◄ Carnets Geol. 4 (A01 fr) ►
![]()
![]()
CNRS UMR 6540, Centre d'Océanologie, Chemin de la
Batterie-des-Lions, 13007 Marseille (France)
CNRS UMR 7093, Laboratoire d'Océanographie de Villefranche; Laboratoire d'Ichtyologie, Muséum National d'Histoire Naturelle, 43 Rue Cuvier, 75231 Paris cedex 05 (France)
Manuscrit en ligne depuis le 12 Avril 2004
![]()
Le benthos profond méditerranéen est caractérisé par une absence d'originalité et une pauvreté dont les raisons sont à rechercher dans l'histoire récente de la faune. Trois types principaux d‘échanges ont été distingués :
• les échanges historiques à travers les changements de faunes depuis le Pliocène et durant les glaciations du Quaternaire ;
• les échanges bathymétriques au sein des étages Bathyal et Abyssal soumis à des conditions très particulières (homothermie, relative oligotrophie, barrière du seuil de Gibraltar, circulation des masses d'eaux) ; plus profonde est l'extension des espèces en Mer Méditerranée et plus large est leur distribution hors Méditerranée ;
• les échanges géographiques avec des affinités étroites
entre Mer Méditerranée et Océan Atlantique. Le cas des espèces endémiques reste un problème biogéographique. Les espèces ont toujours une distribution plus profonde en allant vers
l'Est et leur taille devient plus petite.
Ainsi, la Mer Méditerranée profonde actuelle apparaît comme une mer beaucoup plus jeune qu'aucune autre partie de l'Océan mondial profond.
• Mer Méditerranée ;
• Faune profonde ;
• Bathyal ;
• Abyssal ;
• Échanges ;
• Glaciations.
C.C. & P. (2004).- Faune profonde en Mer Méditerranée : les échanges historiques, géographiques et bathymétriques.- Carnets Geol., Madrid, vol. 4, no. A01 (fr), 10 p. DOI : 10.4267/2042/289
The Mediterranean deep-sea fauna: historical evolution, bathymetric variations and geographical changes.-
The deep-water fauna of the Mediterranean is characterized by an absence of distinctive characteristics and by a relative impoverishment. Both are a result of events after the Messinian salinity crisis (Late Miocene). The three main classes of phenomena involved in producing or recording these effects are analysed and discussed:
• Historical: Sequential faunal changes during the Pliocene and thereafter in particular those during the Quaternary glaciations and still in progress.
• Bathymetric: Changes in the vertical aspects of the Bathyal and Abyssal zones that took place under peculiar conditions,
i.e. homothermy, a relative oligotrophy, the barrier of the Gibraltar sill, and water mass movement. The deeper the habitat of a species in the Mediterranean, the more extensive is its distribution elsewhere.
• Geographical: There are strong affinities and relationships between Mediterranean and Atlantic faunas. Endemic species remain a biogeographical problem. Species always become smaller in size eastward where they occupy a progressively deeper habitat.
Thus, the existing deep Mediterranean Sea appears to be younger than any other deep-sea constituent of the World Ocean.
• Mediterranean
Sea;
• Deep-sea fauna;
• Bathyal;
• Abyssal;
• Changes;
• Glaciations.
Le rebord du plateau continental constitue une limite, séparant le domaine néritique du domaine profond, aussi importante que la ligne côtière. Ce rebord est défini par des caractéristiques à la fois géologiques, physico-chimiques et biologiques (, 1997).
Le domaine profond est divisé en trois étages (Tableau 1), dont l'extension et l'importance en Mer Méditerranée diffèrent de ceux dans l'Océan Mondial. Le domaine profond méditerranéen reste mal connu, probablement parce que le benthos profond est caractérisé par une absence d'originalité et une pauvreté dont les raisons sont à rechercher dans l'histoire récente de la faune. Trois principaux types d‘échanges en sont à l'origine : d'abord, les échanges historiques depuis le Pliocène, surtout pendant les fluctuations climatiques du Quaternaire avec des successions de périodes glaciaires et interglaciaires (Fig. 1
![]() ) ; puis, les échanges verticaux à cause des conditions très particulières de l'environnement profond ; et enfin, les échanges géographiques, avec des affinités entre les bassins occidentaux et orientaux de la Mer Méditerranée, ainsi qu'entre cette mer et le proche Océan Atlantique.
) ; puis, les échanges verticaux à cause des conditions très particulières de l'environnement profond ; et enfin, les échanges géographiques, avec des affinités entre les bassins occidentaux et orientaux de la Mer Méditerranée, ainsi qu'entre cette mer et le proche Océan Atlantique.
