![]()
![]()
Abstract
Key WordsTaxonomy; Obolus; Ungula; Brachiopoda; Cambrian; Ordovician; East Baltic. CitationPopov L.E., Holmer L.E. (2003).- Understanding linguloid brachiopods: Obolus and Ungula as examples.- Carnets de Géologie / Notebooks on Geology, Maintenon, Article 2003/06 (CG2003_A06) Résumé
Mots-Clefs
|
![]()
![]() The linguloid brachiopod Obolus has been the subject of numerous controversies since it was first established by Eichwald in
1829 (see summaries by Mickwitz, 1896; Gorjansky,
1969; Popov et alii, 1989). Recently, Emig
(2002) used Obolus as an example in a proposal for revising linguloid taxonomy, by limiting the "valid" (sensu Emig,
2002) taxonomic characters to a very restricted number of features (see also Emig,
1982; 1983), declaring that previous studies of Obolus and related forms (e.g. Gorjansky,
1969; Popov et alii, 1989; Holmer and Popov,
2000; Puura, 1996) are based mainly on characters that have "no taxonomic value" (sensu Emig,
2002). The proposal by Emig (2002) is based almost entirely on his experience with fossil and Recent members of a single linguloid Family, the Lingulidae (e.g. Emig,
1982; 1983; Biernat and Emig,
1993). Here we argue that the limited morphological diversity of the taxa found within this family is not a useful basis for working out the taxonomy of the 10 other extinct families now recognized within the very variable and diverse Superfamily Linguloidea (Holmer and Popov,
2000).
The linguloid brachiopod Obolus has been the subject of numerous controversies since it was first established by Eichwald in
1829 (see summaries by Mickwitz, 1896; Gorjansky,
1969; Popov et alii, 1989). Recently, Emig
(2002) used Obolus as an example in a proposal for revising linguloid taxonomy, by limiting the "valid" (sensu Emig,
2002) taxonomic characters to a very restricted number of features (see also Emig,
1982; 1983), declaring that previous studies of Obolus and related forms (e.g. Gorjansky,
1969; Popov et alii, 1989; Holmer and Popov,
2000; Puura, 1996) are based mainly on characters that have "no taxonomic value" (sensu Emig,
2002). The proposal by Emig (2002) is based almost entirely on his experience with fossil and Recent members of a single linguloid Family, the Lingulidae (e.g. Emig,
1982; 1983; Biernat and Emig,
1993). Here we argue that the limited morphological diversity of the taxa found within this family is not a useful basis for working out the taxonomy of the 10 other extinct families now recognized within the very variable and diverse Superfamily Linguloidea (Holmer and Popov,
2000).
![]() The present is obviously the key to the past, but we should not let the present restrict our understanding of the past; the fact is that there has been a drastic loss of diversity within many groups such as the Recent linguloid brachiopods. Furthermore, some of the most important new contributions to the understanding of the taxonomy and phylogeny of the Superfamily Linguloidea and related Lingulate taxa come from detailed studies of the shell structure and micro-ornamentation (Recent and fossil), which are proving to be the most valid characters for taxonomic purposes
(e.g. Cusack et alii, 1999; Holmer,
1989, 2001; Williams,
2003; Williams and Cusack, 1999; Williams et alii,
1994, 1997, 2000).
The present is obviously the key to the past, but we should not let the present restrict our understanding of the past; the fact is that there has been a drastic loss of diversity within many groups such as the Recent linguloid brachiopods. Furthermore, some of the most important new contributions to the understanding of the taxonomy and phylogeny of the Superfamily Linguloidea and related Lingulate taxa come from detailed studies of the shell structure and micro-ornamentation (Recent and fossil), which are proving to be the most valid characters for taxonomic purposes
(e.g. Cusack et alii, 1999; Holmer,
1989, 2001; Williams,
2003; Williams and Cusack, 1999; Williams et alii,
1994, 1997, 2000).
![]() Emig
(2002) does not take into account these new developments in the understanding of the ultrastructure of the Lingulate shell and its phylogenetic importance. When dealing with fossil taxa it is also essential to understand something of the geological setting in which the taxa are found, and in particular the stratigraphic context must be considered. Emig
(2002) does not so the geological associations of Obolus and Ungula are summarised below.
Emig
(2002) does not take into account these new developments in the understanding of the ultrastructure of the Lingulate shell and its phylogenetic importance. When dealing with fossil taxa it is also essential to understand something of the geological setting in which the taxa are found, and in particular the stratigraphic context must be considered. Emig
(2002) does not so the geological associations of Obolus and Ungula are summarised below.
![]()
![]() The type species of Obolus, O. apollinis Eichwald, is from a well known locality on the Luga River near the town of Kingisepp (formerly
Jamburg, St. Petersburg or "Leningrad"
District, Russia), where it occurs in several exposures of the so-called "Obolus sands" in the lower Tosna Formation (uppermost Upper Cambrian: Cordylodus proavus - lower C. lindstromi conodont
biozones). Fortunately, the type material of Obolus apollinis Eichwald, as well as that of another obolid species described by Eichwald
(1829) as Obolus (= Ungula) ingrica, is preserved in the Department of Historical Geology of St. Petersburg State University (contrary to the statement by Emig,
2002, p. 8), and Popov and Khazanovitch (in Popov et alii,
1989) selected lectotypes for both species. Mickwitz
(1896) monographed the brachiopods from the "Obolus sands" and erroneously synonymised these two species without checking the type material; this practice was also adopted by most subsequent researchers (e.g. Walcott,
1912; Bulman, 1939; Rowell,
1965, etc.) until Gorjansky (1969) demonstrated the validity of Obolus apollinis and Ungula ingrica as discrete entities.
The type species of Obolus, O. apollinis Eichwald, is from a well known locality on the Luga River near the town of Kingisepp (formerly
Jamburg, St. Petersburg or "Leningrad"
District, Russia), where it occurs in several exposures of the so-called "Obolus sands" in the lower Tosna Formation (uppermost Upper Cambrian: Cordylodus proavus - lower C. lindstromi conodont
biozones). Fortunately, the type material of Obolus apollinis Eichwald, as well as that of another obolid species described by Eichwald
(1829) as Obolus (= Ungula) ingrica, is preserved in the Department of Historical Geology of St. Petersburg State University (contrary to the statement by Emig,
2002, p. 8), and Popov and Khazanovitch (in Popov et alii,
1989) selected lectotypes for both species. Mickwitz
(1896) monographed the brachiopods from the "Obolus sands" and erroneously synonymised these two species without checking the type material; this practice was also adopted by most subsequent researchers (e.g. Walcott,
1912; Bulman, 1939; Rowell,
1965, etc.) until Gorjansky (1969) demonstrated the validity of Obolus apollinis and Ungula ingrica as discrete entities.
