![]()
![]()
C.V. (2005).- Ordovician to Lower Silurian palynomorphs from the Sierras subandinas (Subandean ranges), northwestern Argentina: a preliminary report. In: P. & E. (eds.), Pre-Cambrian to Palaeozoic Palaeopalynology and Palaeobotany.- Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2005/02, Abstract 09 (CG2005_M02/09)
![]() Ordovician; Silurian; palynomorphs; biostratigraphy; palaeogeography; palaeoenvironment.
Ordovician; Silurian; palynomorphs; biostratigraphy; palaeogeography; palaeoenvironment.
![]() Ordovicien ; Silurien ; palynomorphes ; biostratigraphie ; paléogéographie ; paléoenvironnement.
Ordovicien ; Silurien ; palynomorphes ; biostratigraphie ; paléogéographie ; paléoenvironnement.
![]()
![]() Ordovician clastic sediments of the Central Andean Basin, northwestern Argentina, were deposited in a proto-Andean foreland basin on the western margin of Gondwana (,
2003; & ,
2003). The Sierras Subandinas (Subandean Ranges) represent the outermost exposures of
this Ordovician basin, where proximal environmental settings and their related palynological content are
currently being analysed. These platform facies are exposed in the Eastern Cordillera, where palynological knowledge has increased remarkably in recent years (& , 2001; ,
2003). On the other hand palynological information on
the deep marine facies exposed in the Puna is still meager and involves only the
Ordovician-Silurian boundary ( & ,
2004) (Pl. 1
Ordovician clastic sediments of the Central Andean Basin, northwestern Argentina, were deposited in a proto-Andean foreland basin on the western margin of Gondwana (,
2003; & ,
2003). The Sierras Subandinas (Subandean Ranges) represent the outermost exposures of
this Ordovician basin, where proximal environmental settings and their related palynological content are
currently being analysed. These platform facies are exposed in the Eastern Cordillera, where palynological knowledge has increased remarkably in recent years (& , 2001; ,
2003). On the other hand palynological information on
the deep marine facies exposed in the Puna is still meager and involves only the
Ordovician-Silurian boundary ( & ,
2004) (Pl. 1 ![]() ).
).
![]() The outermost portion of the foreland basin, the Sierras Subandinas, is characterized by alternating shallow-marine deltaic systems and estuarine environments, which indicate repeated changes in the coast line
caused by fluctuations in relative sea-level ( & ,
2003) (Pl. 1
The outermost portion of the foreland basin, the Sierras Subandinas, is characterized by alternating shallow-marine deltaic systems and estuarine environments, which indicate repeated changes in the coast line
caused by fluctuations in relative sea-level ( & ,
2003) (Pl. 1 ![]() ). These authors undertook a detailed study of sequence stratigraphy in the Ordovician-earliest Silurian rocks of this region.
However, determination of their age is difficult because of the scarcity in some
units of biostratigraphically useful fossils.
). These authors undertook a detailed study of sequence stratigraphy in the Ordovician-earliest Silurian rocks of this region.
However, determination of their age is difficult because of the scarcity in some
units of biostratigraphically useful fossils.
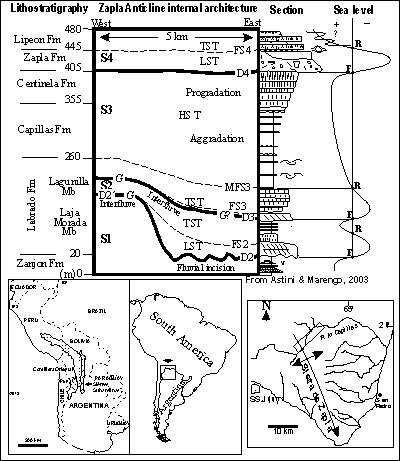
![]() Ordovician and earliest Silurian rocks are well exposed along the Río Capillas, in the Sierra de Zapla,
Jujuy Province (Fig. 1
Ordovician and earliest Silurian rocks are well exposed along the Río Capillas, in the Sierra de Zapla,
Jujuy Province (Fig. 1 ![]() ). The lowest unit is the Zanjón Formation, considered as middle-late Arenig. It is followed in succession by the Labrado Formation with its two sub-units, the late Arenig Laja Morada Member and the early-middle Llanvirn Lagunillas Member; the Capillas Formation, of late Llanvirn-basal Caradoc age; the Centinela Formation, considered to be not older than late Caradoc, and the Zapla Formation of Hirnantian age. The uppermost Ordovician is overlain by the Silurian Lipeón Formation that initiates a separate supercycle ( & ,
2003; et alii,
2003).
