![]()
![]()
C. (2005).- Early diversification of seeds and seed-like structures. In: P. & E. (eds.), Pre-Cambrian to Palaeozoic Palaeopalynology and Palaeobotany.- Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2005/02, Abstract 06 (CG2005_M02/06)
![]() Seeds; diversity; Devonian; Lower Carboniferous; evolution.
Seeds; diversity; Devonian; Lower Carboniferous; evolution.
![]() Graines ; diversité ; Dévonien ; Carbonifère inférieur ; évolution.
Graines ; diversité ; Dévonien ; Carbonifère inférieur ; évolution.
![]()
![]() The seed habit is one of the most important evolutionary acquisition in plant evolution. It allowed colonisation of new ecological niches, leading to a level of diversification never observed before. The seed habit involves occurrence of a single functional megaspore within an indehiscent megasporangium (nucellus), development of an endosporic megagametophyte, enclosure of the nucellus by an integument, and capture of pollen before dispersal from the sporophyte.
The seed habit is one of the most important evolutionary acquisition in plant evolution. It allowed colonisation of new ecological niches, leading to a level of diversification never observed before. The seed habit involves occurrence of a single functional megaspore within an indehiscent megasporangium (nucellus), development of an endosporic megagametophyte, enclosure of the nucellus by an integument, and capture of pollen before dispersal from the sporophyte.
![]() Early ovulate structures are better regarded as preovules (sensu & ,
1993) as their nucellus is surrounded by unfused or partially fused integumentary lobes, and they consequently lack a well-defined micropyle. The functions of the micropyle were than assumed by modifications of the nucellar apex.
Early ovulate structures are better regarded as preovules (sensu & ,
1993) as their nucellus is surrounded by unfused or partially fused integumentary lobes, and they consequently lack a well-defined micropyle. The functions of the micropyle were than assumed by modifications of the nucellar apex.
![]() Many preovules are known from Upper Devonian deposits (see Fig. 1
Many preovules are known from Upper Devonian deposits (see Fig. 1 ![]() ). They show a variety of morphologies, but they all share the same set of modifications of the nucellar apex: the hydrasperman syndrome (,
1986; & ,
1988). The nucellar apex is modified into a pollen chamber (sensu ,
1941) closed by a plinth (sensu ,
1941) and extended by a cylindrical structure (
1996), the salpinx (sensu ,
1941). The pollen chamber contains a central parenchymatous column. We here call lagenostome (sensu ,
1917) the whole modification of the nucellar apex, pollen chamber plus apex.
). They show a variety of morphologies, but they all share the same set of modifications of the nucellar apex: the hydrasperman syndrome (,
1986; & ,
1988). The nucellar apex is modified into a pollen chamber (sensu ,
1941) closed by a plinth (sensu ,
1941) and extended by a cylindrical structure (
1996), the salpinx (sensu ,
1941). The pollen chamber contains a central parenchymatous column. We here call lagenostome (sensu ,
1917) the whole modification of the nucellar apex, pollen chamber plus apex.
![]() All these Upper Devonian types show various degrees of fusion of the integumentary lobes and/or characteristics of the cupule, a set of vegetative segments that usually encloses the pre-ovulate structure. On the basis of the characteristics of these parts, the Upper Devonian preovules can be classified in the following types.
All these Upper Devonian types show various degrees of fusion of the integumentary lobes and/or characteristics of the cupule, a set of vegetative segments that usually encloses the pre-ovulate structure. On the basis of the characteristics of these parts, the Upper Devonian preovules can be classified in the following types.
![]() The
first type of preovule is the Moresnetia-type. It is characterised by a four unit cupule
formed by two successive cruciated dichotomous divisions. Within this group, we notice a progressive fusion of the integumentary lobes, ranging from completely dissected in the Belgian taxon Moresnetia zalesskyi
(1948), emend. et
(1987) [West European VCo biozone] to a nearly completely fused integument in Archaeosperma arnoldii et
(1968) [North American LE-LN biozone]. Xenotheca devonica et
(1915), emend. et
(1999) [West European LL biozone] and Elkinsia polymorpha et alii
(1989), emend. et
(1992) [North American VH biozone] have an intermediate degree of fusion. The degree of fusion of the integument can be observed only on perfectly preserved specimens. This led
(1999) to doubt the justification of
a distinction between the genera Moresnetia, Elkinsia and Xenotheca.
