sauvage, 1880
![]() S. rochebrunei, l'espèce type du genre, a été trouvé dans le Cénomanien du centre-ouest et du sud-ouest de la France ainsi qu'au Portugal, dans la région de Lisbonne (Jonet,
1981 ; Rage, 1984). L'espèce provient d'une douzaine de gisements qui, à l'exception de deux d'entre eux, sont soit Cénomanien inférieur, soit Cénomanien moyen. L'âge précis dans le Cénomanien de Sillac et Basseau, deux gisements charentais aujourd'hui disparus, n'est pas connu. On ne peut donc pas exclure la présence de S. rochebrunei dans le Cénomanien supérieur.
S. rochebrunei, l'espèce type du genre, a été trouvé dans le Cénomanien du centre-ouest et du sud-ouest de la France ainsi qu'au Portugal, dans la région de Lisbonne (Jonet,
1981 ; Rage, 1984). L'espèce provient d'une douzaine de gisements qui, à l'exception de deux d'entre eux, sont soit Cénomanien inférieur, soit Cénomanien moyen. L'âge précis dans le Cénomanien de Sillac et Basseau, deux gisements charentais aujourd'hui disparus, n'est pas connu. On ne peut donc pas exclure la présence de S. rochebrunei dans le Cénomanien supérieur.
Simoliophis libycus Nessov, Zhegallo & Averianov, 1998
![]() Cette espèce a été trouvée en Libye, à Draa Ubari. D'après Nessov et alii
(1998), elle se distingue de S. rochebrunei par son centrum plus étroit, sa neurépine moins haute sur les vertèbres antérieures, la moindre dimension antéropostérieure de la neurépine, et ses paradiapophyses moins développées. L'espèce étant fondée sur un matériel réduit et étant plus petite que S. rochebrunei, il resterait à établir que ces différences ne sont pas de nature ontogénique et/ou liées à la position des vertèbres dans la colonne vertébrale.
Cette espèce a été trouvée en Libye, à Draa Ubari. D'après Nessov et alii
(1998), elle se distingue de S. rochebrunei par son centrum plus étroit, sa neurépine moins haute sur les vertèbres antérieures, la moindre dimension antéropostérieure de la neurépine, et ses paradiapophyses moins développées. L'espèce étant fondée sur un matériel réduit et étant plus petite que S. rochebrunei, il resterait à établir que ces différences ne sont pas de nature ontogénique et/ou liées à la position des vertèbres dans la colonne vertébrale.
![]() Nessov et alii
(1998) avaient considéré l'âge de Draa Ubari comme Santonien-Campanien, mais tous les taxons stratigraphiquement significatifs du gisement sont d'âge Cénomanien, voire plus ancien pour certains (Rage & Cappetta,
2002) ; l'âge précis, dans le Cénomanien, ne peut pas être établi.
Nessov et alii
(1998) avaient considéré l'âge de Draa Ubari comme Santonien-Campanien, mais tous les taxons stratigraphiquement significatifs du gisement sont d'âge Cénomanien, voire plus ancien pour certains (Rage & Cappetta,
2002) ; l'âge précis, dans le Cénomanien, ne peut pas être établi.
Les autres Simoliophis
![]() Nopcsa
(1925) a décrit une série de vertèbres et de côtes du Cénomanien de Baharija (Égypte). Il a attribué ces fossiles à S. rochebrunei, seule espèce alors connue. En fait, en reconstituant la colonne vertébrale, Nopcsa a probablement mélangé des vertèbres de deux serpents. Certaines d'entre elles, des dorsales moyennes et postérieures, appartiennent effectivement à Simoliophis mais des vertèbres considérées comme des dorsales antérieures pourraient appartenir à un serpent distinct (opinion basée sur un matériel non publié). D'autre part, les vertèbres du Simoliophis de Baharija n'appartiennent ni à S. rochebrunei, ni à S. libycus, mais à une espèce non nommée comme le montre la présence d'un sillon très net sur la base du bord antérieur de la neurépine.
Nopcsa
(1925) a décrit une série de vertèbres et de côtes du Cénomanien de Baharija (Égypte). Il a attribué ces fossiles à S. rochebrunei, seule espèce alors connue. En fait, en reconstituant la colonne vertébrale, Nopcsa a probablement mélangé des vertèbres de deux serpents. Certaines d'entre elles, des dorsales moyennes et postérieures, appartiennent effectivement à Simoliophis mais des vertèbres considérées comme des dorsales antérieures pourraient appartenir à un serpent distinct (opinion basée sur un matériel non publié). D'autre part, les vertèbres du Simoliophis de Baharija n'appartiennent ni à S. rochebrunei, ni à S. libycus, mais à une espèce non nommée comme le montre la présence d'un sillon très net sur la base du bord antérieur de la neurépine.
![]() Il n'y a pas d'accord sur l'âge précis de Baharija dans le Cénomanien : Cénomanien inférieur pour certains (Lapparent de Broin,
2000), Cénomanien supérieur pour d'autres (Werner,
1989). Notons toutefois que Dominik (1985), en se fondant sur les ammonites, a suggéré un âge cénomanien supérieur pour une partie au moins de la formation de Baharija.