). The lowest unit is the Zanjón Formation, considered as middle-late Arenig. It is followed in succession by the Labrado Formation with its two sub-units, the late Arenig Laja Morada Member and the early-middle Llanvirn Lagunillas Member; the Capillas Formation, of late Llanvirn-basal Caradoc age; the Centinela Formation, considered to be not older than late Caradoc, and the Zapla Formation of Hirnantian age. The uppermost Ordovician is overlain by the Silurian Lipeón Formation that initiates a separate supercycle ( & ,
2003; et alii,
2003).
![]() This
work analyses the succession of palynological assemblages throughout the Ordovician and lower Silurian, and
attempts to determine the biostratigraphic control of the units, their palaeogeographic affinities, and the palaeoenvironmental changes related to recurrent fluctuations in sea level.
This
work analyses the succession of palynological assemblages throughout the Ordovician and lower Silurian, and
attempts to determine the biostratigraphic control of the units, their palaeogeographic affinities, and the palaeoenvironmental changes related to recurrent fluctuations in sea level.
![]() All units involved in this study yielded palynomorph assemblages that include acritarchs and related marine forms, chitinozoans and cryptospores (Pl. 1
All units involved in this study yielded palynomorph assemblages that include acritarchs and related marine forms, chitinozoans and cryptospores (Pl. 1 ![]() ). Their abundance, diversity and preservation
range widely in accordance with stratigraphic level and sedimentary facies.
). Their abundance, diversity and preservation
range widely in accordance with stratigraphic level and sedimentary facies.
![]() Detailed descriptions and illustrations of the palynological assemblages from all units discussed herein are currently in progress.
Detailed descriptions and illustrations of the palynological assemblages from all units discussed herein are currently in progress.
![]() The Zanjón Formation is characterized by repeated tidal parasequences that shallow-upward into heterolithic and muddy intervals. It yielded a rich acritarch assemblage containing Striatotheca principalis parva 1970, Arbusculidium filamentosum () 1972 emend.
et 1999, Aureotesta clathrata var. simplex ( et alii) emend et alii 1998, Cymatiogalea granulata 1966, Cymatiogalea messaoudensis et alii 1974 var. messaoudensis autonym, Rhopaliophora palmata (
et ) et 1984, Dactylofusa velifera forma brevis 1989, Eisenackidium orientalis
in et alii, 1999, Coryphidium sp., and Vogtlandia sp.. This assemblage can be correlated with acritarchs of the Acoite Formation in the Cordillera Oriental. Consequently, its age is not younger than middle Arenig, although stratigraphic correlation suggests that it includes the late Arenig.
The Zanjón Formation is characterized by repeated tidal parasequences that shallow-upward into heterolithic and muddy intervals. It yielded a rich acritarch assemblage containing Striatotheca principalis parva 1970, Arbusculidium filamentosum () 1972 emend.
et 1999, Aureotesta clathrata var. simplex ( et alii) emend et alii 1998, Cymatiogalea granulata 1966, Cymatiogalea messaoudensis et alii 1974 var. messaoudensis autonym, Rhopaliophora palmata (
et ) et 1984, Dactylofusa velifera forma brevis 1989, Eisenackidium orientalis
in et alii, 1999, Coryphidium sp., and Vogtlandia sp.. This assemblage can be correlated with acritarchs of the Acoite Formation in the Cordillera Oriental. Consequently, its age is not younger than middle Arenig, although stratigraphic correlation suggests that it includes the late Arenig.