Tableau 1 : Comparaison entre l'Océan Mondial et la Mer Méditerranée qui ne représente que 0,82 % de la superficie totale de l'Océan Mondial et 0,35 % de son volume. Le rebord du plateau continental (dont la profondeur est variable selon l'océan ou la mer pris en considération) marque la frontière entre les deux domaines.
|
Domaine néritique
|
Domaine profond
|
|||
| Plateau continental | étage Bathyal | étage Abyssal | étage Hadal | |
| Limites en m | ||||
| Océan Mondial | 0 m à variable | Jusqu'à 3000 m | 3000 à 6000 m | Plus de 6000 m |
| Mer Méditerranée | 0 à 100-110 m | 100-110 à 3000 m | 3000 à 5093 m | - |
| Extension en % de la surface totale | ||||
| Océan Mondial | 7 % | 15 % | 77 % | 1 % |
| Mer Méditerranée | 15 % | 72 % | 13 % | - |
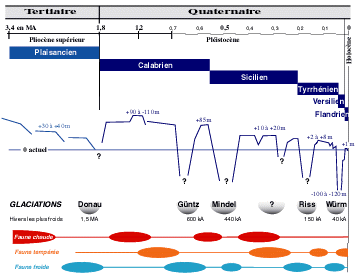
Figure 1 : Principales phases de succession des faunes en Mer Méditerranée depuis le Pliocène durant les diverses périodes glaciaires et interglaciaires (marquées par les variations du niveau de la mer) (modifié, d'après & , 1993). Le nombre réel de glaciations au cours du Pléistocène reste à définir ; il est possible qu'il y ait une ou deux autres glaciations entre celles de Mindel et de Riss. |
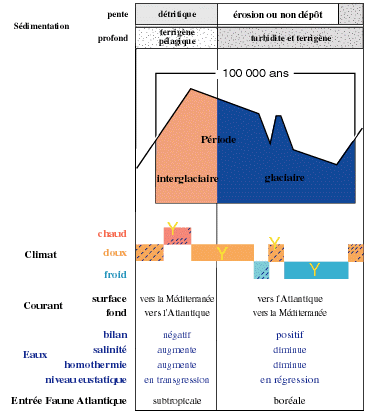
Ces échanges sont tous directement soumis aux caractéristiques du domaine profond méditerranéen, principalement à l'homothermie (de l'ordre de 13°-13,5°C dans le bassin occidental et 14-15°C dans le bassin oriental), à une forte salinité (environ 38 à 38,5 psu*), à la stratification des couches d'eaux et à leur circulation barotropique, aux courants dans les détroits (Gibraltar et
siculo-tunisien) (Fig. 2
![]() ).
).
* Selon les conventions internationales (UNESCO, 1985), les valeurs de salinité n'ont pas d'unité, les données sont exprimées en psu (= "practical salinity unit").
Figure 2 : Résumé des principales caractéristiques d'une glaciation quaternaire (périodes glaciaire et interglaciaire) en Mer Méditerranée. |
Pour comprendre l'évolution de la faune profonde de la Mer Méditerranée, il est indispensable de prendre en compte l'histoire géologique récente de cette mer depuis la fermeture définitive avec l'Océan Indien au Burdigalien moyen (Miocène inférieur) ( et , 1984), alors que son extension actuelle ne sera atteinte qu'à la fin de l'ère Tertiaire.
Au cours du Messinien (Miocène supérieur), la crise la plus importante pour la Mer Méditerranée récente fut la "crise de salinité messinienne" ( et al., 1978), dont les effets provoquèrent une forte réduction de la faune, affectant directement le domaine profond. Ainsi, la plupart des éléments paléo-méditerranéens, notamment la faune tropicale néogène (comme les poissons de l'éocène-Oligocène, des brachiopodes, etc.) disparaissent. Pourtant, plusieurs genres et espèces de cette période sont encore présents actuellement dans la faune méditerranéenne profonde. L'hypothèse la plus vraisemblable, mais elle est loin de faire l'unanimité (voir , 1986 ; et al., 1996 ; , 2000 ; , 2002), est que, durant cette crise, subsistaient des zones profondes (plus de 1 000-1500 m d'eau) où une partie de la faune profonde a pu se maintenir ou survivre ( et al., 1988 ; et al., 1989 ; 1990 ; et , 1993), notamment dans les bassins périphériques (comme dans le Sud-Est de l'Espagne : et al., 1999, ou dans le Sud de l'Italie : et al., 1989). Réfutant la "théorie du bassin profond desséché", (1984), (1984), et (1984), (1990) proposent un modèle de mise en place des "évaporites", selon le système des bassins oscillants avec une liaison continue durant le Messinien avec l'Océan Atlantique par le seuil de Gibraltar et le maintien de plaines abyssales, ce qui corrobore l'hypothèse ci-dessus de la survivance d'une faune profonde durant le Messinien. Néanmoins, cet évènement messinien a provoqué une rupture qui marque le début de l'évolution biologique de cette jeune mer qu'est la Mer Méditerranée.