![]() The monographic study of the brachiopods from the "Obolus sands" by Popov and Khazanovitch (in Popov et alii,
1989) was based on a total of more than
10,000 specimens (from more than 100 localities including core material) with carefully measured biometric data that further demonstrated that Obolus apollinis and Ungula ingrica are disparate in both space and time. At the few localities where Obolus apollinis and Ungula ingrica occur together, the valves of the former species show clear signs of re-deposition (Popov et alii,
1989).
The monographic study of the brachiopods from the "Obolus sands" by Popov and Khazanovitch (in Popov et alii,
1989) was based on a total of more than
10,000 specimens (from more than 100 localities including core material) with carefully measured biometric data that further demonstrated that Obolus apollinis and Ungula ingrica are disparate in both space and time. At the few localities where Obolus apollinis and Ungula ingrica occur together, the valves of the former species show clear signs of re-deposition (Popov et alii,
1989).
![]() Popov and Khazanovitch (in Popov et alii,
1989) resolved the long-standing problems connected with several species of Ungula described by Pander
(1830) from the southern outskirts of St Petersburg. The original Pander collection was lost possibly at the end of the nineteenth century (for details see Jaanusson and Bassett,
1993). However, precise information on the geology of the type area (provided by Pander) made it possible to determine that the type locality of these species is on the Izhora River (near the
abandoned village of
Samsonovka). It is clear that Pander's species are distinct and valid (see Gorjansky,
1969; Popov et alii, 1989,
for details). Of these, Obolus transversus (Pander,
1830), occurs in the Rebrovo Member of the Sablinka Formation
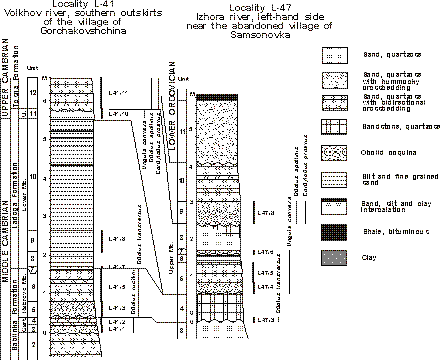
(uppermost Middle Cambrian, Fig. 2
Popov and Khazanovitch (in Popov et alii,
1989) resolved the long-standing problems connected with several species of Ungula described by Pander
(1830) from the southern outskirts of St Petersburg. The original Pander collection was lost possibly at the end of the nineteenth century (for details see Jaanusson and Bassett,
1993). However, precise information on the geology of the type area (provided by Pander) made it possible to determine that the type locality of these species is on the Izhora River (near the
abandoned village of
Samsonovka). It is clear that Pander's species are distinct and valid (see Gorjansky,
1969; Popov et alii, 1989,
for details). Of these, Obolus transversus (Pander,
1830), occurs in the Rebrovo Member of the Sablinka Formation
(uppermost Middle Cambrian, Fig. 2
![]() ), whereas Ungula convexa Pander,
1830, is from the upper member of the Ladoga Formation (Upper Cambrian), where it occurs in association with moderately diverse acritarch and conodont assemblages that date the beds as being within the interval of the Leptoplastus and lower Parabolina spinulosa Biozones of the Scandinavian trilobite standard (Martinsson,
1974). Ungula convexa is the type of Ungula Pander,
1830, which as demonstrated by Popov and Khazanovitch (in Popov et alii,
1989) is different from Obolus (see further below).
), whereas Ungula convexa Pander,
1830, is from the upper member of the Ladoga Formation (Upper Cambrian), where it occurs in association with moderately diverse acritarch and conodont assemblages that date the beds as being within the interval of the Leptoplastus and lower Parabolina spinulosa Biozones of the Scandinavian trilobite standard (Martinsson,
1974). Ungula convexa is the type of Ungula Pander,
1830, which as demonstrated by Popov and Khazanovitch (in Popov et alii,
1989) is different from Obolus (see further below).
![]() Species of Obolus are generally associated with the Skolithos ichnofacies and show a preference for well-aerated shallow marine environments, possibly, with low primary biological productivity. Its relatively thin shells were not resistant to transportation and re-deposition and it is likely that coquinas with the best preserved shells from the Kingisepp quarry, Luga, Lava and Suma rivers were deposited in beach ridges above average sea-level during strong storm events (Artyushkov et alii,
2000).
Species of Obolus are generally associated with the Skolithos ichnofacies and show a preference for well-aerated shallow marine environments, possibly, with low primary biological productivity. Its relatively thin shells were not resistant to transportation and re-deposition and it is likely that coquinas with the best preserved shells from the Kingisepp quarry, Luga, Lava and Suma rivers were deposited in beach ridges above average sea-level during strong storm events (Artyushkov et alii,
2000).
![]() Unlike Obolus, the species of Ungula do not co-occur with Skolithos ichnofacies, and the shells of Ungula are invariably enriched in sulfides and are secondarily phosphatized, which may suggest fossilization in a dysaerobic environment. The host rock often contains thin layers of black bituminous argillite. All these factors suggest that Ungula was adapted to life in shallow waters, characterized by high primary biological productivity, strong daily fluctuations in the content of oxygen, and possibly, periodical eutrophication. The phosphatized shells of Ungula were resistant to re-deposition and they constitute a significant part of the deposits that have been mined commercially in North Estonia (Popov et alii,
1989).
Unlike Obolus, the species of Ungula do not co-occur with Skolithos ichnofacies, and the shells of Ungula are invariably enriched in sulfides and are secondarily phosphatized, which may suggest fossilization in a dysaerobic environment. The host rock often contains thin layers of black bituminous argillite. All these factors suggest that Ungula was adapted to life in shallow waters, characterized by high primary biological productivity, strong daily fluctuations in the content of oxygen, and possibly, periodical eutrophication. The phosphatized shells of Ungula were resistant to re-deposition and they constitute a significant part of the deposits that have been mined commercially in North Estonia (Popov et alii,
1989).
