![]()
![]()
![]() The Mahakam delta is a mixed, fluvial and wave dominated delta located in the eastern part of the island of Borneo (the East Kalimantan province of Indonesia). The distribution of benthic fauna in this delta system is influenced by the combined or antagonistic action of three main parameters: fluvial input of fresh water and sediment, tide, and a strong regional north to south drift current. A model of the present-day faunal distribution has been established taking into account the perturbations induced by the tide and by the regional drift current. This model has been extrapolated into the recent past through the study of shallow cores that permit the reconstruction of late Holocene deltaic regression patterns.
The Mahakam delta is a mixed, fluvial and wave dominated delta located in the eastern part of the island of Borneo (the East Kalimantan province of Indonesia). The distribution of benthic fauna in this delta system is influenced by the combined or antagonistic action of three main parameters: fluvial input of fresh water and sediment, tide, and a strong regional north to south drift current. A model of the present-day faunal distribution has been established taking into account the perturbations induced by the tide and by the regional drift current. This model has been extrapolated into the recent past through the study of shallow cores that permit the reconstruction of late Holocene deltaic regression patterns.
Indonesia, Kalimantan, Mahakam Delta, fluvial input, tide, drift current, benthonic Foraminifera, biofacies, distribution pattern.
Lambert B. (2003).- Micropaleontological investigations in the modern Mahakam delta, East Kalimantan (Indonesia).- Carnets de Géologie / Notebooks on Geology, Maintenon, Article 2003/02 (CG2003_A02)
![]() Recherches micropaléontologiques dans le delta actuel de la Mahakam, Est de lîle de Kalimantan
(Indonésie).- Le delta de la Mahakam est un delta mixte, influencé par les apports fluviaux et la marée, localisé sur la partie est de l'île de Bornéo (province indonésienne de Kalimantan
Est). La répartition de la microfaune benthique est controlée par l'action parfois combinée, parfois antagoniste de trois principaux
facteurs, l'apport fluvial d'eau douce et de sédiments, de la marée et de l'action d'un fort courant balayant du
Nord vers le Sud le détroit de Makassar qui sépare l'île de Bornéo de l'archipel des Célèbes. Un modèle de distribution de la microfaune actuelle a été établi prenant en compte les perturbations induites par l'action de la marée et du courant de dérive. Ce modèle de distribution a pu être extrapolé au passé par le biais de l'étude de carottages superficiels illustrant la régression holocène.
Recherches micropaléontologiques dans le delta actuel de la Mahakam, Est de lîle de Kalimantan
(Indonésie).- Le delta de la Mahakam est un delta mixte, influencé par les apports fluviaux et la marée, localisé sur la partie est de l'île de Bornéo (province indonésienne de Kalimantan
Est). La répartition de la microfaune benthique est controlée par l'action parfois combinée, parfois antagoniste de trois principaux
facteurs, l'apport fluvial d'eau douce et de sédiments, de la marée et de l'action d'un fort courant balayant du
Nord vers le Sud le détroit de Makassar qui sépare l'île de Bornéo de l'archipel des Célèbes. Un modèle de distribution de la microfaune actuelle a été établi prenant en compte les perturbations induites par l'action de la marée et du courant de dérive. Ce modèle de distribution a pu être extrapolé au passé par le biais de l'étude de carottages superficiels illustrant la régression holocène.
![]() Indonésie, Kalimantan, delta de la Mahakam, apports fluviaux, marée, courant
de dérive, Foraminifères benthiques, biofacies, modèle de distribution.
Indonésie, Kalimantan, delta de la Mahakam, apports fluviaux, marée, courant
de dérive, Foraminifères benthiques, biofacies, modèle de distribution.
![]() Recherches micropaléontologiques dans le delta actuel de la Mahakam, Est de lîle de Kalimantan
(Indonésie).- Le delta de la Mahakam est un delta mixte, influencé par les apports fluviaux et la marée, localisé en Indonésie
(sur lîle de Kalimantan et débouchant sur le détroit de Makassar) sous un climat équatorial
humide. Le courant balayant le détroit de Makassar (de direction nord-sud) a pour effet de perturber les dépôts sableux qui se concentrent dans les parties centrale et sud du delta. La fraction pélitique est répartie sur un large périmètre et se retrouve très loin vers le
Sud.
Recherches micropaléontologiques dans le delta actuel de la Mahakam, Est de lîle de Kalimantan
(Indonésie).- Le delta de la Mahakam est un delta mixte, influencé par les apports fluviaux et la marée, localisé en Indonésie
(sur lîle de Kalimantan et débouchant sur le détroit de Makassar) sous un climat équatorial
humide. Le courant balayant le détroit de Makassar (de direction nord-sud) a pour effet de perturber les dépôts sableux qui se concentrent dans les parties centrale et sud du delta. La fraction pélitique est répartie sur un large périmètre et se retrouve très loin vers le
Sud.
![]() La répartition de la microfaune benthique à lintérieur de ce système deltaïque va être influencée par laction conjuguée ou antagoniste de ces trois facteurs
(apport fluvial, marée et courant de dérive).
La répartition de la microfaune benthique à lintérieur de ce système deltaïque va être influencée par laction conjuguée ou antagoniste de ces trois facteurs
(apport fluvial, marée et courant de dérive).
![]() Les apports fluviaux dominants au
Centre et au Sud se traduisent par la présence dun "delta
front" très développé où les cortèges faunistiques sont relativement bien discriminés avec:
Les apports fluviaux dominants au
Centre et au Sud se traduisent par la présence dun "delta
front" très développé où les cortèges faunistiques sont relativement bien discriminés avec:
![]() Linfluence de la marée se traduit dune manière assez spectaculaire par la
"remontée" de microfaunes de la partie externe du "delta front" (Asterorotalia) voire du prodelta (Operculina, Pseudorotalia) à lintérieur des chenaux localisés dans la partie interne du "delta front. Cette observation est fondamentale pour lexploitation des données fossiles où des dépôts sableux chenalisants contenant une microfaune calcaire sont souvent abusivement interprétés comme des dépôts de
plate-forme interne.
Linfluence de la marée se traduit dune manière assez spectaculaire par la
"remontée" de microfaunes de la partie externe du "delta front" (Asterorotalia) voire du prodelta (Operculina, Pseudorotalia) à lintérieur des chenaux localisés dans la partie interne du "delta front. Cette observation est fondamentale pour lexploitation des données fossiles où des dépôts sableux chenalisants contenant une microfaune calcaire sont souvent abusivement interprétés comme des dépôts de
plate-forme interne.
![]() Linfluence du courant de dérive sexerce essentiellement sur la partie nord du delta où il renforce considérablement laction perturbatrice de la marée. On assiste à une véritable migration de microfossiles dont lorigine est parfois
lointaine (Calcarina spengleri (Gmelin) 1791, Calcarina venusta (Brady) 1884, Elphidium
"ibericum" (Schrodt) 1932), provenant des faciès sub-coralligènes situés au Nord et pénétrant très profondément à lintérieur du "delta
front" jusquà atteindre les limites de la partie tidale du delta, le cas ultime sobservant au niveau amont des chenaux de "delta
front" dans lesquels le contraste est saisissant entre le fond (Asterorotalia, Pseudorotalia, Operculina, Calcarina) et les "tidal
flats" bordiers (Trochammina, Ammobaculites, Eggerelloides). Si les facteurs mécaniques sont indéniables (tests roulés, brisés fréquents dans la partie nord du delta), il nest pas exclu quune partie de cette microfaune ait trouvée dans le fond des
chenaux, dont leau est plus salée, un milieu refuge.
Linfluence du courant de dérive sexerce essentiellement sur la partie nord du delta où il renforce considérablement laction perturbatrice de la marée. On assiste à une véritable migration de microfossiles dont lorigine est parfois
lointaine (Calcarina spengleri (Gmelin) 1791, Calcarina venusta (Brady) 1884, Elphidium
"ibericum" (Schrodt) 1932), provenant des faciès sub-coralligènes situés au Nord et pénétrant très profondément à lintérieur du "delta
front" jusquà atteindre les limites de la partie tidale du delta, le cas ultime sobservant au niveau amont des chenaux de "delta
front" dans lesquels le contraste est saisissant entre le fond (Asterorotalia, Pseudorotalia, Operculina, Calcarina) et les "tidal
flats" bordiers (Trochammina, Ammobaculites, Eggerelloides). Si les facteurs mécaniques sont indéniables (tests roulés, brisés fréquents dans la partie nord du delta), il nest pas exclu quune partie de cette microfaune ait trouvée dans le fond des
chenaux, dont leau est plus salée, un milieu refuge.
