![]()
![]()
La section du stratotype de la limite Campanien-Maastrichtien établi à Tercis (SO France) comprend des niveaux carbonatés indurés qui ont été dissociés par acétolyse. Les résidus obtenus ont permis d'observer pour la première fois des microfossiles dégagés dans ces niveaux. Parmi eux figurent en grande quantité des tests classés incertae sedis qui sont illustrés ici en détail pour la première fois. Des observations à la loupe et au microscope optique permettent d'illustrer 36 formes reconstituées sur ordinateur. Des examens complémentaires au microscope électronique précisent la caractérisation des formes permettant de distinguer les caractères propres au groupe homogène des gilianelles, microproblematica comprenant 30 des 36 formes illustrées vis à vis de formes qui ne présentent pas tous ces caractères. D'après la morphologie des tests, un mode de vie en suspension dans l'eau est suggéré pour certains des micro-organismes ayant produit ces tests à l'organisation simple. Quant aux microproblematica nommés gilianelles, une appartenance à un ordre non répertorié de Protozoaires Rhizopodes est proposée. La distribution stratigraphique établie dans la section met en évidence un extraordinaire taux de renouvellement de ces incertae sedis dont beaucoup n'ont laissé de témoignages que pendant moins d'un million d'années.
Micropaléontologie ; microfossiles calcaires ; microproblematica ; gilianelles ; Campanien ; Maastrichtien ; stratotype.
G.S. & A. (2006).- Nouvelles études micropaléontologiques sur le stratotype de la limite Campanien-Maastrichtien à Tercis (SO France) : les gilianelles (microproblematica) extraites par acétolyse.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2006/05 (CG2006_A05)
New micropalaeontological studies on the stratotype of the Campanian-Maastrichtian boundary at Tercis (SW France): the gilianelles (microproblematica) extracted through acetolysis.- The Campanian-Maastrichtian stratotype section at Tercis (SW France) consists mainly of hard calcareous levels. Their treatment using acetolysis left residues among which for the first time well-preserved free microfossils were obtained (These levels had been studied previously only through thin sections). Within these residues, a wealth of calcareous shells of unknown affinity (incertae sedis) occur. These previously undescribed fossils are documented herein with photographs and drawings. Optical observations are synthesised in computer assisted drawings which depict 36 discrete forms. Scanning electron microscopy documented the nannostructures of these tests which led to the recognition of 30 forms with a common organisation and nannostructure. These are the microproblematica group called "gilianelles". The remaining six forms of incertae sedis do not share the same characteristics. According to morphological observations, a planktonic existence is inferred for most of the incertae sedis. The simple organisation and small size of the gilianelles suggest that they may represent a new Order of Rhizopod Protozoans. Their stratigraphical distribution has been established in the stratotype section of the "Grande Carrière" at Tercis (SW France). This distribution shows an extraordinary rate of turn-over with some taxa having a range of less than 1 Ma.
Micropalaeontology; calcareous microfossils; microproblematica; gilianelles; Campanian; Maastrichtian; stratotype.
Additional micropalaeontological characterisation of the global standard stratotype section of the boundary between the Campanian and Maastrichtian stages at Tercis, SW France (, 2001a) has been undertaken using several techniques of acetolysis applied to hard carbonate deposits. These new experimental procedures and the special care devoted to the 0.10 - 0.25 mm residual fraction led to the discovery of unpublished microfossils including a surprisingly diversified series of microproblematica. They are presented in a previous paper (, in press) where a morphological terminology is proposed. In that earlier paper, this new group of microfossils has been shown to be different from any known to date. They are called "gilianelles" with a type species designated under the name Gilianella stellata . In this second contribution, we wish to give a comprehensive illustration of their morphology and the first published data on their nannostructure which is sufficiently discrete to distinguish the gilianelles from other microproblematica. We also consider the ecological aspect and the potential of these microfossils for stratigraphic studies. Taxonomic considerations (formal naming, classification) will be the subject of a third presentation in preparation.
The gilianelles are small compared to the majority of foraminifera. Other characteristics presented in (in press) include the fact that they are all unilocular calcareous shells with an axial symmetry. A unique, small, axial aperture determines an oral face. Expansions are often present in equatorial position (arms, collarette) or aboral position (axial foot, peri-axial feet or collarette).
Pictures made with a digital camera adapted to an ocular of the
stereomicroscope are given first. This technique
(Fig. 1 ![]() ) is
usually able to distinguish forms and allows different views of the same specimen to
be made quickly for tridimensional illustration. For example,
Figure 2
) is
usually able to distinguish forms and allows different views of the same specimen to
be made quickly for tridimensional illustration. For example,
Figure 2 ![]() shows that the 2 faces of the shell of a specimen of the form B2b are not similar.
Figure 3
shows that the 2 faces of the shell of a specimen of the form B2b are not similar.
Figure 3 ![]() presents several different views of a shell of form H2 which consists of a body, 7 arms and 2 peri-axial feet.
presents several different views of a shell of form H2 which consists of a body, 7 arms and 2 peri-axial feet.
More precise information came from microscopic observation at low magnification. A general view of a picking is given in
Figure 4 ![]() :
several discrete microproblematica are present but foraminifera, pithonellids,
and echinoderm remains occur also. The name gilianelle has been restricted to unilocular shells
of which the internal cavity is visible in some broken specimens. This character easily distinguishes
these specimens from the skeletal elements of echinoderms and other remains. From digitalised
photographs, specimens have been selected and rearranged with a computer as shown in
Figure 5
:
several discrete microproblematica are present but foraminifera, pithonellids,
and echinoderm remains occur also. The name gilianelle has been restricted to unilocular shells
of which the internal cavity is visible in some broken specimens. This character easily distinguishes
these specimens from the skeletal elements of echinoderms and other remains. From digitalised
photographs, specimens have been selected and rearranged with a computer as shown in
Figure 5 ![]() .
.
From a collection of about 75 photographs taken with a stereomicroscope and 200 by microscope that together include pictures of about one thousand specimens it has been possible to define and to draw a number of discrete forms using the Blender free software (see www.blender.org). The result of this work is given in 6 plates which illustrate 36 among the forms characterised to date.
As a preliminary step, the microproblematica have been classified into 5 morphological sets: discs, spheres, cupules, cones, kegs. Descriptions have been undertaken as a first approach to diagnosis. They are summarised in this English extended abstract quoted from (in press).
![]() "Seven simple discoid forms (A group) are presently known. Some of them
(
) evolve along the documented section at Tercis (SW France) which nevertheless represents about 5 Ma of deposition. At least five forms (B group) are discoid shells with arms, the number of which varies (from 4 to 15) from a form to another. Often, the number varies also within a given form. Two extra forms have a thick discoid shell with arms (C group).
"Seven simple discoid forms (A group) are presently known. Some of them
(
) evolve along the documented section at Tercis (SW France) which nevertheless represents about 5 Ma of deposition. At least five forms (B group) are discoid shells with arms, the number of which varies (from 4 to 15) from a form to another. Often, the number varies also within a given form. Two extra forms have a thick discoid shell with arms (C group).
![]() Among the spheres (D group) 3 forms have been evidenced.
Among the spheres (D group) 3 forms have been evidenced.
![]() Cupules are diversified with 2-4 forms without any arm or foot (E group). Another group shows thick and short arms (F group). A third group displays arms and feet (G group). A fourth group (H) shows tube-like arms (7 to 9) and feet (2 to 5). A fifth group has blade-like expansions (J
group). (...)
Cupules are diversified with 2-4 forms without any arm or foot (E group). Another group shows thick and short arms (F group). A third group displays arms and feet (G group). A fourth group (H) shows tube-like arms (7 to 9) and feet (2 to 5). A fifth group has blade-like expansions (J
group). (...)
![]() Conoid microproblematica comprise smooth tests such as a pin-like form (K1) and a funnel-shaped form (L1). Other conoid forms are ribbed (M group with 5 distinct forms).
Conoid microproblematica comprise smooth tests such as a pin-like form (K1) and a funnel-shaped form (L1). Other conoid forms are ribbed (M group with 5 distinct forms).
![]() The keg-shaped tests (N group with 3-5 forms) are often finely
ribbed."
The keg-shaped tests (N group with 3-5 forms) are often finely
ribbed."
Scanning electron microscopy made possible a characterization of the shell wall, the surface ornamentation, the expansions.
3.1. Shell wall
The shell wall is
consistently 15-18 µm thick
(Pl. 7 ![]() )
and in 32 of the 36 forms is but a single layer. It is made up of elongated crystals generally oriented perpendicularly to the surface of the wall. A view from inside a broken shell
(Pl. 7, fig. 3
)
and in 32 of the 36 forms is but a single layer. It is made up of elongated crystals generally oriented perpendicularly to the surface of the wall. A view from inside a broken shell
(Pl. 7, fig. 3 ![]() ) suggests the presence of pores in the wall.
) suggests the presence of pores in the wall.
Four forms do not
have all of the characteristics that identify the gilianelles and so are removed from the main set of microproblematica. The shell wall of form L1 is 15 to 20 µm thick as
is usual in gilianelles but has 2 layers. The outer layer has no organisation and
the inner one consists of crystals oriented perpendicularly to the surface as is
the case in the named group gilianelles (Pl. 8, fig. 6 ![]() ).
The crystals of this inner layer are arranged regularly at the surface
(Pl. 8, fig. 4
).
The crystals of this inner layer are arranged regularly at the surface
(Pl. 8, fig. 4 ![]() ). In the foot of form L1, crystals of the
inner layer are axially elongated fibres
(Pl. 8, fig. 5
). In the foot of form L1, crystals of the
inner layer are axially elongated fibres
(Pl. 8, fig. 5 ![]() ).
).
The several forms of group K have no shell walls. However, we have observed specimens of form K1 made up entirely of glaucony at levels 54.9 and 69.0 of the section. Because glaucony is commonly present at Tercis as infillings of the chambers of microfossils , this find suggests that the group K forms are all internal casts.
Another exception is that of group D
of which the shell wall is 25 to 30 µm thick
(Pl. 8, fig. 3 ![]() ).
).
3.2. Surface ornamentation
The external ornamentation of the shells is rich and can be seen using the scanning electron microscope. A constant and original nannostructure
is evident for 30 among the 36 forms illustrated. It is characterised by several
kinds of cloth-like arrangements which may or not combine to form radial or concentric microstructures
(Pl. 9, figs. 1 & 7 ![]() )
)
Forms of group D show a honey comb microstructure with no nannostructural features apparent at the surface. Shells of forms E1, F1 and L1 appear to have a smooth external surface. The absence of shell remains in group K results in their having no known surface ornamentation. These 6 microproblematica are thus not part of the main population which comprises the gilianelles.
3.3. Expansions
The presence or
absence of expansions and their form, position, structure and number permit the differentiation
of the various forms. Some examples are illustrated
(Pl. 9 ![]() ). Arms and feet may have a cross section
(Pl. 9, figs. 4-5
). Arms and feet may have a cross section
(Pl. 9, figs. 4-5 ![]() ); others are tubular (form F1 :
Pl. 9, fig. 6
); others are tubular (form F1 :
Pl. 9, fig. 6 ![]() ) or full
(Pl. 8, fig. 5
) or full
(Pl. 8, fig. 5 ![]() ). Some tubular feet show wings
(Pl. 9, fig. 3
). Some tubular feet show wings
(Pl. 9, fig. 3 ![]() ). Expansions may also be blades
(Pl. 9, fig. 6
). Expansions may also be blades
(Pl. 9, fig. 6 ![]() ).
).
4.1. Ecology
Many of the shells illustrated here are arranged in such a manner that, if
submerged in sea water, they would become oriented with the aperture upward. This results from the relative position of the
centre of gravity and the centre of the effect as explained in
Figure 6 ![]() . The presence, orientation and position of expansions
all seem to contribute to a hydrodynamic equilibrium
that maintains the aperture upward. Those forms have an adaptation that implies a planktonic ecology.
. The presence, orientation and position of expansions
all seem to contribute to a hydrodynamic equilibrium
that maintains the aperture upward. Those forms have an adaptation that implies a planktonic ecology.
Another adaptation to planktonic life is the development of elongated and thin expansions which tends to increase buoyancy. Many expansions of our shells are reminiscent of the expansions observed in planktonic larvae.
Forms A4a and A4b, D, and C1 (we cannot evaluate forms of the group K) do not present such an adaptation and it is probable that they were benthic.
4.2. Temporal distribution of microproblematica
The stratigraphical distribution of the best documented microproblematica from
Tercis is summarised in
Figure 7 ![]() . The time scale is in millions of year
based on estimates by
(2001b) improved following North-America/Europe precise correlation obtained from the study of inoceramids ( & ,
2003). The duration of the geological occurrence of
these microproblematica may be compared to that of Radotruncana calcarata. This
species probably the most precise biostratigraphic marker of the Campanian is also present in the residues of acetolysis and was obtained
free of matrix for the first time from the Campanian-Maastrichtian
stratotype. Several microproblematica have a stratigraphical range as short as that of Radotruncana calcarata and would be excellent stratigraphical markers if found in other places.
. The time scale is in millions of year
based on estimates by
(2001b) improved following North-America/Europe precise correlation obtained from the study of inoceramids ( & ,
2003). The duration of the geological occurrence of
these microproblematica may be compared to that of Radotruncana calcarata. This
species probably the most precise biostratigraphic marker of the Campanian is also present in the residues of acetolysis and was obtained
free of matrix for the first time from the Campanian-Maastrichtian
stratotype. Several microproblematica have a stratigraphical range as short as that of Radotruncana calcarata and would be excellent stratigraphical markers if found in other places.
A nearby section has been investigated in order to document this point. The studied stratotype is located on the northern flank of the Tercis-Angoumé diapiric anticline (, 2001), a similar distribution of microproblematica has been documented on the southern flank. The microproblematica have not been found in upper Campanian flysch in four sections about 100 km apart in the Pyrenees. However, a single similar form was found in another section from Northern Spain, where external platform deposits like those at Tercis occur.