![]() Obolus and Ungula are currently regarded as endemic Baltoscandian taxa, each represented by three successive species (Kaljo et alii,
1986; Popov et alii, 1989). Obolus is represented by two successive Middle Cambrian species, Obolus ruchini Khazanovitch
et Popov,
1984 and Obolus transversus (Pander,
1830)
(Fig. 2
Obolus and Ungula are currently regarded as endemic Baltoscandian taxa, each represented by three successive species (Kaljo et alii,
1986; Popov et alii, 1989). Obolus is represented by two successive Middle Cambrian species, Obolus ruchini Khazanovitch
et Popov,
1984 and Obolus transversus (Pander,
1830)
(Fig. 2
![]() ). No species of Obolus
occur in the lower and medial portions of the Upper Cambrian, but Obolus apollinis Eichwald then appears in the Cordylodus proavus Biozone and ranges into the lower Tremadoc Cordylodus angulatus Biozone. In contrast, Ungula is confined entirely to the Upper Cambrian where it is represented by Ungula inornata (Mickwitz,
1896), Ungula convexa (Pander, 1830) and Ungula ingrica (Eichwald,
1829). The stratigraphical ranges of the most important of these species
are illustrated by two sections in the St Petersburg region
(Fig. 2
). No species of Obolus
occur in the lower and medial portions of the Upper Cambrian, but Obolus apollinis Eichwald then appears in the Cordylodus proavus Biozone and ranges into the lower Tremadoc Cordylodus angulatus Biozone. In contrast, Ungula is confined entirely to the Upper Cambrian where it is represented by Ungula inornata (Mickwitz,
1896), Ungula convexa (Pander, 1830) and Ungula ingrica (Eichwald,
1829). The stratigraphical ranges of the most important of these species
are illustrated by two sections in the St Petersburg region
(Fig. 2
![]() ; see Popov et alii,
1989 for a full account of the brachiopod biostratigraphy). A more detailed summary in English of the lithostratigraphy of the Middle Cambrian to Lower Ordovician deposits and associated faunas in the East Baltic is given by Kaljo et alii
(1986).
; see Popov et alii,
1989 for a full account of the brachiopod biostratigraphy). A more detailed summary in English of the lithostratigraphy of the Middle Cambrian to Lower Ordovician deposits and associated faunas in the East Baltic is given by Kaljo et alii
(1986).
![]()
![]() According to Emig
(2002, p. 2) "the main taxonomic criteria used to discriminate between linguloid taxa were established and figured by Emig
(1982, 1983) and Biernat and Emig
(1993)". He also noted (p. 3) that
"external features as shape, size and dimension ratios of the valves have been demonstrated to have no taxonomic value" although at the same time making an exception for Lingula adamsi with its distinctive subrectangular shell outline. It is important to note that these statements are based only Recent and Mesozoic-Cenozoic
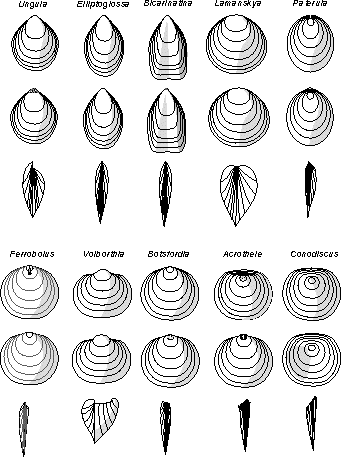
Lingulidae, which do not represent the total morphological diversity of the group (shell shapes, ornament, shell structure, characters of
pseudointerareas, etc.;
Fig. 1
According to Emig
(2002, p. 2) "the main taxonomic criteria used to discriminate between linguloid taxa were established and figured by Emig
(1982, 1983) and Biernat and Emig
(1993)". He also noted (p. 3) that
"external features as shape, size and dimension ratios of the valves have been demonstrated to have no taxonomic value" although at the same time making an exception for Lingula adamsi with its distinctive subrectangular shell outline. It is important to note that these statements are based only Recent and Mesozoic-Cenozoic
Lingulidae, which do not represent the total morphological diversity of the group (shell shapes, ornament, shell structure, characters of
pseudointerareas, etc.;
Fig. 1
![]() ). The post-Palaeozoic members of the
Lingulidae, with their infaunal mode of life in near-shore environments represent only a small fraction of the diversity of shell morphologies and life styles observed in linguloids that lived in Palaeozoic times (Bassett et alii,
1999; Holmer and Popov,
2000). It is not necessary to discuss all aspects of variation in linguloid shell morphologies in this paper, as it is treated in full in the latest edition of the Treatise on Invertebrate Paleontology (Williams,
1997; Holmer and Popov,
2000); however, the wide range of general shell shapes illustrated on
Figure 1
). The post-Palaeozoic members of the
Lingulidae, with their infaunal mode of life in near-shore environments represent only a small fraction of the diversity of shell morphologies and life styles observed in linguloids that lived in Palaeozoic times (Bassett et alii,
1999; Holmer and Popov,
2000). It is not necessary to discuss all aspects of variation in linguloid shell morphologies in this paper, as it is treated in full in the latest edition of the Treatise on Invertebrate Paleontology (Williams,
1997; Holmer and Popov,
2000); however, the wide range of general shell shapes illustrated on
Figure 1
![]() is a good illustration of the diversity exhibited by Palaeozoic
linguloids. It is clear that in many instances, outline, relative length, convexity and proportions of the valves in various linguloid stocks can be used as diagnostic characters. For instance, the strongly dorsibiconvex shell of Volborthia (Fig. 1
is a good illustration of the diversity exhibited by Palaeozoic
linguloids. It is clear that in many instances, outline, relative length, convexity and proportions of the valves in various linguloid stocks can be used as diagnostic characters. For instance, the strongly dorsibiconvex shell of Volborthia (Fig. 1
![]() ) is a good example; it can be identified easily even without knowing details of the micro-ornament or internal morphology (Holmer and Popov,
1995). The dorsibiconvex shell in the genera referred to the Elkaniidae and the Eoobolidae make this feature useful in definition of these families, whereas the families
Lingulasmatidae, Pseudolingulidae and Lingulidae with their specific infaunal mode of life are characterised by exclusively equibiconvex and almost
equivalved, elongated shells. By contrast, the Family Obolidae demonstrates the greatest variability in shell shapes and outlines from one genus to another. For example, there is a distinct tendency towards miniaturisation of the shell in some linguloid lineages (e.g. Elliptoglossinae and
Paterulidae), which would appear to have a most unusual mode of life as far from that of Recent Lingulidae as could be possible. The most extreme example is in Paterula. Shells of this genus range from the Ordovician to the Devonian, invariably demonstrating a very limited variation in the shape of the shell and in its size, which almost never exceeds three millimetres in width. Paterula is often found in sediments enriched in sponge spicules, and the discovery of Silurian sponges with Paterula shells clustering along their oscular margins (Lenz,
1993, fig. 2; brachiopods identified erroneously as Craniops) suggests that the observed association is not a coincidence.
