![]()
![]()
E. & E. (2005).- Main changes in microfossil communities throughout the Upper Proterozoic of Russia. In: P. & E. (eds.), Pre-Cambrian to Palaeozoic Palaeopalynology and Palaeobotany.- Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2005/02, Abstract 04 (CG2005_M02/04)
![]() Microfossils; Upper Proterozoic; East-European and Siberian platforms.
Microfossils; Upper Proterozoic; East-European and Siberian platforms.
![]() Microfossiles ; Protérozoïque supérieur ;
plates-formes est-européenne et sibérienne.
Microfossiles ; Protérozoïque supérieur ;
plates-formes est-européenne et sibérienne.
![]()
![]() More than 50 years of study have resulted in the description of hundreds of taxa of Precambrian microfossils from the different regions of Russia. These forms are usually preserved either as silicified or organic-walled remains and generally are morphologically simple and stratigraphically long-ranging. The systematics of Precambrian microorganisms still needs a serious revision, although efforts by specialists from all over the world to solve these problems have helped to clarify the biological assignment of some taxa.
More than 50 years of study have resulted in the description of hundreds of taxa of Precambrian microfossils from the different regions of Russia. These forms are usually preserved either as silicified or organic-walled remains and generally are morphologically simple and stratigraphically long-ranging. The systematics of Precambrian microorganisms still needs a serious revision, although efforts by specialists from all over the world to solve these problems have helped to clarify the biological assignment of some taxa.
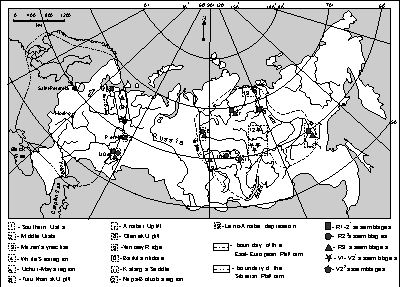
![]() Distinctive, exceptionally diverse and well-preserved microbiotas were found at several locations on the East-European and Siberian Platforms. They occur at discrete stratigraphical levels and are geographically sporadic. However, a comprehensive analysis based on the published data and original material (Fig. 1
Distinctive, exceptionally diverse and well-preserved microbiotas were found at several locations on the East-European and Siberian Platforms. They occur at discrete stratigraphical levels and are geographically sporadic. However, a comprehensive analysis based on the published data and original material (Fig. 1 ![]() ) permitted the drawing of an evolutionary trend in the main taxonomical changes that occurred in the organic world throughout the Upper Proterozoic of Russia.
) permitted the drawing of an evolutionary trend in the main taxonomical changes that occurred in the organic world throughout the Upper Proterozoic of Russia.
![]() For the present study the only records we used were from strata controlled either by isotopic data or other evidence of age. Both silicified and organic-walled microfossils were analyzed taxonomically. Only species with real biological meaning or biostratigraphical significance were used to define noteworthy stages in the development of Upper Proterozoic microorganisms. Five main stratigraphical levels, characterized by the FAD of distinctive species or by the occurrence of new morphological forms have been distinguished (Fig. 2
For the present study the only records we used were from strata controlled either by isotopic data or other evidence of age. Both silicified and organic-walled microfossils were analyzed taxonomically. Only species with real biological meaning or biostratigraphical significance were used to define noteworthy stages in the development of Upper Proterozoic microorganisms. Five main stratigraphical levels, characterized by the FAD of distinctive species or by the occurrence of new morphological forms have been distinguished (Fig. 2 ![]() ).
).
![]() A long interval embracing the Early Riphean and the bulk of the Middle Riphean does not show much diversity in the microfossil populations.
A long interval embracing the Early Riphean and the bulk of the Middle Riphean does not show much diversity in the microfossil populations.
