![]()
![]()
B. (2005).- Correlation and biostratigraphy of the Kortrijk (Sint-Antonius) and Kortrijk (Lust) boreholes (early Silurian, Belgium). In: P. & E. (eds.), Pre-Cambrian to Palaeozoic Palaeopalynology and Palaeobotany.- Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2005/02, Abstract 14 (CG2005_M02/14)
![]() Acritarch; Belgium; biostratigraphy; Llandovery; Silurian.
Acritarch; Belgium; biostratigraphy; Llandovery; Silurian.
![]() Acritarche ; Belgique ; biostratigraphie ; Llandoverien ; Silurien.
Acritarche ; Belgique ; biostratigraphie ; Llandoverien ; Silurien.
![]()
![]() Since their drilling, respectively in 1961 and 1971, the 83W421 Kortrijk (Sint-Antonius) and 83W44 Kortrijk (Lust brewery) boreholes have been studied rather extensively for biostratigraphical information. Three fossil groups are included in these studies: graptolites, chitinozoans and acritarchs.
Since their drilling, respectively in 1961 and 1971, the 83W421 Kortrijk (Sint-Antonius) and 83W44 Kortrijk (Lust brewery) boreholes have been studied rather extensively for biostratigraphical information. Three fossil groups are included in these studies: graptolites, chitinozoans and acritarchs.
![]() (1962, 1981) made the first studies of the
graptolites;
(1990) investigated the chitinozoans; et alii
(1998) revised the graptolites and provided biostratigraphical data on both graptolites and
chitinizoans. Acritarchs have been studied over the years, first by &
(1963), and thereafter by
(1966, 1969),
(1997), &
(1999), (2003) and
(in press). This contribution, based on the data now
available, discusses in chronological order possible correlations between the boreholes
and it reviews the biostratigraphies of graptolites, chitinozoans and acritarchs
as they are understood currently.
(1962, 1981) made the first studies of the
graptolites;
(1990) investigated the chitinozoans; et alii
(1998) revised the graptolites and provided biostratigraphical data on both graptolites and
chitinizoans. Acritarchs have been studied over the years, first by &
(1963), and thereafter by
(1966, 1969),
(1997), &
(1999), (2003) and
(in press). This contribution, based on the data now
available, discusses in chronological order possible correlations between the boreholes
and it reviews the biostratigraphies of graptolites, chitinozoans and acritarchs
as they are understood currently.
![]() (1962, 1981) did not propose a formal correlation between the Kortrijk (Sint-Antonius) and Kortrijk (Lust) boreholes although he states that
(1981, translation): "[
] there is a perfect palaeontological tie between both boreholes". He recognises three, possibly four, of the biozones of &
(1913) in Kortrijk (Sint-Antonius) and two, possibly three, of them in Kortrijk (Lust).
(1962, 1981) did not propose a formal correlation between the Kortrijk (Sint-Antonius) and Kortrijk (Lust) boreholes although he states that
(1981, translation): "[
] there is a perfect palaeontological tie between both boreholes". He recognises three, possibly four, of the biozones of &
(1913) in Kortrijk (Sint-Antonius) and two, possibly three, of them in Kortrijk (Lust).
![]() He gives fairly detailed graptolite distributions in both boreholes. It appears that no single species can be used to correlate the two sections (see Fig. 1
He gives fairly detailed graptolite distributions in both boreholes. It appears that no single species can be used to correlate the two sections (see Fig. 1 ![]() ). So the only correlation possible would
of necessity be based on the
boundary between the sedgwickii and the turriculatus graptolite biozones.
). So the only correlation possible would
of necessity be based on the
boundary between the sedgwickii and the turriculatus graptolite biozones.
![]() In Kortrijk (Lust), this
boundary lies in a barren interval spanning 8.00 m. In Kortrijk (Sint-Antonius),
(1981) places it at -205.00 m. However, a close inspection of graptolite distribution
in this borehole show that a 5.50 m interval exists between the disappearance of M. sedgwickii at
-207.40 m and the first appearance of M. runcinatus at -201.90 m (see data in ,
1981). Thus a correlation of the two boreholes based on
the graptolite ranges reported by
(1962, 1981) involves an error
of as much as 8.00 m.