) is a good example; it can be identified easily even without knowing details of the micro-ornament or internal morphology (Holmer and Popov,
1995). The dorsibiconvex shell in the genera referred to the Elkaniidae and the Eoobolidae make this feature useful in definition of these families, whereas the families
Lingulasmatidae, Pseudolingulidae and Lingulidae with their specific infaunal mode of life are characterised by exclusively equibiconvex and almost
equivalved, elongated shells. By contrast, the Family Obolidae demonstrates the greatest variability in shell shapes and outlines from one genus to another. For example, there is a distinct tendency towards miniaturisation of the shell in some linguloid lineages (e.g. Elliptoglossinae and
Paterulidae), which would appear to have a most unusual mode of life as far from that of Recent Lingulidae as could be possible. The most extreme example is in Paterula. Shells of this genus range from the Ordovician to the Devonian, invariably demonstrating a very limited variation in the shape of the shell and in its size, which almost never exceeds three millimetres in width. Paterula is often found in sediments enriched in sponge spicules, and the discovery of Silurian sponges with Paterula shells clustering along their oscular margins (Lenz,
1993, fig. 2; brachiopods identified erroneously as Craniops) suggests that the observed association is not a coincidence.
![]() The umbonal areas and pseudointerareas of Palaeozoic linguloids show a diversity in features, that are either poorly developed or absent in Mesozoic-Cenozoic Lingulidae and consequently are absent in the list of 'Main Taxonomic Characters' presented by Emig
(2002). Some linguloids (e.g. Lingulellotreta) have a pedicle foramen on the ventral pseudointerarea, whereas in others (e.g. Elliptoglossa and the Dysoristidae) the pedicle groove is completely absent (Holmer and Popov,
2000, figs. 25.1e, 34, 36.2d).
The umbonal areas and pseudointerareas of Palaeozoic linguloids show a diversity in features, that are either poorly developed or absent in Mesozoic-Cenozoic Lingulidae and consequently are absent in the list of 'Main Taxonomic Characters' presented by Emig
(2002). Some linguloids (e.g. Lingulellotreta) have a pedicle foramen on the ventral pseudointerarea, whereas in others (e.g. Elliptoglossa and the Dysoristidae) the pedicle groove is completely absent (Holmer and Popov,
2000, figs. 25.1e, 34, 36.2d).
![]() Thus most of the characters proposed by Emig
(2002) are useful within the Family Lingulidae, but are not applicable directly to other linguloid families, or for the Order Lingulida as whole.
Thus most of the characters proposed by Emig
(2002) are useful within the Family Lingulidae, but are not applicable directly to other linguloid families, or for the Order Lingulida as whole.
![]()
![]() As noted by Emig
(2002, p.6), the diagnosis of any particular taxon cannot be based on features that demonstrably lack taxonomic value; Emig's proposed
"New diagnosis", is re-examined below, and for comparison, we also discuss the
"Previous diagnosis" (Holmer and Popov,
2000).
As noted by Emig
(2002, p.6), the diagnosis of any particular taxon cannot be based on features that demonstrably lack taxonomic value; Emig's proposed
"New diagnosis", is re-examined below, and for comparison, we also discuss the
"Previous diagnosis" (Holmer and Popov,
2000).
![]() "Bi-symmetrical muscle arrangement".- The arrangement of muscle scars is obviously a taxonomically valuable feature; however, the symmetrical arrangement of the muscle scars in both valves of Obolidae cannot be a diagnostic character of Obolus (Emig,
2002, p. 3), because it is actually a plesiomorphic character, at least for all members of the Class Lingulata. In contrast, the asymmetrical musculature of the Lingulidae is an important apomorphic character of the Family
Lingulidae, as pointed out by Holmer and Popov
(2000, p. 36).
"Bi-symmetrical muscle arrangement".- The arrangement of muscle scars is obviously a taxonomically valuable feature; however, the symmetrical arrangement of the muscle scars in both valves of Obolidae cannot be a diagnostic character of Obolus (Emig,
2002, p. 3), because it is actually a plesiomorphic character, at least for all members of the Class Lingulata. In contrast, the asymmetrical musculature of the Lingulidae is an important apomorphic character of the Family
Lingulidae, as pointed out by Holmer and Popov
(2000, p. 36).
![]() "Ventral valve:"
"Ventral valve:"
![]() "Triangular umbonal region".- In reality this is a very general feature, which characterises nearly 90 per cent of the Obolidae as well as all the Eoobolidae and the
Lingulellotretidae, and can be demonstrated as a perfect example of a non-diagnostic character, lacking taxonomic value.
"Triangular umbonal region".- In reality this is a very general feature, which characterises nearly 90 per cent of the Obolidae as well as all the Eoobolidae and the
Lingulellotretidae, and can be demonstrated as a perfect example of a non-diagnostic character, lacking taxonomic value.
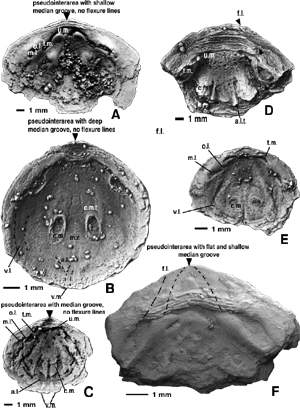
![]() "Pseudointerarea reduced, slightly concave, with elevated flexure lines".- This could be a useful feature for identification of Obolus. However, the ventral pseudointerarea of Obolus is certainly not reduced, and can be said to be of average size
(Fig. 3F
"Pseudointerarea reduced, slightly concave, with elevated flexure lines".- This could be a useful feature for identification of Obolus. However, the ventral pseudointerarea of Obolus is certainly not reduced, and can be said to be of average size
(Fig. 3F ![]() ). The Family Obolidae includes genera with very high
pseudointerareas, e.g. Rebrovia and Lingulepis (Holmer and Popov,
2000, fig. 20.2) and taxa with rudimentary ventral
pseudointerareas, e.g. Apatobolus, Ralfia and Elliptoglossa (Holmer and Popov,
2000, figs. 14.3, 19.1, 25.1).
). The Family Obolidae includes genera with very high
pseudointerareas, e.g. Rebrovia and Lingulepis (Holmer and Popov,
2000, fig. 20.2) and taxa with rudimentary ventral
pseudointerareas, e.g. Apatobolus, Ralfia and Elliptoglossa (Holmer and Popov,
2000, figs. 14.3, 19.1, 25.1).
![]() "Lateral umbonal plates overhanging the internal side".- This is a new term, but in our view the so-called umbonal plates are nothing more than the tracks of the anterior migration of the muscles forming the anterior-lateral muscle fields
(Fig. 3F
"Lateral umbonal plates overhanging the internal side".- This is a new term, but in our view the so-called umbonal plates are nothing more than the tracks of the anterior migration of the muscles forming the anterior-lateral muscle fields
(Fig. 3F ![]() ), and are of little use taxonomically, unless distinct muscle platforms can be recognised
(Fig.
4D
), and are of little use taxonomically, unless distinct muscle platforms can be recognised
(Fig.
4D ![]() ).
).
![]() "Posterior adductor muscle paired, separated by short median septum". Available data suggest (Holmer and Popov,
2000) that the ventral umbonal muscles were paired in all early Palaeozoic
obolides. Therefore this plesiomorphic feature has little value in the discrimination of genera within the
Obolidae.
