![]()
![]()
![]() The Campo Pisano Formation of southwestern Sardinia is represented by an offshore carbonate succession spanning the latest Early to late Middle Cambrian. Paleogeographically, the fauna is characteristic of western Perigondwana, and indicates faunal relations to France, Spain, and Turkey. Microfaunal paleoecology reflects drowning of an isolated carbonate platform at tropical latitudes. Sessile epibenthic filter feeders dominate at the base, succeeded upward by sessile, epibenthic, suspension feeders. Upsection, a shift in the ratio of faunal groups indicates increasing replacement by mobile epibenthos. Autochthonous faunal elements decline near the top where allochthonous taxa become important. In the basal portion of the formation the faunal succession indicates relatively shallow neritic habitats with a moderate influx of suspended sediment, followed by a period of slightly deeper neritic conditions. Probably a shallow bathyal environment was established at the transition to the overlying siliciclastic Cabitza Formation. The bathymetric and ecofacies changes in the Campo Pisano Formation are interpreted as having been caused by a discontinuous rise in eustatic sea-level, probably accompanied by subsidence due to tensional tectonics. The depositional environment was that of a distal open-marine shelf or ramp without strong relief. This interpretation is supported by the lack of significant lateral changes in the fossil assemblages, and an upward deepening of facies. The applied micropaleoecological approach is proven a powerful tool in regional stratigraphy and in the reconstruction of sedimentary realms in the Mediterranean Cambrian, particularly when high-resolution biostratigraphic data and diagnostic sedimentary characteristics are sparse.
The Campo Pisano Formation of southwestern Sardinia is represented by an offshore carbonate succession spanning the latest Early to late Middle Cambrian. Paleogeographically, the fauna is characteristic of western Perigondwana, and indicates faunal relations to France, Spain, and Turkey. Microfaunal paleoecology reflects drowning of an isolated carbonate platform at tropical latitudes. Sessile epibenthic filter feeders dominate at the base, succeeded upward by sessile, epibenthic, suspension feeders. Upsection, a shift in the ratio of faunal groups indicates increasing replacement by mobile epibenthos. Autochthonous faunal elements decline near the top where allochthonous taxa become important. In the basal portion of the formation the faunal succession indicates relatively shallow neritic habitats with a moderate influx of suspended sediment, followed by a period of slightly deeper neritic conditions. Probably a shallow bathyal environment was established at the transition to the overlying siliciclastic Cabitza Formation. The bathymetric and ecofacies changes in the Campo Pisano Formation are interpreted as having been caused by a discontinuous rise in eustatic sea-level, probably accompanied by subsidence due to tensional tectonics. The depositional environment was that of a distal open-marine shelf or ramp without strong relief. This interpretation is supported by the lack of significant lateral changes in the fossil assemblages, and an upward deepening of facies. The applied micropaleoecological approach is proven a powerful tool in regional stratigraphy and in the reconstruction of sedimentary realms in the Mediterranean Cambrian, particularly when high-resolution biostratigraphic data and diagnostic sedimentary characteristics are sparse.
![]() Cambrian, Italy, Sardinia, micropaleontology, microfacies, paleoecology.
Cambrian, Italy, Sardinia, micropaleontology, microfacies, paleoecology.
O. (2006).- Microbiofacies analysis of Cambrian offshore carbonates from Sardinia (Italy): environment reconstruction and development of a drowning carbonate platform.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2006/01 (CG2006_A01)
![]() Analyse
microbiofaciologique de séries carbonatées marines cambriennes de Sardaigne (Italie) :
reconstitution paléoenvironnementale et développement d'une plate-forme
carbonatée ennoyée.- La Formation de Campo Pisano du SW de la
Sardaigne est constituée par une série carbonatée - marine - couvrant un
intervalle de temps qui s'étend du Cambrien inférieur terminal ŕ Cambrien
moyen élevé. Du point de vue de la paléogéographie, la faune y est de type
périgondwanien et présente des affinités occidentales certaines (France,
Espagne et Turquie). L'évolution de la paléoécologie des microfaunes
retranscrit l'ennoiement de cette plate-forme carbonatée isolée en climat
tropical. À la base, les organismes sessiles épibenthiques filtreurs dominent.
En montant dans la série, leur succèdent des formes sessiles épibenthiques
suspensivores. Enfin, vers le sommet, le glissement dans les proportions au sein
des associations fauniques traduit le remplacement croissant des faunes
autochtones par un épibenthos vagile, c'est-ŕ-dire par des faunes allochtones.
Dans la partie basale de la formation, la succession des faunes suggère qu'ŕ
une période initiale marquée par des environnements néritiques relativement
peu profonds, avec de faibles apports détritiques en suspension, a succédé
une période caractérisée par des environnements néritiques légèrement plus
profonds. Le stade bathyal supérieur a vraisemblablement été atteint au toit
de la série, avant le dépôt de la formation silicoclastique de Cabitza. Les
variations écologiques et bathymétriques de la Formation de Campo Pisano
semblent résulter d'une remontée irrégulière du niveau marin, probablement
couplée ŕ une subsidence d'origine tectonique. L'environnement de dépôt
était comparable à celui de la partie distale d'une plate-forme ouverte ou
d'une rampe avec des reliefs peu exprimés comme en atteste l'absence
d'évolution latérale significative des associations fauniques reconnues.
L'analyse micropaléoécologique constitue un outil puissant pour les études
stratigraphiques régionales ainsi que pour les reconstitutions des ceintures de
faciès du Cambrien méditerranéen, en particulier quand figures sédimentaires
caractéristiques et données biostratigraphiques de haute résolution sont
rares ou absentes.
Analyse
microbiofaciologique de séries carbonatées marines cambriennes de Sardaigne (Italie) :
reconstitution paléoenvironnementale et développement d'une plate-forme
carbonatée ennoyée.- La Formation de Campo Pisano du SW de la
Sardaigne est constituée par une série carbonatée - marine - couvrant un
intervalle de temps qui s'étend du Cambrien inférieur terminal ŕ Cambrien
moyen élevé. Du point de vue de la paléogéographie, la faune y est de type
périgondwanien et présente des affinités occidentales certaines (France,
Espagne et Turquie). L'évolution de la paléoécologie des microfaunes
retranscrit l'ennoiement de cette plate-forme carbonatée isolée en climat
tropical. À la base, les organismes sessiles épibenthiques filtreurs dominent.
En montant dans la série, leur succèdent des formes sessiles épibenthiques
suspensivores. Enfin, vers le sommet, le glissement dans les proportions au sein
des associations fauniques traduit le remplacement croissant des faunes
autochtones par un épibenthos vagile, c'est-ŕ-dire par des faunes allochtones.
Dans la partie basale de la formation, la succession des faunes suggère qu'ŕ
une période initiale marquée par des environnements néritiques relativement
peu profonds, avec de faibles apports détritiques en suspension, a succédé
une période caractérisée par des environnements néritiques légèrement plus
profonds. Le stade bathyal supérieur a vraisemblablement été atteint au toit
de la série, avant le dépôt de la formation silicoclastique de Cabitza. Les
variations écologiques et bathymétriques de la Formation de Campo Pisano
semblent résulter d'une remontée irrégulière du niveau marin, probablement
couplée ŕ une subsidence d'origine tectonique. L'environnement de dépôt
était comparable à celui de la partie distale d'une plate-forme ouverte ou
d'une rampe avec des reliefs peu exprimés comme en atteste l'absence
d'évolution latérale significative des associations fauniques reconnues.
L'analyse micropaléoécologique constitue un outil puissant pour les études
stratigraphiques régionales ainsi que pour les reconstitutions des ceintures de
faciès du Cambrien méditerranéen, en particulier quand figures sédimentaires
caractéristiques et données biostratigraphiques de haute résolution sont
rares ou absentes.
![]() Cambrien, Italie, Sardaigne, micropaléontologie, microfaciès, paléoécologie.
Cambrien, Italie, Sardaigne, micropaléontologie, microfaciès, paléoécologie.
![]()
![]() In Early- to Middle Cambrian times carbonate and mixed siliciclastic–carbonate environments were widely distributed on the European Shelf of the low-latitude Perigondwana realm (Morocco, Spain, France, Sardinia, Germany, and the Near– and Middle East; Fig. 1
In Early- to Middle Cambrian times carbonate and mixed siliciclastic–carbonate environments were widely distributed on the European Shelf of the low-latitude Perigondwana realm (Morocco, Spain, France, Sardinia, Germany, and the Near– and Middle East; Fig. 1 ![]() ).
But sedimentary records of these environments are commonly incomplete, due to variations in bed thickness and shelf morphology, together with widespread erosion during the Late Cambrian to Early Ordovician, as in Sardinia (Sardic Phase movements), and possibly also in Germany. Thus, a full reconstruction of early Paleozoic development in this region is still problematic for biostratigraphic, paleogeographic and postdepositional geological processes are not
yet completely defined.
).
But sedimentary records of these environments are commonly incomplete, due to variations in bed thickness and shelf morphology, together with widespread erosion during the Late Cambrian to Early Ordovician, as in Sardinia (Sardic Phase movements), and possibly also in Germany. Thus, a full reconstruction of early Paleozoic development in this region is still problematic for biostratigraphic, paleogeographic and postdepositional geological processes are not
yet completely defined.
![]() Earliest Cambrian sediments on the Gondwana European Shelf are generally not exposed, either because they were not deposited or were removed through tectonism and subsequent erosion. However, from the later Early Cambrian to the late Middle
Cambrian sedimentary successions up to 2000 m or more are well developed (see ,
1974; , 1991; & ,
1995; , 1997; et alii,
2002; et alii,
2003). These deposits show an overall trend upward from siliciclastic to relatively shallow carbonate and mixed carbonate–siliciclastic environments, followed by a siliciclastic cover denoting deepening. Younger Cambrian deposits are only sporadically developed in some areas of the European Shelf. Although the paleontology and sedimentary facies of the carbonate successions (mainly Early Cambrian) and the siliciclastic deposits (Early and Middle Cambrian) are well known, the transition from carbonate to siliciclastic deposition, reflecting the drowning of the carbonate platforms and consequently the reorganization of the Perigondwana realm, has been investigated only minimally in some areas.
Earliest Cambrian sediments on the Gondwana European Shelf are generally not exposed, either because they were not deposited or were removed through tectonism and subsequent erosion. However, from the later Early Cambrian to the late Middle
Cambrian sedimentary successions up to 2000 m or more are well developed (see ,
1974; , 1991; & ,
1995; , 1997; et alii,
2002; et alii,
2003). These deposits show an overall trend upward from siliciclastic to relatively shallow carbonate and mixed carbonate–siliciclastic environments, followed by a siliciclastic cover denoting deepening. Younger Cambrian deposits are only sporadically developed in some areas of the European Shelf. Although the paleontology and sedimentary facies of the carbonate successions (mainly Early Cambrian) and the siliciclastic deposits (Early and Middle Cambrian) are well known, the transition from carbonate to siliciclastic deposition, reflecting the drowning of the carbonate platforms and consequently the reorganization of the Perigondwana realm, has been investigated only minimally in some areas.
![]() The main reason for the limited investigation of this critical interval in Sardinia may lie in the lithologic character of the transition, represented in general by rather monotonous, condensed, often nodular limestones, affected by a strong diagenetic overprint ( & ,
1990; , 2001). Nevertheless, the carbonates are commonly rich in shelly microfossils that usually are an important means of obtaining additional information on depositional conditions. However, here this microfossil
wealth is frequently hard to bring to light. It is an extremely intricate, time-consuming and almost impossible task to extract these fossils from the host
rocks if they are not diagenetically silicified or preserved as steinkerns. This problem exists for nearly all the important fossil groups other than organophosphatic brachiopods and problematic small shelly
fossils that can be extracted by acidic dissolution. Furthermore, organic biota, such as palynomorphs are usually not preserved in the mostly light- and reddish-colored carbonates. These hindrances have led to a paleontological focus on archaeocyaths in Early Cambrian carbonate rocks and on trilobites in Early and Middle Cambrian siliciclastic beds. Therefore, the Middle Cambrian drowning stage of the Sardinian platform has not been properly investigated
paleontologically and geological processes near the Early to Middle Cambrian boundary are not well known.
The main reason for the limited investigation of this critical interval in Sardinia may lie in the lithologic character of the transition, represented in general by rather monotonous, condensed, often nodular limestones, affected by a strong diagenetic overprint ( & ,
1990; , 2001). Nevertheless, the carbonates are commonly rich in shelly microfossils that usually are an important means of obtaining additional information on depositional conditions. However, here this microfossil
wealth is frequently hard to bring to light. It is an extremely intricate, time-consuming and almost impossible task to extract these fossils from the host
rocks if they are not diagenetically silicified or preserved as steinkerns. This problem exists for nearly all the important fossil groups other than organophosphatic brachiopods and problematic small shelly
fossils that can be extracted by acidic dissolution. Furthermore, organic biota, such as palynomorphs are usually not preserved in the mostly light- and reddish-colored carbonates. These hindrances have led to a paleontological focus on archaeocyaths in Early Cambrian carbonate rocks and on trilobites in Early and Middle Cambrian siliciclastic beds. Therefore, the Middle Cambrian drowning stage of the Sardinian platform has not been properly investigated
paleontologically and geological processes near the Early to Middle Cambrian boundary are not well known.
![]() In southwestern Sardinia the drowning stage of the Cambrian carbonate platform is represented by the Campo Pisano Formation. A genetic interpretation of the depositional environment of this succession is problematic because of its limited sedimentological characteristics. Previous environmental interpretations ranged from shallow water, low-energy areas (,
1979) to lagoonal and partly euxinic tracts (,
1979), to ammonitico rosso-type deposits (the typical Tethyan red-colored, nodular, deep water facies; ,
1985; , 1987b; et alii,
1988), that are interpreted as being on tectonically unstable shelves with highs and troughs ( & ,
1990). The fauna of the Campo Pisano Formation is mainly trilobites, echinoderms, poriferans, brachiopods, and chancelloriids. Nevertheless, this fauna has been investigated very scantily because of its extremely poor accessibility. Only some trilobites, a few foraminiferans, poriferan spicules, and hadimopanellids (dermal plates of palaeoscolecid worms) have been the subject of sporadic work ( & ,
1985; , 1985; ,
1991). Systematic studies on mollusks, trilobites, brachiopods and some small shelly fossils were done recently by
(2002), et alii
(2003), &
(2004), and &
(2004).
In southwestern Sardinia the drowning stage of the Cambrian carbonate platform is represented by the Campo Pisano Formation. A genetic interpretation of the depositional environment of this succession is problematic because of its limited sedimentological characteristics. Previous environmental interpretations ranged from shallow water, low-energy areas (,
1979) to lagoonal and partly euxinic tracts (,
1979), to ammonitico rosso-type deposits (the typical Tethyan red-colored, nodular, deep water facies; ,
1985; , 1987b; et alii,
1988), that are interpreted as being on tectonically unstable shelves with highs and troughs ( & ,
1990). The fauna of the Campo Pisano Formation is mainly trilobites, echinoderms, poriferans, brachiopods, and chancelloriids. Nevertheless, this fauna has been investigated very scantily because of its extremely poor accessibility. Only some trilobites, a few foraminiferans, poriferan spicules, and hadimopanellids (dermal plates of palaeoscolecid worms) have been the subject of sporadic work ( & ,
1985; , 1985; ,
1991). Systematic studies on mollusks, trilobites, brachiopods and some small shelly fossils were done recently by
(2002), et alii
(2003), &
(2004), and &
(2004).
![]() This paper presents for the first time a regionally-scaled detailed micropaleontological and
paleoecological analysis of the Campo Pisano Formation. Based on extensive field
work that involved the measurement and sampling of 19 stratigraphic sections, together with the study of about 300 thin-sections, some chemical preparations, and sedimentologic and diagenetic observations, a paleoecologic reconstruction of the drowning of the Sardinian Perigondwanan platform is
feasible and offered here.
This paper presents for the first time a regionally-scaled detailed micropaleontological and
paleoecological analysis of the Campo Pisano Formation. Based on extensive field
work that involved the measurement and sampling of 19 stratigraphic sections, together with the study of about 300 thin-sections, some chemical preparations, and sedimentologic and diagenetic observations, a paleoecologic reconstruction of the drowning of the Sardinian Perigondwanan platform is
feasible and offered here.
