![]()
![]()
de F. de, P., P., C., P. & M. (2007).- Middle Devonian (Givetian) megaspores from Belgium (Ronquières) and Libya (A1-69 borehole). In: P. & E. (eds.), Recent Advances in Palynology.- Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2007/01, Abstract 11 (CG2007_M01/11).
![]() Megaspores;
Middle Devonian; Givetian; Belgium; Libya; biodiversity.
Megaspores;
Middle Devonian; Givetian; Belgium; Libya; biodiversity.
![]() Mégaspores ;
Dévonien moyen ; Givétien ; Belgique ; Libye ; biodiversité.
Mégaspores ;
Dévonien moyen ; Givétien ; Belgique ; Libye ; biodiversité.
![]() Preliminary results are presented on newly discovered megaspore assemblages recovered from Givetian localities at Ronquières, Belgium, and a borehole (A1-69) drilled
in the Ghadames Basin, Libya, by SHELL in 1959. The miospore biostratigraphy of the Ronquières and of A1-69 borehole sections has been published (
de,
2005; et alii,
2004; & ,
1989; et alii,
1992; et alii,
1990).
Preliminary results are presented on newly discovered megaspore assemblages recovered from Givetian localities at Ronquières, Belgium, and a borehole (A1-69) drilled
in the Ghadames Basin, Libya, by SHELL in 1959. The miospore biostratigraphy of the Ronquières and of A1-69 borehole sections has been published (
de,
2005; et alii,
2004; & ,
1989; et alii,
1992; et alii,
1990).
![]() Thirty-six samples have been studied to date. Eighteen are from the "Plan Incliné
de Ronquières" section. Their weight
ranged between 60 to 75 g. Most of the samples found productive are grey sandstone. The eighteen Libyan samples are from the collections of the University of Liège. Their weight ranged from 4 to 23 g. They are grey sandstone and sandy clay.
Thirty-six samples have been studied to date. Eighteen are from the "Plan Incliné
de Ronquières" section. Their weight
ranged between 60 to 75 g. Most of the samples found productive are grey sandstone. The eighteen Libyan samples are from the collections of the University of Liège. Their weight ranged from 4 to 23 g. They are grey sandstone and sandy clay.
![]() The Libyan samples were immersed in 40% hydrofluoric acid for
3 to 5 days and the Belgian sediments from 5 to 13 days. The rock dissolution was facilitated by a 0.5-1 mm sieve placed under the sample. In
consequence, the sediments were constantly surrounded by acid, and the acid insoluble residue
dropped through the sieve mesh to the bottom of the container during processing. The sample is thus not coated by acid-insoluble material.
The Libyan samples were immersed in 40% hydrofluoric acid for
3 to 5 days and the Belgian sediments from 5 to 13 days. The rock dissolution was facilitated by a 0.5-1 mm sieve placed under the sample. In
consequence, the sediments were constantly surrounded by acid, and the acid insoluble residue
dropped through the sieve mesh to the bottom of the container during processing. The sample is thus not coated by acid-insoluble material.
![]() All residues were washed through a 150 µm sieve. The fraction
of the residue less than 150 µm in diameter was re-sieved in order to collect miospores.
All residues were washed through a 150 µm sieve. The fraction
of the residue less than 150 µm in diameter was re-sieved in order to collect miospores.
![]() Megaspores
were located using a dissecting microscope at high magnification (up to 70x). They
were picked with a pipette and then placed over a slide on which lines had been engraved. After examination of the upper surface
of the megaspore using the SEM, a drop of water was placed on the specimens. This
induced the formation of an air bubble under the specimens;
so they could be turned over to observe the opposite surface.
Megaspores
were located using a dissecting microscope at high magnification (up to 70x). They
were picked with a pipette and then placed over a slide on which lines had been engraved. After examination of the upper surface
of the megaspore using the SEM, a drop of water was placed on the specimens. This
induced the formation of an air bubble under the specimens;
so they could be turned over to observe the opposite surface.
![]() Ronquières is located in the Walloon Brabant Province between the cities of Braine-le-Comte and Nivelles (x=50°36'30"N, y=4°13'30"E). The section is
situated at the "Plan Incliné de Ronquières".
Ronquières is located in the Walloon Brabant Province between the cities of Braine-le-Comte and Nivelles (x=50°36'30"N, y=4°13'30"E). The section is
situated at the "Plan Incliné de Ronquières".
![]() The sediments of the
Givetian at Ronquières are on the northern flank of the Namur Syncline. The megaspores were isolated from grey sandstones
in the Bois de Planti Member of the Bois de Bordeaux Formation (
& , 2002;
& , 2002).
Most of these sediments were deposited in a fluvial to near-shore environment. One megaspore-rich level also yielded the proto-ovule Runcaria , recently redescribed by et alii
(2004). This level is no longer accessible.
