![]()
![]()
J., J., A., N. & J. (2005).- New evidence for the Hirnantian (Upper Ordovician) in Belgium? An integrated isotopical, biostratigraphic and sedimentologic approach. In: P. & E. (eds.), Pre-Cambrian to Palaeozoic Palaeopalynology and Palaeobotany.- Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2005/02, Abstract 12 (CG2005_M02/12)
![]() Ordovician; Hirnantian; glaciation; chitinozoans; sedimentology; stable carbon isotopes; Condroz Inlier.
Ordovician; Hirnantian; glaciation; chitinozoans; sedimentology; stable carbon isotopes; Condroz Inlier.
![]() Ordovicien ; Hirnantien ; glaciation ; chitinozoaires ; sédimentologie ; isotopes stables du carbone ; Massif du Condroz.
Ordovicien ; Hirnantien ; glaciation ; chitinozoaires ; sédimentologie ; isotopes stables du carbone ; Massif du Condroz.
![]()
![]() The Hirnantian is an important time in Earth history as it includes one of the major episodes of continental glaciation and marine mass extinction (e.g. et alii,
1994). Positive C-isotope excursions and an eustatic lowering of sea level extend from a level near the base of the Normalograptus extraordinarius-N. ojsuensis Zone (lower Hirnantian) to a level within the upper Hirnantian N. persculptus Zone. C-isotope values decline to pre-glacial levels throughout the extent of uppermost Hirnantian strata. At least two discrete and distinct positive shifts of δ13Corg have been observed globally; they may coincide with two pulses of peak glaciation ( et alii,
2003; see also et alii,
2000).
The Hirnantian is an important time in Earth history as it includes one of the major episodes of continental glaciation and marine mass extinction (e.g. et alii,
1994). Positive C-isotope excursions and an eustatic lowering of sea level extend from a level near the base of the Normalograptus extraordinarius-N. ojsuensis Zone (lower Hirnantian) to a level within the upper Hirnantian N. persculptus Zone. C-isotope values decline to pre-glacial levels throughout the extent of uppermost Hirnantian strata. At least two discrete and distinct positive shifts of δ13Corg have been observed globally; they may coincide with two pulses of peak glaciation ( et alii,
2003; see also et alii,
2000).
![]() This study documents the sedimentology, the δ13C of dispersed organic carbon (δ13Corg) and the chitinozoan biostratigraphy of the presumed Hirnantian (Upper Ordovician) sediments of the Génicot Formation (Puagne Inlier, Condroz Inlier, Belgium). It provides new evidence to support the premise that two conglomerates, present in the mainly silty sediments of the Génicot Formation, may be the lithologic demonstration of a drop in sea level caused by the Hirnantian glaciation ( et alii,
2002; & ,
2004).
This study documents the sedimentology, the δ13C of dispersed organic carbon (δ13Corg) and the chitinozoan biostratigraphy of the presumed Hirnantian (Upper Ordovician) sediments of the Génicot Formation (Puagne Inlier, Condroz Inlier, Belgium). It provides new evidence to support the premise that two conglomerates, present in the mainly silty sediments of the Génicot Formation, may be the lithologic demonstration of a drop in sea level caused by the Hirnantian glaciation ( et alii,
2002; & ,
2004).
![]() The Condroz Inlier (or
"Bande de Sambre-et-Meuse") is a narrow strip of outcropping Ordovician and Silurian rocks, 65 km long and 0.5 to 4 km wide. The southwestern part of the Condroz Inlier, the Puagne Inlier, is located on the community grounds of Sart-Eustache. It is
a tectonic wedge carried along the Midi Overthrust or one of its satellites during Variscan orogeny.
The Condroz Inlier (or
"Bande de Sambre-et-Meuse") is a narrow strip of outcropping Ordovician and Silurian rocks, 65 km long and 0.5 to 4 km wide. The southwestern part of the Condroz Inlier, the Puagne Inlier, is located on the community grounds of Sart-Eustache. It is
a tectonic wedge carried along the Midi Overthrust or one of its satellites during Variscan orogeny.