The
first type of preovule is the Moresnetia-type. It is characterised by a four unit cupule
formed by two successive cruciated dichotomous divisions. Within this group, we notice a progressive fusion of the integumentary lobes, ranging from completely dissected in the Belgian taxon Moresnetia zalesskyi
(1948), emend. et
(1987) [West European VCo biozone] to a nearly completely fused integument in Archaeosperma arnoldii et
(1968) [North American LE-LN biozone]. Xenotheca devonica et
(1915), emend. et
(1999) [West European LL biozone] and Elkinsia polymorpha et alii
(1989), emend. et
(1992) [North American VH biozone] have an intermediate degree of fusion. The degree of fusion of the integument can be observed only on perfectly preserved specimens. This led
(1999) to doubt the justification of
a distinction between the genera Moresnetia, Elkinsia and Xenotheca.
![]() We also include in the Moresnetia-type two slightly different taxa: Kerrya mattenii et
(1989) [West European LE/LN biozone] and Lenlogia krystofovichii
in et alii
(1962) emend. et
(1995) [LL Biozone]. Kerrya mattenii possesses a six unit cupule, but its overall organisation conforms to the Moresnetia-type. Lenlogia krystofovichii is poorly understood, but may be provisionally included in this group because of its apparent structural resemblance.
We also include in the Moresnetia-type two slightly different taxa: Kerrya mattenii et
(1989) [West European LE/LN biozone] and Lenlogia krystofovichii
in et alii
(1962) emend. et
(1995) [LL Biozone]. Kerrya mattenii possesses a six unit cupule, but its overall organisation conforms to the Moresnetia-type. Lenlogia krystofovichii is poorly understood, but may be provisionally included in this group because of its apparent structural resemblance.
![]() The
Moresnetia-type comprises the most ancient type of preovules. Its first representatives are found in the upper Frasnian of Russia ( et alii,
1988). This early
occurrence may be a partial explanation for its wide geographic distribution by late Famennian times (see Fig. 2
The
Moresnetia-type comprises the most ancient type of preovules. Its first representatives are found in the upper Frasnian of Russia ( et alii,
1988). This early
occurrence may be a partial explanation for its wide geographic distribution by late Famennian times (see Fig. 2 ![]() ). This type
ranged across the D/C boundary, for it is represented by the Carboniferous lagenostomalean seeds ovules (,
1993; , 1975).
). This type
ranged across the D/C boundary, for it is represented by the Carboniferous lagenostomalean seeds ovules (,
1993; , 1975).
![]() The second type
of preovule is the acupulate Aglosperma-type. It includes: Aglosperma quadrapartita
et (1996) [West European LL-LE biozone] and Aglosperma avonensis
(1998) [West European VI Biozone]. Their integument is formed by three to four flat lobes fused up to their lower third.
The second type
of preovule is the acupulate Aglosperma-type. It includes: Aglosperma quadrapartita
et (1996) [West European LL-LE biozone] and Aglosperma avonensis
(1998) [West European VI Biozone]. Their integument is formed by three to four flat lobes fused up to their lower third.
![]() An
adaptation to anemochory may be observed in the third and fourth types: the Warsteinia-type and the Dorinnotheca-type. Warsteinia paprothii
(1997) [West European LE biozone] is acupulate. Its integument is made of four winged lobes adnate or fused to the nucellus. The Dorinnotheca-type, represented by Dorinnotheca streelii,
(1996) [West European VCo], is putatively anemochoreous too. Its cupule is composed of eight parts fused proximally so that they form a cup, the segments of which divide to form at least 40 free endings. Each cupule contains only one central ovule. The integument is composed of four free terete lobes (,
1996).