Il n'y a pas d'accord sur l'âge précis de Baharija dans le Cénomanien : Cénomanien inférieur pour certains (Lapparent de Broin,
2000), Cénomanien supérieur pour d'autres (Werner,
1989). Notons toutefois que Dominik (1985), en se fondant sur les ammonites, a suggéré un âge cénomanien supérieur pour une partie au moins de la formation de Baharija.
![]() Plusieurs gisements des Kem Kem (Maroc) ont aussi fourni des vertèbres de Simoliophis qui rappellent celles de S. libycus (travail en cours). Les couches des Kem Kem correspondent au Cénomanien, mais il n'est pas impossible que les niveaux les plus inférieurs correspondent à l'Albien supérieur. Toutefois, les niveaux qui ont fourni les spécimens de Simoliophis sont tous cénomaniens (Sereno et alii,
1996).
Plusieurs gisements des Kem Kem (Maroc) ont aussi fourni des vertèbres de Simoliophis qui rappellent celles de S. libycus (travail en cours). Les couches des Kem Kem correspondent au Cénomanien, mais il n'est pas impossible que les niveaux les plus inférieurs correspondent à l'Albien supérieur. Toutefois, les niveaux qui ont fourni les spécimens de Simoliophis sont tous cénomaniens (Sereno et alii,
1996).
![]() Trois serpents sont incontestablement munis de membres postérieurs : Pachyrhachis problematicus et Haasiophis terrasanctus du Cénomanien basal, ainsi que Eupodophis descouensi du Cénomanien moyen. Tous trois ont été trouvés au Moyen-Orient. Trois autres genres possédaient peut-être de tels membres : Pachyophis (seule espèce : P. woodwardi), Mesophis (seule espèce : M. nopcsai), tous deux du Cénomanien moyen ou plus probablement supérieur, et Simoliophis (trois espèces : S. rochebrunei, S. libycus et une espèce non nommée) du Cénomanien inférieur, moyen et, peut-être supérieur.
Trois serpents sont incontestablement munis de membres postérieurs : Pachyrhachis problematicus et Haasiophis terrasanctus du Cénomanien basal, ainsi que Eupodophis descouensi du Cénomanien moyen. Tous trois ont été trouvés au Moyen-Orient. Trois autres genres possédaient peut-être de tels membres : Pachyophis (seule espèce : P. woodwardi), Mesophis (seule espèce : M. nopcsai), tous deux du Cénomanien moyen ou plus probablement supérieur, et Simoliophis (trois espèces : S. rochebrunei, S. libycus et une espèce non nommée) du Cénomanien inférieur, moyen et, peut-être supérieur.
![]() Il n'est pas impossible que Pachyrhachis, ou Haasiophis, soit synonyme de Simoliophis, ce dernier n'étant connu que par des vertèbres et des côtes isolées. Eupodophis, dont les vertèbres possèdent des tubercules dorsaux, ne peut pas être synonyme de Simoliophis.
Il n'est pas impossible que Pachyrhachis, ou Haasiophis, soit synonyme de Simoliophis, ce dernier n'étant connu que par des vertèbres et des côtes isolées. Eupodophis, dont les vertèbres possèdent des tubercules dorsaux, ne peut pas être synonyme de Simoliophis.
![]()
![]() Il est difficile de ne pas aborder ici la question des affinités des "serpents à pattes" même si les aspects liés à la phylogénie ne sont pas les plus importants dans l'optique de cet article. Nous ne parlerons ici que des trois serpents bipèdes certains.
Il est difficile de ne pas aborder ici la question des affinités des "serpents à pattes" même si les aspects liés à la phylogénie ne sont pas les plus importants dans l'optique de cet article. Nous ne parlerons ici que des trois serpents bipèdes certains.
![]() La position phylétique des taxons bipèdes au sein des serpents fait l'objet d'une polémique acharnée. Deux faits principaux sont à retenir :
La position phylétique des taxons bipèdes au sein des serpents fait l'objet d'une polémique acharnée. Deux faits principaux sont à retenir :
![]() 1) ces trois taxons possèdent des pattes postérieures, petites mais parfaitement formées ; logiquement, il s'agit d'un caractère primitif chez les serpents ;
1) ces trois taxons possèdent des pattes postérieures, petites mais parfaitement formées ; logiquement, il s'agit d'un caractère primitif chez les serpents ;
![]() 2) les trois serpents bipèdes ont un crâne macrostomate ;
2) les trois serpents bipèdes ont un crâne macrostomate ;
or, chez les serpents actuels, cette caractéristique n'apparaît que chez les formes considérées comme étant les plus évoluées, les Macrostomata ; a priori, cette structure devrait être dérivée.
![]() Le nom de structure macrostomate est donné à un dispositif qui permet l'agrandissement de l'ouverture buccale. Cet agrandissement résulte principalement de l'allongement postérieur du supratemporal, ce qui déplace postérieurement le carré, et de l'allongement dorsoventral de ce dernier os. La structure macrostomate permet l'ingestion de très grosses proies, de taille parfois supérieure au diamètre du serpent prédateur.