![]() Une série de petites carottes prélevées dans la partie
sud-est, très fortement progradante, a permis détudier le comportement de la microfaune face à larrivée massive de sable. Une cartographie assez précise de la distribution faunistique associée à la mise en place dune barre sableuse a pu être établie. Grâce au contenu des
carottes, cest lévolution chronologique des associations qui a pu être déterminée. Il ressort de cette étude que les progradations sont extrêmement rapides et très étendues (la barre sableuse représente une surface de cinq kilomètres sur cinq kilomètres pour une épaisseur de quelques mètres).
Une série de petites carottes prélevées dans la partie
sud-est, très fortement progradante, a permis détudier le comportement de la microfaune face à larrivée massive de sable. Une cartographie assez précise de la distribution faunistique associée à la mise en place dune barre sableuse a pu être établie. Grâce au contenu des
carottes, cest lévolution chronologique des associations qui a pu être déterminée. Il ressort de cette étude que les progradations sont extrêmement rapides et très étendues (la barre sableuse représente une surface de cinq kilomètres sur cinq kilomètres pour une épaisseur de quelques mètres).
![]() La variété des biofaciès rencontrés nous oblige à dessiner un modèle de répartition relativement complexe à la hauteur de la diversité des facteurs physico-chimiques influant sur la distribution des tests. Il constitue un outil important pour linterprétation des séries néogènes sous-jacentes au delta
actuel. Ce modèle permet tout à la fois, de mieux apprécier les variations de faciès à petite échelle (à léchelle de quelques kilomètres) et de mieux
mesurer, à la lueur de la complexité des phénomènes observés, les limites de nos propres
interprétations.
La variété des biofaciès rencontrés nous oblige à dessiner un modèle de répartition relativement complexe à la hauteur de la diversité des facteurs physico-chimiques influant sur la distribution des tests. Il constitue un outil important pour linterprétation des séries néogènes sous-jacentes au delta
actuel. Ce modèle permet tout à la fois, de mieux apprécier les variations de faciès à petite échelle (à léchelle de quelques kilomètres) et de mieux
mesurer, à la lueur de la complexité des phénomènes observés, les limites de nos propres
interprétations.
![]()
![]() Located in the eastern central portion of the island of Borneo in the Indonesian province of Kalimantan, the Mahakam river drains the Kutei basin and debouches into the Makassar strait. The climate is humid and tropical with an average rainfall of over 260 cm
(100") a month. Consequently, deposits in the broad, many-mouthed delta of the Mahakam include all types of terrigenous detritus. Its site of deposition is greatly influenced by the strong north-south currents of the Makassar strait and by wave action so the sand-sized fraction is concentrated in the central and southern portions of the delta, thus permitting the classification of the delta as mixed fluvial/tidal. The pelitic fraction is spread over a large area and is transported far to the south.
Located in the eastern central portion of the island of Borneo in the Indonesian province of Kalimantan, the Mahakam river drains the Kutei basin and debouches into the Makassar strait. The climate is humid and tropical with an average rainfall of over 260 cm
(100") a month. Consequently, deposits in the broad, many-mouthed delta of the Mahakam include all types of terrigenous detritus. Its site of deposition is greatly influenced by the strong north-south currents of the Makassar strait and by wave action so the sand-sized fraction is concentrated in the central and southern portions of the delta, thus permitting the classification of the delta as mixed fluvial/tidal. The pelitic fraction is spread over a large area and is transported far to the south.
![]() The distribution of the benthonic microfauna in the deltaic system is controlled by three factors: fluvial input, tidal action and drift currents which may act in parallel or in opposition.
The distribution of the benthonic microfauna in the deltaic system is controlled by three factors: fluvial input, tidal action and drift currents which may act in parallel or in opposition.
![]() For the micropaleontologist the reconstruction of paleoenvironments is a constant preoccupation, particularly for those engaged in exploration for hydrocarbons. A number of contributions have been published on this topic, among them several regarding existing distribution patterns. An understanding of modern environments is paramount and is the key to an understanding of ancient systems. This approach is particularly valid for reconstructing environments of the last ten million years (when taxa were similar to those of existing faunas). This study investigates the distribution of foraminiferal assemblages in the Mahakam delta
(Fig. 1
For the micropaleontologist the reconstruction of paleoenvironments is a constant preoccupation, particularly for those engaged in exploration for hydrocarbons. A number of contributions have been published on this topic, among them several regarding existing distribution patterns. An understanding of modern environments is paramount and is the key to an understanding of ancient systems. This approach is particularly valid for reconstructing environments of the last ten million years (when taxa were similar to those of existing faunas). This study investigates the distribution of foraminiferal assemblages in the Mahakam delta
(Fig. 1
![]() ), particularly those in the active depositional center, and to analyze some of the environmental parameters which may influence microfaunal
distribution. It should be of considerable value for the interpretation of ancient delta deposits in
Kutei.
), particularly those in the active depositional center, and to analyze some of the environmental parameters which may influence microfaunal
distribution. It should be of considerable value for the interpretation of ancient delta deposits in
Kutei.
![]() The Kutei basin has been an oil producer for a long time (since the end of the 19th century). The reservoirs are mainly deltaic sandstones of Early to Late Miocene age. The presence of a modern delta superimposed on its Neogene oil-producing equivalents constituted an ideal opportunity for a comparison of modern and ancient delta systems.
The Kutei basin has been an oil producer for a long time (since the end of the 19th century). The reservoirs are mainly deltaic sandstones of Early to Late Miocene age. The presence of a modern delta superimposed on its Neogene oil-producing equivalents constituted an ideal opportunity for a comparison of modern and ancient delta systems.
![]() This opportunity was first exploited in
1974 by sedimentologists
(TOTAL internal reports: Allen G.P., Cassoudebat
& Thouvenin, 1979). During their field trips, a great number of surface samples were collected (350). In 1992, another field trip, devoted specifically to the mouth bar complex, resulted in the collection of a hundred more surface and core samples.
This opportunity was first exploited in
1974 by sedimentologists
(TOTAL internal reports: Allen G.P., Cassoudebat
& Thouvenin, 1979). During their field trips, a great number of surface samples were collected (350). In 1992, another field trip, devoted specifically to the mouth bar complex, resulted in the collection of a hundred more surface and core samples.
![]() Studies by Coleman
& Wright (1975) and Galloway
(1975) have shown that deltas can be categorized into several types. Differences in morphology and internal geometry are generated by the relative importance of various physical parameters, such as water flow, sediment input, wind, currents, waves and tidal processes. The classification adopted here centers around poles corresponding to the three main classes:
Studies by Coleman
& Wright (1975) and Galloway
(1975) have shown that deltas can be categorized into several types. Differences in morphology and internal geometry are generated by the relative importance of various physical parameters, such as water flow, sediment input, wind, currents, waves and tidal processes. The classification adopted here centers around poles corresponding to the three main classes:
![]() The Mahakam delta is a mixed, tide- and river-dominated delta in a humid equatorial climate.
The Mahakam delta is a mixed, tide- and river-dominated delta in a humid equatorial climate.
![]() Literature devoted to the micropaleontology of deltas as such is scare. In comparison, coastal or estuarine environments are better known (Haman,
1981; Jennings & Nelson,
1992; Jonasson & Patterson,
1992). However some deltaic systems have been studied and provide data relevant to the present study.
Literature devoted to the micropaleontology of deltas as such is scare. In comparison, coastal or estuarine environments are better known (Haman,
1981; Jennings & Nelson,
1992; Jonasson & Patterson,
1992). However some deltaic systems have been studied and provide data relevant to the present study.