An
adaptation to anemochory may be observed in the third and fourth types: the Warsteinia-type and the Dorinnotheca-type. Warsteinia paprothii
(1997) [West European LE biozone] is acupulate. Its integument is made of four winged lobes adnate or fused to the nucellus. The Dorinnotheca-type, represented by Dorinnotheca streelii,
(1996) [West European VCo], is putatively anemochoreous too. Its cupule is composed of eight parts fused proximally so that they form a cup, the segments of which divide to form at least 40 free endings. Each cupule contains only one central ovule. The integument is composed of four free terete lobes (,
1996).
![]() The fifth type is the Condrusia-type represented by three
described species, C. rumex (1948), C.
minor (1948) and C. brevis
in et
alii (1962). They differ from all the others in the characteristics of their cupule which is composed of two flat bilaterally symmetrical wings adpressed against each other. In the two species currently under investigation,
C. rumex and C. minor, the integument is not dissected into lobes and shows a complex organisation with three layers, the middle one consisting of large sclerified cells. The nucellus appears to be fused with the integument. Its apex is modified into a very long salpinx protruding above both the integument and the cupule wings.
The fifth type is the Condrusia-type represented by three
described species, C. rumex (1948), C.
minor (1948) and C. brevis
in et
alii (1962). They differ from all the others in the characteristics of their cupule which is composed of two flat bilaterally symmetrical wings adpressed against each other. In the two species currently under investigation,
C. rumex and C. minor, the integument is not dissected into lobes and shows a complex organisation with three layers, the middle one consisting of large sclerified cells. The nucellus appears to be fused with the integument. Its apex is modified into a very long salpinx protruding above both the integument and the cupule wings.
![]() In
addition, two new types of preovules occur in the Irish Hook Head locality ( & ,
1999; , 2000). They are both radially symmetrical and present the hydrasperman-type of reproduction. Their integument is made of four lobes fused basally and curving inward over the ovule apex. One of the two new preovules presents an integument composed of three distinct layers. However, without published description or illustration, further discussion is impossible.
In
addition, two new types of preovules occur in the Irish Hook Head locality ( & ,
1999; , 2000). They are both radially symmetrical and present the hydrasperman-type of reproduction. Their integument is made of four lobes fused basally and curving inward over the ovule apex. One of the two new preovules presents an integument composed of three distinct layers. However, without published description or illustration, further discussion is impossible.
![]() Some Upper Devonian seed-like structures are
not understood well enough to be placed in one or another group. They are
provisionally included in an heterogeneous type containing the Frasnian seed-like structure Sphinxia wuhania et alii
(1997), the enigmatic Spermolithus devonicus
(1917) and the two poorly described Belgian seeds "Xenotheca" bertrandii
(1948) and Pseudosporogonites hallei
(1948).
Some Upper Devonian seed-like structures are
not understood well enough to be placed in one or another group. They are
provisionally included in an heterogeneous type containing the Frasnian seed-like structure Sphinxia wuhania et alii
(1997), the enigmatic Spermolithus devonicus
(1917) and the two poorly described Belgian seeds "Xenotheca" bertrandii
(1948) and Pseudosporogonites hallei
(1948).
![]() Contrary
to earlier opinion, the preovules of the Upper Devonian are very diverse. This diversity, as well as the sophistication of the hydrasperman reproduction, strongly suggests an origin for early seeds older than the Frasnian. The Givetian taxon Runcaria heinzelinii
1948, emend. et alii
2004, probably represents one of the early stages in this evolution ( et alii,
2004). This taxon already possesses most of the characteristics of the Upper Devonian
preovules (see above). It consists of a short basal cuplike cupule made of four segments. This cupule contains a radially symmetrical megasporangium surrounded by an integument comprising at least 16 free lobes. The megasporangium bears a long distal extension emerging above the integument and enlarged at the top. The apical extension of the megasporangium probably played a role in the capture of microspores. Consequently, the reproductive biology of
Runcaria putatively included anemophilous pollination and dissolution of sporangial cells to allow fertilization ( et alii,
2004). The morphology of its megasporangium and its presumable particular type of reproduction distinguishes Runcaria from all the Famennian early seeds.