Le nom de structure macrostomate est donné à un dispositif qui permet l'agrandissement de l'ouverture buccale. Cet agrandissement résulte principalement de l'allongement postérieur du supratemporal, ce qui déplace postérieurement le carré, et de l'allongement dorsoventral de ce dernier os. La structure macrostomate permet l'ingestion de très grosses proies, de taille parfois supérieure au diamètre du serpent prédateur.
![]() L'association de ces deux caractères, présence de pattes postérieures et structure macrostomate, pose un sérieux problème.
L'association de ces deux caractères, présence de pattes postérieures et structure macrostomate, pose un sérieux problème.
![]() Des analyses phylétiques différentes ont mené à des résultats totalement opposés : les serpents à pattes pourraient être soit
Des analyses phylétiques différentes ont mené à des résultats totalement opposés : les serpents à pattes pourraient être soit
![]() 1) des serpents primitifs, groupe frère de tous les autres serpents (Caldwell & Lee,
1997 ; Lee & Caldwell,
1998 ; Rage & Escuillié,
2000 ; Scanlon & Lee,
2000 ; Lee & Scanlon, 2002), soit
1) des serpents primitifs, groupe frère de tous les autres serpents (Caldwell & Lee,
1997 ; Lee & Caldwell,
1998 ; Rage & Escuillié,
2000 ; Scanlon & Lee,
2000 ; Lee & Scanlon, 2002), soit
![]() 2) des serpents "évolués" (malgré la présence de pattes), groupe frère des Macrostomata modernes (Zaher & Rieppel,
1999 ; Tchernov et alii, 2000 ; Rieppel & Zaher,
2000 ; Zaher & Rieppel, 2000).
2) des serpents "évolués" (malgré la présence de pattes), groupe frère des Macrostomata modernes (Zaher & Rieppel,
1999 ; Tchernov et alii, 2000 ; Rieppel & Zaher,
2000 ; Zaher & Rieppel, 2000).
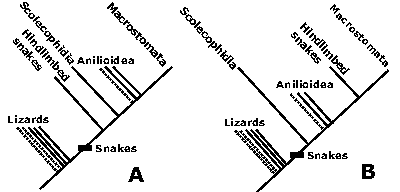
![]() La première hypothèse (les serpents bipèdes sont "primitifs" ;
Fig.
5A
La première hypothèse (les serpents bipèdes sont "primitifs" ;
Fig.
5A
![]() ) suppose soit que la structure macrostomate est apparue deux fois, par convergence (chez les serpents bipèdes d'une part et chez les Macrostomata d'autre part), soit, plus probablement, que la structure macrostomate est primitive chez les serpents (contrairement à ce que l'on croyait) et que chez les serpents non-macrostomates elle a été perdue sous l'influence de la vie fouisseuse (Rage & Escuillié,
2000) ; en effet, tous les serpents non-macrostomates sont fouisseurs, mode de vie qui entraîne une réduction de la taille de la bouche chez les vertébrés.
) suppose soit que la structure macrostomate est apparue deux fois, par convergence (chez les serpents bipèdes d'une part et chez les Macrostomata d'autre part), soit, plus probablement, que la structure macrostomate est primitive chez les serpents (contrairement à ce que l'on croyait) et que chez les serpents non-macrostomates elle a été perdue sous l'influence de la vie fouisseuse (Rage & Escuillié,
2000) ; en effet, tous les serpents non-macrostomates sont fouisseurs, mode de vie qui entraîne une réduction de la taille de la bouche chez les vertébrés.
![]() La seconde hypothèse (les serpents bipèdes sont "évolués" ;
Fig.
5B
La seconde hypothèse (les serpents bipèdes sont "évolués" ;
Fig.
5B
![]() ) conduit à supposer soit que plusieurs groupes de serpents (cinq ?) ont perdu indépendamment leurs pattes postérieures, soit que les pattes postérieures sont réapparues chez les serpents bipèdes. Bien que théoriquement pas absolument impossible, cette dernière possibilité semble très peu probable.
) conduit à supposer soit que plusieurs groupes de serpents (cinq ?) ont perdu indépendamment leurs pattes postérieures, soit que les pattes postérieures sont réapparues chez les serpents bipèdes. Bien que théoriquement pas absolument impossible, cette dernière possibilité semble très peu probable.
![]() Ces divergences à propos de la position phylétique des fossiles bipèdes ne sont pas sans conséquences sur nos conceptions de l'origine des serpents. Etant entendu que les lézards forment le groupe souche des serpents, on a longtemps pensé que ces derniers sont nés à la suite d'une phase fouisseuse ou subfouisseuse (Rage,
1987). Mais les serpents bipèdes étant marins, si ils représentent réellement le groupe frère, ou le groupe souche, des autres serpents l'origine du groupe pourrait s'être située en milieu marin. Cette hypothèse est clairement confortée par le fait que les Mosasauroidea, lézards du Crétacé eux aussi fortement adaptés à la vie marine, sont presque unanimement reconnus comme le groupe le plus étroitement apparenté aux serpents (par exemple, récemment, Lee,
1997 ; Caldwell, 1999 ; Lee & Caldwell,
2000 ; voir toutefois l'opinion divergente de Rieppel & Zaher,
2000). En revanche, si les serpents bipèdes sont des serpents "évolués", ce qui semble peu probable, l'origine des serpents à travers une phase fouisseuse ou subfouisseuse reste plausible.