![]() The Lower Paleozoic strata of southwestern Sardinia represent an autochthonous unit bordered to the east and north by Hercynian traps
(Fig. 2
The Lower Paleozoic strata of southwestern Sardinia represent an autochthonous unit bordered to the east and north by Hercynian traps
(Fig. 2 ![]() ). Prominent Cenozoic grabens separate this part of Sardinia from the remainder of the island and divide the area internally into several geographic regions. Locally, Hercynian granitoids have metamorphosed the Lower Paleozoic rocks.
). Prominent Cenozoic grabens separate this part of Sardinia from the remainder of the island and divide the area internally into several geographic regions. Locally, Hercynian granitoids have metamorphosed the Lower Paleozoic rocks.
![]() Cambrian sediments are widely exposed in southwestern Sardinia
(Fig. 2
Cambrian sediments are widely exposed in southwestern Sardinia
(Fig. 2 ![]() ) and reach a thickness of 1500–2000 m (,
1991). Initial investigations were focused mainly on paleontology and biostratigraphy, but work has now been done on sedimentology, litho- and biofacies, paleogeography and economic aspects (e.g., ,
1886, 1891; ,
1964; , 1972; ,
1979; & ,
1984; , 1985; ,
1987a, 1987b; & ,
1990; , 1991; & ,
1994; , 2002; et alii,
2002; et alii,
2003; & ,
2004; & ,
2004).
) and reach a thickness of 1500–2000 m (,
1991). Initial investigations were focused mainly on paleontology and biostratigraphy, but work has now been done on sedimentology, litho- and biofacies, paleogeography and economic aspects (e.g., ,
1886, 1891; ,
1964; , 1972; ,
1979; & ,
1984; , 1985; ,
1987a, 1987b; & ,
1990; , 1991; & ,
1994; , 2002; et alii,
2002; et alii,
2003; & ,
2004; & ,
2004).
![]() When the deposition of deposits of Cambrian age began in Sardinia is not precisely known: the only area where underlying rocks (Precambrian gneisses) are exposed is the southern part of SW Sardinia,
but here the transition from gneisses to Cambrian sediments is represented by a nonconformity ( & ,
1983). From bottom to top, the Cambrian succession is divided into the Nebida, Gonnesa, and Iglesias groups
(Fig. 3
When the deposition of deposits of Cambrian age began in Sardinia is not precisely known: the only area where underlying rocks (Precambrian gneisses) are exposed is the southern part of SW Sardinia,
but here the transition from gneisses to Cambrian sediments is represented by a nonconformity ( & ,
1983). From bottom to top, the Cambrian succession is divided into the Nebida, Gonnesa, and Iglesias groups
(Fig. 3 ![]() ; ,
1991). The middle to late Early Cambrian Nebida Group consists of clay-, silt- and sandstone in the lower part (Matoppa Formation), and a mixed carbonate–siliciclastic unit in the upper part (Punta Manna Formation). In southwesternmost Sardinia a discrete siliciclastic unit, the so-called "Bithia Formation", is described below the Nebida Group by &
(1983) and et alii
(1987). This placement was doubted by later authors who consider it as slightly heteropic facies of the lower Nebida Group ( & ,
1989). The Nebida Group is widely accepted as indicating a shallowing upward from a deep-water siliciclastic facies to a shallow mixed carbonate–siliciclastic ramp (e.g., et alii,
1985; , 1987; & ,
1994. Alternative interpretations initiate this evolution with a prodelta environment ,
1979, , 1979, and et alii,
1987). The overlying Gonnesa Group (late Early Cambrian) is characterized almost entirely by carbonate deposits with no siliciclastic influx; thus this group represents an isolated carbonate platform (sensu ,
1985; e.g., & ,
1994, and cited literature). Low in this group, higher salinity is denoted by the restricted, dolomitic, very shallow subtidal to intertidal laminites of the Santa Barbara Formation, overlain by shallow subtidal, open-marine limestones of a discrete and different facies (San Giovanni Formation). The youngest Cambrian unit is the Iglesias Group (latest Early Cambrian to Early Ordovician). At the base of this group are the limestones of the Campo Pisano Formation (at times nodular) deposited during platform drowning. It is succeeded by the deeper-water siliciclastics of the Cabitza Formation
(Fig. 3
; ,
1991). The middle to late Early Cambrian Nebida Group consists of clay-, silt- and sandstone in the lower part (Matoppa Formation), and a mixed carbonate–siliciclastic unit in the upper part (Punta Manna Formation). In southwesternmost Sardinia a discrete siliciclastic unit, the so-called "Bithia Formation", is described below the Nebida Group by &
(1983) and et alii
(1987). This placement was doubted by later authors who consider it as slightly heteropic facies of the lower Nebida Group ( & ,
1989). The Nebida Group is widely accepted as indicating a shallowing upward from a deep-water siliciclastic facies to a shallow mixed carbonate–siliciclastic ramp (e.g., et alii,
1985; , 1987; & ,
1994. Alternative interpretations initiate this evolution with a prodelta environment ,
1979, , 1979, and et alii,
1987). The overlying Gonnesa Group (late Early Cambrian) is characterized almost entirely by carbonate deposits with no siliciclastic influx; thus this group represents an isolated carbonate platform (sensu ,
1985; e.g., & ,
1994, and cited literature). Low in this group, higher salinity is denoted by the restricted, dolomitic, very shallow subtidal to intertidal laminites of the Santa Barbara Formation, overlain by shallow subtidal, open-marine limestones of a discrete and different facies (San Giovanni Formation). The youngest Cambrian unit is the Iglesias Group (latest Early Cambrian to Early Ordovician). At the base of this group are the limestones of the Campo Pisano Formation (at times nodular) deposited during platform drowning. It is succeeded by the deeper-water siliciclastics of the Cabitza Formation
(Fig. 3 ![]() ). For detailed description of the origin and development of the early platform stages (Nebida and Gonnesa groups), see the cited literature or the extended summary by &
(1994), and the references cited therein.
). For detailed description of the origin and development of the early platform stages (Nebida and Gonnesa groups), see the cited literature or the extended summary by &
(1994), and the references cited therein.
![]() The upper boundary of the Cambrian succession lies within the siliciclastic Cabitza
Formation which continues to late Tremadocian times. The Cabitza Formation is truncated by a prominent erosional discordance caused by mid-Ordovician Sardic tectonic movements on a regional
scale ( et alii, 1994; et alii,
2002).
The upper boundary of the Cambrian succession lies within the siliciclastic Cabitza
Formation which continues to late Tremadocian times. The Cabitza Formation is truncated by a prominent erosional discordance caused by mid-Ordovician Sardic tectonic movements on a regional
scale ( et alii, 1994; et alii,
2002).
![]() The contact between the Campo Pisano Formation and the underlying massive limestones of the San Giovanni formation is abrupt, sharply defined
(Fig. 3
The contact between the Campo Pisano Formation and the underlying massive limestones of the San Giovanni formation is abrupt, sharply defined
(Fig. 3 ![]() ), and in many localities is marked by a breccia consisting mainly of San Giovanni clasts. But fragments from the basal Campo Pisano occur locally. (e.g., at San Benedetto, Mt. Marganai area, Mt. Tasua, Acquacadda, Terraseo,
Fig. 4
), and in many localities is marked by a breccia consisting mainly of San Giovanni clasts. But fragments from the basal Campo Pisano occur locally. (e.g., at San Benedetto, Mt. Marganai area, Mt. Tasua, Acquacadda, Terraseo,
Fig. 4 ![]() ) ( et alii,
1981; , 1985; ,
1987a, 1987b). The Campo Pisano Formation is mainly wavy-bedded to nodular limestone with a variable clay content and a few quartz grains, although the siliciclastic content is greater locally (e.g., sections at Su Banatzu, Mt. Tasua, Terraseo,
Fig. 4
) ( et alii,
1981; , 1985; ,
1987a, 1987b). The Campo Pisano Formation is mainly wavy-bedded to nodular limestone with a variable clay content and a few quartz grains, although the siliciclastic content is greater locally (e.g., sections at Su Banatzu, Mt. Tasua, Terraseo,
Fig. 4 ![]() ) and continues farther East and South (Sulcis area),
(1985). The most prominent features of the Campo Pisano Formation are: (1) the generally rich fossil content and (2) millimeter- to centimeter-thick bands of insoluble material, generated by diagenetic processes (,
1987b; late diagenetic pressure dissolution: ,
2001). (1979) and
(1985) reported thin clay-rich and silty layers from the Campo Pisano Formation. The degree of diagenetic nodularity of the limestone varies both laterally and vertically. There is no regional or stratigraphic trend in the occurrence or intensity of nodularity, nor any correlation with a particular microfacies (see below).
) and continues farther East and South (Sulcis area),
(1985). The most prominent features of the Campo Pisano Formation are: (1) the generally rich fossil content and (2) millimeter- to centimeter-thick bands of insoluble material, generated by diagenetic processes (,
1987b; late diagenetic pressure dissolution: ,
2001). (1979) and
(1985) reported thin clay-rich and silty layers from the Campo Pisano Formation. The degree of diagenetic nodularity of the limestone varies both laterally and vertically. There is no regional or stratigraphic trend in the occurrence or intensity of nodularity, nor any correlation with a particular microfacies (see below).
![]() The limestone of the Campo Pisano Formation is generally micrite-dominated. The most prominent lithofacies is light grey, yellowish, or reddish bioclastic wackestone (Figs.
4B
The limestone of the Campo Pisano Formation is generally micrite-dominated. The most prominent lithofacies is light grey, yellowish, or reddish bioclastic wackestone (Figs.
4B ![]() ,
5
,
5 ![]() & 6
& 6 ![]() ) with qualitatively and quantitatively distinct fossil content (see next section). Bioclastic floatstone is another important lithofacies that occurs as intercalations at nearly all the sites investigated and in some sections occurs in significant quantities (e.g., Su Corovau, Mt. Serrau;
Fig. 4
) with qualitatively and quantitatively distinct fossil content (see next section). Bioclastic floatstone is another important lithofacies that occurs as intercalations at nearly all the sites investigated and in some sections occurs in significant quantities (e.g., Su Corovau, Mt. Serrau;
Fig. 4 ![]() ). Fossil-bearing mudstone is intercalated in varying amounts
(Fig. 4B
). Fossil-bearing mudstone is intercalated in varying amounts
(Fig. 4B ![]() ). Bioclastic packstone was found as a small layer in only one section (Monte Scorra;
Fig. 4B
). Bioclastic packstone was found as a small layer in only one section (Monte Scorra;
Fig. 4B ![]() ). More packstone occurrences were mentioned by
(1985), (1987b), et alii
(1988) and &
(1994), but these authors did not report grainstone intercalations in the Campo Pisano Formation. Generally, wackestone and floatstone are more common in the western to northwestern areas ranging from the San Benedetto district to Su Corovau and Campo Pisano via Carbonia up to Su Banatzu, whereas mudstone is more abundant to the east and southeast
(Fig. 4
). More packstone occurrences were mentioned by
(1985), (1987b), et alii
(1988) and &
(1994), but these authors did not report grainstone intercalations in the Campo Pisano Formation. Generally, wackestone and floatstone are more common in the western to northwestern areas ranging from the San Benedetto district to Su Corovau and Campo Pisano via Carbonia up to Su Banatzu, whereas mudstone is more abundant to the east and southeast
(Fig. 4 ![]() ). Within the interval of transition to the overlying siliciclastic Cabitza Formation, limestones commonly alternate with thin greenish claystones.
). Within the interval of transition to the overlying siliciclastic Cabitza Formation, limestones commonly alternate with thin greenish claystones.
![]() Distinct (planar) bedding is not developed in the Campo Pisano Formation. Significant early compaction of the sediment was not observed. The fossil content is often random within the sediment
(Figs. 6-1
Distinct (planar) bedding is not developed in the Campo Pisano Formation. Significant early compaction of the sediment was not observed. The fossil content is often random within the sediment
(Figs. 6-1 ![]() ,
6-2
,
6-2 ![]() & 6-7
& 6-7 ![]() ), most probably, the result of shallow bioturbation
(Fig. 6-7
), most probably, the result of shallow bioturbation
(Fig. 6-7 ![]() ), though recognizable burrows were not seen.
), though recognizable burrows were not seen.
![]() The Campo Pisano Formation ranges in thickness from about 30 to100 m, but most of the mapped sections are 40–60 m thick. From the "San Benedetto–Su Banatzu line" the thickness of the sections tends to decrease to the west–northwest as well as to the east–southeast
(Fig. 4A
The Campo Pisano Formation ranges in thickness from about 30 to100 m, but most of the mapped sections are 40–60 m thick. From the "San Benedetto–Su Banatzu line" the thickness of the sections tends to decrease to the west–northwest as well as to the east–southeast
(Fig. 4A ![]() ). The occurrence of breccias at the base of the Campo Pisano Formation is reported from several sites by et alii
(1982), (1987a,
1987b), and et alii
(1988). The distribution of these coarse clastics shows no distinct regional trend.
(1985) and &
(1989) described slumping and mass flow deposits from different locations in the area.
). The occurrence of breccias at the base of the Campo Pisano Formation is reported from several sites by et alii
(1982), (1987a,
1987b), and et alii
(1988). The distribution of these coarse clastics shows no distinct regional trend.
(1985) and &
(1989) described slumping and mass flow deposits from different locations in the area.
![]() Unlike the very pure limestone of the underlying San Giovanni Formation (isolated carbonate platform stage sensu ,
1985) and the overlying siliciclastic Cabitza Formation (cover of the drowned platform), the deposits of the Campo Pisano Formation are characterized by a rich fauna of shelly fossils.
Unlike the very pure limestone of the underlying San Giovanni Formation (isolated carbonate platform stage sensu ,
1985) and the overlying siliciclastic Cabitza Formation (cover of the drowned platform), the deposits of the Campo Pisano Formation are characterized by a rich fauna of shelly fossils.
![]() The most numerous are trilobites, brachiopods and echinoderms, with poriferans and chancelloriids locally common
(Figs. 6-5
The most numerous are trilobites, brachiopods and echinoderms, with poriferans and chancelloriids locally common
(Figs. 6-5 ![]() , 6-6
, 6-6 ![]() , 6-7
, 6-7 ![]() , 6-8
, 6-8 ![]() , 6-9
, 6-9 ![]() , 6-12
, 6-12 ![]() & 7
& 7 ![]() ). The proportion of molluscs (pelagiellid gastropods), hyoliths
(Fig. 6-13
). The proportion of molluscs (pelagiellid gastropods), hyoliths
(Fig. 6-13 ![]() ) and endolithic filamentous microbes is distinctly lower. Phosphatic small shelly fossils (palaeoscolecids, cambroclaves;
Figs. 6-10
) and endolithic filamentous microbes is distinctly lower. Phosphatic small shelly fossils (palaeoscolecids, cambroclaves;
Figs. 6-10 ![]() & 6-11
& 6-11 ![]() ) are present only sporadically in some beds of several sections.
) are present only sporadically in some beds of several sections.
![]() The trilobite content is generally high but varies within sections
(Fig. 7
The trilobite content is generally high but varies within sections
(Fig. 7 ![]() ). The size of the identified trilobite remains ranges from a few millimeters up to one centimeter. Although most of them were recognized in thin sections
(Fig. 6-7
). The size of the identified trilobite remains ranges from a few millimeters up to one centimeter. Although most of them were recognized in thin sections
(Fig. 6-7 ![]() ), some rocks yield silicified remains, so that dissolution to expose three-dimensional pseudomorphs made taxonomic investigation possible
(Fig. 6-9
), some rocks yield silicified remains, so that dissolution to expose three-dimensional pseudomorphs made taxonomic investigation possible
(Fig. 6-9 ![]() ) ( & ,
2004). The chemical digestion procedure typically yields trilobite remains consisting mainly of disarticulated cephala, pygidia, and a range of thoracic elements, but in some cases enrolled specimens and early ontogenetic protaspis and meraspis stages were obtained ( et alii,
2003; & ,
2004). Generally, agnostid and polymeran species are present together. Among agnostids, Dawsonia bohemica
(Fig. 6-7
) ( & ,
2004). The chemical digestion procedure typically yields trilobite remains consisting mainly of disarticulated cephala, pygidia, and a range of thoracic elements, but in some cases enrolled specimens and early ontogenetic protaspis and meraspis stages were obtained ( et alii,
2003; & ,
2004). Generally, agnostid and polymeran species are present together. Among agnostids, Dawsonia bohemica
(Fig. 6-7 ![]() ) and Condylopyge antiqua are very common, whereas Peronopsis is rather rare. Polymerans are represented mainly by Corynexochus and less commonly by Clavigellus, Dorypyge, Agraulos and Protolenus cf. pisidianus. Taxonomic descriptions and discussions of this trilobite fauna are given in &
(2004).