In Kortrijk (Lust), this
boundary lies in a barren interval spanning 8.00 m. In Kortrijk (Sint-Antonius),
(1981) places it at -205.00 m. However, a close inspection of graptolite distribution
in this borehole show that a 5.50 m interval exists between the disappearance of M. sedgwickii at
-207.40 m and the first appearance of M. runcinatus at -201.90 m (see data in ,
1981). Thus a correlation of the two boreholes based on
the graptolite ranges reported by
(1962, 1981) involves an error
of as much as 8.00 m.
![]() In his unpublished
Ph.D. thesis, (1990) used the
base of the turriculatus graptolite Biozone to correlate the two boreholes. Thus, he
equates level -205.00 m in Kortrijk (Sint-Antonius) with level -190.80 m in Kortrijk (Lust).
In his unpublished
Ph.D. thesis, (1990) used the
base of the turriculatus graptolite Biozone to correlate the two boreholes. Thus, he
equates level -205.00 m in Kortrijk (Sint-Antonius) with level -190.80 m in Kortrijk (Lust).
![]() He defines some local biozones in these boreholes. However, his data reveal that no biozone defined in Kortrijk (Sint-Antonius) can be recognized in Kortrijk (Lust) and vice-versa. Thus it appears that this study can neither confirm the chosen correlation based on the appearance of the turriculatus graptolite Biozone, nor propose a discrete correlation based on chitinozoans.
He defines some local biozones in these boreholes. However, his data reveal that no biozone defined in Kortrijk (Sint-Antonius) can be recognized in Kortrijk (Lust) and vice-versa. Thus it appears that this study can neither confirm the chosen correlation based on the appearance of the turriculatus graptolite Biozone, nor propose a discrete correlation based on chitinozoans.
![]() et alii
(1998) revise the graptolites and relate them to 's
(1992, 1993) biozones and subzones. Chitinozoans too are revised and related to the global chitinozoan biozonation of
et alii (1995).
et alii
(1998) revise the graptolites and relate them to 's
(1992, 1993) biozones and subzones. Chitinozoans too are revised and related to the global chitinozoan biozonation of
et alii (1995).
![]() Below the base of the turriculatus s.l. (?
guerichi) Biozone in Kortrijk (Sint-Antonius), determinable graptolites
are absent in a large interval: -220.00 to -250.00 m ( et alii,
1998). In Kortrijk (Lust), a definite guerichi Biozone is present from
-203.50 to -159.00 m and some subzones can be recognised ( et alii,
1998). But as the Kortrijk (Lust) borehole is devoid of graptolites from
-225.50 to -203.50 m, no reasonable correlation can be proposed using this revision of the graptolites.
Below the base of the turriculatus s.l. (?
guerichi) Biozone in Kortrijk (Sint-Antonius), determinable graptolites
are absent in a large interval: -220.00 to -250.00 m ( et alii,
1998). In Kortrijk (Lust), a definite guerichi Biozone is present from
-203.50 to -159.00 m and some subzones can be recognised ( et alii,
1998). But as the Kortrijk (Lust) borehole is devoid of graptolites from
-225.50 to -203.50 m, no reasonable correlation can be proposed using this revision of the graptolites.
![]() As
regards chitinozoans, only the global Eisenackitina dolioliformis Biozone can be recognised in the upper part of the Kortrijk (Sint-Antonius) borehole.
E. dolioliformis does indeed appears at -240.10 m. Unfortunately, et alii
(1998) are not clear about chitinozoan biozonation between
-265.30 and -240.10 m and do not refer to or discuss the limit between the alargada and dolioliformis
biozones.
As
regards chitinozoans, only the global Eisenackitina dolioliformis Biozone can be recognised in the upper part of the Kortrijk (Sint-Antonius) borehole.
