◄ Carnets Geol. 4 (L04) ►
![]()
Early Life Institute, Key Laboratory for Continental Dynamics of the
Ministry Education, Northwest University, Xi'an, 710069 (P.R. China)
Early Life Institute and Department of Geology, Northwest University, Xi'an,
710069; and School of Earth Sciences and Resources, China University of
Geosciences, Beijing, 100083 ( P.R. China).
Early Life Institute and Department of Geology, Northwest University, Xi'an,
710069 ( P.R. China)
Early Life Institute and Department of Geology, Northwest University, Xi'an,
710069 ( P.R. China)
Manuscript online since October 28, 2004
![]()
A succession of developmental types in the lophophores of lingulid brachiopods is reported from the Early Cambrian Chengjiang deposits of South China. These types range from trocholophe, schizolophe to simple coiled spirolophe. Of Atdabanian age, this succession of forms is mirrored in the ontogeny of the lophophore of Recent linguloids, thus demonstrating a close similarity in the development of the lophophores of Cambrian and Recent forms. We illustrate also more than 10 extraordinary specimens with aberrant dispositions of the lophophoral arms that extend to unusual lengths either inside or outside the shell.
Linguloidea;
Brachiopoda;
Lophophore;
Lagerstätte;
Chengjiang;
China;
Early Cambrian;
Atdabanian.
Z.F., D.G., J., J.N. (2004).- New data on the lophophore anatomy of Early Cambrian linguloids from the Chengjiang Lagerstätte, Southwest China.- Carnets Geol., Madrid, vol. 4, no. L04, 7 p. DOI: 10.4267/2042/310
Nouvelles données sur l'anatomie du lophophore chez des linguloïdes du Cambrien inférieur du Lagerstätte de Chengjiang, Sud-Ouest de la Chine.- Une succession de types de lophophore est décrite chez des brachiopodes linguloïdes dans les dépôts du Chengjiang (Cambrien inférieur) au Sud de la Chine. Ces types vont depuis un lophophore trocholophe, au schizolophe et au spirolophe à enroulement spiral simple. Cette succession d'âge Atdabanien existe encore dans le développement du lophophore chez les linguloïdes actuels, ce qui démontre une similarité entre les lophophores au Cambrien et dans l'Actuel. Des dispositions aberrantes des bras lophophoraux qui tendent à s'étendre de façon inhabituelle dans et hors de la coquille sont illustrées dans une dizaine d'exemplaires.
Linguloidea ;
Brachiopoda ;
Lophophore ;
Lagerstätte ;
Chengjiang ;
Chine ;
Cambrien inférieur ;
Atdabanien.
Brachiopoda and Phoronida, together with Ectoprocta possess a tentacular, ciliated, feeding organ, the lophophore, and thus sometimes have been grouped as Lophophorata (, 1977, 1997a). Based on traditional morpho-anatomical characters, brachiopods and phoronids are undoubtedly closely related, and usually regarded as sister groups among deuterostomes (, 2001, 2002). However, molecular phylogenetic studies confirm phoronid-brachiopod monophyly and establish their affinities with protostomes (see among others et al., 1995; et al., 1996; et al., 1998). Brachiopods are exclusively marine bivalved lophophorates. As a clade, the most distinctive feature of this group that supposedly differentiates them from the other lophophore-bearing animals is a filtration system in an isolated cavity (, 1992). However, in view of molecular phylogenetic studies this classification seems no longer tenable. et al. (1998) and (2000) suggest that the phylum Phoronida should be regarded as a subphylum of shell-less brachiopods named Phoroniformea, though this view has been questioned ( and , 2001; , 2002, 2003).
The brachiopod lophophore functions principally as a ciliary pump and feeding organ by creating currents of water through out the mantle cavity that serve for both the capture of food particles and the uptake of oxygen for respiration (, 1976). The shape and disposition of the lophophore are apposite with both the inhalant and exhalant "apertures" of brachiopods and with the orientation of the brachiopod shell (, 1992). As in the phoronids the brachiopod lophophore is composed of a pair of long lophophoral arms or brachia, and is not capable of much extension, even in the inarticulate forms where its musculature is best developed. The brachia are disposed symmetrically about the mouth and variously looped or coiled (, 1992). They are attached to the anterior body wall, sometimes to the dorsal mantle surface. The spirolophe is characteristic of most inarticulate brachiopods. An adult spirolophe must go through trocholophous and schizolophous stages during its development (, 1992). However, fossil records hitherto have provided no information regarding this ontogenic process, albeit some well-defined lophophore tissues were reported recently from the Lower Cambrian of China (Z.F. et al., 2004; X.G. et al., 2003).
