![]()
![]()
![]() Lingula is often considered a "living-fossil" based on its supposed lengthy morphological conservatism
owing to its absence of evolution, and its remarkable survival for more than 550 M.Y. This conclusion is based on the typical apparently unchanged
"linguliform" shape of the shell. However the taxa of the family Lingulidae show morphological evolutionary changes despite the fact that the group appears panchronic among the Recent Brachiopoda. Consequently, traditional opinion that Lingula is a "living-fossil" should be rejected. Diagnoses of the Family Lingulidae and of its three genera are herewith emended.
Lingula is often considered a "living-fossil" based on its supposed lengthy morphological conservatism
owing to its absence of evolution, and its remarkable survival for more than 550 M.Y. This conclusion is based on the typical apparently unchanged
"linguliform" shape of the shell. However the taxa of the family Lingulidae show morphological evolutionary changes despite the fact that the group appears panchronic among the Recent Brachiopoda. Consequently, traditional opinion that Lingula is a "living-fossil" should be rejected. Diagnoses of the Family Lingulidae and of its three genera are herewith emended.
Living-fossil; Lingula; Lingulidae; Brachiopoda; taxonomy.
Emig C. (2003).- Proof that Lingula (Brachiopoda) is not a living-fossil, and emended diagnoses of the Family Lingulidae.- Carnets de Géologie / Notebooks on Geology, Maintenon, Letter 2003/01 (CG2003_L01)
![]() Preuves que Lingula (Brachiopoda) n'est pas un fossile vivant, avec de nouvelles diagnoses pour les taxons de la Famille
des Lingulidae.- Lingula est souvent considérée comme un fossile-vivant, voire le plus ancien actuellement
connu, à cause de son long conservatisme morphologique basé sur une supposée absence d’évolution,
ainsi qu'en raison de sa remarquable survie depuis plus de 550 MA. Cette
assertion est basée sur une forme inchangée de la coquille, dite
"linguliforme". Cependant, les taxons de la famille des Lingulidae montrent des changements évolutifs de la morphologie et de l’anatomie interne bien que ce groupe puisse être considéré comme panchronique au sein des Brachiopoda actuels. Il est démontré que l’opinion
traditionnellement véhiculée selon laquelle les Lingula
seraient des fossiles vivants doit être rejetée. En conséquence, de nouvelles diagnoses sont proposées
pour la famille des Lingulidae et pour les trois genres pouvant s’y rapporter avec certitude.
Preuves que Lingula (Brachiopoda) n'est pas un fossile vivant, avec de nouvelles diagnoses pour les taxons de la Famille
des Lingulidae.- Lingula est souvent considérée comme un fossile-vivant, voire le plus ancien actuellement
connu, à cause de son long conservatisme morphologique basé sur une supposée absence d’évolution,
ainsi qu'en raison de sa remarquable survie depuis plus de 550 MA. Cette
assertion est basée sur une forme inchangée de la coquille, dite
"linguliforme". Cependant, les taxons de la famille des Lingulidae montrent des changements évolutifs de la morphologie et de l’anatomie interne bien que ce groupe puisse être considéré comme panchronique au sein des Brachiopoda actuels. Il est démontré que l’opinion
traditionnellement véhiculée selon laquelle les Lingula
seraient des fossiles vivants doit être rejetée. En conséquence, de nouvelles diagnoses sont proposées
pour la famille des Lingulidae et pour les trois genres pouvant s’y rapporter avec certitude.
![]() Fossile vivant ; Lingula ; Lingulidae ; Brachiopoda ; taxinomie.
Fossile vivant ; Lingula ; Lingulidae ; Brachiopoda ; taxinomie.
![]()
![]() Among the Brachiopoda Lingula, belonging to the Family Lingulidae, is often considered one of the most ancient "living-fossils". This assertion is based on the supposed morphological conservatism of the genus, its lack of morphological change,
i.e. the absence of change in shell shape and structure and of evolution, and its remarkable survival for more than 550 M.Y. All of this because over the years many Palaeozoic species have been referred to Lingula.