Au Pliocène, lors de la transgression plaisancienne (Fig. 1
![]() ), les conditions climatiques (environ +4°C par rapport au présent) étaient relativement stables et les eaux chaudes subtropicales prédominaient ce qui a permis l'entrée par le détroit Gibraltar d'une faune atlantique depuis la baie Ibéro-marocaine et ainsi que le repeuplement des bassins méditerranéens. Ensuite, vers 2,1-2 Ma, le climat est devenu tempéré à froid, ce qui a favorisé l'entrée d'un grand nombre d'espèces des zones tempérées boréales de l'Océan Atlantique permettant ainsi la constitution d'un stock "atlantico-méditerranéen" dominant dans la faune de la Méditerranée.
), les conditions climatiques (environ +4°C par rapport au présent) étaient relativement stables et les eaux chaudes subtropicales prédominaient ce qui a permis l'entrée par le détroit Gibraltar d'une faune atlantique depuis la baie Ibéro-marocaine et ainsi que le repeuplement des bassins méditerranéens. Ensuite, vers 2,1-2 Ma, le climat est devenu tempéré à froid, ce qui a favorisé l'entrée d'un grand nombre d'espèces des zones tempérées boréales de l'Océan Atlantique permettant ainsi la constitution d'un stock "atlantico-méditerranéen" dominant dans la faune de la Méditerranée.
Bien que la limite Plio-Pléistocène soit encore sujette à controverses, elle peut être définie par l'arrivée de ces espèces boréales en Mer Méditerranée à cause de modifications climatiques et océanographiques, notamment durant la régression à la fin du Plaisancien : la température des eaux de surface diminue de plusieurs degrés avec déplacements alternatifs des vents dominants, durant la glaciation de Donau (Fig. 1
![]() ).
).
Ensuite, au Quaternaire (du Pléistocène à nos jours) se succèdent des fluctuations climatiques remarquables - période glaciaire et période interglaciaire - dont les principales sont résumées dans la figure 2. Les profondeurs de la Méditerranée étaient semblables à l'Actuel, le détroit de Gibraltar avait un seuil d'environ 300 m de profondeur et les variations du niveau de la mer étaient dues aux processus glacio-eustatiques de l'Océan Mondial.
En période glaciaire, sous climat froid, de larges biocœnoses se développent sur le plateau continental et la pente avec des espèces circalittorales et bathyales, généralement tempérées à boréales, dont l'introduction en Mer Méditerranée à partir de l'Océan Atlantique est favorisée par le courant de fond dans le détroit de Gibraltar. En revanche, en période inter-glaciaire, sous climat tempéré à chaud (qui ne représente qu'environ 10 % du cycle total de 90 à 125 ka), le courant
de surface, entrant en Mer Méditerranée, tend à introduire des espèces subtropicales de la Province Sénégalienne, tandis que le courant en profondeur, sortant de Méditerranée, rend difficile l'arrivée d'espèces profondes (Figs. 1
![]() - 2
- 2
![]() ).
).
Durant le Calabrien (glaciation de Donau), le Sicilien (glaciation de Mindel) et le Tyrrhénien (glaciation de Riss), un stock d'espèces boréales s'est ajouté en vagues successives au stock atlantico-méditerranéen, surtout d'origine tempérée, arrivé au début du Pléistocène (glaciation de Donau) et au stock d'espèces paléoméditerranéennes survivantes, mais avec disparition du stock pliocène. Ces arrivées sont liées à un climat tempéré avec des saisons bien marquées, avec des températures des eaux de surface de l'ordre de 9-10°C en hiver et 19-20° C en été : or, ces espèces (parfois nommées faune "celtique") vivent encore actuellement dans le Nord-Est de l'Océan Atlantique, comme le mollusque gastéropode Buccinum undatum (qui vit encore en Méditerranée, à plus de 1 000 m) et les bivalves Cyprina (Arctica) islandica, Mya truncata, Panomya spengleri, Modiolus modiolus et Chlamys islandica, ainsi que, probablement, les poissons pleuronectidé Platichthys flesus et rajidé Raia clavata, toujours pêchés en Mer Méditerranée ( et , 1964 ; , 1986). C'est vers la fin du Sicilien, début du Tyrrhénien (glaciation de Riss) que se sont développées les biocœnoses bathyales et abyssales.
Au Tyrrhénien, durant l'interglaciation de Riss-Würm, le niveau des eaux n'est pas monté à plus de 2 à 8 m au-dessus du niveau actuel ; des espèces tempérées chaudes, surtout d'origine sénégalienne, se sont introduites, notamment un stock important de mollusques maintenant disparus.
