![]()
![]()
![]() Limestone-marl alternations are usually interpreted to reflect cyclic paleoenvironmental fluctuations linked to -scale climate variations. However, the impact of diagenesis on lithological differentiation can be overprinted on the primary signal.
Limestone-marl alternations are usually interpreted to reflect cyclic paleoenvironmental fluctuations linked to -scale climate variations. However, the impact of diagenesis on lithological differentiation can be overprinted on the primary signal.
![]() In order to evaluate environmental variations during the deposition of the
Gargasian hemipelagic limestone-marl alternations from the "La
Marcouline" quarry (SE France), we have applied a multi-discipline approach (carbonate geochemistry, micropaleontology and mineralogy).
The work uses of a method of granulometric separation of the main carbonate particles which (1) allows the characterization and quantification of the compositions of the carbonate phases of both lithologies and (2) gives access to the geochemical signatures of similar carbonate (bio-) particles in each
lithology.
In order to evaluate environmental variations during the deposition of the
Gargasian hemipelagic limestone-marl alternations from the "La
Marcouline" quarry (SE France), we have applied a multi-discipline approach (carbonate geochemistry, micropaleontology and mineralogy).
The work uses of a method of granulometric separation of the main carbonate particles which (1) allows the characterization and quantification of the compositions of the carbonate phases of both lithologies and (2) gives access to the geochemical signatures of similar carbonate (bio-) particles in each
lithology.
![]() The quantifications show that micarb constitutes a significant portion (around 40 per cent) of the sediments. Their geochemical signatures indicate that most of them were formed by fine fragmentation of nannoconid tests. Based on these interpretations, our results indicate that nannoconids are the major constituents of both the marly limestone and the marl layers and that both lithologies are similar in composition.
The quantifications show that micarb constitutes a significant portion (around 40 per cent) of the sediments. Their geochemical signatures indicate that most of them were formed by fine fragmentation of nannoconid tests. Based on these interpretations, our results indicate that nannoconids are the major constituents of both the marly limestone and the marl layers and that both lithologies are similar in composition.
![]() The coccolith assemblages (mainly Watznaueria spp.) of the limestone and marl layers are similar and equally well-preserved. Changes in oxygen and carbon isotopic ratios express cyclic fertility and salinity/temperature variations in surface waters. The marls indicate higher fertility and lower salinity (or higher temperature) conditions than those denoted by the marly
limestones.
The coccolith assemblages (mainly Watznaueria spp.) of the limestone and marl layers are similar and equally well-preserved. Changes in oxygen and carbon isotopic ratios express cyclic fertility and salinity/temperature variations in surface waters. The marls indicate higher fertility and lower salinity (or higher temperature) conditions than those denoted by the marly
limestones.
![]() Although diagenesis has not altered these rhythmites no strong evidence has been found for linking classical processes such as dilution or productivity cycles to the composition of the sediments (carbonate constituents and clay mineralogy).
Although diagenesis has not altered these rhythmites no strong evidence has been found for linking classical processes such as dilution or productivity cycles to the composition of the sediments (carbonate constituents and clay mineralogy).
![]() This study demonstrates that La Marcouline succession has retained, as recorded by geochemical proxy, its primary environmental variations. Nevertheless, the homogeneity in both lithotypes of sedimentological parameters (clay mineralogy, sediment composition,
...) seems to indicate that these fluctuations did not affect the ecosystem strongly enough to cause marked differences in the composition of marls and
marly-limestones.
This study demonstrates that La Marcouline succession has retained, as recorded by geochemical proxy, its primary environmental variations. Nevertheless, the homogeneity in both lithotypes of sedimentological parameters (clay mineralogy, sediment composition,
...) seems to indicate that these fluctuations did not affect the ecosystem strongly enough to cause marked differences in the composition of marls and
marly-limestones.
![]() Limestone-marl
alternation, stable isotopes, Aptian, Gargasian, calcareous nannofossils, granulometric
separation.
Limestone-marl
alternation, stable isotopes, Aptian, Gargasian, calcareous nannofossils, granulometric
separation.
C., M. de, M., M. & G. (2007).- Environmental changes during marl-limestone formation: evidence from the Gargasian (Middle Aptian) of La Marcouline Quarry (Cassis, SE France).- Carnets de Géologie / Notebooks on Geology, Brest, Article 2007/01 (CG2007_A01)
![]() Variations des paramètres de l'environnement au cours de la sédimentation d'alternances marne-calcaire : Exemple de la série gargasienne
(Aptien moyen) de la carrière de La Marcouline (Cassis, SE France).- Il est classiquement admis que les systèmes alternants marne-calcaire reflètent des variations climatiques orbito-dépendantes.
Toutefois, des processus diagénétiques conduisant à la différentiation lithologique peuvent avoir masqué ce signal
primaire.
Variations des paramètres de l'environnement au cours de la sédimentation d'alternances marne-calcaire : Exemple de la série gargasienne
(Aptien moyen) de la carrière de La Marcouline (Cassis, SE France).- Il est classiquement admis que les systèmes alternants marne-calcaire reflètent des variations climatiques orbito-dépendantes.
Toutefois, des processus diagénétiques conduisant à la différentiation lithologique peuvent avoir masqué ce signal
primaire.
![]() Afin de caractériser les variations des paramètres environnementaux associés à la formation des alternances marno-calcaires gargasiennes de La Marcouline (SE France), nous avons mené une étude pluridisciplinaire
(géochimique, micropaléontologique et minéralogique) basée sur l'utilisation d'une méthode de séparation granulométrique des particules sédimentaires. Cette approche nous a
permis, d'une part de caractériser et quantifier les compositions des phases carbonatées des deux types de lithologies et d'autre part de disposer des signatures géochimiques de (bio-) particules carbonatées identiques dans les marnes et les calcaires
marneux.
Afin de caractériser les variations des paramètres environnementaux associés à la formation des alternances marno-calcaires gargasiennes de La Marcouline (SE France), nous avons mené une étude pluridisciplinaire
(géochimique, micropaléontologique et minéralogique) basée sur l'utilisation d'une méthode de séparation granulométrique des particules sédimentaires. Cette approche nous a
permis, d'une part de caractériser et quantifier les compositions des phases carbonatées des deux types de lithologies et d'autre part de disposer des signatures géochimiques de (bio-) particules carbonatées identiques dans les marnes et les calcaires
marneux.
![]() Les quantifications ont mis en évidence la présence, en proportions importantes (environ 40% des échantillons) de
micarb. À partir des données géochimiques obtenues sur fractions séparées, nous avons proposé pour ces particules une origine liée à la fragmentation fine de tests
de nannoconidés. Il en résulte donc que les nannoconidés sont les constituants majeurs aussi bien des marnes que des calcaires marneux et qu'ils sont présents en proportions équivalentes dans les deux
lithologies.
Les quantifications ont mis en évidence la présence, en proportions importantes (environ 40% des échantillons) de
micarb. À partir des données géochimiques obtenues sur fractions séparées, nous avons proposé pour ces particules une origine liée à la fragmentation fine de tests
de nannoconidés. Il en résulte donc que les nannoconidés sont les constituants majeurs aussi bien des marnes que des calcaires marneux et qu'ils sont présents en proportions équivalentes dans les deux
lithologies.
![]() Les assemblages de coccolithes
(principalement Watznaueria spp.) ne présentent pas de variations de composition, ni de préservation d'une lithologie à
l'autre. L'évolution de leurs signatures isotopiques signe des fluctuations marquées des conditions de fertilité et de température
et/ou salinité dans la zone photique au cours de la sédimentation de ces
alternances. Les épisodes marneux correspondent à des périodes plus chaudes
et/ou moins salées au cours desquelles les apports de matériel terrigène sont importants induisant une augmentation de la fertilité des eaux de surface par rapport aux périodes de formation des niveaux plus carbonatés.
Les assemblages de coccolithes
(principalement Watznaueria spp.) ne présentent pas de variations de composition, ni de préservation d'une lithologie à
l'autre. L'évolution de leurs signatures isotopiques signe des fluctuations marquées des conditions de fertilité et de température
et/ou salinité dans la zone photique au cours de la sédimentation de ces
alternances. Les épisodes marneux correspondent à des périodes plus chaudes
et/ou moins salées au cours desquelles les apports de matériel terrigène sont importants induisant une augmentation de la fertilité des eaux de surface par rapport aux périodes de formation des niveaux plus carbonatés.
![]() Malgré le fait que la diagenèse ne semble pas avoir joué un rôle significatif dans la mise en place de ces alternances
marno-calcaires, nous n'avons pas pu mettre en évidence de relation nette entre des processus classiques du type cycles de dilution ou de production et les compositions des sédiments
(particules carbonatées et cortèges argileux).