![]() Lankford
(1959) was the first to introduce a zonation based on microfauna distribution. He distinguished six formal assemblages, namely marsh, interdistributary bay, fluvial marine, deltaic marine, sand, and open shelf. Anderson
(1968) completed this early work.
Lankford
(1959) was the first to introduce a zonation based on microfauna distribution. He distinguished six formal assemblages, namely marsh, interdistributary bay, fluvial marine, deltaic marine, sand, and open shelf. Anderson
(1968) completed this early work.
![]() The main contribution was published by Kruit
(1955). This study shows a clear dominance of Ammonia beccarii, associated with Nonion depressulum (Walker & Jacob)
1798. These two species are present in the outer part of the deltaic complex (present-day beaches, abandoned coastal ridges, coastal lagoons). Agglutinated Foraminifera (Trochammina, Haplophragmoides) are restricted to the inner part of the deltaic system: swamps.
The main contribution was published by Kruit
(1955). This study shows a clear dominance of Ammonia beccarii, associated with Nonion depressulum (Walker & Jacob)
1798. These two species are present in the outer part of the deltaic complex (present-day beaches, abandoned coastal ridges, coastal lagoons). Agglutinated Foraminifera (Trochammina, Haplophragmoides) are restricted to the inner part of the deltaic system: swamps.
![]() Most recently the previous works have been completed by Vangerow
(1974).
Most recently the previous works have been completed by Vangerow
(1974).
![]() During the past twenty years, several studies have been published on modern micropaleontological distributions in the Mahakam delta (Oesterle,
1974; Hoibian, 1984) and neighboring areas (Coustillas,
1983; Boichard et alii, 1985; Granier, Villain & Boichard,
1997).
During the past twenty years, several studies have been published on modern micropaleontological distributions in the Mahakam delta (Oesterle,
1974; Hoibian, 1984) and neighboring areas (Coustillas,
1983; Boichard et alii, 1985; Granier, Villain & Boichard,
1997).
![]() In his pioneer work,
Oesterle (TOTAL internal report, 1974) distinguished four biofacies associated respectively with the deltaic plain, inner delta front, outer delta front and prodelta slope. These subdivisions were later used by
Hoibian (1984).
In his pioneer work,
Oesterle (TOTAL internal report, 1974) distinguished four biofacies associated respectively with the deltaic plain, inner delta front, outer delta front and prodelta slope. These subdivisions were later used by
Hoibian (1984).
![]() The present study supplements earlier work by including a sedimentological approach and proposing a micropaleontological model for deltaic clastic deposits that are potential reservoirs for oil.
The present study supplements earlier work by including a sedimentological approach and proposing a micropaleontological model for deltaic clastic deposits that are potential reservoirs for oil.
![]()
![]() The Mahakam delta is located in Indonesia (Fig. 1
The Mahakam delta is located in Indonesia (Fig. 1
![]() ) on the eastern coast of Kalimantan Island (between 0°21 and 1°10 south latitude, and 117°40 east longitude). The coastline of this part of Kalimantan is oriented NNE-SSW, and borders the Makassar strait. This delta, like all modern deltas, is a recent feature, having developed in the last phase of the Holocene transgression, over a 5,000 to 7,000 year period during which delta progradation gave rise to a system comprising approximately 1,300 km2 of subaerial delta plain, 1,000 km2 of delta front and 2,700 km2 of prodelta accumulations
(Allen G.P., Cassoudebat
& Thouvenin, 1979).
) on the eastern coast of Kalimantan Island (between 0°21 and 1°10 south latitude, and 117°40 east longitude). The coastline of this part of Kalimantan is oriented NNE-SSW, and borders the Makassar strait. This delta, like all modern deltas, is a recent feature, having developed in the last phase of the Holocene transgression, over a 5,000 to 7,000 year period during which delta progradation gave rise to a system comprising approximately 1,300 km2 of subaerial delta plain, 1,000 km2 of delta front and 2,700 km2 of prodelta accumulations
(Allen G.P., Cassoudebat
& Thouvenin, 1979).
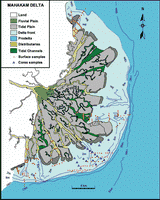
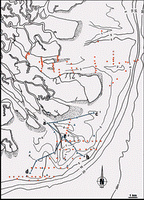
![]() The general morphology of the delta can be divided into three radially concentric systems
(Fig. 2
The general morphology of the delta can be divided into three radially concentric systems
(Fig. 2 ![]() ): the delta plain, delta front, and prodelta (Allen G.P., Cassoudebat & Thouvenin,
1979).
): the delta plain, delta front, and prodelta (Allen G.P., Cassoudebat & Thouvenin,
1979).
![]() The delta plain forms the subaerial part of the delta. It can be subdivided into a fluvial delta plain, and a tidal delta plain. The fluvial delta plain is characterized by highly compacted and well drained ground, covered with hardwood trees. Its width is between 10 and 20 km. The tidal delta plain is characterized by its low elevation that consequently is subject to daily inundations. The plant cover is Nipah palms and mangroves. The tidal deltaic plain is incised by both distributaries and tidal channels. Its width ranges between between 20 and 30 km
(Allen G.P., Cassoudebat
& Thouvenin, 1979).
The delta plain forms the subaerial part of the delta. It can be subdivided into a fluvial delta plain, and a tidal delta plain. The fluvial delta plain is characterized by highly compacted and well drained ground, covered with hardwood trees. Its width is between 10 and 20 km. The tidal delta plain is characterized by its low elevation that consequently is subject to daily inundations. The plant cover is Nipah palms and mangroves. The tidal deltaic plain is incised by both distributaries and tidal channels. Its width ranges between between 20 and 30 km
(Allen G.P., Cassoudebat
& Thouvenin, 1979).
![]() The delta front is an intertidal to shallow subtidal platform. Its width ranges between 8 and 10 km. The topography consists of linear undulations perpendicular to the coast forming bars and shoals. The delta front too is incised by distributary channels. They extend seaward to its outer limit, terminating in a mouth bar. The inner portion is made up of extensive tidal flats.
The delta front is an intertidal to shallow subtidal platform. Its width ranges between 8 and 10 km. The topography consists of linear undulations perpendicular to the coast forming bars and shoals. The delta front too is incised by distributary channels. They extend seaward to its outer limit, terminating in a mouth bar. The inner portion is made up of extensive tidal flats.
![]() The prodelta is a smooth seaward slope, the inner part set off by an abrupt break in the slope at the 5 m isobath. The outer limit is between the 60 and 70 m isobaths. The width of the prodelta shows a sharp asymmetry, an effect of the drift current. Off the southern distributary system it is 30 km wide, but 5 to 15 km wide in the central and northern portions (see
below).
The prodelta is a smooth seaward slope, the inner part set off by an abrupt break in the slope at the 5 m isobath. The outer limit is between the 60 and 70 m isobaths. The width of the prodelta shows a sharp asymmetry, an effect of the drift current. Off the southern distributary system it is 30 km wide, but 5 to 15 km wide in the central and northern portions (see
below).
![]() The repartition of benthonic microfauna seems to be controlled by two parameters:
The repartition of benthonic microfauna seems to be controlled by two parameters:
![]()
![]() In this study, we have selected for investigation taxa well represented in both surface and core
samples (such as Trochammina, Ammonia, Asterorotalia, Pseudorotalia), neglecting the lower frequencies (such as Miliolids). We have considered unimportant the determination to specific
levels of several genera, for example: "Elphidium
group" includes all small species (Elphidium advenum (Cushman) 1922 (Pl.
1, fig. 11
In this study, we have selected for investigation taxa well represented in both surface and core
samples (such as Trochammina, Ammonia, Asterorotalia, Pseudorotalia), neglecting the lower frequencies (such as Miliolids). We have considered unimportant the determination to specific
levels of several genera, for example: "Elphidium
group" includes all small species (Elphidium advenum (Cushman) 1922 (Pl.