Among the Brachiopoda Lingula, belonging to the Family Lingulidae, is often considered one of the most ancient "living-fossils". This assertion is based on the supposed morphological conservatism of the genus, its lack of morphological change,
i.e. the absence of change in shell shape and structure and of evolution, and its remarkable survival for more than 550 M.Y. All of this because over the years many Palaeozoic species have been referred to Lingula.
![]() Arising in the
Early Cambrian, the Superfamily Linguloidea (Brachiopoda,
Lingulata) expanded rapidly from the Medial Cambrian to the beginning of the Ordovician (Babin et alii, 1992; Emig, 1984a). After Devonian times only the Family Lingulidae subsists and it seems to be represented from the Carboniferous to the Cretaceous
only by
Lingularia, and since the Tertiary, or possibly from the Late Cretaceous, by the two extant genera Lingula and Glottidia.
Arising in the
Early Cambrian, the Superfamily Linguloidea (Brachiopoda,
Lingulata) expanded rapidly from the Medial Cambrian to the beginning of the Ordovician (Babin et alii, 1992; Emig, 1984a). After Devonian times only the Family Lingulidae subsists and it seems to be represented from the Carboniferous to the Cretaceous
only by
Lingularia, and since the Tertiary, or possibly from the Late Cretaceous, by the two extant genera Lingula and Glottidia.
![]() Thus, it is reasonable to inquire into the validity of the traditional opinion that Lingula represents a lineage of "living-fossils" that originated during the Cambrian, as well as the idea that Lingula has survived without significant morphological changes since the
Early Palaeozoic.
Thus, it is reasonable to inquire into the validity of the traditional opinion that Lingula represents a lineage of "living-fossils" that originated during the Cambrian, as well as the idea that Lingula has survived without significant morphological changes since the
Early Palaeozoic.
![]()
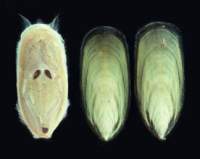
![]() The living-fossil argument rests on a single piece of evidence; the typical shape of the shell - namely
"linguliform" (from the common shape in Lingula; Fig. 1
The living-fossil argument rests on a single piece of evidence; the typical shape of the shell - namely
"linguliform" (from the common shape in Lingula; Fig. 1
![]() , Table 1
, Table 1 ![]() )
- with smooth external faces marked only by growth lines (Emig, 1983a). This form is generally the only
"character" remaining in fossil lingulides that has led many authors to refer specimens
ipso facto to Lingula, regardless of the age of the outcrop, from the
Earliest Cambrian to the present. Nevertheless, the "linguliform" shell indicates only an endobiont life in burrows (Emig, 1982a, 1997a). This mode of existence is very common in the life history of many Palaeozoic genera and families of the Order Lingulida whereas Lingula sensu
stricto appeared only at the beginning of the Tertiary or perhaps in the
Late Cretaceous.
)
- with smooth external faces marked only by growth lines (Emig, 1983a). This form is generally the only
"character" remaining in fossil lingulides that has led many authors to refer specimens
ipso facto to Lingula, regardless of the age of the outcrop, from the
Earliest Cambrian to the present. Nevertheless, the "linguliform" shell indicates only an endobiont life in burrows (Emig, 1982a, 1997a). This mode of existence is very common in the life history of many Palaeozoic genera and families of the Order Lingulida whereas Lingula sensu
stricto appeared only at the beginning of the Tertiary or perhaps in the
Late Cretaceous.
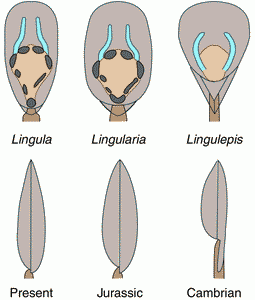
![]() The persistence of this linguliform shell has been and is still interpreted by many palaeontologists as indicating morphological stability through the whole of the Phanerozoic record, although both the shell and its content have evolved since the Palaeozoic along with the phylogenetical characters of the Lingulida (Biernat and Emig, 1993; Emig, 2002).