The sediments of the
Givetian at Ronquières are on the northern flank of the Namur Syncline. The megaspores were isolated from grey sandstones
in the Bois de Planti Member of the Bois de Bordeaux Formation (
& , 2002;
& , 2002).
Most of these sediments were deposited in a fluvial to near-shore environment. One megaspore-rich level also yielded the proto-ovule Runcaria , recently redescribed by et alii
(2004). This level is no longer accessible.
![]() During Middle
Devonian times Belgium was in the Southern Hemisphere on the Euramerican Plate at 20°S.
During Middle
Devonian times Belgium was in the Southern Hemisphere on the Euramerican Plate at 20°S.
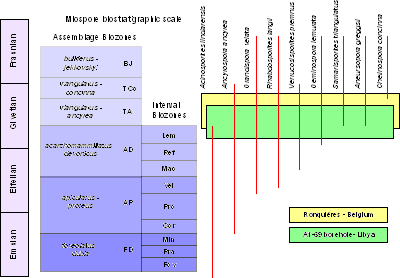
![]() The stratigraphic range of the biostratigraphically most characteristic miospores from
the "Plan Incliné de Ronquières" are illustrated in Figure 1
The stratigraphic range of the biostratigraphically most characteristic miospores from
the "Plan Incliné de Ronquières" are illustrated in Figure 1 ![]() . The miospore assemblage includes, among other species, Acinosporites lindlarensis 1968, Ancyrospora ancyrea var. ancyrea 1965, Grandispora velata () 1971, Rhabdosporites langii () 1960, Verrucosisporites premnus () 1965, Geminospora lemurata 1962, Chelinospora concinna 1965, Samarisporites triangulatus 1965 and Aneurospora greggsii () 1974.
. The miospore assemblage includes, among other species, Acinosporites lindlarensis 1968, Ancyrospora ancyrea var. ancyrea 1965, Grandispora velata () 1971, Rhabdosporites langii () 1960, Verrucosisporites premnus () 1965, Geminospora lemurata 1962, Chelinospora concinna 1965, Samarisporites triangulatus 1965 and Aneurospora greggsii () 1974.
![]() Those species indicate that the sediments of Ronquières range from the uppermost part of the Lem Interval Zone (G. lemurata Interval Zone) of the AD Oppel Zone (A.
acanthomammillatus - D. devonicus Oppel Zone) to the lowermost part of the TCo Oppel Zone (S. triangulatus - C. concinna Oppel Zone) ( et alii,
1987).
The Ronquières locality is thus middle to late Givetian in age.
Those species indicate that the sediments of Ronquières range from the uppermost part of the Lem Interval Zone (G. lemurata Interval Zone) of the AD Oppel Zone (A.
acanthomammillatus - D. devonicus Oppel Zone) to the lowermost part of the TCo Oppel Zone (S. triangulatus - C. concinna Oppel Zone) ( et alii,
1987).
The Ronquières locality is thus middle to late Givetian in age.
![]() Nineteen megaspore morphotypes were recognized. The specimens are black and have been altered by thermal diagenesis and damaged by pyritization. This megaspore assemblage is the richest ever described
of Givetian age ( de,
2005).
Nineteen megaspore morphotypes were recognized. The specimens are black and have been altered by thermal diagenesis and damaged by pyritization. This megaspore assemblage is the richest ever described
of Givetian age ( de,
2005).
![]() The megaspores are 200 to 800 µm in diameter. Most are circular;
a few others are subtriangular in outline.
The labra of the trilete mark can be very high (up to 35 µm) to nonexistent and curvaturae may or may not be present. Their ornamentation can consist of grana, coni, spinae, large appendices, or
of a reticulum. The maximum length of the ornamentation ranges from 1.5 to 71.6 µm.
The megaspores are 200 to 800 µm in diameter. Most are circular;
a few others are subtriangular in outline.
The labra of the trilete mark can be very high (up to 35 µm) to nonexistent and curvaturae may or may not be present. Their ornamentation can consist of grana, coni, spinae, large appendices, or
of a reticulum. The maximum length of the ornamentation ranges from 1.5 to 71.6 µm.
![]() Contagisporites optivus () var. optivus 1971
(Pl. 1, fig. 4
Contagisporites optivus () var. optivus 1971
(Pl. 1, fig. 4 ![]() ), and Corystisporites multispinosus 1965
(Pl. 1, fig. 5
), and Corystisporites multispinosus 1965
(Pl. 1, fig. 5 ![]() ) have been identified from this assemblage.
) have been identified from this assemblage.
![]() The A1-69 borehole is located in the Ghadames Basin
of northwestern Libya
(X=29°03'50", Y=13°40'13"E). The 18 samples studied are from the
interval 1,074 feet (327 m) to 1,486 feet (453 m).