E. dolioliformis does indeed appears at -240.10 m. Unfortunately, et alii
(1998) are not clear about chitinozoan biozonation between
-265.30 and -240.10 m and do not refer to or discuss the limit between the alargada and dolioliformis
biozones.
![]() The dolioliformis Biozone is recognised in Kortrijk (Lust) from
-215.50 to -148.30 m ( et alii,
1998), i.e. it occupies all of the borehole
with the exception of the lowermost 10 m (-225.50 to -215.50 m). Those ten meters
yielded only one barren sample and another with no indicative chitinozoans (,
1990).
The dolioliformis Biozone is recognised in Kortrijk (Lust) from
-215.50 to -148.30 m ( et alii,
1998), i.e. it occupies all of the borehole
with the exception of the lowermost 10 m (-225.50 to -215.50 m). Those ten meters
yielded only one barren sample and another with no indicative chitinozoans (,
1990).
![]() Thus, the revisions of et alii
(1998) do not permit any correlation of the two boreholes, mainly because of the presence of intervals either barren or with no meaningful biota.
Thus, the revisions of et alii
(1998) do not permit any correlation of the two boreholes, mainly because of the presence of intervals either barren or with no meaningful biota.
![]() An early palynological investigation of Kortrijk (Sint-Antonius) by the author revealed that acritarchs in samples below -150 m are too mature thermally for convenient study with the notable exception of levels at
-260.50 m, -267.20 m and -285.00 m where stratigraphically interesting species were recognised (,
1997). These are Elektoriskos williereae, Tylotopalla robustispinosa, Multiplicisphaeridium fisherii and Tunisphaeridium tentaculaferum.
An early palynological investigation of Kortrijk (Sint-Antonius) by the author revealed that acritarchs in samples below -150 m are too mature thermally for convenient study with the notable exception of levels at
-260.50 m, -267.20 m and -285.00 m where stratigraphically interesting species were recognised (,
1997). These are Elektoriskos williereae, Tylotopalla robustispinosa, Multiplicisphaeridium fisherii and Tunisphaeridium tentaculaferum.
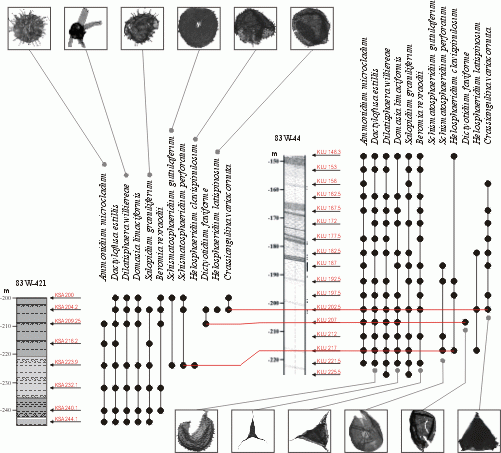
![]() Above
-150.00 m, diversity is high with a minimum of 26 species, a mean of 65 species and a maximum of 89 species. Inspection of the stratigraphic ranges of acritarchs reveals four species with a potential for the correlation of the two boreholes (see Fig. 2
Above
-150.00 m, diversity is high with a minimum of 26 species, a mean of 65 species and a maximum of 89 species. Inspection of the stratigraphic ranges of acritarchs reveals four species with a potential for the correlation of the two boreholes (see Fig. 2 ![]() and ,
in press). These species are Crassiangulina variacornuta, Dictyotidium faviforme, Helosphaeridium clavispinulosum and Schismatosphaeridium perforatum.
and ,
in press). These species are Crassiangulina variacornuta, Dictyotidium faviforme, Helosphaeridium clavispinulosum and Schismatosphaeridium perforatum.
Crassiangulina variacornuta is known from Gondwana and Balonia. It does not appear prior to the Telychian on the Balonian palaeocontinent ( et alii, 2003). New information from Argentina indicate an early appearance of this species in Gondwana during late Aeronian times ( & , in press).