In this paper, we provide additional information on the ontogeny of the lophophore in Early Cambrian lingulids, and illustrate some aberrant dispositions of lophophores that differ markedly from those depicted by Z.F. et al. (2004).
All the specimens show typical Chengjiang preservation (
et al., 1991). As in the phoronids, after weathering the fossils are
reddish or dark yellow and are embedded in grayish-green and grayish-yellow
mudstone intercalated with grayish-brown silty shale. All of them were recovered
from the Ercai village and Hill Jianshan localities at Haikou, Kunming, where
the earliest-known agnathan Haikouichthys ( et al.,
2003a) and
the enigmatic yunnanozoan Haikouella jianshanensis ( et al.,
2003b) were found. The strata containing the soft-bodied fossils belong to the
Yu'anshan Member (Eoredlichia Zone), i.e. the upper part of Early
Cambrian Heilinpu (formerly Qiongzhusi) Formation, exposed in a wide area around
Kunming, Yunnan. Details of the localities and stratigraphy were given in X.L.
et al. (2001). So far, nearly 1,000 specimens of the two lingulid taxa,
Lingulella chengjiangensis and Lingulellotreta malongensis, have
been collected from these localities by the work-team of the Early Life
Institute (prefixed ELI); all the specimens are deposited in ELI, Northwest
University, Xi'an, China. In our lingulid collections, around 80 specimens
reveal partial or paired lophophoral imprints, in some cases with well-defined
tentacles (Z.F. et al., 2004). In this collection there are only 20
specimens where the lophophoral imprints do not display the usual spiral coil
(Z.F. et al., 2004). Of these apparently aberrant dispositions of
lophophores, 6 appear rounded (Fig. 1 A
![]() - B
- B
![]() and I
and I
![]() -
J
-
J
![]() ), 2 are crescent-shaped
with an introversion of the anterior edge (Fig. 1 C
), 2 are crescent-shaped
with an introversion of the anterior edge (Fig. 1 C
![]() and
K
and
K
![]() ), and the remaining
lophophores seem to display some extension of the lophophoral arms which are
either elongated inside, or protrude from the shell (Fig. 2 A-J
), and the remaining
lophophores seem to display some extension of the lophophoral arms which are
either elongated inside, or protrude from the shell (Fig. 2 A-J
![]() ).
).
The lophophore of Lingulellotreta malongensis has
recently been documented by Z.F. et al. (2004), who considered it to
correspond to an early spirolophe stage. However, new material from the same
localities shows that some lophophores of L. malongensis do not
invariably take the shape of simple spiral coils as depicted by Z.F. et
al. (2004). They appear either as rounded or crescent-shaped loops
distinctly delimited by two concentric brownish strands (Fig. 1 A
![]() - B
- B
![]() and I
and I
![]() -
J
-
J
![]() ).
This disposition is closely reminiscent of a taxolophe-trocholophe stage of
development (, 1992). In Fig. 1 C
).
This disposition is closely reminiscent of a taxolophe-trocholophe stage of
development (, 1992). In Fig. 1 C
![]() a rounded lophophore has an anterior-median
indentation, dividing the lophophore into two lobes: it probably represents a
schizolophe stage of development (Fig. 1 C
a rounded lophophore has an anterior-median
indentation, dividing the lophophore into two lobes: it probably represents a
schizolophe stage of development (Fig. 1 C
![]() and
K
and
K
![]() ). These lingulid shells
(Fig. 1 A
). These lingulid shells
(Fig. 1 A
![]() - B
- B
![]() - C
- C
![]() ) appear to be smaller (approximately 5mm long) and shorter than those with
an early spirolophe (Z.F. et al., 2004). Based on a single specimen,
the lophophore of the obolid Lingulella chengjiangensis has also been
described by X.G. et al. (2003) as a schizolophe stage. However, in the
light of evidence from more than 20 specimens of L. chengjiangensis
numbered ELI C-0030-0146, the lophophores of L. chengjiangensis, like
those of Lingulellotreta malongensis, include trocholophes (Fig. 1
F
) appear to be smaller (approximately 5mm long) and shorter than those with
an early spirolophe (Z.F. et al., 2004). Based on a single specimen,
the lophophore of the obolid Lingulella chengjiangensis has also been
described by X.G. et al. (2003) as a schizolophe stage. However, in the
light of evidence from more than 20 specimens of L. chengjiangensis
numbered ELI C-0030-0146, the lophophores of L. chengjiangensis, like
those of Lingulellotreta malongensis, include trocholophes (Fig. 1
F ![]() ),
schizolophes (X.G. et al., 2003) and the less intricately coiled
spirolophes (Fig. 1 G
),
schizolophes (X.G. et al., 2003) and the less intricately coiled
spirolophes (Fig. 1 G
![]() - H
- H
![]() and
L
and
L ![]() ). All these types of lophophores are mirrored in the
ontogeny of the lophophores of Recent lingulids (, 1992). Lophophore
ontogeny in Cambrian lingulids is, therefore, homologous to that of extant
lingulids, and has remained fairly constant over 530 Ma. Thus, the early stages
of lophophoral ontogeny and development appear to be plesiomorphic characters
inherited from an ancestral form.