4.3. Biological affinities of microproblematica
Since June 2005, we have circulated pictures of gilianelles among palaeontologists. No one form was recognised but several hypotheses have been suggested. Following appropriate discussion, all these hypotheses have been rejected by (in press) including microcrinoids (, 1999; & , 2001), calcareous dinoflagellates ( & , 1994), pseudarcellids -Tertiary micro enigmatica- (, 1989), calcitised radiolarians (pers. comm., T. ), bryozoans (, 1990, 1991). In addition, the microproblematica of Tercis are not thecamoebians in which the shell (theca) is neither calcareous, nor ornamented like that of gilianelles ( et alii, 2003). As far as forms of group D are concerned, their appearance is similar to that of some foraminifera such as Favulina hexagona () as illustrated by (2006, fig. 28) ; the size is similar (the scale given in 's figure is 10 times too large) but the shell wall illustrated by in another species of the same genus is only 4 µm thick; it is 30 µm thick in the forms of group D.
The set of shells illustrated was built by simple unicellular organisms smaller than most foraminifers. The simple calcareous shell and small size suggest that the set pertains to the phylum Protozoa. Among its group, the Class Rhizopoda is a possible repository. But the set is not ascribable to any of the known Orders (Foraminifera, Radiolaria, Heliozoa).
4.4. Discussion
The morphological grouping established after optical observation is not supported by the study using a scanning electron microscopic. For example, conoid forms (K, L, M) include infillings, shell with double and mono layered walls while some cupules (E2, E3, G) display an ornamentation similar to that of cones (group M) with respect to striated bodies and a peri-oral lip with radial combs.
Some forms of microproblematica must be removed from the group established by designation of the type species Gilianella stellata . These removals are as follows : group K (infillings of unknown shells), E1 and F1 (surface of the shell without cloth-like nannostructure), L1 (double layered wall). The 30 others microproblematica have a single chambered small shell, a fundamentally axial symmetry, a single aperture, a mono-layered wall, 15-20 µm thick made up of elongated crystals perpendicular to the surface, and a cloth-like nannostructure.
As now redefined, the gilianelles include only the 30 forms illustrated here from the outcrop at Tercis where more will be found in the future.
We have brought to the attention of micropalaeontologists a comprehensive documentation of a variety of Campanian-Maastrichtian microproblematica present in the stratotype of Tercis as determined by the use of several techniques of observation.
This paper illustrates for the first time the precise nature of expansions thus allowing a preliminary classification based on a clear differentiation of the various forms of microproblematica. Within these groupings, the gilianelles display specific nannostructures at the surface of their shell.
Preliminary diagnoses have been proposed; they will facilitate the next step of the research: to establish a rigorous taxonomy (classification and naming of forms). A second step, yet to be undertaken, involves the search for similar microfossils in other outcrops of the same age (Campanian-Maastrichtian) with the same facies (continental carbonate platform of the Tethyan Realm).
We hope that this presentation will encourage others to carry out research in these fields in the near future, for the complete elucidation of this topic will require a period of time far greater than that remaining to us.
La section de Tercis (Landes, France) a servi à élaborer le point stratotypique mondial (PSM) de la limite Campanien-Maastrichtien ( & , 2001a). À cette fin, une récolte systématique de longue haleine fut entreprise pour la recherche de fossiles dans de nombreux groupes qui furent étudiés par un ou plusieurs experts lors d'une étude coopérative (, 2001a).
Parmi les microfossiles et, singulièrement, les microfossiles calcaires, il restait un besoin d'amélioration lié au fait qu'une large portion de la section type, la Grande Carrière de Tercis, était constituée de niveaux carbonatés indurés (moins de 10 % d'argile : , 2001c). Après avoir épuisé les niveaux plus argileux qui donnaient des restes abondants mais ponctuels, nous avons cherché à dissocier les niveaux compacts.
Des traitements par acétolyse variés ont été appliqués pendant 3 années (2004-2006), ce qui a permis de découvrir des restes non encore observés. La distribution des espèces déjà répertoriées fut complétée. De nouveaux foraminifères benthiques et planctoniques, des ostracodes, des éléments squelettiques d'échinodermes (échinides, holothuries, ophiures ) ont été isolée. Durant ces recherches, des restes incertae sedis qui n'avaient pas retenu l'attention auparavant furent observés régulièrement puis systématiquement recherchés et collectés.
Ces restes font l'objet de la présente contribution. Dans une première contribution (, sous presse), une présentation de ces incertae sedis a été faite. Elle établit une terminologie morphologique qui est reprise ici pour décrire les tests. Le terme de gilianelles (microproblematica) a été proposé pour désigner un partie de ces incertae sedis avec comme type l'espèce nouvelle Gilianella stellata. L'intérêt de les nommer fut établi après avoir démontré qu'il était distinct de tout ce qui était connu auparavant. Dans la présente contribution, la seconde sur le sujet, nous illustrons en détail l'aspect morphologique et apportons les premières informations sur la nannostructure de l'ensemble des incertae sedis. Enfin, nous envisageons des aspects biologiques de ces microfossiles et ce qu'ils peuvent apporter au géologue. Nous reportons à une future contribution les aspects taxonomiques : présentation formelle des espèces et classification.
Les gilianelles ont des tests de petite taille. Dans un premier temps, nous les avons recherchées dans toutes les fractions des roches dissociées ; par la suite, nous nous sommes focalisés sur la fraction comprise entre 100 et 250 µm.
Les autres caractéristiques des gilianelles furent précisées par ailleurs (, sous presse) : il s'agit de tests calcaires constitués d'un corps délimitant une loge unique, entière ; la symétrie est axiale. Un orifice unique, petit, axial, détermine une face orale ; cet orifice peut être porté par ou entouré d'une structure particulière (col haut ou lèvre fine).
Des expansions viennent parfois orner ce corps en position équatoriale comme en position aborale. Lorsqu'elles sont présentes, les expansions équatoriales, qualifiées de bras, donnent une organisation rayonnée. Les rayons sont en nombre varié selon les formes et, souvent, à l'intérieur d'une même forme. Parfois, une expansion équatoriale continue forme un bourrelet ; elle se développe en une collerette délimitant clairement la face orale de la face aborale dans plusieurs formes.
En position aborale, il existe parfois une expansion axiale nommée pied. Toutefois, l'expansion aborale peut être péri-axiale (bourrelet ou collerette cylindrique). Cette expansion peut être remplacée par des pieds, irrégulièrement disposés sur un cercle situé près de l'axe ou à mi-rayon, tandis que la collerette péri-axiale peut elle-même se prolonger de diverticules en forme de palette ; l'organisation rayonnée devient irrégulière.
Étant donnée leur petite taille, l'utilisation du microscope électronique est la seule façon de caractériser complètement ces restes. Cependant, outre la lourdeur du travail, des restrictions diverses obligent à envisager et développer des moyens d'illustration plus immédiats. Hormis la question du financement, l'illustration au moyen du microscope électronique pose la question de la manipulation de microfossiles fragilisés par les dissociations chimiques. En outre, les spécimens traités en vue de cet examen sont perdus pour constituer un matériel type lorsque des taxons nouveaux sont concernés. Enfin, il est souhaitable de proposer aux géologues des figurations susceptibles de les informer sur l'existence d'un groupe qu'ils pourraient observer eux-mêmes et prendre en compte.
Pour toutes ces raisons, nous proposons ci-après une iconographie composée de vues à la loupe, au microscope et de dessins assistés par ordinateur.
2.1. Vues à la loupe binoculaire
Les clichés ont été réalisés en adaptant simplement un boîtier numérique sur un des tubes de la loupe binoculaire. Ces clichés sont suffisants pour distinguer, caractériser et reconnaître un bon nombre de formes.
La Figure 1 ![]() illustre le fait que, si la qualité des images n'est pas parfaite, beaucoup de spécimens peuvent être caractérisés et des modèles morphologiques proposés. Lors de telles observations, il n'est pas toujours évident de repérer l'orifice unique qui est une des caractéristiques du groupe. En effet, les deux pôles des tests comportent parfois des structures (dépression, bouton ou picot en relief,
) dont la relation avec la cavité intérieure n'est pas évidente. Pour lever l'incertitude, un test est plongé dans une solution diluée d'acide acétique. L'expérience a montré que la cavité du test contenait du carbonate attaqué préférentiellement provoquant l'émission de gaz carbonique lequel empruntait l'orifice naturel ; cet orifice était ainsi mis en évidence par l'échappement de bulles au pôle oral.
illustre le fait que, si la qualité des images n'est pas parfaite, beaucoup de spécimens peuvent être caractérisés et des modèles morphologiques proposés. Lors de telles observations, il n'est pas toujours évident de repérer l'orifice unique qui est une des caractéristiques du groupe. En effet, les deux pôles des tests comportent parfois des structures (dépression, bouton ou picot en relief,
) dont la relation avec la cavité intérieure n'est pas évidente. Pour lever l'incertitude, un test est plongé dans une solution diluée d'acide acétique. L'expérience a montré que la cavité du test contenait du carbonate attaqué préférentiellement provoquant l'émission de gaz carbonique lequel empruntait l'orifice naturel ; cet orifice était ainsi mis en évidence par l'échappement de bulles au pôle oral.
En outre, les prises de vue à la loupe binoculaire ont l'avantage de permettre d'observer et d'illustrer, un même spécimen sur toutes ses faces avec un résultat quasi instantané. La
Figure 2 ![]() montre que les deux faces d'un individu de la forme B2b ne sont pas semblables. La
Figure 3
montre que les deux faces d'un individu de la forme B2b ne sont pas semblables. La
Figure 3 ![]() rassemble des vues permettant de comprendre l'organisation d'un spécimen de la forme H2 doté d'un corps en capsule, de 7 bras et de 2 pieds plantés de côté, laquelle n'est pas évidente pour un objet de moins de 0,1 mm. Ces premières observations et prises de vues (environ 75 furent faites) ont permis d'ordonner les multiples formes rencontrées.
rassemble des vues permettant de comprendre l'organisation d'un spécimen de la forme H2 doté d'un corps en capsule, de 7 bras et de 2 pieds plantés de côté, laquelle n'est pas évidente pour un objet de moins de 0,1 mm. Ces premières observations et prises de vues (environ 75 furent faites) ont permis d'ordonner les multiples formes rencontrées.
2.2. Observation et illustration au microscope optique
Afin de préciser la morphologie des restes, des observations et prises de vues ont été entreprises à faible grossissement au microscope optique dans des cellules de tri éclairées par dessus. Le faible grossissement est rendu nécessaire par la faible profondeur de champ de ce moyen d'observation. Un peu plus de 200 clichés numériques furent réalisés au cours des deux années passées. Lorsque des grossissements plus forts étaient exigés pour certains détails, ils furent effectués numériquement sur des vues à grande résolution avec plus de succès qu'avec un objectif plus fort.
La
Figure 4 ![]() montre une vue générale d'une cellule de tri bien garnie. Elle illustre la variété des microfossiles disponibles : diverses gilianelles mais aussi quelques foraminifères, des pithonelles, des restes d'échinodermes. Quelques tests brisés laissent voir une paroi ; ce caractère distingue les tests des éléments squelettiques (spicules, ossicules,
). Nous n'avons rapporté aux gilianelles que les formes ayant montré une cavité distincte. Parmi les spicules, il y a des éléments d'holothurie (sclérites en grille,
Fig. 4
montre une vue générale d'une cellule de tri bien garnie. Elle illustre la variété des microfossiles disponibles : diverses gilianelles mais aussi quelques foraminifères, des pithonelles, des restes d'échinodermes. Quelques tests brisés laissent voir une paroi ; ce caractère distingue les tests des éléments squelettiques (spicules, ossicules,
). Nous n'avons rapporté aux gilianelles que les formes ayant montré une cavité distincte. Parmi les spicules, il y a des éléments d'holothurie (sclérites en grille,
Fig. 4 ![]() ). D'autres éléments, spicules? non identifiés et communément observés, sont formés d'un centre sub-sphérique de 80 µm de diamètre hérissé, dans toutes les directions, d'expansions cylindriques transparentes (8 ou 10) dont la longueur conservée dépasse parfois 200 µm de part et d'autre du corps central
(Fig. 4
). D'autres éléments, spicules? non identifiés et communément observés, sont formés d'un centre sub-sphérique de 80 µm de diamètre hérissé, dans toutes les directions, d'expansions cylindriques transparentes (8 ou 10) dont la longueur conservée dépasse parfois 200 µm de part et d'autre du corps central
(Fig. 4 ![]() : 3 spécimens au dessus et à droite de l'échelle).
: 3 spécimens au dessus et à droite de l'échelle).
À partir des clichés bruts obtenus au microscope optique et illustrant un millier de spécimens environ, des montages sont réalisés pour caractériser et classer les formes présentes. La plupart du temps, les clichés sont doublés avec mise au point à différents niveaux tandis qu'une même vue est souvent prise sous divers éclairages rasants afin de mettre en évidence l'ornementation de faible relief. La
Figure 5 ![]() offre un aperçu des illustrations utiles à la réalisation de l'étape suivante : la reconstitution tridimensionnelle. Les informations obtenues au microscope sont rapprochées de celles recueillies à la loupe binoculaire : comparer, par exemple, les vues orale et aborales de la forme A2b
(Fig. 5
offre un aperçu des illustrations utiles à la réalisation de l'étape suivante : la reconstitution tridimensionnelle. Les informations obtenues au microscope sont rapprochées de celles recueillies à la loupe binoculaire : comparer, par exemple, les vues orale et aborales de la forme A2b
(Fig. 5 ![]() ) et les vues de la même forme dans la
Figure 2
) et les vues de la même forme dans la
Figure 2 ![]() .
.
2.3. Reconstitution tridimensionnelle
Le logiciel gratuit Blender (adresse internet :
www.blender.org) est efficace pour modéliser les formes en trois dimensions. Le point de départ de chaque forme est une sphère ; elle est sculptée, modelée, ponctuellement étirée, pour reproduire les caractéristiques observées sur la forme du test à représenter
(Animation 1 ![]() ). Selon la complexité du volume recherché, l'élaboration de ces modèles prend de quelques minutes à plusieurs heures. Ces tentatives de représentation nous obligent à retourner à l'observation optique pour vérifier les caractéristiques de toutes les facettes
(Animation 2
). Selon la complexité du volume recherché, l'élaboration de ces modèles prend de quelques minutes à plusieurs heures. Ces tentatives de représentation nous obligent à retourner à l'observation optique pour vérifier les caractéristiques de toutes les facettes
(Animation 2 ![]() ). Le logiciel permet également de créer des animations.
). Le logiciel permet également de créer des animations.