Malgré le fait que la diagenèse ne semble pas avoir joué un rôle significatif dans la mise en place de ces alternances
marno-calcaires, nous n'avons pas pu mettre en évidence de relation nette entre des processus classiques du type cycles de dilution ou de production et les compositions des sédiments
(particules carbonatées et cortèges argileux).
![]() Cette étude montre donc que la série de La Marcouline a conservé un signal primaire lié à des variations environnementales clairement enregistrées par les marqueurs géochimiques
(salinité, température et fertilité). Néanmoins, il semble que ces variations n'aient pas suffisamment perturbé l'environnement pour que cela se traduise dans les paramètres sédimentologiques
(matériel terrigène, assemblages biologiques, etc.).
Cette étude montre donc que la série de La Marcouline a conservé un signal primaire lié à des variations environnementales clairement enregistrées par les marqueurs géochimiques
(salinité, température et fertilité). Néanmoins, il semble que ces variations n'aient pas suffisamment perturbé l'environnement pour que cela se traduise dans les paramètres sédimentologiques
(matériel terrigène, assemblages biologiques, etc.).
![]() Alternances marno-calcaires, isotopes stables,
Aptien, Gargasien, nannofossiles calcaires, séparation granulométrique.
Alternances marno-calcaires, isotopes stables,
Aptien, Gargasien, nannofossiles calcaires, séparation granulométrique.
![]() Marl-limestone alternations from the Gargasian (Middle
Aptian) of southeastern France are characterised by a gradual transition from one lithotype to the other. The area around Cassis provides well-exposed and laterally continuous outcrops to study marl-limestone alternations. General geological information regarding the litho- and biostratigraphy of the Gargasian of La Marcouline Quarry are presented in et alii
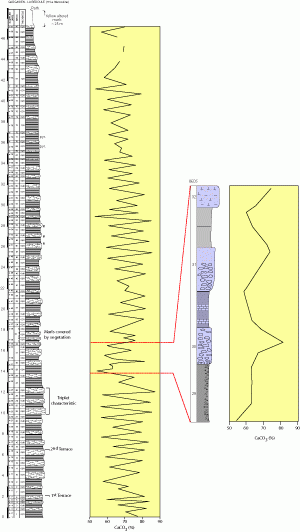
(2004). The section is 50 meters thick (Fig. 1
Marl-limestone alternations from the Gargasian (Middle
Aptian) of southeastern France are characterised by a gradual transition from one lithotype to the other. The area around Cassis provides well-exposed and laterally continuous outcrops to study marl-limestone alternations. General geological information regarding the litho- and biostratigraphy of the Gargasian of La Marcouline Quarry are presented in et alii
(2004). The section is 50 meters thick (Fig. 1 ![]() ) and consists of regular alternation of light-colored marly limestones and grey marls spanning the early Gargasian (,
1966; , 1987; ,
1997; et alii,
2004). The entire section has been investigated in terms of chemostratigraphy (
et alii, in press) and cyclostratigraphy (,
2006). Frequencies in tune with
cycles, deduced from spectral analysis, suggest an orbitally-forced nature for these marl-limestone alternations (,
2006). The aim of this work is to understand the changes occurring in the marine environment during the sedimentation of a marl-limestone couplet by means of geochemical analysis (δ18O and δ13C) on fine fractions isolated from both lithotypes. Differential diagenetic processes, and the possible variation of carbonate producers in limestones and marls may makes it difficult to obtain an unambiguous geochemical signature for both lithologies. In order to prevent possible artefacts related to these differences (,
2006), we have tried to analyse the same constituents isolated from both lithologies by using the technique of granulometric separation developed by et alii
(2001). This method allowed us to obtain homogeneous fractions of the same type of constituent and then to study the isotopic signature of all the carbonate particles of the sediments.
) and consists of regular alternation of light-colored marly limestones and grey marls spanning the early Gargasian (,
1966; , 1987; ,
1997; et alii,
2004). The entire section has been investigated in terms of chemostratigraphy (
et alii, in press) and cyclostratigraphy (,
2006). Frequencies in tune with
cycles, deduced from spectral analysis, suggest an orbitally-forced nature for these marl-limestone alternations (,
2006). The aim of this work is to understand the changes occurring in the marine environment during the sedimentation of a marl-limestone couplet by means of geochemical analysis (δ18O and δ13C) on fine fractions isolated from both lithotypes. Differential diagenetic processes, and the possible variation of carbonate producers in limestones and marls may makes it difficult to obtain an unambiguous geochemical signature for both lithologies. In order to prevent possible artefacts related to these differences (,
2006), we have tried to analyse the same constituents isolated from both lithologies by using the technique of granulometric separation developed by et alii
(2001). This method allowed us to obtain homogeneous fractions of the same type of constituent and then to study the isotopic signature of all the carbonate particles of the sediments.
![]() The present study focuses on the couplets 29 to 32 of et alii
(2004), corresponding to the parasequence 4 defined in et alii
(in press). The sampled section
(Fig. 1
The present study focuses on the couplets 29 to 32 of et alii
(2004), corresponding to the parasequence 4 defined in et alii
(in press). The sampled section
(Fig. 1 ![]() ) consists of 0.3 to 0.6 m thick light-colored marly limestones (60% < CaCO3 < 82%), regularly alternating with 0.7 m thick grey marls (54% < CaCO3 < 72%).
) consists of 0.3 to 0.6 m thick light-colored marly limestones (60% < CaCO3 < 82%), regularly alternating with 0.7 m thick grey marls (54% < CaCO3 < 72%).
![]() The protocol of separation is described in et alii
(2001) and (2002). This method, based on granulometric separations of the various carbonate particles, was applied to samples from both lithologies. The constituents of a sample must first be disaggregated into a suspension. Consequently, the protocol can be split into two processes. A preliminary phase is a physical disaggregation of the rock and a wet sieving to remove larger microfossils. In a second phase, the resulting suspension is size-separated in a sequence of filtration steps. In view of the composition of the samples, we chose to apply four filtration steps in an ultrasonic bath through etched-polycarbonate membrane filters, successively 12, 8, 5 and 3 µm micropores in diameter. Finally, the smallest particles (< 3 µm) were concentrated via centrifugation. Smear slides were then prepared from each separated fraction. In each fraction, the surface area of 200 randomly selected carbonate particles was measured in LM using a cross-grid graticule to quantify the composition of each fraction. Combining the surface area of particles and their mean thickness, measured using SEM, the volume of particles was evaluated. Assuming that the density of all carbonate minerals is almost homogeneous, this method led to estimates of the relative weight of the different types of carbonate particle in each fraction. Based on replicate counts, the cumulative error for a given component is less than 10% ( et alii,
2004). Fractions are regarded as pure when one type of particles represents at least 80% of the volume of the carbonates, while they are considered mixed when there are two or three major components.
The protocol of separation is described in et alii
(2001) and (2002). This method, based on granulometric separations of the various carbonate particles, was applied to samples from both lithologies. The constituents of a sample must first be disaggregated into a suspension. Consequently, the protocol can be split into two processes. A preliminary phase is a physical disaggregation of the rock and a wet sieving to remove larger microfossils. In a second phase, the resulting suspension is size-separated in a sequence of filtration steps. In view of the composition of the samples, we chose to apply four filtration steps in an ultrasonic bath through etched-polycarbonate membrane filters, successively 12, 8, 5 and 3 µm micropores in diameter. Finally, the smallest particles (< 3 µm) were concentrated via centrifugation. Smear slides were then prepared from each separated fraction. In each fraction, the surface area of 200 randomly selected carbonate particles was measured in LM using a cross-grid graticule to quantify the composition of each fraction. Combining the surface area of particles and their mean thickness, measured using SEM, the volume of particles was evaluated. Assuming that the density of all carbonate minerals is almost homogeneous, this method led to estimates of the relative weight of the different types of carbonate particle in each fraction. Based on replicate counts, the cumulative error for a given component is less than 10% ( et alii,
2004). Fractions are regarded as pure when one type of particles represents at least 80% of the volume of the carbonates, while they are considered mixed when there are two or three major components.
![]() The oxygen and carbon isotope composition of bulk carbonate and of the various separated fractions were measured for all samples. The extraction of CO2 was done by reaction with anhydrous orthophosphoric acid at 50°C. Isotopic analyses were performed in a mass spectrometer Finnigan Delta E. The oxygen and carbon isotope values are expressed in per mil relative to the V-PDB standard reference. The analytical precision is estimated to 0.1‰ for oxygen and 0.05‰ for carbon.