). All these types of lophophores are mirrored in the
ontogeny of the lophophores of Recent lingulids (, 1992). Lophophore
ontogeny in Cambrian lingulids is, therefore, homologous to that of extant
lingulids, and has remained fairly constant over 530 Ma. Thus, the early stages
of lophophoral ontogeny and development appear to be plesiomorphic characters
inherited from an ancestral form.
Another noteworthy fact, identified in more than 10 specimens
(see: Fig. 2 A-J
![]() ) is lophophoral arms that are either elongated inside or extend
outside the shell. In specimen ELI C-0078 (Fig. 2 A
) is lophophoral arms that are either elongated inside or extend
outside the shell. In specimen ELI C-0078 (Fig. 2 A
![]() and G
and G
![]() ) paired lophophoral
imprints arise from the anterior-medial visceral region. One extends forward and
curves outward inside the shell, and the other projects laterally outward, and
then curves backward outside the shell. On the outer portion of this lophophoral
arm are some reddish-brown grains that may represent organic particles
interpreted as nutrient, or at least particles filtered by the tentacles. In
specimen ELI C-0040 (Fig. 2 B
) paired lophophoral
imprints arise from the anterior-medial visceral region. One extends forward and
curves outward inside the shell, and the other projects laterally outward, and
then curves backward outside the shell. On the outer portion of this lophophoral
arm are some reddish-brown grains that may represent organic particles
interpreted as nutrient, or at least particles filtered by the tentacles. In
specimen ELI C-0040 (Fig. 2 B
![]() and H
and H
![]() ), a pair of lophophoral tubes protrude
from both sides of the shell. They are disposed symmetrically about the midline.
In the center of the paired lophophoral axes (Fig. 2 A
), a pair of lophophoral tubes protrude
from both sides of the shell. They are disposed symmetrically about the midline.
In the center of the paired lophophoral axes (Fig. 2 A
![]() - B
- B
![]() and G
and G
![]() - H
- H
![]() ) runs a dark
strand. It is taken to mark the hollow lophophoral canal (or lophophoral
coelomic cavity) (Fig. 2 A
) runs a dark
strand. It is taken to mark the hollow lophophoral canal (or lophophoral
coelomic cavity) (Fig. 2 A
![]() - B
- B
![]() and G
and G
![]() - H
- H
![]() ). In Fig. 2 C
). In Fig. 2 C
![]() - D
- D
![]() , one lophophoral
arm outside the shell appears as a coiled counterclockwise inwards imprint (Fig. 2 C
, one lophophoral
arm outside the shell appears as a coiled counterclockwise inwards imprint (Fig. 2 C
![]() - D
- D
![]() and I
and I
![]() ); the aspect of the other arm is unknown because it is absent in the
broken specimen. Nonetheless, it is in the aberrant single lophophoral axis that
the tentacles are clearly visible, and closely arranged in a single palisade (Fig. 2 C
); the aspect of the other arm is unknown because it is absent in the
broken specimen. Nonetheless, it is in the aberrant single lophophoral axis that
the tentacles are clearly visible, and closely arranged in a single palisade (Fig. 2 C
![]() - D
- D
![]() and I
and I
![]() ). As seen in
Fig. 2 E
). As seen in
Fig. 2 E
![]() and J
and J
![]() , one lophophoral arm appears freely
coiled, whereas the other one apparently spreads antero-laterally inside the
shell (Fig. 2 E
, one lophophoral arm appears freely
coiled, whereas the other one apparently spreads antero-laterally inside the
shell (Fig. 2 E
![]() and J
and J
![]() ). In addition, in the specimen ELI L-0127 the paired
lophophoral arms appear to cross each other (Fig. 2 F
). In addition, in the specimen ELI L-0127 the paired
lophophoral arms appear to cross each other (Fig. 2 F
![]() ). However, none of the
brachiopod shells were crushed and deformed laterally, and their marginal edge
is also well preserved. The state of preservation suggests that the animal is
most probably in an undisturbed life position, possibly as a result of
storm-induced rapid burial. This is likely because the organic component of the
shell is so quickly hydrolysed or digested by microbes and the exposed apatitic
framework so easily abraded hydro-dynamically (, 1990), that fossilization
of lingulid shells could only have taken place after catastrophic changes
provoking rapid burial were developed in the post-mortem environment (,
1997b).