![]() The most representative assemblages of this age
(1,650-1,250 Ma), all of them on the Siberian platform, are from the Billyakh Group of the Anabar Uplift ( et alii,
2001; et alii, 1995 and others), the Kyutingde and Debengda
formations of the Olenek Uplift, and the Omackhta Fm of the Uchur-Maya region. But more diverse Early Riphean microbiotas were described from the Burzyan Group of the Bashkirian meganticlinorium, Southern Urals, adjacent to the East-European Platform.
The most representative assemblages of this age
(1,650-1,250 Ma), all of them on the Siberian platform, are from the Billyakh Group of the Anabar Uplift ( et alii,
2001; et alii, 1995 and others), the Kyutingde and Debengda
formations of the Olenek Uplift, and the Omackhta Fm of the Uchur-Maya region. But more diverse Early Riphean microbiotas were described from the Burzyan Group of the Bashkirian meganticlinorium, Southern Urals, adjacent to the East-European Platform.
![]() These assemblages are dominated by cyanobacteria. The most common are the ellipsoidal akinetes Archaeollipsoides (Nostocacea or Stigonematacea), chroococcacean cyanobacterium Eoentophysalis (Entophysalidacea) and empty sheaths Siphonophycus (Oscillatoriacea or Nostocacea). In addition, the less abundant taxa of various cyanophyta such as Circumvaginalis, Oscillatoriopsis, Filiconstrictosus, Partitiofilum, Palaeolyngbia, Eosynechococcus, Sphaerophycus, Myxococcoides and some others occur in these assemblages. Although the representatives of Archaeollipsoides, Eoentophysalis and Siphonophycus were reported from the older sediments (approximately 1,900-2,000 Ma) of the Belcher Supergroup and the Epworth Group of Canada, they became the dominant components of microbiotas only in the Early-Middle Riphean. The list of common taxa present in these assemblages includes the following: Leiosphaeridia, Eomarginata, Simia, Pterospermopsimorpha, Satka, Konderia, Germinosphaera, Palaeopleurocapsa and others.
These assemblages are dominated by cyanobacteria. The most common are the ellipsoidal akinetes Archaeollipsoides (Nostocacea or Stigonematacea), chroococcacean cyanobacterium Eoentophysalis (Entophysalidacea) and empty sheaths Siphonophycus (Oscillatoriacea or Nostocacea). In addition, the less abundant taxa of various cyanophyta such as Circumvaginalis, Oscillatoriopsis, Filiconstrictosus, Partitiofilum, Palaeolyngbia, Eosynechococcus, Sphaerophycus, Myxococcoides and some others occur in these assemblages. Although the representatives of Archaeollipsoides, Eoentophysalis and Siphonophycus were reported from the older sediments (approximately 1,900-2,000 Ma) of the Belcher Supergroup and the Epworth Group of Canada, they became the dominant components of microbiotas only in the Early-Middle Riphean. The list of common taxa present in these assemblages includes the following: Leiosphaeridia, Eomarginata, Simia, Pterospermopsimorpha, Satka, Konderia, Germinosphaera, Palaeopleurocapsa and others.
![]() The most diverse organic-walled assemblage is from the Ust'-Il'ya and Kotuikan
formations of the above-mentioned Billyakh Group (1,490-1,250 Ma). Here, in addition to cyanobacteria and the remains of what are probably eukaryotic microorganisms, there are large spherical forms of Chuaria circularis , 1899 (400-800 µm in diameter), large filamentous fragments of Elatera (Eosolena) and spheroids bearing one long process (considered as reproductive structure of fungi or
algae) of Caudosphaera ( et alii,
2001).
The most diverse organic-walled assemblage is from the Ust'-Il'ya and Kotuikan
formations of the above-mentioned Billyakh Group (1,490-1,250 Ma). Here, in addition to cyanobacteria and the remains of what are probably eukaryotic microorganisms, there are large spherical forms of Chuaria circularis , 1899 (400-800 µm in diameter), large filamentous fragments of Elatera (Eosolena) and spheroids bearing one long process (considered as reproductive structure of fungi or
algae) of Caudosphaera ( et alii,
2001).