Dictyotidium faviforme is known from Balonia and Laurentia where it ranges from the upper Llandovery to the Ludlow (, 1967; & , 1972; , 1989; , 2001).
Helosphaeridium clavispinulosum is known from Balonia and Gondwana. It appears first in the Aeronian (convolutus graptolite Biozone) and ends in the Ludlow (, 1970; , 1987; , 2001)
Schismatosphaeridium perforatum is recorded on Balonia and Laurentia where it seems to be restricted to the upper Llandovery to Wenlock ( et alii, 1965; & , 1972; , 1982; & , 1982; , 1987; , 1989). One specimen tentatively referred to this species is recorded in the basal part of the Hemse Formation, of the lower Ludlow of Gotland (, 1989).
![]() Thus it appears that a correlation based on the first appearance of
Helosphaeridium clavispinulosum would be hazardous because this species appears in Balonia earlier than its first occurrence in the boreholes.
Dictyotidium faviforme was not recorded frequently enough to provide a reliable basis for correlation. In
(in press), Crassiangulina variacornuta is preferred to
Schismatosphaeridium perforatum because it is recorded more consistently than the latter in the boreholes. Moreover the First Appearence Datum of
Crassiangulina variacornuta is better assessed in Balonia than that of Schismatosphaeridium perforatum. Consequently, based on the first occurrences of
Crassiangulina variacornuta.
(in press) correlates level -204.20 m in Kortrijk (Sint-Antonius) with level
-202.50 in Kortrijk (Lust).
Thus it appears that a correlation based on the first appearance of
Helosphaeridium clavispinulosum would be hazardous because this species appears in Balonia earlier than its first occurrence in the boreholes.
Dictyotidium faviforme was not recorded frequently enough to provide a reliable basis for correlation. In
(in press), Crassiangulina variacornuta is preferred to
Schismatosphaeridium perforatum because it is recorded more consistently than the latter in the boreholes. Moreover the First Appearence Datum of
Crassiangulina variacornuta is better assessed in Balonia than that of Schismatosphaeridium perforatum. Consequently, based on the first occurrences of
Crassiangulina variacornuta.
(in press) correlates level -204.20 m in Kortrijk (Sint-Antonius) with level
-202.50 in Kortrijk (Lust).
![]() Tunisphaeridium tentaculaferum is present from sample
-285.00 on. This species appears in zone 2 of ( & ,
1984), i.e. the Oppilatala eoplanktonica Biozone ( & ,
1987). The Ammonidium microcladum Biozone was not recognised although it should be present for it is roughly equivalent to the sedgwickii graptolite Biozone ( et alii,
1996; et alii,
1997). The Dactylofusa estillis Biozone is recognised from
-244.10 m upward in Kortrijk (Sint-Antonius). Its occurrence in levels attributed to the guerichi graptolite Biozone confirms its extension in time into strata of Telychian age, contrary to the findings of et alii
(1996). (in press) proposes the subdivision of the D. estillis Biozone into two subzones, the upper D. estillis Subzone being the concurrent range zone between D.
estillis and Crassiangulina variacornuta. This upper Dactylofusa estilis
Subzone is also recognised in the 50E134 Steenkerke borehole in levels attributed to the crispus graptolite Biozone (,
2003). This is consistent with the findings of et alii
(1997).
Tunisphaeridium tentaculaferum is present from sample
-285.00 on. This species appears in zone 2 of ( & ,
1984), i.e. the Oppilatala eoplanktonica Biozone ( & ,
1987). The Ammonidium microcladum Biozone was not recognised although it should be present for it is roughly equivalent to the sedgwickii graptolite Biozone ( et alii,
1996; et alii,
1997). The Dactylofusa estillis Biozone is recognised from
-244.10 m upward in Kortrijk (Sint-Antonius). Its occurrence in levels attributed to the guerichi graptolite Biozone confirms its extension in time into strata of Telychian age, contrary to the findings of et alii
(1996). (in press) proposes the subdivision of the D. estillis Biozone into two subzones, the upper D. estillis Subzone being the concurrent range zone between D.
estillis and Crassiangulina variacornuta. This upper Dactylofusa estilis
Subzone is also recognised in the 50E134 Steenkerke borehole in levels attributed to the crispus graptolite Biozone (,
2003). This is consistent with the findings of et alii
(1997).