). However, none of the
brachiopod shells were crushed and deformed laterally, and their marginal edge
is also well preserved. The state of preservation suggests that the animal is
most probably in an undisturbed life position, possibly as a result of
storm-induced rapid burial. This is likely because the organic component of the
shell is so quickly hydrolysed or digested by microbes and the exposed apatitic
framework so easily abraded hydro-dynamically (, 1990), that fossilization
of lingulid shells could only have taken place after catastrophic changes
provoking rapid burial were developed in the post-mortem environment (,
1997b).
The aberrant disposition of lophophores is not easily
explained. Several hypotheses may be proposed. The atypical forms may have been
caused by oxygen deprivation during a living burial produced by turbidity
currents ( and W.T. , 2001). The symmetrical protrusion of both brachia
from the shell (Fig. 2 B
![]() and H
and H
![]() ) suggests this conjecture. However, this proposal
is considered less likely because of the fact, as seen in Fig. 2 E
) suggests this conjecture. However, this proposal
is considered less likely because of the fact, as seen in Fig. 2 E
![]() and J
and J
![]() , that one of lophophoral arms is coiled as usual while the other appears
to form an elongated extension. Furthermore, in one specimen (Fig. 2 A
, that one of lophophoral arms is coiled as usual while the other appears
to form an elongated extension. Furthermore, in one specimen (Fig. 2 A
![]() and G
and G
![]() ),
both the lophophoral arms are broadly spread. They extend asymmetrically toward
one side and then protrude out of the shell, rather than diverge rapidly toward
two sides (Fig. 2 A
),
both the lophophoral arms are broadly spread. They extend asymmetrically toward
one side and then protrude out of the shell, rather than diverge rapidly toward
two sides (Fig. 2 A
![]() and G
and G
![]() ). Such a disposition appears truly aberrant
compared with the normal (Fig. 1 H
). Such a disposition appears truly aberrant
compared with the normal (Fig. 1 H
![]() and
L
and
L ![]() ). In addition, this interpretation of the
imprints may be less tenable if the paired lophophoral arms cross each other
(Fig. 2 F
). In addition, this interpretation of the
imprints may be less tenable if the paired lophophoral arms cross each other
(Fig. 2 F
![]() ).
The distinctive layout of lophophoral arms may be the result of
post-mortem relaxation and re-distribution, possibly consequent on the
weight of a layer of covering silt for instance. This seems a simple
explanation, but may be inconsistent with the coiled shape of most lophophores
(Fig. 1 A-L
).
The distinctive layout of lophophoral arms may be the result of
post-mortem relaxation and re-distribution, possibly consequent on the
weight of a layer of covering silt for instance. This seems a simple
explanation, but may be inconsistent with the coiled shape of most lophophores
(Fig. 1 A-L
![]() ). In addition, post-mortem change cannot account for the
different degrees of spread in the brachia of the several discrete animals
because their integuments were not crushed or strongly distorted laterally,
although flattened dorso-ventrally. The most probable hypothesis is that the
lophophores of Cambrian lingulids were capable of some extensional movement.
Both lophophoral arms could stretch inside and freely extend out of the mantle
cavity under certain specific conditions. The known series of aberrant
lophophore patterns (Fig. 2
). In addition, post-mortem change cannot account for the
different degrees of spread in the brachia of the several discrete animals
because their integuments were not crushed or strongly distorted laterally,
although flattened dorso-ventrally. The most probable hypothesis is that the
lophophores of Cambrian lingulids were capable of some extensional movement.