Lors de la dernière glaciation, celle de Würm (de ≈70 000 à 18 000 ans BP) au Flandrien (Holocène), le niveau de la mer était descendu de -100 à -120 m par rapport au niveau actuel (Fig. 1
![]() ; Tableau 2) avec des conditions très difficiles pour les biocœnoses bathyales et abyssales et des entrées limitées d'espèces atlantiques en Mer Méditerranée. Les thanatocoenoses des faunes malacologiques "froides" celtiques (notamment les mollusques bivalves Chlamys septemradiata et Cyprina (Arctica) islandica) sont contemporaines de la glaciation de Würm (datations : 13 000 - 9 800 ± 300 ans BP ; , 1974). Elles se trouvent actuellement entre 90 et 600 m de profondeur (généralement entre 200-300 m), mais elles se sont développées lorsque le niveau de la mer était à environ - 80 m. Chlamys septemradiata était aussi présente en Mer Egée, mais elle a disparu de la Mer Méditerranée, excepté en Mer d'Alboran où elle vit actuellement. Elle ne tolère pas des salinités de plus de 36,5 psu, ce qui explique qu'elle ne s'étend pas plus à l'Est. Cyprina (Arctica) islandica, très fréquente dans les gisements fossiles, a aussi été éliminée par la salinité élevée de la Mer Méditerranée.
; Tableau 2) avec des conditions très difficiles pour les biocœnoses bathyales et abyssales et des entrées limitées d'espèces atlantiques en Mer Méditerranée. Les thanatocoenoses des faunes malacologiques "froides" celtiques (notamment les mollusques bivalves Chlamys septemradiata et Cyprina (Arctica) islandica) sont contemporaines de la glaciation de Würm (datations : 13 000 - 9 800 ± 300 ans BP ; , 1974). Elles se trouvent actuellement entre 90 et 600 m de profondeur (généralement entre 200-300 m), mais elles se sont développées lorsque le niveau de la mer était à environ - 80 m. Chlamys septemradiata était aussi présente en Mer Egée, mais elle a disparu de la Mer Méditerranée, excepté en Mer d'Alboran où elle vit actuellement. Elle ne tolère pas des salinités de plus de 36,5 psu, ce qui explique qu'elle ne s'étend pas plus à l'Est. Cyprina (Arctica) islandica, très fréquente dans les gisements fossiles, a aussi été éliminée par la salinité élevée de la Mer Méditerranée.
Tableau 2 : Évolution paléo-océanographique des eaux ouest-méditerranéennes depuis la dernière glaciation de Würm (d'après , 1987).
| Age en K ans BP | -18 | -15 | -12 | -10 | -0,8 | Actuel |
| Niveau de la mer (- m) | 120 | 90 | 70 | 50 | 20 | 0 |
| Température (°C) | 8 | 9 | 10 | 8,5 | 13 | 13 |
| Salinité (psu) | 40,1 | 39 | 38,5 | 38,9 | 37 | 37,9 |
| Temps de renouvellement (ans) | 133 | 124 | 91 | 79 | 97 | 75 |
Dans le bassin méditerranéen oriental, les formations de sapropèles (de 2,5 à 70 cm d'épaisseur selon les régions, et , 1988) ont eu lieu entre 25 000 - 23 000 ans BP (S2) et 9 000 - 6 800 ans BP (S1); elles sont mises en place grâce à de fortes arrivée d'eaux douces (Nil, Mer Noire), provoquant une stratification des eaux (une couche d'eaux douces sur des eaux froides fortement salées) et par une sédimentation de matériel organique et terrigène (en partie d'origine éolienne) provoquant la consommation de tout l'oxygène en profondeur. Cette anoxie se développe en période interglaciaire dans l'étage Bathyal (sous 600-1 000 m) et restreint la colonisation tout en provoquant une extinction sous cette profondeur. Puis, suit une recolonisation en période glaciaire ( et , 1992) ; néanmoins, des températures et salinités plus élevées que dans le bassin occidental, empêchent la pénétration d'espèces sténothermes et sténohalines du Bathyal ou même du plateau continental malgré un courant favorable en profondeur au niveau du seuil siculo-tunisien.
Selon (1994), les sapropèles ont été mis en place dans un type de circulation anti-estuarine avec réduction de l'excès d'évaporation face à l'arrivée d'eau douce dans le bassin oriental. Cette réduction a été imposée par une intensification de l'effet des moussons de l'Océan Indien avec une augmentation du débit du Nil et par le système de dépressions depuis le bassin occidental avec une augmentation des précipitations et une réduction de l'évaporation. L'absence de sapropèles dans le bassin occidental peut être attribuée à l'absence de stratification des eaux.