The oxygen and carbon isotope composition of bulk carbonate and of the various separated fractions were measured for all samples. The extraction of CO2 was done by reaction with anhydrous orthophosphoric acid at 50°C. Isotopic analyses were performed in a mass spectrometer Finnigan Delta E. The oxygen and carbon isotope values are expressed in per mil relative to the V-PDB standard reference. The analytical precision is estimated to 0.1‰ for oxygen and 0.05‰ for carbon.
![]() The non-carbonate fraction of the samples is mainly made of clays and quartz. In both marls and limestones, the argillaceous fraction is dominated by illite and kaolinite
(respectively they average 44% and 42% of the clay fraction; ,
2006).
The non-carbonate fraction of the samples is mainly made of clays and quartz. In both marls and limestones, the argillaceous fraction is dominated by illite and kaolinite
(respectively they average 44% and 42% of the clay fraction; ,
2006).
![]() Four types of carbonate components are found in the samples: (1) foraminifera debris, (2) calcareous nannofossils, (3) calcite microparticles (smaller than 5 µm), and (4) carbonate macrocrystals (larger than 5 µm).
Four types of carbonate components are found in the samples: (1) foraminifera debris, (2) calcareous nannofossils, (3) calcite microparticles (smaller than 5 µm), and (4) carbonate macrocrystals (larger than 5 µm).
Low magnesian-calcite foraminifera are the largest carbonate particles. They represent a minor constituent of the sediments. Only few tests and fragments included in aggregates were seen in the largest fractions (> 20µm).
Calcareous nannofossils are a heterogeneous assemblage made up of coccoliths, coccospheres and nannoconids. Coccoliths are present in several of the fractions (12-8 µm, 8-5 µm, 5-3 µm and < 3 µm). The well-preserved nannofossil assemblages from both lithologies are dominated by Watznaueria barnesae but include a few specimens of Rhagodiscus spp., Biscutum spp. and Zeughrabdotus spp.. Because of the size range of these species, none dominate in any separated fraction. Nannoconus spp. dominate the 8-5 µm size spectrum.
Calcite microparticles (smaller than 5 µm) are considered of uncertain origin because they do not exhibit any obvious biogenic microstructure in optical and electronical microscopy observations. From a morphologic point of view these particles are similar to the micarb described by and (1983), (1992), et alii (1994) and and (2002).
Carbonate macrocrystals are calcitic and ankeritic particles of uncertain origin. They range in size between 12 and 5 µm.
![]() In order to isolate these components, five filtration steps were used (Table 1
In order to isolate these components, five filtration steps were used (Table 1 ![]() ). The >20 µm fraction consists of
aggregates (fragments of bulk sediments) containing foraminifera. The 12, 8 and 3 µm fractions are made up of a mixture of the several kinds of carbonate particles of the sediments. The 8-5 µm fraction can be considered as pure in Nannoconus spp. (87% of the volume of the fraction) and the (< 3 µm) fraction is a concentration of micarb (90% of the volume of the fraction).
). The >20 µm fraction consists of
aggregates (fragments of bulk sediments) containing foraminifera. The 12, 8 and 3 µm fractions are made up of a mixture of the several kinds of carbonate particles of the sediments. The 8-5 µm fraction can be considered as pure in Nannoconus spp. (87% of the volume of the fraction) and the (< 3 µm) fraction is a concentration of micarb (90% of the volume of the fraction).
![]() The composition of the carbonate fraction of both marly limestones and marls appear to be quite similar (Table 2A
The composition of the carbonate fraction of both marly limestones and marls appear to be quite similar (Table 2A ![]() ). Micarb are the main carbonate particles (around 58% in marls and 63% in limestones). The biogenic part of the carbonate fraction is dominated by nannoconids (respectively 25% and 20% of the volume of the carbonate fractions of marls and limestones), while coccoliths are systematically less abundant (they average 8%).
). Micarb are the main carbonate particles (around 58% in marls and 63% in limestones). The biogenic part of the carbonate fraction is dominated by nannoconids (respectively 25% and 20% of the volume of the carbonate fractions of marls and limestones), while coccoliths are systematically less abundant (they average 8%).
![]() As almost 40% of the volume of marls and marly limestones is micarb (They average 36.5% in marls and 42% in limestone, Table 2B
As almost 40% of the volume of marls and marly limestones is micarb (They average 36.5% in marls and 42% in limestone, Table 2B ![]() ), it appears that in order to understand the changes occurring in marine environments during limestone-marl formation, it is crucial to determine the origin and genesis of these microparticles.
), it appears that in order to understand the changes occurring in marine environments during limestone-marl formation, it is crucial to determine the origin and genesis of these microparticles.
![]() The biogenic composition of marls and marly limestones is nearly identical: around 15% Nannoconus spp. and 5% coccoliths (Table 2B
The biogenic composition of marls and marly limestones is nearly identical: around 15% Nannoconus spp. and 5% coccoliths (Table 2B ![]() ). This result does not agree with the classical opinion (see for example & ,
1991) that during the Early Cretaceous, carbonates in limestones were derived mainly from nannoconids and those in marls from coccoliths.
). This result does not agree with the classical opinion (see for example & ,
1991) that during the Early Cretaceous, carbonates in limestones were derived mainly from nannoconids and those in marls from coccoliths.
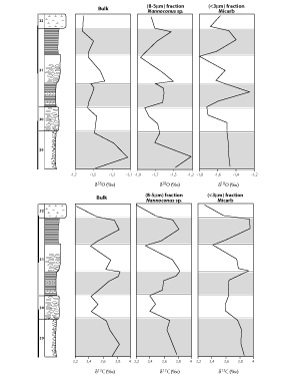
![]() The bulk carbonate of both marly limestones and marls have mean δ18O ratios of about -1.5‰. The values range from -1.6‰ to -1.5‰ in limestones and from -1.6‰ to -1.13‰ in marls (Fig. 2
The bulk carbonate of both marly limestones and marls have mean δ18O ratios of about -1.5‰. The values range from -1.6‰ to -1.5‰ in limestones and from -1.6‰ to -1.13‰ in marls (Fig. 2 ![]() ). Carbon isotopes of bulk carbonate indicate a more significant correlation with lithology than oxygen (Fig. 2
). Carbon isotopes of bulk carbonate indicate a more significant correlation with lithology than oxygen (Fig. 2 ![]() ). Limestone δ13C values (from 3.2‰ to 3.7‰, average 3.5‰) are slightly lower than those in marls (from 3.5‰ and 3.8‰, average 3.7‰).
). Limestone δ13C values (from 3.2‰ to 3.7‰, average 3.5‰) are slightly lower than those in marls (from 3.5‰ and 3.8‰, average 3.7‰).
![]() As the 8-5 µm fraction is dominated up to 87% by Nannoconus spp. and the fine fraction by micarb (90% of the fraction), direct isotope analysis of these two separated fractions give access to the signatures of these two kinds of carbonate particles (Fig. 2
As the 8-5 µm fraction is dominated up to 87% by Nannoconus spp. and the fine fraction by micarb (90% of the fraction), direct isotope analysis of these two separated fractions give access to the signatures of these two kinds of carbonate particles (Fig. 2 ![]() ). The nannoconid-dominated pure fractions exhibit δ18O and δ13C trends relatively similar to those of bulk carbonates
(Fig. 2
). The nannoconid-dominated pure fractions exhibit δ18O and δ13C trends relatively similar to those of bulk carbonates
(Fig. 2 ![]() ). The mean δ18O values are respectively around -1.7‰ (from -1.9‰ to -1.5‰) in marly limestones and -1.5‰ (-1.7‰ to -1.3‰) in marls. Mean δ13C ratios of the nannoconid-dominated fraction are similar in both lithologies, around 3.7‰ (from 3.5‰ to 3.9‰).
). The mean δ18O values are respectively around -1.7‰ (from -1.9‰ to -1.5‰) in marly limestones and -1.5‰ (-1.7‰ to -1.3‰) in marls. Mean δ13C ratios of the nannoconid-dominated fraction are similar in both lithologies, around 3.7‰ (from 3.5‰ to 3.9‰).
![]() The micarb-dominated fine fraction records an isotopic trend generally similar to that of the nannoconids with slight differences noticeable in those derived from marls. Mean oxygen isotope ratios in both lithologies are around -1.5‰ (Fig. 2
The micarb-dominated fine fraction records an isotopic trend generally similar to that of the nannoconids with slight differences noticeable in those derived from marls. Mean oxygen isotope ratios in both lithologies are around -1.5‰ (Fig. 2 ![]() ) (varying between -1.8‰ and -1.3‰). Micarb recorded δ13C ratios varying between 3.3‰ and 3.95‰. The mean values are about 3.6‰ in the marly limestones and 3.8‰ in the marls.