![]() The Upper Ordovician Series of the Condroz Inlier consist of the Fosses Formation (upper Caradoc to upper Rawtheyan, middle to upper Ashgill) and the Génicot Formation (Hirnantian-lowermost Silurian?). The Fosses Formation is divided into two members (,
1968 after , 1934): the lower Bois de Presles Member, consisting of macrofossiliferous calcareous shale and some limestone levels and the upper Faulx-les-Tombes Member, consisting of green-grey mottled siltstone.
The Upper Ordovician Series of the Condroz Inlier consist of the Fosses Formation (upper Caradoc to upper Rawtheyan, middle to upper Ashgill) and the Génicot Formation (Hirnantian-lowermost Silurian?). The Fosses Formation is divided into two members (,
1968 after , 1934): the lower Bois de Presles Member, consisting of macrofossiliferous calcareous shale and some limestone levels and the upper Faulx-les-Tombes Member, consisting of green-grey mottled siltstone.
![]() The section studied is located north of the Étang du Diable, in the castle park of Sart-Eustache. Its outcrop extends from the southern old stone entrance of the park to 125 m north of it. It includes the Bois de Presles Member, a reduced expression of the Faulx-les-Tombes Member of the Fosses Formation, and the Génicot Formation, upper portion not exposed.
The section studied is located north of the Étang du Diable, in the castle park of Sart-Eustache. Its outcrop extends from the southern old stone entrance of the park to 125 m north of it. It includes the Bois de Presles Member, a reduced expression of the Faulx-les-Tombes Member of the Fosses Formation, and the Génicot Formation, upper portion not exposed.
![]() The microfacies of the Bois de Presles Member consists of bioturbated clayey micaceous silty mudstones and wackestones with various bioclasts (bryozoans, brachiopods, ostracods, pelecypods and echinoderms) and abundant unidentified microbioclasts. The matrix is finely recrystallized into a homogeneous calcite microspar. Microscopic organic debris is abundant. In the less-burrowed sediments, bioclasts form both thin (< 1 mm) and thicker (a few millimeters) packstone-grainstone levels. The grains display parallel lamination and local cross-bedding. These levels represent distal to intermediate tempestites sensu
(1985) and are interruptions in the background sedimentation, which took place in a quiet open marine environment. The environment is typical of an outer ramp setting, near the level of the normal storm wave base. Based on similar settings in the Palaeozoic of Belgium ( & ,
1995), the inferred depth of water was between 40 and 60 m.
The microfacies of the Bois de Presles Member consists of bioturbated clayey micaceous silty mudstones and wackestones with various bioclasts (bryozoans, brachiopods, ostracods, pelecypods and echinoderms) and abundant unidentified microbioclasts. The matrix is finely recrystallized into a homogeneous calcite microspar. Microscopic organic debris is abundant. In the less-burrowed sediments, bioclasts form both thin (< 1 mm) and thicker (a few millimeters) packstone-grainstone levels. The grains display parallel lamination and local cross-bedding. These levels represent distal to intermediate tempestites sensu
(1985) and are interruptions in the background sedimentation, which took place in a quiet open marine environment. The environment is typical of an outer ramp setting, near the level of the normal storm wave base. Based on similar settings in the Palaeozoic of Belgium ( & ,
1995), the inferred depth of water was between 40 and 60 m.
![]() The basal part of the Génicot Formation consists of micaceous siltstones. About 21 m above the base are two obliquely-laminated calcareous fine sandstone to siltstone beds several dm thick. Thin-section data place the depositional environment of these beds as between the storm and normal wave bases, for cross-bedding is well developed in the siltstones and the laminar structure is thicker than in the Fosses Formation.
The basal part of the Génicot Formation consists of micaceous siltstones. About 21 m above the base are two obliquely-laminated calcareous fine sandstone to siltstone beds several dm thick. Thin-section data place the depositional environment of these beds as between the storm and normal wave bases, for cross-bedding is well developed in the siltstones and the laminar structure is thicker than in the Fosses Formation.