À partir du volume obtenu, des "clichés" sont réalisés avec une caméra virtuelle sous des angles choisis et un éclairage réglable à partir de plusieurs sources dans l'espace. Ces clichés sous différents angles, sont enregistrés sous un format image de type JPEG, TIF, ... (pour ce travail environ 100 Ko par fichier TIF). Les images sont ensuite importées dans le logiciel Adobe Illustrator® afin d'assembler des planches.
Les résultats de ce travail sont donnés dans 6 planches qui précisent les caractères des 36 formes d'incertae sedis représentées ici. La même échelle est conservée partout pour une comparaison aisée. Au cours de l'illustration, certaines de ces formes n'ont pas révélé toutes les caractéristiques que nous avons attribuées aux gilianelles ce qui est précisé chaque fois.
Les microfossiles de nature incertaine ont été regroupés en cinq ensembles morphologiques (, sous presse) : disques, sphères, cupules, cônes, tonnelets. Les descriptions ci-après sont issues de la seule observation optique. Il s'agit d'une première approche de la diagnose ce qui ne permet pas d'éviter une certaine monotonie pour un exposé qui peut être sauté sans affecter la compréhension de notre étude.
Neuf formes en disque (groupe A) sont illustrées
(Planches 1-6 ![]() ); d'autres existent mais ne sont pas encore fermement caractérisées.
); d'autres existent mais ne sont pas encore fermement caractérisées.
![]() - A1a est un grand disque (souvent 200 µm) à profil sub-symétrique par rapport à l'équateur. La forme rappelle une épaisse galette bretonne. En lumière rasante, les faces orales et aborales sont généralement ornées de 3 rides dans la moitié distale ; l'orifice est discret. A1a est présente dans toute la section de la Grande Carrière et au delà, dans la partie plus haute du Maastrichtien (,
2001d) ; son abondance est localement élevée (jusqu'à 10.000 individus/ 100 g).
- A1a est un grand disque (souvent 200 µm) à profil sub-symétrique par rapport à l'équateur. La forme rappelle une épaisse galette bretonne. En lumière rasante, les faces orales et aborales sont généralement ornées de 3 rides dans la moitié distale ; l'orifice est discret. A1a est présente dans toute la section de la Grande Carrière et au delà, dans la partie plus haute du Maastrichtien (,
2001d) ; son abondance est localement élevée (jusqu'à 10.000 individus/ 100 g).
![]() - A1b est un petit disque (autour de 140 µm) à profil dissymétrique comparable à celui d'une tourte. La face orale est en dôme bas, sans ride et avec une dépression circulaire sub-périphérique. L'orifice axial est discret. La face aborale s'élève rapidement depuis l'équateur donnant au disque un aspect tronqué en profil. Une dépression circulaire périphérique et une large expansion péri-axiale sont caractéristiques de cette face. L'abondance de A1b est difficilement perceptible car cette petite forme n'est reconnaissable qu'en profil tandis que des variations d'aspect de la face orale laissent penser qu'il y aurait plusieurs formes.
- A1b est un petit disque (autour de 140 µm) à profil dissymétrique comparable à celui d'une tourte. La face orale est en dôme bas, sans ride et avec une dépression circulaire sub-périphérique. L'orifice axial est discret. La face aborale s'élève rapidement depuis l'équateur donnant au disque un aspect tronqué en profil. Une dépression circulaire périphérique et une large expansion péri-axiale sont caractéristiques de cette face. L'abondance de A1b est difficilement perceptible car cette petite forme n'est reconnaissable qu'en profil tandis que des variations d'aspect de la face orale laissent penser qu'il y aurait plusieurs formes.
![]() - A2a est un disque dont le corps, de taille voisine de celle de A1b, est prolongé d'une collerette équatoriale. La collerette à contour circulaire, translucide, est souvent endommagée; sa largeur atteint 30 µm ; elle laisse parfois deviner des rayons. La face orale est en dôme bas ; la face aborale est plus haute avec une collerette péri-axiale large et peu élevée. Cette forme est présente, sans être commune, entre les cotes 50 et 110 de la section de la Grande Carrière.
- A2a est un disque dont le corps, de taille voisine de celle de A1b, est prolongé d'une collerette équatoriale. La collerette à contour circulaire, translucide, est souvent endommagée; sa largeur atteint 30 µm ; elle laisse parfois deviner des rayons. La face orale est en dôme bas ; la face aborale est plus haute avec une collerette péri-axiale large et peu élevée. Cette forme est présente, sans être commune, entre les cotes 50 et 110 de la section de la Grande Carrière.
![]() - A2b est une forme semblable à A2a mais nettement étirée dans un diamètre du plan équatorial (diamètre long du corps : environ 140 µm). La symétrie bilatérale obtenue est quasi unique pour le groupe. Cet aspect à contour elliptique est souligné par une collerette plus étendue dans la longueur que dans la largeur. A2b est présente dès la base de la section (parfois plus de 1000 individus / 100 g) et accompagne la forme précédente entre les cotes 50 et 69.
- A2b est une forme semblable à A2a mais nettement étirée dans un diamètre du plan équatorial (diamètre long du corps : environ 140 µm). La symétrie bilatérale obtenue est quasi unique pour le groupe. Cet aspect à contour elliptique est souligné par une collerette plus étendue dans la longueur que dans la largeur. A2b est présente dès la base de la section (parfois plus de 1000 individus / 100 g) et accompagne la forme précédente entre les cotes 50 et 69.
![]() - A3 est une forme en disque avec un corps lenticulaire simple, semblable à une soucoupe volante, de profil symétrique où chaque face est un dôme bas d'environ 150 µm de diamètre. Ce corps est prolongé d'une large collerette équatoriale qui atteint parfois 60 µm, fine, transparente, de contour circulaire. Cette forme rare est présente dans 2 niveaux bas du Campanien "moyen".
- A3 est une forme en disque avec un corps lenticulaire simple, semblable à une soucoupe volante, de profil symétrique où chaque face est un dôme bas d'environ 150 µm de diamètre. Ce corps est prolongé d'une large collerette équatoriale qui atteint parfois 60 µm, fine, transparente, de contour circulaire. Cette forme rare est présente dans 2 niveaux bas du Campanien "moyen".
![]() - A4a est une grande forme en disque épais (près de 200 µm), de profil très dissymétrique. La face orale, très haute, tronconique, est marquée, sur son plateau, d'une structure en pustule au centre de laquelle se situe l'orifice, discret. La face aborale ne s'abaisse que légèrement en dessous de l'équateur puis s'invagine en une large dépression; il en résulte un profil rappelant celui d'un Kugelhopf (en Alsace) ou d'un pastis landais (pour se rapprocher de Tercis). L'équateur est marqué d'un bourrelet épais qui ceinture le disque et s'affine en un liseré transparent vers l'extérieur. Cette forme très reconnaissable, souvent légèrement repliée comme un béret rabattu sur les 2 oreilles, est présente en abondance dans les niveaux bas de la carrière. La discrétion de l'orifice ne permet pas une orientation immédiate. L'examen de spécimens plus ou moins attaqués par l'acide qui agrandit l'orifice situé au centre du "bouton" marquant l'une des faces, a solutionné le problème.
- A4a est une grande forme en disque épais (près de 200 µm), de profil très dissymétrique. La face orale, très haute, tronconique, est marquée, sur son plateau, d'une structure en pustule au centre de laquelle se situe l'orifice, discret. La face aborale ne s'abaisse que légèrement en dessous de l'équateur puis s'invagine en une large dépression; il en résulte un profil rappelant celui d'un Kugelhopf (en Alsace) ou d'un pastis landais (pour se rapprocher de Tercis). L'équateur est marqué d'un bourrelet épais qui ceinture le disque et s'affine en un liseré transparent vers l'extérieur. Cette forme très reconnaissable, souvent légèrement repliée comme un béret rabattu sur les 2 oreilles, est présente en abondance dans les niveaux bas de la carrière. La discrétion de l'orifice ne permet pas une orientation immédiate. L'examen de spécimens plus ou moins attaqués par l'acide qui agrandit l'orifice situé au centre du "bouton" marquant l'une des faces, a solutionné le problème.
![]() - A4b est une forme semblable à A4a mais son profil est sub-symétrique avec la face orale souvent dominante, renforcée qu'elle est par l'appareil péri-oral. A4b succède à A4a dans la section, les deux formes s'excluant stratigraphiquement ce qui confère à ce doublet de formes une intéressante implication biostratigraphique. A4b est souvent abondante dans la zone à Radotruncana calcarata et un peu plus haut (forme dominante avec 2.000 individus / 100 g).
- A4b est une forme semblable à A4a mais son profil est sub-symétrique avec la face orale souvent dominante, renforcée qu'elle est par l'appareil péri-oral. A4b succède à A4a dans la section, les deux formes s'excluant stratigraphiquement ce qui confère à ce doublet de formes une intéressante implication biostratigraphique. A4b est souvent abondante dans la zone à Radotruncana calcarata et un peu plus haut (forme dominante avec 2.000 individus / 100 g).
![]() - A6 est une grande forme en disque régulier (autour de 210 µm). Les deux faces tronconiques, de hauteur moyenne, donnent un profil symétrique marqué par un bourrelet équatorial aigu. L'orifice est au centre d'une légère dépression tandis que la face aborale est marquée par une collerette basse et large, péri-axiale. Cette forme n'est connue que dans le niveau le plus élevé de la carrière (cote 172).
- A6 est une grande forme en disque régulier (autour de 210 µm). Les deux faces tronconiques, de hauteur moyenne, donnent un profil symétrique marqué par un bourrelet équatorial aigu. L'orifice est au centre d'une légère dépression tandis que la face aborale est marquée par une collerette basse et large, péri-axiale. Cette forme n'est connue que dans le niveau le plus élevé de la carrière (cote 172).
![]() - A7 se présente comme une sorte de palet fin, plus pâle que les autres formes, de grande taille (près de 190 µm). Les 2 faces ne se distinguent que par la présence d'un orifice discret, circulaire, de diamètre inférieur à 20 µm sur la face orale. La forme est circonscrite aux quinze mètres inférieurs de la section de la Grande Carrière.
- A7 se présente comme une sorte de palet fin, plus pâle que les autres formes, de grande taille (près de 190 µm). Les 2 faces ne se distinguent que par la présence d'un orifice discret, circulaire, de diamètre inférieur à 20 µm sur la face orale. La forme est circonscrite aux quinze mètres inférieurs de la section de la Grande Carrière.
![]() - B1a est classée dans un groupe B caractérisé par un corps discoïde peu élevé, équipé de bras développés au niveau équatorial. B1a comporte 6 bras courts disposés régulièrement autour du corps de 180 µm de diamètre et perpendiculairement à l'axe de symétrie. Sur la face orale, l'orifice est porté par un léger col. Cette face est marquée par une légère dépression annulaire à mi-distance entre l'axe et la périphérie. La face aborale comporte le même arrangement. Le profil est symétrique par rapport au plan équatorial. La forme est rare dans un niveau situé à mi-carrière (cote 79,8). Des spécimens d'aspect très proche sont observés plus bas ; ils présentent tous 5 bras, régulièrement répartis eux-aussi. Le faible nombre de spécimens récoltés ne permet pas de décider s'il s'agit d'une variation intra-spécifique ou de deux formes distinctes à deux niveaux différents.
- B1a est classée dans un groupe B caractérisé par un corps discoïde peu élevé, équipé de bras développés au niveau équatorial. B1a comporte 6 bras courts disposés régulièrement autour du corps de 180 µm de diamètre et perpendiculairement à l'axe de symétrie. Sur la face orale, l'orifice est porté par un léger col. Cette face est marquée par une légère dépression annulaire à mi-distance entre l'axe et la périphérie. La face aborale comporte le même arrangement. Le profil est symétrique par rapport au plan équatorial. La forme est rare dans un niveau situé à mi-carrière (cote 79,8). Des spécimens d'aspect très proche sont observés plus bas ; ils présentent tous 5 bras, régulièrement répartis eux-aussi. Le faible nombre de spécimens récoltés ne permet pas de décider s'il s'agit d'une variation intra-spécifique ou de deux formes distinctes à deux niveaux différents.
![]() - B2a possède un corps en disque plus élevé que B1a et plus petit (diamètre autour de 130 µm). Sur la face orale, l'orifice est situé au sommet d'un dôme large. La face aborale a une forme identique, sans orifice. Une dépression annulaire étroite borde les deux faces et le profil est généralement symétrique. Typiquement 8 bras bien développés et en tube rayonnent dans le plan équatorial (cf.
Animation 2
- B2a possède un corps en disque plus élevé que B1a et plus petit (diamètre autour de 130 µm). Sur la face orale, l'orifice est situé au sommet d'un dôme large. La face aborale a une forme identique, sans orifice. Une dépression annulaire étroite borde les deux faces et le profil est généralement symétrique. Typiquement 8 bras bien développés et en tube rayonnent dans le plan équatorial (cf.
Animation 2 ![]() ). La forme est présente dans les 80 mètres inférieurs de la section de la Grande Carrière de Tercis. Des formes à 9 bras constituent plus de 50 % des spécimens dans certains niveaux. De rares spécimens comptent 7 bras, voire 10. B2a est la forme type du groupe des gilianelles sous le nom de Gilianella stellata (,
sous presse).
). La forme est présente dans les 80 mètres inférieurs de la section de la Grande Carrière de Tercis. Des formes à 9 bras constituent plus de 50 % des spécimens dans certains niveaux. De rares spécimens comptent 7 bras, voire 10. B2a est la forme type du groupe des gilianelles sous le nom de Gilianella stellata (,
sous presse).
![]() - B2b a un corps plus grand que B2a (diamètre autour de 150 µm). D'aspect très semblable, elle ne comporte que 4 bras. Les rares spécimens observés proviennent des niveaux bas de la carrière où la forme B2a, plus abondante, montre 8 bras de façon assez constante.
- B2b a un corps plus grand que B2a (diamètre autour de 150 µm). D'aspect très semblable, elle ne comporte que 4 bras. Les rares spécimens observés proviennent des niveaux bas de la carrière où la forme B2a, plus abondante, montre 8 bras de façon assez constante.
![]() Le groupe B comporte d'autres formes plus grandes et avec plus de bras (jusqu'à 15). Leur rareté et le nombre variable de bras observé pour B2a ne permettent pas encore de faire la part entre variations intra- et inter-spécifique.