) (varying between -1.8‰ and -1.3‰). Micarb recorded δ13C ratios varying between 3.3‰ and 3.95‰. The mean values are about 3.6‰ in the marly limestones and 3.8‰ in the marls.
![]() In addition to micarb and Nannoconus spp., the samples contain coccoliths. Due to their size ranges, we were not able to isolate them in pure fractions. Coccoliths were concentrated in mixed fractions that also contain Nannoconus spp. and micarb. As the isotopic compositions of nannoconids and micarb have been determined from analysis of pure fractions, we were able to calculate the isotopic signature of coccoliths using the following formula:
In addition to micarb and Nannoconus spp., the samples contain coccoliths. Due to their size ranges, we were not able to isolate them in pure fractions. Coccoliths were concentrated in mixed fractions that also contain Nannoconus spp. and micarb. As the isotopic compositions of nannoconids and micarb have been determined from analysis of pure fractions, we were able to calculate the isotopic signature of coccoliths using the following formula:
![]() δi = [ (NCi) * δ8-5µm fraction ] + [ (MICi) * δ< 3µm fraction ] + [ (CCi) * δCC ]
δi = [ (NCi) * δ8-5µm fraction ] + [ (MICi) * δ< 3µm fraction ] + [ (CCi) * δCC ]
![]() in a given sample:
in a given sample:
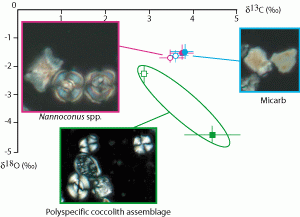
![]() The results of the calculations show that values range from +2.8‰ to +5.6‰ for carbon isotopes and from -4.6‰ to -1.9‰ for oxygen (Fig. 3
The results of the calculations show that values range from +2.8‰ to +5.6‰ for carbon isotopes and from -4.6‰ to -1.9‰ for oxygen (Fig. 3 ![]() ). Coccolith assemblages from marls yield significantly higher carbon isotope values (4.4‰ average) than the ones from marly limestones (mean value 2.9‰). On the contrary, coccolith assemblages from marls yield significantly lower oxygen isotope values (-4.2‰ average) than the ones from marly limestones (-2.3‰ average).
). Coccolith assemblages from marls yield significantly higher carbon isotope values (4.4‰ average) than the ones from marly limestones (mean value 2.9‰). On the contrary, coccolith assemblages from marls yield significantly lower oxygen isotope values (-4.2‰ average) than the ones from marly limestones (-2.3‰ average).
![]() The isotopic signatures of bulk carbonate and biogenic particles (Nannoconus spp. and heterogeneous coccolith assemblages) display similar trends throughout the section (Figs. 2
The isotopic signatures of bulk carbonate and biogenic particles (Nannoconus spp. and heterogeneous coccolith assemblages) display similar trends throughout the section (Figs. 2 ![]() - 3
- 3 ![]() ). Because no evidence of overgrowth or dissolution was observed in LM, we can consider that carbonates are well-preserved and therefore that differences in the isotope trends of Nannoconus spp. and coccoliths are mainly linked to changes in environment. The parallel evolution of the isotope signatures of bulk carbonate and biogenic particles indicates that bulk carbonate records a primary signature reflecting environmental changes in the upper part of the water column. Burial diagenesis seems not to have modified the isotope record of bulk carbonate.
). Because no evidence of overgrowth or dissolution was observed in LM, we can consider that carbonates are well-preserved and therefore that differences in the isotope trends of Nannoconus spp. and coccoliths are mainly linked to changes in environment. The parallel evolution of the isotope signatures of bulk carbonate and biogenic particles indicates that bulk carbonate records a primary signature reflecting environmental changes in the upper part of the water column. Burial diagenesis seems not to have modified the isotope record of bulk carbonate.
![]() Coccolithophores dwell and secrete coccolith plates only in the photic zone ( & ,
1973), thus they record an isotopic signal characteristic of the shallow mixed-layer of oceans ( et alii,
1980; et alii,
1986). Paleotemperature equations (,
1973) show that calcite δ18O is controlled both by temperature and by sea-water δ18O. Due to the absence of icecaps during the Early Cretaceous (,
1983; et alii, 1985; ,
1990; , 1982), variations of sea-water δ18O are probably linked only to fluctuations in salinity. Thus, the lower δ18O values, recorded mainly by coccolithophores and to a minor extent by nannoconids, in the marly sediments, may reflect both surface water freshening caused by enhanced river runoff and increases in temperature. Higher δ13C values recorded in marls may be related to an increase in light carbon uptake linked to an augmentation of the fertility of surface water because of the input of nutrients into the photic zone.
Coccolithophores dwell and secrete coccolith plates only in the photic zone ( & ,
1973), thus they record an isotopic signal characteristic of the shallow mixed-layer of oceans ( et alii,
1980; et alii,
1986). Paleotemperature equations (,
1973) show that calcite δ18O is controlled both by temperature and by sea-water δ18O. Due to the absence of icecaps during the Early Cretaceous (,
1983; et alii, 1985; ,
1990; , 1982), variations of sea-water δ18O are probably linked only to fluctuations in salinity. Thus, the lower δ18O values, recorded mainly by coccolithophores and to a minor extent by nannoconids, in the marly sediments, may reflect both surface water freshening caused by enhanced river runoff and increases in temperature. Higher δ13C values recorded in marls may be related to an increase in light carbon uptake linked to an augmentation of the fertility of surface water because of the input of nutrients into the photic zone.
![]() In both lithologies
(Fig. 3
In both lithologies
(Fig. 3 ![]() ), mean δ18O values of the heterogeneous coccolith fraction are significantly lower than the δ18O values measured on nannoconids. Two explanations may account for the isotopic differences recorded for these two biological groups:
), mean δ18O values of the heterogeneous coccolith fraction are significantly lower than the δ18O values measured on nannoconids. Two explanations may account for the isotopic differences recorded for these two biological groups:
![]() - A non-equilibrium isotopic fractionation induced by physiologic factors. A species-specific isotope disequilibrium has been recognized for several species of modern coccolithophorids (see & ,
2004, et alii, 1980,
and et alii,
1986, for a synthesis). A similar oxygen isotopic disequilibrium might have existed between coccolithophorids and nannoconids in geological times, but this possible factor is difficult to demonstrate for extinct groups.
- A non-equilibrium isotopic fractionation induced by physiologic factors. A species-specific isotope disequilibrium has been recognized for several species of modern coccolithophorids (see & ,
2004, et alii, 1980,
and et alii,
1986, for a synthesis). A similar oxygen isotopic disequilibrium might have existed between coccolithophorids and nannoconids in geological times, but this possible factor is difficult to demonstrate for extinct groups.
![]() - The properties of the waters in which skeletal carbonate was precipitated by coccolithophorids and nannoconids might have been different. This implies a possible difference in bathymetric habitat and/or season of growth.
- The properties of the waters in which skeletal carbonate was precipitated by coccolithophorids and nannoconids might have been different. This implies a possible difference in bathymetric habitat and/or season of growth.
![]() Assuming that the oxygen isotopic signatures of coccoliths and nannoconids reflect the depth at which calcification occurs, Nannoconus spp. seem to have calcified in a cooler and deeper water mass than coccolithophorids. Moreover, the difference in the amplitude of the δ18O and δ13C shifts (Δδ18Om-l, Δδ13Cm-l) of marls (m) and marly limestones (l) recorded by coccoliths are significantly higher (on average Δδ18Om-l = -1.9‰, Δδ13Cm-l = 1.5‰) than those of Nannoconus spp. (Δδ18Om-l and Δδ13Cm-l average 0.2‰). This observation suggests that the oxygen isotopic signatures of coccoliths and nannoconids were controlled mainly by environmental conditions with respect to differences in vital effect. The watermasses in which nannoconids thrived seem to have been more stable than those in which coccoliths calcified. Thus it may be reasonably stated that Nannoconus spp. calcified in deeper water masses than
coccolithophorids. This geochemical result is in good agreement with paleontological studies of
(1994) and
and (1997) that interpret nannoconids as deep-dwellers, inhabiting the lower photic zone like the extant coccolithophore Florisphaera profunda ( & ,
1990).
