◄ Carnets Geol. 4 (L05) ►
![]()
Hellenic Ministry of Culture, Ephorate of PalaeoanthropologySpeleology
Ardittou 34B, 11636 Athens (Greece)
Manuscript online since November 25, 2004
![]()
This paper reports the discovery of an incomplete proboscidean mandible near the village of Maroniá in eastern Crete. The fragment described here includes the first molar (m1) of a deinothere, that because of its large size is identified as Deinotherium giganteum. The specimen was found in shallow-water marine sediments. The presence of Deinotherium on the island, together with other Miocene vertebrate faunas, suggests that during that epoch Crete was connected to the mainland by a wide land bridge.
Greece;
Crete;
Late Miocene;
Proboscidea;
Deinotherium.
A. (2004).- On a Deinotherium (Proboscidea) finding in the Neogene of Crete.- Carnets Geol., Madrid, vol. 4, no. L05, 7 p. DOI: 10.4267/2042/311
Découverte de Deinotherium (Proboscidea) dans le Néogène de Crète.- Près du village Maroniá, en Crète orientale, une formation marine miocène a livré une mandibule incomplète de Proboscidien. Ce fragment comprend une première molaire (m1) de grande dimension attribuable à l'espèce Deinotherium giganteum. La présence d'un Déinothère dans la faune des mammifères miocènes de l'île de Crète montre qu'à cette époque il existait une liaison relativement large et durable avec le continent.
Grèce ;
Crète ;
Miocène supérieur ;
Proboscidea ;
Deinotherium.
Crete is well known for its faunas of large mammals in Quaternary times, mainly elephants, deer and hippos. These faunas are endemic and ecologically unbalanced due to the fact that Crete was already an island. To reach it from the mainland demanded the ability, possessed only by these larger mammals, to swim well ( & , 1991; , 1994; , 1996; et al., 1996). However, Miocene larger mammal faunas reported from the island (recently revised by , 1996) have a clearly continental aspect, thus indicating that during the Miocene epoch Crete was not an island. These faunas comprise a gomphotheriid, cf. Prohyrax hendeyi, Microstonyx cf. major, cf. Propotamochoerus palaeochoerus, Taucanamo ? / Yunnanochoerus ? sp., Dorcatherium naui, cf. Dorcabune anthracotherioides, cf. Pliocervus pentelici, Bovidae indet., Hipparion sp. and Carnivora indet. ( et al., 1970; et al., 1971; & , 1972; & , 1974; , 1976; & , 1978; , 1996). Their age (often determined with the aid of micromammals) spans Middle to Late Miocene times (MN 6 to MN 11 , 1996).
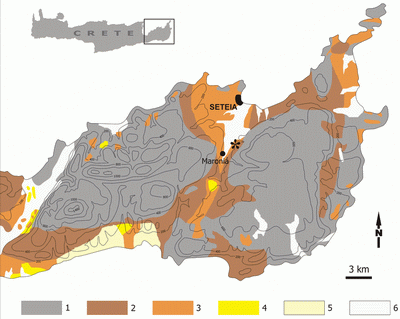
The new report of a large mammal in the Neogene of Crete is from a site situated south of the town of Seteía (Prefecture of Lasíthi, Eastern Crete) in the vicinity of the village of Maroniá, WNW of the small settlement of Sfakiá
(Fig. 1
![]() ). The only fossil of a mammal found at the site, a Deinotherium dentary fragment, was collected about seven years ago by Mr. Ioannis , a resident of Seteía. This is the first deinotheriid found in Crete. In May 2004 the specimen was delivered to the Archaeological Museum of Seteía, where it will be on exhibit soon.
). The only fossil of a mammal found at the site, a Deinotherium dentary fragment, was collected about seven years ago by Mr. Ioannis , a resident of Seteía. This is the first deinotheriid found in Crete. In May 2004 the specimen was delivered to the Archaeological Museum of Seteía, where it will be on exhibit soon.