![]() A half-meter above the upper calcareous sandstone bed is a conglomerate with macrofossils: brachiopods, trilobites,
etc. This conglomerate displays small-scale foresets, made up of subcentimetric angular and subrounded microbreccias consisting of clasts of homogeneous carbonate mudstone. These elements make the oblique bedding more obvious. The conglomerate reflects a lowering of sea level associated with an erosional episode.
A half-meter above the upper calcareous sandstone bed is a conglomerate with macrofossils: brachiopods, trilobites,
etc. This conglomerate displays small-scale foresets, made up of subcentimetric angular and subrounded microbreccias consisting of clasts of homogeneous carbonate mudstone. These elements make the oblique bedding more obvious. The conglomerate reflects a lowering of sea level associated with an erosional episode.
![]() A
5 m-thick grey micaceous siltstone intercalated with greenish-grey mudstone separates the lower from upper conglomerate. The latter is more than 2 m thick and contains limestone boulders up to 0.85 m in diameter on their major axis. Several fining upward sequences can be distinguished in this conglomerate. Solitary rugose coral heads occur in it. Thin sections of a cobble from this conglomeratic interval reveal the presence of coarse-grained rudstones with corroded and perforated bioclasts (bryozoans, 'pitted' echinoderms, molluscs and trilobites) and with dissolution and irregular vadose cavities of karstic origin. The same sample includes a subrounded microbreccia, similar in composition to the one in the lower conglomerate. Homogeneous mudstone fragments from the outer ramp occur in this microbreccia. A cobble from the basal level of the upper conglomerate contains laminar bioclastic (crinoids, ostracods) grainstone beds with abundant irregular peloids and micritized grains, a few of which enclose Girvanella (Cyanobacteria), indicative of a euphotic environment (or one close to such an environment). This 'coquina' or bioclastic level is similar to the proximal tempestites of
(1985). As the conglomerate contains pebbles, cobbles and boulders from the outer ramp, along with indicators of euphotic and karstic environments, a drop in sea level of at least 40 to 60 m can be inferred for this portion of the section.
A
5 m-thick grey micaceous siltstone intercalated with greenish-grey mudstone separates the lower from upper conglomerate. The latter is more than 2 m thick and contains limestone boulders up to 0.85 m in diameter on their major axis. Several fining upward sequences can be distinguished in this conglomerate. Solitary rugose coral heads occur in it. Thin sections of a cobble from this conglomeratic interval reveal the presence of coarse-grained rudstones with corroded and perforated bioclasts (bryozoans, 'pitted' echinoderms, molluscs and trilobites) and with dissolution and irregular vadose cavities of karstic origin. The same sample includes a subrounded microbreccia, similar in composition to the one in the lower conglomerate. Homogeneous mudstone fragments from the outer ramp occur in this microbreccia. A cobble from the basal level of the upper conglomerate contains laminar bioclastic (crinoids, ostracods) grainstone beds with abundant irregular peloids and micritized grains, a few of which enclose Girvanella (Cyanobacteria), indicative of a euphotic environment (or one close to such an environment). This 'coquina' or bioclastic level is similar to the proximal tempestites of
(1985). As the conglomerate contains pebbles, cobbles and boulders from the outer ramp, along with indicators of euphotic and karstic environments, a drop in sea level of at least 40 to 60 m can be inferred for this portion of the section.
![]() Above the conglomerates, grey micaceous siltstones and fine sandstones occur. Samples at the upper limit of the second conglomerate, from immediately above it and from a calcareous sandstone level near the top of the section are bioturbated and relatively restricted in fauna (only crinoids, mollusks and microbioclasts). Sedimentary structures in these terminal beds are similar in aspect to those of the calcareous sandstones below the upper conglomerate (cross-bedding, plane parallel laminations) and like them suggest environments between wave and storm base levels.
Above the conglomerates, grey micaceous siltstones and fine sandstones occur. Samples at the upper limit of the second conglomerate, from immediately above it and from a calcareous sandstone level near the top of the section are bioturbated and relatively restricted in fauna (only crinoids, mollusks and microbioclasts). Sedimentary structures in these terminal beds are similar in aspect to those of the calcareous sandstones below the upper conglomerate (cross-bedding, plane parallel laminations) and like them suggest environments between wave and storm base levels.