Assuming that the oxygen isotopic signatures of coccoliths and nannoconids reflect the depth at which calcification occurs, Nannoconus spp. seem to have calcified in a cooler and deeper water mass than coccolithophorids. Moreover, the difference in the amplitude of the δ18O and δ13C shifts (Δδ18Om-l, Δδ13Cm-l) of marls (m) and marly limestones (l) recorded by coccoliths are significantly higher (on average Δδ18Om-l = -1.9‰, Δδ13Cm-l = 1.5‰) than those of Nannoconus spp. (Δδ18Om-l and Δδ13Cm-l average 0.2‰). This observation suggests that the oxygen isotopic signatures of coccoliths and nannoconids were controlled mainly by environmental conditions with respect to differences in vital effect. The watermasses in which nannoconids thrived seem to have been more stable than those in which coccoliths calcified. Thus it may be reasonably stated that Nannoconus spp. calcified in deeper water masses than
coccolithophorids. This geochemical result is in good agreement with paleontological studies of
(1994) and
and (1997) that interpret nannoconids as deep-dwellers, inhabiting the lower photic zone like the extant coccolithophore Florisphaera profunda ( & ,
1990).
![]() The fact that the amplitude of oxygen and carbon isotope fluctuations recorded by coccoliths is higher than that recorded by nannoconids during the deposition of marl and limestone supports the existence in Early Cretaceous oceans of an upper photic zone, inhabited by coccolithophorids, which was more affected by environmental fluctuations than the lower photic zone, in which nannoconids thrived.
The fact that the amplitude of oxygen and carbon isotope fluctuations recorded by coccoliths is higher than that recorded by nannoconids during the deposition of marl and limestone supports the existence in Early Cretaceous oceans of an upper photic zone, inhabited by coccolithophorids, which was more affected by environmental fluctuations than the lower photic zone, in which nannoconids thrived.
![]() If we assume that oxygen isotope ratio fluctuations are related only to temperature variations, the
(1973) paleotemperature equation indicates that, the lower δ18O recorded in marls samples, would reflect a temperature increase of about 7°C in the surface layer of the sea, and of only 0.8°C in the deeper part of the photic zone. Conversely, if we interpret the oxygen isotope fluctuations in terms of both sea-water δ18O and salinity variations, assuming that salinity increases by 1‰ when the δ18O of sea-water rises by 1.2‰ (,
1987), during the formation of marly layers the salinity of surface water might have lowered about 1.5‰ and about 0.2‰ in the lower photic zone. Regarding alternations in the Pliocene succession of South Sicily, et alii
(in press) have estimated the respective roles of temperature and salinity variations in causing fluctuations in the carbonate δ18O values by combining temperatures derived from the alkenone unsaturation ratio (Uk'37) and coccoliths δ18O data. In the present study, unfortunately we could not extract alkenones from the sediments, so it was not possible to distinguish the respective roles of these two parameters in the lowering of the δ18O recorded in marls. However, as an increase of 7°C in a gradational change from limestones to marls is quite unrealistic, the oxygen isotope record is more likely to be due to the coupled effect of an increase in temperature and a decrease in salinity.
If we assume that oxygen isotope ratio fluctuations are related only to temperature variations, the
(1973) paleotemperature equation indicates that, the lower δ18O recorded in marls samples, would reflect a temperature increase of about 7°C in the surface layer of the sea, and of only 0.8°C in the deeper part of the photic zone. Conversely, if we interpret the oxygen isotope fluctuations in terms of both sea-water δ18O and salinity variations, assuming that salinity increases by 1‰ when the δ18O of sea-water rises by 1.2‰ (,
1987), during the formation of marly layers the salinity of surface water might have lowered about 1.5‰ and about 0.2‰ in the lower photic zone. Regarding alternations in the Pliocene succession of South Sicily, et alii
(in press) have estimated the respective roles of temperature and salinity variations in causing fluctuations in the carbonate δ18O values by combining temperatures derived from the alkenone unsaturation ratio (Uk'37) and coccoliths δ18O data. In the present study, unfortunately we could not extract alkenones from the sediments, so it was not possible to distinguish the respective roles of these two parameters in the lowering of the δ18O recorded in marls. However, as an increase of 7°C in a gradational change from limestones to marls is quite unrealistic, the oxygen isotope record is more likely to be due to the coupled effect of an increase in temperature and a decrease in salinity.
![]() Calcite particles known as micarb are the dominant carbonate components in the samples of La Marcouline Quarry section studied here. Isotope signatures of micarb-dominated fine fractions are different from those of the coccoliths values but very close to those of nannoconids (Fig. 3
Calcite particles known as micarb are the dominant carbonate components in the samples of La Marcouline Quarry section studied here. Isotope signatures of micarb-dominated fine fractions are different from those of the coccoliths values but very close to those of nannoconids (Fig. 3 ![]() ). This geochemical similarity causes us consider micarb as the result of Nannoconus spp. fragmentation. SEM observations and mineralogical analysis (Differential Thermic Analysis-Thermogravimetry technique, et alii,
2005; , 2006; et alii,
2006) support this hypothesis. This interpretation is in agreement with the observations made by
and (1990) concerning the fragility of Nannoconus spp. because of the easy disaggregation of their elements.
and (1983),
(1992), et alii
(1994) and
and (2002) have already postulated a biogenic origin for these microparticles from nannofossil disaggregation. For the
Lower Cretaceous sequence of La Marcouline, geochemical data demonstrate the relationship between nannoconids and micarb. Assuming that the majority of micarb represent nannoconid fragments, the tests of these organisms may represent up to 80% of the volume of the carbonate fraction which corresponds to about 43% of the volume of the sediment in marl and limestone samples.
). This geochemical similarity causes us consider micarb as the result of Nannoconus spp. fragmentation. SEM observations and mineralogical analysis (Differential Thermic Analysis-Thermogravimetry technique, et alii,
2005; , 2006; et alii,
2006) support this hypothesis. This interpretation is in agreement with the observations made by
and (1990) concerning the fragility of Nannoconus spp. because of the easy disaggregation of their elements.
and (1983),
(1992), et alii
(1994) and
and (2002) have already postulated a biogenic origin for these microparticles from nannofossil disaggregation. For the
Lower Cretaceous sequence of La Marcouline, geochemical data demonstrate the relationship between nannoconids and micarb. Assuming that the majority of micarb represent nannoconid fragments, the tests of these organisms may represent up to 80% of the volume of the carbonate fraction which corresponds to about 43% of the volume of the sediment in marl and limestone samples.
![]() There are two end-member models in the current literature for the origin of limestone-marl alternations. They could reflect either a primary environmental signal, or alternatively they could be created merely by diagenesis.
There are two end-member models in the current literature for the origin of limestone-marl alternations. They could reflect either a primary environmental signal, or alternatively they could be created merely by diagenesis.
![]() Current models for the primary origin of limestone-marl alternations infer that at least one component of the original deposit, either the carbonate or the non-carbonate fraction, or both, fluctuated in time (,
1982). These factors include fluctuations in:
Current models for the primary origin of limestone-marl alternations infer that at least one component of the original deposit, either the carbonate or the non-carbonate fraction, or both, fluctuated in time (,
1982). These factors include fluctuations in:
biogenic production of carbonates by calcareous plankton (productivity cycles, , 1982; , 1983);
Export of shallow-water platform carbonate to deeper settings (, 1952; & , 1976; & , 1998; & , 2002);
Input of fine terrigeneous sediment (dilution cycles, , 1982; et alii, 1984);
Degree of saturation of calcium carbonate in sea-water (dissolution cycles, et alii, 1981; , 1982);
Availability of oxygen on the sea floor as it affects the degree of preservation of organic matter (redox cycles, et alii, 1984).
![]() In this study, the higher δ13C values recorded by coccoliths in the marl samples suggest that in the photic zone conditions increasing fertility can result from a greater input of nutrients in times of enhanced runoff (Fig. 3
In this study, the higher δ13C values recorded by coccoliths in the marl samples suggest that in the photic zone conditions increasing fertility can result from a greater input of nutrients in times of enhanced runoff (Fig. 3 ![]() ). This view is supported by the lower oxygen isotope ratios recorded in marls (Fig. 3
). This view is supported by the lower oxygen isotope ratios recorded in marls (Fig. 3 ![]() ) that correlate with a lowering of surface salinity during their deposition. In such conditions, in the marl layers the production of calcareous nannofossils may have been enhanced beyond that of the limestones.
) that correlate with a lowering of surface salinity during their deposition. In such conditions, in the marl layers the production of calcareous nannofossils may have been enhanced beyond that of the limestones.