Both lophophoral arms could stretch inside and freely extend out of the mantle
cavity under certain specific conditions. The known series of aberrant
lophophore patterns (Fig. 2
![]() ) accords well with this supposition, so it may be a
valid explanation of the phenomena observed. The presence of nutrient particles
on the outer lophophoral arm (Fig. 2 A
) accords well with this supposition, so it may be a
valid explanation of the phenomena observed. The presence of nutrient particles
on the outer lophophoral arm (Fig. 2 A
![]() and G
and G
![]() ) and the normal (counterclockwise)
coil of the lophophoral arm outside the shell (Fig. 2 C
) and the normal (counterclockwise)
coil of the lophophoral arm outside the shell (Fig. 2 C
![]() - D
- D
![]() and I
and I
![]() ) are congruent
with a probable extensional motion of the lophophore for nutrient capture.
) are congruent
with a probable extensional motion of the lophophore for nutrient capture.
Brachiopods and phoronids were commonly grouped together with bryozoans (, 1959; , 1977, 1997a; , 1995, 1996; et al., 1995), and sometimes with pterobranch hemichordates (, 1987, 2001, 2002; , 2002) because they possess a ciliated tentacular structure that invades the mesocoelomic cavity and surrounds the mouth but not the anus. This grouping is based primarily on the inferred homology of their ciliated tentacular feeding structures (, 1959; , 1996; , 1987, 2002). 18S rDNA sequence data suggest, however, that pterobranchs and lophophorates (consisting of the brachiopods, phoronids, and bryozoans) are in different metazoan subkingdoms and are not closely related (, 1995, 1996; et al., 1996). If so, the shared possession of a lophophore-like apparatus may be the result of evolutionary convergence (, 1996). Within the Bilateria the placement of the lophophorate bryozoans is perhaps the most puzzling (, 2002; et al., 1995; et al., 2000; and , 2001; and , 2004). Based on morphological analyses (, 1987, 2001, 2002), bryozoans were repeatedly separated from the phoronid-brachiopod clade, and are not monophyletic with brachiopods and phoronids. Nevertheless, the filter-feeding process in phoronids and bryozoans is remarkably similar (, 2002; and , 2002). Morphological analyses indicate indisputably that brachiopods and phoronids are closely related (, 2001, 2002). They share numerous developmental and anatomical traits that are largely in accordance with the molecular phylogenetic data ( et al., 1995; et al., 1998; et al., 1998; , 2000).
The series of forms in the development of lophophores during
Atdabanian times (see Fig. 1
![]() ) indicate that Early Cambrian lingulids are
represented in the larval-juvenile developmental stage of their Recent
relatives. Hence, their inhalant and exhalant currents could resemble those of
the schizolophous and early spirolophous stages (, 1974; ,
1992), and
most probably differ from the three apertures (the central for exhalant current
water, and the outer two for inhalant current) of Recent Lingula because
of the absence of a pointed medially lophophore filter cone and three
pseudosiphons of setae. In addition, the fossils illustrated herein (Fig. 1
) indicate that Early Cambrian lingulids are
represented in the larval-juvenile developmental stage of their Recent
relatives. Hence, their inhalant and exhalant currents could resemble those of
the schizolophous and early spirolophous stages (, 1974; ,
1992), and
most probably differ from the three apertures (the central for exhalant current
water, and the outer two for inhalant current) of Recent Lingula because
of the absence of a pointed medially lophophore filter cone and three
pseudosiphons of setae. In addition, the fossils illustrated herein (Fig. 1
![]() - 2
- 2
![]() )
indicate that like Recent linguliforms, Early Cambrian lingulids lack median
tentacles. Lophophore palisades and the brachial lip in the trocholophe stage
are of critical importance to an understanding of brachiopod phylogeny (
et al., 1995). Unfortunately, data regarding these aspects cannot be
confidently demonstrated in the collected specimens. Continuing investigation of
the unique Chengjiang deposits may yield information bearing on these
issues.
)
indicate that like Recent linguliforms, Early Cambrian lingulids lack median
tentacles. Lophophore palisades and the brachial lip in the trocholophe stage
are of critical importance to an understanding of brachiopod phylogeny (
et al., 1995). Unfortunately, data regarding these aspects cannot be
confidently demonstrated in the collected specimens. Continuing investigation of
the unique Chengjiang deposits may yield information bearing on these
issues.