) and Condylopyge antiqua are very common, whereas Peronopsis is rather rare. Polymerans are represented mainly by Corynexochus and less commonly by Clavigellus, Dorypyge, Agraulos and Protolenus cf. pisidianus. Taxonomic descriptions and discussions of this trilobite fauna are given in &
(2004).
![]() In large part brachiopods too are preserved as silicified pseudomorphs, but at one locality (Gutturu Pala) there are phosphatic shells. Two groups of linguliformeans are recorded in great quantity: lingulides (Lingulella, Schmidtites) and acrotretides (Opisthotreta, Vandalotreta, Linnarssonella).
In large part brachiopods too are preserved as silicified pseudomorphs, but at one locality (Gutturu Pala) there are phosphatic shells. Two groups of linguliformeans are recorded in great quantity: lingulides (Lingulella, Schmidtites) and acrotretides (Opisthotreta, Vandalotreta, Linnarssonella).
![]() Echinoderm (Fig.
6-6
Echinoderm (Fig.
6-6 ![]() ) remains occur generally as disarticulated plates that often preserve the original netlike stereome structure. Phosphatized plates, segments and barrel-shaped elements are obtained by acid digestion
(Fig. 6-12
) remains occur generally as disarticulated plates that often preserve the original netlike stereome structure. Phosphatized plates, segments and barrel-shaped elements are obtained by acid digestion
(Fig. 6-12 ![]() ). The rarity of barrel-shaped elements is interpreted to indicate (1) a rather small proportion of stemmed forms and (2) that the stems were probably rather short. Much more widely distributed are polygonal thecal plates, commonly with sutural pores (epispires) for respiration (,
1992). Frequent annulate-segmented sclerites may represent brachiole or ambulacral parts.
(1985) stated that most of the Campo Pisano Formation echinoderm remains represent eocrinoids,
but a minor content of cincta (carpoideans) may be present too. Unfortunately, most were recognized in thin-sections so taxonomy is rendered difficult. But, epispires (typical in eocrinoids, cincta and some edrioasteriods) and the estimated ratios of sclerite morphologies: ([1] dominance of small probable brachiole/ambulacral elements, [2] many thecal plates, [3] rare stem segments, [4] a lack of typical stylocone segments sensu & ,
2005, and of [5] cinctan marginal plates and [6] axis channels sensu ,
1998),do indeed point to a probable dominance of eocrinoids. However, such a differentiation is not of key importance for basic paleoecological interpretations.
). The rarity of barrel-shaped elements is interpreted to indicate (1) a rather small proportion of stemmed forms and (2) that the stems were probably rather short. Much more widely distributed are polygonal thecal plates, commonly with sutural pores (epispires) for respiration (,
1992). Frequent annulate-segmented sclerites may represent brachiole or ambulacral parts.
(1985) stated that most of the Campo Pisano Formation echinoderm remains represent eocrinoids,
but a minor content of cincta (carpoideans) may be present too. Unfortunately, most were recognized in thin-sections so taxonomy is rendered difficult. But, epispires (typical in eocrinoids, cincta and some edrioasteriods) and the estimated ratios of sclerite morphologies: ([1] dominance of small probable brachiole/ambulacral elements, [2] many thecal plates, [3] rare stem segments, [4] a lack of typical stylocone segments sensu & ,
2005, and of [5] cinctan marginal plates and [6] axis channels sensu ,
1998),do indeed point to a probable dominance of eocrinoids. However, such a differentiation is not of key importance for basic paleoecological interpretations.
![]() Disarticulated monaxons, tetractines, and pentactines are the most frequent sponge remains
(Fig. 6-8
Disarticulated monaxons, tetractines, and pentactines are the most frequent sponge remains
(Fig. 6-8 ![]() ).
But, only the monaxons are occasionally abundant in discrete layers
(Fig. 6-5
).
But, only the monaxons are occasionally abundant in discrete layers
(Fig. 6-5 ![]() ).
(1985) reported polyactinellid heteractinides from the Pisano Formation (Phobetractina polymorpha). Another heteractinide, Eiffelia araniformis, and the possible calcarean Dodecaactinella triradiata were identified.
).
(1985) reported polyactinellid heteractinides from the Pisano Formation (Phobetractina polymorpha). Another heteractinide, Eiffelia araniformis, and the possible calcarean Dodecaactinella triradiata were identified.
![]() Chancelloriids, a systematically problematic
'sponge-like'group, are lumped as coeloscleritophorans together with halkieriids, wiwaxiids, and siphonogonuchitids by &
(1981), but considered as a polyphyletic, non-poriferan group of unknown systematic affinity by
(1999) and et alii
(2002). In the Sardinian samples most of the generally poorly preserved chancelloriids are represented by three- or four-rayed (Allonnia-type) and rosette-like (Chancelloria-type) sclerites. Many of the sclerites are single disarticulated rays. Identified forms include Allonnia tripodophora, Chancelloria sardinica, and Archiasterella pentactina.
(1985) mentions Chancelloria maroccana and Archiasterella sp. as occurring in the Campo Pisano Formation.
Chancelloriids, a systematically problematic
'sponge-like'group, are lumped as coeloscleritophorans together with halkieriids, wiwaxiids, and siphonogonuchitids by &
(1981), but considered as a polyphyletic, non-poriferan group of unknown systematic affinity by
(1999) and et alii
(2002). In the Sardinian samples most of the generally poorly preserved chancelloriids are represented by three- or four-rayed (Allonnia-type) and rosette-like (Chancelloria-type) sclerites. Many of the sclerites are single disarticulated rays. Identified forms include Allonnia tripodophora, Chancelloria sardinica, and Archiasterella pentactina.
(1985) mentions Chancelloria maroccana and Archiasterella sp. as occurring in the Campo Pisano Formation.
![]() Hyolithelminths (probable polychaetes), are nearly straight phosphatic open tubes with a more-or-less coiled proximal part and a round cross-section. The few specimens, up to 3 mm long, collected from near the base of the Campo Pisano Formation, are identified as Hyolithellus cf. filiformis ( et alii,
2003). A few specimens of the first known Sardinian bradoriid were found at the same stratigraphic level. Their subtriangular shape, phosphatic mineralogy, and other surface morphologic features identify them as Hipponicharion ichnusum ( & ,
2004).
Hyolithelminths (probable polychaetes), are nearly straight phosphatic open tubes with a more-or-less coiled proximal part and a round cross-section. The few specimens, up to 3 mm long, collected from near the base of the Campo Pisano Formation, are identified as Hyolithellus cf. filiformis ( et alii,
2003). A few specimens of the first known Sardinian bradoriid were found at the same stratigraphic level. Their subtriangular shape, phosphatic mineralogy, and other surface morphologic features identify them as Hipponicharion ichnusum ( & ,
2004).
![]() Molluscs are represented by pelagiellid and hyolith steinkerns with partially preserved pseudomorph shells. Though the systematic affinity of pelagiellids is debated ( & ,
2000; , 2001; et alii,
2002), the Sardinian specimens all belong to Pelagiella subangulata (for detailed description see ,
2002). Both hyolithomorphs and orthothecimorphs are present in the Campo Pisano Formation, but the only genus identified thus far is Microcornus
(Fig. 6-13
Molluscs are represented by pelagiellid and hyolith steinkerns with partially preserved pseudomorph shells. Though the systematic affinity of pelagiellids is debated ( & ,
2000; , 2001; et alii,
2002), the Sardinian specimens all belong to Pelagiella subangulata (for detailed description see ,
2002). Both hyolithomorphs and orthothecimorphs are present in the Campo Pisano Formation, but the only genus identified thus far is Microcornus
(Fig. 6-13 ![]() ).
).
![]() So called "small shelly fossils" (phosphatic microproblematica) are represented by hadimopanellids and cambroclaves. The hadimopanellids (considered to be palaeoscolecids; et alii,
1990; & ,
1993) are represented by very well preserved specimens of Hadimopanella
(Fig. 6-10
So called "small shelly fossils" (phosphatic microproblematica) are represented by hadimopanellids and cambroclaves. The hadimopanellids (considered to be palaeoscolecids; et alii,
1990; & ,
1993) are represented by very well preserved specimens of Hadimopanella
(Fig. 6-10 ![]() ), abundant in discrete horizons of the lower portion of the Su Corovau section
(Fig. 4
), abundant in discrete horizons of the lower portion of the Su Corovau section
(Fig. 4 ![]() ). In addition, some poorly preserved sclerites are known from the Campo Pisano Formation type section (Hadimopanella cf. oezgueli; see & ,
1984).
(1985) also mention the occurrence of hadimopanellids in the Gutturu Pala section
(Fig. 4
). In addition, some poorly preserved sclerites are known from the Campo Pisano Formation type section (Hadimopanella cf. oezgueli; see & ,
1984).
(1985) also mention the occurrence of hadimopanellids in the Gutturu Pala section
(Fig. 4 ![]() ), but gives no detailed information or figures. Cambroclaves
(Fig. 6-11
), but gives no detailed information or figures. Cambroclaves
(Fig. 6-11 ![]() ) are represented by minute sclerites of probable wormlike animals. They came from the same locality and levels as the hadimopanellids ( & ,
2003).
) are represented by minute sclerites of probable wormlike animals. They came from the same locality and levels as the hadimopanellids ( & ,
2003).
![]() Microbes occur in the Campo Pisano Formation too. The endolithic cyanobacterium Endoconchia angusta was observed between the steinkerns and shell pseudomorphs of molluscs, and some single specimens of the possible oscillatoriacean Girvanella problematica are reported from the Campo Pisano Formation by &
(1984). The lack of other microbial fossils may be to the result of taphonomic and
diagenetic phenomena.
Microbes occur in the Campo Pisano Formation too. The endolithic cyanobacterium Endoconchia angusta was observed between the steinkerns and shell pseudomorphs of molluscs, and some single specimens of the possible oscillatoriacean Girvanella problematica are reported from the Campo Pisano Formation by &
(1984). The lack of other microbial fossils may be to the result of taphonomic and
diagenetic phenomena.
![]() The microfauna of the Campo Pisano Formation is chaotically oriented
(Fig. 6-7
The microfauna of the Campo Pisano Formation is chaotically oriented
(Fig. 6-7 ![]() ). There is no parallel "bedding", distinct layering, or size gradation. Even though no clear indication of burrows exists, the chaotic orientation of the biota in this succession, which was deposited at a low rate of sedimentation ( et alii,
1995), might be interpreted to be a result of bioturbation. Nevertheless, because of the lack of infaunal burrows or other deep
traces and the absence of fecal pellets, a rather shallow bioturbation near the sediment surface can be assumed. The lack of surface traces of bioturbation may have been caused by intensive bioerosion due to low sedimentation rates, or by late diagenetic pressure dissolution (,
2001). Most of the fossils are preserved as calcite pseudomorphs.
). There is no parallel "bedding", distinct layering, or size gradation. Even though no clear indication of burrows exists, the chaotic orientation of the biota in this succession, which was deposited at a low rate of sedimentation ( et alii,
1995), might be interpreted to be a result of bioturbation. Nevertheless, because of the lack of infaunal burrows or other deep
traces and the absence of fecal pellets, a rather shallow bioturbation near the sediment surface can be assumed. The lack of surface traces of bioturbation may have been caused by intensive bioerosion due to low sedimentation rates, or by late diagenetic pressure dissolution (,
2001). Most of the fossils are preserved as calcite pseudomorphs.
![]() Mechanical abrasion of the fossil remains was not seen. Even very fine and fragile structures on cambroclaves and microbrachiopods are perfectly preserved
(Figs. 6-10
Mechanical abrasion of the fossil remains was not seen. Even very fine and fragile structures on cambroclaves and microbrachiopods are perfectly preserved
(Figs. 6-10 ![]() & 6-11
& 6-11 ![]() ). That portion of the fauna that had scleritomes covering or embedded in soft
tissue is represented exclusively by disarticulated sclerites (poriferans, chancelloriids, echinoderms, etc.). Postmortem microbial decomposition of organic matter may have led to disarticulation in situ rather than as an accumulation caused by currents or slides. Too, trilobites in thin sections commonly show articulations (if the orientation of the section permits their recognition). These observations, together with the absence of lithoclasts and the near-total absence of coarse siliciclastics suggest that taphocoenosis was autochthonous. Cone-shaped fossils such as hyoliths occasionally show cone-in-cone stacking, thus indicating some bottom currents with low to moderate energy. Generally, bioclasts become fewer and smaller immediately below the transition to the overlying Cabitza Formation.
). That portion of the fauna that had scleritomes covering or embedded in soft
tissue is represented exclusively by disarticulated sclerites (poriferans, chancelloriids, echinoderms, etc.). Postmortem microbial decomposition of organic matter may have led to disarticulation in situ rather than as an accumulation caused by currents or slides. Too, trilobites in thin sections commonly show articulations (if the orientation of the section permits their recognition). These observations, together with the absence of lithoclasts and the near-total absence of coarse siliciclastics suggest that taphocoenosis was autochthonous. Cone-shaped fossils such as hyoliths occasionally show cone-in-cone stacking, thus indicating some bottom currents with low to moderate energy. Generally, bioclasts become fewer and smaller immediately below the transition to the overlying Cabitza Formation.
![]() There is no clear correlation between the occurrence of certain fossil groups and carbonate textural types (mudstone, floatstone, wackestone, packstone). Sections in the southeastern part of the study area tend to have fewer fossils than sections in the northwest. Generally, the various fossil groups show a similar pattern of stratigraphic distribution everywhere: at most localities fossil content is highest at the base of the Campo Pisano Formation. In nearly every section, the Campo Pisano Formation begins with a poriferan spiculite (locally as mass accumulations; e.g., at Su Banatzu, Terraseo West, Santa Barbara, Monte Scorra, Su Corovau, Monte Serrau;
Figs. 4
There is no clear correlation between the occurrence of certain fossil groups and carbonate textural types (mudstone, floatstone, wackestone, packstone). Sections in the southeastern part of the study area tend to have fewer fossils than sections in the northwest. Generally, the various fossil groups show a similar pattern of stratigraphic distribution everywhere: at most localities fossil content is highest at the base of the Campo Pisano Formation. In nearly every section, the Campo Pisano Formation begins with a poriferan spiculite (locally as mass accumulations; e.g., at Su Banatzu, Terraseo West, Santa Barbara, Monte Scorra, Su Corovau, Monte Serrau;
Figs. 4 ![]() & 7
& 7 ![]() ), whereas other fossil groups are clearly under–represented (rare brachiopods, small trilobites or echinoderms and single hyolithelminths and bradoriids), or totally absent. This remarkable initial biofacies is referred to here as
'biofacies unit 1' (bu1; Figs.
6-5
), whereas other fossil groups are clearly under–represented (rare brachiopods, small trilobites or echinoderms and single hyolithelminths and bradoriids), or totally absent. This remarkable initial biofacies is referred to here as
'biofacies unit 1' (bu1; Figs.
6-5 ![]() ,
7
,
7 ![]() & 8
& 8 ![]() ).
).
![]() The overlying
'biofacies unit 2' (bu2;
Figs. 7
The overlying
'biofacies unit 2' (bu2;
Figs. 7 ![]() & 8
& 8 ![]() ) is characterized by an abrupt onset of echinoderm remains (probably mostly eocrinoids, ,
1985; Figs.