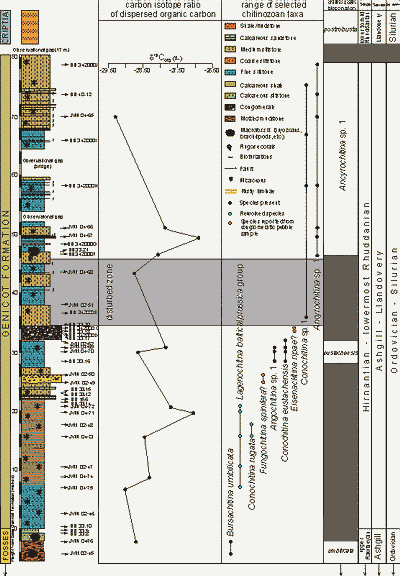
![]() The carbon isotope ratio of the Dispersed Organic Carbon (DOC) was measured in 13 samples of the Sart-Eustache section (samples with prefix JVM 04- and BB 342000F; see Fig. 1
The carbon isotope ratio of the Dispersed Organic Carbon (DOC) was measured in 13 samples of the Sart-Eustache section (samples with prefix JVM 04- and BB 342000F; see Fig. 1 ![]() ). The methodology used is described in et alii
(2004). Samples were dried and their carbonate content was measured. They were then carefully cleaned in the laboratory, to remove surface oxidation and exclude potential sources of degraded organic matter and fossil carbonates, if any. After powdering, the samples were treated with 10% HCl for 20 min. to 2 h to remove the remaining carbonates. The samples without carbonate underwent the same treatment. Soluble salts were removed by repetitive centrifuging (7000 revolutions per second) until a neutral solution was obtained. Finally, the residue was dried at 90°C and re-powdered. Fractions of the resulting powder were measured with a standard LECO carbon analyzer (CS-200) to determine total organic carbon (TOC) and SO3 contents. Quantities required for analysis were calculated on the basis of the TOC values.
). The methodology used is described in et alii
(2004). Samples were dried and their carbonate content was measured. They were then carefully cleaned in the laboratory, to remove surface oxidation and exclude potential sources of degraded organic matter and fossil carbonates, if any. After powdering, the samples were treated with 10% HCl for 20 min. to 2 h to remove the remaining carbonates. The samples without carbonate underwent the same treatment. Soluble salts were removed by repetitive centrifuging (7000 revolutions per second) until a neutral solution was obtained. Finally, the residue was dried at 90°C and re-powdered. Fractions of the resulting powder were measured with a standard LECO carbon analyzer (CS-200) to determine total organic carbon (TOC) and SO3 contents. Quantities required for analysis were calculated on the basis of the TOC values.
![]() Isotopic measurements were carried out on bulk samples with an Optima-isochrome system at the Royal Holloway University of London (U.K.). For each sample, 20 to 29 milligram was loaded into a purified tin cup. The dropped samples were flash combusted at 1800°C under O2 atmosphere. The gases released were oxidised with O2 and the catalysts, and then transported by He flow to the mass spectrometer to be analysed as CO2 gas. The signals corresponding to masses 44, 45, and 46 were measured, and isotopic compositions were calculated from them and compared to an isotopic international standard (NBS-21: -28.16‰) and a laboratory standard (GF: -23.95‰). Organic 13C values are reported as a proportion of 12C in δ13C notation normalized to the international PDB standard (V-PDB, Vienna Peedee belemnite). Calculated reproducibility is within 0.1‰.
Isotopic measurements were carried out on bulk samples with an Optima-isochrome system at the Royal Holloway University of London (U.K.). For each sample, 20 to 29 milligram was loaded into a purified tin cup. The dropped samples were flash combusted at 1800°C under O2 atmosphere. The gases released were oxidised with O2 and the catalysts, and then transported by He flow to the mass spectrometer to be analysed as CO2 gas. The signals corresponding to masses 44, 45, and 46 were measured, and isotopic compositions were calculated from them and compared to an isotopic international standard (NBS-21: -28.16‰) and a laboratory standard (GF: -23.95‰). Organic 13C values are reported as a proportion of 12C in δ13C notation normalized to the international PDB standard (V-PDB, Vienna Peedee belemnite). Calculated reproducibility is within 0.1‰.