![]() The uppermost Middle Riphean
(1,250-1,000 Ma) is marked by
the appearance of several new forms. The stalk-forming cyanobacterium Polybessurus bipatitus , 1975, ex et alii, 1987 occurs in silicified microbiotas from the Southern Urals and the Turukhansk Uplift, Siberia (,
1994 and others). Among organic-walled microfossils the first appearances of the spherical striate eukaryote form Valeria lophostriata , 1982 and of the spiral prokaryotes Obruchevella and Glomovertella are documented from the Katanga saddle, the Baikit syneclise and the Patom Uplift of the Southern Siberia (,
2001). The acanthomorphic acritarch Tappania, assigned by
(2005) to fungi, recently was reported from the uppermost Middle Riphean sediments of
southwestern Siberia (,
2001). Tappania was first described in the early Mesoproterozoic Ruyang Group of China (,
1998) and the Roper Group of Ausralia ( et alii,
2001) but differs in various aspects from the younger (~800 Ma) Tappania described by
(, pers. comm.). The branching multicellular talloms of Ulophyton, provisionally attributed by (pers. comm.) to red algae, occur in the coeval strata of the Totta Fm in the Uchur-Maya region.
The uppermost Middle Riphean
(1,250-1,000 Ma) is marked by
the appearance of several new forms. The stalk-forming cyanobacterium Polybessurus bipatitus , 1975, ex et alii, 1987 occurs in silicified microbiotas from the Southern Urals and the Turukhansk Uplift, Siberia (,
1994 and others). Among organic-walled microfossils the first appearances of the spherical striate eukaryote form Valeria lophostriata , 1982 and of the spiral prokaryotes Obruchevella and Glomovertella are documented from the Katanga saddle, the Baikit syneclise and the Patom Uplift of the Southern Siberia (,
2001). The acanthomorphic acritarch Tappania, assigned by
(2005) to fungi, recently was reported from the uppermost Middle Riphean sediments of
southwestern Siberia (,
2001). Tappania was first described in the early Mesoproterozoic Ruyang Group of China (,
1998) and the Roper Group of Ausralia ( et alii,
2001) but differs in various aspects from the younger (~800 Ma) Tappania described by
(, pers. comm.). The branching multicellular talloms of Ulophyton, provisionally attributed by (pers. comm.) to red algae, occur in the coeval strata of the Totta Fm in the Uchur-Maya region.
![]() The base of the Upper Riphean (approximately 1,000 Ma) is one of the most important levels in the evolution of Upper Proterozoic microbiotas. This is expressed
by incoming morphological innovations, including a remarkable increase in the diversity and size of eukaryotes.
The base of the Upper Riphean (approximately 1,000 Ma) is one of the most important levels in the evolution of Upper Proterozoic microbiotas. This is expressed
by incoming morphological innovations, including a remarkable increase in the diversity and size of eukaryotes.
![]() The most distinctive and diverse microbiotas are known from the Uchur-Maya Region (the Lakhanda and Ui
groups, ,
1981, 1990 and others), the Turukhansk Uplift (the Derevninya, Shorikha and Miroedikha
formations, et alii,
1998; , 2001 and others) and the Yenisey Ridge (the Seryi Kluch and Dashka
formations, ,
2000) of Siberia, from the Southern Urals (the Zil'merdak, Nugush, and Inzer
formations, et alii,
2003 and others) and the Mesen' syneclise (Vashkin Fm.) of the East-European Platform ( et alii,
2004).
The most distinctive and diverse microbiotas are known from the Uchur-Maya Region (the Lakhanda and Ui
groups, ,
1981, 1990 and others), the Turukhansk Uplift (the Derevninya, Shorikha and Miroedikha
formations, et alii,
1998; , 2001 and others) and the Yenisey Ridge (the Seryi Kluch and Dashka
formations, ,
2000) of Siberia, from the Southern Urals (the Zil'merdak, Nugush, and Inzer
formations, et alii,
2003 and others) and the Mesen' syneclise (Vashkin Fm.) of the East-European Platform ( et alii,
2004).