"Posterior adductor muscle paired, separated by short median septum". Available data suggest (Holmer and Popov,
2000) that the ventral umbonal muscles were paired in all early Palaeozoic
obolides. Therefore this plesiomorphic feature has little value in the discrimination of genera within the
Obolidae.
![]() "Dorsal valve:"
"Dorsal valve:"
![]() "Rounded umbonal region with reduced concave, pseudointerarea with flexure lines".- A rounded umbonal region is present in many Palaeozoic linguloids and is again of little use as a taxonomic character. Moreover, the dorsal pseudointerarea of Obolus does not have any flexure lines. The so-called flexure lines illustrated and discussed by Emig
(2002, figs. 1, 3) are in fact not flexure lines but simply the outer boundaries of the median groove.
"Rounded umbonal region with reduced concave, pseudointerarea with flexure lines".- A rounded umbonal region is present in many Palaeozoic linguloids and is again of little use as a taxonomic character. Moreover, the dorsal pseudointerarea of Obolus does not have any flexure lines. The so-called flexure lines illustrated and discussed by Emig
(2002, figs. 1, 3) are in fact not flexure lines but simply the outer boundaries of the median groove.
![]() "Lateral umbonal plates overhanging the internal side".- See comments above.
"Lateral umbonal plates overhanging the internal side".- See comments above.
![]() "Posterior adductor muscle unpaired".- The expression of the umbonal scar in Obolus is variable in appearance due to taphonomic processes, and although the dorsal umbonal scar in some specimens may appear to be unpaired, other specimens have imprints of a paired dorsal umbonal muscle.
"Posterior adductor muscle unpaired".- The expression of the umbonal scar in Obolus is variable in appearance due to taphonomic processes, and although the dorsal umbonal scar in some specimens may appear to be unpaired, other specimens have imprints of a paired dorsal umbonal muscle.
![]()
![]() "Shell circular to rounded triangular, dorsibiconvex to subequally biconvex".-
It can be demonstrated
(Fig. 1
"Shell circular to rounded triangular, dorsibiconvex to subequally biconvex".-
It can be demonstrated
(Fig. 1
![]() ) that shell shape and convexity are useful diagnostic features in recognizing genera such as Obolus, which is quite different from a number of genera with strongly elongated shells (e.g. Aksarinaia, Anomaloglossa, Leontiella, Leptembolon, Lindinella, Lingulepis, Lingulella, Chakassilingula, Oepikites, Rebrovia, Palaeoglossa, Schmidtites, Spinilingula). Moreover, some obolid genera are nearly equivalved and others like Schmidtites are
ventribiconvex.
) that shell shape and convexity are useful diagnostic features in recognizing genera such as Obolus, which is quite different from a number of genera with strongly elongated shells (e.g. Aksarinaia, Anomaloglossa, Leontiella, Leptembolon, Lindinella, Lingulepis, Lingulella, Chakassilingula, Oepikites, Rebrovia, Palaeoglossa, Schmidtites, Spinilingula). Moreover, some obolid genera are nearly equivalved and others like Schmidtites are
ventribiconvex.
![]() "Ventral propareas with deep, narrow pedicle groove" (Figs. 3F
"Ventral propareas with deep, narrow pedicle groove" (Figs. 3F ![]() , 6D
, 6D ![]() ).- This character is important in distinguishing Obolus from other genera within the
Obolinae; e.g. in Anomaloglossa the pedicle groove is widely triangular (see Holmer and Popov,
2000, p. 40-54, for numerous other examples).
).- This character is important in distinguishing Obolus from other genera within the
Obolinae; e.g. in Anomaloglossa the pedicle groove is widely triangular (see Holmer and Popov,
2000, p. 40-54, for numerous other examples).
![]() "Dorsal pseudointerarea lacking flexure lines".- See above (Fig.
4A-C
"Dorsal pseudointerarea lacking flexure lines".- See above (Fig.
4A-C ![]() ).
).
![]() "Visceral areas of both valves weakly thickened, extending to mid-valve; dorsal median ridge vestigial or absent".- The relative thickness of the visceral area is an important character in, e.g. discriminating the genus from taxa with raised muscle platforms in one or both valves (e.g. Dicellomus, Fordinia, and Ungula). The relative length of the visceral area has also been demonstrated to be an important character by Biernat and Emig
(1993), since it is obviously related directly to the size of the lophophoral cavity. In a similar way, the presence or absence of a dorsal median ridge is an obvious important taxonomic character
(e.g. Leontiella Holmer and Popov,
2000, fig. 16.2a).
"Visceral areas of both valves weakly thickened, extending to mid-valve; dorsal median ridge vestigial or absent".- The relative thickness of the visceral area is an important character in, e.g. discriminating the genus from taxa with raised muscle platforms in one or both valves (e.g. Dicellomus, Fordinia, and Ungula). The relative length of the visceral area has also been demonstrated to be an important character by Biernat and Emig
(1993), since it is obviously related directly to the size of the lophophoral cavity. In a similar way, the presence or absence of a dorsal median ridge is an obvious important taxonomic character
(e.g. Leontiella Holmer and Popov,
2000, fig. 16.2a).
![]() "Vascula lateralia of both valves submarginal, arcuate" (Figs. 3F
"Vascula lateralia of both valves submarginal, arcuate" (Figs. 3F ![]() ,
4A-C
,
4A-C ![]() ).- The shape of the mantle canals within the Obolidae is clearly of taxonomic importance, and some genera have straight proximal ventral vascula lateralia, which can be divergent or subparallel (e.g. Palaeobolus and Dicellomus).
).- The shape of the mantle canals within the Obolidae is clearly of taxonomic importance, and some genera have straight proximal ventral vascula lateralia, which can be divergent or subparallel (e.g. Palaeobolus and Dicellomus).
![]() In addition to the earlier diagnosis by Holmer and Popov
(2000), current studies (work currently in progress) reveal that the first formed shell in Obolus (Fig.
5A-D
In addition to the earlier diagnosis by Holmer and Popov
(2000), current studies (work currently in progress) reveal that the first formed shell in Obolus (Fig.
5A-D ![]() ) is finely pitted.
) is finely pitted.
![]()
![]() The species Obolus apollinis Eichwald; Obolus ruchini Khazanovitch
et Popov and Ungula transversa Pander were synonymised by Emig
(2002). However, his paper contains neither illustrations nor any detailed discussion stating the reasons for the revision. Ungula convexa Pander was also synonymised with Obolus apollinis by the same author, but again without illustrations or detailed discussion. The reasons for retaining these species discrete are summarized below.