![]() Thirteen samples were analysed, more analyses are in progress. The δ13Corg values
measured until now range from -29.3‰ to -26.9‰ (Fig. 1
Thirteen samples were analysed, more analyses are in progress. The δ13Corg values
measured until now range from -29.3‰ to -26.9‰ (Fig. 1 ![]() ). The top of the Fosses Formation and the lower part of the Génicot Formation show relatively low values (-29.0‰ to -28.3‰). Below the calcareous sandstone levels, δ13C values increase to -27.0‰ in JVM 04-71. Between the conglomerates, a lower sample (JVM 04-70) shows a relatively low value (-28.6‰), whereas a higher sample (JVM 04-69) shows a relatively high value (-27.9‰). Above the conglomerates, δ13C values
appear to increase again from -28.7‰ in JVM 04-68 to -26.9‰ in JVM 04-67. Above this sample, values decrease again to -26.9‰ in JVM 04-65, near the top of the section.
). The top of the Fosses Formation and the lower part of the Génicot Formation show relatively low values (-29.0‰ to -28.3‰). Below the calcareous sandstone levels, δ13C values increase to -27.0‰ in JVM 04-71. Between the conglomerates, a lower sample (JVM 04-70) shows a relatively low value (-28.6‰), whereas a higher sample (JVM 04-69) shows a relatively high value (-27.9‰). Above the conglomerates, δ13C values
appear to increase again from -28.7‰ in JVM 04-68 to -26.9‰ in JVM 04-67. Above this sample, values decrease again to -26.9‰ in JVM 04-65, near the top of the section.
![]() In addition to the samples of &
(2004), chitinozoans from 14 new samples (1 from the Fosses and 13 from the Génicot Formation, identified as JVM 04-xx, BB 342000F and JVM 02-62 were studied, see Fig. 1
In addition to the samples of &
(2004), chitinozoans from 14 new samples (1 from the Fosses and 13 from the Génicot Formation, identified as JVM 04-xx, BB 342000F and JVM 02-62 were studied, see Fig. 1 ![]() . They aided in setting more precise limits to the biozones recognised in &
(2004).
. They aided in setting more precise limits to the biozones recognised in &
(2004).
![]() In the upper part of the Fosses Formation, the Bursachitina umbilicata et alii
(in press) chitinozoan biozone is recognised up to JVM 04-76. This zone is defined in the type Ashgill area, (Cautley District, England, Avalonia) and corresponds to the late Rawtheyan ( et alii,
in press). As the Génicot Formation overlies the Fosses Formation, we assume it to be post-Rawtheyan in age.
In the upper part of the Fosses Formation, the Bursachitina umbilicata et alii
(in press) chitinozoan biozone is recognised up to JVM 04-76. This zone is defined in the type Ashgill area, (Cautley District, England, Avalonia) and corresponds to the late Rawtheyan ( et alii,
in press). As the Génicot Formation overlies the Fosses Formation, we assume it to be post-Rawtheyan in age.
![]() The Génicot Formation yields poorly preserved chitinozoan assemblages. Below the conglomerates, most chitinozoan determinations are generic. Noteworthy is the presence of a few fragments of Conochitina rugata nomen nudum and of the Lagenochitina baltica-prussica group. These chitinozoans are characteristic in the Pusgillian-mid-Rawtheyan interval of the type Ashgill area, beneath the Bursachitina umbilicata biozone ( et alii,
in press). Their presence here, poorly preserved and few in number, indicates that some part of the Fosses Formation was reworked.