6-6
) is characterized by an abrupt onset of echinoderm remains (probably mostly eocrinoids, ,
1985; Figs.
6-6 ![]() & 7
& 7 ![]() ) which are dominant here. Trilobites and brachiopods are conspicuous accompanying faunal elements while sponge spicules are of minor importance. New (in comparison to bu1), but minor elements of the biota are chancelloriids, pelagiellids, and (rare) hyoliths. Of particular paleoecologic and biostratigraphic interest is the restriction in the occurrence of phosphatic small shelly fossils to this biofacies and stratigraphic level
(Figs. 6-10
) which are dominant here. Trilobites and brachiopods are conspicuous accompanying faunal elements while sponge spicules are of minor importance. New (in comparison to bu1), but minor elements of the biota are chancelloriids, pelagiellids, and (rare) hyoliths. Of particular paleoecologic and biostratigraphic interest is the restriction in the occurrence of phosphatic small shelly fossils to this biofacies and stratigraphic level
(Figs. 6-10 ![]() & 6-11
& 6-11 ![]() ).
).
![]() Transition into
'biofacies unit 3' (bu3;
Figs. 7
Transition into
'biofacies unit 3' (bu3;
Figs. 7 ![]() & 8
& 8 ![]() ) is gradual. Bu3 is characterized by about the same fossil groups as bu2, but trilobites dominate the fauna
(Fig. 6-7
) is gradual. Bu3 is characterized by about the same fossil groups as bu2, but trilobites dominate the fauna
(Fig. 6-7 ![]() ). Echinoderms are also important constituents, and brachiopods, poriferans, hyoliths, and chancelloriids are occasionally present. This faunal composition continues upward to the base of the Cabitza Formation,
but fewer and smaller fossils are present in the last few decimeters of the Campo Pisano Formation, designated here as
'biofacies unit 4' (bu4;
Figs. 7
). Echinoderms are also important constituents, and brachiopods, poriferans, hyoliths, and chancelloriids are occasionally present. This faunal composition continues upward to the base of the Cabitza Formation,
but fewer and smaller fossils are present in the last few decimeters of the Campo Pisano Formation, designated here as
'biofacies unit 4' (bu4;
Figs. 7 ![]() & 8
& 8 ![]() ).
).
![]() All sections of the Campo Pisano Formation show more-or-less the same general trends of biofacies described above, albeit with local minor variations in fossil content and distribution
(Fig. 7
All sections of the Campo Pisano Formation show more-or-less the same general trends of biofacies described above, albeit with local minor variations in fossil content and distribution
(Fig. 7 ![]() ). In several regions, e.g., Su Banatzu, the Terraseo region, the Mt. Scorra–Masua region, and at Mt. Serrau the content of fossils increases in the upper levels of bu3
(Fig. 7
). In several regions, e.g., Su Banatzu, the Terraseo region, the Mt. Scorra–Masua region, and at Mt. Serrau the content of fossils increases in the upper levels of bu3
(Fig. 7 ![]() ). At Gutturu Pala, sponge spicules are prominent and reach a second maximum at these same levels
(Fig. 7
). At Gutturu Pala, sponge spicules are prominent and reach a second maximum at these same levels
(Fig. 7 ![]() ).
Throughout the entire succession at Masua, poriferans are much more abundant than
they are at other localities.
).
Throughout the entire succession at Masua, poriferans are much more abundant than
they are at other localities.
![]() The fossil groups of the Campo Pisano Formation occupied a variety of habitats and display a number of feeding strategies. Completely infaunal forms are represented by hadimopanellids
(Fig. 6-10
The fossil groups of the Campo Pisano Formation occupied a variety of habitats and display a number of feeding strategies. Completely infaunal forms are represented by hadimopanellids
(Fig. 6-10 ![]() ), usually interpreted as palaeoscolecids, for which a deposit-feeding strategy is assumed (e.g., & ,
2001). Whether or not some of the agnostid trilobites were also deposit feeders cannot be determined with certainty. The lack of eyes and small size alone do not validate such an interpretation. Unquestionable infaunal trace fossils have not been found in the formation. Semi-infaunal forms, represented by hylolihs with a rounded cross-section are found only rarely in the Campo Pisano
Formation (, 2002).
), usually interpreted as palaeoscolecids, for which a deposit-feeding strategy is assumed (e.g., & ,
2001). Whether or not some of the agnostid trilobites were also deposit feeders cannot be determined with certainty. The lack of eyes and small size alone do not validate such an interpretation. Unquestionable infaunal trace fossils have not been found in the formation. Semi-infaunal forms, represented by hylolihs with a rounded cross-section are found only rarely in the Campo Pisano
Formation (, 2002).
![]() Most of the fauna in the Campo Pisano Formation is epibenthic. Sessile and semi-sessile epibenthic forms are represented by poriferans, chancelloriids, most of the echinoderms, and brachiopods (Figs.
6-8
Most of the fauna in the Campo Pisano Formation is epibenthic. Sessile and semi-sessile epibenthic forms are represented by poriferans, chancelloriids, most of the echinoderms, and brachiopods (Figs.
6-8 ![]() & 6-12
& 6-12 ![]() ). For hyolithelminths a secondary epibenthic mode of life is probable. Whereas the filter-feeding sponges ingested mainly very small food particles and bacteria, the other sessile epibenthos lived on suspended detritus and microorganisms. The infaunal and sessile-epifaunal organisms noted above all suggest a soft firm- to mud ground habitat. There is no evidence for hardgrounds.
). For hyolithelminths a secondary epibenthic mode of life is probable. Whereas the filter-feeding sponges ingested mainly very small food particles and bacteria, the other sessile epibenthos lived on suspended detritus and microorganisms. The infaunal and sessile-epifaunal organisms noted above all suggest a soft firm- to mud ground habitat. There is no evidence for hardgrounds.
![]() Mobile epibenthic biota are also common (most of the trilobites, some hyoliths, pelagiellids, bradoriids and some small shelly fossils such as cambroclaves; Figs. 6-9
Mobile epibenthic biota are also common (most of the trilobites, some hyoliths, pelagiellids, bradoriids and some small shelly fossils such as cambroclaves; Figs. 6-9 ![]() , 6-11
, 6-11 ![]() & 6-13
& 6-13 ![]() ). Some of the echinoderms too may have had only limited movement (cinctans, perhaps stylophorans, some edrioasteroids; ,
1992; , 1993; ,
1998; & ,
2001; & ,
2003; & ,
2005). A few of the epibenthos could have adopted a sporadic or nektobenthic life-style (some trilobites and bradoriids). All these groups are commonly viewed as low-level suspension feeders, but deposit feeding (e.g., some trilobites and possibly some carpoid echinoderms) or grazing (some molluscs) might have been feasible.
). Some of the echinoderms too may have had only limited movement (cinctans, perhaps stylophorans, some edrioasteroids; ,
1992; , 1993; ,
1998; & ,
2001; & ,
2003; & ,
2005). A few of the epibenthos could have adopted a sporadic or nektobenthic life-style (some trilobites and bradoriids). All these groups are commonly viewed as low-level suspension feeders, but deposit feeding (e.g., some trilobites and possibly some carpoid echinoderms) or grazing (some molluscs) might have been feasible.
![]() Direct indication of predation was not found. However, indirect evidence for this mode of feeding may be provided by the cambroclaves, of which the skeletal elements have been interpreted as the protective external armor of a mobile epibenthic animal ( & ,
1991). Exclusively nektic elements of the Campo Pisano Formation fauna are unknown (even though some animals may have had such behavior during their early ontogeny). Whether or not some of the biota were scavengers is not apparent. So far, the only representative of the primary producers identified is Girvanella (oscillatoriacean
cyanobacterium; & ,
1984).
Direct indication of predation was not found. However, indirect evidence for this mode of feeding may be provided by the cambroclaves, of which the skeletal elements have been interpreted as the protective external armor of a mobile epibenthic animal ( & ,
1991). Exclusively nektic elements of the Campo Pisano Formation fauna are unknown (even though some animals may have had such behavior during their early ontogeny). Whether or not some of the biota were scavengers is not apparent. So far, the only representative of the primary producers identified is Girvanella (oscillatoriacean
cyanobacterium; & ,
1984).
![]() The geographic (lateral) and stratigraphic (vertical) distribution of the biofacies units (bu1 to bu4) and the trophic and habitat characteristics of the fauna are fundamental to an interpretation of the depositional environment of the Campo Pisano Formation.
The geographic (lateral) and stratigraphic (vertical) distribution of the biofacies units (bu1 to bu4) and the trophic and habitat characteristics of the fauna are fundamental to an interpretation of the depositional environment of the Campo Pisano Formation.
![]() To summarize: in the 19 sections investigated the basal unit of the Campo Pisano Formation is overwhelmingly dominated by sessile epibenthic filter feeders (spiculite biofacies, bu1;
Figs. 7
To summarize: in the 19 sections investigated the basal unit of the Campo Pisano Formation is overwhelmingly dominated by sessile epibenthic filter feeders (spiculite biofacies, bu1;
Figs. 7 ![]() & 8
& 8 ![]() ). Sessile and mobile suspension feeders are rarely present in bu1. Upsection, a low-level suspension-feeder fauna becomes important (echinoderm biofacies, bu2;
Figs. 7
). Sessile and mobile suspension feeders are rarely present in bu1. Upsection, a low-level suspension-feeder fauna becomes important (echinoderm biofacies, bu2;
Figs. 7 ![]() & 8
& 8 ![]() ). The dominant fossils of bu2 are sessile forms (most echinoderms, brachiopods), but mobile elements are also common (trilobites). The proportion of poriferans decreases significantly. In the accompanying fauna, distinctive elements include a rare infaunal biota (hadimopanellids) and other fossils, which suggest adaptations for protection against predators (cambroclaves). The upper Campo Pisano Formation (bu3;
Figs. 7
). The dominant fossils of bu2 are sessile forms (most echinoderms, brachiopods), but mobile elements are also common (trilobites). The proportion of poriferans decreases significantly. In the accompanying fauna, distinctive elements include a rare infaunal biota (hadimopanellids) and other fossils, which suggest adaptations for protection against predators (cambroclaves). The upper Campo Pisano Formation (bu3;
Figs. 7 ![]() & 8
& 8 ![]() ) is characterized by mobile epibenthic suspension feeders and by an increase of probable nektic or planktic elements (greater proportion of agnostids). The proportion of sessile organisms decreases significantly. In the uppermost Campo Pisano Formation (bu4) an increase of transported elements (indicated by small broken bioclasts;
Figs. 7
) is characterized by mobile epibenthic suspension feeders and by an increase of probable nektic or planktic elements (greater proportion of agnostids). The proportion of sessile organisms decreases significantly. In the uppermost Campo Pisano Formation (bu4) an increase of transported elements (indicated by small broken bioclasts;
Figs. 7 ![]() & 8
& 8 ![]() ) is interpreted here as indicative of a more pelagic origin. Figure
8
) is interpreted here as indicative of a more pelagic origin. Figure
8 ![]() depicts this paleoecological succession of biofacies along with habitat characteristics, feeding strategies and bathymetry.
depicts this paleoecological succession of biofacies along with habitat characteristics, feeding strategies and bathymetry.
![]() The age of the Campo Pisano Formation was determined primarily by trilobites, but, because of their scarcity, the stratigraphic resolution obtained is low.
The age of the Campo Pisano Formation was determined primarily by trilobites, but, because of their scarcity, the stratigraphic resolution obtained is low.
![]() The assignment of the trilobites from the Campo Pisano Formation to the Middle Cambrian (,
1972) was refined by
(1991) and et alii
(1995) who identified two stratigraphically sequential assemblages. The oldest trilobite fauna of the Campo Pisano Formation (CP1 fauna of et alii,
1995), located several meters above the base of the formation, includes Protolenus (Protolenus) cf. pisidianus that indicates the Early to Middle Cambrian boundary interval (for discussion see et alii,
1995). The base of the Campo Pisano Formation therefore is in the uppermost Early Cambrian in the Mediterranean sense (for discussion of the problematic interregional correlation of the Early–Middle Cambrian boundary see & ,
2004), whereas the immediately overlying portion of the formation (early CP2 fauna of et alii,
1995) correlates with the Paradoxides mureroensis Zone (Leonian, early Middle Cambrian; early to middle Agdzian of the western Gondwana scale of & ,
2004;
Fig. 3
The assignment of the trilobites from the Campo Pisano Formation to the Middle Cambrian (,
1972) was refined by
(1991) and et alii
(1995) who identified two stratigraphically sequential assemblages. The oldest trilobite fauna of the Campo Pisano Formation (CP1 fauna of et alii,
1995), located several meters above the base of the formation, includes Protolenus (Protolenus) cf. pisidianus that indicates the Early to Middle Cambrian boundary interval (for discussion see et alii,
1995). The base of the Campo Pisano Formation therefore is in the uppermost Early Cambrian in the Mediterranean sense (for discussion of the problematic interregional correlation of the Early–Middle Cambrian boundary see & ,
2004), whereas the immediately overlying portion of the formation (early CP2 fauna of et alii,
1995) correlates with the Paradoxides mureroensis Zone (Leonian, early Middle Cambrian; early to middle Agdzian of the western Gondwana scale of & ,
2004;
Fig. 3 ![]() ). The middle portion of the Campo Pisano Formation (middle CP2 fauna of et alii,
1995) yields Calodiscus foveolatus, some oryctocepalids, dorypygids and others and corresponds to the Badulesia Substage of the mid-Middle Cambrian (early Caesaraugustan; et alii,
1995;
Fig. 3
). The middle portion of the Campo Pisano Formation (middle CP2 fauna of et alii,
1995) yields Calodiscus foveolatus, some oryctocepalids, dorypygids and others and corresponds to the Badulesia Substage of the mid-Middle Cambrian (early Caesaraugustan; et alii,
1995;
Fig. 3 ![]() ). The upper part of the Campo Pisano Formation (late CP2 fauna of et alii,
1995) contains, among others trilobites, Pardailhania hispida and Ptychagnostus sp., relating to the Pardailhania to basal Solenopleuropsis Substages (middle Caesaraugustan) of the mid- Middle Cambrian. The transition from the Campo Pisano Formation to the Cabitza Formation should thus be in the late Caesaraugustan (late Solenopleuropsis Substage of the late Middle Cambrian;
Fig. 3
). The upper part of the Campo Pisano Formation (late CP2 fauna of et alii,
1995) contains, among others trilobites, Pardailhania hispida and Ptychagnostus sp., relating to the Pardailhania to basal Solenopleuropsis Substages (middle Caesaraugustan) of the mid- Middle Cambrian. The transition from the Campo Pisano Formation to the Cabitza Formation should thus be in the late Caesaraugustan (late Solenopleuropsis Substage of the late Middle Cambrian;
Fig. 3 ![]() ).
).
![]() Non-trilobite age determinations are consistent with the trilobite biostratigraphy. Lingulid and acrotretid brachiopods from the upper Campo Pisano Formation (especially Linnarssonella sp.) imply a late Middle Cambrian age ( & ,
2004). Poriferans, chancelloriids, hyoliths (Microcornus), pelagiellids, and small shelly fossils (e.g., Hipponicharion) are in accordance with a late Early to Middle Cambrian age ( et alii,
1990; ,
2001; , 2002; & ,
2002; et alii, 2003).
Non-trilobite age determinations are consistent with the trilobite biostratigraphy. Lingulid and acrotretid brachiopods from the upper Campo Pisano Formation (especially Linnarssonella sp.) imply a late Middle Cambrian age ( & ,
2004). Poriferans, chancelloriids, hyoliths (Microcornus), pelagiellids, and small shelly fossils (e.g., Hipponicharion) are in accordance with a late Early to Middle Cambrian age ( et alii,
1990; ,
2001; , 2002; & ,
2002; et alii, 2003).
![]() Precise correlation of sections in the study area is hindered by a lack of trilobite material. However, on a regional scale this problem can be somewhat mitigated through the use of ecostratigraphic data. For example, in the Campo Pisano type section, the abrupt appearance of
'biofacies unit 2' (bu2) is the equivalent of the top of the Protolenus (Protolenus) cf. pisidianus level. This distinct change is interpreted as a regional environmental reorganization. For the study area, this means that the base of bu2 coincides with the base of the Middle Cambrian (in the Mediterranean sense) and that bu1 is latest Early Cambrian in age.