Le groupe B comporte d'autres formes plus grandes et avec plus de bras (jusqu'à 15). Leur rareté et le nombre variable de bras observé pour B2a ne permettent pas encore de faire la part entre variations intra- et inter-spécifique.
![]() - C1a est classé dans un groupe C caractérisé par un corps en disque lenticulaire épais armé de bras équatoriaux petits. C1a, assez grande (diamètre du corps :190 µm), a un profil axial nettement dissymétrique dont la face orale très développée, tronconique, est couronnée d'un col haut et large, légèrement étranglé à la base. L'orifice est grand en haut du col. Le plan équatorial montre 6 bras courts en épine. La face aborale, en dôme bas, est déprimée au centre. C1a est rare dans les 11 premiers mètres de la section de la Grande Carrière de Tercis.
- C1a est classé dans un groupe C caractérisé par un corps en disque lenticulaire épais armé de bras équatoriaux petits. C1a, assez grande (diamètre du corps :190 µm), a un profil axial nettement dissymétrique dont la face orale très développée, tronconique, est couronnée d'un col haut et large, légèrement étranglé à la base. L'orifice est grand en haut du col. Le plan équatorial montre 6 bras courts en épine. La face aborale, en dôme bas, est déprimée au centre. C1a est rare dans les 11 premiers mètres de la section de la Grande Carrière de Tercis.
![]() - C1b est une forme très voisine (diamètre 160 µm) de C1a
(Pl. 3
- C1b est une forme très voisine (diamètre 160 µm) de C1a
(Pl. 3 ![]() ) mais son col n'est pas étranglé à la base et la face aborale ne paraît pas déprimée au centre. Typiquement, 5 bras en épine donnent au pourtour équatorial un aspect pentagonal. Des spécimens comportant 6 bras existent. Cette forme est distinguée car sa distribution est disjointe de celle de la précédente : C1b n'est connue que dans la zone à Radotruncana calcarata et a ainsi vécu environ 1,5 Ma après la forme C1a.
) mais son col n'est pas étranglé à la base et la face aborale ne paraît pas déprimée au centre. Typiquement, 5 bras en épine donnent au pourtour équatorial un aspect pentagonal. Des spécimens comportant 6 bras existent. Cette forme est distinguée car sa distribution est disjointe de celle de la précédente : C1b n'est connue que dans la zone à Radotruncana calcarata et a ainsi vécu environ 1,5 Ma après la forme C1a.
![]() - C2 possède un corps discoïde très élevé, presque sphérique à première vue, plus petit que C1 (autour de 120 µm). La face orale en dôme élevé, régulier, porte un orifice ourlé d'un léger col. La face aborale, un peu plus élevée, montre souvent un dôme moins régulier, avec un palier à mi-chemin de l'extrémité aborale. Cette dernière est marquée par une expansion basse et épaisse péri-apicale semblable à une pustule. Le plan équatorial comporte un nombre variable d'épines. Les spécimens à 7 bras dominent dans certains niveaux ; dans d'autres, ce sont les spécimens à 8 bras avec des individus à 9 bras voire jusqu'à 11. Cette forme est présente sur environ 30 mètres à la base de la section de Tercis.
- C2 possède un corps discoïde très élevé, presque sphérique à première vue, plus petit que C1 (autour de 120 µm). La face orale en dôme élevé, régulier, porte un orifice ourlé d'un léger col. La face aborale, un peu plus élevée, montre souvent un dôme moins régulier, avec un palier à mi-chemin de l'extrémité aborale. Cette dernière est marquée par une expansion basse et épaisse péri-apicale semblable à une pustule. Le plan équatorial comporte un nombre variable d'épines. Les spécimens à 7 bras dominent dans certains niveaux ; dans d'autres, ce sont les spécimens à 8 bras avec des individus à 9 bras voire jusqu'à 11. Cette forme est présente sur environ 30 mètres à la base de la section de Tercis.
![]() - D1a appartient à un groupe dont le corps, sans expansion ni équateur marqué, est globalement sphérique, de 160 à 200 µm de diamètre. Le test dont la cavité est sphérique, présente un découpage en pentagones ou hexagones rappelant l'assemblage d'un ballon de cuir cousu ou des cellules d'un rayon de miel. Un orifice légèrement à nettement détaché du reste de la sphère détermine un pôle oral. La forme représentée
Planche 3
- D1a appartient à un groupe dont le corps, sans expansion ni équateur marqué, est globalement sphérique, de 160 à 200 µm de diamètre. Le test dont la cavité est sphérique, présente un découpage en pentagones ou hexagones rappelant l'assemblage d'un ballon de cuir cousu ou des cellules d'un rayon de miel. Un orifice légèrement à nettement détaché du reste de la sphère détermine un pôle oral. La forme représentée
Planche 3 ![]() est caractérisée par un petit nombre de surfaces organisées latitudinalement. Elle est présente tout le long de la section. Les mêmes prélèvements livrent des sphères comportant un nombre de surfaces bien plus grand. Parfois, les bourrelets entre les surfaces se renforcent et s'arment d'excroissances transformant la sphère en bogue hirsute. Qu'il s'agisse d'une variation morphologique continue ou discontinue n'est pas encore établi. Par ailleurs, la singularité de ces restes évoque un groupe particulier qui s'intègre difficilement par rapport aux autres taxons présentés. L'étude de la nannostructure confirmera ce point (voir plus loin).
est caractérisée par un petit nombre de surfaces organisées latitudinalement. Elle est présente tout le long de la section. Les mêmes prélèvements livrent des sphères comportant un nombre de surfaces bien plus grand. Parfois, les bourrelets entre les surfaces se renforcent et s'arment d'excroissances transformant la sphère en bogue hirsute. Qu'il s'agisse d'une variation morphologique continue ou discontinue n'est pas encore établi. Par ailleurs, la singularité de ces restes évoque un groupe particulier qui s'intègre difficilement par rapport aux autres taxons présentés. L'étude de la nannostructure confirmera ce point (voir plus loin).
![]() - E1 fait partie momentanément d'un groupe (E) de formes dont le corps est en forme de cupule ; c'est-à-dire que la face orale est réduite à un plan légèrement concave ou convexe tandis que la face aborale est développée en coupe. La particularité du groupe composite E par rapport aux groupes F, G, H, est l'absence de bras développés. Le corps de E1 (diamètre et hauteur 150 µm) comporte une face orale élevée en volcan sommé d'un grand orifice (Ø 25 µm) souligné d'une lèvre étroite. Le plan équatorial ne montre pas d'expansion distincte mais quelques festons comme des amorces avortées. La face aborale comporte une collerette péri-axiale discrète. Cette petite forme lisse, au profil en urne, est présente en faible nombre dans les premiers mètres de la section de Tercis.
- E1 fait partie momentanément d'un groupe (E) de formes dont le corps est en forme de cupule ; c'est-à-dire que la face orale est réduite à un plan légèrement concave ou convexe tandis que la face aborale est développée en coupe. La particularité du groupe composite E par rapport aux groupes F, G, H, est l'absence de bras développés. Le corps de E1 (diamètre et hauteur 150 µm) comporte une face orale élevée en volcan sommé d'un grand orifice (Ø 25 µm) souligné d'une lèvre étroite. Le plan équatorial ne montre pas d'expansion distincte mais quelques festons comme des amorces avortées. La face aborale comporte une collerette péri-axiale discrète. Cette petite forme lisse, au profil en urne, est présente en faible nombre dans les premiers mètres de la section de Tercis.
![]() - E2b a la même forme en urne que E1 et une petite taille (diamètre proche de 100 µm). à la face orale, sub-plane, l'orifice est souligné par une lèvre d'où partent environ 8 crêtes (6 à 10) terminées par des amorces de bras en épine à la périphérie. La face aborale est caractérisée par des stries dont certaines rejoignent les épines notées plus haut. Quatre de ces stries se rassemblent au pôle aboral pour former un pied en ailettes. Cette petite forme striée n'est repérée que dans les niveaux bas de la carrière.
- E2b a la même forme en urne que E1 et une petite taille (diamètre proche de 100 µm). à la face orale, sub-plane, l'orifice est souligné par une lèvre d'où partent environ 8 crêtes (6 à 10) terminées par des amorces de bras en épine à la périphérie. La face aborale est caractérisée par des stries dont certaines rejoignent les épines notées plus haut. Quatre de ces stries se rassemblent au pôle aboral pour former un pied en ailettes. Cette petite forme striée n'est repérée que dans les niveaux bas de la carrière.
![]() - E3a est une grande forme (près de 180 µm en hauteur et diamètre) en forme d'urne striée. La face orale est semblable à celle de E2b ; l'orifice de diamètre 40 µm, est entouré d'une lèvre d'où partent 7 à 10 rayons formant crêtes. à la périphérie, une étroite collerette est visible. Sur la face aborale, les crêtes se prolongent sur une collerette péri-axiale. L'ensemble donne un profil en pot de fleur à flanc grossièrement tronconique. E3a se récolte dans les mêmes niveaux que la forme précédente.
- E3a est une grande forme (près de 180 µm en hauteur et diamètre) en forme d'urne striée. La face orale est semblable à celle de E2b ; l'orifice de diamètre 40 µm, est entouré d'une lèvre d'où partent 7 à 10 rayons formant crêtes. à la périphérie, une étroite collerette est visible. Sur la face aborale, les crêtes se prolongent sur une collerette péri-axiale. L'ensemble donne un profil en pot de fleur à flanc grossièrement tronconique. E3a se récolte dans les mêmes niveaux que la forme précédente.
![]() - E3b se distingue de la précédente par un corps de petite taille (diamètre : 120 µm) et de forme plutôt globuleuse. Elle est striée et se prolonge, dans la zone péri-axiale de la face aborale, d'une collerette formant jupe. La forme se trouve dans les premiers mètres de la section. Il n'est pas exclu que des intermédiaires existent. En outre, des formes voisines, plus rares, ont été observées.
- E3b se distingue de la précédente par un corps de petite taille (diamètre : 120 µm) et de forme plutôt globuleuse. Elle est striée et se prolonge, dans la zone péri-axiale de la face aborale, d'une collerette formant jupe. La forme se trouve dans les premiers mètres de la section. Il n'est pas exclu que des intermédiaires existent. En outre, des formes voisines, plus rares, ont été observées.
![]() - F1 est caractérisée par un corps assez grand (autour de 160 µm) en cupule lisse armée de 5 bras (localement plutôt 6, exceptionnellement 4 ou 7) forts et courts, tubulaires. Sur la face orale en dôme bas, lisse, l'orifice est discret. La face aborale en dôme élevé, légèrement conique, laisse supposer le départ d'un pied axial très court. La forme est rare mais constante dans les niveaux de la zone à Radotruncana calcarata entre les cotes 47 et 58.
- F1 est caractérisée par un corps assez grand (autour de 160 µm) en cupule lisse armée de 5 bras (localement plutôt 6, exceptionnellement 4 ou 7) forts et courts, tubulaires. Sur la face orale en dôme bas, lisse, l'orifice est discret. La face aborale en dôme élevé, légèrement conique, laisse supposer le départ d'un pied axial très court. La forme est rare mais constante dans les niveaux de la zone à Radotruncana calcarata entre les cotes 47 et 58.
![]() - G0 fait partie d'un groupe de petites formes (diamètre 110 µm) au corps nettement sphérique, caractérisé par la présence constante de bras nets et d'une expansion aborale. G0 montre typiquement 6 bras fins, probablement tubulaires. La face orale est presque hémisphérique et l'orifice axial est mis en évidence par une lèvre peu élevée. La face aborale comporte une expansion péri-axiale en collerette fine. 1 ou 2 ébauches de "rame" sont parfois observables. Les quelques éléments répondant à cette description rendue difficile par la taille réduite des spécimens proviennent des 25 premiers mètres de la section de Tercis.
- G0 fait partie d'un groupe de petites formes (diamètre 110 µm) au corps nettement sphérique, caractérisé par la présence constante de bras nets et d'une expansion aborale. G0 montre typiquement 6 bras fins, probablement tubulaires. La face orale est presque hémisphérique et l'orifice axial est mis en évidence par une lèvre peu élevée. La face aborale comporte une expansion péri-axiale en collerette fine. 1 ou 2 ébauches de "rame" sont parfois observables. Les quelques éléments répondant à cette description rendue difficile par la taille réduite des spécimens proviennent des 25 premiers mètres de la section de Tercis.
![]() - G1 est une forme au corps petit (diamètre : 120 µm). La face orale en dôme bas comporte une ornementation voisine de celles de E2 et E3. Les crêtes radiales se prolongent et arment des bras forts à section cruciforme généralement par 6 à 7 (de rares individus n'ont que 4 bras inégalement répartis tandis que d'autres présentent, rarement, jusqu'à 10 bras). La face aborale est ridée (rides allongées axialement) et se prolonge d'un pied net, lui aussi à section cruciforme et dont la taille atteint parfois celle du corps. Le profil est celui d'un artichaut orné de cornes. La forme présente et commune dans toute la partie médiane de la section de Tercis, comporte peut-être des variétés non distinguées jusqu'à présent.
- G1 est une forme au corps petit (diamètre : 120 µm). La face orale en dôme bas comporte une ornementation voisine de celles de E2 et E3. Les crêtes radiales se prolongent et arment des bras forts à section cruciforme généralement par 6 à 7 (de rares individus n'ont que 4 bras inégalement répartis tandis que d'autres présentent, rarement, jusqu'à 10 bras). La face aborale est ridée (rides allongées axialement) et se prolonge d'un pied net, lui aussi à section cruciforme et dont la taille atteint parfois celle du corps. Le profil est celui d'un artichaut orné de cornes. La forme présente et commune dans toute la partie médiane de la section de Tercis, comporte peut-être des variétés non distinguées jusqu'à présent.
![]() - H2 appartient au groupe H caractérisé par des formes au corps en cupule et aux bras et pieds tubulaires multiples. Les gilianelles atteignent ici le sommet de leur complexité morphologique. Le corps minuscule de H2 ne dépasse pas 80 µm de diamètre. La face orale est plutôt conique, basse ; elle est terminée par un orifice aussi gros que celui des autres formes (autour de 20 µm). Un ensemble de 7 à 9 bras inégalement développés mais toujours longs part de l'équateur et se dirige légèrement vers le haut. La face aborale, tronconique, courte, se termine par une zone dans laquelle s'insèrent 2 à 5 pieds péri-axiaux, aussi forts que les bras et dont le développement et la disposition ne sont pas réguliers. H2 existe en petit nombre dans la zone à Radotruncana calcarata et un peu au dessus.