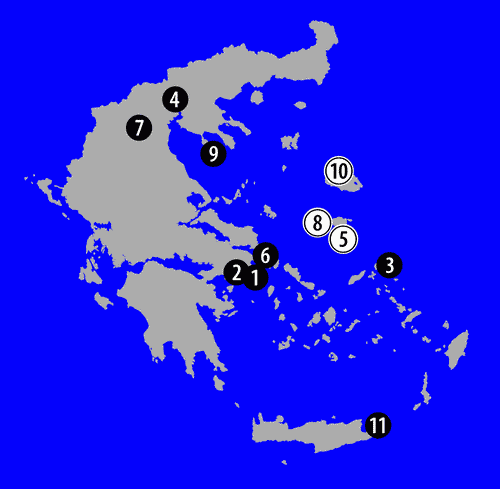
Fossil deinotheres have been reported from other localities in Greece
(Fig. 2
![]() ). Smaller and geologically older specimens, classified in Prodeinotherium , 1930 (,
1973; , 1999; ,
2002a), have been found on the islands of Chíos, Psará and Lésbos in the eastern Aegean Sea (,
1940; & ,
1975; et al., 2003). Larger and younger specimens, classified in
Deinotherium ,
1829, are more common and more widely disseminated. They are represented in some of the classical
"Pikermian"-type faunas (Pikérmi, Sámos, Halmyropótamos, Pýrgos Vassilíssis, Vathýlakkos ,
1862; , 1970; et al.,
1982), as well as in other less well-known localities (a not tracked down locality in Central Macedonia and Haghía Paraskeví ,
1970; & ,
1994). Another deinothere site was discovered recently in Eastern Crete ( et al.,
in press). This locality too is near Seteía (Gela section), less than 10 km north of the Maroniá site, where the specimen described here was found. The Gela site yielded several dental and postcranial specimens, all from one large individual of Deinotherium giganteum.
). Smaller and geologically older specimens, classified in Prodeinotherium , 1930 (,
1973; , 1999; ,
2002a), have been found on the islands of Chíos, Psará and Lésbos in the eastern Aegean Sea (,
1940; & ,
1975; et al., 2003). Larger and younger specimens, classified in
Deinotherium ,
1829, are more common and more widely disseminated. They are represented in some of the classical
"Pikermian"-type faunas (Pikérmi, Sámos, Halmyropótamos, Pýrgos Vassilíssis, Vathýlakkos ,
1862; , 1970; et al.,
1982), as well as in other less well-known localities (a not tracked down locality in Central Macedonia and Haghía Paraskeví ,
1970; & ,
1994). Another deinothere site was discovered recently in Eastern Crete ( et al.,
in press). This locality too is near Seteía (Gela section), less than 10 km north of the Maroniá site, where the specimen described here was found. The Gela site yielded several dental and postcranial specimens, all from one large individual of Deinotherium giganteum.
|
Figure 1: Geological map of the area of Seteía, based on et al.
(1977). An asterisk indicates the Maroniá locality. |
|
Figure 2: Map of Greece, indicating the Deinotheriidae bearing localities: white circles indicate the presence of Prodeinotherium bavaricum; black circles indicate the presence of Deinotherium giganteum. |
Geologically, eastern Crete consists of a predominantly Mesozoic Alpine succession that is expressed in a fairly high and steep relief, and of Neogene deposits that form hilly lowlands. The Maroniá area is in a graben of neotectonic origin, filled by Neogene, shallow-water marine sediments that overlie a pre-Neogene series. The Neogene deposits of the Seteía District consist primarily of marls, clays, marly limestones and sandstones. They are rich in invertebrate fossils (Foraminifera, Bivalvia, Gastropoda, Echinodermata, etc.) that indicate a Middle Miocene to Pliocene age (, 1966; , 1973; et al., 1993). Cranial and postcranial remains of a Miocene sirenian, Metaxytherium cuvieri, were found at the village of Achládia, WNW of Maroniá ( & , 2004). (1973) recognises seven Neogene lithostratigraphic formations in the Seteía region, two of which are present around Maroniá: an underlying Skopí Formation, mainly conglomerates, sands, silts and clays with Crassostrea, Ostrea and plant remains, and the Faneroméni Formation, consisting mainly of limestones and marls with foraminiferans, sponges, algae and molluscs. The fossil content of these Neogene deposits indicates a low energy littoral shallow-water milieu (, 1973; & , 2004).