Ces divergences à propos de la position phylétique des fossiles bipèdes ne sont pas sans conséquences sur nos conceptions de l'origine des serpents. Etant entendu que les lézards forment le groupe souche des serpents, on a longtemps pensé que ces derniers sont nés à la suite d'une phase fouisseuse ou subfouisseuse (Rage,
1987). Mais les serpents bipèdes étant marins, si ils représentent réellement le groupe frère, ou le groupe souche, des autres serpents l'origine du groupe pourrait s'être située en milieu marin. Cette hypothèse est clairement confortée par le fait que les Mosasauroidea, lézards du Crétacé eux aussi fortement adaptés à la vie marine, sont presque unanimement reconnus comme le groupe le plus étroitement apparenté aux serpents (par exemple, récemment, Lee,
1997 ; Caldwell, 1999 ; Lee & Caldwell,
2000 ; voir toutefois l'opinion divergente de Rieppel & Zaher,
2000). En revanche, si les serpents bipèdes sont des serpents "évolués", ce qui semble peu probable, l'origine des serpents à travers une phase fouisseuse ou subfouisseuse reste plausible.
![]() Il resterait à savoir si les serpents bipèdes forment un clade ou un groupe paraphylétique, souche d'un autre groupe ou de tous les autres serpents. Cette question, soulevée par Rage
& Escuillié
(2000), n'a pas encore pu être réglée.
Il resterait à savoir si les serpents bipèdes forment un clade ou un groupe paraphylétique, souche d'un autre groupe ou de tous les autres serpents. Cette question, soulevée par Rage
& Escuillié
(2000), n'a pas encore pu être réglée.
![]()
![]() Il est frappant de constater que tous les serpents à pattes, ou pouvant avoir eu des pattes, ont été trouvés dans le Cénomanien. Il est vrai, aussi, que les rares fossiles trouvés dans des niveaux plus anciens, c'est-à-dire l'Albien supérieur, se présentent sous forme de vertèbres isolées, mal conservées, et que l'on ne peut pas établir s'ils avaient des pattes. Quoi qu'il en soit, il faut remarquer que les serpents à pattes et ceux qui sont supposés en avoir eu possèdent des vertèbres dont la morphologie est très particulière et se distingue de celle de tous les autres serpents. Or ce type de vertèbre n'a été trouvé que dans le Cénomanien. On peut donc penser que les serpents à pattes ne sont présents que dans cet étage.
Il est frappant de constater que tous les serpents à pattes, ou pouvant avoir eu des pattes, ont été trouvés dans le Cénomanien. Il est vrai, aussi, que les rares fossiles trouvés dans des niveaux plus anciens, c'est-à-dire l'Albien supérieur, se présentent sous forme de vertèbres isolées, mal conservées, et que l'on ne peut pas établir s'ils avaient des pattes. Quoi qu'il en soit, il faut remarquer que les serpents à pattes et ceux qui sont supposés en avoir eu possèdent des vertèbres dont la morphologie est très particulière et se distingue de celle de tous les autres serpents. Or ce type de vertèbre n'a été trouvé que dans le Cénomanien. On peut donc penser que les serpents à pattes ne sont présents que dans cet étage.
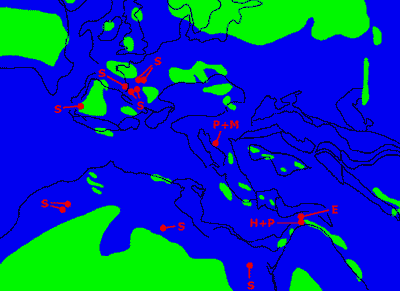
![]() D'autre part, géographiquement, ces serpents montrent aussi une étonnante concentration. Ils proviennent tous de la "partie méditerranéenne" de la Téthys ou de dépendances immédiates : marges nord, est et sud de la Méditerranée actuelle et extension jusqu'à la transition bassin aquitain/bassin parisien
(Fig.
6
D'autre part, géographiquement, ces serpents montrent aussi une étonnante concentration. Ils proviennent tous de la "partie méditerranéenne" de la Téthys ou de dépendances immédiates : marges nord, est et sud de la Méditerranée actuelle et extension jusqu'à la transition bassin aquitain/bassin parisien
(Fig.
6
![]() ).
).