- H2 appartient au groupe H caractérisé par des formes au corps en cupule et aux bras et pieds tubulaires multiples. Les gilianelles atteignent ici le sommet de leur complexité morphologique. Le corps minuscule de H2 ne dépasse pas 80 µm de diamètre. La face orale est plutôt conique, basse ; elle est terminée par un orifice aussi gros que celui des autres formes (autour de 20 µm). Un ensemble de 7 à 9 bras inégalement développés mais toujours longs part de l'équateur et se dirige légèrement vers le haut. La face aborale, tronconique, courte, se termine par une zone dans laquelle s'insèrent 2 à 5 pieds péri-axiaux, aussi forts que les bras et dont le développement et la disposition ne sont pas réguliers. H2 existe en petit nombre dans la zone à Radotruncana calcarata et un peu au dessus.
![]() - J1 appartient au groupe J caractérisé par des expansions (bras et pieds) en forme de palette
(Pl. 5
- J1 appartient au groupe J caractérisé par des expansions (bras et pieds) en forme de palette
(Pl. 5 ![]() ). Le corps a environ 120 µm de diamètre. La face orale est peu élevée; la face aborale l'est beaucoup plus. 4 bras perpendiculaires partent de l'équateur et se dirigent très légèrement vers le haut; ils sont reliés par une étroite collerette transparente comme les bras. La base de la face aborale est marquée par une collerette péri-apicale prolongée ou non de 1 à 3 bras fragiles. Cette forme est présente dans 2 niveaux situés environ 5 et 11 m au dessus de la cote 0 de la section de Tercis.
). Le corps a environ 120 µm de diamètre. La face orale est peu élevée; la face aborale l'est beaucoup plus. 4 bras perpendiculaires partent de l'équateur et se dirigent très légèrement vers le haut; ils sont reliés par une étroite collerette transparente comme les bras. La base de la face aborale est marquée par une collerette péri-apicale prolongée ou non de 1 à 3 bras fragiles. Cette forme est présente dans 2 niveaux situés environ 5 et 11 m au dessus de la cote 0 de la section de Tercis.
![]() - J2 est une forme similaire à J1 mais avec plus de bras (5 à 8) répartis très inégalement autour de l'équateur. La face aborale est moins élevée que celle de la forme J1 donnant à ces tests un aspect plus aplati
(Pl. 5
- J2 est une forme similaire à J1 mais avec plus de bras (5 à 8) répartis très inégalement autour de l'équateur. La face aborale est moins élevée que celle de la forme J1 donnant à ces tests un aspect plus aplati
(Pl. 5 ![]() ). Elles sont présentes dans tous les niveaux traités entre 10 et 25 m au dessus de la cote 0 de la section de Tercis (avec jusqu'à 100-200 individus / 100 g).
). Elles sont présentes dans tous les niveaux traités entre 10 et 25 m au dessus de la cote 0 de la section de Tercis (avec jusqu'à 100-200 individus / 100 g).
![]() - K1 est classé dans un groupe dont le corps a la forme d'une punaise qui a, ici, un diamètre et une hauteur proches de 200 µm. La face orale est en dôme bas, régulier; son sommet montre un orifice (?) axial au contour incertain. La face aborale est en cône aux flancs concaves. Il n'y a pas de cavité individualisée ni de paroi dans ces restes et le rattachement aux gilianelles est incertain. K1 est assez commun dans le niveau 79,8 de la section de Tercis (~1000 individus/ 100 g).
- K1 est classé dans un groupe dont le corps a la forme d'une punaise qui a, ici, un diamètre et une hauteur proches de 200 µm. La face orale est en dôme bas, régulier; son sommet montre un orifice (?) axial au contour incertain. La face aborale est en cône aux flancs concaves. Il n'y a pas de cavité individualisée ni de paroi dans ces restes et le rattachement aux gilianelles est incertain. K1 est assez commun dans le niveau 79,8 de la section de Tercis (~1000 individus/ 100 g).
![]() - K2a présente un corps plus petit (diamètre : 150 µm; hauteur : 160 µm) dont la face aborale présente un flanc légèrement convexe. Sans cavité ni paroi distincte, son appartenance au groupe des gilianelles est aussi incertaine. K2a est rare dans un niveau bas de la section.
- K2a présente un corps plus petit (diamètre : 150 µm; hauteur : 160 µm) dont la face aborale présente un flanc légèrement convexe. Sans cavité ni paroi distincte, son appartenance au groupe des gilianelles est aussi incertaine. K2a est rare dans un niveau bas de la section.
![]() - L1 est une forme au corps petit (diamètre et hauteur autour de 100 µm), en cône inversé. La face orale est légèrement concave avec son orifice axial peu marqué (pas plus de 10 µm quand on peut le deviner). Une collerette transparente part de la périphérie du corps et prolonge la concavité de la face orale vers le haut. La face aborale est prolongée par un pied axial aussi long que le corps, solide, plein. Le profil obtenu est celui d'un entonnoir. Curieusement, l'extrémité du pied est systématiquement déjetée par rapport à l'axe dans certains niveaux ; dans d'autres il est approximativement axial. Cette curieuse forme est présente constamment et commune voire abondante (5 à 10.000 individus / 100 g) depuis la cote -5 de la section jusqu'à la base de la zone à Rd. calcarata.
- L1 est une forme au corps petit (diamètre et hauteur autour de 100 µm), en cône inversé. La face orale est légèrement concave avec son orifice axial peu marqué (pas plus de 10 µm quand on peut le deviner). Une collerette transparente part de la périphérie du corps et prolonge la concavité de la face orale vers le haut. La face aborale est prolongée par un pied axial aussi long que le corps, solide, plein. Le profil obtenu est celui d'un entonnoir. Curieusement, l'extrémité du pied est systématiquement déjetée par rapport à l'axe dans certains niveaux ; dans d'autres il est approximativement axial. Cette curieuse forme est présente constamment et commune voire abondante (5 à 10.000 individus / 100 g) depuis la cote -5 de la section jusqu'à la base de la zone à Rd. calcarata.
![]() - M1 fait partie d'un groupe caractérisé par un corps de grande taille, franchement conique, nettement strié et dont la face orale est parcourue de crêtes radiales partant d'une lèvre péri-orale élevée. La forme M1 (diamètre et hauteur autour de 180 µm) est dépourvue de collerette équatoriale ; la face aborale est formée d'un cône élevé, strié, dont le profil, légèrement convexe, ne se prolonge pas par un pied.
- M1 fait partie d'un groupe caractérisé par un corps de grande taille, franchement conique, nettement strié et dont la face orale est parcourue de crêtes radiales partant d'une lèvre péri-orale élevée. La forme M1 (diamètre et hauteur autour de 180 µm) est dépourvue de collerette équatoriale ; la face aborale est formée d'un cône élevé, strié, dont le profil, légèrement convexe, ne se prolonge pas par un pied.
![]() - M2a a généralement une taille plus élevée que M1 (diamètre et hauteur dépassant parfois 200 µm). La face orale, similaire à celle de M1, se prolonge par une collerette entière, armée du prolongement des crêtes radiales. La face aborale, conique, se prolonge d'un pied tubulaire large (diamètre : 40 µm) armé d'ailettes ; celles-ci prolongent les crêtes qui déterminent la striation de cette face.
- M2a a généralement une taille plus élevée que M1 (diamètre et hauteur dépassant parfois 200 µm). La face orale, similaire à celle de M1, se prolonge par une collerette entière, armée du prolongement des crêtes radiales. La face aborale, conique, se prolonge d'un pied tubulaire large (diamètre : 40 µm) armé d'ailettes ; celles-ci prolongent les crêtes qui déterminent la striation de cette face.
![]() - M2b est proche de M2a
(Pl. 6
- M2b est proche de M2a
(Pl. 6 ![]() ) mais les crêtes radiales de la face orale s'interrompent au niveau de la collerette équatoriale, entière. Le pied atteint une longueur de 100 µm donnant à certains spécimens l'aspect d'une girolle.
) mais les crêtes radiales de la face orale s'interrompent au niveau de la collerette équatoriale, entière. Le pied atteint une longueur de 100 µm donnant à certains spécimens l'aspect d'une girolle.
![]() - M3 est une forme de même aspect et taille que M2a. La collerette équatoriale se prolonge au niveau des crêtes radiales de la face orale : son contour devient découpé. Certains spécimens donnent l'impression d'avoir des bras larges à la base. Le pied est parcouru de crêtes développées en ailettes (généralement 5). Cette forme très ornée est présente à la cote 69 de la section de Tercis où tous les spécimens sont atteints d'un foisonnement des expansions.
- M3 est une forme de même aspect et taille que M2a. La collerette équatoriale se prolonge au niveau des crêtes radiales de la face orale : son contour devient découpé. Certains spécimens donnent l'impression d'avoir des bras larges à la base. Le pied est parcouru de crêtes développées en ailettes (généralement 5). Cette forme très ornée est présente à la cote 69 de la section de Tercis où tous les spécimens sont atteints d'un foisonnement des expansions.
![]() - M5 est une forme dont la taille est voisine de 200 µm. Elle est caractérisée par des crêtes péri-orales très nombreuses, serrées, peu distinctes ; certaines (5 à 9), plus longues, dépassent de la masse péri-orale et atteignent le début de la collerette équatoriale où elles s'interrompent. Cette collerette demeure ainsi sans renfort. Cette forme existe dans les niveaux terminaux de la section de la Grande Carrière de Tercis. La répartition relative des formes M reste à préciser car il n'est pas aisé de les distinguer lors du tri sous la loupe binoculaire. Le groupe M est présent dans toute la coupe et même au delà dans les niveaux du Maastrichtien supérieur en dehors de la carrière ( & ,
2001b).
- M5 est une forme dont la taille est voisine de 200 µm. Elle est caractérisée par des crêtes péri-orales très nombreuses, serrées, peu distinctes ; certaines (5 à 9), plus longues, dépassent de la masse péri-orale et atteignent le début de la collerette équatoriale où elles s'interrompent. Cette collerette demeure ainsi sans renfort. Cette forme existe dans les niveaux terminaux de la section de la Grande Carrière de Tercis. La répartition relative des formes M reste à préciser car il n'est pas aisé de les distinguer lors du tri sous la loupe binoculaire. Le groupe M est présent dans toute la coupe et même au delà dans les niveaux du Maastrichtien supérieur en dehors de la carrière ( & ,
2001b).
![]() - N1 est une gilianelle dont la forme ne permet pas de définir un équateur. Son test est petit (diamètre : 125 µm), marqué par une face orale très peu convexe séparée d'une face aborale par un flanc haut coupant perpendiculairement ces deux faces. La striation de ces flancs, visible à la loupe binoculaire, distingue les formes du groupe N des abondantes structures sphériques de même taille observées dans les résidus d'acétolyse (essentiellement des pithonelles, et alii,
2001). Le profil est celui d'un cylindre en tonnelet court, terminé, aux deux extrémités, d'un dôme bas. La portion aborale du cylindre comporte un léger étranglement. La distribution est large mais malaisée à établir car les spécimens sont discrets parmi les abondantes pithonelles.
- N1 est une gilianelle dont la forme ne permet pas de définir un équateur. Son test est petit (diamètre : 125 µm), marqué par une face orale très peu convexe séparée d'une face aborale par un flanc haut coupant perpendiculairement ces deux faces. La striation de ces flancs, visible à la loupe binoculaire, distingue les formes du groupe N des abondantes structures sphériques de même taille observées dans les résidus d'acétolyse (essentiellement des pithonelles, et alii,
2001). Le profil est celui d'un cylindre en tonnelet court, terminé, aux deux extrémités, d'un dôme bas. La portion aborale du cylindre comporte un léger étranglement. La distribution est large mais malaisée à établir car les spécimens sont discrets parmi les abondantes pithonelles.
![]() - N2b est une forme morphologiquement plus proche de celle des pithonelles. Sa taille est petite avec un diamètre autour de 140 µm. Son profil est celui d'une sphère aplatie au pôle oral seulement, lequel est occupé par un orifice de diamètre voisin de 20 µm. En vue polaire, des stries concentriques entourent l'orifice tandis que la périphérie est finement découpée d'une quarantaine de cannelures dues aux ridules des flancs allongées axialement.
- N2b est une forme morphologiquement plus proche de celle des pithonelles. Sa taille est petite avec un diamètre autour de 140 µm. Son profil est celui d'une sphère aplatie au pôle oral seulement, lequel est occupé par un orifice de diamètre voisin de 20 µm. En vue polaire, des stries concentriques entourent l'orifice tandis que la périphérie est finement découpée d'une quarantaine de cannelures dues aux ridules des flancs allongées axialement.
L'examen au microscope électronique permet d'observer des caractères nannostructuraux (nannostructure désigne l'échelle de quelques microns) qui vont se montrer déterminants pour caractériser le groupe des gilianelles parmi les incertae sedis illustrés. Trois caractères sont abordés : la paroi, l'aspect de surface, les expansions.
3.1. Paroi du test
La Planche
7 ![]() rassemble une sélection de vues de la paroi des incertae sedis de Tercis qui est représentative de celles observées sur la vaste majorité des formes. Cette paroi est épaisse de 15 à 20 µm ; la même épaisseur est visible dans une section de la base d'un bras de la forme F1
(Pl. 7, fig. 6
rassemble une sélection de vues de la paroi des incertae sedis de Tercis qui est représentative de celles observées sur la vaste majorité des formes. Cette paroi est épaisse de 15 à 20 µm ; la même épaisseur est visible dans une section de la base d'un bras de la forme F1
(Pl. 7, fig. 6 ![]() ). Notre estimation antérieure de 7 à 10 µm d'après les observations optiques (,
sous presse) était sous estimée. L'organisation des cristallites de la paroi est constante avec des éléments orientés perpendiculairement à la surface. On peut donc parler d'homogénéité de la structure de la paroi de ces tests.
). Notre estimation antérieure de 7 à 10 µm d'après les observations optiques (,
sous presse) était sous estimée. L'organisation des cristallites de la paroi est constante avec des éléments orientés perpendiculairement à la surface. On peut donc parler d'homogénéité de la structure de la paroi de ces tests.
