◄ Carnets Geol. 4 (L03) ►
![]()
Carl von Ossietzky Universität Oldenburg,
Institute for Chemistry and Biology of the Marine Environment, Arbeitsgruppe Geomikrobiologie,
PO Box 2503, D 26111 Oldenburg, Germany
Carl von Ossietzky Universität Oldenburg, Institute for Chemistry and Biology of the Marine Environment, Arbeitsgruppe Geomikrobiologie, PO Box 2503, D 26111 Oldenburg, Germany
Carl von Ossietzky Universität Oldenburg, Institute for Chemistry and Biology of the Marine Environment, Arbeitsgruppe Geomikrobiologie, PO Box 2503, D 26111 Oldenburg, Germany
Manuscript online since June 20, 2004
![]()
The in vitro production of ooid-like structures as possible precursors of oolites has been observed in laboratory cultures of spherical microbial communities isolated from the Wadden Sea (North Sea). The microbial spherulites consist of aggregated benthic diatoms (Navicula perminuta) enveloped by layers of filamentous cyanobacteria of the genus Phormidium and a halo-like biofilm of heterotrophic bacteria. The development of the structures takes several months and these configurations appear to be stable, before they calcify. The precipitation starts on the surface of the spheres as clouds of small scattered crystals, which later increase in size and aggregate to form hollow spheres around the microbial assemblage. Here we report for the first time carbonate precipitation in defined spherical microbial communities.
Carbonate precipitate;
Cyanobacteria;
Diatoms;
Ooids;
Spherulites;
Microbial association.
Brehm U., Palinska K.A., Krumbein W.E. (2004).- Laboratory cultures of calcifying biomicrospheres generate ooids - A contribution to the origin of oolites.- Carnets Geol., Madrid, vol. 4, no. L03, 6 p. DOI: 10.4267/2042/309
Ooidbildung durch carbonatfällende Biomikrosphären - Laboruntersuchungen zur Entstehung von Oolithen.- Die in vitro Entstehung von ooidähnlichen Strukturen als möglichen Vorläufern der Oolithe, konnte in sphärischen mikrobiellen Gemeinschaften im Labor beobachtet und dokumentiert werden. Die Biomikrosphären wurden aus dem Wattenmeer (Nordsee) isoliert. Die Sphären bestehen aus einer Ansammlung von benthischen Diatomeen (Navicula perminuta), die mit mehreren Lagen fädiger Cyanobakterien der Art Phormidium und einer äußeren Hülle von heterotrophen Bakterien umgeben sind. Die Strukturentwicklung zieht sich über mehrere Monate hin und scheint stabil zu sein, ehe die Calcifizierung eintritt. Die Kristallisierung beginnt auf der Oberfläche der Sphären in Form von vereinzelten kleinen Kristallansammlungen, die sich später vergrößern, zusammenwachsen und damit eine Hohlkugel um die mikrobielle Gemeinschaft bilden. Wir berichten hier zum ersten Mal über die Carbonatausfällung in sphärischen mikrobiellen Gemeinschaften.
Carbonatausfällung;
Cyanobakterien;
Diatomeen;
Ooide;
Sphären;
mikrobielle
Gemeinschaft.
Production d'ooïdes à partir de cultures en laboratoire de biomicrosphères se calcifiant - Une contribution à la genèse des oolithes.- La production in vitro de structures semblables aux ooïdes a été observée à partir de cultures d'associations microbiennes sphériques récoltées en mer de Wadden (Mer du Nord). Les sphérules microbiennes sont des agrégats de diatomées benthiques (Navicula perminuta) entourés de couches de cyanobactéries filamenteuses du genre Phormidium et d'une auréole de bactéries hétérotrophes. Le développement de ces structures prend plusieurs mois et ces organisations sont stables avant qu'elles ne se calcifient. La calcification débute à la surface des sphères sous forme de nuages de petits cristaux isolés qui vont ensuite se développer et s'agglomérer pour constituer des sphères creuses autour des associations microbiennes. Il s'agit de la première observation de calcifications au sein de communautés microbiennes de forme sphérique.