The persistence of this linguliform shell has been and is still interpreted by many palaeontologists as indicating morphological stability through the whole of the Phanerozoic record, although both the shell and its content have evolved since the Palaeozoic along with the phylogenetical characters of the Lingulida (Biernat and Emig, 1993; Emig, 2002).
![]() If some important Lingulida anatomical features such as the lophophore (Zhang et alii, 2003)
arose before the Cambrian and remained in the Lingulata, others with taxonomic value have evolved since then among the Family Lingulidae:
e.g. the reduction of the pseudointerareas with only vestigial propareas, an asymmetrical disposition of the body muscles, convergent main mantle canals (vascula lateralia) to become subparallel anteriorly, the absence of the median secondary mantle canals (vascula media), a single posterior adductor muscle on both ventral and dorsal side
(Fig. 2
If some important Lingulida anatomical features such as the lophophore (Zhang et alii, 2003)
arose before the Cambrian and remained in the Lingulata, others with taxonomic value have evolved since then among the Family Lingulidae:
e.g. the reduction of the pseudointerareas with only vestigial propareas, an asymmetrical disposition of the body muscles, convergent main mantle canals (vascula lateralia) to become subparallel anteriorly, the absence of the median secondary mantle canals (vascula media), a single posterior adductor muscle on both ventral and dorsal side
(Fig. 2
![]() , 3
, 3
![]() ). All these characters are a part of the diagnosis of the Family Lingulidae
(Table 1
). All these characters are a part of the diagnosis of the Family Lingulidae
(Table 1 ![]() ).
).
![]() Lingularia,
Lingula and Glottidia belong unquestionably to this family. Furthermore a number of evolutionary changes listed
below took place in these three genera respectively (Fig. 2
Lingularia,
Lingula and Glottidia belong unquestionably to this family. Furthermore a number of evolutionary changes listed
below took place in these three genera respectively (Fig. 2
![]() , 3
, 3
![]() ; Table 1
; Table 1 ![]() ):
):
![]() These evolutionary changes do not support the view of Recent lingulides as "living-fossils" that have survived and undergone little significant morphological change since the Palaeozoic. On the contrary, no Palaeozoic or Mesozoic species is assignable to Lingula sensu
stricto.
These evolutionary changes do not support the view of Recent lingulides as "living-fossils" that have survived and undergone little significant morphological change since the Palaeozoic. On the contrary, no Palaeozoic or Mesozoic species is assignable to Lingula sensu
stricto.
![]() The current taxonomy revision of the Brachiopoda calls into question other families, not just the lingulides (Biernat and Emig, 1993; Emig, 2002). Until very recently soft parts were rarely considered to have taxonomic value, even in extant taxa where their anatomy is evident. This neglect occurred because skeletal characteristics were considered to be of primary importance by palaeontologists who found that using these
"biological constraints" would make it impossible to describe or to redefine many fossil species. Because morpho-anatomical characters are not always available or are difficult to recognize, it seems best to avoid new descriptions (Emig, 2002). Furthermore when
making a cladistic analysis many systematists forget that all the phylogenetical characters have to be tested and that these criteria must also be applied in descriptions of taxa. Following the traditional path increases confusion among the taxa, and Lingula as well as Obolus are among the most prominent examples, for they have endured for nearly two centuries.
The current taxonomy revision of the Brachiopoda calls into question other families, not just the lingulides (Biernat and Emig, 1993; Emig, 2002). Until very recently soft parts were rarely considered to have taxonomic value, even in extant taxa where their anatomy is evident. This neglect occurred because skeletal characteristics were considered to be of primary importance by palaeontologists who found that using these
"biological constraints" would make it impossible to describe or to redefine many fossil species. Because morpho-anatomical characters are not always available or are difficult to recognize, it seems best to avoid new descriptions (Emig, 2002). Furthermore when
making a cladistic analysis many systematists forget that all the phylogenetical characters have to be tested and that these criteria must also be applied in descriptions of taxa. Following the traditional path increases confusion among the taxa, and Lingula as well as Obolus are among the most prominent examples, for they have endured for nearly two centuries.