The Deinotherium specimen was found in the Faneroméni Formation (sensu , 1973), in a pebble bed intercalated in white marls, which include several Ostrea banks. Based on biostratigraphic data (1973, p. 569) reports a Late Miocene age for the Faneroméni Formation.
Order: Proboscidea , 1811
Suborder: Deinotherioidea , 1921
Family: Deinotheriidae , 1845
Genus: Deinotherium , 1829
Deinotherium giganteum , 1829
Material: right dentary fragment with the first molar (m1), MAP-1
(Fig. 3
![]() ).
).
|
Figure 3: Deinotherium giganteum, right mandibular part with m1; a, occlusal view; b, labial view; c, lingual view. Scale: 50 mm. |
Locality: Maroniá, Prefecture of Lasíthi, E Crete.
Stratigraphic level: Upper Miocene, Faneroméni Formation.
Description: The dentary fragment MAP-1 is too incomplete to provide clues about the morphology of the mandible. The first molar is preserved in good condition, except for a part of the enamel on the lingual side, which was detached post-mortem. As is the case in all deinotheriid first molars, the molar of MAP-1 is trilophodont (it consists of three transverse lophids) and is elongated. The lophids are curved, the concave side facing anteriorly. The tooth is almost pristine, presenting only slight attrition (wear facets are hardly visible at the posterior side of the lophids). The cutting edges of the lophids retain traces of crenulations, which are very clear on the unworn distal lophid. The enamel is thick, ranging in thickness from 1 mm at the sides of the crown, to 4 mm on the occlusal surface. The mesial (first) lophid (protolophid) is the most prominent, while the distal one is the least. Lophids are separated by deep and wide valleys, which are open at both sides and deeper labially. The anterior valley is a little wider than the posterior one. There is a mesial crenulated cingulid, which is well developed, especially on the labial half of the mesial side. The talonid, which is also crenulated, is weak. The dimensions of MAP-1 are given in Table 1.
Table 1: Dental dimensions in mm of the specimen MAP-1. L: maximal length; W1,2,3: maximal widths of the first, second and third lophid respectively; H: maximal crown height (at the metaconid).
| L | W1 | W2 | W3 | H | |
| MAP-1 | 105.3 | 70.5 | 70.5 | 64.9 | 51.5 |
The family Deinotheriidae is an extinct proboscidean group that originated in Africa, where it was already present in the Late Oligocene and survived until the Early Pleistocene. Deinotheres soon dispersed in Europe and southwest Asia during the Early Miocene, where they survived until the Pliocene ( & , 1962; , 2002a). They are characterised by typical proboscidean cranial and postcranial morphology, including large size, short body on long columnar limbs with short distal elements, pneumatised cranial bones, retracted nasals, the inferred presence of a proboscis, and the presence of elongate incisors that form tusks. They retain some ancestral characters - a low skull and bunodont dentition with deciduous and permanent tooth generations that erupt vertically. However, deinotheres show a unique derived character: the lack of upper tusks and the presence of downturned, vertically developed tusks in the mandible ( & , 1962; , 1988; et al., 1996; , 2002a).