Calcification ;
Cyanobactéries ;
Diatomées ;
Ooïdes ;
Sphérulites ;
Association microbienne.
Precipitation of calcium carbonate is widespread in microbial communities forming biofilms and microbial mats. The laminated structure of these communities consists of layers of carbonate which outlast the microbial colony that produced them. Fossilized remains of these communities in which particles of other sediment are also included are known as stromatolites. They have a long fossil record since early Proterozoic and still flourish in particular in the reefs of the Bahamas and Australia (e.g. Visscher et al., 2002).
The typical stromatolitic structure is laminated. Each lamina represents a horizon of former microbial biofilm or mat (Kalkowsky, 1908). Associated mineral particles (precipitates and detrital grains) are overgrown and sometimes entirely coated by microbial assemblages (Riding and Awramik, 2000). Small (mm size), spherical to oval concentrically laminated carbonate bodies or aggregates, which form in shallow tropical seas are called ooids and are known to become consolidated into rocks called oolitic limestones (oolith, Rogenstein; Kalkowsky, 1908). The genesis of ooid grains is still enigmatic. The alternative explanations are confronted along the lines of predominantly abiotic vs. biogenic origin of ooid grains and the associated carbonate precipitates.
The principal biochemical processes that have been recognized to affect the degree of carbonate saturation and therefore may cause carbonate precipitation include:
In this study we show the microbially induced formation of ooids in the laboratory.
Filamentous cyanobacteria of the genus Phormidium, diatoms (Navicula perminuta) and heterotrophic bacteria were isolated from Wadden Sea (North Sea) microbial mats. Species identification was based on both morphological features and the sequencing of the 16Sr RNA gene fragment (data not shown).
The isolates were grown on artificial seawater medium ASNIII solidified with 1% of Bacto Agar, prepared according to Rippka et al. (1979). The Petri Dishes were maintained at 18ºC, and 120 µmol photons m-2 s-1 (Osram tungsten light tubes) and with a light/dark cycle of 12/12 h. For the control experiments, the same medium and conditions of incubation were used, without organisms.
The cultures used in our experiments were not axenic. We worked on microbial community consisting of cyanobacteria, diatoms, and bacteria. However, filamentous cyanobacteria were always the dominant species regulating the development of the spheres.
In order to accelerate bacterial activity the experiments were set up with signal substance BHL (Butyroyl-Homoserinlacton) in a final concentration of 10 mM (Brehm et al., 2003).
Light microscopy was performed on an inverted microscope (Zeiss Axiovert).
Samples for TEM were prepared as described previously (Palinska and Krumbein, 2000).
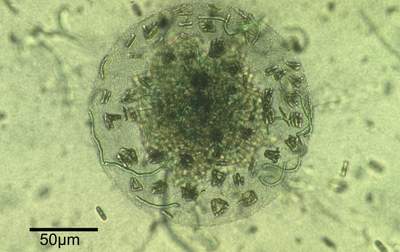
Distinct spherical structures (Fig. 1
![]() ) developed in culture by aggregation of cells of filamentous cyanobacteria (Phormidium sp.), heterotrophic bacteria and benthic diatoms (Navicula perminuta), persist for an extended time and may suggest a symbiotic relationship (Brehm et al.,
2003). Biomicrospheres isolated from a microbial mat of the Wadden Sea
(German Bight) have now been cultured and systematically transferred in the laboratory for more than four years (Brehm et al.,
2003).
Interestingly, the same type of biomicrospheres has also been repeatedly observed and isolated from
fresh, microbial mat samples. Invariably after a cultivation period of two to three weeks a community of one cyanobacterium species (Phormidium sp.), several well defined heterotrophic species of bacteria and a diatom (Navicula perminuta) created biomicrospheres 40-400 µm in
diameter. Under laboratory conditions the first step in the formation of biomicrospheres is the appearance of a thin 1-3 µm thick
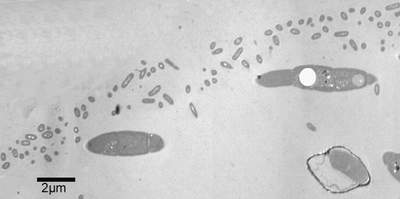
envelope. This spherical envelope is always observed and documented using light- and transmission electron microscopy
(Fig. 2
) developed in culture by aggregation of cells of filamentous cyanobacteria (Phormidium sp.), heterotrophic bacteria and benthic diatoms (Navicula perminuta), persist for an extended time and may suggest a symbiotic relationship (Brehm et al.,
2003). Biomicrospheres isolated from a microbial mat of the Wadden Sea
(German Bight) have now been cultured and systematically transferred in the laboratory for more than four years (Brehm et al.,
2003).