![]() Early work on the graptolites (,
1962, 1981) of the Kortrijk (Sint-Antonius) and Kortrijk (Lust) wells suggested a possible correlation with an imprecision of
8.00 m. This correlation is founded on the limit between the sedgwickii and the turriculatus graptolite
biozones of &
(1913). Because of the revision of graptolite taxonomy and distributions together with the application of 's
(1992, 1993) biozonation scheme, this correlation is no longer possible, principally because many long intervals are barren.
Early work on the graptolites (,
1962, 1981) of the Kortrijk (Sint-Antonius) and Kortrijk (Lust) wells suggested a possible correlation with an imprecision of
8.00 m. This correlation is founded on the limit between the sedgwickii and the turriculatus graptolite
biozones of &
(1913). Because of the revision of graptolite taxonomy and distributions together with the application of 's
(1992, 1993) biozonation scheme, this correlation is no longer possible, principally because many long intervals are barren.
![]() Our current knowledge of chitinozoan distribution does not provide a means of correlation between the two boreholes. The local biozonation proposed by (,
1990) does not appear to be applicable between the
boreholes although the two are near each other (~600 m). Global chitinozoan biozones ( et alii,
1995) cannot be used for the correlatable portions of
both boreholes are in the dolioliformis Biozone.
Our current knowledge of chitinozoan distribution does not provide a means of correlation between the two boreholes. The local biozonation proposed by (,
1990) does not appear to be applicable between the
boreholes although the two are near each other (~600 m). Global chitinozoan biozones ( et alii,
1995) cannot be used for the correlatable portions of
both boreholes are in the dolioliformis Biozone.
![]() Acritarchs provide a sounder basis for correlation. Among four species potentially useful for this purpose, Crassiangulina variacornuta is retained because of its well-defined FAD, potential world-wide recognition, and consistency of occurrence ( et alii,
2003; , in press). The imprecision of this correlation is represented by the largest interval between the
uppermost sample with C. variacornuta and the lowest one lacking it. The
gap is 5.05 m in Kortrijk (Sint-Antonius) and is 4.50 m in Kortrijk (Lust). Therefore the maximum
possible error in correlation is 5.05 m.
Acritarchs provide a sounder basis for correlation. Among four species potentially useful for this purpose, Crassiangulina variacornuta is retained because of its well-defined FAD, potential world-wide recognition, and consistency of occurrence ( et alii,
2003; , in press). The imprecision of this correlation is represented by the largest interval between the
uppermost sample with C. variacornuta and the lowest one lacking it. The
gap is 5.05 m in Kortrijk (Sint-Antonius) and is 4.50 m in Kortrijk (Lust). Therefore the maximum
possible error in correlation is 5.05 m.
![]() The correlation proposed by
(in press) coincides precisely with the lithologic pattern and is geometrically coherent. So this correlation is well-supported and we can confidently present an integrated biostratigraphical scheme for graptolites, chitinozoans and acritarchs
covering the whole of the sequences present in the two boreholes (Fig. 3
The correlation proposed by
(in press) coincides precisely with the lithologic pattern and is geometrically coherent. So this correlation is well-supported and we can confidently present an integrated biostratigraphical scheme for graptolites, chitinozoans and acritarchs
covering the whole of the sequences present in the two boreholes (Fig. 3 ![]() ).
).
![]() Samples from the Kortrijk boreholes were collected by M. and B. in 1996. The director of the Geological survey of Belgium kindly provided permission to study this material and Walter guided M. and B. through the core library. Samples were processed by B. . The use of facilities in the Laboratoire de Paléobotanique, Paléopalynologie et Micropaléontologie at the Univesité de Liège is gratefully acknowledged. This work was partly supported by a
Ph.D. grant from the FRIA.