Contrary
to earlier opinion, the preovules of the Upper Devonian are very diverse. This diversity, as well as the sophistication of the hydrasperman reproduction, strongly suggests an origin for early seeds older than the Frasnian. The Givetian taxon Runcaria heinzelinii
1948, emend. et alii
2004, probably represents one of the early stages in this evolution ( et alii,
2004). This taxon already possesses most of the characteristics of the Upper Devonian
preovules (see above). It consists of a short basal cuplike cupule made of four segments. This cupule contains a radially symmetrical megasporangium surrounded by an integument comprising at least 16 free lobes. The megasporangium bears a long distal extension emerging above the integument and enlarged at the top. The apical extension of the megasporangium probably played a role in the capture of microspores. Consequently, the reproductive biology of
Runcaria putatively included anemophilous pollination and dissolution of sporangial cells to allow fertilization ( et alii,
2004). The morphology of its megasporangium and its presumable particular type of reproduction distinguishes Runcaria from all the Famennian early seeds.
![]() and
(2003) described
Spermasporites allenii, a Givetian seed-megaspore. The "seed-megaspores" are obligatory tetrads, with a unique functional and three aborted megaspores.
Spermasporites allenii seems to be enclosed in a sporangium. Whether or not this sporangium is indehiscent remains unknown. The relative ultrastructural simplicity of the outer megaspore layer and the presence of monomegaspory (sensu & ,
1994) led and
(2003) to suggest gymnospermous affinities for
Spermasporites. As the sporangium contains both micro- and megaspores, the structural differences and the uncertainties about dehiscence
seem to us rather suggestive of a lineage discrete from that of the seed plants. This taxon might be another example of the iterative (sensu & ,
1994) aspect of heterospory.
and
(2003) described
Spermasporites allenii, a Givetian seed-megaspore. The "seed-megaspores" are obligatory tetrads, with a unique functional and three aborted megaspores.
Spermasporites allenii seems to be enclosed in a sporangium. Whether or not this sporangium is indehiscent remains unknown. The relative ultrastructural simplicity of the outer megaspore layer and the presence of monomegaspory (sensu & ,
1994) led and
(2003) to suggest gymnospermous affinities for
Spermasporites. As the sporangium contains both micro- and megaspores, the structural differences and the uncertainties about dehiscence
seem to us rather suggestive of a lineage discrete from that of the seed plants. This taxon might be another example of the iterative (sensu & ,
1994) aspect of heterospory.
![]() After Devonian times, a gap is observed in the seed record. This remains enigmatic
(it was probably related to lack of adequate deposits) for a lot of vegetative parts assignable to seed-plants are found, but reproductive organs are
recorded either not at all or very rarely (, pers. com.). It is only at the PC biozone that an increase in the seed record is observed. A very peculiar type of seed,
Coumiasperma remyi ( et ,
1991) has been reported once. It is atypical because the usual hydrasperman nucellar apical modification is absent. This ovule is interpreted as showing adaptation to aquatic dispersal ( & ,
1991). More classical hydrasperman types are also described as
Gnetopsis hispida et
(1977) and
Lagenospermum imparirameum (1939) emend. et
(1977). Some other taxa of the Lydienne Formation are also mentioned ( & ,
1990; , 1856;
& ,
1997), but their affinities remain uncertain. However, some have been compared to the Moresnetia morphological group ( & ,
1990) or to some Carboniferous genera.