![]() The lower subdivision of the Labrado Formation, the Laja Morada Member, records exposure during a lowering of relative sea level indicated by subaerial features and a gradual change toward purple-red colours near the top. Consequently, the only palynomorphs present are leiospheres and other prasinophycean algae
such as Cymatiosphaera, all devoid of stratigraphic value. The upper Lagunillas Member, representing restricted estuarine facies, yielded a poorly diversified assemblage with some acritarchs such as ? Aremoricanium simplex
et 1971, in addition to leiospheres.
The lower subdivision of the Labrado Formation, the Laja Morada Member, records exposure during a lowering of relative sea level indicated by subaerial features and a gradual change toward purple-red colours near the top. Consequently, the only palynomorphs present are leiospheres and other prasinophycean algae
such as Cymatiosphaera, all devoid of stratigraphic value. The upper Lagunillas Member, representing restricted estuarine facies, yielded a poorly diversified assemblage with some acritarchs such as ? Aremoricanium simplex
et 1971, in addition to leiospheres.
![]() Transition to the open marine deposits of the Capilla Formation is indicated by a maximum flooding surface. This formation yielded a rich and well-preserved acritarch assemblage containing such taxa as Arbusculidium filamentosum, Striatotheca
spp., Arkonia sp., Dactylofusa cf. D. striatogranulata et alii
1974, Ericanthea pollicipes et 1977, Liliosphaeridium cf. L. intermedium () et alii 1995 and Leprotolypa evexa 1979. It also contains cryptospores of the
morphon Dyadospora murusattenuata et
1979 sensu et alii 1996, and Sphaerasaccus glabellus et alii 2000. The latest record of Arbusculidium filamentosum, independently dated, is probably lowermost Llanvirn.
But other less well age-constrained records of its occurrence attain the Llanvirn
and may reach the Caradoc strata ( et alii,
2003). The genus Striatotheca may also extend up to the Caradoc ( &
,
2004). The genus Arkonia, and the species Ericanthea pollicipes and Liliosphaeridium cf. L. intermedium were recorded from Llanvirn strata ( et alii,
2003; & ,
2004) and Dactylofusa striatogranulata, previously known only in the Ashgill has recently been found in the middle Arenig (uppermost Lower Ordovician) of Argentina ( et alii,
in press). Furthermore, Leprotolypa evexa is unknown below the base of the Caradoc ( & ,
2004). Although there is some dispersion in the stratigraphic ranges of
the acritarch species, a late Llanvirn-earliest Caradoc age assignment for the Capilla
Formation based on the presence of Sacabambaspis janvieri (Vertebrata), roughly agrees with the age inferred from the palynomorph assemblages.
Transition to the open marine deposits of the Capilla Formation is indicated by a maximum flooding surface. This formation yielded a rich and well-preserved acritarch assemblage containing such taxa as Arbusculidium filamentosum, Striatotheca
spp., Arkonia sp., Dactylofusa cf. D. striatogranulata et alii
1974, Ericanthea pollicipes et 1977, Liliosphaeridium cf. L. intermedium () et alii 1995 and Leprotolypa evexa 1979. It also contains cryptospores of the
morphon Dyadospora murusattenuata et
1979 sensu et alii 1996, and Sphaerasaccus glabellus et alii 2000. The latest record of Arbusculidium filamentosum, independently dated, is probably lowermost Llanvirn.
But other less well age-constrained records of its occurrence attain the Llanvirn
and may reach the Caradoc strata ( et alii,
2003). The genus Striatotheca may also extend up to the Caradoc ( &
,
2004). The genus Arkonia, and the species Ericanthea pollicipes and Liliosphaeridium cf. L. intermedium were recorded from Llanvirn strata ( et alii,
2003; & ,
2004) and Dactylofusa striatogranulata, previously known only in the Ashgill has recently been found in the middle Arenig (uppermost Lower Ordovician) of Argentina ( et alii,
in press). Furthermore, Leprotolypa evexa is unknown below the base of the Caradoc ( & ,
2004). Although there is some dispersion in the stratigraphic ranges of
the acritarch species, a late Llanvirn-earliest Caradoc age assignment for the Capilla
Formation based on the presence of Sacabambaspis janvieri (Vertebrata), roughly agrees with the age inferred from the palynomorph assemblages.