1, fig. 11 ![]() ),
E. crispum (Linné) 1767 (Pl. 1, fig. 9
),
E. crispum (Linné) 1767 (Pl. 1, fig. 9 ![]() ), "Pseudorotalia
group" includes mainly Pseudorotalia conoides associated with a few Pseudorotalia schroeteriana (Parker & Jones) 1862, "Nonion
group" includes Nonion boueanum (Orbigny) 1846, Nonion scaphum (Fitchel & Moll) 1798, Nonion sp.
), "Pseudorotalia
group" includes mainly Pseudorotalia conoides associated with a few Pseudorotalia schroeteriana (Parker & Jones) 1862, "Nonion
group" includes Nonion boueanum (Orbigny) 1846, Nonion scaphum (Fitchel & Moll) 1798, Nonion sp.
![]() The following relationships between microfauna and facies were
established:
The following relationships between microfauna and facies were
established:
![]() The main characteristic of this area is the complex interaction between several factors:
The main characteristic of this area is the complex interaction between several factors:
![]() The sandy beaches contain a specific assemblage dominated by Calcarina spengleri
(Pl. 2, fig. 6
The sandy beaches contain a specific assemblage dominated by Calcarina spengleri
(Pl. 2, fig. 6 ![]() ) and Calcarina venusta
(Pl. 2, fig. 4
) and Calcarina venusta
(Pl. 2, fig. 4 ![]() )
associated with Elphidium "ibericum" and Operculina gaymardi. The origin of microfossils belonging to the genus Calcarina is traced to the nearby northern
patch reefs.
)
associated with Elphidium "ibericum" and Operculina gaymardi. The origin of microfossils belonging to the genus Calcarina is traced to the nearby northern
patch reefs.
![]() In contrast, the fauna of the muddy inner shelf is very different with a clear predominance of Asterorotalia trispinosa (deltaic influx).
In contrast, the fauna of the muddy inner shelf is very different with a clear predominance of Asterorotalia trispinosa (deltaic influx).
![]() The sandy bar located at the northern limit of the deltaic system presents a mixed association with Calcarina venusta, Asterorotalia trispinosa and the Elphidium group, thus comprising another biofacies.
The sandy bar located at the northern limit of the deltaic system presents a mixed association with Calcarina venusta, Asterorotalia trispinosa and the Elphidium group, thus comprising another biofacies.
![]() The tidal deltaic plain in this area is composed of muddy distributary channels and associated organic mud. The samples contain only agglutinated species (Trochammina group, Arenoparrella mexicana)
(Fig. 3-A
The tidal deltaic plain in this area is composed of muddy distributary channels and associated organic mud. The samples contain only agglutinated species (Trochammina group, Arenoparrella mexicana)
(Fig. 3-A ![]() ,
4-A
,
4-A ![]() ).
).
![]() The mud (tidal) flats located on either side of the distributary channel
(Fig. 3-B
The mud (tidal) flats located on either side of the distributary channel
(Fig. 3-B ![]() ,
4-B
,
4-B ![]() )
contain a more diversified assemblage of agglutinated Foraminifera (Trochammina group plus Trochammina globigeriniformis (Parker & Jones) 1865, Ammobaculites agglutinans, Eggerelloides scabrum). Channel bottoms contain a calcareous microfauna comprising various taxa: Asterorotalia trispinosa, Operculina gaymardi, Pseudorotalia group, Elphidium "ibericum" .
)
contain a more diversified assemblage of agglutinated Foraminifera (Trochammina group plus Trochammina globigeriniformis (Parker & Jones) 1865, Ammobaculites agglutinans, Eggerelloides scabrum). Channel bottoms contain a calcareous microfauna comprising various taxa: Asterorotalia trispinosa, Operculina gaymardi, Pseudorotalia group, Elphidium "ibericum" .
![]() The tidal flats contain an association composed of only two taxa (Asterorotalia trispinosa and the Elphidium group).
The tidal flats contain an association composed of only two taxa (Asterorotalia trispinosa and the Elphidium group).
![]() The fauna of the muddy delta front varies in composition depending on the location of the sample. The dominant taxon is Asterorotalia trispinosa. The samples located very close to the channel contain agglutinated taxa and Ammonia beccarii. The samples located within the channel contain a more varied calcareous microfauna: Asterorotalia trispinosa, Pseudorotalia group, Operculina gaymardi, Ammonia annectens, Elphidium "ibericum" , Calcarina spengleri (center of
Fig. 3
The fauna of the muddy delta front varies in composition depending on the location of the sample. The dominant taxon is Asterorotalia trispinosa. The samples located very close to the channel contain agglutinated taxa and Ammonia beccarii. The samples located within the channel contain a more varied calcareous microfauna: Asterorotalia trispinosa, Pseudorotalia group, Operculina gaymardi, Ammonia annectens, Elphidium "ibericum" , Calcarina spengleri (center of
Fig. 3 ![]() ).
).
![]() Because of the important fluvial influx the southern part of the delta is an active depositional center. The primary consequence is the existence of widespread mud (tidal) flats and the presence of a relatively wide delta
front.
Because of the important fluvial influx the southern part of the delta is an active depositional center. The primary consequence is the existence of widespread mud (tidal) flats and the presence of a relatively wide delta
front.
![]() In the mud flats associated with channels the tidal deltaic plain contains only agglutinated species: the Trochammina group, Ammotium salsum (Fig.
5
In the mud flats associated with channels the tidal deltaic plain contains only agglutinated species: the Trochammina group, Ammotium salsum (Fig.
5 ![]() ).
).
![]() The tidal flats contain a more diversified agglutinated microfauna: the Trochammina group, Ammobaculites agglutinans, Eggerelloides scabrum. In the outer part of these tidal flats we note the presence of Asterorotalia trispinosa.
The tidal flats contain a more diversified agglutinated microfauna: the Trochammina group, Ammobaculites agglutinans, Eggerelloides scabrum. In the outer part of these tidal flats we note the presence of Asterorotalia trispinosa.
![]() In the inner delta front
the bottom of the channels (Fig.
6 A, B, C
In the inner delta front
the bottom of the channels (Fig.
6 A, B, C ![]() ) contains a relatively more diversified calcareous microfauna: Ammonia beccarii, Asterorotalia trispinosa, Pseudorotalia group, Operculina gaymardi.
) contains a relatively more diversified calcareous microfauna: Ammonia beccarii, Asterorotalia trispinosa, Pseudorotalia group, Operculina gaymardi.
![]() In the sandy mouth bars Ammonia beccarii predominates.
In the sandy mouth bars Ammonia beccarii predominates.
![]() In the delta front
muds Asterorotalia trispinosa is ascendant.
In the delta front
muds Asterorotalia trispinosa is ascendant.
![]() The delta front - prodelta boundary too is dominated by Asterorotalia trispinosa associated with the Pseudorotalia group, Elphidium "ibericum" , and Operculina gaymardi.
The delta front - prodelta boundary too is dominated by Asterorotalia trispinosa associated with the Pseudorotalia group, Elphidium "ibericum" , and Operculina gaymardi.
![]() Beach deposits contain an association dominated by Ammonia beccarii which here is associated with Asterorotalia trispinosa and the Elphidium group.
Beach deposits contain an association dominated by Ammonia beccarii which here is associated with Asterorotalia trispinosa and the Elphidium group.
![]() A large mouth bar occupies most of this
area (Fig. 7
A large mouth bar occupies most of this
area (Fig. 7 ![]() ). This bar was cored in order to get a better understanding of progradational processes (see below).
). This bar was cored in order to get a better understanding of progradational processes (see below).
![]() The tidal flats contain only an agglutinated assemblage: Trochammina group, Ammobaculites agglutinans, Eggerelloides scabrum associated with rare Ammonia beccarii (6 chambers).
The tidal flats contain only an agglutinated assemblage: Trochammina group, Ammobaculites agglutinans, Eggerelloides scabrum associated with rare Ammonia beccarii (6 chambers).
![]() The inner sandy delta front is colonized by Ammonia beccarii (9 chambers).
The inner sandy delta front is colonized by Ammonia beccarii (9 chambers).
![]() The outer delta front presents a more diversified association with the appearance of Asterorotalia trispinosa and the Elphidium group.
The outer delta front presents a more diversified association with the appearance of Asterorotalia trispinosa and the Elphidium group.