Their distinctive morphology led to the classification of the family in a separate suborder, Deinotherioidea. However, several opinions have been expressed about taxonomy at the genus and species level (, 1936; , 1957; & , 1962; , 1973, , 2002a; among others). Deinotheres were originally referred to a single genus, Deinotherium , 1829. Nevertheless, important anatomical (cranial and postcranial) differences between the earlier, smaller forms and the later larger forms supported the grouping of the former ones in another genus, Prodeinotherium , 1930. The division of the family into two genera is generally accepted and is adopted in recent taxonomical works (, 1973; et al., 1996; , 1999; , 2002a). The distinction between Prodeinotherium and Deinotherium is based mainly on size, the morphology of the p3, snout and cranial roof shape, inclination of the occiput, relative position of the occipital condyles, development of the scapular spine and disposition of the carpals and tarsals. Deinotherium has also a more cursorially adapted skeleton than Prodeinotherium. The dental morphology does not offer any diagnostic character (except for p3), and the distinction of genus from dental material is based primarily on size. More recently, a third deinothere genus, Chilgatherium, was erected to accommodate dentally primitive, very diminutive specimens from the Late Oligocene site of Chilga, Ethiopia ( et al., 2004). The Deinotheriidae present a constant trend towards larger size, which culminated in the gigantic size of late representatives of Deinotherium.
The history of specific classification in the Deinotheriidae is more complicated: apart from the recently described Chilgatherium harrisi et al., 2004, 31 species have been erected (for the most part during the 19th century), based primarily on morphological and metrical diversity in the dental elements of specimens from African, European and Asian localities (, 2002a). However, more recent studies ( & , 1962; , 1973; , 2002a) have shown that most of these differences (often in minor dental features) are due to intra- and not interspecific variation. Today, only six of these species are considered valid (, 1973, 1978; , 2002a), of which two are confined to Europe: Prodeinotherium bavaricum , 1831 of Early to Middle Miocene age, and Deinotherium giganteum of Middle Miocene to Pliocene age. Some authors (e.g. , 1999) also accept another, not as well known Late Miocene Pliocene species, Deinotherium gigantissimum , 1892, which is of gigantic size and palaeobiogeographically confined to Europe. Most authors, however, consider it to be a large-sized variety of Deinotherium giganteum ( & , 1962; , 1978; , 2002a).
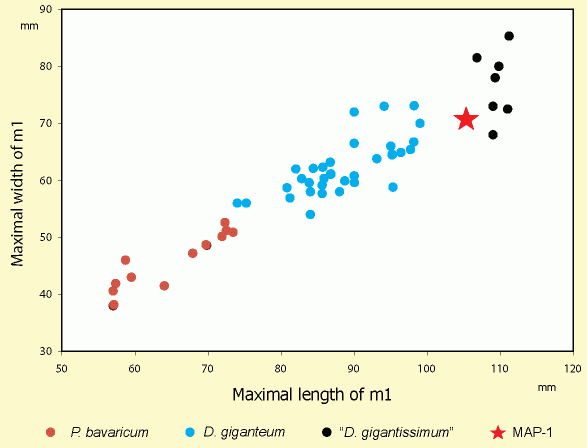
The specimen reported here is characterised by its large size. Compared to published m1 measurements, MAP-1 appears to be much larger than any known specimen of Prodeinotherium bavaricum, and is even larger than certain Deinotherium giganteum from German, French, Hungarian and Austrian localities
(Fig. 4
![]() ). However, there are two specimens, referred by
(1832) and (1936, p. 109) to Deinotherium giganteum, which are comparable in size to MAP-1. The former (plotted in
Fig. 4
). However, there are two specimens, referred by
(1832) and (1936, p. 109) to Deinotherium giganteum, which are comparable in size to MAP-1. The former (plotted in
Fig. 4
![]() ), slightly longer and narrower than MAP-1, has dimensions close to those of Deinotherium gigantissimum (a species erected many years after 's publication); the latter has a length of 104 mm (not plotted in
Fig. 4
), slightly longer and narrower than MAP-1, has dimensions close to those of Deinotherium gigantissimum (a species erected many years after 's publication); the latter has a length of 104 mm (not plotted in
Fig. 4
![]() , as does not give any width measurement). Actually, the dimensions of MAP-1 seem to be slightly smaller, but very near those of first lower molars attributed to Deinotherium gigantissimum. A metrical comparison with the other Cretan deinothere find is not possible, as this material does not include a lower first molar ( et al.,
in press).