Pour et (1964), l'évolution des peuplements profonds méditerranéens durant la période plio-quaternaire apparaît plus liée aux modifications des régimes de courants dans le détroit de Gibraltar qu'aux variations de température des eaux, au moins pour expliquer les vagues successives d'espèces tantôt boréales (celtiques), tantôt tropicales ou tempérées chaudes. Pour d'autres auteurs (cf. , 1970), ce sont les facteurs salinité et hydrodynamisme, plutôt que la température, qui ont réglé la pénétration des faunes "chaudes" ou "froides". Les peuplements profonds appartiennent essentiellement à l'étage Bathyal et leur introduction et développement se fait surtout en période glaciaire où les conditions (température, salinité, apport nutritionnel, courants, etc.) sont favorables à des espèces "tempérées-froides" ; leur arrivée dépend de leurs caractéristiques et exigences écologiques en fonction des conditions de milieu présentes.
Dans la distribution verticale du benthos profond méditerranéen, l'homothermie profonde est le facteur le plus important : il n'y a pas de variations de température pour limiter, en profondeur, la faune à divers niveaux comme dans l'Océan Atlantique. Beaucoup d'espèces ne sont eurybathes en Mer Méditerranée que parce qu'elles sont eurythermes ou sténothermes chaudes. Aussi, les limites de distribution sont régies par d'autres facteurs, tels que salinité, granulométrie, pression, nourriture disponible, hydrodynamisme. Et, cette variété de conditions locales conduit à un étage Bathyal plus hétérogène que celui de l'Océan Atlantique (, 1989 ; et , 1993).
Les biocœnoses bathyales de la Mer Méditerranée sont très similaires à celles de la pente dans le NE de l'Océan Atlantique, mais appauvries en richesse spécifique et en abondance, à cause :
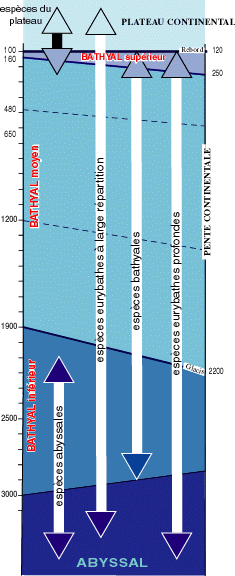
L'étage Bathyal comporte plusieurs sous-étages dont les limites méditerranéennes (Fig. 4
![]() ) correspondent à celles décrites dans le Bathyal océanique, mais avec modification de la faune. Les facteurs en cause seraient : la nourriture disponible couplée avec l'énergie nécessaire pour la collecte en raison de la diminution de la faune, notamment de l'endobenthos et du macroplancton, avec la profondeur (, 1997).
) correspondent à celles décrites dans le Bathyal océanique, mais avec modification de la faune. Les facteurs en cause seraient : la nourriture disponible couplée avec l'énergie nécessaire pour la collecte en raison de la diminution de la faune, notamment de l'endobenthos et du macroplancton, avec la profondeur (, 1997).
Pour le macrobenthos abyssal, on peut conclure qu'en dépit d'un fort pourcentage d'espèces eurybathes, une véritable faune abyssale est représentée par environ 20 à 30 espèces. Dans le bassin occidental, où la profondeur ne dépasse pas les 3 000 m, les formes abyssales sont moins abondantes. Au contraire, celles-ci semblent dominantes dans la fosse abyssale de Matapan (5 093 m ; bassin oriental).
La macrofaune comportant actuellement quelque 7 200 espèces (soit environ 5,6 % de la faune marine mondiale), est loin d'être entièrement connue, bien que la faune méditerranéenne soit celle qui a été et est la plus étudiée ( et , 2000).
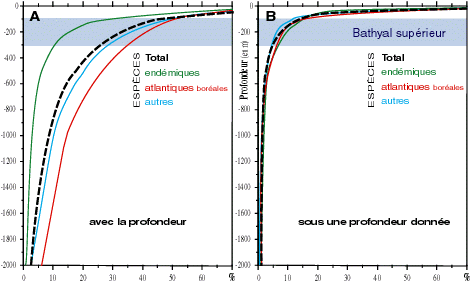
La comparaison entre la distribution bathymétrique des espèces (Fig. 3A
![]() ) et celle des espèces vivant strictement sous une profondeur donnée (Fig. 3B
) et celle des espèces vivant strictement sous une profondeur donnée (Fig. 3B
![]() ) fournit une estimation du degré d'eurybathie de la faune à une même profondeur ( & , 1983) : ce degré diminue graduellement avec la profondeur (ainsi, 25 % des espèces vivent au-delà de 50 m, 15 % sous 100 m, 1 % sous 2 000 m). En outre, avec la profondeur, jusqu'à 2 000 m, le pourcentage des espèces présentes dans les eaux littorales augmente : par exemple, près de 40 % des espèces vivant entre 0 et 50 m ont une distribution plus ou moins étendue en profondeur, et, sous 500 m, il y a 6 fois plus d'espèces vivant également dans les hauts niveaux que d'espèces strictement profondes. Cette distribution eurybathe coïncide avec la distribution des mêmes espèces hors de la Mer Méditerranée.