![]() More than 50 species of microfssils are described from the Upper Riphean assemblages of Russia. The spherical acanthomorphic acritarch Trachyhystrichosphaera is of a particular interest, because it appears and becomes common in the Upper Riphean assemblages of every region. In the Lakhanda biota from the Neryuen Fm of the Lakhanda Group (the Uchur-Maya region, eastern Siberia) diverse and abundant microorganisms belonging to fungi, algae and several problematic groups are known, including possible animals ( & ,
2002). Some taxa (Eosacchoromyces, Mucorites and Mycosphaeroides, ,
1990) are classified as fungi while some others have definite algal affinities. The latter include green algae, such as Archaeoclada (attributed to
Siphonocladales), Lomentunella (probably belonging to Ulotrichales), Palaeovaucheria (assigned to
Vaucheriales of a xanthophyte algae), and Ulophyton and Bangiomorpha interpreted as red algae (,
2000). In the silicified Siberian microbiotas there are the coccoidal microfossils Glenobotrydion, Palaeastrum and Myxococcoides attributed to green algae of the order
Chlorococcales (,
2000 and others).
More than 50 species of microfssils are described from the Upper Riphean assemblages of Russia. The spherical acanthomorphic acritarch Trachyhystrichosphaera is of a particular interest, because it appears and becomes common in the Upper Riphean assemblages of every region. In the Lakhanda biota from the Neryuen Fm of the Lakhanda Group (the Uchur-Maya region, eastern Siberia) diverse and abundant microorganisms belonging to fungi, algae and several problematic groups are known, including possible animals ( & ,
2002). Some taxa (Eosacchoromyces, Mucorites and Mycosphaeroides, ,
1990) are classified as fungi while some others have definite algal affinities. The latter include green algae, such as Archaeoclada (attributed to
Siphonocladales), Lomentunella (probably belonging to Ulotrichales), Palaeovaucheria (assigned to
Vaucheriales of a xanthophyte algae), and Ulophyton and Bangiomorpha interpreted as red algae (,
2000). In the silicified Siberian microbiotas there are the coccoidal microfossils Glenobotrydion, Palaeastrum and Myxococcoides attributed to green algae of the order
Chlorococcales (,
2000 and others).
![]() The microfossil assemblages of Early? - Late Vendian age (0.6-0.55 Ma) are recorded from the Kursov and Parshin
formations of the Nepa-Botuoba region and from the Turkut Fm of the Lena-Anabar depression of Siberia ( & ,
1989; , 2004). There is
but one report of what is probably a contemporaneous microbiota from the Vychegoda Fm of the Mesen' syneclise, in the
northeastern part of the East-European Platform (, pers. comm.). These assemblages are stratigraphically below the first appearance of Metazoa. The large spherical acritarchs (100 - 200 µm in diameter, probably green algae, ,
2004), ornamented with various processes, are the most characteristic features of these assemblages. Among them
species of the genera Tanarium (as Goniosphaeridium), Cavaspina, Appendisphaera (as Meghystrichosphaeridium), Ericiasphaera and Multifrosphaeridium have been identified, as well as numerous algal filaments like Talakania, Glomovertella (= Volyniella) and Obruchevella.
The microfossil assemblages of Early? - Late Vendian age (0.6-0.55 Ma) are recorded from the Kursov and Parshin
formations of the Nepa-Botuoba region and from the Turkut Fm of the Lena-Anabar depression of Siberia ( & ,
1989; , 2004). There is
but one report of what is probably a contemporaneous microbiota from the Vychegoda Fm of the Mesen' syneclise, in the
northeastern part of the East-European Platform (, pers. comm.). These assemblages are stratigraphically below the first appearance of Metazoa. The large spherical acritarchs (100 - 200 µm in diameter, probably green algae, ,
2004), ornamented with various processes, are the most characteristic features of these assemblages. Among them
species of the genera Tanarium (as Goniosphaeridium), Cavaspina, Appendisphaera (as Meghystrichosphaeridium), Ericiasphaera and Multifrosphaeridium have been identified, as well as numerous algal filaments like Talakania, Glomovertella (= Volyniella) and Obruchevella.