![]() The Centinela Formation contains mainly leiospheres, pre-cryptospores and simple
acanthomorph acritarchs, and testifies to a possible progradation of localized deltaic complexes. At its upper
limit a regional unconformity correlated with a major eustatic fall in sea-level caused by the Hirnantian
glaciation separates the Centinela Formation from the uppermost Ordovician Zapla Formation.
The Centinela Formation contains mainly leiospheres, pre-cryptospores and simple
acanthomorph acritarchs, and testifies to a possible progradation of localized deltaic complexes. At its upper
limit a regional unconformity correlated with a major eustatic fall in sea-level caused by the Hirnantian
glaciation separates the Centinela Formation from the uppermost Ordovician Zapla Formation.
![]() The palynomorph assemblages of the Zapla glacial horizon are dominated by marine elements such as Villosacapsula sp., subordinate
acanthomorph acritarchs (Polygonium, Micrhystridium, Multiplicisphaeridium) and leiospheres, but also contain terrestrial cryptospores (mainly tetrads). Stratigraphically, the Zapla Formation is dated latest Ordovician, probably corresponding to the Hirnantian Glaciation. However, chitinozoans indicate an Aeronian s.l. to early Telychian age ( & ,
2001). Acritarch assemblages are meager and not well
preserved so they do not provide accurate stratigraphic control. In addition, transgressive-regressive events frequently caused reworking of fossils. Nevertheless, it should be noted that no typical Silurian forms were found among acritarchs or cryptospores. Further investigations, particularly on chitinozoans from the levels
under discusssion, may shed some light on this controversy.
The palynomorph assemblages of the Zapla glacial horizon are dominated by marine elements such as Villosacapsula sp., subordinate
acanthomorph acritarchs (Polygonium, Micrhystridium, Multiplicisphaeridium) and leiospheres, but also contain terrestrial cryptospores (mainly tetrads). Stratigraphically, the Zapla Formation is dated latest Ordovician, probably corresponding to the Hirnantian Glaciation. However, chitinozoans indicate an Aeronian s.l. to early Telychian age ( & ,
2001). Acritarch assemblages are meager and not well
preserved so they do not provide accurate stratigraphic control. In addition, transgressive-regressive events frequently caused reworking of fossils. Nevertheless, it should be noted that no typical Silurian forms were found among acritarchs or cryptospores. Further investigations, particularly on chitinozoans from the levels
under discusssion, may shed some light on this controversy.
![]() The Lipeón Formation initiates a new supercycle with the deposition of oolitic ironstones during the transgression that followed the Hirnantian glaciation. These Silurian deposits correspond to a series of high-frequency sea-level fluctuations. In the area
studied the lowermost levels of the Lipeón Formation have yielded abundant, diverse and well-preserved palynomorphs. The assemblage is dominated by acritarchs and prasinophyte algae, but also contains cryptospores and chitinozoans. Acritarch species include Domasia trispinosa 1960, Domasia elongata 1960, Dactylofusa estillis
et 1972, Dactylofusa maranhensis
et 1965, Beromia rexroadii 1996, Baiomeniscus camurus 1970, Carminella maplewoodensis 1968, Eupoikilofusa striatifera () 1970, among others. Cryptospores are represented by tetrads and also Imperfectotriletes vavrdovae () et alii 2000 and Laevolancis chibrikovae et alii 2000. The acritarch assemblage shares some species with those described by &
(1982) from the lower part of the unit, dated late Llandovery-early Wenlock, and also
has species in common with acritarch assemblages from the Vargas Peña Formation, in Paraguay ( & ,
1997). The palynomorphs of the Lipeón Formation have particularly strong affinities with the assemblage of the Tianguá Formation, in the Brazilian Parnaíba Basin ( et alii,
2001). In the Subandean Ranges, the lower part of this unit was referred to the early Llandovery (Parakidograptus acuminatus- Atavograptus atavus graptolite biozones) by et alii (2002). On the other hand, in the Río Capillas section, chitinozoans indicate a late Telychian age for the lower part of this formation ( & , 2001). Nevertheless, the acritarch assemblage of the Lipeón Formation suggests an age probably near the Aeronian/Telychian boundary and not older than late Aeronian ( & , in press).