So the bu1–bu2 transition represents an ecostratigraphic level permitting recognition of the Early–Middle Cambrian boundary in southwestern Sardinia. Using this model, a correlation of the basal portions of most of the sections is possible although trilobites are absent. The validity of this technique has been confirmed recently by finding the Protolenus (Protolenus) cf. pisidianus index taxon in the Su Corovau section (, pers. comm., 2004;
Fig. 4
Precise correlation of sections in the study area is hindered by a lack of trilobite material. However, on a regional scale this problem can be somewhat mitigated through the use of ecostratigraphic data. For example, in the Campo Pisano type section, the abrupt appearance of
'biofacies unit 2' (bu2) is the equivalent of the top of the Protolenus (Protolenus) cf. pisidianus level. This distinct change is interpreted as a regional environmental reorganization. For the study area, this means that the base of bu2 coincides with the base of the Middle Cambrian (in the Mediterranean sense) and that bu1 is latest Early Cambrian in age.
So the bu1–bu2 transition represents an ecostratigraphic level permitting recognition of the Early–Middle Cambrian boundary in southwestern Sardinia. Using this model, a correlation of the basal portions of most of the sections is possible although trilobites are absent. The validity of this technique has been confirmed recently by finding the Protolenus (Protolenus) cf. pisidianus index taxon in the Su Corovau section (, pers. comm., 2004;
Fig. 4 ![]() ) at precisely the level predicted by ecostratigraphy.
) at precisely the level predicted by ecostratigraphy.
![]() The Campo Pisano trilobites have a western Gondwanan affinity
(Fig. 1
The Campo Pisano trilobites have a western Gondwanan affinity
(Fig. 1 ![]() ). Geographical relationships exist with southern France (Montagne Noire), northern Spain and southern Turkey, i.e. the Mediterranean region of the so-called "Atlantic Faunal Province" sensu
(1972) and (1994). Associations with the northern region of the "Atlantic Faunal Province" seem to be more distant. Biogeographic relations with the Far East as interpreted by
(1972), (1991) and et alii
(2003) for some Early Cambrian faunas have not yet been found among the trilobites of the Campo Pisano Formation. But, some small shelly fossils (cambroclaves) may indicate biogeographic relationships with the Asiatic realm. Until recently, cambroclaves were known only from Kazakhstan, China, and Australia. But, the latest finds from the late Early Cambrian of Germany (,
1994) and early Middle Cambrian of Sardinia ( & ,
2003), both on the western Gondwana shelf, represent the oldest and youngest representatives of this group known thus far and may have some paleogeographic significance. The mollusc Pelagiella subangulata shows a similar paleobiogeographic distribution along this "Palaeotethyan belt" (,
1989; , 2001; ,
2002). Small shelly fossils offer further support for close connections between the Mediterranean Cambrian areas: e.g., the bradoriid Hipponicharion is a good index taxon for the western Gondwanan late Early and early Middle Cambrian beds of Germany, Spain, Morocco, and Sardinia (,
1994; & ,
2002). In summary, a good faunal exchange among and connections between the western Gondwanan paleogeographic regions is clearly established.
). Geographical relationships exist with southern France (Montagne Noire), northern Spain and southern Turkey, i.e. the Mediterranean region of the so-called "Atlantic Faunal Province" sensu
(1972) and (1994). Associations with the northern region of the "Atlantic Faunal Province" seem to be more distant. Biogeographic relations with the Far East as interpreted by
(1972), (1991) and et alii
(2003) for some Early Cambrian faunas have not yet been found among the trilobites of the Campo Pisano Formation. But, some small shelly fossils (cambroclaves) may indicate biogeographic relationships with the Asiatic realm. Until recently, cambroclaves were known only from Kazakhstan, China, and Australia. But, the latest finds from the late Early Cambrian of Germany (,
1994) and early Middle Cambrian of Sardinia ( & ,
2003), both on the western Gondwana shelf, represent the oldest and youngest representatives of this group known thus far and may have some paleogeographic significance. The mollusc Pelagiella subangulata shows a similar paleobiogeographic distribution along this "Palaeotethyan belt" (,
1989; , 2001; ,
2002). Small shelly fossils offer further support for close connections between the Mediterranean Cambrian areas: e.g., the bradoriid Hipponicharion is a good index taxon for the western Gondwanan late Early and early Middle Cambrian beds of Germany, Spain, Morocco, and Sardinia (,
1994; & ,
2002). In summary, a good faunal exchange among and connections between the western Gondwanan paleogeographic regions is clearly established.
![]() Microfacies analysis of the Campo Pisano Formation demonstrates that many macroscopic features, e.g., color and nodularity, are of little value in such studies. After extensive field and microfacies investigations, the following characteristics were found to be essential for environmental interpretation: (1) microfacies, (2) geographic and vertical distribution of biota, (3) taxonomic variety, (4) non-biogenous components, (5) ichnology, (6) regional variation in stratigraphic thickness, and (7) the characteristics of underlying and overlying strata and of the formational limits.
Microfacies analysis of the Campo Pisano Formation demonstrates that many macroscopic features, e.g., color and nodularity, are of little value in such studies. After extensive field and microfacies investigations, the following characteristics were found to be essential for environmental interpretation: (1) microfacies, (2) geographic and vertical distribution of biota, (3) taxonomic variety, (4) non-biogenous components, (5) ichnology, (6) regional variation in stratigraphic thickness, and (7) the characteristics of underlying and overlying strata and of the formational limits.
![]() Within the study area, the thickness of the Campo Pisano Formation increases slightly from southeast to northwest as far as the San Benedetto to Su Banatzu line
(Figs. 4
Within the study area, the thickness of the Campo Pisano Formation increases slightly from southeast to northwest as far as the San Benedetto to Su Banatzu line
(Figs. 4 ![]() & 7
& 7 ![]() ). Thickness decreases northwest of this line. Because deposition was almost entirely autochthonous (see above), this region is assumed to be a carbonate depocenter – a weakly elevated, low-energy area – where conditions for carbonate precipitation were favorable
(Fig. 4
). Thickness decreases northwest of this line. Because deposition was almost entirely autochthonous (see above), this region is assumed to be a carbonate depocenter – a weakly elevated, low-energy area – where conditions for carbonate precipitation were favorable
(Fig. 4 ![]() ). The range in number and quantity of the autochthonous biota shows a similar distribution
(Fig. 7
). The range in number and quantity of the autochthonous biota shows a similar distribution
(Fig. 7 ![]() ), suggesting that this area was favorable to life and hence a preferred habitat. This view agrees well with the inference that it was a weak high. For a part of the Early Cambrian, a similar, but high-energy shallow area was proposed by et alii
(1985), but it was sited some 5–10 km to the west, coinciding approximately with the present southwestern Sardinian coastline. The weak rise during the Middle Cambrian may represent a later stage of this topographic elevation, relocated by a rise in relative sea-level (see below).
), suggesting that this area was favorable to life and hence a preferred habitat. This view agrees well with the inference that it was a weak high. For a part of the Early Cambrian, a similar, but high-energy shallow area was proposed by et alii
(1985), but it was sited some 5–10 km to the west, coinciding approximately with the present southwestern Sardinian coastline. The weak rise during the Middle Cambrian may represent a later stage of this topographic elevation, relocated by a rise in relative sea-level (see below).
![]() The abrupt arrival of the silty Campo Pisano Formation over the shallow, isolated carbonate platform of the Gonnesa Group
(Figs. 3
The abrupt arrival of the silty Campo Pisano Formation over the shallow, isolated carbonate platform of the Gonnesa Group
(Figs. 3 ![]() & 5-1
& 5-1 ![]() ) is indicated by its initial spiculite biofacies (bu1;
Figs. 6-5
) is indicated by its initial spiculite biofacies (bu1;
Figs. 6-5 ![]() & 7
& 7 ![]() ) in nearly every section, sometimes as a gradual transition, in other places associated with breccias (see ,
1987b; and the discussion of these breccias below). The sudden influx of siliciclastic material into the depositional area with the arrival of the Campo Pisano Formation is best interpreted as the aftermath of the completion of the fill of a sediment-trapping basin between an erosional area and the isolated carbonate platform, This is the interpretation of et alii
(1985) ("Eastern Sulcis basin"), that accounts for fine siliciclastic sediment reaching a formerly isolated depositional area. Suspension currents were presumably the primary agent for the transport of the very fine-grained siliciclastics of the Campo Pisano Formation. Infrequent storms may have occurred, but the hummocky cross-bedding, allochthonous shell layers, reworked and transported fauna, and extraclasts commonly produced by storms were not observed. These facts support the suggestion by previous authors that extensional tectonics combined with a relative rise in sea-level affected the area and led to the "drowning" of the platform (,
1987a, 1987b; et alii,
1988; & ,
1989; & ,
1990). The brief dominance of filter feeders (bu1;
Figs. 7
) in nearly every section, sometimes as a gradual transition, in other places associated with breccias (see ,
1987b; and the discussion of these breccias below). The sudden influx of siliciclastic material into the depositional area with the arrival of the Campo Pisano Formation is best interpreted as the aftermath of the completion of the fill of a sediment-trapping basin between an erosional area and the isolated carbonate platform, This is the interpretation of et alii
(1985) ("Eastern Sulcis basin"), that accounts for fine siliciclastic sediment reaching a formerly isolated depositional area. Suspension currents were presumably the primary agent for the transport of the very fine-grained siliciclastics of the Campo Pisano Formation. Infrequent storms may have occurred, but the hummocky cross-bedding, allochthonous shell layers, reworked and transported fauna, and extraclasts commonly produced by storms were not observed. These facts support the suggestion by previous authors that extensional tectonics combined with a relative rise in sea-level affected the area and led to the "drowning" of the platform (,
1987a, 1987b; et alii,
1988; & ,
1989; & ,
1990). The brief dominance of filter feeders (bu1;
Figs. 7 ![]() & 8
& 8 ![]() ) at the base of the Campo Pisano Formation reflects the successful adaptation of one group of organisms to the newly increased depth and to the sudden rise in the quantity of suspended material along with more influential water currents, and in addition the advantages gained by a rapid adaptation to new ecological niches.
) at the base of the Campo Pisano Formation reflects the successful adaptation of one group of organisms to the newly increased depth and to the sudden rise in the quantity of suspended material along with more influential water currents, and in addition the advantages gained by a rapid adaptation to new ecological niches.
![]() The transition from bu1 (spiculite biofacies) to bu2 (echinoderm biofacies) is a conspicuous change of milieu in the lower Campo Pisano Formation affecting the entire depositional area
(Figs. 7
The transition from bu1 (spiculite biofacies) to bu2 (echinoderm biofacies) is a conspicuous change of milieu in the lower Campo Pisano Formation affecting the entire depositional area
(Figs. 7 ![]() & 8
& 8 ![]() ). It is interpreted as reflecting an increase in water depth. The persistence and strong dominance of sessile epibenthic suspension feeders in this biofacies
(Fig. 8
). It is interpreted as reflecting an increase in water depth. The persistence and strong dominance of sessile epibenthic suspension feeders in this biofacies
(Fig. 8 ![]() ) suggest a permanent increase in the influx of suspended nutrients. Reasons for this change might include a reorganization in current patterns or a wetter climate that would intensify chemical erosion and sediment transport in the hinterland and thus an increase in the input of siliciclastic sediment. These changes may have been caused by a shift of this region to a higher paleogeographic latitude ( et alii,
1992; et alii,
1992; & ,
2001).
) suggest a permanent increase in the influx of suspended nutrients. Reasons for this change might include a reorganization in current patterns or a wetter climate that would intensify chemical erosion and sediment transport in the hinterland and thus an increase in the input of siliciclastic sediment. These changes may have been caused by a shift of this region to a higher paleogeographic latitude ( et alii,
1992; et alii,
1992; & ,
2001).
![]() The next change in the biofacies of the stratigraphic succession, from bu2 to bu3
(Figs. 7
The next change in the biofacies of the stratigraphic succession, from bu2 to bu3
(Figs. 7 ![]() & 8
& 8 ![]() ), took place without a conspicuous change in lithology; trilobites (especially agnostids) became more numerous, while the sessile epibenthos declined
(Fig. 8
), took place without a conspicuous change in lithology; trilobites (especially agnostids) became more numerous, while the sessile epibenthos declined
(Fig. 8 ![]() ). This change is best attributed to an ongoing, steady rise in relative sea-level.
). This change is best attributed to an ongoing, steady rise in relative sea-level.
![]() Near the lithological transition to the overlying siliciclastic Cabitza Formation
(Fig. 7
Near the lithological transition to the overlying siliciclastic Cabitza Formation
(Fig. 7 ![]() ), the environment changed again. At this level the fossil content as a whole decreases significantly and among the trilobites and brachiopods, only small, mostly broken parts remain (bu4); this fragmentation is assumed to have been caused by the action of currents that transported the fossils. The accompanying lithological change is interpreted as evidence of a second distinct eustatic pulse that caused the depositional environment to deepen
(Fig. 8
), the environment changed again. At this level the fossil content as a whole decreases significantly and among the trilobites and brachiopods, only small, mostly broken parts remain (bu4); this fragmentation is assumed to have been caused by the action of currents that transported the fossils. The accompanying lithological change is interpreted as evidence of a second distinct eustatic pulse that caused the depositional environment to deepen
(Fig. 8 ![]() ). However, it is improbable that the depth of water exceeded that of an outer shelf because of the general persistence of similar lithofacies in overlying strata. The existence of deeper conditions may not be taken as clearly expressed lithologically until a fully siliciclastic regime was established in the Cabitza Formation. et alii
(1995) have demonstrated that younger portions of the Cabitza Formation were deposited in relatively shallow waters near storm wave base. This supports the view that the uppermost Campo
Pisano and the lower Cabitza Formations were not much deeper than the overlying strata, because extreme fluctuations in bathymetry in such a short time are unrealistic. Hence, the rather rapid lithological change from the carbonates of the Campo Pisano Formation to the siliciclastic sediment of the Cabitza Formation probably does not indicate a dramatic increase in water depth, but rather reflects a drift to a higher latitude (cooling and increased precipitation).
). However, it is improbable that the depth of water exceeded that of an outer shelf because of the general persistence of similar lithofacies in overlying strata. The existence of deeper conditions may not be taken as clearly expressed lithologically until a fully siliciclastic regime was established in the Cabitza Formation. et alii
(1995) have demonstrated that younger portions of the Cabitza Formation were deposited in relatively shallow waters near storm wave base. This supports the view that the uppermost Campo
Pisano and the lower Cabitza Formations were not much deeper than the overlying strata, because extreme fluctuations in bathymetry in such a short time are unrealistic. Hence, the rather rapid lithological change from the carbonates of the Campo Pisano Formation to the siliciclastic sediment of the Cabitza Formation probably does not indicate a dramatic increase in water depth, but rather reflects a drift to a higher latitude (cooling and increased precipitation).