Samples from the Kortrijk boreholes were collected by M. and B. in 1996. The director of the Geological survey of Belgium kindly provided permission to study this material and Walter guided M. and B. through the core library. Samples were processed by B. . The use of facilities in the Laboratoire de Paléobotanique, Paléopalynologie et Micropaléontologie at the Univesité de Liège is gratefully acknowledged. This work was partly supported by a
Ph.D. grant from the FRIA.
![]()
F.H. & M.D.C.R. (1972).- North American Silurian palynofacies and their spatial arrangement: Acritarchs.- Palaeontographica, Abteilung B, Stuttgart, vol. 138, n° 5-6, p. 107-180.
J.R., C.J.N., R.A., D., D.G. & J.A. (1997).- Geology of the country around Llanilar and Rhayader.- Memoir for 1:50 000 geological sheets 178 and 179 (England & Wales), British Geological Survey, London, 267 p.
K.J. (1982).- Early Wenlock acritarchs from the Knockgardner and Straiton Grit formations of Knockgardner, Ayrshire.- Scottish Journal of Geology, Edinburgh, vol. 18, n° 4, p. 267-273.
K.J. & R.J. (1982).- A preliminary investigation of palynological assemblages from the early Silurian of Ringerike.- Contributions from the University of Oslo, n° 278, p. 105-108.
K.J. & D.G. (1987).- The Silurian carbonate shelf microflora: acritarch distribution in the Much Wenlock Limestone Formation. In: M.B. (ed.), Micropalaeontology of Carbonate Environments.- Ellis Horwood, Chichester, p. 266-287.
G.L. & E.M.R. (1913).- A monograph of British graptolites.- Palaeontographical Society Monographs, London, vol. 171, 539 p.
P.J. & K.J. (1984).- The Llandovery Series of the type area. Appendix I. Acritarchs. In: L.R.M., N.H., R.B., J.T. & P.D. (eds.), The Llandovery Series of the type area.- Bulletin of the British Museum of Natural History, London, (Geology), vol. 38, n° 3, p. 174-176.
A. (1989).- Acritarches et kystes d'algues Prasinophycées du Silurien de Gotland, Suède.- Palaeontographia Italica, Pisa, vol. 76, p. 57-302.
R. (1962).- Le Tarannonien à Graptolites reconnu sous Courtrai (Flandre Occidentale).- Bulletin de la Société belge de Géologie, Bruxelles, vol. 70, p. 174-185.
R. (1981).- Le Llandoverien à Graptolites reconnu sous Courtrai.- Professional Papers, Service géologique de Belgique, Bruxelles, vol. 184, 4 p.
T.R. (1970).- The acritarchs and chitinozoa from the Wenlock and Ludlow Series of the Ludlow and Millichope areas, Shropshire.- Palaeontographical Society Monographs, London, vol. 124, part 1, p. 1-100.
D.K. (1992).- Middle and Upper Llandovery graptolites from western mid-Wales. Part 1.- Monograph of the Palaeontological Society, London, vol. 146, p. 1-55.
D.K. (1993).- Middle and Upper Llandovery graptolites from western mid-Wales. Part 2.- Monograph of the Palaeontological Society, London, vol. 147, p. 56-180.
F. (1966).- Les Acritarches du sondage de la brasserie Lust, à Kortrijk (Courtrai) (Silurien belge).- Bulletin de la Société belge de Géologie, Bruxelles, vol. 74, n° 3, p. 354-400.
F. (1969).- Les acritarches de l'Ordovicien et du Silurien belges. Détermination et valeur stratigraphique.- Institut Royal des Sciences Naturelles de Belgique, Bruxelles, 175 p.