En dehors de ces observations communes aux 10 groupes A, B, C, E, F, G, H, J, M, N, quatre formes présentent des nannostructures différentes
(Pl. 8 ![]() ). Dans la forme L1, la paroi des tests reste épaisse de 15 à 20 µm mais elle est constituée de deux couches distinctes. La couche extérieure ne montre pas d'organisation claire tandis que la couche intérieure est formée de cristallites perpendiculaires à la surface
(Pl. 8, fig. 6
). Dans la forme L1, la paroi des tests reste épaisse de 15 à 20 µm mais elle est constituée de deux couches distinctes. La couche extérieure ne montre pas d'organisation claire tandis que la couche intérieure est formée de cristallites perpendiculaires à la surface
(Pl. 8, fig. 6 ![]() ). Vus de l'extérieur, les cristallites de la paroi de la face orale présentent un arrangement dessinant des croix en surface
(Pl. 8, fig. 4
). Vus de l'extérieur, les cristallites de la paroi de la face orale présentent un arrangement dessinant des croix en surface
(Pl. 8, fig. 4 ![]() ). Dans le pied, sous la couche extérieure, les cristallites sont organisés en fibres allongées dans le sens de l'axe
(Pl. 8, fig. 5
). Dans le pied, sous la couche extérieure, les cristallites sont organisés en fibres allongées dans le sens de l'axe
(Pl. 8, fig. 5 ![]() ).
).
Pour le groupe D, la paroi est toujours constituée de cristallites allongés perpendiculaires à la surface mais elle est nettement plus épaisse (25 à 30 µm
: Pl. 8, fig. 3 ![]() ).
Enfin, la paroi du test n'a pu être examinée pour le groupe K. Nous proposons une explication : on a noté plus haut que les restes carbonatés de K1 provenaient du niveau 79,8 ; cependant, un reste exactement semblable (en punaise avec une trace d'orifice? au centre de la face orale) a été observée à la cote 69 mais sous forme glauconitisée. Or, la glauconie est commune en remplissage de tests dans la section de Tercis. Cette trouvaille suggère que les formes du groupe K seraient des moulages internes de tests particulièrement fragiles puisque toujours dissous.
).
Enfin, la paroi du test n'a pu être examinée pour le groupe K. Nous proposons une explication : on a noté plus haut que les restes carbonatés de K1 provenaient du niveau 79,8 ; cependant, un reste exactement semblable (en punaise avec une trace d'orifice? au centre de la face orale) a été observée à la cote 69 mais sous forme glauconitisée. Or, la glauconie est commune en remplissage de tests dans la section de Tercis. Cette trouvaille suggère que les formes du groupe K seraient des moulages internes de tests particulièrement fragiles puisque toujours dissous.
3.2. Ornementation externe des tests
Nous distinguerons maintenant deux sortes de tests : ceux des gilianelles à l'ornementation riche et les autres tests. L'ornementation des gilianelles comporte souvent des microstructures radiales ou concentriques qui se combinent pour donner à chaque forme un aspect extérieur propre sous la loupe binoculaire. L'observation au microscope électronique révèle que ces microstructures sont liées à un arrangement particulier de remarquables nannostructures dessinant un réseau tel un véritable napperon au crochet. La forme A1a
(Pl. 9, fig. 1 ![]() ) est caractéristique de ce type de double ornementation : nannostructure en réseau visible seulement au microscope électronique dessinant une ornementation radiale ou concentrique visible à la loupe. Des arrangements diversifiés ont été observés selon les formes. Toujours très discrètes et peu visibles à la loupe, les microstructures sont souvent absente. Ainsi, dans la forme A7, la surface paraît lisse à la loupe tandis que le microscope électronique révèle une belle nannostructure en grille
(Pl. 9, fig. 2
) est caractéristique de ce type de double ornementation : nannostructure en réseau visible seulement au microscope électronique dessinant une ornementation radiale ou concentrique visible à la loupe. Des arrangements diversifiés ont été observés selon les formes. Toujours très discrètes et peu visibles à la loupe, les microstructures sont souvent absente. Ainsi, dans la forme A7, la surface paraît lisse à la loupe tandis que le microscope électronique révèle une belle nannostructure en grille
(Pl. 9, fig. 2 ![]() ). Le point commun nouveau révélé par le microscope électronique est une nannostructure en napperon très distincte en surface des tests de gilianelles.
). Le point commun nouveau révélé par le microscope électronique est une nannostructure en napperon très distincte en surface des tests de gilianelles.
Certaines formes illustrées ici n'en comportent pas. Ainsi, pour le groupe D, les bourrelets arrangés en rayon de miel ne sont pas liés à l'arrangement particulier d'une nannostructure ; il n'y a pas de nannostructure en napperon, observation qui s'ajoute à celle d'une paroi plus épaisse que celle des autres formes présentées. De même, les tests des formes E1, F1 et L1 ne montrent pas de nannostructure organisée bien visible en surface. Enfin, les restes des formes du groupe K
(Pl. 8, fig. 1 ![]() ) ne présentent pas de nannostructure organisée en surface, observation qui rejoint l'absence de paroi.
) ne présentent pas de nannostructure organisée en surface, observation qui rejoint l'absence de paroi.
Il reste que 30 des 36 formes illustrées dans ce travail ont acquis un nouvelle caractéristique commune grâce à l'examen au microscope électronique à balayage.
3.3. Les expansions
La présence ou l'absence, la forme, la position et la structure des expansions qui ornent les tests de gilianelles permettent de les distinguer entre elles. Les quelques exemples illustrés dans la
Planche 9 ![]() précisent les caractéristiques des divers types d'expansion. Bras et pieds peuvent être des cornières à section cruciforme
(Pl. 9, figs. 4-5
précisent les caractéristiques des divers types d'expansion. Bras et pieds peuvent être des cornières à section cruciforme
(Pl. 9, figs. 4-5 ![]() ). Certaines expansions (bras ou pieds) sont cylindriques et creuses
(forme F1 : Pl. 7, fig. 6
). Certaines expansions (bras ou pieds) sont cylindriques et creuses
(forme F1 : Pl. 7, fig. 6 ![]() ). Chez L1 qui ne présente pas les nannostructures des gilianelles, le pied est plein
(Pl. 8, fig. 5
). Chez L1 qui ne présente pas les nannostructures des gilianelles, le pied est plein
(Pl. 8, fig. 5 ![]() ). D'autres expansions sont des tubes renforcés d'ailettes
(Pl. 9, fig. 3
). D'autres expansions sont des tubes renforcés d'ailettes
(Pl. 9, fig. 3 ![]() ) ; d'autres encore sont des palettes
(Pl. 9, fig. 6
) ; d'autres encore sont des palettes
(Pl. 9, fig. 6 ![]() ). Quant aux collerettes, elles sont fines et se situent soit à la limite face orale et face aborale, soit à la base de la face aborale auquel cas elles sont plus ou moins développées depuis un simple bourrelet péri-axial jusqu'à une véritable jupe. Elles peuvent, aussi être prolongées de palettes
(Pl. 9, fig. 6
). Quant aux collerettes, elles sont fines et se situent soit à la limite face orale et face aborale, soit à la base de la face aborale auquel cas elles sont plus ou moins développées depuis un simple bourrelet péri-axial jusqu'à une véritable jupe. Elles peuvent, aussi être prolongées de palettes
(Pl. 9, fig. 6 ![]() ).
).
4.1. Morphologie du test et milieu de vie
L'orientation d'un test libre en milieu marin est déterminée par la position relative de leurs centres de gravité et de poussée. Le centre de gravité est le point d'équilibre des masses composant le test. Le centre de poussée (d') d'un corps plongé dans un liquide est le point où toutes les forces génèrent l'effort de poussée c'est-à-dire le centre de gravité du fluide déplacé. Prenons la forme lisse L1 ; le long de l'axe de symétrie, le centre de gravité du test est éloigné de l'orifice, près du pied plein qui fait quille. Le centre de poussée est plus près de l'orifice; dès que le test penche d'un côté, il est immédiatement remis par la poussée d' avec l'orifice vers le haut et le pied vers le bas. Une observation similaire pour beaucoup de formes nous a conduit à orienter tous les tests avec la face orale vers le haut.
Pour les formes à expansions et ornementation, celles-ci jouent leur rôle dans l'équilibre des masses du test. Sur la face orale, ces ornementations sont souvent moins prononcées que sur la face aborale laquelle est, le plus souvent, plus développée. Plus dense, elle abaisse le centre de gravité de son côté. Les formes A1b, A2, C2, illustrent ce point de vue : les faces aborales montrent des escaliers, des collerettes, des renforts en bouton absents sur l'autre face. Si ces formes sont plongées dans un liquide, la face orale se positionnera vers le haut.
Dans les formes G, L, M, la géométrie du test est telle que la face orale, quasi plane, s'oppose à la face aborale qui s'organise en un véritable plomb de rappel qui se dirigera naturellement vers le bas
(Fig. 6 ![]() et
Animation 3
et
Animation 3 ![]() ).
).
Pour les formes E ou N, l'orifice est situé sur une face plus plane tandis que la face aborale est plus bombée avec parfois un large dôme apical qui abaisse le centre de gravité par rapport au centre de poussée; le test se positionnera ainsi, naturellement, l'orifice vers le haut s'il est plongé dans un liquide.
Dans le cas des formes à bras et pieds (G, H, J), alors que les bras sont dans le plan de la face orale ou légèrement redressés, l'orientation des pieds est nettement rapprochée de celle de l'axe de symétrie ce qui doit conduire naturellement à orienter le test plongé dans l'eau avec les pieds vers le bas. De même, lorsque les formes de type B ont des bras dans le plan équatorial, il n'existe pas de pieds mais seulement un dôme aboral large; au contraire, dès que les bras sont plus redressés, un pied apparaît dans l'axe de symétrie pour compenser la masse des bras.
La logique d'équilibre ainsi mise en évidence suggère que ces formes étaient effectivement libres en milieu liquide avec les dispositions adaptatives nécessaires pour maintenir une position constante. Une adaptation à la vie marine planctonique est donc probable, adaptation renforcée par la localisation de l'orifice car celle-ci implique, si l'on admet que ces tests sont ceux d'êtres unicellulaires, la présence de cytoplasme pouvant dépasser par cet orifice et, par sa densité moindre que celle du test, renforcer la position d'équilibre l'orifice vers le haut.
Une autre adaptation à la vie planctonique ou, du moins, à une mobilisation passive est la flottabilité. Les formes du plancton sont communément équipées d'appendices augmentant la surface de contact avec l'eau. Avec leurs expansions, leur fine collerette équatoriale ou péri-apicale, leurs stries ou ailettes et leur surface découpée, la plupart des gilianelles montrent une tendance à multiplier la surface de contact avec le fluide ambiant; ceci facilite leur maintien au sein des eaux et augmente la prise offerte aux courants susceptibles de les disperser.
Quelques formes (A4, C1, D) ne présentent pas cette adaptation. On peut suggérer qu'elles auraient été benthiques avec la face aborale disposée contre un support tandis que l'orifice est généralement porté par un long col ou un bouton afin de l'éloigner du substrat et favoriser la fonction nutritive. Rappelons que les formes A4 et C1 appartiennent clairement aux gilianelles par la nature de leurs nannostructures de surface ce qui n'est pas le cas de la forme D.
4.2. Durée d'existence
La durée d'existence des diverses formes illustrées est très variable. La
Figure 7 ![]() utilise, les conclusions proposées par
(2001b) améliorées d'après les corrélations fines Europe-Amérique du Nord établies d'après l'étude des inocérames ( et ,
2003). D'après la datation numérique retenue, la forme A1a aurait une durée d'existence supérieure à 8 Ma. Il n'est pas exclu qu'une étude plus détaillée conduise à tronçonner cet apparent continuum. Plus communes sont les formes dont la présence n'est attestée que sur une durée de 2 à 3 millions d'années telles A2b, A4a, B2a, L1. Les tests nommés ici A3, A4b, A7, F1, H2 sont présents en nombre suffisant pour affirmer que leur existence dans le bassin de dépôt aturien n'a pas dépassé 1 Ma. Cette durée est équivalente à celle de l'une des plus courtes biozones de foraminifères planctoniques du Crétacé Supérieur. Celui-ci : Radotruncana calcarata, par sa forme reconnaissable et sa courte durée d'existence (moins de 1 Ma) est considéré comme un excellent marqueur stratigraphique. Il y aurait donc, parmi les microfossiles énigmatiques illustrés dans la section de Tercis, un grand nombre de marqueurs stratigraphiques remarquablement précis
mais locaux pour l'instant.
utilise, les conclusions proposées par
(2001b) améliorées d'après les corrélations fines Europe-Amérique du Nord établies d'après l'étude des inocérames ( et ,
2003). D'après la datation numérique retenue, la forme A1a aurait une durée d'existence supérieure à 8 Ma. Il n'est pas exclu qu'une étude plus détaillée conduise à tronçonner cet apparent continuum. Plus communes sont les formes dont la présence n'est attestée que sur une durée de 2 à 3 millions d'années telles A2b, A4a, B2a, L1. Les tests nommés ici A3, A4b, A7, F1, H2 sont présents en nombre suffisant pour affirmer que leur existence dans le bassin de dépôt aturien n'a pas dépassé 1 Ma. Cette durée est équivalente à celle de l'une des plus courtes biozones de foraminifères planctoniques du Crétacé Supérieur. Celui-ci : Radotruncana calcarata, par sa forme reconnaissable et sa courte durée d'existence (moins de 1 Ma) est considéré comme un excellent marqueur stratigraphique. Il y aurait donc, parmi les microfossiles énigmatiques illustrés dans la section de Tercis, un grand nombre de marqueurs stratigraphiques remarquablement précis
mais locaux pour l'instant.
4.3. Nature des microfossiles incertae sedis de la section de Tercis
Depuis Juin 2005, nous avons fait circuler dans la communauté des paléontologistes, des images des formes décrites ici. Une grande perplexité et une variété d'affiliations avec des groupes connus a été proposée par les correspondants auxquels nous nous sommes adressés. Après discussion des diverses possibilités, nous avons conclu (, sous presse) qu'aucune des affiliations suggérées : les micro-crinoïdes ( 1999 ; & , 2001), les dinoflagellés calcaires (par exemple Heptasphaera michaeli : & , 1994), les Pseudarcellidés -autres problematica du Tertiaire- (, 1989), les radiolaires calcitisés (comm. personnelle T. ), les bryozoaires (, 1990, 1991) n'était convaincante. Nous ne reprendrons la question ici que pour ajouter qu'il ne s'agit pas, non plus, de thécamoebiens dont le test n'est ni calcaire ni ornementé comme celui des gilianelles ( et alii, 2003). En outre, les formes D, qui ne montrent pas tous les caractères des gilianelles pourraient être rapprochées de certains foraminifères comme Favulina hexagona () tel qu'illustré par (2006, fig. 28) ; la taille est similaire (l'échelle donnée par la figure citée est trop grande) mais la paroi du test ne dépasse pas 4 µm d'après la figure 43 du même travail alors que celle de la forme D atteint 30 µm.