![]() In the identification of genera and species of Linguloid taxa soft-bodied characters or their scars and impressions are required because external shell characters have no suprageneric taxonomical value. The diagnoses of the family Lingulidae, and of its three genera that can be recognized with certainty, have been emended on Table 1
In the identification of genera and species of Linguloid taxa soft-bodied characters or their scars and impressions are required because external shell characters have no suprageneric taxonomical value. The diagnoses of the family Lingulidae, and of its three genera that can be recognized with certainty, have been emended on Table 1 ![]() based on the recent original descriptions (see Emig, 2002).
Diagnoses made recently by Holmer and Popov (2000) do not correspond to the original descriptions; these authors did not use many of the taxonomic characters established previously by Emig (1982b, 1983b) to describe the extant lingulide species, and applied by Biernat and Emig (1993) to the fossil genus Lingularia and its species,
and to Obolus by Emig (2002).
based on the recent original descriptions (see Emig, 2002).
Diagnoses made recently by Holmer and Popov (2000) do not correspond to the original descriptions; these authors did not use many of the taxonomic characters established previously by Emig (1982b, 1983b) to describe the extant lingulide species, and applied by Biernat and Emig (1993) to the fossil genus Lingularia and its species,
and to Obolus by Emig (2002).
![]()
![]() As a group the living lingulides approach closely a steady state (Emig, 1984b, 1989) in view to the following characteristics:
As a group the living lingulides approach closely a steady state (Emig, 1984b, 1989) in view to the following characteristics:
![]() Most of the species of the Superfamily Linguloidea
share an infaunal way of life as a plesiomorphic state in contrast to the epifaunal one
of all other brachiopod taxa.
Most of the species of the Superfamily Linguloidea
share an infaunal way of life as a plesiomorphic state in contrast to the epifaunal one
of all other brachiopod taxa.
![]() From this set of characteristics Lingula is obviously the most primitive representative among the Brachiopoda (about 420 species) From the ancestral infaunal behaviour it has retained the following features (Fig. 1
From this set of characteristics Lingula is obviously the most primitive representative among the Brachiopoda (about 420 species) From the ancestral infaunal behaviour it has retained the following features (Fig. 1
![]() , 4
, 4
![]() , 5
, 5
![]() , 6
, 6 ![]() , 7
, 7 ![]() ):
):
![]() Despite a low specific diversity (respectively 7 and 5 species), the two extant genera show a
broad geographical distribution on the continental shelf in many temperate and intertropical biocoenoses: Lingula in Asia, in Oceania, and in Africa; Glottidia on the American continent (Emig, 1997b).
Despite a low specific diversity (respectively 7 and 5 species), the two extant genera show a
broad geographical distribution on the continental shelf in many temperate and intertropical biocoenoses: Lingula in Asia, in Oceania, and in Africa; Glottidia on the American continent (Emig, 1997b).
![]()
![]() Although the Recent Lingulidae constitute a panchronic group, the evaluation of the bradytelic evolution of a rather poorly preserved group with a low potential for fossilisation is under development based on the recent descriptions of anatomical structures in the fossil taxa (Biernat and Emig, 1993; Jin et
alii, 1993; Kowalevski and Flessa, 1996; Emig, 2002; Zhang et alii, 2003). Albeit they have kept surficial aspects derived from ancestral life, species of Lingula show significant evolutionary differences and can no longer be considered as "living-fossils"! Consequently, the traditional opinions regarding Lingula must be rejected because the diagnoses of the Lingulidae given here are based on recent morphological findings and evolutionary novelties.