The species Obolus apollinis Eichwald; Obolus ruchini Khazanovitch
et Popov and Ungula transversa Pander were synonymised by Emig
(2002). However, his paper contains neither illustrations nor any detailed discussion stating the reasons for the revision. Ungula convexa Pander was also synonymised with Obolus apollinis by the same author, but again without illustrations or detailed discussion. The reasons for retaining these species discrete are summarized below.
![]()
![]() In summary, Emig
(2002) has provided no satisfactory basis for his radical revisions to the existing taxonomy of the Cambrian to earliest Ordovician Obolidae of the East Baltic. Ungula (with U. convexa as type species) represent a separate taxon, which can be distinguished readily from Obolus and other obolid genera. Thus there is no reason to change the type species of the genus as proposed by Emig
(2002, p. 5).
In summary, Emig
(2002) has provided no satisfactory basis for his radical revisions to the existing taxonomy of the Cambrian to earliest Ordovician Obolidae of the East Baltic. Ungula (with U. convexa as type species) represent a separate taxon, which can be distinguished readily from Obolus and other obolid genera. Thus there is no reason to change the type species of the genus as proposed by Emig
(2002, p. 5).
![]() It is also important to point out that notwithstanding Emig's
(2002, p. 8) comments, the type material of the taxa described by Popov et alii
(1989) is accessible in the CNIGR Museum (Sredniy Pr. 74, 199106 St Petersburg, Russia), where all the specimens are catalogued and properly
curated. Moreover, the most important Middle to Upper Cambrian obolid brachiopod localities in the vicinity of St Petersburg, including the type localities of most of the discussed species, are also easily accessible for re-study, and any proposed revision of the taxonomy of the fauna of the "Obolus sands" obviously needs to include a careful re-study of both the type material and the type sections, something which is entirely lacking from the brief study by Emig
(2002).
It is also important to point out that notwithstanding Emig's
(2002, p. 8) comments, the type material of the taxa described by Popov et alii
(1989) is accessible in the CNIGR Museum (Sredniy Pr. 74, 199106 St Petersburg, Russia), where all the specimens are catalogued and properly
curated. Moreover, the most important Middle to Upper Cambrian obolid brachiopod localities in the vicinity of St Petersburg, including the type localities of most of the discussed species, are also easily accessible for re-study, and any proposed revision of the taxonomy of the fauna of the "Obolus sands" obviously needs to include a careful re-study of both the type material and the type sections, something which is entirely lacking from the brief study by Emig
(2002).
![]() We agree wholeheartedly that
"systematics and taxonomy must propose new ideas and test
hypotheses", but these should build upon, and extend previous studies, and in our view the study by Emig
(2002) does not represent "a first step in better understanding in linguloid
taxonomy".
We agree wholeheartedly that
"systematics and taxonomy must propose new ideas and test
hypotheses", but these should build upon, and extend previous studies, and in our view the study by Emig
(2002) does not represent "a first step in better understanding in linguloid
taxonomy".
Acknowledgements
![]() We thank Prof. Michael G. Bassett (National Museum of Wales, Cardiff), Prof. Arthur J. Boucot (Oregon State University, Corvallis),
and Prof. Fernando Alvarez (Universidad de Oviedo, Oviedo) for critical comments on the manuscript.
L.E. Popov's work is financed by fellowships from the Royal Society of London and the National Museum of Wales. Grants from the Swedish Research Council
(VR) have supported L.E. Holmer's work.
We thank Prof. Michael G. Bassett (National Museum of Wales, Cardiff), Prof. Arthur J. Boucot (Oregon State University, Corvallis),
and Prof. Fernando Alvarez (Universidad de Oviedo, Oviedo) for critical comments on the manuscript.
L.E. Popov's work is financed by fellowships from the Royal Society of London and the National Museum of Wales. Grants from the Swedish Research Council
(VR) have supported L.E. Holmer's work.
![]()
Artyushkov E.A., Lindström M., Popov L.E. (2000).- Relative sea-level changes in Baltoscandia in the Cambrian and Early Ordovician: the predominance of tectonic factors and the absence of large scale eustatic fluctuations.- Tectonophysics, Amsterdam, Vol. 320, pp. 375-407.
Bassett M. G., Popov L.E., Holmer L.E. (1999).- Organophosphatic brachiopods: patterns of biodiversification and extinction in the early Palaeozoic.- Géobios, Lyon, Vol. 32(1), pp. 145-163.
Biernat G., Emig C.C. (1993).- Anatomical distinctions of the Mesozoic lingulide brachiopods.- Acta Palaeontologica Polonica, Warsaw, Vol. 38, N° 1/2, pp. 1-20.
Cusack M., Williams A., Buckman, J.O. (1999).- Chemico-structural evolution of linguloid brachiopod shells.- Palaeontology, Oxford, Vol. 42, pp. 799-840.
Bulman O.M. (1939).- Muscle systems of some inarticulate brachiopods.- Geological Magazine, Cambridge, Vol. 76, pp. 434-444.
Eichwald E. (1829).- Zoologia specialis, quam expositis animalibus tum vivis, tum fossilibus potissimum Rossiae in universum et Poloniae in specie, in usum lectionum publicarum in Universitate Caesarea Vilnensi. Volume 1.- Josephi Zawadski, Vilnae, 314 p.
Emig C.C. (1977).- Réflexions sur la taxonomie des espèces du genre Lingula (Brachiopodes, Inarticulés).- Comptes Rendus de l'Académie des Sciences, Paris, Vol. 285, pp. 523-525.
Emig C.C. (1982).- Taxonomie du genre Lingula (Brachiopodes, Inarticulés).- Bulletin du Muséum National d'Histoire Naturelle, Paris, Sér. 4, Vol. 4, N° 3/4, pp. 337-367.
Emig C.C. (1983).- Taxonomie du genre Glottidia (Brachiopodes Inarticulés).- Bulletin du Muséum National d'Histoire Naturelle, Paris, Sér. 4, Vol. 5 (Sect. 4), N° 2, pp. 469-489.
Emig C.C. (2002).- Tools for linguloid taxonomy: the genus Obolus (Brachiopoda) as an example.- Carnets de Géologie - Notebooks on Geology, Maintenon, Article 2002/01 (CG2002_A01), 9 p.
Gorjansky V.Yu. (1969).- Inarticulate brachiopods of the Cambrian and Ordovician of the northwest Russian Platform [in Russian].- Ministerstvo Geologii RSFSR, Severo-Zapadnoe Territorialnoe Geologicheskoe Upravlenie, Leningrad, pp. 1-173.
Holmer L.E. (1989).- Middle Ordovician phosphatic inarticulate brachiopods from Västergötland and Dalarna, Sweden.- Fossils and Strata, Oslo, Vol. 26, pp. 1-172.
Holmer L.E. (2001).- Phylogeny and classification: Linguliformea and Craniiformea.- The paleontological Society Papers, New Haven, Vol. 7, pp. 11-26.