The aberrant lophophore layouts discussed herein suggest
extensibility in the Cambrian lingulid lophophore under certain stringent
conditions. The extended paired feeding apparatuses (Fig. 2 A
![]() and G
and G
![]() ) are
surprisingly reminiscent of lophophore-like tentacular feeding arms of
rhabdopleurid pterobranchs. This high degree of ostensible similarity presumably
reflects a certain selective pressure on these brachiopod taxa from the
surroundings at that time, which resulted in morphological convergence
(, 1996). Brachiopods have two coelomic cavities in the lophophore, a
large brachial canal which is restricted to the base of the lophophore and a
small brachial canal with extension into each tentacle (, 2002). In the
fossil brachiopods (Fig. 2 A
) are
surprisingly reminiscent of lophophore-like tentacular feeding arms of
rhabdopleurid pterobranchs. This high degree of ostensible similarity presumably
reflects a certain selective pressure on these brachiopod taxa from the
surroundings at that time, which resulted in morphological convergence
(, 1996). Brachiopods have two coelomic cavities in the lophophore, a
large brachial canal which is restricted to the base of the lophophore and a
small brachial canal with extension into each tentacle (, 2002). In the
fossil brachiopods (Fig. 2 A
![]() - B
- B
![]() and G
and G
![]() - H
- H
![]() ) the large brachial canal appear to be
visible as a dark lineation in the centre of either lophophoral axis, and the
presence of the small brachial canal is suggested by the well-defined tentacles
(Fig. 2 C
) the large brachial canal appear to be
visible as a dark lineation in the centre of either lophophoral axis, and the
presence of the small brachial canal is suggested by the well-defined tentacles
(Fig. 2 C
![]() - D
- D
![]() and I
and I
![]() ). However, Phoronids have a single coelomic cavity in the
lophophore, that lacks a brachial lip as well as cartilage-like connective
tissue ( et al., 1995; ,
2002). Thus it is unlikely that
the phoronid lophophore represents a derivation of the brachiopod lophophore and
if the molecular data are correct ( et al., 1995; et
al., 1998, 2003; ,
2000; and ,
2004), it is
reasonable to assume that the respiratory function of the lophophore of the
Brachiopoda now known only in an isolated cavity (the shell) could be an
apomorphy derived from a stem lophophore ancestor.
). However, Phoronids have a single coelomic cavity in the
lophophore, that lacks a brachial lip as well as cartilage-like connective
tissue ( et al., 1995; ,
2002). Thus it is unlikely that
the phoronid lophophore represents a derivation of the brachiopod lophophore and
if the molecular data are correct ( et al., 1995; et
al., 1998, 2003; ,
2000; and ,
2004), it is
reasonable to assume that the respiratory function of the lophophore of the
Brachiopoda now known only in an isolated cavity (the shell) could be an
apomorphy derived from a stem lophophore ancestor.
We are grateful to Dr. Susan (Queensland Museum) and Dr. Nestor J. for helpful revision of the draft manuscript and improving the English. Special thanks are due to Simon for examination of the fossil lingulid photographs and for advice in Xi'an. The first author (Z.F. ) owes thanks to Dr. Bernard L. for kindly suggestions and to Dr. Leonid for some important publications. The manuscript was reviewed by Dr. Christian C. , and Leonid , who greatly improved it. Technical assistance from Professor S. J. , N. in Northwest University, Xi'an, is acknowledged. We also thank H.X. , Y.B. for help with fieldwork, and J.P. , M.R. for preparation of the fossils. This work is supported by the National Natural Science Foundation of China (programs 40332016 and 30270207) and the Ministry of Sciences and Technology of China (program G. 2000077702).
L.E. & W.T. (2001).- Stratigraphy, paleontology, and depositional setting of the Chengjiang Lagerstätte (Lower Cambrian), Yunnan, China.- Palaeoworld, Nanjing, vol. 13, p. 66-86.
S.H. (1974).- Observations on the ciliary feeding mechanism of the brachiopod Crania anomala.- Journal of Zoology, London, vol. 173, pp. 441-449.
B.L. (2000).- Monophyly of brachiopods and phoronids: reconciliation of molecular evidence with Linnaean classification (the subphylum Phoroniformea nov.).- Proceedings of the Royal Society, London, Series B, vol. 267, pp. 225-231.
B.L., A. & T. (1998).- Molecular phylogeny of brachiopod and phoronids based on nuclear-encoded small subunit ribosomal RNA gene sequences.- Proceedings of the Royal Society, London, Series B, vol. 353, pp. 2039-2061.
B.L., L.E. & C. (2003).- The brachiopod fold: A neglected body plan hypothesis.- Palaeontology, vol. 46, n° 1, pp. 59-65.
C.C. (1976).- Le lophophore structure significative des Lophophorates - Brachiopoda, Bryozoa, Phoronida.-Zoologica Scripta, vol. 5, pp. 133-137.
C.C. (1977).- Un nouvel embranchement: les Lophophorates.- Bulletin de la Société zoologique de France, vol. 102, pp. 341-344.