, as does not give any width measurement). Actually, the dimensions of MAP-1 seem to be slightly smaller, but very near those of first lower molars attributed to Deinotherium gigantissimum. A metrical comparison with the other Cretan deinothere find is not possible, as this material does not include a lower first molar ( et al.,
in press).
The morphology of MAP-1 does not differ from that typical of deinotheriids, for the molars of all members of the family show great morphological homogeneity.
If we accept Deinotherium gigantissimum as a valid species, then the size of MAP-1 places it among the largest D. giganteum and the smallest D. gigantissimum. So a specific attribution is difficult, given that there is no discontinuity in the range of sizes between these species ( & , 1962). If we accept the synonymy of D. gigantissimum and D. giganteum (the most common contemporary trend), MAP-1 is a large specimen of Deinotherium giganteum, but is considerably smaller than the largest reported measurements for the species (length: 112 mm, width: 90 mm, according to & , 1962). As the synonymy of the two species has been generally accepted, the specimen MAP-1 is referred to the species Deinotherium giganteum.
|
Figure 4: First lower molar dimensions diagram of MAP-1, compared to European Deinotheriidae samples. Data according to (1832), (1957), & (1962), (1988), (1993), (2002b) and et al. (2003). |
In Eurasia, Deinotherium giganteum has a broad stratigraphical distribution: from the Middle Miocene (MN 6) to the Pliocene, although the upper limit is not known precisely. (1990) places the last occurrence of the species in Central and Western Europe in MN 13, while (in press) reports the presence of Deinotherium in Russia until the end of the Ruscinian (MN 15). Given the deinotheriid trend towards larger size during the Miocene (, 1957; , 1978; , 1999; , 2002a) and the large size of the new deinotheriid specimen, a Late Miocene or even later age can be assumed. Published marine biostratigraphical data for the Maroniá area place its Neogene sediments in the Upper Miocene (, 1973), thus limiting the age of the find. Moreover, a Pliocene age for MAP-1 is not plausible, as an extensive marine transgression during that epoch broke Crete up into small islands ( et al., 1996). This resulted in the submergence of large tracts of land, leaving areas insufficient to sustain a population of large mammals. The continental Middle to Late Miocene mammalian faunas found in Crete, together with the new Deinotherium find, show that the island was connected to the mainland by a land bridge at that time. The discovery of Deinotherium at another site in the Seteía region ( et al., in press) suggests that deinotheres were probably not visitors by happenstance but were established residents on the island.
The morphology of the deinotheriid cheek teeth, which are lophodont, brachyodont and simple in shape, indicate that the deinotheres were primarily folivores, consumers of soft, non-abrasive vegetation. This feeding adaptation commonly implies a forested environment. Another feature that points to a closed habitat is the increased mobility of the head as compared to that of the Elephantoids (, 1973, 1978). Conversely, the skeleton of Deinotherium, in contrast to that of Prodeinotherium, exhibits cursorial adaptations, such as the relative elongation of the distal limb elements. This feature, together with the very large size of the animal and that deinotheres may have had to travel widely to find adequate forage, suggests a landscape more open than that of a forest. Thus, the presence of the Deinotherium giganteum tooth in the Maroniá area makes plausible the inference that an open forest or woodland landscape prevailed in Eastern Crete during the Late Miocene.
The author thanks the reviewers of this paper, Sevket , Emmanuel and William J. , whose suggestions and comments improved the manuscript, as well as Nestor , who emended the English text.
L., H., S.E. & N.K. (1970).- Der erste Mastodon-Fund der Insel Kreta.- Annales Géologiques des Pays Helléniques, Athènes, v. XXI, p. 167-177.
F.-M. & F. (1962).- Les Déinothéridés d'Europe.- Annales de Paléontologie, Paris, v. XLVIII, p. 11-56.