The Lipeón Formation initiates a new supercycle with the deposition of oolitic ironstones during the transgression that followed the Hirnantian glaciation. These Silurian deposits correspond to a series of high-frequency sea-level fluctuations. In the area
studied the lowermost levels of the Lipeón Formation have yielded abundant, diverse and well-preserved palynomorphs. The assemblage is dominated by acritarchs and prasinophyte algae, but also contains cryptospores and chitinozoans. Acritarch species include Domasia trispinosa 1960, Domasia elongata 1960, Dactylofusa estillis
et 1972, Dactylofusa maranhensis
et 1965, Beromia rexroadii 1996, Baiomeniscus camurus 1970, Carminella maplewoodensis 1968, Eupoikilofusa striatifera () 1970, among others. Cryptospores are represented by tetrads and also Imperfectotriletes vavrdovae () et alii 2000 and Laevolancis chibrikovae et alii 2000. The acritarch assemblage shares some species with those described by &
(1982) from the lower part of the unit, dated late Llandovery-early Wenlock, and also
has species in common with acritarch assemblages from the Vargas Peña Formation, in Paraguay ( & ,
1997). The palynomorphs of the Lipeón Formation have particularly strong affinities with the assemblage of the Tianguá Formation, in the Brazilian Parnaíba Basin ( et alii,
2001). In the Subandean Ranges, the lower part of this unit was referred to the early Llandovery (Parakidograptus acuminatus- Atavograptus atavus graptolite biozones) by et alii (2002). On the other hand, in the Río Capillas section, chitinozoans indicate a late Telychian age for the lower part of this formation ( & , 2001). Nevertheless, the acritarch assemblage of the Lipeón Formation suggests an age probably near the Aeronian/Telychian boundary and not older than late Aeronian ( & , in press).
![]() Although still in a very preliminary stage, the analyses in this contribution provide biostratigraphic, palaeogeographic and palaeonvironmental information about the palynomorph assemblages of the Central Andean
Basin. They cover almost all of Ordovician, the Ordovician/Silurian transition and the
Lower Silurian.
Although still in a very preliminary stage, the analyses in this contribution provide biostratigraphic, palaeogeographic and palaeonvironmental information about the palynomorph assemblages of the Central Andean
Basin. They cover almost all of Ordovician, the Ordovician/Silurian transition and the
Lower Silurian.
![]() The acritarch assemblages of the Zanjón Formation provide the first age constraint for this unit.
For the Capillas Formation, acritarch assemblages support the age suggested by other fossils.
The palynomorph assemblages of the Zapla and Lipeón formations allow an independent age assessment.
The acritarch assemblages of the Zanjón Formation provide the first age constraint for this unit.
For the Capillas Formation, acritarch assemblages support the age suggested by other fossils.
The palynomorph assemblages of the Zapla and Lipeón formations allow an independent age assessment.
![]() From a palaeogeographic viewpoint, it is significant that acritarchs from the Zanjón Formation clearly show affinities to those of the peri-Gondwana province. Acritarchs of the lower Silurian also have Gondwanian affinities.
From a palaeogeographic viewpoint, it is significant that acritarchs from the Zanjón Formation clearly show affinities to those of the peri-Gondwana province. Acritarchs of the lower Silurian also have Gondwanian affinities.
![]() Palynological assemblages recovered from the several depositional settings vary in diversity, composition and abundance, thus indicating
the existence of a relationship between palynomorphs and local
palaeoenvironments.
Palynological assemblages recovered from the several depositional settings vary in diversity, composition and abundance, thus indicating
the existence of a relationship between palynomorphs and local
palaeoenvironments.