The Génicot Formation yields poorly preserved chitinozoan assemblages. Below the conglomerates, most chitinozoan determinations are generic. Noteworthy is the presence of a few fragments of Conochitina rugata nomen nudum and of the Lagenochitina baltica-prussica group. These chitinozoans are characteristic in the Pusgillian-mid-Rawtheyan interval of the type Ashgill area, beneath the Bursachitina umbilicata biozone ( et alii,
in press). Their presence here, poorly preserved and few in number, indicates that some part of the Fosses Formation was reworked.
![]() Between the conglomerates, two characteristic species occur: Conochitina eustachensis &
2004 and Angochitina sp. 1. Unfortunately, these species cannot be correlated with chitinozoan species found elsewhere and may be endemic.
Between the conglomerates, two characteristic species occur: Conochitina eustachensis &
2004 and Angochitina sp. 1. Unfortunately, these species cannot be correlated with chitinozoan species found elsewhere and may be endemic.
![]() The 10 m interval immediately above the conglomerates is disturbed, as a result of slumping or tectonic activity and contain no identifiable or characteristic species. From BB 342000F up to the uppermost samples of the Génicot Formation, however, almost all samples yield Ancyrochitina sp. 1. Some of the specimens are very close to Ancyrochitina ellisbayensis &
2000, reported from the late Hirnantian Upper Laframboise Member of the Ellis Bay Formation (Anticosti Island, Canada, Laurentia) ( & ,
2000). It is most probable that this species occurs also in the upper part of the upper Hirnantian N. persculptus Zone and the lowermost Silurian praematurus Zone (lower subzone of the ascensus
Zone)
(, pers. comm.).
The 10 m interval immediately above the conglomerates is disturbed, as a result of slumping or tectonic activity and contain no identifiable or characteristic species. From BB 342000F up to the uppermost samples of the Génicot Formation, however, almost all samples yield Ancyrochitina sp. 1. Some of the specimens are very close to Ancyrochitina ellisbayensis &
2000, reported from the late Hirnantian Upper Laframboise Member of the Ellis Bay Formation (Anticosti Island, Canada, Laurentia) ( & ,
2000). It is most probable that this species occurs also in the upper part of the upper Hirnantian N. persculptus Zone and the lowermost Silurian praematurus Zone (lower subzone of the ascensus
Zone)
(, pers. comm.).
![]() Chitinozoan assemblages from the overlying Criptia Group contain Belonechitina postrobusta
1980 and Conochitina electa
1980 and can be correlated with the Cystograptus vesiculosus and Monograptus cyphus graptolite zones, mid to late Rhuddanian (early Llandovery) ( et alii,
2002).
Chitinozoan assemblages from the overlying Criptia Group contain Belonechitina postrobusta
1980 and Conochitina electa
1980 and can be correlated with the Cystograptus vesiculosus and Monograptus cyphus graptolite zones, mid to late Rhuddanian (early Llandovery) ( et alii,
2002).
![]() The sediments of the Fosses Formation represent distal to intermediate tempestites sensu
(1985), and demonstrate a disturbance of the background
sedimentation which took place in a quiet, open marine environment. The environment of the succession is that typical of an outer ramp setting (40-60 m depth), near the normal storm wave base level. The lower calcareous sandstone
beds were deposited in a locality between the fair-weather and the storm-wave base, so indicate a lowering in sea level. This conclusion is corroborated by the presence of reworked chitinozoans below the conglomerates of the Génicot Formation. The lower conglomerate itself reflects a marked fall in sea level associated with an erosional episode. The upper conglomerate contains clasts of outer ramp, euphotic and karstic environments, indicating a fall in sea level of at least 40 to 60m. Above the conglomerates, deposition in the area was restored to that of an outer ramp sited between fair-weather and storm wave bases.
The sediments of the Fosses Formation represent distal to intermediate tempestites sensu
(1985), and demonstrate a disturbance of the background
sedimentation which took place in a quiet, open marine environment. The environment of the succession is that typical of an outer ramp setting (40-60 m depth), near the normal storm wave base level. The lower calcareous sandstone
beds were deposited in a locality between the fair-weather and the storm-wave base, so indicate a lowering in sea level. This conclusion is corroborated by the presence of reworked chitinozoans below the conglomerates of the Génicot Formation. The lower conglomerate itself reflects a marked fall in sea level associated with an erosional episode. The upper conglomerate contains clasts of outer ramp, euphotic and karstic environments, indicating a fall in sea level of at least 40 to 60m. Above the conglomerates, deposition in the area was restored to that of an outer ramp sited between fair-weather and storm wave bases.