) fournit une estimation du degré d'eurybathie de la faune à une même profondeur ( & , 1983) : ce degré diminue graduellement avec la profondeur (ainsi, 25 % des espèces vivent au-delà de 50 m, 15 % sous 100 m, 1 % sous 2 000 m). En outre, avec la profondeur, jusqu'à 2 000 m, le pourcentage des espèces présentes dans les eaux littorales augmente : par exemple, près de 40 % des espèces vivant entre 0 et 50 m ont une distribution plus ou moins étendue en profondeur, et, sous 500 m, il y a 6 fois plus d'espèces vivant également dans les hauts niveaux que d'espèces strictement profondes. Cette distribution eurybathe coïncide avec la distribution des mêmes espèces hors de la Mer Méditerranée.
Figure 3 : Distribution verticale de 3613 espèces d'invertébrés benthiques en Mer Méditerranée : A. pourcentage des espèces de chaque groupe récoltées à une profondeur donnée, quel que soit l'intervalle de leur distribution verticale ; B. pourcentage des espèces de chaque groupe vivant strictement sous une profondeur donnée (d'après les données de Médifaune, & , 1983). |
L'endémisme méditerranéen d'abord estimé à 15-16 % des espèces, est aujourd'hui de l'ordre de 28 à 29 %, mais il diminue rapidement avec la profondeur : environ 15 % à 200 m, 13 % à 500 m, 14 % à 1 000 m et 19 % à 2 000 m ( et al., 1992). En effet, quand la profondeur augmente, l'originalité de la faune décroît et il est difficile de proposer une hypothèse valable pour l'origine de cet endémisme ; en outre, plus de 75 % des espèces endémiques profondes ne sont connues que par un très petit nombre d'individus dans une ou quelques stations seulement. Les espèces endémiques profondes correspondent souvent à des espèces proches du Nord-Est Atlantique, ce qui pose les problèmes de leurs relations taxonomiques et de leur distribution. Au niveau des genres, à quelques exceptions près, il apparaît que les genres endémiques vivent presque exclusivement dans les eaux littorales. Si on admet un parallélisme entre le rang hiérarchique d'un taxon endémique et le temps passé par ce taxon dans son nouvel environnement, l'absence pratiquement totale de genres et de familles endémiques au sein de la faune profonde démontre clairement sa jeunesse.
L'homothermie en profondeur avec une température relativement élevée (environ 13°C dans le bassin occidental), alliée à une forte salinité plus de 38 psu, représente une barrière pour un grand nombre d'espèces. En y ajoutant toutes les conséquences des fluctuations climatiques quaternaires, on trouve l'explication de la relative pauvreté et de la faible originalité de la faune. Les espèces sténothermes froides sont absentes et plusieurs groupes taxonomiques typiquement profonds sont très rares (comme les éponges Hexactinellides et les Holothuries Elasipodes). La distribution bathymétrique des principaux stocks d'espèces dans le domaine profond est représentée sur la figure 4
![]() .
.
Figure 4 : Répartition bathymétrique des diverses catégories d'espèces (modifié, d'après , 1970 ; , 1970) avec les limites des étages Bathyal (et sous-étages) et Abyssal et leurs variations (en trait pointillé les sous-limites dans le Bathyal moyen). |
Les origines et l'importance des stocks d'espèces méditerranéennes (pas de données pour le domaine profond) sont les suivants :
| Origine atlantique | 50,2 % |
| Espèces endémiques | 28,6 % |
| Origine atlantico-pacifique | 16,8 % |
| Origine indo-pacifique | 4,4 % |
La faune profonde est principalement composée d'espèces eurybathes qui ont aussi une très large répartition géographique extra-méditerranéenne, avec un postulat général "plus profonde est leur extension méditerranéenne et plus large est leur distribution hors Méditerranée". Ainsi, près de 70 % de ces espèces sont communes avec la province boréale de l'Océan Atlantique (20 % sous 1 000 m) où elles sont généralement néritiques (comme les échinodermes Leptometra celtica et Plutonaster bifrons, le crustacé Amphilepsis norvegica, le polychète Nephtys ciliata, les mollusques Dentalium agile, Anamathia rissoana, etc.). Et, près de 35 % des espèces sont connues des deux côtés de l'Atlantique Nord. L'affinité étroite entre les espèces profondes congénériques méditerranéennes et atlantiques atteste de relations phylogénétiques étroites et suggère que les espèces endémiques bathyales méditerranéennes actuelles aient eu des ancêtres de même habitat, arrivés de l'Océan Atlantique quand les conditions étaient favorables.
Ceci démontre que le seuil de Gibraltar n'a pas toujours été et n'est pas une barrière infranchissable pour coloniser ou recoloniser le domaine profond, même pour des espèces strictement profondes, comme entre autres pour les crustacés Décapodes Polychelidés, un petit groupe strictement profond et les seuls "fossiles vivants" de la Mer Méditerranée (, 1987 ; et , 1992). Mais les groupes taxonomiques typiquement bathyaux ou abyssaux sont manquants, ainsi que des espèces sténothermes froides qui ailleurs représentent la majeure partie de la faune profonde.