H. & N.K. (1975).- Der erste Säugetierfund aus dem Neogen der griechischen Insel Psara (Ostägäis).- Annales Géologiques des Pays Helléniques, Athènes, v. XXVI, p. 109-117.
M. & L. (1974).- Découverte de Dorcatherium puyhauberti et (Mammalia) dans les faciès continentaux de base de la molasse tertiaire de Crète (Grèce): Conséquences stratigraphiques et tectoniques.- Compte Rendu Sommaire des Séances de la Société Géologique de France, Paris, v. 1974, n° 1, p. 11-12.
N., C.W., J.E., J., W., E. & A. (1977).- General Geological Map of Greece: Crete Island, 1:200,000.- Institute of Geological and Mining Research, Athens.
H. & J.E. (1972).- Late Miocene rodents from the Pandanassa Formation (Prov. Rethymnon), Crete, Greece.- Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, Amsterdam, v. 75, n° 1, p. 54-60.
H., P.Y. & W.J. (1971).- Mammalia and Foraminifera from the Neogene of Kastellios Hill (Crete). A correlation of continental and marine biozones I.- Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, Amsterdam, v. 74, n° 5, p. 1-22.
M.D. (1994).- Late Cenozoic paleogeography and faunal succession of mammals in Crete.- Bulletin de la Société Spéléologique de Grèce, Athènes, v. XXI, p. 301-317.
M.D., N.K., L.E.M. & P.Y. (1982).- The evolution of the elephants.- Editions of the Laboratory of Geology and Palaeontology, University of Athens, Athens, 85 p.
M. (1993).- Deinotheres (Proboscidea, Mammalia) of Hungary.- Annales Historico-Naturales Musei Nationalis Hungarici, Budapest, v. 85, p. 3-17.
A. (1862).- Animaux fossiles et géologie de l'Attique.- E. Savy, Paris, 476 p.
U.B. (1999).- Order Proboscidea. In: G.E. & K. (eds.), The Miocene land mammals of Europe.- Dr. Friedrich Pfeil, München, p. 157-168.
F.M. (1973).- The Neogene and Quaternary deposits in the Sitia District of Eastern Crete.- Annales Géologiques des Pays Helléniques, Athènes, v. XXIV, p. 527-572.
I.E. (1957).- Die Prinzipien der Artbestimmung bei Dinotherium.- Palaeontographica, Stuttgart, v. 108, p. 131-185.
J.M. (1973).- Prodeinotherium from Gebel Zelten, Libya.- Bulletin of the British Museum (Natural History) , London, v. 23, n° 5, p. 285-348.
J.M. (1978).- Deinotherioidea and Barytherioidea. In: V.J. & H.B.S. (eds.), Evolution of African Mammals.- Harvard University Press, Cambridge, p. 315-332.
K. (2002a).- Systematics and taxonomy of the European Deinotheriidae (Proboscidea, Mammalia).- Annalen des Naturhistorischen Museums in Wien, Wien, v. 103A, p. 237-250.
K. (2002b).- Deinotheriidae (Proboscidea, Mammalia) dental remains from the Miocene of Lower Austria and Burgenland.- Annalen des Naturhistorischen Museums in Wien, Wien, v. 103A, p. 251-285.
J.-J. (1832).- Description d'ossements fossiles de mammifères inconnus jusqu'à présent, qui se trouvent au Muséum grand-ducal de Darmstadt.- Heyer, Darmstadt, 122 p.
G.D., N. & O. (2003).- Prodeinotherium bavaricum (Proboscidea, Mammalia) from Lesvos island, Greece; the appearance of deinotheres in the eastern Mediterranean.- Géobios, Lyon, v. 36, n° 3, p. 305-315.
S.E. (1976).- Ein erster Fund von Pliohyrax aus dem Vallesium von Kreta / Griechenland.- Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, Stuttgart, v. 1976, n° 3, p. 157-162.