![]() The most representative Late Vendian microbiotas (0.55-0.535 Ma) in Russia are from the Redkin and Kotlin Horizons of the East-European Platform. They are also recorded from the Buton Fm of the Serebryan Group, Middle Urals ( et alii,
1989) and from the White Sea region where strata also yielded an ediacarian macrofossil. In Siberia, late Vendian acritarch assemblages are documented from the Nemakit-Daldyn Fm of the Anabar Uplift ( & ,
1989). A diverse assemblage of silicified microfossils has been found in the Yudoma Group of the Uchur-Maya Region.
The most representative Late Vendian microbiotas (0.55-0.535 Ma) in Russia are from the Redkin and Kotlin Horizons of the East-European Platform. They are also recorded from the Buton Fm of the Serebryan Group, Middle Urals ( et alii,
1989) and from the White Sea region where strata also yielded an ediacarian macrofossil. In Siberia, late Vendian acritarch assemblages are documented from the Nemakit-Daldyn Fm of the Anabar Uplift ( & ,
1989). A diverse assemblage of silicified microfossils has been found in the Yudoma Group of the Uchur-Maya Region.
![]() In contrast to the early Vendian assemblages, those of the younger Vendian are dominated by cyanobacteria: Obruchevella and Glomovertella (= Volyniella) are
the most common. Filamentous algae Pomoria rhomboidalis (, 1985), , 1989, Striatella coriaceae , 1983, Palaeolyngbia sp., Oscillatoriopsis
sp., Siphonophycus sp. and the rare small acritarch Micrhystridium are associated with the cyanobacteria. A typical Late Vendian taxon is Cochleatina. The name is apt for it decribes chitinoid spirals with a complex morphology. In addition to those found on the east-European Platform, these fossils are recorded from the Nemakit-Daldyn Fm of the Anabar Uplift of Siberia and are interpreted as reproductive structures of algae adapted to subaerial environments (,
1996). Late Vendian fungi are represented by Vendomyces major ,
1993, which had been assigned by
(1993) to Chytridiomycetes. This form occurs in the Khatyspyt Fm of the Olenek Uplift, in the Nemakit-Daldyn Fm of the Anabar Uplift of Siberia and in the Kotla Fm of northwestern Russia. Other characteristic components of the Late Vendian biotas are the macro remains of multicellular Vendotaenia, a formal group of
Metaphyta and a specific bacterial organism Primoflagella attributed to Actinomycetes (,
1988). An important biological event of the uppermost Precambrian is the first appearance of calcareous algae, which in the Cambrian become one of the main bioherm-forming organisms of Siberia.
In contrast to the early Vendian assemblages, those of the younger Vendian are dominated by cyanobacteria: Obruchevella and Glomovertella (= Volyniella) are
the most common. Filamentous algae Pomoria rhomboidalis (, 1985), , 1989, Striatella coriaceae , 1983, Palaeolyngbia sp., Oscillatoriopsis
sp., Siphonophycus sp. and the rare small acritarch Micrhystridium are associated with the cyanobacteria. A typical Late Vendian taxon is Cochleatina. The name is apt for it decribes chitinoid spirals with a complex morphology. In addition to those found on the east-European Platform, these fossils are recorded from the Nemakit-Daldyn Fm of the Anabar Uplift of Siberia and are interpreted as reproductive structures of algae adapted to subaerial environments (,
1996). Late Vendian fungi are represented by Vendomyces major ,
1993, which had been assigned by
(1993) to Chytridiomycetes. This form occurs in the Khatyspyt Fm of the Olenek Uplift, in the Nemakit-Daldyn Fm of the Anabar Uplift of Siberia and in the Kotla Fm of northwestern Russia. Other characteristic components of the Late Vendian biotas are the macro remains of multicellular Vendotaenia, a formal group of
Metaphyta and a specific bacterial organism Primoflagella attributed to Actinomycetes (,
1988). An important biological event of the uppermost Precambrian is the first appearance of calcareous algae, which in the Cambrian become one of the main bioherm-forming organisms of Siberia.