After Devonian times, a gap is observed in the seed record. This remains enigmatic
(it was probably related to lack of adequate deposits) for a lot of vegetative parts assignable to seed-plants are found, but reproductive organs are
recorded either not at all or very rarely (, pers. com.). It is only at the PC biozone that an increase in the seed record is observed. A very peculiar type of seed,
Coumiasperma remyi ( et ,
1991) has been reported once. It is atypical because the usual hydrasperman nucellar apical modification is absent. This ovule is interpreted as showing adaptation to aquatic dispersal ( & ,
1991). More classical hydrasperman types are also described as
Gnetopsis hispida et
(1977) and
Lagenospermum imparirameum (1939) emend. et
(1977). Some other taxa of the Lydienne Formation are also mentioned ( & ,
1990; , 1856;
& ,
1997), but their affinities remain uncertain. However, some have been compared to the Moresnetia morphological group ( & ,
1990) or to some Carboniferous genera.
![]() After the CM biozone [Lower Carboniferous], we observe a rapid and important expansion of the seed plants. The number of genera for isolated seeds increases (,
1975); a lot of types are known. This is the beginning of the worldwide dominance of seed plants.
After the CM biozone [Lower Carboniferous], we observe a rapid and important expansion of the seed plants. The number of genera for isolated seeds increases (,
1975); a lot of types are known. This is the beginning of the worldwide dominance of seed plants.
![]()
E.A.N. & R.H. (1915).- On some fossil plants from the Devonian rocks of North Devon.- Proceedings of the Cambridge Philosophical Society, vol. 18, p. 89104.
C.A. (1939).- Lagenospermum imparirameum sp. nov., a seedbearing fructification from the Missipian of Pennsylvania and Virginia.- Bulletin of the Torrey Botanical Club, Lawrence, vol. 6, p. 297-303.
R.M. & W.A. (1994).- Heterospory: the most iterative key innovation in the evolutionary history of the plant kingdom.- Biological Reviews, Cambridge, vol. 69, p. 345-417.
C.J. (1993).- Gymnospermophyta. In: M.J. (ed.), The Fossil Record 2.- London, Chapman & Hall.
R., E., M. & J. (1993).- Late Famennian to Namurian in the Eastern Ardenne, Belgium.- Subcommission on Carboniferous Stratigraphy, Guidebook, Services associés de paléontologie de l'Université de Liège, 60 p.
M. (1996).- Dorinnotheca streelii , gen. et sp. nov., a new early seed plant from the upper Famennian of Belgium.- Review of Palaeobotany and Palynology, Amsterdam, vol. 93, n° 1-4, p. 217-233.
M. & S.E. (1987).- Typification and redescription of Moresnetia zalesskyi , 1948, an early seed plant from the Upper Famennian of Belgium.- Bulletin de l'Institut Royal des Sciences naturelles de Belgique, Bruxelles, (Sciences de la Terre), vol. 57, p. 183-199.
J. & N.P. (1991).- A new permineralized seed-like structure from the basalmost Carboniferous of France.- Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen, Stuttgart, vol. 183, n° 1-3, p. 103-120.
P.G. & J.E. (1977).- Two Early Missipian seeds from the Price Formation of southwestern Virginia.- Brittonia, New York, vol. 29, n° 3, p. 332-351.
P., B., M., M. & P. (2004).- Runcaria, a Middle Devonian Seed Plant Precursor.- Science, Washington, vol. 306, p. 856-858.
W.T. (1941).- On Salpingostoma dasu: a new Carboniferous seed from East Lothian.- Transactions of the Royal Society of Edinburgh, Edinburgh, vol. 60, n° 12, p. 427-464.
J. (1996).- Famennian-Tournaisian plant assemblages from South-West Britain.- Ph.D. thesis (unpublished), University of Wales, Cardiff.
J. (1998).- Spermatophyte preovules from the basal Carboniferous of the Avon Gorge, Bristol.- Palaeontology, London, vol. 41, part 5, p. 1077-1091.
J. & D. (1995).- A new Late Devonian acupulate preovule from the Taff Gorge, South Wales.- Review of Palaeobotany and Palynology, Amsterdam, vol. 93, n° 1-4, p. 235-252.
J. & D. (1999).- New data on Xenotheca devonica & , an enigmatic early seed plant cupule with preovules. In: M.H. & A.R. (eds.), The Evolution of Plant Architecture.- Royal Botanical Gardens, Kew, p. 75-90.