![]() Si les serpents à pattes sont bien primitifs (groupe frère ou groupe souche de tous les autres serpents) comme nous le pensons avec Caldwell
& Lee
(1997) et Scanlon & Lee (2000), cette concentration dans le temps et l'espace mérite un intérêt certain. Les serpents seraient nés en milieu aquatique, marin (Nopcsa,
1923 ; Caldwell, 1999 ; Rage,
2000) bien que Lee et alii (1999) aient admis que les serpents à pattes pourraient éventuellement être devenus marins secondairement. Si les serpents sont effectivement nés en milieu marin, nous pouvons alors nous demander si cette "partie méditerranéenne" de la Téthys n'aurait pas été le berceau des serpents. Quant à leur limitation au Cénomanien, nous ne pouvons avancer aucune explication ; toutefois, un problème taphonomique ne peut pas être définitivement écarté.
Si les serpents à pattes sont bien primitifs (groupe frère ou groupe souche de tous les autres serpents) comme nous le pensons avec Caldwell
& Lee
(1997) et Scanlon & Lee (2000), cette concentration dans le temps et l'espace mérite un intérêt certain. Les serpents seraient nés en milieu aquatique, marin (Nopcsa,
1923 ; Caldwell, 1999 ; Rage,
2000) bien que Lee et alii (1999) aient admis que les serpents à pattes pourraient éventuellement être devenus marins secondairement. Si les serpents sont effectivement nés en milieu marin, nous pouvons alors nous demander si cette "partie méditerranéenne" de la Téthys n'aurait pas été le berceau des serpents. Quant à leur limitation au Cénomanien, nous ne pouvons avancer aucune explication ; toutefois, un problème taphonomique ne peut pas être définitivement écarté.
![]()
![]() L'un des traits les plus marquants des serpents à pattes est la pachyostose, c'est-à-dire l'"épaississement" osseux non pathologique, qui touche une grande partie de la colonne vertébrale. Hoffstetter
(1955) avait avancé deux hypothèses expliquant le, ou les, rôle(s) possible(s) de cette pachyostose :
L'un des traits les plus marquants des serpents à pattes est la pachyostose, c'est-à-dire l'"épaississement" osseux non pathologique, qui touche une grande partie de la colonne vertébrale. Hoffstetter
(1955) avait avancé deux hypothèses expliquant le, ou les, rôle(s) possible(s) de cette pachyostose :
![]() a) augmentation de la masse des tissus formateurs d'érythrocytes, donc de la capacité
respiratoire,
a) augmentation de la masse des tissus formateurs d'érythrocytes, donc de la capacité
respiratoire,
![]() b) rôle de lest corrigeant la position du corps dans
l'eau.
b) rôle de lest corrigeant la position du corps dans
l'eau.
Après Hoffstetter, la possibilité d'un rôle dans la formation des érythrocytes n'a pas été reprise (ce qui ne veut pas dire que cette hypothèse est fausse), en revanche Buffrénil & Rage
(1993) et Scanlon et alii (1999) ont considéré que la pachyostose jouait effectivement un rôle hydrostatique. En effet, la zone pachyostotique apparaît comme un lest situé approximativement dans la région qu'occupaient les poumons ; elle devait intervenir dans le contrôle de la flottaison. Plus exactement, d'après Scanlon et alii
(1999), grâce à la pachyostose, la densité moyenne de l'animal était probablement proche de celle de l'eau et le centre de gravité devait être voisin du centre de flottaison. En étudiant Pachyrhachis, ils concluaient que ce serpent n'était pas un animal de surface et que la pachyostose permettait sans doute le maintien en profondeur.
![]() Buffrénil
& Rage
(1993), se basant sur Simoliophis (l'existence de pattes chez certains serpents n'étant pas connue à l'époque), ainsi que Scanlon et alii
(1999), d'après Pachyrhachis, ont atteint des conclusions très semblables en ce qui concerne le mode de vie de ces serpents. Ces formes vivaient dans des eaux de faible profondeur, parmi des récifs, dans des lagons, et ne devaient pas être de vrais plongeurs. Ils étaient peu rapides, peu mobiles et devaient se nourrir d'animaux fixés ou, eux aussi peu mobiles ; ils pouvaient sans doute aussi bloquer les formes plus rapides dans des anfractuosités. D'après Scanlon et alii
(1999), ils devaient pouvoir chasser en embuscade.
Buffrénil
& Rage
(1993), se basant sur Simoliophis (l'existence de pattes chez certains serpents n'étant pas connue à l'époque), ainsi que Scanlon et alii
(1999), d'après Pachyrhachis, ont atteint des conclusions très semblables en ce qui concerne le mode de vie de ces serpents. Ces formes vivaient dans des eaux de faible profondeur, parmi des récifs, dans des lagons, et ne devaient pas être de vrais plongeurs. Ils étaient peu rapides, peu mobiles et devaient se nourrir d'animaux fixés ou, eux aussi peu mobiles ; ils pouvaient sans doute aussi bloquer les formes plus rapides dans des anfractuosités. D'après Scanlon et alii
(1999), ils devaient pouvoir chasser en embuscade.
![]() Les serpents à pattes présentent une anatomie tellement homogène dans ses grandes caractéristiques que les principaux traits du mode de vie tirés de Pachyrhachis et Simoliophis doivent être valables pour tous.