Although the Recent Lingulidae constitute a panchronic group, the evaluation of the bradytelic evolution of a rather poorly preserved group with a low potential for fossilisation is under development based on the recent descriptions of anatomical structures in the fossil taxa (Biernat and Emig, 1993; Jin et
alii, 1993; Kowalevski and Flessa, 1996; Emig, 2002; Zhang et alii, 2003). Albeit they have kept surficial aspects derived from ancestral life, species of Lingula show significant evolutionary differences and can no longer be considered as "living-fossils"! Consequently, the traditional opinions regarding Lingula must be rejected because the diagnoses of the Lingulidae given here are based on recent morphological findings and evolutionary novelties.
![]() In conclusion, the notion of "living-fossils" is very probably erroneous. In fact such a concept appears more journalistic than scientific, for it is based for the most part on a unique characteristic maintained over time, like the lingulide shape and the coelocanth fin and scale pattern. In any event only a species can be referred to as a living-fossil - supra-specific taxa are excluded automatically.
In conclusion, the notion of "living-fossils" is very probably erroneous. In fact such a concept appears more journalistic than scientific, for it is based for the most part on a unique characteristic maintained over time, like the lingulide shape and the coelocanth fin and scale pattern. In any event only a species can be referred to as a living-fossil - supra-specific taxa are excluded automatically.
![]() I am most grateful to Nestor J. Sander (USA), Art Boucot (Oregon State University, Corvallis, USA) and Fernando Alvarez (Universidad de Oviedo, Spain) for comments
on and improvements in the English of the earlier version.
I am most grateful to Nestor J. Sander (USA), Art Boucot (Oregon State University, Corvallis, USA) and Fernando Alvarez (Universidad de Oviedo, Spain) for comments
on and improvements in the English of the earlier version.
![]()
Arnaud P.M., Emig C.C. (1987).- La population, unité fonctionnelle de la biocoenose. In: Biologie des Populations.- Actes du Colloque National du Centre National de la Recherche Scientifique, Lyon 1986, pp. 69-72.
Babin C., Delance J.H., Emig C.C., Racheboeuf P.R. (1992).- Brachiopodes et Mollusques Bivalves : concurrence ou indifférence ?- Géobios, Mémoire spécial, Lyon, Vol. 14, pp. 35-44.
Biernat G., Emig C.C. (1993).- Anatomical distinctions of the Mesozoic lingulide brachiopods.- Acta Palaeontologica Polonica, Warsaw, Vol. 38, n° 1/2, pp. 1-20.
Emig C.C. (1982a).- Terrier et position des Lingules (Brachiopodes, Inarticulés).- Bulletin de la Société zoologique de France, Paris, Vol. 107, n° 2, pp. 185-194.
Emig C.C. (1982b).- Taxonomie du genre Lingula (Brachiopodes, Inarticulés).- Bulletin du Muséum national d'Histoire naturelle de Paris, Paris, (4), Vol. 4, Section A, n° 3/4, pp. 337-367.
Emig C.C. (1983a).- Sur les relations du panchronisme avec les conditions écologiques : le cas des Lingules (Brachiopodes, Inarticulés).- Bulletin de la Société zoologique de France, Paris, Vol. 108, n° 4, pp. 558-561.
Emig C.C. (1983b).- Taxonomie du genre Glottidia (Brachiopodes, Inarticulés).- Bulletin du Muséum national d'Histoire naturelle de Paris, Paris, (4), Vol. 5, Section A, n° 2, pp. 469-489.
Emig C.C. (1984a).- On the origin of the Lophophorates.- Zeitschrift für zoologische Systematik und Evolutionforschung, Frankfurt am Main, Vol. 22, n° 2, pp. 91-94.
Emig C.C. (1984b).- Pourquoi les Lingules (Brachiopodes, Inarticulés) ont survécu à la transition Secondaire-Tertiaire.- Bulletin du Comité des Travaux historiques et scientifiques, Section Sciences, Paris, Vol. 6, pp. 87-94.
Emig C.C. (1989).- Les lingules fossiles, représentants d'écosystèmes oligotypiques ?- Atti 3° Simposio de Ecologia e Paleoecologia de Comunità bentoniche (Catania, Italia, 1985), pp. 117-121.