![]() The interpretation of the depositional environments of the Campo Pisano Formation used in previous models was based on facies and sedimentological studies (,
1979, 1987a, 1987b; et alii,
1988; & ,
1990; and others). The resulting model for the Campo Pisano limestone was a type of nodular deep water facies ("ammonitico rosso" lithofacies) deposited on pelagic topographic highs on a broken, drowned platform, while troughs between these highs would serve as areas of deposition for calcareous clays and silt. Nevertheless, judging from the microfacies and paleoecological results presented here, this model must be modified. In the 19 studied sections
(Fig. 4A
The interpretation of the depositional environments of the Campo Pisano Formation used in previous models was based on facies and sedimentological studies (,
1979, 1987a, 1987b; et alii,
1988; & ,
1990; and others). The resulting model for the Campo Pisano limestone was a type of nodular deep water facies ("ammonitico rosso" lithofacies) deposited on pelagic topographic highs on a broken, drowned platform, while troughs between these highs would serve as areas of deposition for calcareous clays and silt. Nevertheless, judging from the microfacies and paleoecological results presented here, this model must be modified. In the 19 studied sections
(Fig. 4A ![]() ), nodular, wavy-bedded, silty limestones, and massive carbonate banks alternate repeatedly and irregularly
(Figs. 5-3
), nodular, wavy-bedded, silty limestones, and massive carbonate banks alternate repeatedly and irregularly
(Figs. 5-3 ![]() ,
5-4
,
5-4 ![]() ,
5-5
,
5-5 ![]() ,
5-6
,
5-6 ![]() ,
5-7
,
5-7 ![]() ,
5-8
,
5-8 ![]() & 5-9
& 5-9 ![]() ). Closely adjacent sections do not show the same patterns of vertical distribution for these lithotypes, so that their sequence cannot be used for correlation. Furthermore, biofacies do not correlate with these lithofacies. In the discrete more or less widely separated sections although there is no difference in the sequential position of the zones established by the autochthonous fossils, this fossil zonation has no relationship to the lithology of the approximately corresponding stratigraphic levels: that is whether or not the same fossils (qualitatively and quantitatively) are enclosed in nodular, wavy-bedded, silty or massive limestones. The origin of these textural types is related to diagenetic processes (,
1987b; , 2001) that have accentuated minor differences in the lithology of the primary sediment (e.g., siliciclastic content, permeability), and early
cementation; the direction and velocity of minor currents may also be involved. The relative uniformity of biofacies over the entire are of study demonstrates that during deposition of the Campo Pisano Formation bottom conditions that could have caused ecological differences were almost completely absent. However, a slight difference in bottom topography may have led to small regional differences in siliciclastic content (an increase to the East) as reported by et alii
(1988). Semi-quantitative analysis of sections from several localities found quartz content to be below 2 % that
(2003) regarded as probably aeolian. Nevertheless, this weak eastward increase in siliciclastic content in the Campo Pisano limestones may support the interpretation of et alii
(1988), that a hinterland lay to the East, in sharp contrast to the views of et alii
(1987), who postulates a basin that direction.
). Closely adjacent sections do not show the same patterns of vertical distribution for these lithotypes, so that their sequence cannot be used for correlation. Furthermore, biofacies do not correlate with these lithofacies. In the discrete more or less widely separated sections although there is no difference in the sequential position of the zones established by the autochthonous fossils, this fossil zonation has no relationship to the lithology of the approximately corresponding stratigraphic levels: that is whether or not the same fossils (qualitatively and quantitatively) are enclosed in nodular, wavy-bedded, silty or massive limestones. The origin of these textural types is related to diagenetic processes (,
1987b; , 2001) that have accentuated minor differences in the lithology of the primary sediment (e.g., siliciclastic content, permeability), and early
cementation; the direction and velocity of minor currents may also be involved. The relative uniformity of biofacies over the entire are of study demonstrates that during deposition of the Campo Pisano Formation bottom conditions that could have caused ecological differences were almost completely absent. However, a slight difference in bottom topography may have led to small regional differences in siliciclastic content (an increase to the East) as reported by et alii
(1988). Semi-quantitative analysis of sections from several localities found quartz content to be below 2 % that
(2003) regarded as probably aeolian. Nevertheless, this weak eastward increase in siliciclastic content in the Campo Pisano limestones may support the interpretation of et alii
(1988), that a hinterland lay to the East, in sharp contrast to the views of et alii
(1987), who postulates a basin that direction.
![]() Breccias exist in the beds marking the changeover from the Gonnesa platform to the Campo Pisano offshore carbonates ( et alii,
1981; , 1987a,
1987b; et alii,
1988). These breccias
(Fig. 5-2
Breccias exist in the beds marking the changeover from the Gonnesa platform to the Campo Pisano offshore carbonates ( et alii,
1981; , 1987a,
1987b; et alii,
1988). These breccias
(Fig. 5-2 ![]() ) have usually been interpreted as due to tensional tectonics and/or karstification.
(1987b) described two discrete types: (a) breccias consisting of well sorted clasts (up to 20 cm), and (b) unsorted megabreccias. The first type (a – Mt. Agruxiau type) interpreted as karst-related with a red hematitic and/or marly matrix sometimes yielding Campo Pisano fossils. The second (b – Mt. San Giovanni type) was called a collapse breccia with a marly matrix, and lacking Campo Pisano Formation clasts. A karstic origin for these breccias was doubted by et alii
(1988). For some authors these breccias indicate a profound relief on the sea floor ( et alii,
1984; , 1985; ,
1987b; et alii,
1988; & ,
1990). However, significant relief with highs and lows would promote distinct differentiation in the biofacies of the autochthonous fauna. But such differentiation does not exist. Biotic assemblages occur at the same stratigraphic levels over the whole area, and there are no marked lateral differences in their composition. The characteristics of the breccias mentioned above (clast content and shape, matrix, thickness, stratigraphic position) are easily explained as typical of fault-related internal brecciation
(Fig. 9
) have usually been interpreted as due to tensional tectonics and/or karstification.
(1987b) described two discrete types: (a) breccias consisting of well sorted clasts (up to 20 cm), and (b) unsorted megabreccias. The first type (a – Mt. Agruxiau type) interpreted as karst-related with a red hematitic and/or marly matrix sometimes yielding Campo Pisano fossils. The second (b – Mt. San Giovanni type) was called a collapse breccia with a marly matrix, and lacking Campo Pisano Formation clasts. A karstic origin for these breccias was doubted by et alii
(1988). For some authors these breccias indicate a profound relief on the sea floor ( et alii,
1984; , 1985; ,
1987b; et alii,
1988; & ,
1990). However, significant relief with highs and lows would promote distinct differentiation in the biofacies of the autochthonous fauna. But such differentiation does not exist. Biotic assemblages occur at the same stratigraphic levels over the whole area, and there are no marked lateral differences in their composition. The characteristics of the breccias mentioned above (clast content and shape, matrix, thickness, stratigraphic position) are easily explained as typical of fault-related internal brecciation
(Fig. 9 ![]() ). Breccias of this origin are typically monomictic (or nearly monomictic) and part of their marl matrix may be derived from overlying strata, so can contain fossils from overlying beds injected with the marls into the still unconsolidated internal breccias below (,
1988). Breccias of this kind originate beneath a sedimentary cover that is under a tensional tectonic regime ( & ,
1983; , 1988). et alii
(1990) observed transitions from the Sardinian breccias in the upper levels of the Gonnesa platform into unbrecciated rocks below, a phenomenon characteristic of such an origin ,
1988). et alii
(1990) were the first to interpret the breccias at the Gonnesa–Campo Pisano transition as flexure-related rocks caused by tension. A reinterpretation of lithological features, taking into consideration regional tectonics (e.g., ,
1987b; & ,
1989; & ,
1990), and paleoecologic data presented here indicate that when these breccias formed the morphologic gradient was rather low (in contrast to the gradient inferred for other types of breccias and debris flows that developed on the platform slopes during older Gonnesa time, et alii,
1988). If the gradient was low, a horst-and-graben environment for the breccias of Campo Pisano time cannot be sustained
(Fig. 9
). Breccias of this origin are typically monomictic (or nearly monomictic) and part of their marl matrix may be derived from overlying strata, so can contain fossils from overlying beds injected with the marls into the still unconsolidated internal breccias below (,
1988). Breccias of this kind originate beneath a sedimentary cover that is under a tensional tectonic regime ( & ,
1983; , 1988). et alii
(1990) observed transitions from the Sardinian breccias in the upper levels of the Gonnesa platform into unbrecciated rocks below, a phenomenon characteristic of such an origin ,
1988). et alii
(1990) were the first to interpret the breccias at the Gonnesa–Campo Pisano transition as flexure-related rocks caused by tension. A reinterpretation of lithological features, taking into consideration regional tectonics (e.g., ,
1987b; & ,
1989; & ,
1990), and paleoecologic data presented here indicate that when these breccias formed the morphologic gradient was rather low (in contrast to the gradient inferred for other types of breccias and debris flows that developed on the platform slopes during older Gonnesa time, et alii,
1988). If the gradient was low, a horst-and-graben environment for the breccias of Campo Pisano time cannot be sustained
(Fig. 9 ![]() ).
).
![]() There are two opinions regarding the depth of water under which the Campo Pisano Formation was deposited. The fossil content has been interpreted as a neritic shelf fauna, but the nodular texture of the carbonates has been thought to indicate depth up to several hundreds of meters (e.g., ,
1985; , 1985). Shallower conditions were assumed by
(1979, "shallow water"). The Campo Pisano carbonates are typical Cambrian offshore sediments, deposited in the photic zone below storm wave base
(Figs. 8
There are two opinions regarding the depth of water under which the Campo Pisano Formation was deposited. The fossil content has been interpreted as a neritic shelf fauna, but the nodular texture of the carbonates has been thought to indicate depth up to several hundreds of meters (e.g., ,
1985; , 1985). Shallower conditions were assumed by
(1979, "shallow water"). The Campo Pisano carbonates are typical Cambrian offshore sediments, deposited in the photic zone below storm wave base
(Figs. 8 ![]() & 9
& 9 ![]() ). No indicators of wave and tidal activity, erosion, transport of clasts from shallower areas, or gradation, or hummocky bedding was observed. Rarely, thin allochthonous shell layers occur (, , pers. comm.); possibly they are indicative of storms. All these features are typical in distal ramp or shelf environments, which, in the Cambrian are usually characterized paleoecologically by a dominance of echinoderms and sponges and by intensive bio-retexturing (,
2002). The source of carbonate mud in such environments could be (1) adjacent shallow platforms (if allochthonous), or (2) biotic precipitation by planktic microbial organisms (if autochthonous) or (3) a mixture of both (,
1986; et alii,
1995; ,
2002). An autochthonous microbial origin is favoured for the Campo Pisano carbonates. No shallow platform remnants "able to deliver carbonate ooze are known from that time" & ,
1989). Furthermore, the high mud content points to a rather low slope on the sea floor, a conclusion compatible with the interpretation from fossils of a shelf/ramp environment.
). No indicators of wave and tidal activity, erosion, transport of clasts from shallower areas, or gradation, or hummocky bedding was observed. Rarely, thin allochthonous shell layers occur (, , pers. comm.); possibly they are indicative of storms. All these features are typical in distal ramp or shelf environments, which, in the Cambrian are usually characterized paleoecologically by a dominance of echinoderms and sponges and by intensive bio-retexturing (,
2002). The source of carbonate mud in such environments could be (1) adjacent shallow platforms (if allochthonous), or (2) biotic precipitation by planktic microbial organisms (if autochthonous) or (3) a mixture of both (,
1986; et alii,
1995; ,
2002). An autochthonous microbial origin is favoured for the Campo Pisano carbonates. No shallow platform remnants "able to deliver carbonate ooze are known from that time" & ,
1989). Furthermore, the high mud content points to a rather low slope on the sea floor, a conclusion compatible with the interpretation from fossils of a shelf/ramp environment.
![]() Although "neritic" conditions are strongly indicated by the fauna, by the lithology ( et alii,
1995), and by the context of syntectonic evolution ( & ,
1989; et alii,
1987), absolute water depths are hard to calculate because of limited knowledge concerning rates of subsidence, sediment budgets and the range of rises in sea-level. Nevertheless, because of (1) the shallow subtidal to intertidal conditions indicated for the underlying upper Gonnesa Group ( et alii,
1985; et alii,
1987; , 1994), (2) the conclusions of &
(1990) and et alii
(1995), that the overlying lower Cabitza Formation indicates a deeper shelf, and (3) the two regional well-defined paleoecologic changes in the Campo Pisano Formation indicate a two-step deepening of the whole area during Campo Pisano time
(Fig. 8
Although "neritic" conditions are strongly indicated by the fauna, by the lithology ( et alii,
1995), and by the context of syntectonic evolution ( & ,
1989; et alii,
1987), absolute water depths are hard to calculate because of limited knowledge concerning rates of subsidence, sediment budgets and the range of rises in sea-level. Nevertheless, because of (1) the shallow subtidal to intertidal conditions indicated for the underlying upper Gonnesa Group ( et alii,
1985; et alii,
1987; , 1994), (2) the conclusions of &
(1990) and et alii
(1995), that the overlying lower Cabitza Formation indicates a deeper shelf, and (3) the two regional well-defined paleoecologic changes in the Campo Pisano Formation indicate a two-step deepening of the whole area during Campo Pisano time
(Fig. 8 ![]() ).Consequently, it is with considerable confidence that the siliciclastic lower Cabitza Formation is assigned a depth below the photic zone and the Campo Pisano Formation itself is considered to have recorded an intermittent deepening of the sea within the photic zone
(Figs. 8
).Consequently, it is with considerable confidence that the siliciclastic lower Cabitza Formation is assigned a depth below the photic zone and the Campo Pisano Formation itself is considered to have recorded an intermittent deepening of the sea within the photic zone
(Figs. 8 ![]() & 9
& 9 ![]() ).
).![]()
![]() The late Early to late Middle Cambrian Campo Pisano Formation of southwestern Sardinia is represented by a succession of nodular, wavy-bedded, silty, and massive offshore limestones
(Fig. 5
The late Early to late Middle Cambrian Campo Pisano Formation of southwestern Sardinia is represented by a succession of nodular, wavy-bedded, silty, and massive offshore limestones
(Fig. 5 ![]() ) deposited during the drowning of the isolated Sardinian Early Cambrian carbonate platform. After a distinct rise in relative sea-level that caused the isolated carbonate platform conditions of the underlying Gonnesa Group to end
(Fig. 9
) deposited during the drowning of the isolated Sardinian Early Cambrian carbonate platform. After a distinct rise in relative sea-level that caused the isolated carbonate platform conditions of the underlying Gonnesa Group to end
(Fig. 9 ![]() ), but with water-depth still relatively shallow, the initial Campo Pisano fauna developed with epibenthic filter feeders (sponges) dominant and with a moderate amount of suspended input from the hinterland
(Fig. 7
), but with water-depth still relatively shallow, the initial Campo Pisano fauna developed with epibenthic filter feeders (sponges) dominant and with a moderate amount of suspended input from the hinterland
(Fig. 7 ![]() ). This fossil community predominated in the lower beds of the Campo Pisano Formation in nearly all of the 19 sections mapped.
). This fossil community predominated in the lower beds of the Campo Pisano Formation in nearly all of the 19 sections mapped.
![]() Higher in the succession poriferans persist as an important element of the fauna only in some westernmost areas. Upsection, echinoderms come to dominate the biofacies (mainly sessile epibenthic eocrinoids and other low-level echinoderm suspension feeders) and they in turn are succeeded by a trilobite-dominated biofacies in which the mobile epibenthos becomes increasingly important upward as the depth of water increases
(Figs. 7
Higher in the succession poriferans persist as an important element of the fauna only in some westernmost areas. Upsection, echinoderms come to dominate the biofacies (mainly sessile epibenthic eocrinoids and other low-level echinoderm suspension feeders) and they in turn are succeeded by a trilobite-dominated biofacies in which the mobile epibenthos becomes increasingly important upward as the depth of water increases
(Figs. 7 ![]() & 8
& 8 ![]() ). Near the boundary with the deeper water Cabitza Formation an increase in the strength of currents is suggested by the fact that the fossils are all fragmentary and have been transported. This trend to less proximal and deeper-water sedimentation is apparent over the entire working area for it is expressed in both stratigraphy and paleoecology. Regional paleoecologic trends
(Figs. 4
). Near the boundary with the deeper water Cabitza Formation an increase in the strength of currents is suggested by the fact that the fossils are all fragmentary and have been transported. This trend to less proximal and deeper-water sedimentation is apparent over the entire working area for it is expressed in both stratigraphy and paleoecology. Regional paleoecologic trends
(Figs. 4 ![]() & 7
& 7 ![]() ) are areally extensive and not pronounced, so they are thought to have been caused by a rise in sea level rather than by tensional block-tectonics and fracturing on the sea floor. The data are interpreted as indicating the existence of a weak rise (low-energy swell) trending SE–NW that served as a carbonate depocenter and is thought to have been a preferred habitat for the Campo Pisano biota
(Fig. 4B
) are areally extensive and not pronounced, so they are thought to have been caused by a rise in sea level rather than by tensional block-tectonics and fracturing on the sea floor. The data are interpreted as indicating the existence of a weak rise (low-energy swell) trending SE–NW that served as a carbonate depocenter and is thought to have been a preferred habitat for the Campo Pisano biota
(Fig. 4B ![]() ). There is no evidence of a high-energy shallow area anywhere. The breccias are classed as "internal" that is formed below a cover of sediments while tensional tectonics were active
(Fig. 9
). There is no evidence of a high-energy shallow area anywhere. The breccias are classed as "internal" that is formed below a cover of sediments while tensional tectonics were active
(Fig. 9 ![]() ).