![]() and (2002) have demonstrated in upper Jurassic limestone-marl alternations that trophic fluctuations are well recorded by variations in the composition of coccolith assemblages. If so, similar characteristics should be observable on the couplet scale if trophic changes were in fact the controlling process. In the samples studied, the composition of the sediments (55% nannoconids and 6%
coccoliths, mainly Watznaueria spp. in both lithologies) does not display clear evidence of fluctuations. One possible explanation for this homogeneity is a diagenetic bias.
and (2002) have demonstrated in upper Jurassic limestone-marl alternations that trophic fluctuations are well recorded by variations in the composition of coccolith assemblages. If so, similar characteristics should be observable on the couplet scale if trophic changes were in fact the controlling process. In the samples studied, the composition of the sediments (55% nannoconids and 6%
coccoliths, mainly Watznaueria spp. in both lithologies) does not display clear evidence of fluctuations. One possible explanation for this homogeneity is a diagenetic bias.
![]() The dominance of Nannoconus
spp. observed in the samples could be a result of selective dissolution of less resistant
coccoliths. Watznaueria spp. and Nannoconus spp. are considered dissolution resistant (,
1976;
& , 1986; ,
1992; et alii,
1992; & ,
1995), and their higher concentrations in deposits depleted in other coccoliths can be attributed to diagenetic alteration. Most
nannoconid-rich deposits usually contain coccolith assemblages that are
undiversified. This relationship suggests that community dynamics are controlled by competitive displacement for the use of resources as nutrients.
The dominance of Nannoconus
spp. observed in the samples could be a result of selective dissolution of less resistant
coccoliths. Watznaueria spp. and Nannoconus spp. are considered dissolution resistant (,
1976;
& , 1986; ,
1992; et alii,
1992; & ,
1995), and their higher concentrations in deposits depleted in other coccoliths can be attributed to diagenetic alteration. Most
nannoconid-rich deposits usually contain coccolith assemblages that are
undiversified. This relationship suggests that community dynamics are controlled by competitive displacement for the use of resources as nutrients.
![]() SEM observation of coccolith species considered highly susceptible to dissolution, such as Biscutum
spp. (, 1976) show little evidence of dissolution in our samples. If the above arguments are valid, it would appear that our calcareous nannofossil assemblages are unchanged from their original composition. These observations do not favour the idea that cyclic productivity alone is responsible for the distributions observed.
SEM observation of coccolith species considered highly susceptible to dissolution, such as Biscutum
spp. (, 1976) show little evidence of dissolution in our samples. If the above arguments are valid, it would appear that our calcareous nannofossil assemblages are unchanged from their original composition. These observations do not favour the idea that cyclic productivity alone is responsible for the distributions observed.
![]() As oxygen and carbon isotopic data suggest, dilution cycles, that is to say periodic changes in the supply of non-carbonate material in addition to steady carbonate productivity, are conceivable. If dilution cycles were the dominant cause of limestone marl alternations, et alii
(1977) consider that there might exist a difference in the mineralogy of clay assemblages between periods of strong (marl layers) and weak dilution
(marly limestones). In our study, clay mineralogy is identical in high- and low-carbonate samples, mainly kaolinite and
illite. This fact does not rule out dilution but is a strong indication that dilution cycles are not the main processes responsible for the formation of these sedimentary cycles.
As oxygen and carbon isotopic data suggest, dilution cycles, that is to say periodic changes in the supply of non-carbonate material in addition to steady carbonate productivity, are conceivable. If dilution cycles were the dominant cause of limestone marl alternations, et alii
(1977) consider that there might exist a difference in the mineralogy of clay assemblages between periods of strong (marl layers) and weak dilution
(marly limestones). In our study, clay mineralogy is identical in high- and low-carbonate samples, mainly kaolinite and
illite. This fact does not rule out dilution but is a strong indication that dilution cycles are not the main processes responsible for the formation of these sedimentary cycles.
![]() The rhythmic alternations examined could have undergone differential diagenesis processes. The model proposed by
and (1996) assume an aragonite dissolution in marl layers and calcite reprecipitation in limestone beds in shallow marine burial setting. This model could explain the high degree of preservation of
(calcitic) calcareous nannofossils in the samples studied. Moreover, the paleogeographic setting of La Marcouline succession is consistent with the idea of aragonite export from adjacent shallow-water platforms. An estimate of the composition of the precursor sediments using the numerical model of et alii
(2001) indicates that it would have contained about 4 per cent aragonite. The main difficulty in accepting this hypothesis is that, according to et alii
(2001), such a small quantity of initial aragonite is not enough to generate the formation of limestone marl alternations.
The rhythmic alternations examined could have undergone differential diagenesis processes. The model proposed by
and (1996) assume an aragonite dissolution in marl layers and calcite reprecipitation in limestone beds in shallow marine burial setting. This model could explain the high degree of preservation of
(calcitic) calcareous nannofossils in the samples studied. Moreover, the paleogeographic setting of La Marcouline succession is consistent with the idea of aragonite export from adjacent shallow-water platforms. An estimate of the composition of the precursor sediments using the numerical model of et alii
(2001) indicates that it would have contained about 4 per cent aragonite. The main difficulty in accepting this hypothesis is that, according to et alii
(2001), such a small quantity of initial aragonite is not enough to generate the formation of limestone marl alternations.
![]() Because the calcareous nannofossils are preserved to the same degree of excellence in both lithotypes, the "La Marcouline" section has provided a fine opportunity to study paleoenvironmental conditions prevailing during the deposition of these rhythmites.
Because the calcareous nannofossils are preserved to the same degree of excellence in both lithotypes, the "La Marcouline" section has provided a fine opportunity to study paleoenvironmental conditions prevailing during the deposition of these rhythmites.
![]() The geochemical results obtained on bulk carbonate and separated biogenic- dominated fine fractions show that there was no late burial diagenesis in these sediments. Consequently, bulk carbonate provides a reliable approximation of primary fluctuations in the sea water during the formation of marl-limestone alternations.
The geochemical results obtained on bulk carbonate and separated biogenic- dominated fine fractions show that there was no late burial diagenesis in these sediments. Consequently, bulk carbonate provides a reliable approximation of primary fluctuations in the sea water during the formation of marl-limestone alternations.
![]() Isotopic and mineralogic data show that the calcite microparticles are derived mainly from the fragmentation of Nannoconus spp. tests. This implies that during the
early Gargasian of the Cassis area nannoconids were the main producers of pelagic carbonates (up to 50% of the total sediment), while coccoliths represent only 6% of the sediment. From a comparison of the ratios of stable isotopes of coccoliths and coexisting nannoconids, we infer that the Nannoconus spp. inhabited an ecological niche within the lower photic zone which is consistent with the hypothesis of
(1994) and (2003).
Isotopic and mineralogic data show that the calcite microparticles are derived mainly from the fragmentation of Nannoconus spp. tests. This implies that during the
early Gargasian of the Cassis area nannoconids were the main producers of pelagic carbonates (up to 50% of the total sediment), while coccoliths represent only 6% of the sediment. From a comparison of the ratios of stable isotopes of coccoliths and coexisting nannoconids, we infer that the Nannoconus spp. inhabited an ecological niche within the lower photic zone which is consistent with the hypothesis of
(1994) and (2003).
![]() Even using this multiproxy approach, no single primary mechanism can be proposed as responsible for the formation of these Gargasian rhythmites. Based on coccolith isotopic data, we have identified a possible interaction of nutrient levels, terrestrial influx, and temperature and salinity that could have initiated productivity and dilution cycles causing rhythmites. But these processes remain difficult to confirm with regard to their effects on the composition of the two lithologies.
Even using this multiproxy approach, no single primary mechanism can be proposed as responsible for the formation of these Gargasian rhythmites. Based on coccolith isotopic data, we have identified a possible interaction of nutrient levels, terrestrial influx, and temperature and salinity that could have initiated productivity and dilution cycles causing rhythmites. But these processes remain difficult to confirm with regard to their effects on the composition of the two lithologies.
![]() We are grateful to the reviewers A. and E. for their
constructive criticism of the manuscript. Special thanks are due Nestor
for language corrections and improvement of the text. We would also
like to thank Nathalie (Laboratoire Biominéralisations et
Paleoenvironnements, UPMC) for her efficient technical assistance in this work.
This study was financially supported by UPMC research grants (BQR 2004/2006).
We are grateful to the reviewers A. and E. for their
constructive criticism of the manuscript. Special thanks are due Nestor
for language corrections and improvement of the text. We would also
like to thank Nathalie (Laboratoire Biominéralisations et
Paleoenvironnements, UPMC) for her efficient technical assistance in this work.
This study was financially supported by UPMC research grants (BQR 2004/2006).
M.A., W.E., D. & P.A. (1984).- Rhythmic bedding events in Mesozoic-Cenozoic pelagic carbonate sequences: the primary and diagenetic origin of -like cycles.- In: A., J., J., G. & B. (eds.), and climate.- Riedel, Hingham, p. 191-222.