![]() Apparently, elevated δ13C values occur in parts of the Génicot Formation. These may represent the positive δ13C shift, recorded worldwide and associated with the Hirnantian glaciation and large ice caps on Gondwana ( et alii,
2003). In the δ13Corg values reported from Arctic Canada (Laurentia; et alii,
2003), two distinct peaks can be seen in the interval of positive excursion. These may be coincident with pulses of peak glaciation (cf. et alii,
2000). Two peaks seem also to be present in our data: one below the calcareous sandstone and the lower conglomerate; the other above the upper conglomerate. These peaks are defined on but a small number of samples so their interpretation as coincident events is highly tentative. New analyses are in progress
Apparently, elevated δ13C values occur in parts of the Génicot Formation. These may represent the positive δ13C shift, recorded worldwide and associated with the Hirnantian glaciation and large ice caps on Gondwana ( et alii,
2003). In the δ13Corg values reported from Arctic Canada (Laurentia; et alii,
2003), two distinct peaks can be seen in the interval of positive excursion. These may be coincident with pulses of peak glaciation (cf. et alii,
2000). Two peaks seem also to be present in our data: one below the calcareous sandstone and the lower conglomerate; the other above the upper conglomerate. These peaks are defined on but a small number of samples so their interpretation as coincident events is highly tentative. New analyses are in progress
![]() Stratigraphically,
the Génicot Formation is situated between the Fosses Formation and the Criptia Group. The youngest known sediments of the Fosses Formation are dated late Rawtheyan on the basis of the Bursachitina umbilicata biozone in the type Ashgill area. Some chitinozoans from the upper part of the Génicot Formation resemble closely A. ellisbayensis from the upper Hirnantian Laframboise Member of Anticosti Island, Canada. The oldest sediments dated by chitinozoans of the Silurian Criptia Group are of mid Rhudannian age, because of the presence of Belonechitina postrobusta. Worldwide, the early Silurian is trangressive and generally is associated with the deposition of black shales. The lithology of the Génicot Formation is markedly different from that of these shales for as a whole it is far coarser. Consequently, we think that the Génicot Formation is of Hirnantian - lowermost Silurian age and that its sediments reflect the drop in sea-level associated with the glaciation of that time.
Stratigraphically,
the Génicot Formation is situated between the Fosses Formation and the Criptia Group. The youngest known sediments of the Fosses Formation are dated late Rawtheyan on the basis of the Bursachitina umbilicata biozone in the type Ashgill area. Some chitinozoans from the upper part of the Génicot Formation resemble closely A. ellisbayensis from the upper Hirnantian Laframboise Member of Anticosti Island, Canada. The oldest sediments dated by chitinozoans of the Silurian Criptia Group are of mid Rhudannian age, because of the presence of Belonechitina postrobusta. Worldwide, the early Silurian is trangressive and generally is associated with the deposition of black shales. The lithology of the Génicot Formation is markedly different from that of these shales for as a whole it is far coarser. Consequently, we think that the Génicot Formation is of Hirnantian - lowermost Silurian age and that its sediments reflect the drop in sea-level associated with the glaciation of that time.
![]() The authors would like to acknowledge the reviewers for their useful comments and careful reading. We are also grateful to A. for language advice. Thanks to S. for the preparation of the palynological samples, Baron d' for permission to work on his property, T. for chitinozoan discussions, and A. and O. for help on the field. The first author is supported by the BOF of the Ghent University. This work is a contribution to the IGCP 503 project.
The authors would like to acknowledge the reviewers for their useful comments and careful reading. We are also grateful to A. for language advice. Thanks to S. for the preparation of the palynological samples, Baron d' for permission to work on his property, T. for chitinozoan discussions, and A. and O. for help on the field. The first author is supported by the BOF of the Ghent University. This work is a contribution to the IGCP 503 project.