Les serpents à pattes présentent une anatomie tellement homogène dans ses grandes caractéristiques que les principaux traits du mode de vie tirés de Pachyrhachis et Simoliophis doivent être valables pour tous.
![]() Enfin, inévitablement, se pose la question de la fonction des membres postérieurs chez ces serpents. Trop réduits, ils ne pouvaient certainement pas jouer un rôle réel dans la locomotion. Ont été suggérés un rôle d'ancrage du corps dans les fissures ou leur intervention lors de l'accouplement (Scanlon et alii,
1999). Mais, étant simplement des membres en voie de régression totale, peut-être n'avaient-ils aucune fonction.
Enfin, inévitablement, se pose la question de la fonction des membres postérieurs chez ces serpents. Trop réduits, ils ne pouvaient certainement pas jouer un rôle réel dans la locomotion. Ont été suggérés un rôle d'ancrage du corps dans les fissures ou leur intervention lors de l'accouplement (Scanlon et alii,
1999). Mais, étant simplement des membres en voie de régression totale, peut-être n'avaient-ils aucune fonction.
![]() Cet article correspond à une communication présentée au Colloque sur le Cénomanien organisé à Rouen (Octobre 2001) par B. Ferré,
M. Fouray et J. Tabouelle. Nous remercions tout particulièrement N.J. Sander qui a traduit ce texte en anglais dans l'urgence. E. Buffetaut, M.W. Caldwell et M.S.Y. Lee ont fait d'utiles commentaires et B. Granier a réalisé un important travail éditorial.
Cet article correspond à une communication présentée au Colloque sur le Cénomanien organisé à Rouen (Octobre 2001) par B. Ferré,
M. Fouray et J. Tabouelle. Nous remercions tout particulièrement N.J. Sander qui a traduit ce texte en anglais dans l'urgence. E. Buffetaut, M.W. Caldwell et M.S.Y. Lee ont fait d'utiles commentaires et B. Granier a réalisé un important travail éditorial.
![]()
Bolkay S.J. (1925).- Mesophis nopcsai n.g. n.sp. ein neues, schlangenähnliches reptil aus der unteren Kreide (Neocom) von Bilek-Selista (Ost-Hercegovina).- Glasnik zemaljskog Muzeja u Bosni i Hercegovini, Sarajevo, t. 37, pp. 125-135.
Buffrénil V. de & Rage J.C. (1993).- La "pachyostose" vertébrale de Simoliophis (Reptilia, Squamata) : données comparatives et considérations fonctionnelles.- Annales de Paléontologie, Paris, vol. 79, pp. 315-335.
Caldwell M.W. (1999).- Squamate phylogeny and the relationships of snakes and mosasauroids.- Zoological Journal of the Linnean Society, Oxford, vol. 125, pp. 155-147.
Caldwell M.W. & Lee M.S.Y. (1997).- A snake with legs from the marine Cretaceous of the Middle East.- Nature, London, t. 386, pp. 705-709.
Chalifa Y. & Tchernov E. (1982).- Pachyamia latimaxillaris, new genus and species (Actinopterygii : Amiidae), from the Cenomanian of Jerusalem.- Journal of Vertebrate Paleontology, Los Angeles, vol. 2, pp. 269-285.
Cuny G., Jaeger J.J., Mahboubi M. & Rage J.C. (1990).- Les plus anciens serpents (Reptilia, Squamata) connus. Mise au point sur l'âge géologique des serpents de la partie moyenne du Crétacé.- Comptes rendus des séances de l'Académie des Sciences, Paris, II, t. 311, pp. 1267-1272.
Dominik W. (1985).- Stratigraphie und Sedimentologie (Geochemie, Schwermineralanalyse) der Oberkreide von Bahariya und ihre Korrelation zum Dakhla-Becken (Western-Desert, Ägypten). -Berliner Geowissenschaftliche Abhandlungen, Berlin A, vol. 62, pp. 1-172.
Gardner J.D. & Cifelli R.L. (1999).- A primitive snake from the Cretaceous of Utah.- Special Papers in Palaeontology, London, nº 60, pp. 87-100.
Haas G. (1979).- On a new Snakelike Reptile from the Lower Cenomanian of Ein Jabrud, near Jerusalem.- Bulletin du Muséum national d'Histoire Naturelle de Paris, Paris, t. 1, C, pp. 51-64.
Haas G. (1980).- Remarks on a new ophiomorph reptile from the lower Cenomanian of Ein Jabrud, Israel.- In : Aspects of Vertebrate History (Jacobs L.L., Ed.), Museum of Northern Arizona Press, Flagstaff, pp. 177-192.
Hoffstetter R. (1955).- Squamates de type moderne.- Traité de Paléontologie, 5. Masson, Paris, pp. 606-662.
Hoffstetter R. (1959).- Un serpent terrestre dans le Crétacé du Sahara.- Bulletin de la Société géologique de France, Paris, (7), t. I, pp. 897-902.
Jonet S. (1981).- Contribution à l'étude des vertébrés du Crétacé portugais et spécialement du Cénomanien de l'Estremadure.- Comunicações dos Serviços Geológicos de Portugal, Lisboa, t. 67, pp. 191-300.