Holmer L.E., Popov L.E. (1995).- The elkaniide brachiopod Volborthia from the Lower Ordovician of Baltoscandia.- Paläontologisches Zeitschrift, Stuttgart, Vol. 69, pp. 211-219.
Holmer L.E., Popov L.E. (2000).- Lingulida. In: Kaesler R.L. (Ed.), Revised Brachiopoda.- Treatise on Invertebrate Paleontology, Geological Society of America, New York, and University of Kansas, Lawrence, Part H, Vol. 2, pp. 32-95.
Jaanusson V., Bassett M.G. (1993): Orthambonites and related Ordovician brachiopod genera.- Palaeontology, Oxford, Vol. 36, pp. 21-63.
Kaljo D., Borovko N., Heinsalu H., Khazanovitch K., Mens K., Popov L., Sergeyeva S., Sobolevskaya R., Viira V. (1986).- The Cambrian-Ordovician boundary in the Baltic-Ladoga clint area.- Eesti NSV Teaduste Akadeemia Toimetised. Geoloogia, Tallinn, Vol. 35, pp. 97-108.
Khazanovitch K.K., Popov L.E., In: Khazanovitch K.K., Popov L.E., Melnikova L.M. (1984).- Inarticulate brachiopods, ostracodes (bradoriids) and hyolithelminths from the Sablinka Formation of the Leningrad District [in Russian].- Paleontologicheskiy Zhurnal, Moscow, Vol. 1984, N° 4, pp. 33-47.
Lenz A.C. (1993).- A Silurian sponge-inarticulate brachiopod life? association.- Journal of Palaeontology, Lawrence, Vol. 67, pp. 138-139.
Martinsson A. (1974).- The Cambrian of Norden.- Lower Palaeozoic Rocks of the World, John Wiley and Sons, New York, Vol. 2, pp. 185-283.
Mickwitz A. (1896).- Über die Brachiopodengattung Obolus Eichwald.- Mémoires de l'Académie Impériale des Sciences de St Pétersbourg, Classe physicomathématique, St-Pétersbourg, Vol. 4, N° 2, pp. 1-216.
Pander C.H. (1830).- Beiträge zur Geognosie des Russischen Reiches.- St. Petersburg, 165 pp.
Popov L., Khazanovitch K.K., Borovko N.G., Sergeyeva S.P., Sobolevskaya R.F. (1989).- The Key sections and stratigraphy of the phosphate bearing Obolus beds of the North-East of the Russian platform [in Russian].- Nauka Trudy, Leningrad, Vol. 18, pp. 1-222.
Puura I. (1996).- Lingulate brachiopods and biostratigraphy of the Cambrian-Ordovician boundary beds in Baltoscandia.- Dr. Thesis, Univ. Uppsala, 136 pp.
Rowell A.J. (1965).- Inarticulata. In: Moore R.C. (Ed.), Brachiopoda.- Treatise on Invertebrate Paleontology, Geological Society of America, New York, and University of Kansas, Lawrence, Part H, pp. 260-296.
Walcott C.D. (1912).- Cambrian Brachiopoda.- Monographs of the United States Geological Survey, Washington, Vol. 51, pp. 812.
Williams A. (2003).- Microscopic imprints of the juvenile shells of Palaeozoic linguliform brachiopods.- Palaeontology, Oxford, Vol. 46, pp. 67-92.
Williams A., Cohen B.L., Cusack M., Long S. (2000).- Provenance of Atlantic lingulid brachiopods.- Palaeontology, Oxford, Vol. 43, pp. 999-1018.
Williams A., Cusack M. (1999).- Evolution of a rhythmic lamination in the organophosphatic shells of brachiopods.- Journal of Structural Biology, Amsterdam, Vol. 126, pp. 227-240.
Williams A., Cusack M., Buckman J. O. (1998).- Chemico-structural phylogeny of the discinoid brachiopod shell.- Philosophical Transactions of the Royal Society of London, London, Series B, 353, pp. 2005-2038.
Williams A., Cusack M., Mackay S. (1994).- Collagenous chitinophosphatic shell of the brachiopod Lingula.- Philosophical Transactions of the Royal Society of London, London, Series B, 346, pp. 223-266.
Williams A. et alii (1997).- In: Kaesler R.L. (Ed.), Revised Brachiopoda.- Treatise on Invertebrate Paleontology, Geological Society of America, New York, and University of Kansas, Lawrence, Part H, Vol. 1, 539 pp.
![]()
Click on thumbnail to enlarge the image.
Figure 1: Examples of variations in shell outline and relative valve convexity in selected Palaeozoic lingulide genera.
Click on thumbnail to enlarge the image.
Figure 2: Columnar sections through the Cambrian-Ordovician boundary beds in the East Baltic at the Volkhov and Izhora rivers (see Popov et alii, 1989, for details) showing of distribution of discussed relevant species.

Click on thumbnail to enlarge the image.
Figure 3: A-E, variations in the external shape of the dorsal valve and ornament in species of Obolus and Ungula. A, Ungula convexa (Pander, 1830), NMW 2001.39G.4; Upper Cambrian, Ladoga Formation, locality B-2, Unit C2 (Popov et alii, 1989, fig. 3), on the right (facing downstream) bank of the Izhora River near the abandoned village of Samsonovka, St Petersburg District; note smooth shell with few growth rings and lack of median sulcus. B, Ungula inornata (Mickwitz, 1896), CNIGR 160/12348; Upper Cambrian, upper Ulgase Formation, Sample E60/3 (Popov et alii, 1989, fig. 10), outskirts of the village of Kallavere, North Estonia; note weak umbonal sulcus. C, Obolus transversus (Pander, 1830), NMW 2001.39G.51; Middle Cambrian, Sablinka Formation, Rebrovo Member, Sample L-41/3 (Popov et alii, 1989, fig. 2), right-hand side of Volkhov River in southern outskirts of the village of Gorchakovshchina, St Petersburg District; note lack of median sulcus. D, Obolus ruchini Khazanovitch et Popov, 1984; CNIGR 16/12348; Middle Cambrian, Sablinka Formation, Gertovo Member, Sample L6/1 (Popov et alii, 1989, fig. 3), right bank of the Sarya River about 700 m downstream from the northern outskirts of the village of Voibokalo; note the very shallow median sulcus, which can be traced in nearly all dorsal valves of Obolus from Gertovo Member. E, Obolus apollinis Eichwald, 1829; NMW 2001.39G.119; Upper Cambrian, Suurjogi Member, sample 1653/25 (Popov et alii, 1989, fig. 1), borehole no. 1653 south east of Rakvere, North Estonia; note lack of sulcus. F, View of ventral valve interior of Obolus apollinis Eichwald, 1829; CNIGR 2/12348; Upper Cambrian, lower Tosna Formation, sample L1/3 (Popov et alii, 1989, fig. 3), right-hand side of lava River in southern outskirts of the village of Gorodishche; note well developed ventral pseudointerarea with narrow pedicle groove and deep flexure lines (f.l.), pair of weak diverging furrows of pedicle nerve impression (p.n.i.) flanked by paired impression of umbonal muscle scar (u.m.), posterolateral muscle fields bearing attachment scars of transmedian (t.m.) and anterior lateral (a.l.) muscles; paired anterior muscle fields formed by combined attachment scars of central, middle lateral and anterior lateral muscles (a.m.f.) and weakly impressed marginal vascula lateralia (v.l.); depressions under the muscle scars clearly formed as a result of post-mortem degradation of the organic shell matter and subsequent collapse of phosphatic layers.