Interestingly, the same type of biomicrospheres has also been repeatedly observed and isolated from
fresh, microbial mat samples. Invariably after a cultivation period of two to three weeks a community of one cyanobacterium species (Phormidium sp.), several well defined heterotrophic species of bacteria and a diatom (Navicula perminuta) created biomicrospheres 40-400 µm in
diameter. Under laboratory conditions the first step in the formation of biomicrospheres is the appearance of a thin 1-3 µm thick
envelope. This spherical envelope is always observed and documented using light- and transmission electron microscopy
(Fig. 2
![]() ) and is probably produced by the heterotrophic
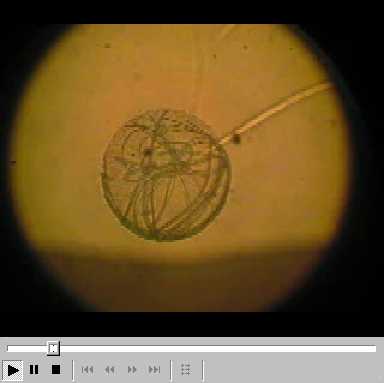
bacteria. When the spheres appear they are recognized by filamentous
cyanobacteria that rapidly approach and forcefully penetrate into them
(see Video file
) and is probably produced by the heterotrophic
bacteria. When the spheres appear they are recognized by filamentous
cyanobacteria that rapidly approach and forcefully penetrate into them
(see Video file ![]() ). All trichomes arrange themselves in the shape of a thin spherical film inside the
biomicrosphere. N. perminuta eventually sneaks in with the Phormidium trichomes and by massive multiplication fill the whole interior of the sphere
(frustules and EPS). After twelve weeks the peripheral part of the cyanobacterial coating of the sphere turn
"sclerotic", i.e. tiny scleres or sclera form a layer at or near the outer surface.
). All trichomes arrange themselves in the shape of a thin spherical film inside the
biomicrosphere. N. perminuta eventually sneaks in with the Phormidium trichomes and by massive multiplication fill the whole interior of the sphere
(frustules and EPS). After twelve weeks the peripheral part of the cyanobacterial coating of the sphere turn
"sclerotic", i.e. tiny scleres or sclera form a layer at or near the outer surface.
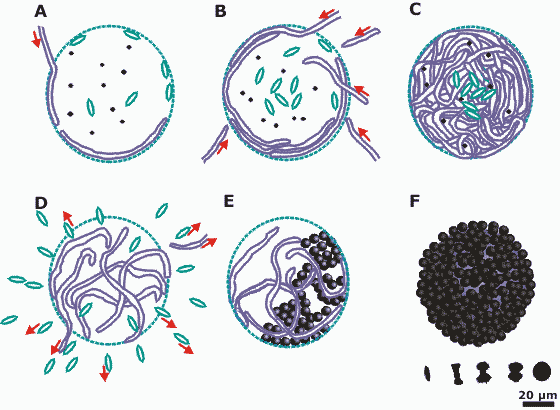
The spheres promote calcification in the surface layers and ultimately produce ooid-like hollow carbonate structures
(Fig. 3 A-F
![]() ). In control runs without microorganisms no carbonate precipitation was
observed.
). In control runs without microorganisms no carbonate precipitation was
observed.