![]() The tidal deltaic plain is composed of muddy channels. The microfauna is represented by agglutinated Foraminifera: the Trochammina group and numerous Arenoparrella mexicana.
The tidal deltaic plain is composed of muddy channels. The microfauna is represented by agglutinated Foraminifera: the Trochammina group and numerous Arenoparrella mexicana.
![]() The tidal flats are always dominated by agglutinated species: the Trochammina group, Ammobaculites agglutinans, Eggerelloides scabrum. As in other areas the channel bottoms contain a calcareous microfauna: Asterorotalia trispinosa, Ammonia beccarii
(Fig. 8-A
The tidal flats are always dominated by agglutinated species: the Trochammina group, Ammobaculites agglutinans, Eggerelloides scabrum. As in other areas the channel bottoms contain a calcareous microfauna: Asterorotalia trispinosa, Ammonia beccarii
(Fig. 8-A ![]() ,
8-B
,
8-B ![]() ).
).
![]() The muddy inner
delta front is dominated by Ammonia beccarii, with Asterorotalia trispinosa, the Elphidium group and the Nonion group increasing in number progressively seaward.
The muddy inner
delta front is dominated by Ammonia beccarii, with Asterorotalia trispinosa, the Elphidium group and the Nonion group increasing in number progressively seaward.
![]() The outer muddy delta front is dominated by Asterorotalia trispinosa.
The outer muddy delta front is dominated by Asterorotalia trispinosa.
![]() The boundary between the delta front and the prodelta is characterized by the presence of the Pseudorotalia group and Operculina gaymardi.
The boundary between the delta front and the prodelta is characterized by the presence of the Pseudorotalia group and Operculina gaymardi.
![]() In the southern and eastern parts of the Mahakam delta, the distribution of the several taxa seems to follow the same
pattern (Fig. 9
In the southern and eastern parts of the Mahakam delta, the distribution of the several taxa seems to follow the same
pattern (Fig. 9 ![]() ).
).
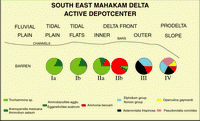
Biofacies Ia (Tidal Plain: mudflats, channels, 0-6 m water depth)
![]() The tidal deltaic plain contains only agglutinated Foraminifera represented by:
The tidal deltaic plain contains only agglutinated Foraminifera represented by:
Biofacies Ib (Tidal Plain: tidal flats, 0-2 m water depth)
![]() The tidal flats located on either side of the delta front channels present a more diversified group of arenaceous Foraminifera including:
The tidal flats located on either side of the delta front channels present a more diversified group of arenaceous Foraminifera including:
![]() In the northern parts of the Mahakam delta, Trochammina globigeriniformis
(Pl. 1, fig. 7
In the northern parts of the Mahakam delta, Trochammina globigeriniformis
(Pl. 1, fig. 7 ![]() )
is present.
)
is present.
Biofacies IIa (delta front, 0-2 m water depth)
![]() The first significant calcareous benthonic foraminifer to appear in the inner delta front is Ammonia beccarii (9 chambered specimens). The complete association is composed of the following species:
The first significant calcareous benthonic foraminifer to appear in the inner delta front is Ammonia beccarii (9 chambered specimens). The complete association is composed of the following species:
Biofacies IIb (delta front, 0-1 m water depth)
![]() In sandy bars deposited in the delta front, the number of Ammonia beccarii increases. Frequently the samples contain only numerous Ammonia specimens.
In sandy bars deposited in the delta front, the number of Ammonia beccarii increases. Frequently the samples contain only numerous Ammonia specimens.
Biofacies III (delta front, 1-5 m water depth)
![]() The outer part of the delta front presents a more diversified association with the appearance of Asterorotalia trispinosa
(Pl. 1, fig. 13
The outer part of the delta front presents a more diversified association with the appearance of Asterorotalia trispinosa
(Pl. 1, fig. 13 ![]() ),
the Elphidium group (Pl. 1, fig. 10
),
the Elphidium group (Pl. 1, fig. 10 ![]() ), and the Nonion
group (Pl. 1, fig. 12
), and the Nonion
group (Pl. 1, fig. 12 ![]() ).
).
Biofacies IV (prodelta, water depth> 5 m)
![]() Samples taken along the prodelta slope were characterized by a greater diversity of species:
Samples taken along the prodelta slope were characterized by a greater diversity of species:
![]() The general pattern of distribution observed in the southern and eastern parts of the delta does not fully apply in the northern parts due to the specific characteristics discussed above. This illustrates the need for caution and restraint when trying to apply directly and rigidly a modem model to ancient sediments.
The general pattern of distribution observed in the southern and eastern parts of the delta does not fully apply in the northern parts due to the specific characteristics discussed above. This illustrates the need for caution and restraint when trying to apply directly and rigidly a modem model to ancient sediments.
![]()
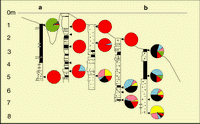
![]() Eighteen cores were taken in the eastern distributary part of the delta in order to investigate the most recent regression phase. Three representative profiles were
studied (Fig. 10
Eighteen cores were taken in the eastern distributary part of the delta in order to investigate the most recent regression phase. Three representative profiles were
studied (Fig. 10 ![]() ). The retained area is an active sandy mouth bar deposit. The general aspect of the sandy bar corresponds to a triangular body 5 to 6 km wide. This body is bounded to the SW by another sandy bar and to the NE by the muddy facies of the delta front. The front of the bar is adjacent to the edge of the prodelta. The first profile corresponds to a longitudinal section of the bar. The other two profiles correspond to transverse sections.
). The retained area is an active sandy mouth bar deposit. The general aspect of the sandy bar corresponds to a triangular body 5 to 6 km wide. This body is bounded to the SW by another sandy bar and to the NE by the muddy facies of the delta front. The front of the bar is adjacent to the edge of the prodelta. The first profile corresponds to a longitudinal section of the bar. The other two profiles correspond to transverse sections.
![]() Four cores are located on this NW-SE profile in a continuation of the main distributary
channel (Fig. 11
Four cores are located on this NW-SE profile in a continuation of the main distributary
channel (Fig. 11 ![]() ).
).
![]() The surface samples show the general trend from a mudflat facies (Trochammina sp., Ammobaculites agglutinans, Eggerelloides scabrum) to upper prodelta shales (Asterorotalia trispinosa, Ammonia beccarii, Pseudorotalia conoides, etc.).
The surface samples show the general trend from a mudflat facies (Trochammina sp., Ammobaculites agglutinans, Eggerelloides scabrum) to upper prodelta shales (Asterorotalia trispinosa, Ammonia beccarii, Pseudorotalia conoides, etc.).
![]() The change in microfaunal distribution is relatively clear. In core (a), the mudflat facies of the upper part overlies a silty facies containing populations of Ammonia beccarii. In the following cores the sandy bar overlies a sandy, silty bioturbated facies containing a more diversified microfauna: Pseudorotalia conoides, Operculina gaymardi, Asterorotalia trispinosa, etc.. In the last core (b), the A. trispinosa shales are superimposed on a silty facies dominated by O. gaymardi and P. conoides.
The change in microfaunal distribution is relatively clear. In core (a), the mudflat facies of the upper part overlies a silty facies containing populations of Ammonia beccarii. In the following cores the sandy bar overlies a sandy, silty bioturbated facies containing a more diversified microfauna: Pseudorotalia conoides, Operculina gaymardi, Asterorotalia trispinosa, etc.. In the last core (b), the A. trispinosa shales are superimposed on a silty facies dominated by O. gaymardi and P. conoides.
![]() This profile of four cores is parallel to the present-day mud
flat (Fig. 12
This profile of four cores is parallel to the present-day mud
flat (Fig. 12 ![]() ). Surface samples along the profile indicate a mudflat facies. Foraminiferal assemblages are dominated by arenaceous forms associated to varying degrees with Ammonia beccarii. The sandy and silty facies present below are mostly dominated by A.
beccarii. However, the most distal core (c) presents a slightly different distribution. The relatively high frequency of Asterorotalia trispinosa and small Elphidium in this core, especially in the muddy facies, is noteworthy.