A.L. (1988).- The Middle and Late Devonian floras of northern Eurasia.- Transactions of the Palaeontological Institute, Moskva, vol. 227, 176 p. (in Russian).
T. (1917).- Spermolithus devonicus gen. sp. nov., and other pteridosperms from the Upper Devonian beds of Kiltorkan.- Proceedings of the Royal Dublin Society, vol. 15, p. 245-254.
S.D. (2000).- Anatomically preserved gymnosperms from the Late Devonian of Ireland.- Botany 2000 "New Frontiers in Botany" (Portland, August 6-10, 2000), Abstract 17.
S. & L. (1999).- A new seed from the Upper Devonian of Southern Ireland.- XVI International Botanical Congress (Saint-Louis, August 1-7, 1999), Abstract.
V.A. & T.V. (1995).- Moresnetia-like plants from the Upper Devonian of Minusink basin, Siberia.- Paleontological Journal, Moscow, vol. 29, n° 1A, p. 35-43.
V.G., N.M. & G.P. (1962).- Main Devonian plants of the Altay-Sayan mountain region.- Vsesoyuznogo Nauchno-Issledovatel'skogo, Geologicheskogo Instituta (VSEGEI), Trudy, (Novaya Seriya), Leningrad, n° 70, 189 p.
C.-S., J. & A.R. (1997).- Frasnian (Upper Devonian) evidence for multiple origins of seed-like structures.- Botanical Journal of the Linnean Society, London, vol. 123, p. 133-146.
A.G. (1975).- Further observation on some Lower Carboniferous Seeds and Cupules.- Transactions of the Royal Society of Edinburgh, Edinburgh, vol. 69, n° 12, p. 267-293.
J.E. & A.R. (2003).- A Mid Devonian seed-megaspore from East Greenland an the origin of the seed plants.- Palaeontology, London, vol. 46, part 4, p. 647-670.
B. & N.P. (1997).- A Lower Carboniferous plant assemblage from Thuringia (Germany): compressions.- Review of Palaeobotany and Palynology, Amsterdam, vol. 97, n° 3-4, p. 361-379.
J.M. & C.B. (1968).- Archaeosperma arnoldii - a cupulate seed from the Upper Devonian of North America.- Contributions from the University of Michigan Museum of Paleontology, vol. 22, n° 10, p. 139-154.
G.W. (1986).- Classifying the earliest Gymnosperms. In: R.A. & B.A. (eds.), Systematic and Taxonomic Approaches in Palaeobotany.- Oxford University Press, Oxford, p. 137-161.
G.W. & S.E. (1988).- Biology of ancestral gymnosperms. In: C.B. (ed.), Origin and Evolution of Gymnosperms.- Columbia University Press, New York, p. 85-134.
G.W., S.E. & W.H. (1989).- Elkinsia gen. nov., a Late Devonian Gymnosperm with cupulate ovules.- Botanical Gazette, Chicago, vol. 150, n° 2, p. 170-189.
G.W. & D.C. (1989).- Pullaritheca longii gen. nov. and Kerryia mattenii gen. et sp. nov., Lower Carboniferous cupules with ovules of the Hydrasperma tenuis-type.- Review of Palaeobotany and Palynology, Amsterdam, vol. 60, n° 3-4, p. 295-309.
N.P. (1997).- Late Devonian winged preovules and their implication for the adaptative radiation of early seed plants.- Palaeontology, London, vol. 40, part 2, p. 575-595.
N.P. & J. (1990).- A Lower Carboniferous plant assemblage from La Serre (Montagne Noire, France). Part II. Gymnosperms.- Review of Palaeobotany and Palynology, Amsterdam, vol. 63, n° 1-2, p. 91-115.
C.R. (2002).- Paleomap website.- http://www.scotese.com
D.H. (1917).- The heterangiums of the British Coal Measures.- Botanical Journal of the Linnean Society, London, vol. 44, p. 59-105.