Pour l'ensemble de la faune méditerranéenne, la tendance générale est un appauvrissement spécifique en allant de l'Ouest vers l'Est (Tableau 3). Les espèces vivant dans les deux bassins sont toujours plus profondes dans le bassin oriental, ce qui est une tendance générale dans tous les océans, c'est-à-dire une distribution bathymétrique des espèces profondes moins profondes sur la côte ouest que sur la côte est (, 1987).
Tableau 3 : Appauvrissement de la faune de l'Ouest vers l'Est (total des espèces connues - données compilées de divers auteurs et pour les polychètes de et al., 2002).
| Bassin occidental | Bassin oriental | Mer Noire | ||||
| Adriatique | Ionienne | Levantine | égée | |||
| Total des espèces en % | 92 | 54 | - | |||
| Espèces profondes en % | 97 | 33 | 20 | - | ||
| Total des espèces de Polychètes | 884 | 527 | 528 | 451 | 597 | 310 |
| et total % | 85 | 51 | 51 | 44 | 57 | 30 |
L'appauvrissement qualitatif plus fort dans la faune profonde permet de suggérer plusieurs explications possibles :
La Méditerranée profonde actuelle apparaît comme une mer beaucoup plus jeune qu'aucune autre partie de l'Océan Mondial profond. Sa faune est principalement composée de groupes anciens (primitifs ?) au sein des embranchements, tandis qu'une fraction plus réduite comprend les types les plus spécialisés de leur groupe.
Cette mer est un important centre d'évolution avec de futures spéciations et la création de nouvelles espèces endémiques à cause de ses particularités physiques à forts gradients. La Méditerranée constitue un laboratoire naturel remarquable et unique pour étudier les processus récents de colonisation en fonction de l'histoire spécifique de chacun des grands bassins méditerranéens. Les études sur la faune profonde devraient être particulièrement favorisées.
Nous adressons tous nos remerciements à Nestor J. (USA) pour ses commentaires et son aide pour l'amélioration de la version anglaise, ainsi que les trois lecteurs-arbitres Jean-Claude (Station Marine d'Arcachon, Université de Bordeaux 1, France), Alain (Département des Sciences de la Terre, Université de Bretagne occidentale, Brest, France) et Jean-Loup (Centre Scientifique et Technique, Total , Pau) pour leurs suggestions et remarques constructives.
P. & J.E. (1992).- Population characteristics of the deep-sea lobsters Polycheles typhlops and Stereomastis sculpta (Decapoda: Polychelidae) in a bathyal mud community of the Mediterranean Sea.- Marine Biology, New York, Vol. 114, pp. 109-117.
C., G., P., V., C., A. & A. (2002).- Seascape biodiversity patterns along the Mediterranean and the Black Sea : lessons from the biogeography of benthic polychaetes.- Marine Ecology Progress Series, Oldendorf (Luhe), Vol. 244, pp. 139-152.
D., G. & G. (1992).- Mise au point sur les connaissances concernant le benthos profond méditerranéen.- Oebalia - International Journal of Marine Biology and Oceanography, Taranto, suppl. Vol. 17, pp. 21-36.
P., I., C., M. & H. (1989).- Présence de faunes bathyales atlantiques dans le Pliocène et le Pléistocène de Méditerranée (détroit de Messine, Italie).- Bulletin de la Société géologique de France, Paris, (8), Vol. 5, N° 4, pp. 787-796.
A., P., A. & G. (1988).- Intérêt paléobiologique des mollusques messiniens de Melilla (NE Maroc).- Revue de Paléobiologie, Genève, Vol. 7, N° 2, pp. 335-358.
C.N. & C. (2000).- Marine biodiversity of the Mediterranean Sea: Situation, problems and prospects for future research.- Marine Pollution Bulletin, Oxford, Vol. 40, pp. 367-376.
P.-L. (2000).- Of sills and straits: a quantitative assessment of the Messinian salinity crisis.- Deep-Sea Research I, Oxford, 47, pp. 1429-1460.
H.A. & L.R. (1988).- Late Pleistocene to Recent sediment deposition in the central and western Mediterranean.- Deep-Sea Research, Oxford, Vol. 35, pp. 749-766.
G. (1984).- Transposition des données sur les marais salants aux grandes accumulations évaporitiques du passé.- Géologie méditerranéenne, Marseille, Vol. 9, N° 4, pp. 563-591.
G. (1990).- Le Messinien de la Méditerranée... vingt ans après.- Géologie de la France, Orléans, Vol. 3-4, pp. 3-58.
C. (1970).- Ecologie de l'étage bathyal dans la Méditerranée occidentale.- Mémoires de l'Institut Océanographique de Monaco, Vol. 2, pp. 1-146.