). Surface samples along the profile indicate a mudflat facies. Foraminiferal assemblages are dominated by arenaceous forms associated to varying degrees with Ammonia beccarii. The sandy and silty facies present below are mostly dominated by A.
beccarii. However, the most distal core (c) presents a slightly different distribution. The relatively high frequency of Asterorotalia trispinosa and small Elphidium in this core, especially in the muddy facies, is noteworthy.
![]() This profile, including four cores, is oriented SW-NE and is located on two of the
present-day sandy mouth bars (Fig. 13
This profile, including four cores, is oriented SW-NE and is located on two of the
present-day sandy mouth bars (Fig. 13 ![]() ). Sediments are mostly sands and silty shales. Ammonia beccarii is by far the most abundant microfossil.
). Sediments are mostly sands and silty shales. Ammonia beccarii is by far the most abundant microfossil.
![]() The micropaleontological study of the cores confirms the general trend inferred from analysis of the surface samples. It supports a strong relationship between the occurrence of Ammonia beccarii and an influx of sand and a high level of correlation between muddy facies and the occurrence of small species of Elphidium (associated with Asterorotalia trispinosa).
The micropaleontological study of the cores confirms the general trend inferred from analysis of the surface samples. It supports a strong relationship between the occurrence of Ammonia beccarii and an influx of sand and a high level of correlation between muddy facies and the occurrence of small species of Elphidium (associated with Asterorotalia trispinosa).
![]() An important result is the appreciation of the rate of sedimentation. As the average depth of the cores is five meters, we may estimate the advance of the sand body as having been between three and four kilometers during the time that three or four meters of sediment accumulated. Bearing in mind that 30 to 40 kilometers of new land has formed in 10,000 years, we can estimate that the time required for the full development of the sandy bar was less than 1,000 years, its sediments deposited at a rate of 3 to 4 millimeters per year.
An important result is the appreciation of the rate of sedimentation. As the average depth of the cores is five meters, we may estimate the advance of the sand body as having been between three and four kilometers during the time that three or four meters of sediment accumulated. Bearing in mind that 30 to 40 kilometers of new land has formed in 10,000 years, we can estimate that the time required for the full development of the sandy bar was less than 1,000 years, its sediments deposited at a rate of 3 to 4 millimeters per year.
![]()
![]() It is most important to the petroleum geologist to be able to interpret the relationship between fossil distribution, sedimentological processes and the resulting lithology. Identical lithologies may contain discrete microfaunal associations; a unique association may be found in several lithologies.
It is most important to the petroleum geologist to be able to interpret the relationship between fossil distribution, sedimentological processes and the resulting lithology. Identical lithologies may contain discrete microfaunal associations; a unique association may be found in several lithologies.
![]() In the Mahakam delta, the two main lithologies are sands and compacted mud
("shale"). Their microfaunal content depends on their locations relative to the deltaic
influx (Fig. 14 A
In the Mahakam delta, the two main lithologies are sands and compacted mud
("shale"). Their microfaunal content depends on their locations relative to the deltaic
influx (Fig. 14 A ![]() , 14 B
, 14 B ![]() ). We have seen earlier the importance of allochthonous factors (tide and current) on the areal distribution of faunas especially in the northern part of the delta.
). We have seen earlier the importance of allochthonous factors (tide and current) on the areal distribution of faunas especially in the northern part of the delta.
Pyritic organic muds more or less rich in wood remains
![]() This type of sediment is found only in the delta itself.
This type of sediment is found only in the delta itself.
![]() In the southern and eastern part, of the delta the association present in the organic mudflats is composed of the Trochammina group, Ammobaculites agglutinans, Eggerelloides scabrum, Ammonia beccarii (6 chambers).
In the southern and eastern part, of the delta the association present in the organic mudflats is composed of the Trochammina group, Ammobaculites agglutinans, Eggerelloides scabrum, Ammonia beccarii (6 chambers).
![]() In the northern part of the delta faunal content content varies with locality:
In the northern part of the delta faunal content content varies with locality:
Sands
![]() Sandy facies are found in the delta itself (distributary channels) and on the
delta front (channels and bars).
Sandy facies are found in the delta itself (distributary channels) and on the
delta front (channels and bars).
Shales, clays
![]() These sediments are associated with the delta front and upper prodelta. The dominant species is Asterorotalia trispinosa everywhere in the delta.
These sediments are associated with the delta front and upper prodelta. The dominant species is Asterorotalia trispinosa everywhere in the delta.
![]() From all these observations it is possible to propose a general model of distribution, particularly for the southern and eastern parts of the delta; the northern part is more specific in nature. This model is now widely used for the interpretation of Upper Neogene deltaic sediments in the Mahakam
basin (Fig. 15
From all these observations it is possible to propose a general model of distribution, particularly for the southern and eastern parts of the delta; the northern part is more specific in nature. This model is now widely used for the interpretation of Upper Neogene deltaic sediments in the Mahakam
basin (Fig. 15 ![]() ).
).
![]() This model presents a tridimensional distribution of micropaleontological associations thus providing an accurate means of determining the nature and location of sand reservoirs and the shaly facies associated with them.
This model presents a tridimensional distribution of micropaleontological associations thus providing an accurate means of determining the nature and location of sand reservoirs and the shaly facies associated with them.
![]()
![]() The distribution of the benthonic microfauna in the deltaic system is controlled by three factors: fluvial input, tidal action and drift currents which may act in parallel or in opposition.
The distribution of the benthonic microfauna in the deltaic system is controlled by three factors: fluvial input, tidal action and drift currents which may act in parallel or in opposition.
![]() Fluvial input, dominant in the center and south, is indicated by the presence of a widely developed delta front, in which faunal suites are relatively well segregated:
Fluvial input, dominant in the center and south, is indicated by the presence of a widely developed delta front, in which faunal suites are relatively well segregated:
![]() The influence of the tide is shown rather spectacularly by the migration up-river of microfaunas from the outer part of the delta front (Asterorotalia), and even from the prodelta (Operculina, Pseudorotalia) along channels located in the inner part of the delta front. This observation is fundamental for the accurate interpretation and use of fossil data, for sand deposits with a calcareous microfauna are often misinterpreted as inner platform deposits.
The influence of the tide is shown rather spectacularly by the migration up-river of microfaunas from the outer part of the delta front (Asterorotalia), and even from the prodelta (Operculina, Pseudorotalia) along channels located in the inner part of the delta front. This observation is fundamental for the accurate interpretation and use of fossil data, for sand deposits with a calcareous microfauna are often misinterpreted as inner platform deposits.
![]() Drift currents are most effective in the northern part of the delta where they reinforce markedly disturbances produced by tides. Together they allowed a veritable migration of microfossils deep into the delta front, some from a considerable distance (Calcarina spengleri (Gmelin) 1791, Calcarina venusta (Brady) 1884, Elphidium "ibericum" (Schrodt) 1932), their habitat a sub-reefal facies to the north. These forms occur far shoreward in delta front deposits. The extreme is shown by the innermost reaches of delta front channels where the contrast is striking between the channel fauna (Asterorotalia, Pseudorotalia, Operculina, Calcarina) and that of the bordering tidal flats (Trochammina, Ammobaculites, Eggerelloides). Although some of the fossils in the channels have undoubtedly been subjected to abrasion (tests rolled, often broken in the northern part of the delta) it is possible that a part of this microfauna found a a refuge in the bottom of the channels where the water was saltier.
Drift currents are most effective in the northern part of the delta where they reinforce markedly disturbances produced by tides. Together they allowed a veritable migration of microfossils deep into the delta front, some from a considerable distance (Calcarina spengleri (Gmelin) 1791, Calcarina venusta (Brady) 1884, Elphidium "ibericum" (Schrodt) 1932), their habitat a sub-reefal facies to the north. These forms occur far shoreward in delta front deposits. The extreme is shown by the innermost reaches of delta front channels where the contrast is striking between the channel fauna (Asterorotalia, Pseudorotalia, Operculina, Calcarina) and that of the bordering tidal flats (Trochammina, Ammobaculites, Eggerelloides). Although some of the fossils in the channels have undoubtedly been subjected to abrasion (tests rolled, often broken in the northern part of the delta) it is possible that a part of this microfauna found a a refuge in the bottom of the channels where the water was saltier.