C.C. (1990).- Examples of post-mortality alteration in Recent brachiopod shells and (paleo) ecological consequences.- Marine Biology, vol. 104, pp. 233-238.
C.C. (1992).- Functional disposition of the lophophore in living Brachiopods.- Lethaia, vol. 25, pp. 291-302.
C.C. (1997a).- Les Lophophorates constituent-ils un embranchement ?- Bulletin de la Société zoologique de France, vol. 122, pp. 279-288.
C.C. (1997b).- Ecology of inarticulated brachiopods. In: R.L. (eds.), Treatise on Invertebrate Paleontology, Part H, Brachiopoda.- Geol. Soc. Amer. & Univ. Kansas Press, Boulder, Kansas, vol. 1, pp. 473-495.
A., D., M. & K. (1998).- Molecular phylogeny of metazoan intermediate filament proteins.- Journal of Molecular Evolution, vol. 47, pp. 751-762.
G., D.L., M., W. & W.C. (2000).- Triploblastic relationships with emphasis on the acoelomates and the position of Gnathostomulida, Cycliophora, Plathelminthes and Chaetognatha: A combined approach of 18S rDNA sequecnces and morphology.- Systematic Biology, v. 49, pp. 539-562.
K.M. (1995).- The phylogenetic position of the pterobranch Hemichordates based on 18S rDNA sequence data.- Molecular phylogenetics and Evolution. vol. 4, n° I, pp. 72-76.
K.M. (1996).- Convergence in the feeding apparatuses of Lophophorates and pterobranch hemichordates revealed by 18S rDNA: An interpretation.- Biological Bulletin, vol. 190, pp. 1-5.
K.M., J.D., A.M.A., S.M., D.M. & J.A. (1995).- Evidence from 18S ribosomal DNA that the lophophorates are protostome animals. Science, vol. 267, pp. 1641-1643.
X.G., L. & J. (1991).- Composition and preservation of the Chengjaing fauna a Lower Cambrian soft-bodied biota.- Zoologica Scripta, vol. 20, pp. 395-411.
L.E., L.E., M.G. & J. (1995).- Phylogenetic analysis and ordinal classification of the brachiopoda.- Palaeontology, vol. 38, n° 4, 713-741.
L.H. (1959).- The invertebrates: Smaller coelomate groups, Chaetognatha, Hemichordata, Pogonophora, Phoronida, Ectoprocta, Brachiopoda, Sipunculida, the Coelomate Bilateria.- McGraw-Hill, New York.
P.S. & H.U. (2002).- On ciliary sieving and pumping in bryozoans.- Journal of Sea Research, vol. 48, pp. 181-195.
L.Y., B., R.D., U., P. & J.R. (1996).- 18S rDNA suggests that Entoprocta are protostomes, unrelated to Ectoprocta.- Journal of molecular Evolution, vol. 42, pp. 552-559.
C. (1987).- Structure and function of metazoan ciliary bands and their phylogenetic significance.- Acta zoologica, vol. 68, pp. 205-262.
C. (2001).- Animal evolution: Interrelationships of the living phyla, 2nd edition. Oxford Univ. Press, Oxford. pp. 385-406.
C. (2002).- The phylogenetic position of Entoprocta, Ectoprocta, Phoronida, and Brachiopoda.-Integrative and Comparative Biology, vol. 42, n° 3, pp. 685-691.
C. (2003).- Defining phyla: morphological and molecular clues to metazoan evolution.- Evolution and Development, vol. 4, pp. 386-393.
Y.L. & K.M. (2004).- Evidence from Hox genes that bryozoans are lophotrochozoans.- Evolution and Development, vol. 6, n° 4, pp. 275-281.
K. & D.J. (2001).- Animal phylogeny and the ancestry of bilaterians: inferences from morphology and 18S rDNA sequences.- Evolution and Development, vol. 3, pp. 170-205.
L.E. (1992).- The Cambrian radiation of brachiopods. In: J.H. and P.W. (eds.), Origin and early evolution of the Metazoa.- Plenum Press, New York, pp. 399-423.
H.U. (2002).- Methods of ciliary filter feeding in adult Phoronis muelleri (phylum Phoronida) and in its free-swimming actinotroch larva.- Marine Biology, vol. 141, pp. 75-87.
D.G., S., J., Z.F., K., P., L., X.L., J.N., Y. & H.Q. (2003a).- Head and backbone of the Early Cambrian vertebrate Haikouichthys. Nature, vol. 421, pp. 526-529.