![]() The last compilation on Precambrian microphytofossils from the former Soviet Union by et alii
(1989) provided descriptions of more than 300 species
assigned to 150 genera, and suggested recognition of three main epochs in their evolution: the Early-Middle Riphean, Late Riphean and Vendian. Since that time there have been many new finds of rich microbiotas previously unknown in Russia. Taxonomical concepts have changed slightly and the stratigraphical distribution of characteristic taxa had been modified. Although the suggested biological affinities of many of the acritarchs cited above remain to be confirmed, the Upper Proterozoic strata of Russia contain a very rich record of the evolution of the early biosphere, thus permitting the recognition and definition of five stratigraphical levels (R1-R21, R22, R3, V1?-V21 and V22) each characterized by the appearance of new taxa.
The last compilation on Precambrian microphytofossils from the former Soviet Union by et alii
(1989) provided descriptions of more than 300 species
assigned to 150 genera, and suggested recognition of three main epochs in their evolution: the Early-Middle Riphean, Late Riphean and Vendian. Since that time there have been many new finds of rich microbiotas previously unknown in Russia. Taxonomical concepts have changed slightly and the stratigraphical distribution of characteristic taxa had been modified. Although the suggested biological affinities of many of the acritarchs cited above remain to be confirmed, the Upper Proterozoic strata of Russia contain a very rich record of the evolution of the early biosphere, thus permitting the recognition and definition of five stratigraphical levels (R1-R21, R22, R3, V1?-V21 and V22) each characterized by the appearance of new taxa.
![]()
M. (1993).- Drevneyshiy khitridiomitset (Mycota, Chytridiomycetes incertae sedis) iz verkhnego venda Vostochno Evropeyskoy platformy [The oldest chytrid (Mycota, Chytridiomycetes incertae sedis) from the Upper Vendian of the East European Platform].- In: B.S. (ed.), Fauna i ekosistemy geologicheskogo proshlogo [Fauna and ecosystems of geological past].- Nauka, Moskva, p. 21-33 (in Russian).
M. (1996).- Redescription of the enigmatic microfossil Cochleatina from the Upper Vendian of the European Platform.- Paleontological Journal, Moscow, vol. 29, n° 2A (1995), p. 50-80.
N.J. (2000).- Bangiomorpha pubescens n. gen., n. sp.: implications for the evolution of sex, multicellularity and the Mesoproterozoic-Neoproterozoic radiation of eukaryotes.- Paleobiology, Chicago, vol. 26, n° 3, p. 386-404.
N.J. (2005).- Probable Proterozoic fungi.- Paleobiology, Chicago, vol. 31, n° 1, p. 165-182.
M. (1988).- Vendotenidy Vostochni-Evrope'skoy Platformy.- Nauka, Leningrad, 144 p. (in Russian).
V., M. & F. (1989).- Pervaya nahodka obruchevell v vendkih otlozheniyah Srednego Urala.- Doklady Akademii Nauk, Moskva, vol. 309, n° 3, p. 701-705 (in Russian).
T.N. (1981).- Nitchatye mikroorganizmy lahandinskoy svity reki Mai.- Paleontologicheskiy Zhurnal, Moskva, n° 2, p. 126-131 (in Russian).
T.N. (1990).- Organic world billion year ago.- Nauka, Leningrad, 49 p.
T.N. & V. (2002).- O nahodkah rifeyskih dimorfnyh organizmov.- Paleontologicheskiy Zhurnal, Moskva, n° 3, p. 3-10 (in Russian).