![]()
![]() Financial support for this study was provided by the ANPCyT-FONCyT (PICT 99 No 5387, PICT 2000 No 8920) and the CONICET. The author is grateful to J.H.G. (PETROBRAS, Brazil) for reviewing the English version. C. (Sheffield, UK) and P. (Liège, Belgium) are thanked for their advice concerning miospores and A. (Brest, France) for the review of the manuscript and useful suggestions The author benefited from a NFSR post-doctoral grant at the University of Liège.
Financial support for this study was provided by the ANPCyT-FONCyT (PICT 99 No 5387, PICT 2000 No 8920) and the CONICET. The author is grateful to J.H.G. (PETROBRAS, Brazil) for reviewing the English version. C. (Sheffield, UK) and P. (Liège, Belgium) are thanked for their advice concerning miospores and A. (Brest, France) for the review of the manuscript and useful suggestions The author benefited from a NFSR post-doctoral grant at the University of Liège.
A., C.V. & R.A. (in press).- Chitinozoans and acritarchs from the Ordovician peri-Gondwana volcanic arc of the Famatina System, northwestern Argentina.- Review of Palaeobotany and Palynology, Amsterdam.
R.A. (2003).- Ordovician basins of Argentina. In: J.L. (ed.), Ordovician fossils of Argentina.- Secretaría de Ciencia y Tecnología, Universidad Nacional de Córdoba, Córdoba, p. 1-74.
R.A. & L. (2003).- Sequence stratigraphy of Perigondwanic Ordovician clastics at the outermost reach of the Protoandean Foreland, Sierras Subandinas, Argentina.- 3rd Latin American Congress of Sedimentology (June 8-11, 2003), Belém, Extended Abstracts Volume.
R.A., L. & C.V. (2003).- The Ordovician stratigraphy of the Sierras Subandinas (Subandean Ranges) in northwest Argentina and its bearing on an integrated foreland basin model for the Ordovician of the Central Andean Region. In: G.L., M.S. & S.H. (eds.), Ordovician from the Andes. Proceedings of the 9th International Symposium on the Ordovician System.- INSUGEO, Serie de Correlación Geológica, Tucumán, 17, p. 381-386.
P. & F. (1982).- Conodontes et Acritarches de l'Ordovicien inférieur et acritarches du Silurien inférieur de la partie septentrionale de la Cordillère Argentine.- Bulletin de l'Institut Royal des Sciences naturelles de Belgique, Bruxelles, (Sciences de la Terre), vol. 53, n° 4, p. 1-21.
Y. & P.R. (2001).- Silurian and Middle Devonian Chitinozoa from the Zapla and Santa Bárbara Ranges, Tarija Basin, northwestern Argentina.- Ameghiniana, Buenos Aires, 38(1), p. 35-50.
A., J.H.G., L.P., Y. & P. (2001).- Palynological characterization and dating of the Tianguá Formation, Serra Grande Group, northern Brazil. In: J.H.G. & G.J.S. (eds.), Correlação de Sequências Paleozóicas Sul-Americanas.- Ciência-Técnica-Petróleo. Seção: Exploração de Petróleo, Rio de Janeiro, 20, p. 25-40.
R.B., G., M., M.A. & C.R. (2002).- Talacastograptus, an unusual biserial graptolite, and other Silurian forms from Argentina and Bolivia.- Ameghiniana, Buenos Aires, vol. 39 (3), p. 343-350.
C.V. (2003).- Ordovician acritarchs from northwestern argentina: new insight into the biostratigraphy and paleonvironmental aspects of the Central Andean Basin and Famatina. In: G.L., M.S. & S.H. (eds.), Ordovician from the Andes. Proceedings of the 9th International Symposium on the Ordovician System.- INSUGEO, Serie de Correlación Geológica, Tucumán, 17, p. 125-130.
C.V. & B.A. (2001).- Review of acritarch biostratigraphy in the Arenig of Eastern Cordillera, northwestern Argentina. New data and calibration with the graptolite zonation. In: R.H. (ed.), Contributions to Geology and Palaeontology of Gondwana.- Geological Institute, University of Cologne, p. 421-439.
C.V. & B.A. (in press).- Aeronian (Llandovery, Lower Silurian) palynomorphs and graptolites from the Lipeón Formation, Eastern Cordillera, north-west Argentina.- Geobios, Lyon.