).
![]() The biofacies and the sedimentary criteria: thickness, mud-, fossil-, and clastic content indicate that the area gradually drowned as one unit; it was undifferentiated bathymetrically
(Figs. 8
The biofacies and the sedimentary criteria: thickness, mud-, fossil-, and clastic content indicate that the area gradually drowned as one unit; it was undifferentiated bathymetrically
(Figs. 8 ![]() & 9
& 9 ![]() ). It is classed as a neritic open-marine shelf or ramp. An intermittent rise of relative sea-level, accompanied by a tensional tectonic regime, along with a move into higher paleogeographic latitudes, led to a regional uniformity in facies. The sudden rise of relative sea-level at the transition from the Gonnesa platform to the Campo Pisano offshore carbonates caused the entire area to be under about the same depth of water
(Fig. 8
). It is classed as a neritic open-marine shelf or ramp. An intermittent rise of relative sea-level, accompanied by a tensional tectonic regime, along with a move into higher paleogeographic latitudes, led to a regional uniformity in facies. The sudden rise of relative sea-level at the transition from the Gonnesa platform to the Campo Pisano offshore carbonates caused the entire area to be under about the same depth of water
(Fig. 8 ![]() ). Great morphological differences within the area, caused by tectonism during the isolated platform stage (Gonnesa Group), are interpreted here as having been leveled by Campo Pisano times through relief-filling redeposition during late Gonnesa times and by offshore sedimentation during early Campo Pisano times
(Fig. 9
). Great morphological differences within the area, caused by tectonism during the isolated platform stage (Gonnesa Group), are interpreted here as having been leveled by Campo Pisano times through relief-filling redeposition during late Gonnesa times and by offshore sedimentation during early Campo Pisano times
(Fig. 9 ![]() ). So the model presented here resolves seeming contradictions between sedimentologic and paleontologic criteria.
). So the model presented here resolves seeming contradictions between sedimentologic and paleontologic criteria.
![]() The techniques of micropaleoecological analysis presented here are a very useful tool for modeling the configuration and evolution of Perigondwanan depositional areas during the Cambrian. The method can be successfully applied to the reconstruction of regional geological processes for that time, in particular where biostratigraphic data and sedimentary manifestations are fragmentary or unavailable.
The techniques of micropaleoecological analysis presented here are a very useful tool for modeling the configuration and evolution of Perigondwanan depositional areas during the Cambrian. The method can be successfully applied to the reconstruction of regional geological processes for that time, in particular where biostratigraphic data and sedimentary manifestations are fragmentary or unavailable.
![]() I express my warmest thanks to Thilo (Heidelberg, Germany), Maria (Naples, Italy), and Anna (Siena, Italy) for encouragement, discussions, and joint field trips. Thanks also to Gian Luigi (Cagliari, Sardinia) and Michal (Plzeň, Czech Republic) for collaboration on special fossil groups and to Petra (Regensburg, Germany) and Yvonne (Leipzig, Germany) for analytic work. I am very grateful to Pierre (Darwin, Australia), Bosiljka (Northampton, Massachusetts), James F. (Springfield, Missouri), Paul (Sudbury, Canada), and Nestor (Modesto, California) for important remarks on the manuscript and for linguistic help. Jan and Michael (both Freiberg) are gratefully thanked for technical help. The project was generously supported by the German Research Foundation (project EL 144/1
'Comparative studies of facies and paleoecology of the final stage of a Cambrian platform evolution: the small shelly fauna of the Campo Pisano
Fm, SW-Sardinia').
I express my warmest thanks to Thilo (Heidelberg, Germany), Maria (Naples, Italy), and Anna (Siena, Italy) for encouragement, discussions, and joint field trips. Thanks also to Gian Luigi (Cagliari, Sardinia) and Michal (Plzeň, Czech Republic) for collaboration on special fossil groups and to Petra (Regensburg, Germany) and Yvonne (Leipzig, Germany) for analytic work. I am very grateful to Pierre (Darwin, Australia), Bosiljka (Northampton, Massachusetts), James F. (Springfield, Missouri), Paul (Sudbury, Canada), and Nestor (Modesto, California) for important remarks on the manuscript and for linguistic help. Jan and Michael (both Freiberg) are gratefully thanked for technical help. The project was generously supported by the German Research Foundation (project EL 144/1
'Comparative studies of facies and paleoecology of the final stage of a Cambrian platform evolution: the small shelly fauna of the Campo Pisano
Fm, SW-Sardinia').
![]()
J.J., O., G., A.W.A. & J.H. (2003).– Palaeogeographical controls on the Cambrian trilobite immigration and evolutionary patterns reported in the western Gondwana margin.– Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, v. 195, p. 5–35.
A., T. & M. (1990).– Stratabound ore deposits related to synsedimentary tectonics: South-west Sardinia (Italy) and Sierra de Aracena (Spain); a comparison.– Geologische Rundschau, Berlin–Heidelberg, v. 79/2, p. 373–386.
M., D.W.J. & D. (1995).– Carbonate ramp depositional systems from late Jurassic epeiric platform (Iberian Basin, Spain): a combined computer modelling and outcrop analysis.– Sedimentology, Oxford, v. 42, p. 75–94.
T. & M. (1989).– Tectonic control on the formation of a carbonate platform: the Cambrian of southwestern Sardinia. In: P.D., J.L., J.F. & R.F. (eds.), Controls on carbonate platform and basin development.– Society of Economic Paleontologists and Mineralogists Special Publications, Tulsa, v. 44, p. 107–122.
T. & M. (1994).– Sedimentological, stratigraphical and ore deposits field guide of the autochthonous Cambro–Ordovician of southwestern Sardinia.– Memoire descrittive della carta geologica d'Italia, Servizio Geologico Nazionale, Roma, v. XLVIII, 434 p.
T., M. & M. (1985).– The Lower Cambrian of SW-Sardinia: from a clastic tidal shelf to an isolated carbonate platform.– Facies, Erlangen, v. 12, p. 113–140.
T., T. & M. (1988).– Rise and fall of an isolated, unstable carbonate platform: the Cambrian of Southwestern Sardinia.– Geologische Rundschau, Berlin–Heidelberg, v. 77, p. 389–416.
T., , M. & R. (1994).– The Sardic unconformity. In: T. & M. (eds.), Sedimentological, stratigraphical and ore deposits field guide of the autochthonous Cambro–Ordovician of southwestern Sardinia.– Memoire descrittive della carta geologica d'Italia.– Servizio Geologico Nazionale, Roma, v. XLVIII, p. 109–114.
F. (1974).– Geology of Jordan.– Gebrüder Borntraeger, Stuttgart, 196 p.
S. & V.V. (1981).– Coeloscleritophora – a major group of enigmatic Cambrian metazoans. In: M.E. (ed.), Short papers for the second international symposium on the Cambrian system.– United States Geological Survey Open-File Report, v. 81-743, p. 19–21.
S., S., B.J., P.A. & B.N. (1990).– Early Cambrian fossils from South Australia.– Association of Australasian Palaeontologists, Brisbane, v. Memoir 9, p. 186–190.
M., T., A. & G. (1981).– Tettonica, sedimentazione e mineralizzazioni delle brecce al bordo sud-orientale della piattaforma carbonatica Cambrica (Sulcis, Sardegna).– Memorie della Società Geologica Italiana, Roma, v. 22, p. 111–122.
J.G. (1886).– Die Versteinerungen des Cambrischen Schichtensystems der Insel Sardinien – Erste Abteilung.– Nova Acta der Kaiserlichen Leopoldinisch-Carolinischen Deutschen Akademie der Naturforscher, Halle, v. 51, p. 3–149.
J.G. (1891).– Die Versteinerungen des Cambrischen Schichtensystems der Insel Sardinien – Zweite Abteilung.– Nova Acta der Kaiserlichen Leopoldinisch-Carolinischen Deutschen Akademie der Naturforscher, Halle, v. 56, p. 427–529.
M.D. (1989).– China and the Palaeotethyan Belt (India, Pakistan, Iran, Kazakhstan, and Mongolia). In: J.W. & M.D. (eds.), The Precambrian–Cambrian boundary.– Oxford Monographs on Geology and Geophysics, Oxford, v. 12, p. 40–74.
G., T. & B. (1984).– Late Precambrian–Cambrian geodynamic setting and tectono-sedimentary evolution of Sardinia (Italy).– Bolletino della Società Geologica Italiana, Roma, v. 103, p. 121–128.
A. & R. (1984).– Middle Cambrian foraminifera and other microfossils from SW Sardinia.– Bolletino della Società Paleontologica Italiana, Modena, v. 23, p. 149–160.
S. & A.B. (2005).– Palaeoanatomy and biological affinities of a Cambrian deuterostome (Stylophora).– Nature, London, v. 438, p. 351–354.
T. (1979).– The Cambrian of Sardinia.– Memoire della Società Geologica Italiana, Roma, v. 20, p. 163–187.
T. & A. (1990).– Carbonate deposition during early rifting: the Cambrian of Sardinia and the Triassic-Jurassic of Tuscany, Italy. In: M.E., J.L., P.D., J.R. & , J.F. (eds.), Carbonate platforms-facies, sequences and evolution.– International Association of Sedimentologists Special Publications, v. 9, p. 9–37.
S. & M. (1991).– Cambroclaves and Paracarinachitids, early skeletal problematica from the Lower Cambrian of south China.– Palaeontology, London, v. 34, p. 357–397.
P., F. & A. (1992).– Palaeogeographic and geodynamic evolution of the Gondwana continental margins during the Cambrian.– Terra Nova, Oxford, v. 4, p. 657–667.
F. (1964).– Archaeocyatha – contribution à l'étude des faunes cambriennes du Maroc, de Sardaigne et de France.– Notes et Memoires du Service géologique du Maroc, Rabat, v. 179, 371 p.
O. (1994).– Lower Cambrian carbonates from eastern Germany: palaeontology, stratigraphy and palaeogeography.– Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, v. 191, p. 69–93.
O. (1997).– Biostratigraphic data of the German Cambrian – present state of knowledge.– Freiberger Forschungshefte – psf, Freiberg, v. C466, p. 155–165.
O. (2000).– Die kambrische "Görlitz-Fauna": Charakteristik und Bedeutung für die stratigraphische und paläogeographische Rekonstruktion Mitteleuropas.– Zeitschrift für geologische Wissenschaften, Berlin, v. 28, p. 11–32.
O. (2001).– Fazies und Genese kambrischer Knollenkalke des nördlichen Iglesiente (Gutturu Pala, SW-Sardinien, Italien).– Zentralblatt für Geologie und Paläontologie, Stuttgart, v. 1–2, p. 33–54.
O. (2002).– First record of Cambrian pelagiellids from Sardinia.– Freiberger Forschungshefte – psf, Freiberg, v. C497, p. 19–27.
O. & G.L. (2004).– Cambrian microfauna and palaeoecology of the Campo Pisano Formation at Gutturu Pala (Iglesiente, SW Sardinia, Italy).– Bolletino della Società Paleontologica Italiana, Modena, v. 43, p. 383–401.
O. & T. (2003).– Cambroclaves from the Cambrian of Sardinia (Italy) and Germany: constraints for the architecture of western Gondwana and the palaeogeographical and paleoecological potential of cambroclaves.– Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, v. 195, p. 55–71.
O., Y. & P. (2003).– Biofazies, Paläoökologie und Ablagerungsmodell des Finalstadiums einer kambrischen Karbonatplattform-Entwicklung: die Fauna der Campo Pisano Formation SW-Sardiniens.– Freiberger Forschungshefte – psf, Freiberg, v. C499, p. 1–33.
W.-P. (1993).– Systematik und Funktionsmorphologie mittelkambrischer Cincta (Carpoidea, Echinodermata).– Beringeria, Würzburg, v. 7, p. 3–190.
M. (1994).– Entwicklung der unterkambrischen Gonnesa-Carbonatplattform: Fazies und synsedimentäre Tektonik der Punta Manna-Formation und der Santa Barbara-Formation im westlichen Iglesiente (Südwest-Sardinien / Italien).– Freiburger geowissenschaftliche Beiträge, Freiburg, v. 7, 225 p.
H. (ed.) (1988).– Sediment-Petrologie Teil II: Sedimente und Sedimentgesteine.– E. Schweizerbart'sche Verlagsbuchhandlung (Nägele u. Obermiller), Stuttgart, 1141 p.
H. & D.K. (1983).– Relations between submarine fissures, internal breccias and mass flows during Triassic and earlier rifting periods.– Geologische Rundschau, Berlin–Heidelberg, v. 72, p. 53–66.
A. (1979).– Analisi sedimentologica preliminare della parte superiore della formazione di Gonnesa e della parte inferiore della formazione di Cabitza (Cambriano inferiore e medio della Sardegna).– Memoire della Società Geologica Italiana, Roma, v. 20, p. 395–404.
A. (1987a).– Anomalies at the transition between Gonnesa Fm and Cabitza Fm (Lower to Middle Cambrian; southwestern Sardinia).– IGCP Project n° 5, Newsletters, Padova–Bordeaux, v. 7, p. 52–53.
A. (1987b).– Depositional and paleogeographic evolution of the Cambrian in South-Western Sardinia.– IGCP Project n° 5, Newsletters, Padova–Bordeaux, v. 7, p. 151–166.
A., N., F. & P. (1987).– Shelf to basin transition in the Cambrian–Lower Ordovician of Sardinia (Italy).– Geologische Rundschau, Berlin–Heidelberg, v. 76/3, p. 827–836.
G. & E. (1995).– The Cambrian of the Moroccan Atlas region.– Beringeria special issue, Würzburg, v. 2, p. 7–46.
G. & E. (2004).– A unified Lower–Middle Cambrian chronostratigraphy for West Gondwana.– Acta Geologica Polonica, Warszawa, v. 54, p. 179–218.
R. & I. (2002).– Biostratigraphy and palaeobiogeography of the Cambrian genus Hipponicharion (Ostracoda).– Paläontologische Zeitschrift, Stuttgart, v. 76, p. 65–74.
A.P. & J.S. (2000).– Cambrian monoplacophoran molluscs (Class Helcionellida).– American Malacological Bulletin, Houston, v. 15, p. 139–145.
T.E. & J. (2001).– Ecologic radiation of Cambro–Ordovician echinoderms. In: A.Yu. & R. (eds.), The ecology of the Cambrian radiation.– Perspectives in Paleobiology & Earth History.– Columbia University Press, New York, p. 428–444.
Y. (2003).– Paläoökologie und environment-Rekonstruktion mittelkambrischer Abfolgen SW-Sardiniens (Campo Pisano Formation).– unpublished diploma-thesis, TU Bergakademie Freiberg, Freiberg, 92 p.
C.R. (1986).– Facies and bedding sequences in shelf-storm deposited carbonates. Fayettville Shale and Pitkin Limestone (Mississippian), Arkansas.– Journal of Sedimentary Petrology, Tulsa, v. 56, p. 123–137.
I., P., M. & K.J. (1990).– The problematic Hadimopanella, Kaimanella, Milaculum and Utahphospha identified as sclerites of Palaeoscolecida.– Lethaia, Oslo, v. 23, p. 11–17.
D., M. & M. (2002).– New well-preserved scleritomes of chancelloriidae from the Early Cambrian Yuanshan Formation (Chengjiang, China) and the Middle Cambrian Wheeler Shale (Utah, USA) and paleobiological implications.– Journal of Paleontology, Ithaka, v. 76, p. 596–606.
B. & H.H. (1983).– The Infrakambrian Bithia-Formation – its facies development in SW-Sardinia.– Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, Stuttgart, v. 24, p. 369– 384.