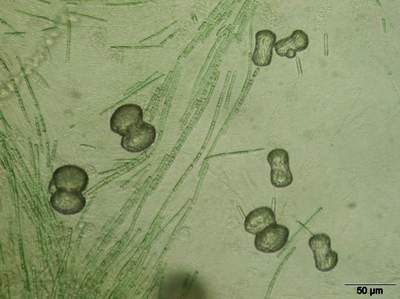
The calcium carbonate precipitates in many
forms: microscopic carbonate needles, wheat seed-shaped grains, small rods,
dumbbells, simple balls and joined balls
(Fig. 4
![]() ). Fractal growth influences not only the size of the fractals but also the inclination of the next generation (Krumbein,
1983; Busch et al., 1999). All stages from sticks to balls can exist at the same time. The solids merge mutually and build
shells and complex structures. In the laboratory these precipitates of carbonate are closely connected with the appearance of structured cyanobacterial assemblages
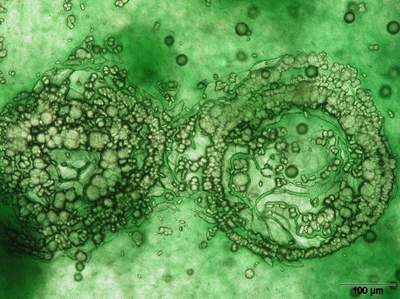
(Fig. 5
). Fractal growth influences not only the size of the fractals but also the inclination of the next generation (Krumbein,
1983; Busch et al., 1999). All stages from sticks to balls can exist at the same time. The solids merge mutually and build
shells and complex structures. In the laboratory these precipitates of carbonate are closely connected with the appearance of structured cyanobacterial assemblages
(Fig. 5
![]() , 6
, 6
![]() ). After two or three months of cultivation the spheres appear as multilayered circular assemblages in which several belts of carbonates are precipitated
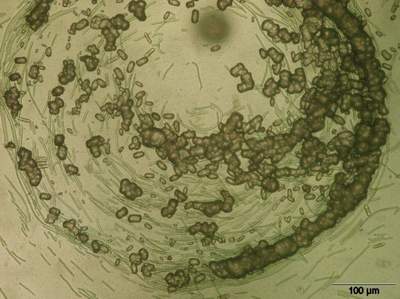
(Fig. 7
). After two or three months of cultivation the spheres appear as multilayered circular assemblages in which several belts of carbonates are precipitated
(Fig. 7
![]() ).
).
The term "oolite" was introduced and defined by Brueckmann (1721) using material collected a few kilometer away from the type locality where the terms "stromatolite" and "ooid" were introduced and defined more than 175 years later by Kalkowsky (1908). Interestingly Brueckmann (1721), Kalkowsky (1908) and Ludwig and Theobald (1852) suggested identical conditions of formation for ooids and stromatolites. However, ooids would often be washed out of stromatolitic microbial mats and deposited elsewhere. The formation of a typical ooid is connected with a nucleation center, which can be of biotic or abiotic origin (Kühl et al., 2003). Here we supply laboratory evidence that the aforementioned authors correctly analyzed the situation despite the vast literature on calm water benthic stromatolites contrasted with agitated water planktonic ooids and oolites. The formation of microbial mat derived ooids and oolites has never before been demonstrated in laboratory experiments.
Our studies were focused on carbonate precipitation in spherical microbial communities
(Fig. 1
![]() ): assemblages of cyanobacteria (Phormidium sp.), heterotrophic bacteria and diatoms (Navicula perminuta) (Brehm,
2001; Brehm et al., 2003).
Precipitation occurs where masses of bacteria are enclosed and concentrated in spherical
envelopes.
): assemblages of cyanobacteria (Phormidium sp.), heterotrophic bacteria and diatoms (Navicula perminuta) (Brehm,
2001; Brehm et al., 2003).
Precipitation occurs where masses of bacteria are enclosed and concentrated in spherical
envelopes.
The envelopes are permeable only for cyanobacteria; other organisms cannot penetrate
them. Within the sphere diatoms accumulate in the centre and cyanobacteria surround
them. In this way a lamination is established, comparable to the lamination of common benthic biofilms. A similar phenomenon of microsphere formation has already been reported by Fox et al.