C.V. & N.E. (2004).- Cryptospore assemblages from the Ordovician/Silurian boundary in the Puna, North-West Argentina.- Palaeontology, London, vol. 47, part 4, p. 1037-1061.
M., L. & A. (2003).- Lower Yushanian to lower Zhejiangian palynology of the Yangtze Gorges area (Daping and Huanghuachang sections), Hubei Province, South China.- Palaeontographica, Abteilung B, Stuttgart, vol. 266, p. 1-160.
M. & A. (2004).- Biostratigraphy, taxonomic diversity, and patterns of morphological evolution of Ordovician acritarchs (organic-walled microphytoplakton) from the northern Gondwana margin in relation to palaeoclimatic and palaeogeographic changes.- Earth-Science Reviews, Amsterdam, 67, p. 267-311.
G.D. & M. (1997).- The stratigraphic and paleoecologic importance of acritarchs, chitinozoans and spores from the Silurian Vargas Peña Shale, Paraguay.- 2nd European Meeting on the Palaeontology and Stratigraphy of South America (Heildelberg, September 2-4, 1997), Poster.
![]()
Click on thumbnail to enlarge the image.
Figure 1: Location map of the study area, in the Sierras Subandinas, north-west Argentina. Ordovician and Lower Silurian stratigraphy of the Sierra de Zapla, recognized sequences, lithologies and sea-level curve from & , 2003.
![]()

Click on thumbnail to enlarge the image.
Plate 1: The palynological slides are housed in the Paleopalynological Slide Collection of the Unit of Paleopalynology, IANIGLA, CRICyT, Mendoza. Specimen locations are referred to using England Finder coordinates.
1- Eisenackidium orientalis in et alii, 1999. Zanjón Fm, 7999d, U33/1.
2- Cymatiogalea granulata 1966. Zanjón Fm, 7999d, K41/2.
3- Dactylofusa velifera forma brevis 1989. Zanjón Fm, 8000d, N35/3.
4- Cymatiogalea messaoudensis et alii 1974 var. messaoudensis autonym. Zanjón Fm, 7999c, E34/0.
5- Striatotheca principalis parva 1970. Zanjón Fm, 7587i, A24/1.
6- Aureotesta clathrata var. simplex ( et alii) emend et alii 1998. Zanjón Fm, 7999d, C37/2.
7- ? Aremoricanium simplex et 1971. Labrado Fm, Lagunilla Member, 7595a, M26/1.
8- Arkonia sp. Capillas Fm, 7589g, N38/0.
9- Liliosphaeridium cf. L. intermedium () et alii 1995. Capillas Fm, 7589c, W45/4.
10- Ericanthea pollicipes et 1977. Capillas Fm, 7589h, V40/2.
11- Leprotolypa evexa 1979. Capillas Fm, 7588c, B22/1.
12- Morphon Dyadospora murusattenuata
et 1979 sensu et alii 1996. Fm. Capillas, 7589h, F23/0.
13- Sphaerasaccus glabellus et alii 2000. Capillas Fm, 7589h, P36/1.
14- Villosacapsula sp. Zapla Fm, 7590e, B23/0.
15- Tetrad 1. Zapla Fm, 7590d, W35/3.
16- Tetrad 2. Zapla Fm, 7583d, S28/0.
17- Carminella maplewoodensis 1968. Lipeón Fm, 7592d, W43/0.
18- Dactylofusa maranhensis et 1965. Lipeón Fm, 7582e, Y34/2.
19- Domasia elongata 1960. Lipeón Fm, 7582e, L33/2.
20- Dactylofusa estillis et 1972. Lipeón Fm, 7592g, P30/0.
21- Beromia rexroadii 1996. Lipeón Fm, 7582e, M27/0.
22- Baiomeniscus camurus 1970. Lipeón Fm, 7592g, Y38/0.
23- Laevolancis chibrikovae et alii 2000. Lipeón Fm, 7592f, Y44/2.
24- Imperfectotriletes vavrdovae () et alii
2000. Lipeón Fm, 7582e, F25/2.
![]()