E.M. (1996).- A gazetteer of Cretan paleontological localities. In: D.S. (ed.), Pleistocene and Holocene fauna of Crete and its first settlers.- Prehistory Press, Madison, p. 1-32.
J.J.M. & J.E. (1978).- A Microstonyx tooth from Eastern Crete; paleogeographical implications of Cretan Tortonian mammal associations.- Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, Amsterdam, v. 81, n° 4, p. 416-424.
A. & E. (2004).- Presence of Metaxytherium cuvieri in the Miocene sediments of Seteía, Crete.- 10th Congress of the Geological Society of Greece, Thessaloniki, 2004, Abstracts, p. 152-153.
A., M.-R., E., A. & S. (1993).- Données nouvelles sur la stratigraphie du Néogène dans le graben néotectonique de Sitia (Crète orientale, Grèce).- Bulletin of the Geological Society of Greece, Athens, v. XXIX, p. 17-31.
P. (1990).- Updating of MN zones. In: E.H., V. & P. (eds.), European Neogene mammal chronology.- Plenum Press, New York, p. 73-90.
J.K. (1970).- Die Pikermifauna von Halmyropotamos (Euböa, Griechenland) 2. Teil: Osteologie.- Annales Géologiques des Pays Helléniques, v. XXI, p. 217-306.
H.F. (1936).- Proboscidea a monograph of the discovery, evolution, migration and extinction of the mastodonts and elephants of the world, Volume I: Moeritherioidea, Deinotherioidea, Mastodontoidea.- American Museum Press, New York, 802 p.
I. (1940).- Eine obermiocäne Fauna von Chios.- Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Stuttgart, v. 83, p. 304-365.
N., P. & C. (in press).- A Deinotherium giganteum , 1829 (Proboscidea, Deinotheriidae) from the middle Miocene of Siteia (East Crete, Greece).- Journal of Vertebrate Paleontology, New York.
W.J., J. & D.T. (2004).- New large-bodied mammals from the late Oligocene site of Chilga, Ethiopia.- Acta Palaeontologica Polonica, Warszawa, v. 49, n° 3, p. 365-392.
J., R.M., N., R.J.G. & J.M. (1996).- The earliest proboscideans: general plan, taxonomy, and palaeoecology. In: J. & P. (eds.), The Proboscidea: evolution and paleoecology of elephants and their relatives.- Oxford University Press, New York, p. 57-75.
P.Y., M.D. & J. (1996).- The palaeogeography and faunal evolution of the land mammals of Crete. In: D.S. (ed.), Pleistocene and Holocene fauna of Crete and its first settlers.- Prehistory Press, Madison, p. 61-67.
N.K. (1966).- Der Neogen von Ostkreta.- Annales Géologiques des Pays Helléniques, Athènes, v. XVI, p. 249-314.
N.K. (1970).- Ein Dinotherium-Fund in Zentralmakedonien (Griechenland).- Annales Géologiques des Pays Helléniques, Athènes, v. XXI, p. 334-341.
G. & M.D. (1991).- Fossil Vertebrates in Cretan caves.- 6th International Cretological Congress, Chania, 1991, Proceedings, v. B, p. 377-392.
H. (1988).- Contributions à l'étude du gisement Miocène supérieur de Montredon (Hérault) Les grands Mammifères: 7 - Les Proboscidiens Deinotheriidae.- Palæovertebrata, Montpellier, Mémoire extraordinaire 1988, p. 135-175.
E. & J.K. (1994).- Deinotherium giganteum (Proboscidea) from Kassandra peninsula (Chalkidiki, Macedonia, Greece).- Géobios, Lyon, v. 27, n° 5, p. 633-640.
J. (1996).- Pre-Pleistocene land mammals from Crete. In: D.S. (ed.), Pleistocene and Holocene fauna of Crete and its first settlers.- Prehistory Press, Madison, p. 69-79.
I. (in press).- On Pliocene faunas with Proboscideans in the territory of the former Soviet Union.- Quaternary International, Oxford, v. 126-128 (2005), p. 93-105.