E.J., A.H. & M.R. (2001).- Morphological and ecological complexity in early eukaryotic ecosystems.- Nature, London, 412, p. 67-69.
M. (2005).- Taxonomic review of some Ediacarian acritarhs from the Siberian Platform.- Precambrian Research, Amsterdam, vol. 136, p. 283-307.
K. (2000).- Okremnehhye mikrobioty Verhnego Rifeya Eniseyskogo kryazha (Vostochnaya Sibir').- Novosti paleontologii i stratigraphii, Novosibirsk, n° 2-3, p. 7-31 (in Russian).
K. (2001).- Mikrofossilii i strstigraphiya Verhnego Rifeya yugo-zapadnoy chasti Sibirskoy Platformy.- Ph. D. Thesis, Novosibirsk, 24 p. (in Russian).
V. & N. (1989).- Talsinskiy komplex akritarh Nepsko-Botuobinskoy antiklizy.- In: N.A. (ed.), Fitostratigraphia i morfologiya spor drevnih rasteniy neftegazonosnyh provinsty SSSR.- Nauka, Leningrad, p. 5-11 (in Russian).
V. (1994).- Microfossils in cherts from the Middle Riphean (Mesoproterozoic) Avzyan Formation, southern Ural Mountains, Russian Federation.- Precambrian Research, Amsterdam, vol. 1, n° 2, p. 231-254.
V., A. & G. (1995).- Paleobiology of the Mesoproterozoic Billyakh Group, Anabar Uplift, Northern Siberia.- Paleontological Society, Memoir 39, Tulsa, 37 p.
V. (2001).- Paleobiology of the Neoproterozoic (Upper Riphean) Shorikha and Burovaya Silicified microbiotas, Turukhansk Uplift, Siberia.- Journal of Paleontology, Tulsa, vol. 75, n° 2, p. 427-448.
A., P. & N. (1998).- Miroedihinskaya mikrobiota verhnego rifeya Sibiri. Soobsheniye 1. Sostav i fastial'no-ecologocheskoye raspredeleniye organostennyh mikrofossiliy.- Stratigrafiya, Geologicheskaya Korrelyatsiya, Moskva, vol. 6, n° 5, p. 15-38 (in Russian).
A., P. & N. (2001).- Geohronologicheskiy i biostratigraphicheskiy podhody k rekonstuktsii istorii dokembrijskoy bioty: novye nahodki mikrofossiliy v rifee zapadnogo sklona Anabarskogo podnyatiya.- Doklady Akademii Nauk, Moskva, vol. 378, n° 4, p. 511-517 (in Russian).
A., V., N. & N. (2003).- Mikrofossilii tipovogo razreza verhnego rifeya (karatavskaya seruya Yuzhnogo Urala).- Stratigrafiya, Geologicheskaya Korrelyatsiya, Moskva, vol. 11, n° 6, p. 20-45 (in Russian).
A., D., Yu., N. & E. (2004).- Microfossils and Riphean Stratigraphy in the North European Platform (Mezen Syneclise).- Stratigraphy and Geological Correlation, Moscow, vol. 12, n° 6, p. 553-571.
T.V., N.S. & T.N. (1989).- Mikrofossilii dokembrija SSSR (Precambrian Microfossils in USSR).- Nauka, Leningrad, 190 p. (in Russian).
L.-M. (1998).- Acanthomorphic acritarchs from Meso-Neoproterozoic shales of the Ruyang Group, Shanxi, China.- Review of Palaeobotany and Palynology, Amsterdam, vol. 98, n° 1-2, p. 15-25.
![]()
Click on thumbnail to enlarge the image.
Figure 1: Geographical locations of the most distinctive Upper Proterozoic microbiotas of Russia.

Click on thumbnail to enlarge the image.
Figure 2: Cambrian acritarch-based biostratigraphy of the East-European Platform and stratigraphical positions of recorded acritarch data from the Siberian Platform.
![]()