G., J.P., F., A. & M.F. (1996).- Alternate interpretation of the Messinian salinity crisis: Controversy resolved?- Geology, Boulder, Vol. 24, N° 4, pp. 363-366.
I. (1990).- Biogeografia dello zoobenthos del Mediterraneo: origine e problematiche.- Oebalia - International Journal of Marine Biology and Oceanography, Taranto, suppl. Vol. 16, pp. 31-49.
Z. (1987).- An analysis of the zoogeography of the decapod crustaceans (Crustacea, Decapoda) of the Mediterranean.- Oceanology, Washington, Vol. 27, pp. 352-356.
C.C. (1989).- Distributional patterns along the Mediterranean continental margin (Upper Bathyal) using Gryphus vitreus (Brachiopoda) densities.- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, Vol. 71, pp. 253-256.
C.C. (1997).- Bathyal zones on the Mediterranean continental slope: An attempt.- Publicaciones Especiales del Instituto Español de Oceanografía, Madrid, Vol. 23, pp. 23-33.
G. & L. (1983).- The deep Mediterranean benthos. In: M. & V. (eds.), Mediterranean marine ecosystems.- NATO Conf. Ser. (I Ecol.), Plenum Press, New-York, vol. 8, pp. 109-145.
C. (1974).- Essai sur la géologie du précontinent de la Provence occidentale.- Thèse de Doctorat-es-Sciences, Université d'Aix-Marseille, 219 p.
B.S. & M. (1992).- The Levantine bathyal - a desert at the bottom of the sea?- Rapport de la Commission internationale de la Mer Méditerranée, Monaco, Vol. 33, pp. 39.
K. J., L., D., M.B., A., R.E., R.B., F., C. & R. (1978).- History of the Mediterranean salinity crisis. In: K.J., L. et al. (eds.), Initial Report of the Deep Sea Drilling Project, Washington, Vol. 42, N° 1, pp. 1053-1078.
A. (1984).- Sur la valeur de quelques hypothèses relatives à la genèse des grandes séries salines.- Revue de Géologie dynamique et de Géographie physique, Paris, Vol. 25, N° 3, pp. 149-156.
A. & P. (1984).- Les bassins oscillants : un modèle de genèse des séries salines.- Bulletin des Sciences géologiques, Strasbourg, Vol. 37, N° 3, pp. 267-282.
L. & C.C. (1993).- La faune benthique profonde de Méditerranée. In: F.R. (ed.), Symposium Mediterranen Seas 2000.- Istituto de Scienze ambientale marina, S. Margherita Ligure, pp. 397-428.
J.M. & J. (1964).- Nouveau manuel de bionomie benthique.- Recueil des Travaux de la Station Marine d'Endoume, Marseille, Vol. 31, fasc. 47, pp. 5-137.
J. (1987).- Was there any deep water formation in the Mediterranean sea before 8000 years BP?- Deep-Sea Research, Oxford, Vol. 34, pp. 309-312.
S. (1986).- The significance of marine boreal molluscs in the early Pleistocene faunas of the Mediterranean area.- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, Vol. 52, pp. 267-289.
D. (1970).- Bionomie benthique de deux canyons sous-marins de la mer Catalane: le rech du Cap et le rech Lacaze-Duthiers.- Thèse de Doctorat-es-Sciences, Université de Paris, 251 pp.
R., J.C. & J.M. (1999).- Late Miocene Mediterranean desiccation: topography and significance of the "Salinity Crisis" erosion surface on-land in southeast Spain.- Sedimentary Geology, Amsterdam, Vol. 123, pp. 1-7.
E.J. (1994).- Review and new aspects concerning the formation of eastern Mediterranean sapropels.- Marine Geology, Amsterdam, Vol. 122, pp. 1-28.
J.M. (1986).- Les évaporites miocènes de la Méditerranée et de la mer Rouge et leurs enseignements pour l'interprétation des grandes accumulations évaporitiques d'origine marine.- Bulletin de la Société géologique de France, Paris, (8), Vol. 2, N° 3, pp. 511-520.
F.F. & F.R. (1984).- Paleogeography and palinspastic reconstruction of the Neogene of the Mediterranean and Paratethys. In: J.E. & A.H.F. (eds.), The geological evolution of the eastern Mediterranean.- Blackwell scientific Publications, Oxford, pp. 659-668.
M. (2002).- The Mediterranean benthos from late Miocene up to present: ten million years of dramatic climatic and geologic vicissitudes.- Biologia Marina Mediterranea, Genova, Vol. 9, pp. 445-463.
O.N. (1987).- Brachiopods collected by Benthedi-cruise in the Mozambique Channel.- Bulletin du Muséum National d'Histoire naturelle de Paris, 4e série, Vol. 9, section A, N° 3, pp. 551-563.