E.J. (1983).- A warm equable Cretaceous: the nature of the problem.- Earth-Science Reviews, Amsterdam, vol. 19, p. 305-338.
C. (2006).- Variations des paramètres de l'environnement océanique au cours de la sédimentation d'un doublet marne-calcaire. Approches géochimique, minéralogique et micropaléontologique.- Thèse de Doctorat n° 2006-05, Université Pierre et Marie Curie, Paris, 252 p.
C., M. de, F., M., M.A. & U. (in press).- Coccolith δ18O and alkenone records in middle Pliocene orbitally-controlled deposits: high frequency temperature and salinity variations of sea surface water.- Geochemistry, Geophysics, Geosystems (G3), Washington, DOI 10.1029/2006gc001483.
B.E., H.J., D.W. & J. (2000).- Temperature influence on the carbon isotopic composition of Globigerina bulloides and Orbulina universa (planktonic foraminifera).- Marine Micropaleontology, Amsterdam, vol. 38, p. 213-228.
P.L. de (1983).- Aspects of the Middle Cretaceous pelagic sedimentation in Southern Europe.- Thesis Univ. Utrecht, Geologica Ultraiectina, Mededelingen van het Instituut voor Aardwetenschappen der Rijksuniversiteit te Utrecht, n° 31, 112 p.
J.G. (1997).- L'Aptien et l'Albien de la fosse Vocontienne (des bordures au bassin). Évolution de la sédimentation et enseignements sur les évènements anoxiques.- Publication - Société Géologique du Nord, Lille, vol. 25, 614 p.
G. & D. (1991).- Les nannoconidés, indicateurs environnementaux des océans et mers épicontinentales du Jurassique terminal et du Crétacé inférieur.- Oceanologica Acta, Paris, vol. 14, n° 4, p. 333-356.
R., E. & I. (1992).- Barremian-Aptian calcareous plankton biostratigraphy from the Gorgo Cerbara section (Marche, central Italy) and implications for plankton evolution.- Cretaceous Research, Oxford, vol. 13, p. 517-537.
H.E. & R.M. (1983).- Diagenesis of deep-sea carbonates.- In: G. & G.V. (eds.), Diagenesis in sediments and sedimentary rocks. 2.- Development in Sedimentology, Amsterdam, 25B, p. 213-288.
W.E., J.V. & P. (1981).- Tertiary carbonate-dissolution cycles on the Sierra Leone Rise, eastern equatorial Atlantic Ocean.- Marine Geology, Amsterdam, vol. 39, p. 81-101.
W.E., J.V., L.F., P. & E. (1977).- Cyclic sedimentation along the continental margin of Northwest Africa.- In: Y., E. et alii (eds.), Initial Reports of the Deep Sea Drilling Project, Washington, vol. 41, p. 965-989.
W.C., J.C., P.L., L.E. & R.R.L. (1980).- Coccoliths in Pleistocene-Holocene nannofossil assemblages.- Nature, London, vol. 285, p. 222-223.
W.C., P.L., L.E. & J.C. (1986).- Stable isotopic composition of coccoliths.- Marine Micropaleontology, Amsterdam, vol. 10, p. 1-8.
G. (1982).- Limestone-marl cycles (periodites): diagnosis, significance, causes – a review. In: G. & A. (eds.), Cyclic and event stratification.- Springer-Verlag, p. 8-53.
E. (1992).- Calcareous nannofossil distribution in pelagic rhythmic sediments (Aptian-Albian Piobbico core, central Italy).- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 97, n° 3-4, p. 455-484.
E. (1994).- Nannofossils and superplumes: The Early Aptian "nannoconids crisis".- Paleoceanography, Washington, vol. 9, n° 3, p. 483-501.
A.G. (1982).- Long-term oscillations recorded in stratigraphy.- In: W.H. & J.C. (eds.), Climate in Earth history.- National Academic Press, Washington, p. 97-104.
G. (1987).– Dynamique du Bassin subalpin méridional de l'Aptien au Cénomanien.- Thèse, Université Pierre et Marie Curie, Paris VI, Mémoires des Sciences de la Terre, École des Mines de Paris, n° 4, 370 p.
J.O. (2003).- Reconstructing nutricline dynamics of mid-Cretaceous oceans: evidence from calcareous nannofossils from the Niveau Paquier black shale (SE France).- Marine Micropaleontology, Amsterdam, vol. 47, p. 307-321.
E. & B. (2002).- Contribution of calcareous nannoplankton to carbonate deposition: a new approach applied to the Lower Jurassic of central Italy.- Marine Micropaleontology, Amsterdam, vol. 45, p. 175-190.
F. (2002).- Mise au point d'un protocole de séparation des assemblages de nannofossiles calcaires. Apports à la micropaléontologie et à la géochimie des producteurs carbonatés pélagiques. Application à la crise Crétacé-Tertiaire.- Thèse, Université Pierre et Marie Curie, Paris VI, 248 p.
F., S., E., M. & S. (2001).- Mise au point d'un protocole expérimental de séparation granulométrique d'assemblages de nannofossiles calcaires : application paléoécologique et géochimique.- Bulletin de la Société Géologique de France, Paris, vol. 172, n° 4, p. 437-446.
F., M. de, M. & S. (2004).- Remaniement des nannofossiles calcaires maastrichtiens dans les sédiments du Danien basal de Bidart (France) : arguments isotopiques (carbone et oxygène).- Revue de Micropaléontologie, Paris, vol. 47, p. 145-152.
B. & A. (1990a).- Nutricline variations in the equatorial Atlantic coincident with the younger Dryas.- Paleoceanography, Washington, vol. 5, p. 997-1008.
B. & A. (1990b).- Precessional forcing of nutricline dynamics in the equatorial Atlantic.- Science, Washington, vol. 249, p. 766-769.
M. (1966).- Étude stratigraphique et micropaléontologique du Crétacé inférieur de la "fosse vocontienne".- Documents des Laboratoires de Géologie de la Faculté des Sciences de Lyon, n° 15, 369 p.
M., G., W., M. & J.-P. (2004).- Le Gargasien (Aptien moyen) de Cassis-La Bédoule (stratotype historique de l'Aptien inférieur, SE France) : localisation géographique et corrélations stratigraphiques.- Carnets de Géologie / Notebooks on Geology, Maintenon, Note brève 2004/02, p. 1-4.
A. & C. (1996).- The formation of micritic limestones and the development of limestone-marl alternations in the Silurian of Gotland, Sweden.- Facies, Erlangen, vol. 34, p. 159-176.
A., H., M. & J.J.G. (2001).- The mineralogical composition of precursor sediments of calcareous rhythmites: a new approach.- International Journal of Earth Sciences, Berlin, vol. 90, p. 795-812.
D. & G. (1990).- L'importance des Schizosphères, Stomiosphères, Conusphaera et Nannoconus dans la genèse des calcaires fins pélagiques du Jurassique et du Crétacé inférieur.- Bulletin de la Société Géologique de France, Paris, vol. 43, p. 63-93.
D., G., A., Y. & A.M. (1994).- Contribution fondamentale des Coccolithophoridées à la constitution des calcaires fins pélagiques du Jurassique moyen et supérieur.- In: 3e Symposium International sur la stratigraphie du Jurassique, Poitiers, 1991.- Geobios, Lyon, vol. 17, p. 701-721.
H. & S. (1973).- The distribution of oceanic coccolithophorids in the Pacific.- Deep-Sea Research, Oxford, vol. 20, p. 355-374.
A., F., C., M., M. de & M. (2006).- Caractérisation de biominéraux carbonatés dans les fractions fines des sédiments par DRX et ATD.- Journées SFMC - Biominéralisations 2006, Nancy, July 2006.
B. & A. (1998).- Depositional sequences in deep-shelf environments formed through carbonate-mud import from shallow platform (Late Oxfordian, German Swabian Alb and eastern Swiss Jura).- Eclogae geologicae Helveticae, Basel, vol. 91, p. 149-169.
B. & E. (2002).- The carbonate signal and calcareous nannofossil distribution in an Upper Jurassic section (Balingen-Tieringen, Late Oxfordian, southern Germany).- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, vol. 179, p. 71-96.
W.L. & J.D. (1976).- Late Pleistocene faunal and temperature patterns of the Colombia Basin, Caribbean Sea.- In: R.M. & J.D. (eds.), Investigation of Late Quaternary paleoceanography and paleoclimatology.- Geological Society of America, Memoir, Boulder, vol. 145, p. 201-220.
M., M. de, L., C., M. & G. (in press).- Inorganic geochemistry of the Gargasian from La Marcouline quarry (Cassis, France).- Carnets de Géologie / Notebooks on Geology, Brest.