(1959). They demonstrated that when placed in
water certain proteins spontaneously self-organize into structures, known as microspheres
Calcium carbonates precipitate in the laminations formed by cyanobacteria and associated heterotrophs following the form of
the organism's
organization: in stromatolites as horizontal laminations and in the biomicrospheres studied here as concentric
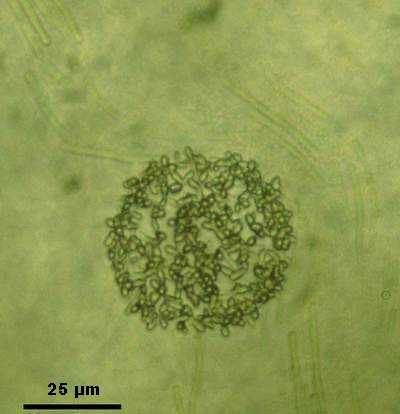
layers. The geometry of the biofilm determines the shape and size of the carbonate layer. Subsequently a calcisphere or
spherulite, composed of numerous small calcite crystals will form; the first step in the development of an ooid
(Fig. 8
![]() ).
).
This structure is comparable to that of fossil oolites and suggests a common genesis for calcispheres and oolites.
Oolites are always built up in several carbonate layers in which each layer represents a separate population of organisms. These fossilized concentric oolitic layers are preserved in carbonate rocks. The main processes in the development of the aggregates studied in the laboratory are limited to diffusion and/or cluster-cluster mechanisms. The aggregates range from 10 to 40 µm in diameter. In sparsely populated areas sparsely disseminated carbonates precipitate. They may be the result of several discrete chemical reactions.
About three months elapse before the first carbonates crystallize in the Petri dishes. The calcification of the biomicrospheres was always observed to start on the spherulitic surface in the form of numerous discrete calcite crystals. Ongoing precipitation leads to the covering of the surface by carbonates. At the same time our control runs without microorganisms never showed any precipitation.
Our investigations indicate that the building of oolites is biologically induced and external nuclei are not necessary to create spherulites. The microorganisms create the basis for their structures in complete independence.
The content of this article was presented originally at the 8th International Symposium on Fossil Algae organized by J.C. Braga and J. Aguirre in Granada, Spain (September, 2003). The authors are most grateful to N.J. Sander (USA) for improvements in the English of the preliminary version. In addition they thank S. Golubic and two anonymous reviewers for their comments and suggestions. The DFG project Pa 842/1-1 is acknowledged as well.
Brehm U. (2001).- Untersuchungen zur mikrobiologisch induzierten Strukturbildung-rezente und fossile Beispiele im Vergleich.- Ph.D. Thesis, Oldenburg, 132 p.
Brehm U., Krumbein W.E., Palinska K.A. (2003).- Microbial spheres: a novel cyanobacterial-diatom symbiosis.- Naturwissenschaften, Berlin, 90, pp. 136-140.
Browne K., Golubic S., Seong-Joo L. (2000).- Shallow marine microbial carbonate deposits. In: Riding E.D., Awramik S.M. (Eds.): Microbial Sediments.- Springer, Berlin, pp. 233-249.
Brueckmann F.E. (1721).- Specimen physicum exhibens historiam naturalem, oolithi seu ovariorum piscium & concharum in Saxa.- Mutatorum, Helmestadii, Salomoni & Schnorrii, 21 p.
Busch S., Dolhaine H., DuChesne A., Heinz S., Hochrein O., Laeri F., Podebrad O., Vietze U., Weiland Th., Kniep R. (1999).- Biomimetic morphogenesis fluorapatite-gelantin composites: Fractal growth, the question of intrinsic electric fields, core/shell assemblies, hollow spheres and reorganization of denatured collagen.- European Journal of Inorganic Chemistry, Weinheim, pp. 1643-1653.
Fox S.W., Harada K., Kendrick J. (1959).- Production of spherules from synthetic proteinoid and hot water.- Science, Washington, 129, p. 1221.
Kalkowsky E. (1908).- Oolith und Stromatolith im Norddeutschen Buntsandstein.- Zeitschrift der Deutschen Geologischen Gesellschaft, Stuttgart, 60, pp. 84-125.