Click on thumbnail to enlarge the image.
Figure 4: Main features of dorsal valve interior of Obolus and Ungula. A, Obolus transversus (Pander, 1830), CNIGR 22/12348; Middle Cambrian, Sablinka Formation, Rebrovo Member, Sample L-41/3 (Popov et alii, 1989, fig. 2), right-hand side of Volkhov River in southern outskirts of the village of Gorchakovshchina, St Petersburg District; note that mature specimen has low dorsal pseudointerarea occupied mainly weakly defined pedicle groove and lacks flexure lines, with paired umbonal muscle scars (u. m.) divided by short median ridge and posterolateral muscle fields slightly raised above the valve floor with attachment scars of transmedian (t.m.), oblique lateral (o.l.) and middle lateral (m.l.) muscles. B, Obolus apollinis Eichwald, 1829; IGT, B1706; Upper Cambrian, lower Tosna Formation, sample L1/3 (Popov et alii, 1989, fig. 3), right-hand side of lava River in southern outskirts of the village of Gorodishche, St Petersburg District; note narrow dorsal pseudointerarea, mainly occupied by median groove, very weak median ridge (m.r.) in anterior half of the visceral area, central muscle scars (c.m.) on the end of distinct tracks (c.m.t.) slightly raised above valve floor, but not forming platforms, anterior lateral muscle scars (a.l.) near the margins of the visceral area, proximal ends of weakly impressed, subparallel vascula media (v.m.) and submarginal, arcuate vascula lateralia (v.l.). C, Obolus ruchini Khazanovitch et Popov, 1984; CNIGR 13/12348; Middle Cambrian, Sablinka Formation, Gertovo Member, Sample L6/1 (Popov et alii, 1989, fig. 3), right-hand side of Sarya River about 700 m downstream from the northern outskirts of the village of Voibokalo, St Petersburg District; note muscle scars and impressions of mantle canals. D-F, Ungula convexa Pander, 1830, Upper Cambrian, Ladoga Formation; D, CNIGR 126/12348, sample L9/31b (Popov et alii, 1989, fig. 4), Naziya River, left-hand side near the abandoned village of Novaya, St Petersburg District; mature specimen (cf. fig. 4A-B) with high pseudointerarea bearing broad flattened median groove separated from flexure lines (f.l.) on propareas, paired umbonal muscle scars and transmedian muscle scars on raised platforms; distinct, raised tracks of central (c.m.t.) and anterior lateral muscles (a.l.t.) in the median part of the shell represent a distinctive pattern markedly different from Obolus; E, CNIGR 129/12348; sample L47/4 (Popov et alii, 1989, fig. 5), Izhora River, left-hand side, about 800 m upstream of the locality B2; juvenile specimens with raised muscle tracks and platforms not yet formed, but with well impressed muscle scars and mantle canals; F, NMW 2001.39G.7; locality B-2, Unit C2 (Popov et alii, 1989, fig. 3), right hand side of Izhora River near the abandoned village of Samsonovka, St Petersburg District; showing distinction between margins of median groove and position of flexure lines on dorsal pseudointerarea.

Click on thumbnail to enlarge the image.
Figure 5: A-B, Obolus ruchini Khazanovitch et Popov, 1984; NMW 2001.102, dorsal valve; Middle Cambrian, Sablinka Formation, Gertovo Member, Sample L-40/5 (Popov et alii, 1989, fig. 2), right-hand side of Volkhov River in southern outskirts of the village of Gorchakovshchina, St Petersburg District; note that the slightly exfoliated surface of the dorsal valve still preserves remnants of a finely pitted juvenile ("larval") shell, about 1.5 mm cross. C-D, Obolus apollinis Eichwald, 1829; NMW 2001,39G.114, ventral valve; Upper Cambrian, lower Tosna Formation, Cordylodus proavus conodont biozone, sample 782/3, quarry on right-hand side of Tosna River downstream from the Sablino caves, St Petersburg District; note well preserved semi-cylindroid and hemispherical pits (pt.) on the juvenile ("larval") shell (D). E-F, Ungula sp.; NMW 2001.39G.109, ventral valve; Middle-Upper Cambrian, lower Ladoga Formation, sample L17/9 (Popov et alii, 1989, fig. 2), Syas River, right-hand side, 200 m upstream of the village of Rebrovo, St Petersburg District; note well preserved smooth juvenile ("larval") shell.

Click on thumbnail to enlarge the image.
Figure 6: SEM microphotographs of fractured shells of Ungula and Obolus showing basic differences in microstructure under posterolateral muscle fields of the ventral valve (all abbreviations, are the same as on Figs. 3-5, unless otherwise stated). A-C, Ungula convexa (Pander, 1830), NMW 2001.39G.5, ventral valve; Upper Cambrian, Ladoga Formation, locality B-2, Unit C2 (Popov et alii, 1989, fig. 3), right hand side of Izhora River near the abandoned village of Samsonovka, St Petersburg District; A oblique lateral view showing pedicle grove (p.g.) flexure lines and muscle scars; B, symmetrical baculate set showing compactly placed baculi usually exceeding 1 µm in diameter; C, general view through the broken shell showing alternating baculate sets (b.s.), thick compact laminae (c.l.) and membranous laminae (m.l.). D-F, Obolus apollinis Eichwald, 1829, NMW 2001.39G.109, ventral valve; Upper Cambrian, Tosna Formation, Cordylodus proavus conodont biozone, sample L47/8 (Popov et alii, 1989, fig. 5), Izhora River, left-hand side, about 800 m upstream of the locality B2; D, oblique lateral view showing pedicle grove, flexure lines and muscle scars; E, general view of baculate set showing relation with compact lamina; F, general view of the shell fracture showing alternating symmetrical baculate sets with loosely spaced inclined baculi and thin compact laminae; note that sizes of major structural elements (e.g. size of baculi and compact laminae) in the Obolus shell are about one order of magnitude smaller than in Ungula.
![]()

{kind=link}