Emig C.C. (1997a).- Ecology of the inarticulated brachiopods. In: Kaesler R.L. (ed.), Treatise on Invertebrate Paleontology. Part H. Revised Brachiopoda.- Geological Society of America, Boulder, and University of Kansas, Lawrence, Vol. 1, pp. 473-495.
Emig C.C. (1997b).- Biogeography of the inarticulated brachiopods. In: Kaesler R.L. (ed.), Treatise on Invertebrate Paleontology. Part H. Revised Brachiopoda.- Geological Society of America, Boulder, and University of Kansas, Lawrence, Vol. 1, pp. 497-502.
Emig C.C. (2002).- Tools for linguloid taxonomy: the genus Obolus (Brachiopoda) as an example.- Carnets de Géologie/Notebooks on Geology, Maintenon, Article 2002/01, https://carnetsgeol.net/cg/ CG2002_A01/index.html
Holmer L.E., Popov L.E. (2000).- Class Lingulata. In: Kaesler R.L. (ed.), Treatise on Invertebrate Paleontology. Part H. Revised Brachiopoda.- Geological Society of America, Boulder, and University of Kansas, Lawrence, Vol. 2, pp. 30-146.
Jin Y., Hou X., Wang W. (1993).- Lower Cambrian pediculate lingulides from Yunnan, China.- Journal of Paleontology, Tulsa, Vol. 67, pp. 788-798.
Kowalevski M., Flessa K.W. (1996).- Improving with age: The fossil record of lingulide brachiopods and the nature of taphonomic megabiases.- Geology, Boulder, Vol. 24, n° 11, pp. 977-980.
Zhang X., Hou X., Emig C.C. (2003).- Evidence of lophophore diversity in Early Cambrian Brachiopoda.- Proceedings of the Royal Society of London, Biological Sciences, London, Biology Letter, DOI: 10.1098/rsbl.2003.0013.
![]()
Click on thumbnail to enlarge the image.
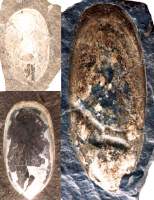
Figure 1: Linguloid specimens from the Ordovician (collection NMNH, Smithsonian Institution, Washington DC, USA) often described as Lingula species.
Click on thumbnail to enlarge the image.
Figure 2: Shell outline and internal view of ventral valve (along with the body muscles involved in valve movements and the main mantle canals) and a lateral view of the shell (after Emig, 1977a; Biernat and Emig, 1993; Jin et alii, 1993).
Click on thumbnail to enlarge the image.
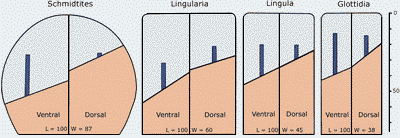
Figure 3: Diagram of the extensions (minimum-maximum) of the lophophoral cavity and of the mean extension of the mantle canals based on average shell dimensions in Glottidia, Lingula and Lingularia (see also Biernat and Emig, 1993) and for comparison in the obolid Schmidtites celatus from the Upper Cambrian of Estonia (see Emig, 2002).
Click on thumbnail to enlarge the image.
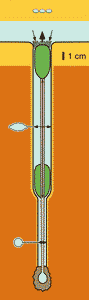
Figure 4: Diagram of a burrow of a lingulide (after Emig, 1982a) with the animal in life position (= feeding one) and after retraction into its burrow.
Click on thumbnail to enlarge the image.
Figure 5: Lingularia siberica (from the Triassic of Northern Siberia) - shell length about 0.7 cm.
Click on thumbnail to enlarge the image.
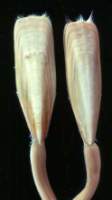
Figure 6: Lingula anatina (from Sumatra) - shell length about 4.5 cm.
Click on thumbnail to enlarge the image.
Figure 7: Glottidia palmeri (from the Gulf of California, Mexico) - shell length about 4.5 cm.
![]()
Click on thumbnail to enlarge the table.
Table 1: Systematic part: Emended diagnoses directly derived from the original descriptions and diagnoses, to replace the diagnoses of Holmer and Popov (2000).
![]()