La plupart des tests présentés suggèrent qu'ils ont été sécrétés pour abriter un être très simple, plus petit que la majorité des foraminifères. Ils vivaient au large, sur le rebord externe du plateau continental du Bassin Aturien ( et alii, 2001). Ce type de test et cette taille suggèrent l'Embranchement des Protozoaires. A l'intérieur de ceux-ci, la Classe des Rhizopodes est possible puisqu'on y trouve des groupes sécrétant des tests calcaires. Au-delà, on ne connaît pas, parmi les Ordres connus (Foraminifères, Radiolaires, ) de groupe présentant des caractéristiques proches pour des tests calcaires.
4.4. Discussion
Afin de faciliter la reconnaissance des multiples formes observées dans les résidus d'acétolyse, nous avons adopté un "classement" morphologique en cinq groupes distingués à la loupe. Les observations nannostructurales rapportées ici montrent que ces groupements ne tiennent pas pour certaines catégories : des tests à surface lisse (E1) et à nannostructure en réseau (E2) se retrouvent dans le même groupe tandis que des formes probablement affiliées sont dans des groupes différents. Les formes en cône (K, L, M), par exemple, regroupent des tests de signification différente tandis que des formes en cupules (E2, E3, G1 par exemple) sont probablement proches de formes en cône (groupe de formes M par exemple) d'après l'ornementation de surface commune (la "striation" visible à la loupe, la lèvre péri-orale d'où partent des crêtes radiales).
L'un des facteurs de distinction des divers taxons est leur taille. Celle-ci varie peu pour une même forme. C'est sur ce critère de différence de taille que certains tests en disque (A1a par rapport à A1b, par exemple) ou en cône (E3a par rapport à E3b, par exemple) ont été distingués lors de l'étude à la loupe.
L'utilisation du dessin assisté par ordinateur indique qu'il suffit parfois d'une modeste modification d'une matrice, d'un gonflement, de l'ajout d'une expansion ou d'une ride pour passer d'une forme à l'autre. Cet outil a ainsi mis en évidence des parentés qui serviront lors de la classification.
Quelques formes (6 sur 36) posent problème et doivent être disjointes du groupe établi par la désignation de l'espèce type (Gilianella stellata) lors de l'étude précédente (, sous presse). Ces formes sont les suivantes : groupe K (moulages de tests de caractéristiques inconnues) , E1 et F1 (surface des tests sans nannostructure en réseau), L1 (paroi à 2 couches). Les 30 autres formes appartiennent au même ensemble de microproblematica; elles ont un test uniloculaire et petit, une symétrie fondamentalement axiale, un orifice unique, une paroi monocouche de 15 à 20 µm d'épaisseur constituée de cristaux allongés perpendiculairement à la surface, une nannostructure de surface très découpée en réseau.
Ainsi redéfini, les gilianelles rassemblent plus d'une trentaine d'espèces dans le seul gisement étudié en détail sur le flanc nord de l'anticlinal de Tercis-Angoumé. D'autres investigations ont porté sur un affleurement situé sur le flanc sud (, 2001), au lieu-dit Peyrata ; nous avons constaté la présence des mêmes formes se succédant dans le même ordre. C'est par des études de ce type que l'on précisera le potentiel corrélatif et l'intérêt (autre que celui de mettre en évidence une biodiversité non reconnue jusqu'ici) de ces microproblematica pour le stratigraphe.
Nous illustrons ici pour la première fois en détail la morphologie des microproblematica de Tercis. Cette illustration avait d'abord pour but de fixer nos observations par l'individualisation de formes aux caractères constants et distincts de ceux des autres formes ainsi que de leur éventuelle variabilité. Au fur et à mesure de l'accumulation des illustrations, réalisée conjointement avec un retour aux observations complémentaires des résidus, les distinctions entre formes se sont précisées et affinées.
La microscopie électronique a conduit à une caractérisation précise des expansions illustrant une diversité remarquable des formes au sein d'une organisation constante. Elle a aussi permis de déceler d'intéressantes nannostructures illustrées ici pour la première fois. L'ornementation de surface des gilianelles est une caractéristique commune déterminante pour les distinguer d'autres micro-enigmatica voire pour les identifier spécifiquement. L'étude de la nannostructure a mis en évidence que la forme L1 et les groupes D et K se détachaient de l'ensemble des gilianelles.
Ce point acquis, il était intéressant de mettre à la disposition des micropaléontologistes des documents qui témoignent de ce qui est visible lors des tris usuels. Plusieurs dizaines de formes ont ainsi été distinguées ce qui permettra de mieux compléter un bilan dont nous pensons qu'il pourrait porter à une cinquantaine la "microfaune" nouvelle de Tercis.
L'étude de la répartition des formes dans le temps met en évidence l'extraordinaire taux de renouvellement de ces microfossiles qui pourraient ainsi acquérir un rôle utile dans la caractérisation des dépôts du Crétacé Supérieur.
Il paraît prioritaire de nommer ces formes en réalisant une taxonomie aussi rigoureuse que possible. Nous avons avancé leur diagnose. Dans un deuxième temps, la recherche de tests semblables dans des couches de même âge et de même faciès (plate-forme continentale crétacée de la Téthys à l'abri des apports détritiques) devra être envisagée. Nous avons tenté d'avancer dans cette direction. Nos recherches en cours dans le bassin du flysch pyrénéen, en France et en Espagne, ont montré que ces microproblematica n'y étaient pas présents; par contre, les faciès plus carbonatés présents en Navarre sont plus prometteurs : 2 spécimens dans un faciès semblable à celui de Tercis.
Claude a contribué à la réalisation des clichés au microscope optique. Sylvie a bien voulu pallier à notre pénurie de matériel pour les acétolyses à chaud. Omar a su rendre parlantes nos préparations de matériel par sa manipulation experte du microscope électronique. Bruno (Université Brest), David et Franco (Université Dalhousie) ont bien voulu nous faire part de commentaires experts sur une version antérieure de cette présentation. La version finale des textes en anglais doit beaucoup aux efforts de Nestor .
Ces travaux ont été partiellement financés par l'intermédiaire de la fédération CEPAGE du CNRS.
P.-Y., G.S., E. & J.-M. (2001).- Microfaciès des sédiments du Campanien et du Maastrichtien de Tercis les Bains (Landes, France). In : G.S. (réd., 2001a), chap. B1b, p. 113-119.
G. (1989).- Les pseudarcellidés, groupe original de microfossiles incertae sedis du Paléogène. Remarques sur la classification et description de trois espèces nouvelles.- Revue de Micropaléontologie, Paris, vol. 31, N° 4, p. 211-224.
M., G.S. & B. (2001).- Geology and Late Cretaceous palaeogeography of the geological site at Tercis les Bains (Landes, France). In : G.S. (réd., 2001a), chap. A4, p. 47-59.
L. (2006).- Glossaire illustré des termes utilisés dans la recherche sur les Foraminifères.- Carnets de Géologie - Notebooks on Geology, Brest, Memoir 2006/02 (CG2006_M02), 126 p.
J.W.M. (1999).- Late Cretaceous-Early Palaeogene echinoderms and the K/T boundary in the southeast Netherlands and northeast Belgium - Part 2: Crinoids.- Scripta Geologica, Leiden, vol. 116B, p. 59-255, 46 pl.
J.W.M. & G.S. (2001).- Campanian-Maastrichtian crinoids (Echinodermata) from Tercis les Bains (Landes). In : G.S. (réd., 2001a): The Campanian - Maastrichtian stage boundary: characterisation at Tercis les Bains (France): correlation with Europe and other continents.- Developments in Palaeontology and Stratigraphy Series, 19, Elsevier Sciences Publ., Amsterdam, p. 635-644.
H. & J. (1994).- Calcareous phytoplankton from the Barremian/Aptian boundary interval from NW Germany.- Cretaceous Research, London, vol. 15, Issue 6, p. 739-763.
F.S., L., D.B. & B.E.
(2003).- The Thecamoebian Bibliography, 2nd Edition.-
Palaeontologia electronica, London, Vol. 6, N° 1, 107 p., 2.1MB
http://palaeo-electronica.org/paleo/2003_1/biblio/issue1_03.htm
G.S., rédacteur (2001a).- The Campanian - Maastrichtian stage boundary. Characterisation at Tercis les Bains (France) and correlation with Europe and other continents.- IUGS Special Publication (monograph) Series, 36 ; Developments in Palaeontology and Stratigraphy Series, 19, Elsevier Sciences Publ. Amsterdam, xxviii + 881 p.
G.S. (2001b).- Numerical age calibration of the Campanian-Maastrichtian succession at Tercis les Bains (Landes, France) and in the Bottaccione Gorge (Italy). In : G.S. (réd., 2001a), chap. E5c, 775-782.
G.S. (2001c).- Descriptive litholostratigraphy of the Campanian-Maastrichtian succession at Tercis les Bains (SW France). In : G.S. (réd. 2001a), chap. B1a, p. 85-109.
G.S. (2001d).- The Campanian-Maastrichtian boundary: characterization at Tercis (Landes, SW France). In : G.S. (réd., 2001a), chap. F1, p. 785-804.
G.S. (in press).- Sur des microfossiles calcaires problématiques du stratotype de la limite Campanien-Maastrichtien de Tercis (Sud-Ouest, France) : les gilianelles.- Comptes Rendus Palevol, Paris, doi : 10.1016/j.crpv.2006.10.001 (Corrected Proof, Available online 27 November 2006).
G.S. & M.A. (2001a).- The global Campanian-Maastrichtian stage boundary at Tercis les Bains, Landes, SW France.- Episodes, Beijing, Vol. 24, N° 4, p. 229-238.
G.S. & M.A. (2001b).- Outcrops of the geological site at Tercis les Bains (Landes, France). In : G.S. (réd., 2001a), chap. A5, p. 60-67.
G.S. & I. (2003).- Sur les inocérames de Tercis (Landes, France) : le meilleur outil corrélatif entre Europe et Amérique du Nord autour de la limite Campanien-Maastrichtien / Inoceramids of the site of Tercis (Landes, France): the best correlative tool between Europe and North America across the Campanian-Maastrichtian boundary.- Comptes Rendus Géosciences, Paris, Vol. 335, p. 239-246.
A. (1990).- Nouveaux microproblematica : Yvonniella parva n. sp., Voorthuyseniella bearnensis n. sp. et Pseudarcella sp. dans l'Éocène et l'Oligocène d'Aquitaine (Sud-Ouest de la France).- Revue de Micropaléontologie, Paris, Vol. 33, N° 2, p. 115-122.
A. (1991).- Unifissurinella, new genus of microproblematica from the Upper Eocene of southwestern France.- Micropaleontology, New York, Vol. 37, N° 1, p. 95-97.
M. (2001). The site at Tercis les Bains (Landes), stratotype of the Campanian-Maastrichtian limit: insights from tectonic studies. In : G.S. (réd., 2001a), chap. A6, p. 68-81.

Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 1 : Prise de vue à la loupe binoculaire d'un séparat de fossiles non encore cités de Tercis. niveau 50,0, palier II, Grande Carrière de Tercis; résidu d'acétolyse à chaud puis à froid, fraction 100-250 µm. Plusieurs formes de microproblematica sont visibles : A4b (3 grands disques), B2a (multiples disques à 8 bras = Gilianella stellata), C1b (disque épais à 5 bras courts), L1 (4 petits entonnoirs), M2a (1 cône strié) avec des restes de foraminifères et d'échinodermes (2 plaques à symétrie d'ordre 5). (barre = 200 µm, cliché AL avec boîtier numérique posé sur oculaire de loupe binoculaire).
Picked sample viewed under a stereomicroscope. Level 50.0, floor II, Tercis Grande Carrière section, SW France; hot then cold acetolysis; size: 100-250 µm. Several forms of microproblematica are present: A4b (3 large discs), B2a (many 8-armed discs = Gilianella stellata), C1b (thick disc with 5 arms), L1 (4 small funnels), M2a (1 striated cone) together with remains of foraminifers and echinoderms (2 plates with pentagonal symmetry). (bar = 200 µm, picture AL by digital camera on the ocular).
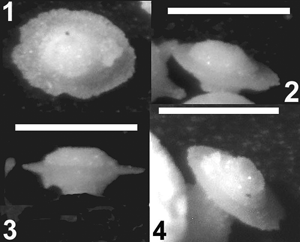
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 2 : Vues à la loupe binoculaire de la gilianelle A2b. Résidu d'acétolyse à froid, niveau 62,5, palier II, Tercis. 1 - vue aborale ; 2 - vue cavalière orale ; 3 - profil face orale en haut ; 4 - vue cavalière aborale. (barres = 200 µm, cliché AL avec boîtier numérique posé sur oculaire de loupe binoculaire).
Stereomicroscope views of gilianelle A2b. Level 62.5, floor II, Tercis; cold acetolysis residue. 1 - aboral view, 2 - oblique view oral surface, 3 - profile oral face up, 4 - oblique view aboral surface. (bars = 200 µm, picture AL with digital camera on the ocular).
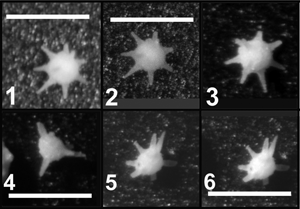
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 3 : Vues à la loupe binoculaire de la gilianelle H2. Résidu d'acétolyse à froid, niveau 79,8, palier II, Tercis. 1 à 3 : vues aborales montrant les 7 bras et 1 ou 2? ou 3? pieds vers l'observateur; 4 : vue de profil face orale vers le NE, 2 pieds superposés vers le SW; 5- 6 : profils légèrement cavalier, face orale vers le NW, 2 pieds sont distincts vers le S et l'E. (barre = 200 µm, cliché AL avec boîtier numérique posé sur oculaire de loupe binoculaire).