P.H. & K.R. (1986).- Middle Cretaceous calcareous nannofossil biogeography and preservation in the Atlantic and Indian oceans: Implications for paleoceanography.- Marine Micropaleontology, Amsterdam, vol. 10, p. 235-266.
A.A. & S. (1997).- Braarudosphaera blooms and anomalous enrichments of Nannoconus: evidence from the Turonian South Atlantic, Santo Basin, Brazil.- Journal of Nannoplankton Research, London, vol. 19, p. 51-55.
E. (1952).- Chemische Untersuchungen zur Bankung im unteren Malm Schwabens.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgardt, vol. 95, p. 337-370.
N.J. (1973).- Attainment of isotopic equilibrium between ocean water and the benthonic foraminifera genus Uvigerina: Isotopic change in the ocean during the last glacial.- Colloque International du C.N.R.S., Gif-sur-Yvette, n° 219, p. 203-209.
N.J. (1987).- Oxygen isotopes, ice volume and sea-level.- Quaternary Science Reviews, Amsterdam, vol. 6, p. 183-190.
F., P., M., A., R., F., M.J., P., C.P., C. & J. (2005).- Search for past life on Mars: Physical and chemical characterization of minerals of biotic and abiotic origin: part 1 – Calcite.- Geophysical Research Letters, Washington, vol. 32, n° 23, p. 1-5, L23205, doi: 10.1029/2005GL023403.
H.M. & P. (2004).- Coccolithophore-based geochemical proxies. In: H. & J. (eds.) Coccolithophores: from molecular processes to global impact.- Springer Verlag, p. 529-562.
H.R. (1976).- Mesozoic calcareous nannoplankton biostratigraphy of marine sediments.- Marine Micropaleontology, Amsterdam, vol. 1, p. 325-362.
H. (1990).- Siliciclastics in the Early Cretaceous Tethys and North Atlantic oceans: documents of periodic greenhouse climate conditions.- Memorie della Società Geologica Italiana, Roma, vol. 44, p. 59-69.
J.R. & T.J. (1995).- Nannofossil assemblages, fine fraction stable isotopes, and the paleoceanography of the Valanginian-Barremian (Early Cretaceous) North Sea Basin.- Paleoceanography, Washington, vol. 10, p. 815-839.
Click on thumbnail to enlarge the image.
Figure 1: Presentation of the lithology and the carbonate fluctuations of "La Marcouline" quarry section ( et alii, 2004). Detail of the section selected for this study.
Figure 1 : Représentation schématique et évolution à long terme du CaCO3 des cycles sédimentaires de la carrière de La Marcouline ( et alii, 2004). Détail de la portion de coupe présentée dans cette étude.
Click on thumbnail to enlarge the image.
Figure 2: Evolution of the oxygen and carbon isotopic ratios (‰ vs PDB) of bulk carbonate, Nannoconus spp. (8-5 µm fraction) and micarb (< 3 µm fraction) throughout the studied section.
Figure 2 : Évolutions des rapports isotopiques de l'oxygène et du carbone du carbonate total, des Nannoconus spp. (fraction 8-5 µm) et des micarb (fraction < 3 µm) en fonction de la lithologie.
Click on thumbnail to enlarge the image.
Figure 3: Composition of the oxygen and carbon isotopes of the main carbonate components (‰ PDB). Solid color symbols indicate the mean isotopic values of marls, open ones those of marly limestones. The range of isotopic variations are also reported.
Figure 3 : Rapports isotopiques du carbone et de l'oxygène des principaux constituants carbonatés (‰ PDB). Les valeurs moyennes mesurées dans les échantillons marneux et carbonatés sont représentées en figurés respectivement pleins et vides. Sont également indiqués les intervalles de variation des données isotopiques.
| Fractions | Size spectrum (mm) |
Unknown carbonate particles | Biogenic particles | |||
| Calcitic Macrocrystals |
Ankeritic Macrocrystals |
Micarb | Coccoliths | Nannoconids | ||
| 12 | > 12 | 49% | 17% | 34% | ||
| 8 | 8-12 | 25% | 5% | 2% | 68% | |
| 5 | 5-8 | 2% | 10% | 1% | 87% | |
| 3 | 3-5 | 60% | 7% | 33% | ||
| < 3 | < 3 | 90% | 10% | |||
Click on thumbnail to enlarge the image.
Table 1: Mean composition of the separated fractions and size spectrum of the main carbonate particles of the sediments.
Tableau 1 : Spectres de taille des principales particules carbonatées trouvées dans les sédiments et compositions moyennes des fractions séparées.
| Unknown carbonate particles | Biogenic particles | |||||
| Calcitic Macrocrystals |
Ankeritic Macrocrystals |
Micarb | Coccoliths | Nannoconids | Foraminifera | |
| MARLS | 6% | 2% | 58% | 8% | 25% | 1% |
| MARLY LIMESTONES | 6% | 2% | 63% | 8% | 20% | 1% |
Click on thumbnail to enlarge the image.
Table 2: Mean composition of the carbonate fraction (A) and of the sediment (B) related to lithology.
Tableau 2 : Compositions moyennes des phases carbonatées (A) et des échantillons de marnes et de calcaires marneux (B).
| Sample | Lithology | % CaCO3 |
% Macro- crystal |
% Micarb |
% Nannoconus spp. |
% coccolith |
δ18O (8-5) | δ13C (8-5) | δ18O (< 3) | δ13C (< 3) | δ18O (coccoliths) calculated |
δ13C (coccoliths) calculated |
| 29-1 | marl | 54 | 7 | 60 | 25 | 8 | -1.47 | 3.78 | -1.47 | 3.86 | -4.6 | 5.6 |
| 29-2 | marl | 63 | 6 | 50 | 37 | 6 | -1.3 | 3.9 | -1.48 | 3.82 | ||
| 29-3 | marl | 63 | 6 | 60 | 24 | 8 | -1.7 | 3.63 | -1.49 | 3.82 | -2.5 | 4.7 |
| 29-4 | marl | 66 | 7 | 58 | 26 | 7 | -1.65 | 3.67 | -1.5 | 3.77 | -4.4 | 4.3 |
| 30-1 | marly limestone | 63 | 7 | 56 | 29 | 6 | -1.67 | 3.38 | -1.5 | 3.6 | -2.6 | 2.9 |
| 30-2 | marly limestone | 67 | 7 | 60 | 25 | 7 | -1.76 | 3.46 | -1.71 | 3.6 | -1.9 | 2.8 |
| 30-3 | marly limestone | 82 | 8 | 66 | 17 | 8 | -1.81 | 3.38 | -1.72 | 3.64 | ||
| 31-1 | marl | 67 | 7 | 60 | 22 | 10 | -1.61 | 3.57 | -1.59 | 3.64 | ||
| 31-3 | marl | 60 | 6 | 62 | 24 | 7 | -1.61 | 3.8 | -1.58 | 3.93 | ||
| 31-4 | marl | 58 | 7 | 61 | 21 | 10 | -1.69 | 3.83 | -1.57 | 3.93 | -4.6 | 4.3 |
| 31-5 | marly limestone | 60 | 6 | 61 | 24 | 8 | -1.5 | 3.81 | -1.62 | 3.78 | ||
| 31-6 | marly limestone | 66 | 5 | 69 | 16 | 9 | -1.62 | 3.72 | -1.52 | 3.76 | -2.6 | 2.9 |
| 31-7 | marly limestone | 74 | 5 | 63 | 16 | 14 | -1.86 | 3.31 | -1.8 | 3.41 | ||
| 31-8 | marl | 72 | 7 | 58 | 25 | 8 | -1.81 | 3.55 | -1.62 | 3.59 | -3.1 | 3.4 |
| 31-9 | marl | 64 | 6 | 60 | 24 | 8 | -1.68 | 3.81 | -1.41 | 3.95 | ||
| 31-10 | marl | 60 | 6 | 60 | 24 | 8 | -1.53 | 3.72 | -1.48 | 3.94 | ||
| 32-1 | marly limestone | 66 | 6 | 67 | 18 | 8 | -1.75 | 3.48 | -1.68 | 3.56 | -2.3 | 2.9 |
Click on thumbnail to enlarge the image.
Appendix: Compositions of the carbonated fractions of each sample studied and, carbon and oxygen isotopic ratios (in per mil vs PDB) of the main calcareous components.
Annexe : Composition des phases carbonatées de chaque échantillon étudié ainsi que les rapports isotopiques (en pour mille par rapport au standard PDB) du carbone et de l'oxygène des différents constituants carbonatés.