R. & G.W. (1992).- Characterizing the most primitive seed ferns. I. A reconstruction of Elkinsia polymorpha.- International Journal of Plant Sciences, Chicago, vol. 153, n° 4, p. 602-621.
W.N. & G.W. (1993).- Paleobotany and the evolution of plants.- Cambridge University Press, Cambridge, 535 p.
F. (1948).- Végétaux du Dévonien supérieur de la Belgique.- Mémoires du Musée Royal d'Histoire Naturelle de Belgique, Bruxelles, vol. 110, 84 p.
F. (1968).- Végétaux mésodévoniens récoltés aux confins du Massif du Brabant (Belgique).- Institut Royal des Sciences Naturelles de Belgique, Mémoires, Bruxelles, vol. 159, p. 1-49.
M., K., S., W. & P. (1987).- Spore stratigraphy and correlation with faunas and floras in the type marine Devonian of the Ardenne-Rhenish regions.- Review of Palaeobotany and Palynology, Amsterdam, vol. 50, n° 3, p. 211-229.
F. (1856).- Beitrag zur Palaeontologie des Thüringer Waldes. II.- Schiefer und Sandstein Flora.- Denkschriften der Akademie der Wissenschaften in Wien, vol. 11, p. 139186.
![]()
Click on thumbnail to enlarge the image.
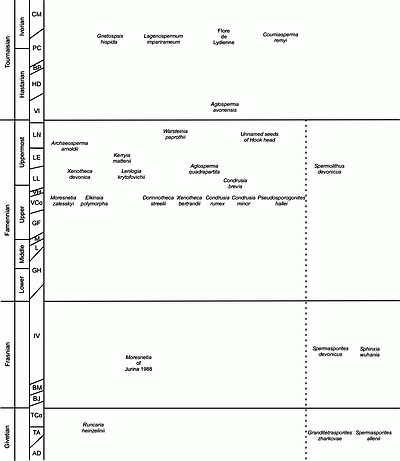
Figure 1: Stratigraphic distribution of Devonian and Lower Carboniferous ovules and ovule-like structures. Dotted line separates preovules (left) from ovule-like structures (right). See text for references. Stratigraphic scale modified from et alii (1993) and et alii (1987). AD. Acinosporites acanthomammilathus - Densosporites evonicus; BJ. Verrucosisporites bulliferus - Cirratriradites jekhowskyi; BM. Verrucosisporites bulliferus - Lophozonotriletes media; P. Spelaeotrilete balteatus - Rugospora polyptycha; CM. Schopfites claviger - Auroraspora macra; GF. Grandispora gracilis - G. famenensis; GH. Grandispora gracilis - A. hirtus; HD. Kraeuzelisporites hibernicus - Umbonatisporites distinctus; LE. Retispora lepidophyta - Indotriradites explanatus; LL. Retispora lepidophyta - Knoxisporites literatus; TA. Samarisporites triangulatus - Acyspora var. ancyrea; TCo. Samarisporites triangulatus - Chelinospora concinna; L; M; PC. Spelaeotriletes pretiosus - Raistrickia clavata; VCo. Diducites versabilis - Grandispora cornuta; VI. Vallatisporites allatus - Retusotriletes incohatus; LN. Retispora lepidophyta - Verucosiporites nitidus; VH. Apiculiretusispora verrucosa - Vallatisporites hystricosus.
Click on thumbnail to enlarge the image.
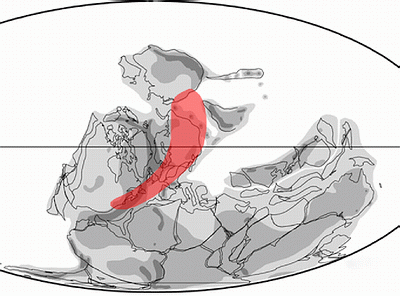
Figure 2 (modified from , 2002): In red the geographic area covered by the Moresnetia-type during the late Famennian. Data from & (1968), & (1987), (1988), et alii (1989), & (1989), & (1995) and & (1999).
![]()