E., G. & K.E. (2002).– Latest Early Cambrian small shelly fossils, trilobites, and hatch hill dysaerobic interval on the Québec continental slope.– Journal of Paleontology, Ithaka, v. 76, p. 287–305.
F., A., W., A., G.L. & E. (2002).– A general view on the post-Sardic Ordovician sequenze from SW Sardinia.– Rendiconti della Società Paleontologica Italiana, Modena, v. 1, p. 51–68.
B. & O. (2003).– Palaeogeographical and palaeoecological aspects of the Cambro–Ordovician radiation of echinoderms in Gondwanan Africa and peri-Gondwanan Europe.– Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, v. 195, p. 73–97.
E., A. & K. (1993).– The Lower–Middle Cambrian stages and stratotypes from the Iberian Peninsula, a revision.– Geological Magazine, Cambridge, v. 130, p. 817–833.
E., R., T., J.A., J.M. & E. (2002).– Cambrian. In: W. & T. (eds.), The Geology of Spain.– Geological Society London, p. 17–29.
A., G.L. & F. (1995).– The Cambrian and Early Ordovician of south-western Sardinia. In: A. (ed.), "Sardinia 95" – 6th Paleobenthos International Symposium.– Rendiconti del Seminario della Facolta di Scienze dell'Universita di Cagliari, supplement v. LXV, Cagliari, p. 63–81.
W.S., C.R. & M.D. (1992).– Early Cambrian continental reconstructions.– Journal of the Geological Society London, London, v. 149, p. 599–606.
D. (1999).– Die frühe Evolution der Porifera.– Münchner Geowissenschaftliche Abhandlungen, Reihe A, München, v. 37, p. 1–72.
M. & O. (2004).– Cambrian lingulid and acrotretid brachiopods from the Iglesiente area (Campo Pisano Formation, southwestern Sardinia).– Rivista Italiana di Paleontologia e Stratigrafia, Milano, v. 110, p. 581–589.
H. (1985).– Neue heteractinide Spongien (Calcispongea) aus dem Unter- und Mittelkambrium Südwestsardiniens.– Berichte des naturwissenschaftlich medizinischen Vereins Innsbruck, Innsbruck, v. 72, p. 7–32.
K.J. & I. (1993).– Palaeoscolecid worms from the Middle Cambrian of Australia.– Palaeontology, Ithaka, v. 36, p. 549–592.
P.Yu. (2001).– Molluscs and siphonoconchs. In: E.M., J.B., A.Y. & A.Y. (eds.), The Cambrian biostratigraphy of the Stansbury basin, South Australia.– Transactions of the Russian Acadademy of Sciences, Moskva, v. 282, p. 133–203.
G.L. (1991).– Trilobites du Cambrien inférieur du SW de la Sardaigne (Italie).– Palaeontographia Italica, Pisa, v. 78, p. 1–174.
G.L. (1994).– The Iglesias Group in the type-area of Campo Pisano–Cabitza. In: T. & M. (eds.), Sedimentological, stratigraphical and ore deposits field guide of the autochthonous Cambro–Ordovician of southwestern Sardinia.– Memorie descrittive della carta geologica d'Italia, Servizio Geologico Nazionale, v. XLVIII, p. 304–306.
G.L., F. & A. (2002).– The Cambrian and Early Ordovician of SW Sardinia (Italy).– Rendiconti della Società Paleontologica Italiana, Modena, v. 1, p. 37–49.
F. (1972).– Cambrian trilobite faunas of Sardinia.– Atti della Accademia Nazionale dei Lincei Memorie, Classe di Scienze fisiche, mattematiche e naturali, Roma, (serie VIII), v. XI, sezione Iia, p. 1–100.
J.F. (1985).– Carbonate platform facies models.– The American Association of Petroleum Geologists Bulletin, Tulsa, v. 69, n° 1, p. 1–21.
H.G. (2002).– Sedimentary Environments: Processes, Facies and Stratigraphy.– Blackwell Science, 688 p.
A. (1987).– Fazies, Diagenese und Metamorphose der unterkambrischen Nebida Formation und ihrer Liegendschichten im Sulcis und Iglesiente (Südwest-Sardinien).– unpublished PhD thesis, Universität Heidelberg, 129 p.
T. (1985).– Fazies, Geochemie und Paläogeographie der unter- bis mittelkambrischen Gonnesa Formation sowie der basalen Cabitza Formation des Sulcis (SW Sardinien, Italien): Der Zerfall einer Karbonatplattform.– unpublished PhD thesis, Albert-Ludwigs-Universität, Freiburg, 154 p.
K. (1972).– Das Kambrium der acadobaltischen Faunenprovinz – Gegenwärtiger Kenntnisstand und Probleme.– Zentralblatt für Geologie und Paläontologie, Stuttgart, v. II, p. 1–91.
K.B. & I.D. (2001).– Global facies distributions from late Vendian to mid-Ordovician. In: A.Yu. & R. (eds.), The ecology of the Cambrian radiation.– Perspectives in Paleobiology & Earth History.– Columbia University Press, New York, p. 47–68.
J. (1992).– Radiation of echinoderms. In: J.H. & P.W. (eds.), Origin and early evolution of the metazoa.– Plenum Press, New York and London, p. 375–398.
R. & B. (2001).– Palaeoscolecid sclerites from the Upper Cambrian Mila Formation of the Shamirzad section, Alborz Mountains, northern Iran.– Acta Geologica Polonica, Warszawa, v. 51/2, p. 101–107.
I. (1972).– Las litofacies carbonatadas del Cámbrico de la Zona Cantábrica (NE España) y su distribución paleogeográfica.– Trabajos de Geologia, Oviedo, v. 5, p. 1–118.
B. (1998).– Einführung in die Paläobiologie, Teil 3: Spezielle Paläontologie (Würmer, Arthropoden, Lophophoraten, Echinodermen).– E. Schweizerbart'sche Verlagsbuchhandlung (Nägele u. Obermiller), Stuttgart, 666 p.
![]()
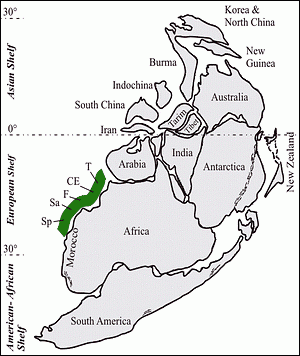
Click on thumbnail to enlarge the image.
Figure 1: Paleogeographic map of Gondwana during late Early to early Middle Cambrian times with the tentative location of the Mediterranean province in dark gray (modified after et alii, 1992). T – Turkey, CE – Central Europe, F – France, Sa – Sardinia, Sp – Spain.
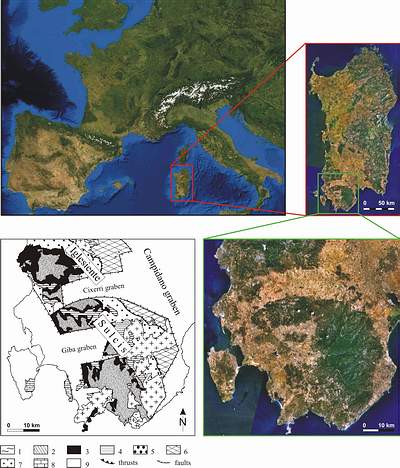
Click on thumbnail to enlarge the image.
Figure 2: Location and simplified geological map of the work area in southwestern Sardinia. The two pictures at the bottom show the same segment and well correlation of main geological structures and those in satellite view (map modified after et alii, 1988; satellite photographs from NASA World Wind). 1 – Precambrian, 2 – Nebida Group (Early Cambrian), 3 – Gonnesa Group (Early Cambrian), 4 – Iglesias Group (late Early Cambrian to Early Ordovician), 5 – Early Ordovician to Devonian, 6 – allochthonous Paleozoic, 7 – Variscan granites, 8 – Mesozoic, 9 – Cenozoic.
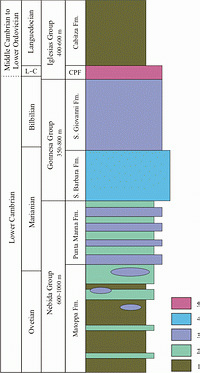
Click on thumbnail to enlarge the image.
Figure 3: Stratigraphic succession of the Cambrian of SW Sardinia related to Spanish substages. 1 – claystone/siltstone, 2 – sandstone, 3 – limestone, 4 – microbial dolostone, 5 – nodular limestone, CPF – Campo Pisano Formation, L–C: Leonian and Caesaraugustan stages.
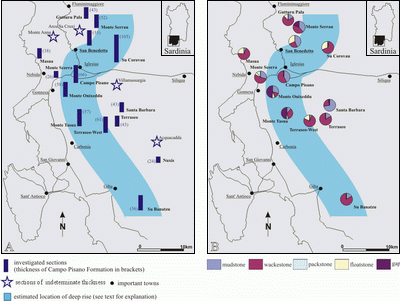
Click on thumbnail to enlarge the image.
Figure 4: Plan of southwestern Sardinia showing the location of the measured sections of the Campo Pisano Formation, their thickness (A); and the relative amounts of the four carbonate lithotypes comprising them (B). The area occupied by suspected weak sea-floor high is in blue (see text for explanation). On the postulated high sections are thicker and the percentage of autochthonous carbonates greater, so the high is presumed to have favoured carbonate production.

Click on thumbnail to enlarge the image.
Figure 5:
1 – Field view of the San Benedetto section. Massive light limestones on the left represent the San Giovanni
Fm (isolated platform, Gonnesa Group); with sharp transition (red arrow), the Campo Pisano
Fm follows (yellowish/reddish nodular banks of the drowning platform, thickness here about 45 m); to the right (yellow arrow) the Cabitza shales (deep siliciclastic cover of the platform; overgrown by vegetation) follow.
2 – Internal breccia at Arcu Sa Cruxi locality near San Benedetto. Note the big clasts
imbedded in reddish matrix. Camera lid for scale.
3 – Macroscopic view of a typical Campo Pisano nodular limestone (Campo Pisano type section). Note the colour changes parallel to the bedding plane and the change in the degree of nodularity vertically. Hammer for scale.
4 & 5 – Nodular texture of the Campo Pisano Fm in mesoscopic view. If the carbonate is removed by erosion, only siliciclastic bands accumulated by diagenetic pressure solution remain
(4, Gutturu Pala section); on fresh surfaces the carbonate nodules and stages of origin of pseudo-components are visible
(5, Gutturu Pala section). Camera lid for scale.
6 – More or less isolated carbonate nodules enclosed in residual siliciclastic bands are rather rare, but, indicate the great role played by diagenetic pressure solution (Masua section). Hammer for scale.
7 – Wavy bedded to slightly nodular textures in the Campo Pisano Fm with nests of residual siliciclastics instead of bands (Gutturu Pala section). Scale bar is 10 cm.
8 – Massive texture of the Campo Pisano Fm indicated by a limestone bank (white arrows) within the wavy to slightly nodular succession at Monte Scorra section. Hammer for scale.
9 – "Shaly" texture of the Campo Pisano Fm at Su Banatzu section. Hammer for scale.

Click on thumbnail to enlarge the image.
Figure 6:
1 – Origin of nodular texture by pressure solution (photomicrograph, Campo Pisano
Fm, Monte Serrau section). Note the snug fit of the pseudo-components. Scale bar is 1 mm.
2 – Photomicrograph of a siliciclastic band showing at the left a continuous lateral transition into limestone (the siliciclastic band seems to vanish. So, the band is not a primary sedimentary layer, but, a pressure solution suture. Nevertheless, the band can include carbonate relicts (pseudo-clasts). Masua section. Scale bar is 1 mm.
3 – Photomicrograph of platy texture in the Campo Pisano Fm described by previous authors as "calcareous slates" (Terraseo-West section). Scale bar is 1 mm.
4 – Photomicrograph of a rapid alternation of limestone and thin siliciclastic laminae typical of the transition from Campo Pisano
Fm to the Cabitza Fm (Gutturu Pala section). Scale bar is 1 mm.
5 – Massive layers of sponge spicules (spiculite wackestone) common at the base of the Campo Pisano
Fm (biofacies unit 1). Photomicrograph, Masua section. Scale bar is 1 mm.
6 – Echinoderm wackestone is characteristic for biofacies unit 2. Stereome preservation in the form of individual calcite crystals can be seen. Photomicrograph, San Benedetto section. Scale bar is 1 mm.
7 – Trilobites predominate in the upper portion of the sections (biofacies unit 3), sometimes they indicate bioturbation (see orientation of fossils). Trilobite wackestone. Photomicrograph, Gutturu Pala section. Scale bar is 1 mm.
8 – Sponge spicule from the Masua section (biofacies unit 1). SEM-micrograph. Scale bar is 0.1 mm.
9 – Silicified trilobite cephalon (Dawsonia bohemica) from biofacies unit 3 (Gutturu Pala section). SEM-micrograph. Scale bar is 1 mm.
10 – Phosphatic palaeoscolecid Hadimopanella sp. ("small shelly fossil") from biofacies unit 2 (Su Corovau section). SEM-micrograph. Scale bar is 0.03 mm.
11 – Phosphatic cambroclave ("small shelly fossil") from biofacies unit 2 (Su Corovau section). SEM-micrograph. Scale bar is 0.1 mm.
12 – Phosphatically preserved echinoderm ossicle from biofacies unit 2 (Su Corovau section). Note the stereome structure and the distinct lumen. SEM-micrograph. Scale bar is 0.1 mm.
13 – Hyolith Microcornus sp. from Su Corovau section (biofacies unit 2). SEM-micrograph. Scale bar is 0.1 mm.
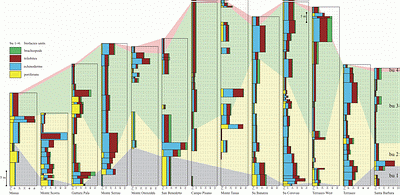
Click on thumbnail to enlarge the image.
Figure 7: Fossil content (from thin sections) of stratigraphic sections of the Campo Pisano Formation arranged from West (Masua section) to East (Santa Barbara). Some sections shown on
Fig. 4 ![]() are not included because of a stronger metamorphic overprint and/or local tectonics. Some tectonic events cause an interruption in the continuity of the fossil record. The units so affected are designated.
are not included because of a stronger metamorphic overprint and/or local tectonics. Some tectonic events cause an interruption in the continuity of the fossil record. The units so affected are designated.
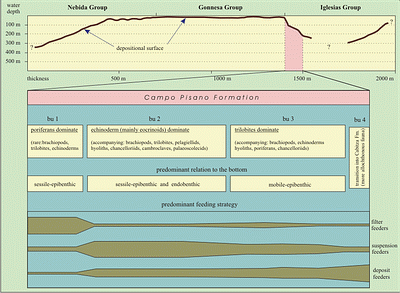
Click on thumbnail to enlarge the image.
Figure 8: Bathymetry and thickness of the Sardinian Cambrian deposits and paleoecological succession (character of habitats and the dominant feeding strategy) in the Campo Pisano Formation that together indicate an intermittent drowning process.
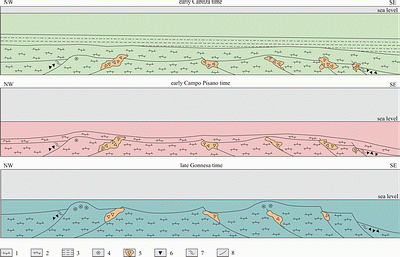
Click on thumbnail to enlarge the image.
Figure 9: Interpretation of development stages at different time slices: During late Gonnesa time, the carbonate platform was already segmented by tensional tectonics (origin of internal breccias) resulting in a significant bottom topography; at steep flanks mass flows occurred. Early in Campo Pisano times the sea level rose and tensional-related faults were still active locally. This led to the continued development of internal breccias and bottom topography was only moderately accidented. Early in Cabitza times the rise in sea level continued and deeper marine clastics covered the drowned carbonate series. 1 – San Giovanni Formation, 2 – Campo Pisano Formation, 3 – Cabitza Formation, 4 – oolites, 5 – internal breccia, 6 – external breccia, 7 – mass flow, 8 – fault.
![]()