![]() A series of shallow cores, taken in the southeastern, strongly prograding part of the delta, allowed us to study the reaction of the microfauna to a massive influx of sand. They made possible a reasonably precise map of the effects of the formation of a (massive) sandy bar on the distribution of the fauna. A study of the cores allowed the determination of changes in faunal associations over time. They showed that progradations are both extremely rapid and widespread (the sandy bar covers 25 square kilometers and is several meters thick).
A series of shallow cores, taken in the southeastern, strongly prograding part of the delta, allowed us to study the reaction of the microfauna to a massive influx of sand. They made possible a reasonably precise map of the effects of the formation of a (massive) sandy bar on the distribution of the fauna. A study of the cores allowed the determination of changes in faunal associations over time. They showed that progradations are both extremely rapid and widespread (the sandy bar covers 25 square kilometers and is several meters thick).
![]() The great diversity in the facies encountered forced us to design a relatively complex model of faunal distribution in conformity with the wide diversity in physical and chemical factors that controlled foraminiferal habitats. This model is an important tool for interpreting the Neogene series underlying the
present-day delta. Not only does it allow a better evaluation of small-scale variations in facies (on the order of a few kilometers) but also, because of the complexity of the phenomena observed, reminds us of the limitations of our interpretations.
The great diversity in the facies encountered forced us to design a relatively complex model of faunal distribution in conformity with the wide diversity in physical and chemical factors that controlled foraminiferal habitats. This model is an important tool for interpreting the Neogene series underlying the
present-day delta. Not only does it allow a better evaluation of small-scale variations in facies (on the order of a few kilometers) but also, because of the complexity of the phenomena observed, reminds us of the limitations of our interpretations.
![]() The writer is
grateful to Total Fina Elf for permission to publish on this topic. Many thanks
to the micropaleontologists who have supplied helpful and pertinent information
to the author: Jean-Pierre Debenay, Bruno Granier, Daniel Michoux, Jean-Michel
Villain, Jean-Louis Volat. Drafting and technical support furnished by Cécile
Laporte, Michelle Roux, Bruno Granier, and Nestor J. Sander has been most
appreciated.
The writer is
grateful to Total Fina Elf for permission to publish on this topic. Many thanks
to the micropaleontologists who have supplied helpful and pertinent information
to the author: Jean-Pierre Debenay, Bruno Granier, Daniel Michoux, Jean-Michel
Villain, Jean-Louis Volat. Drafting and technical support furnished by Cécile
Laporte, Michelle Roux, Bruno Granier, and Nestor J. Sander has been most
appreciated.
![]()
Allen G.P. (1974).- Sedimentological study of the Modern Mahakam Delta, preliminary field report.- TOTAL internal report.
Allen G.P. (1976).- Sedimentological study of the Modem Mahakam Delta, preliminary field report, supplementary boring program.- TOTAL internal report.
Allen G.P., Cassoudebat M., Thouvenin J. (1976).- Sedimentological study of the Mahakam Delta, phase II.- TOTAL internal report.
Allen G.P., Laurier D., Thouvenin J. (1979).- Étude sédimentologique du delta de la Mahakam.- TOTAL Compagnie française des Pétroles, Notes et Mémoires, Paris, N° 15, 156 p. (+ 2 maps)
Anderson J.B. (1968).- Ecology of Foraminifera from Mississipi Sound and surrounding waters.- J. Alabama Acad. Sci., (25), 1: pp. 31-34.
Boichard R., Burollet P.F., Lambert B., Villain J.-M (1985).- La plate-forme carbonatée du Pater Noster, Est de Kalimantan (Indonésie), étude sédimentologique et écologique.- TOTAL Compagnie française des Pétroles, Notes et Mémoires, Paris, N° 20, 101 p.
Coleman J.M., Wright L.D. (1975).- Modern river deltas: variability of processes and sand bodies. In: Deltas (Broussard M.L., ed.), Houston Geological Society, pp. 99-149.
Coustillas F. (1983).- Les facies récents de la plate-forme orientale de Kalimantan (Indonésie) et leur contenu micropaléontologique (Foraminifères benthiques).- Thèse doct., Université Bordeaux, 188 p. (+ 1 vol.: "annexes")
Galloway W. E. (1975).- Process framework for describing the morphologic and stratigraphic evolution of deltaic depositional systems. In: Deltas (Broussard M.L., ed.), Houston Geological Society, pp. 87-98.
Granier B., Villain J.-M., Boichard R. (1997).- Biohermes holocènes à Halimeda au large du delta de la Mahakam, Kalimantan (Indonésie)- Le concept de "section condensée dilatée". In: Carbonates intertropicaux. Actes de la Réunion spécialisée de la Société géologique de France du 3 et 4 mai 1993 à la mémoire du Professeur Gabriel Lucas.- Mémoires de la Société géologique de France, Paris, (n. s.), N° 169 (1996), pp. 225-230.
Haman D. (1981).- Modern Textularina (Foraminifera) from the Balize Delta, Louisiana. In: Proceedings of the First Workshop on Arenaceous Foraminifera, 7-9 September 1981. Continental Shelf Institute (Norway), Pub. N° 108, pp. 59-76.
Hoibian T. (1984).- La microfaune benthique traceur de lévolution dun système deltaïque sous climat equatorial : le delta de la Mahakam (Kalimantan).- Thèse doct., Université Bordeaux, 169 p.
Jennings A.E., Nelson A.E. (1992).- Foraminiferal assemblage zones in Oregon tidal marshes - relation to marsh floral zones and sea level.- Journal of Foraminiferal Research, New York, 22, (1), pp. 13-29.
Jonasson K.E., Patterson R.T. (1992).- Preservation potential of salt marsh foraminifera from the Fraser River delta, British Columbia.- Micropaleontology, New York, (38), N° 3, pp. 289-301.
Kruit C. (1955).- Sediments of the Rhône delta: I- Grain size and microfauna.- Verhandelingen van het Koninklijk Nederlands geologisch minjbouwkundig, Geol. series, XV: pp. 357-514.
Lankford R.R. (1959).- Distribution and ecology of Foraminifera from East Mississipi delta margin.- American Association of Petroleum Geologists, Bulletin, Tulsa, Vol. 43, N° 9, pp. 2068-2099.
Oesterle H. (1974).- Foraminiferal facies in the Modem Mahakam Delta.- TOTAL internal report.
Vangerow E.F. (1974).- Récentes observations écologiques des foraminifères dans la zone saumâtre de lembouchure du Rhône.- Revue de Micropaléontologie, Paris, (17), N° 2, pp. 95-106.
![]()
Click on thumbnail to enlarge the image.
Figure 1: Location map of the studied area.
Click on thumbnail to enlarge the image.
Figure 2: Mahakam delta sedimentological features and sampling locations.
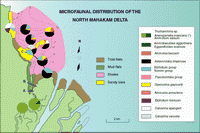
Click on thumbnail to enlarge the image.
Figure 3: North Mahakam delta: microfaunal distribution shows the foraminifera assemblages of three profiles (A, B, C) including channel sections. Note the dominance of Asterorotalia in the muddy delta front facies and the presence of transported calcareous taxa (Calcarina) in the sands of beaches and bars.
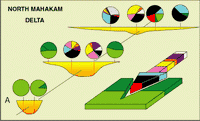
Click on thumbnail to enlarge the image.
Figure 4: North Mahakam delta: lateral distribution of the assemblages across tidal delta and
delta front channels. The main feature is the penetration of calcareous taxa (colored arrow) in the bottom channels (drift current, tide). For colors coding, see Fig.
3
![]() .
.
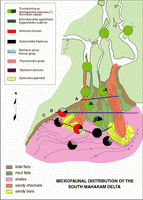
Click on thumbnail to enlarge the image.
Figure 5: South Mahakam delta: microfaunal distribution. In this area, the mud and tidal flats are very well developed. The main result is the wide development of arenaceous biofacies. Asterorotalia and small Elphidium predominate in the muddy delta front facies, the sandy beaches and bars contain populations of Ammonia.