Lapparent de Broin F. de (2000).- The oldest pre-podocnemidid turtle (Chelonii, Pleurodira), from the early Cretaceous, Ceará State, Brasil, and its environment.- Treballs del Museu de Geologia de Barcelona, Barcelona, nº 9, pp. 43-95.
Lee M.S.Y. (1997).- The phylogeny of varanoid lizards and the affinities of snakes. -Philosophical Transactions of the Royal Society of London, London, B, vol. 352, pp. 53-91.
Lee M.S.Y. & Caldwell M.W. (1998).- Anatomy and relationships of Pachyrhachis problematicus, a primitive snake with hindlimbs.- Philosophical Transactions of the Royal Society of London, London, B, vol. 353, pp. 1521-1552.
Lee M.S.Y., Caldwell M.W. & Scanlon J.D. (1999).- A second primitive marine snake : Pachyophis woodwardi from the Cretaceous of Bosnia-Herzegovina.- Journal of Zoology, London, vol. 248, pp. 509-520.
Lee M.S.Y. & Caldwell M.W. (2000).- Adriosaurus and the affinities of mosasaurs, dolichosaurs, and snakes.- Journal of Paleontology, Lawrence, vol. 74, pp. 915-937.
Lee M.S.Y. & Scanlon J.D. (2002).- Snake phylogeny based on osteology, soft anatomy and ecology.- Biological Review, Cambridge, vol. 77, pp. 333-401.
McDowell S.B. (1987).- Systematics.- In : Snakes. Ecology, and evolutionary Biology (Seigel R.A., Collins J.T. & Novak S.S., Eds.), McMillan, New York, pp. 3-50.
Nessov L.A., Zhegallo V.I. & Averianov A.O. (1998).- A new locality of Late Cretaceous snakes, mammals and other vertebrates in Africa (western Libya).- Annales de Paléontologie, Paris, vol. 84, pp. 265-274.
Nopcsa F. (1923).- Vorläufige Notiz über die Pachyostose und Osteosklerose einiger mariner Wirbeltiere.- Anatomischer Anzeiger, Jena, t. 56, pp. 353-359.
Nopcsa F. (1925).- Die Symoliophis-Reste.-In : Ergbenisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens, II.- Abhandlungen der Bayerischen Akademie der Wissenschaften, Mathematisch-naturwissenschaftliche Abteilung, München, vol. 30, pp. 1-27.
Nopcsa F. (1931).- Osteologia Reptilium fossilium et recentium. II : Appendix.- Fossilium Catalogus I. Animalia, Berlin, Pars 50, pp. 1-62.
Rage J.C. (1984).- Serpentes.- In : Handbuch der Paläoherpetologie (Wellnhofer P., Ed.), part 11. Gustav Fischer Verlag, Stuttgart/New York, xii + 80 p.
Rage J.C. (1987).- Fossil History.- In : Snakes. Ecology, and evolutionary Biology (Seigel R.A., Collins J.T. & Novak S.S., Eds.), McMillan, New York, pp. 51-76.
Rage J.C. (1988).- Un serpent primitif (Reptilia, Serpentes) dans le Cénomanien (base du Crétacé supérieur).- Comptes rendus des séances de l'Académie des Sciences, Paris, II, t. 307, pp. 1027-1032.
Rage J.C. (2000).- Phylogénie et origine des serpents.- Bulletin de la Société herpétologique de France, Paris, nº 96, pp. 57-69.
Rage J.C. & Cappetta H. (2002).- Vertebrates from the Cenomanian, and the geological age of the Draa Ubari fauna (Libya).- Annales de Paléontologie, Paris, vol. 88, pp. 79-84.
Rage J.C. & Escuillié F. (2000).- Un nouveau serpent bipède du Cénomanien (Crétacé). Implications phylétiques.- Comptes rendus des séances de l'Académie des Sciences, Paris, IIa, t. 330, pp. 513-520.
Rage J.C. & Escuillié F. (2002).- Eupodophis, new name for the genus Podophis Rage and Escuillié, 2000, an extinct bipedal snake, preoccupied by Podophis Wiegmann, 1843 (Lacertilia, Scincidae).- Amphibia-Reptilia, Leyden, vol. 23, pp. 232-233.
Rage J.C. & Richter A. (1994).- A snake from the Lower Cretaceous (Barremian) of Spain : The oldest known snake.- Neues Jarbuch für Geologie und Paläontologie, Monatshefte, Stuttgart, fasc. 9, pp. 561-565.
Rieppel O. & Zaher H. (2000).- The intramandibular joint in squamates, and the phylogenetic relationships of the fossil snake Pachyrhachis problematicus Haas.- Fieldiana, Chicago, nº 3, pp. 1-69.
Sauvage H.E. (1880).- Sur l'existence d'un reptile du type ophidien dans les couches à Ostrea columba des Charentes.- Comptes rendus hebdomadaires des séances de l'Académie des Sciences, Paris, t. 91, pp. 671-672.