Krumbein W.E. (1983).- Microbial Geochemistry.- Blackwell, Oxford, 330 p.
Kühl M., Fenchel T., Kazmierczak J. (2003).- Growth, structure and calcification potential of an artificial cyanobacterial mat. In: Krumbein W.E., Paterson D.M., Zavarzin G.A. (Eds.): Fossil and recent biofilms.- Kluwer, London, pp. 77-102.
Ludwig R., Theobald G. (1852).- über die Mitwirkung der Pflanzen bei der Abscheidung des kohlensauren Kalkes.- Annalen der Physik und Chemie, Leipzip, 87, pp. 91-107.
Palinska K.A., Krumbein W.E. (2000).- Perforation patterns in filamentous cyanobacteria.- Journal of Phycology, Corvallis, 36, pp. 139-145
Riding R.E., Awramik S.M. (2000).- Microbial sediments.- Springer, Berlin, 331 p.
Rippka R., Deruelles J,. Waterbury J.B., Herdman M., Stanier R.Y. (1979).- Generic assignments, strain histories and properties of pure cultures of cyanobacteria.- Journal of General Microbiology, Reading, Vol. 111, pp. 1-61.
Visscher P.T., Reid R.P., McKenzie J.A., Vasconcelos C. (2002).- Geomicrobial mechanisms of carbonate precipitation: Novel insights from laminated structures.- Abstract, 12th Annual Goldschmidt Conference 2002, Davos, Switzerland.- Geochimica et Cosmochimica Acta, Oxford, 66 (15A), Suppl. 1, p. 808.
Click on thumbnail to enlarge the image.
Figure 1: Microbial biosphere. Assemblage of filamentous cyanobacteria, heterotrophic bacteria and diatoms within a sphere. Diameter of the sphere 166 µm.
Click on thumbnail to enlarge the image.
Figure 2: Transmission electron microscopy of the outer part of the sphere. The envelope consisting of heterotrophic bacteria and their excretes form the surface of the sphere. Below the envelope two filaments of Phormidium sp. and one diatom (lower right side) are documented. The space between organisms is filled by EPS (Extracellular Polymeric Substances).
Click on thumbnail to enlarge the image.
Figure 3: Schematic biomicrosphere development stages. Cyanobacteria are represented as long filaments, diatoms as rice-shaped grains, and hetrotrophic bacteria as small dots. Five different stages of carbonate crystallization are shown at the right, lower edge. The final stage of the carbonate crystallization appears in E and F. A-B Initial stage; Cyanobacteria passing through the spherical, bacterial envelope and encompassing the diatoms. C Densely packed biomicrosphere with diatoms concentrated in the centre and cyanobacteria surrounding them. D Late stage of the biomicrosphere. Diatoms and some cyanobacteria abandon the sphere. E-F Carbonate precipitation following the emplacement of cyanobacteria. F Calcified biomicrosphere.
Click on thumbnail to enlarge the image.
Figure 4: Different stages of carbonate deposition in biomicrospheres. In the background filaments of cyanobacteria are visible.
Click on thumbnail to enlarge the image.
Figure 5: Calcium carbonate precipitate, composed of numerous spherically arranged layers of small calcite crystals. Intergrown crystals are connected to cyanobacterial filaments.
Click on thumbnail to enlarge the image.
Figure 6: Concentric layers of calcium carbonate following the cyanobacterial orientation.
Click on thumbnail to enlarge the image.
Figure 7: Late stage of calcification. Two coalescent spheres of carbonate grains.
Click on thumbnail to enlarge the image.
Figure 8: Spherulitic surface of the biomicrosphere. Numerous, discrete calcite crystals are visible.
Click on thumbnail to play a low-resolution video file (wmv =
762 KB)
or click on the icon ![]() to play a high-resolution video file
(mpg =
8,606 KB)
to play a high-resolution video file
(mpg =
8,606 KB)
or click on the icon ![]() to
download another high-resolution video file (avi = 4,774 KB)
to
download another high-resolution video file (avi = 4,774 KB)
Video File: Cyanobacterial movement inside a biomicrosphere.