D.G., S., Z.F., J.N., J., L., X.L., K. & Y. (2003b).- A new species of yunnanozoan with implications for deuterostome evolution. Science, vol. 299, pp. 1380-1384.
X.G., X.G. & C.C. (2003).- Evidence of lophophore diversity in Early Cambrian Brachiopoda.- Proceedings of the Royal Society, London, Series B, (Suppl.), vol. 270, pp. S65-S68.
X.L., D.G., Y. & J. (2001).- New sites of Chengjiang fossils: crucial windows on the Cambrian explosion.- Journal of the Geological Society, vol. 158, pp. 211-218.
Z.F., J., X.L., J.N. & D.G. (2004).- Soft tissue preservation in the Lower Cambrian linguloid brachiopod from South China.- Acta Palaeontologica Polonica, vol. 49, n° 2, pp. 259-266.
Click on thumbnail to enlarge the image.
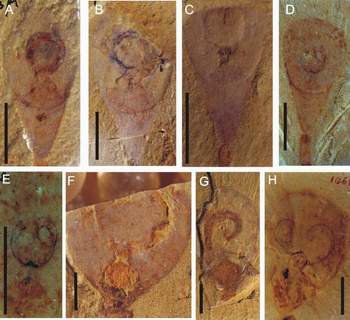
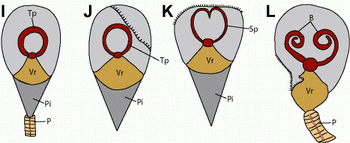
Figure 1: Distinctive disposition of lingulid lophophores from the Lower Cambrian Chengjiang deposits, South China. (A-E) lophophores of the lingulellotretid Lingulellotreta malongensis. (F-H) lophophores of the obolid Lingulella chengjiangensis. (I-L) interpretative drawings of figures shown here. (A) ELI L-0137A, a circular trocholophore imprint; (B) ELI L-0157, an impressed trocholophore imprint; (C) ELI L-0290, a schizolophe imprint with an anterior-median indentation; (D-E) ELI L-0081, a juvenile spirolophe; (E) enlargement of the lophophore of (D); (F) ELI C-0188B, imprint of a recessed round trocholophore; (G) ELI C-0030B, a single coiled lophophoral arm; (H) ELI C-0146B, showing freely coiled paired brachia; (I) Sketch of (A); (J) Sketch of (B); (K) Sketch of (C); (L) Sketch of (H).
Abbreviations: B, lophophoral arm; Vr, visceral region; Pi, pseudointerarea; P, pedicle; Tp, trocholophe; Sp, schizolophe. Scale bars 2 mm.
Click on thumbnail to enlarge the image.
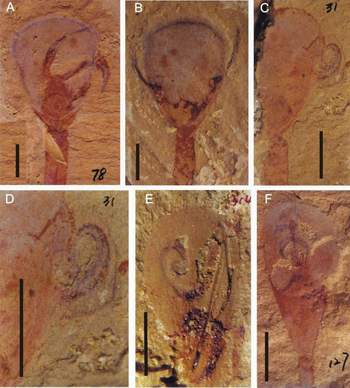
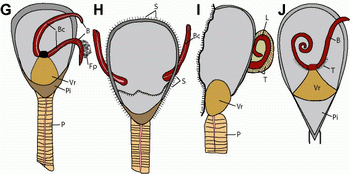
Figure 2: Aberrant disposition of lingulid lophophores from the Lower Cambrian, China. (A-D) of Lingulella chengjiangensis. (E-F) of Lingulellotreta malongensis. (G-J) interpretative drawings. (A) ELI C-0078, note the extended paired lophophoral arms, with the right one apparently outside the shell; (B) ELI C-0040, showing the symmetrically protruded lophophoral tubes and finely impressed setal fringe; (C) ELI C-0031, imperfectly preserved specimen, showing a coiled lophophore protruding from the shell; (D) details of the lophophore in (C), showing the well-defined tentacles; (E) ELI L-0314 a flattened specimen, showing the left arm freely coiled and the right one extended anterolaterally; (F) ELI L-0127, showing the paired crossed lophophoral arms preserved in the sediment infilling between valves; (G) Sketch of (A); (H) Sketch of (B); (I) Sketch of (C); (J) Sketch of (E).
Abbreviations: B, lophophoral arm; Fp, food particle?; Vr, visceral region; Pi, pseudointerarea; P, pedicle; S, mantle setae; Bc, lophophoral canal; T, tentacle; L, lophophore. Scale bars 2 mm.