Click on thumbnail to enlarge the image.
Figure 6: South Mahakam delta: lateral distribution of the microfauna across
delta front channels. Note the extent of arenaceous taxa (green arrow) associated with the mudflats advance and in contrast the deep penetration of calcareous taxa like Operculina never seen in the
delta front itself (colored arrow). For colors coding, see Fig.
5 ![]() .
.
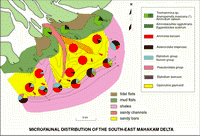
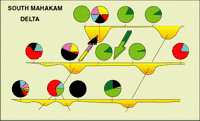
Click on thumbnail to enlarge the image.
Figure 7: South East Mahakam delta: microfaunal distribution. The dominance of Ammonia beccarii is associated with the relative importance of sand bodies (mouth bars) in this area (red arrows indicate the sandy progradation, the green arrow shows the advance of mud flats overlying the sands (in red dashes, an old distributary channel).
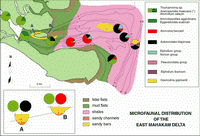
Click on thumbnail to enlarge the image.
Figure 8: East Mahakam delta: microfaunal distribution (interdistributary area). As in the previously studied areas, note the presence (A and B profiles) of calcareous taxa in the channel bottoms.
Click on thumbnail to enlarge the image.
Figure 9: South East Mahakam: delta biofacies distribution. This sketch shows the general distribution of the main Foraminiferal taxa in relation to the topographical and sedimentological pattern.
Click on thumbnail to enlarge the image.
Figure 10: Location of cores (A) and associated surface samples taken in the Tunu mouth bar.
Click on thumbnail to enlarge the image.
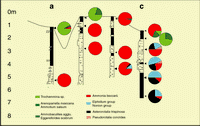
Figure 11: Tunu mouth bar, profile 1. This "dip" profile illustrates the progradational process (for colors coding, see Fig.
9 ![]() ).
).
Click on thumbnail to enlarge the image.
Figure 12: Tunu mouth bar, profile 2. "Strike" profile located along the tidal flats (see Fig.
10 ![]() ). We note the difference in microfaunal composition between the surface mudflats (arenaceous, Ammonia) and
delta front shales (Asterorotalia, Elphidium, Nonion). As do the surface samples, the core sands contain numerous
Ammonia beccarii.
). We note the difference in microfaunal composition between the surface mudflats (arenaceous, Ammonia) and
delta front shales (Asterorotalia, Elphidium, Nonion). As do the surface samples, the core sands contain numerous
Ammonia beccarii.
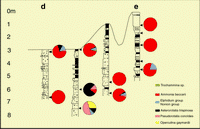
Click on thumbnail to enlarge the image.
Figure 13: Tunu mouth bar, profile 3. This profile represents the sandy mouth bar; Ammonia predominates in the samples However, in the deepest level of the core, the presence of a more diversified biofacies suggests a more distal influence (for colors coding,
see Fig. 12 ![]() ).
).
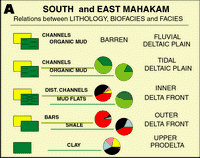
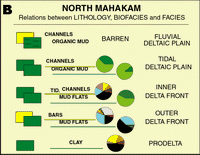
Click on thumbnail to enlarge the image.
Figures 14 A and B: Relations between lithology, biofacies and lithological facies. Depending on their location in the delta, identical lithologies have different associations (especially marked between the North and the other
areas). For colors coding, see Fig. 12 ![]() .
.
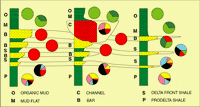
Click on thumbnail to enlarge the image.
Figure 15: Biofacies distribution in the regressive deposits of the Mahakam delta. Most useful is information regarding the association of sandy sediments and various calcareous taxa (including large benthonics like Operculina). In many previous studies this association was interpreted as indicating inner shelf sand deposits. In fact this association indicates
delta front channels
(for colors coding, see Fig. 12 ![]() ).
).
![]()

Click on thumbnail to enlarge the image.
![]() Fig. 1a, 1b: Miliammina fusca (Brady) 1870
Fig. 1a, 1b: Miliammina fusca (Brady) 1870
![]() Fig. 2: Arenoparrella mexicana (Kornfeld) 1931
Fig. 2: Arenoparrella mexicana (Kornfeld) 1931
![]()
![]() Fig. 2a: umbilical view
Fig. 2a: umbilical view
![]()
![]() Fig. 2b: spiral view
Fig. 2b: spiral view
![]() Fig. 3: Trochammina sp.
Fig. 3: Trochammina sp.
![]()
![]() Fig. 3a: umbilical view
Fig. 3a: umbilical view
![]()
![]() Fig. 3b: spiral view
Fig. 3b: spiral view
![]() Fig. 4a, 4b: Ammotium salsum (Cushman & Brönnimann) 1948
Fig. 4a, 4b: Ammotium salsum (Cushman & Brönnimann) 1948
![]() Fig. 5: Eggerelloides scabrum (Williamson) 1858
Fig. 5: Eggerelloides scabrum (Williamson) 1858
![]() Fig. 6: Ammobaculites agglutinans (Orbigny) 1846
Fig. 6: Ammobaculites agglutinans (Orbigny) 1846
![]() Fig. 7: Trochammina globigeriniformis (Parker & Jones) 1865
Fig. 7: Trochammina globigeriniformis (Parker & Jones) 1865
![]()
![]() Fig. 7a: spiral view
Fig. 7a: spiral view
![]()
![]() Fig. 7b: umbilical view
Fig. 7b: umbilical view
![]() Fig. 8: Ammonia beccarii (Linné) 1758 (9 chambers)
Fig. 8: Ammonia beccarii (Linné) 1758 (9 chambers)
![]()
![]() Fig. 8a: spiral view
Fig. 8a: spiral view
![]()
![]() Fig. 8b: umbilical view
Fig. 8b: umbilical view
![]() Fig. 9: Elphidium crispum (Linné) 1767
Fig. 9: Elphidium crispum (Linné) 1767
![]() Fig. 10: Elphidium sp.
Fig. 10: Elphidium sp.
![]() Fig. 11: Elphidium advenum (Cushman) 1922
Fig. 11: Elphidium advenum (Cushman) 1922
![]() Fig. 12: Nonion sp.
Fig. 12: Nonion sp.
![]() Fig. 13: Asterorotalia trispinosa (Thalmann) 1933
Fig. 13: Asterorotalia trispinosa (Thalmann) 1933

Click on thumbnail to enlarge the image.
![]() Fig. 1: Pseudorotalia conoides
(Orbigny) 1826
Fig. 1: Pseudorotalia conoides
(Orbigny) 1826
![]()
![]() Fig. 1a: umbilical view
Fig. 1a: umbilical view
![]()
![]() Fig. 1b: profile view
Fig. 1b: profile view
![]()
![]() Fig. 1c: spiral view
Fig. 1c: spiral view
![]() Fig. 2: Ammonia annectens (Parker & Jones) 1865
Fig. 2: Ammonia annectens (Parker & Jones) 1865
![]()
![]() Fig. 2a: umbilical view
Fig. 2a: umbilical view
![]()
![]() Fig. 2b: profile view
Fig. 2b: profile view
![]()
![]() Fig. 2c: spiral view
Fig. 2c: spiral view
![]() Fig. 3: Operculina complanata (Defrance) 1822
Fig. 3: Operculina complanata (Defrance) 1822
![]() Fig. 4: Calcarina venusta (Brady) 1884
Fig. 4: Calcarina venusta (Brady) 1884
![]()
![]() Fig. 4a: spiral view
Fig. 4a: spiral view
![]()
![]() Fig. 4b: umbilical view
Fig. 4b: umbilical view
![]() Fig. 5: Elphidium aff. craticulatum (Fitchel & Moll) 1798
Fig. 5: Elphidium aff. craticulatum (Fitchel & Moll) 1798
![]() Fig. 6: Calcarina spengleri (Gmelin) 1791
Fig. 6: Calcarina spengleri (Gmelin) 1791
![]()