![]()
T. (1985).- Storm depositional systems.- In: G.M., H.J. & A. (eds.), Lecture notes in Earth Sciences.- Springer Verlag, Berlin, p. 1-174.
P.J., J.D., G.A.F., D.B.R., D.G.F., T., L. & T. (1994). Bathymetric and isotopic evidence for a short-lived Late Ordovician glaciation in a greenhouse period.- Geology, Boulder, vol. 22, p. 295-298.
P.J., G.A., L. D., J.D., T., T. & J. (2003).- High-resolution stable isotope stratigraphy of Upper Ordovician sequences: constraints on timing of bioevents and environmental changes associated with mass extinction and glaciation.- Geological Society of America, Bulletin, Boulder, vol. 115, p. 89-104.
A., J., T., B., S. & M. (2002).- The Lower Palaeozoic stratigraphy and sedimentology of the Brabant Massif in the Dyle and Orneau valleys and of the Condroz Inlier at Fosses: an excursion guidebook.- Geologica Belgica, Liège, vol. 5, n° 3-4, p. 71-143.
R., C., T., E. & P.D. (2004).- Paleocene-Eocene carbon isotope excursion in organic carbon and pedogenic carbonate: direct comparison in a continental stratigraphic section.- Geology, Boulder, vol. 32, p. 553-556.
F. (1968).- Les acritarches de l'Ordovicien et du Silurien belge. Détermination et valeur stratigraphique.- Mémoires de l'Institut Royal des Sciences naturelles de Belgique, Bruxelles, vol. 160, p. 1-176.
M.J., C. & S.H. (2003).- Correlation of graptolite biozones, chitinozoan biozones and carbon isotope curves through the Hirnantian.- In: G.L., M.S. & S.H. (eds.): Ordovician from the Andes.- Proceedings of the 9th International Symposium on the Ordovician System, INSUGEO, Serie de Correlación Geológica, Tucumán, 17, p. 101-104.
P. (1934).- La stratigraphie du Silurien de la bande Sambre-et-Meuse.- Mémoires de l'Académie Royale de Belgique, Bruxelles, Classe des Sciences, 2ème série, vol. 13, n° 2, p. 1-408.
V. (1980).- Novye vidy khitinozoy iz nichnego Llandovery Estonii (New Chitinozoan species from the Lower Llandoverian of Estonia).- Eesti NSV Teaduste Akadeemia Toimetised, Geoloogia, Tallin, vol. 29, n° 3, p. 98-107.
A. & R. (1995).- Sédimentation de rampe mixte silico-carbonatée des couches de transition eiféliennes-givétiennes franco-belges. Première partie : microfaciès et modèle sédimentaire.- Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine, Pau, vol. 19, n° 2, p. 329-375
A. & A. (2000).- Chitinozoan zonation of the Late Ordovician and the Early Silurian of the Island of Anticosti, Québec, Canada.- Review of Palaeobotany and Palynology, Amsterdam, vol. 109, n° 2, p. 85-111.
O.E., J.A., R.J., J.N. & J. (2000).- Calibrating the Late Ordovician glaciation and mass extinction by the eccentricity cycles of Earth's orbit.- Geology, Boulder, vol. 28, p. 967-970.
T., B. & J. (in press).- Upper Ordovician Chitinozoan biostratigraphy from the Type Ashgill Area (Cautley district) and the Pus Gill section (Dufton district, Cross Fell Inlier), Cumbria, Northern England.- Geological Magazine, Cambridge.
J. & J. (2004).- Chitinozoan bio-and lithostratigraphical study of the Ashgill Fosses and Génicot Formations (Condroz Inlier, Belgium).- Review of Palaeobotany and Palynology, Amsterdam, vol. 130, n° 1-4, p. 241-267.
![]()
Click on thumbnail to enlarge the image.
Figure 1: δ13Corg isotope curve and chitinozoan distribution in the Parc de Sart-Eustache section. Chitinozoan biozonation and age assignment are figured on the right.
![]()