S.G., A. & R. (1996).- Acritarch and Prasinophyte Stratigraphy. In: J. & Mc D.C. (eds.), Palynology: principles and applications.- American Association of Stratigraphic Palynologists Foundation, vol. 2, Chapter 16, p. 493-529.
G.L. (2001).- Acritarchs and prasinophyte algae of the Elton Group, Ludlow Series of the type area.- Monograph of the Palaeontological Society, London, n° 616 for vol. 155, 154 p.
C. & B.A. (in press).- Aeronian (middle lower Silurian) palynomorphs and graptolites from the Eastern Cordillera, North-West Argentina.- Geobios, Villeurbanne.
G. (1967).- Mikrofossilien des oberen Llandovery von Dalarne (Schweden).- Kölner Geologische Hefte, Köln, Heft 13, p. 175-187.
M. (1987).- Early Silurian acritarchs and prasinophycean algae from the Ringerike district, Oslo region (Norway).- Review of Palaeobotany and Palynology, Amsterdam, vol. 52, n° 2-3, p. 137-159.
F., J. & S.A.J. (1965).- Evaluation of some Acritarchous Hystricosphere genera.- Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen, Stuttgart, vol. 123, p. 167-201.
F. & Y. (1963).- Les Hystricosphères ou mieux les Acritarches du Silurien belge. Sondage de la Brasserie Lust à Courtrai (Kortrijk).- Bulletin de la Société belge de Géologie, Bruxelles, vol. 71, n° 3, p. 450-481.
G. (1990).- Litho- en Biostratigrafiesche studie met Chitinozoa in het westelijk deel van het Massief van Brabant.- Ph.D. thesis (unpublished), Rijksuniversiteit Gent, 83 + 108 p.
G., J., J. & T. (1998).- Chitinozoa and graptolite biozonation of the Aeronian and lower Telychian in the Brabant Massif (Belgium).- Temas Geologico-Mineros ITGE, Madrid, vol. 23, p. 135-136.
J., V., F., P., S. & G. (1995).- A global Chitinozoa biozonation for the Silurian.- Geological Magazine, Cambridge, vol. 132, n° 6, p. 651-666.
B. (1997).- Étude palynologique des acritarches du sondage Kortrijk 83 W-421, Silurien (Aeronian) du Massif du Brabant, Belgique.- Unpublished M.Sc. thesis (unpublished), Université de Liège, 66 + 40 p.
B. (2003).- Methodological approach to unravel acritarch distribution in chosen sections and boreholes in the late Aeronian-early Telychian from the United Kingdom and Belgium.- Ph.D. thesis (unpublished), Université de Liège, 255 + 250 p.
B. (in press).- Correlation of the 83W-421 Kortrijk (Sint-Antonius) and 83W-44 Kortrijk (Lust) boreholes with acritarchs (late Aeronian-early Telychian, Silurian, Belgium).- Geologica Belgica, Liège.
B., K.J. & A. (2003).- Crassiangulina variacornuta sp. nov. from the late Llandovery and its bearing on Silurian and Devonian acritarch taxonomy.- Bulletin de la Société Géologique de France, vol. 174, n° 1, p. 67-81.
B. & P. (1999).- Biometric study of some Domasia species (Acritarcha) from the Silurian of the Brabant Massif, Belgium.- Bollettino della Società Paleontologica Italiana, Modena, vol. 38, n° 2-3, p. 381-395.
![]()

Click on thumbnail to enlarge the image.
Figure 1: Graptolite distribution in the two boreholes with reference to the biozonation of & (1913). Names of graptolites as in the original papers.
Click on thumbnail to enlarge the image.
Figure 2: Distribution of chosen acritarchs in the two boreholes together with three possible correlations (red lines) discussed in (in press). Photomicrographs are not to scale.

Click on thumbnail to enlarge the image.
Figure 3: Integrated biostratigraphy of the Kortrijk (Sint-Antonius) and Kortrijk (Lust) boreholes based on the correlations of (in press) and data from et alii (1998) and (1997, in press). Letters in the black circles refer to lithologic units defined by (1981).
![]()