Sauvage H.E. (1898).- Vertébrés fossiles du Portugal. Contribution à l'étude des poissons et des reptiles du Jurassique et du Crétacique.- Memorias Commissão do Serviço Geológico de Portugal, Lisboa, 1897-1898, pp. 1-46.
Scanlon J.D., Lee M.S.Y., Caldwell M.W. & Shine R. (1999).- The palaeocology of the primitive snake Pachyrhachis.- Historical Biology, Chur, vol. 13, pp. 127-152.
Scanlon J.D. & Lee M.S.Y. (2000).- The Pleistocene snake Wonambi and the early evolution of snakes.- Nature, London, vol. 403, pp. 416-420.
Sereno P.C., Dutheil D.B., Iarochene M., Larsson H.C.E., Lyon G.H., Magwene P.M., Sidor C.A., Varrichio D.J. & Wilson J.A. (1996).- Predatory dinosaurs from the Sahara and late Cretaceous faunal differentiation.- Science, Washington, vol. 272, pp. 986-991.
Sliskovic T. (1970).- Die stratigraphische Lage der Schichten mit Pachyophiidae aus Seliste bei Bileca (Ostherzegowina).- Bulletin scientifique, Conseil des Académies des Sciences et des Arts de la RSF de Yougoslavie, Zagreb, t. 15, pp. 389-390.
Tchernov E., Rieppel O., Zaher H., Polcyn M.J. & Jacobs L.L. (2000).- A fossil snake with limbs.- Science, Washington, vol. 287, pp. 2010-2012.
Voigt S. (1996).- Paläobiogeographie oberkretazicher Inoceramen und Rudisten - Ozeanographische und Klimatologische Konsequenzen einer neuen Paläogeographie.- Münchner Geowissenschaftliche Abhandlungen, München, A, vol. 31, pp. 1-101.
Wallach V. (1984).- A new name for Ophiomorphus colberti Haas, 1980.- Journal of Herpetology, Riverside, vol. 18, p. 239.
Werner C. (1989).- Die Elasmobranchier-Fauna des Gebel Dist Member der Bahariya Formation (OberCenoman) der Oase Bahariya, Ägypten.- Paleo Ichthyologica, München, vol. 5, pp. 1-112.
Zaher H. & Rieppel O. (1999).- The phylogenetic relationships of Pachyrhachis problematicus, and the evolution of limblessness in snakes (Lepidosauria, Squamata).- Comptes rendus des séances de l'Académie des Sciences, Paris, IIa, t. 329, pp. 831-837.
Zaher H. & Rieppel O. (2000).- A brief history of snakes.- Herpetological Review, Saint-Louis, vol. 31, pp. 73-76.
![]()
Cliquer sur la miniature pour agrandir l'image.
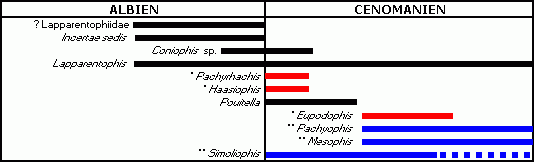
Figure 1 : Répartition stratigraphique des plus anciens serpents (*serpents bipèdes avérés ; **serpents supposés bipèdes). Remarque : en dehors de Simoliophis, chaque taxon provient d'un seul gisement ; chaque trait correspond donc à une extension possible maximale.

Cliquer sur la miniature pour agrandir l'image.
Figure 2 : Eupodophis descouensi. Al Nammoura, Liban, Cénomanien. Vue ventrale. L'animal a été cassé et la partie postérieure s'est fossilisée près de la tête. La longueur totale était d'environ 85 cm.

Cliquer sur la miniature pour agrandir l'image.
Figure 3 : Eupodophis descouensi. Al Nammoura, Liban, Cénomanien. Régions sacrée et caudale avec le membre postérieur ; ce dernier est représenté par le fémur, le tibia et la fibula, plus quelques os plus distaux.
Cliquer sur la miniature pour agrandir l'image.
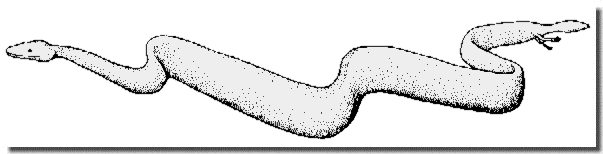
Figure 4 : Reconstitution de Eupodophis descouensi (noter la taille et la position des membres).
Cliquer sur la miniature pour agrandir l'image.
Figure 5 : Relations des serpents bipèdes. Ces serpents pourraient être "primitifs" (A) ou "évolués" (B).
Cliquer sur la miniature pour agrandir l'image.
Figure 6 : Répartition géographique des serpents bipèdes et présumés bipèdes. Reconstitution de la région "méditerranéenne" au Cénomanien d'après Voigt (1996), simplifiée. E : Eupodophis ; H+P : Haasiophis et Pachyrhachis ; P+M : Pachyophis et Mesophis ; S : Simoliophis (en raison de leur proximité, tous les gisements ayant fourni Simoliophis en France, au Portugal et au Maroc n'ont pas été placés).
![]()