Stereomicroscope views of gilianelle H2. Level 79.8, floor II, Tercis; cold acetolysis residue. 1-3: 3 aboral views showing 7 arms and 1? to 3? feet, 4: profile with oral face to the NE, and 2 superposed feet to the SW; 5-6 : slightly oblique profile, oral face to the NW, 2 discrete feet towards S and E. (bar = 200 µm, picture AL with digital camera on the ocular).

Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 4 : Vue au microscope optique d'un tri de microfossiles inédits de Tercis. Grande Carrière, palier II, niveau 54,9, résidu d'acétolyse à froid, fraction 100-250 µm. Noter la présence de foraminifères, d'éléments d'échinodermes, gilianelles diverses dont les formes A1a, A2b, A4b, B2 (dont Gilianella stellata), C1b, F1, G1, M2, spicules. (barre = 200 µm, cliché GSO MIV129 boîtier numérique sur photomicroscope digitalisé).
Picked specimens viewed under an optical microscope. Level 54.9, floor II, Tercis; 100- 250 µm size fraction; cold acetolysis residue. Note the presence of some foraminifers, echinoderm plates, microproblematica including forms A1a, A2b, A4b, B2 (including Gilianella stellata), C1b, F1, G1, M2, spicules. (bar = 200 µm, picture GSO MIV129 digital camera on a photomicroscope).

Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 5 : Vues au microscope optique à partir d'un tri de microfossiles inédits de Tercis. Grande Carrière, palier II, niveau 11,0 ; résidu d'acétolyse à froid, 100-250 µm. Les formes caractéristiques sont identifiées comme décrit dans les planches. (barre = 200 µm, clichés GSO 72 à 82, boîtier numérique sur photomicroscope).
Selected specimens viewed under an optical microscope. Level 11.0, floor II, Tercis; 100-250 µm size fraction; cold acetolysis residue. Forms are identified using the code shown in our three-dimensional plates of drawings. (bar = 200 µm, picture GSO 72 to 82 digital camera on a photomicroscope).
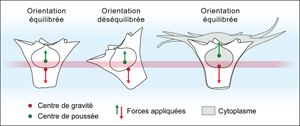
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 6 : Orientation de la forme M3 dans le milieu. La cavité du test est représentée. Dès qu'un déplacement survient, les centre de gravité et de poussée s'éloignent de la verticale et les forces qui s'y appliquent entraînent un rappel vers la position d'équilibre (schéma médian). A gauche test seul; à droite test avec cytoplasme hypothétique. La barre horizontale sert de repére relatif d'un schéma à l'autre.
Orientation of form M3 in sea-water. The drawings show the shell chamber. When movement occurs, the centre of gravity and the centre of the effect deviate from the vertical and these forces lead to a return to the position of equilibrium. Shell without a hypothetical cytoplasm left and center; shell with hypothetical cytoplasm to the right. The horizontal bar serves as a basis for comparing attitudes.
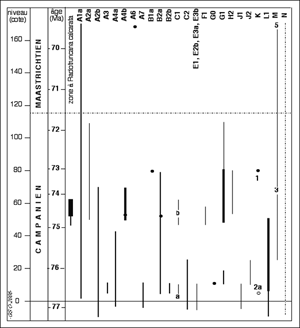
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 7 : Distribution stratigraphique des microproblematica extraits de 32 niveaux carbonatés dissociés par acétolyse dans la section principale de la Grande Carrière de Tercis (Landes, France).
Stratigraphic distribution of microproblematica collected from 32 carbonate levels of the main section of the Tercis Grande Carrière (Landes, France).
Planches/Plates 1-6 : Représentations modélisées des formes de microproblematica extraites de niveaux carbonatés dissociés par acétolyse de la section de Tercis. Vues reconstituées à partir d'observations optiques. On a coloré différemment les 5 catégories morphologiques distinguées : disque (A ; B ; C), sphère (D), cupule (E ; F ; G ; H ; J), cône (K ; L ; M), tonnelet (N). Chaque forme est représentée, de gauche à droite, en profil cavalier oral, vue orale, profil et vue aborale. L'échelle est commune aux 36 formes.
Models of the forms of microproblematica collected from carbonate levels of the Tercis main section (stratotype of the Campanian-Maastrichtian boundary, SW France). The backgrounds of five morphological sets are in different colours: discs (A; B; C), spheres (D), cupules (E; F; G; H; J), cones (K; L; M), kegs (N). Each form is drawn as if viewed from a discrete position, from left to right: oblique view of the oral face from above, oral view, profile and aboral views. The scale is common to all 36 forms.

Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
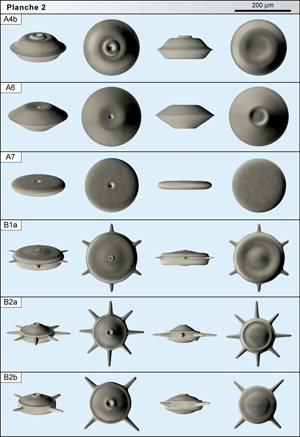
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
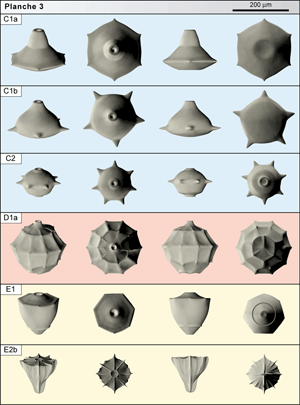
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
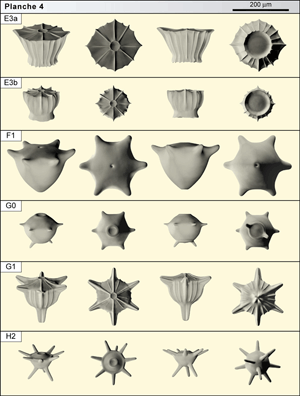
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.

Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.

Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
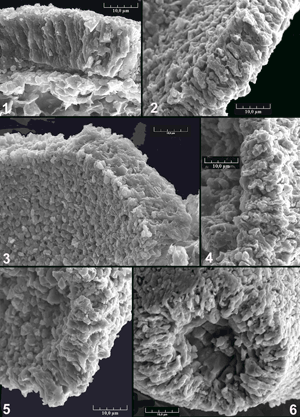
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche/Plate 7 : Nannostructures de la paroi des
microproblematica extraits de niveaux carbonatés dissociés par acétolyse ; Grande Carrière à Tercis ; observations en microscopie électronique à
balayage. (barre d'échelle commune = 10 µm)
1- paroi de la forme A1 b (extérieur en haut) ; niveau 172 ;
2- paroi de la forme A4a
(extérieur en bas à droite) ; niveau 5,0 ; 3- paroi et vue de l'intérieur de la forme A1 b ; niveau 172 ;
4- paroi de la forme B2a
(extérieur à droite) ; niveau 50,25 ; 5- paroi de la forme C1a
(extérieur en bas à droite) ; niveau 5,0 ; 6- section basale d'un bras de la forme F1 ; niveau 47,2.
Nannostructures of microproblematica collected from carbonate levels
in the Tercis section; broken specimens. (scanning electron microscope; scale bars = 10 µm)
1- shell wall of form A1 b (exterior at the top); level 172 ; 2- shell wall of form A4a
(exterior at the bottom right); level 5.0; 3- shell wall viewed from inside, form A1 b; level 172;
4- shell wall of B2a (exterior on the right); level 50.25; 5- shell wall of C1a
(exterior on the bottom right); level 5.0; 6- basal section of an arm of form F1; level 47.2.
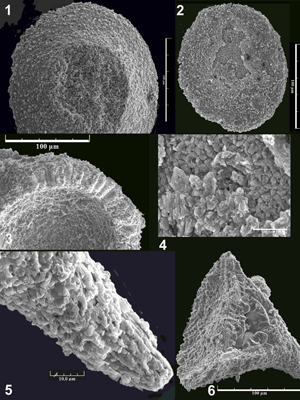
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche/Plate 8 : Nannostructures particulières de formes énigmatiques de Tercis.
(microscopie électronique à balayage ; échelle 10 ou 100 µm selon le cliché)
1- Vue aborale de la forme K1 dont la portion aborale est sectionnée ;:pas de paroi différenciée, pas d'ornementation de surface ; niveau 79,8 ;
2- Vue orale de la forme L1 ; noter l'orifice très discret au centre et la zone desquamée, grossie dans la vue 4 ; niveau 11,0 ;
3- section de la forme D1a
(paroi deux fois plus épaisse que celle des formes rapportées aux gilianelles ; niveau 89,0 ;
4- nannostructure de la forme L1
(grossissement de la vue 2) ; la paroi montre 2 couches ; à la surface de la couche intérieure, les cristallites ont une organisation en croix délimitant des lumières (comparer avec la vue intérieure de la paroi de A1b :
Pl. 7, fig. 3 ![]() ) ;
5- Pied de la forme L1 ; noter les 2 nannostructures différentes : sans organisation en surface et en cristallites très allongés dessous ; niveau 5,0 ;
6- Section de la forme L1 ; la cavité interne du test, emplie de gros
cristaux, s'interrompt au niveau du pied qui est plein ; la paroi de la face orale concave est constituée de deux couches ; à la périphérie de la face
orale, on ne voit que l'amorce de la collerette.
) ;
5- Pied de la forme L1 ; noter les 2 nannostructures différentes : sans organisation en surface et en cristallites très allongés dessous ; niveau 5,0 ;
6- Section de la forme L1 ; la cavité interne du test, emplie de gros
cristaux, s'interrompt au niveau du pied qui est plein ; la paroi de la face orale concave est constituée de deux couches ; à la périphérie de la face
orale, on ne voit que l'amorce de la collerette.
Nannostructures of microproblematica collected from carbonate levels from the Tercis section.
(scanning electron microscope; scale 10 or 100 µm)
1- Aboral view of form K1 with aboral portion of the "shell" removed ; no shell wall, no ornamentation; level 79.8;
2- Oral view of form L1; note the small aperture with a desquamated zone
around it, enlarged 4 times in view 4; level 11.0; 3- broken shell of form D1a (shell wall twice as thick as
in typical gilianelles; level 89.0; 4- nannostructure of form L1 (view 2 enlarged); the shell wall shows 2 layers; the surface of the internal layer shows well arranged crystals (compare to the shell wall of A1b:
Pl. 7, fig. 3 ![]() );
5- Foot of form L1; note the 2 discrete nannostructures: no organisation in the
outer layer, good organisation in the inner layer; level 5.0; 6- Broken form L1; the chamber of the shell is full of large crystals; the shell wall of the oral face
consists of 2 layers; the collarette around the oral face is destroyed.
);
5- Foot of form L1; note the 2 discrete nannostructures: no organisation in the
outer layer, good organisation in the inner layer; level 5.0; 6- Broken form L1; the chamber of the shell is full of large crystals; the shell wall of the oral face
consists of 2 layers; the collarette around the oral face is destroyed.

Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche/Plate 9 : Ornementation de surface et expansions des microproblematica de la Grande Carrière de Tercis.
(microscopie électronique à balayage ; barre d'échelle commune = 100 µm)
1- ornementation de la forme A1a (concentrique et radiale); vue aborale sans d'orifice ; niveau 5,0 ;
2- ornementation de la forme A7
(réseau de mailles en grille); vue orale (orifice présent) ; niveau 5,0 ;
3- ornementation de la forme M3 ; vue aborale (pied en tube armé de 5
ailettes, corps strié en vue optique) ; niveau 69,0 ; 4- ornementation de la forme G1 ; vue aborale (pied à section en
croix) ; niveau 69,0 ; 5- ornementation de la forme G1 ; vue de profil (bras à section en
croix, orifice entouré d'une lèvre d'où partent des crêtes
rayonnantes) ; niveau 11,0 ; 6- ornementation de la forme J1 ; vue cavalière aborale
(collerette aborale d'où part l'amorce d'un pied en palette (en haut à gauche), un autre est détruit (à
droite); collerette orale d'où partent 4 bras perpendiculaires en palettes, seuls les 2 opposés à gauche et à droite sont préservés) ; niveau 5,0 ;
7- ornementation et expansions de la forme A2b ; vue de profil dans l'axe long
(collerette équatoriale horizontale et collerette aborale vers le bas) ; niveau 11, 0.
Surface ornamentation and expansions of microproblematica of the Grande Carrière at Tercis. (scanning electron
microscope; scale bars = 100 µm)
1- ornamentation of form A1a; aboral view (no aperture); level 5.0; 2- ornamentation of form A7; oral view (with aperture); level 5.0;
3- ornamentation of form M3; aboral view (tubular foot with 5 wings, striated body in optical view); level 69.0;
4- ornamentation of form G1; aboral view
(note the foot with a cross-like section); level 69.0; 5- ornamentation of form G1; Profile view (arm with a cross-like section, aperture surrounded by a lip from
which combs radiate); level 11.0; 6- ornamentation of form J1; oblique aboral view
(aboral collarette with blade like foot (top left), another on the right has
been destroyed; oral collarette oral with 4 perpendicular arms; only 2 are preserved; level 5.0;
7- ornamentation and expansions of form A2b; profile view (equatorial collarette and aboral collarette
at the lower end); level 11.0.
Click on thumbnail to play a high-resolution video file (mpg = 3,424 KB)
or click on the icon ![]() to download another high-resolution video file (avi = 3,456 KB)
to download another high-resolution video file (avi = 3,456 KB)
Animation/Video 1 : Manipulation du logiciel Blender. Essai pour la gilianelle N1.
Trial of Blender software on gilianelle N1.
Click on thumbnail to play a high-resolution video file (mpg = 1,248 KB)
or click on the icon ![]() to download another high-resolution video file (avi = 3,947 KB)
to download another high-resolution video file (avi = 3,947 KB)
Animation/Video 2 : Panoramique tridimensionnelle pour la gilianelle Gilianella stellata.
Three dimensional panoramic views of the gilianelle Gilianella stellata.
Click on thumbnail to play a high-resolution video file (mpg = 3,218 KB)
or click on the icon ![]() to download another high-resolution video file (avi = 5,256 KB)
to download another high-resolution video file (avi = 5,256 KB)
Animation/Video 3 : Reconstitution de déplacement passif pour la gilianelle M3 plongée dans le milieu marin.
Reconstruction of the passive movement of gilianelle M3 in sea water.