The A1-69 borehole is located in the Ghadames Basin
of northwestern Libya
(X=29°03'50", Y=13°40'13"E). The 18 samples studied are from the
interval 1,074 feet (327 m) to 1,486 feet (453 m).
![]() The
samples are from the Ouenine II Formation of the Aywanat Wanin Group (,
1988). The Ouenine II Formation consists of
more than 80 m of a greyish green finely bedded argillaceous sequence with three interbedded sandy units. It is a typical deltaic facies.
The
samples are from the Ouenine II Formation of the Aywanat Wanin Group (,
1988). The Ouenine II Formation consists of
more than 80 m of a greyish green finely bedded argillaceous sequence with three interbedded sandy units. It is a typical deltaic facies.
![]() The paleogeographic position of Libya during the Middle Devonian
was at 40°S on the northern margin of the Gondwana plate.
The paleogeographic position of Libya during the Middle Devonian
was at 40°S on the northern margin of the Gondwana plate.
![]() The stratigraphic range of the most characteristic miospores in borehole A1-69 is
shown on Figure 1
The stratigraphic range of the most characteristic miospores in borehole A1-69 is
shown on Figure 1 ![]() .
.
![]() The
miospore assemblage includes, among other species, A. lindlarensis, Grandispora protea () 1980, Emphanisporites rotatus
() 1973, Grandispora megaformis () 1973, R. langii () 1960, Grandispora libyensis 1980, V. premnus () 1965 and S. triangulatus. This indicates that the Libyan sediments range from the uppermost part of the Lem Interval Zone
to the TA Oppel Zone (S. triangulatus - A. ancyrea Oppel Zone). They are thus middle to late Givetian in age. Hence the samples from Ronquières and Libya are contemporaneous.
The
miospore assemblage includes, among other species, A. lindlarensis, Grandispora protea () 1980, Emphanisporites rotatus
() 1973, Grandispora megaformis () 1973, R. langii () 1960, Grandispora libyensis 1980, V. premnus () 1965 and S. triangulatus. This indicates that the Libyan sediments range from the uppermost part of the Lem Interval Zone
to the TA Oppel Zone (S. triangulatus - A. ancyrea Oppel Zone). They are thus middle to late Givetian in age. Hence the samples from Ronquières and Libya are contemporaneous.
![]() Twelve megaspore morphotypes have been differentiated. Megaspores range in color from light orange to dark brown and are very
well preserved. A few specimens have been altered by pyrite.
Twelve megaspore morphotypes have been differentiated. Megaspores range in color from light orange to dark brown and are very
well preserved. A few specimens have been altered by pyrite.
![]() Most
of the megaspores are circular, but
some are subtriangular in outline. The labra of the trilete mark of some specimens are very high (up to 47.9 µm). The contact area is either poorly
defined or is well marked by a circular thickening, a depression, or
ornamentation. All megaspores from this borehole are ornamented. The length of the ornament ranges from 3.6 to 186.3 µm.
Most
of the megaspores are circular, but
some are subtriangular in outline. The labra of the trilete mark of some specimens are very high (up to 47.9 µm). The contact area is either poorly
defined or is well marked by a circular thickening, a depression, or
ornamentation. All megaspores from this borehole are ornamented. The length of the ornament ranges from 3.6 to 186.3 µm.
![]() Only Contagisporites optivus var. optivus
(Pl. 1, fig. 1
Only Contagisporites optivus var. optivus
(Pl. 1, fig. 1 ![]() ) and Heliotriletes longispinosus
et 1981
(Pl. 1, fig. 2
) and Heliotriletes longispinosus
et 1981
(Pl. 1, fig. 2 ![]() ) have been identified. Some megaspores from this borehole are so well preserved that we are able to see the ultrastructure of the outer wall and to
distinguish differences between the exine layers.
) have been identified. Some megaspores from this borehole are so well preserved that we are able to see the ultrastructure of the outer wall and to
distinguish differences between the exine layers.
![]() We found some very large megaspores with a maximum diameter of 1000 µm and
a length of up to 2000 µm. Some of these megaspores possess a hologula that may reach 1000 µm. They have a spinate or reticulate ornamentation on the central body. The hologula is smooth
(Pl. 1, fig. 6
We found some very large megaspores with a maximum diameter of 1000 µm and
a length of up to 2000 µm. Some of these megaspores possess a hologula that may reach 1000 µm. They have a spinate or reticulate ornamentation on the central body. The hologula is smooth
(Pl. 1, fig. 6 ![]() ).
).
![]() Sediments from both localities are coeval and deposited in
very similar paleoenvironments: continental to deltaic for Ronquières and clearly deltaic for the A1-69 borehole. The
palaeoclimate of each region was discrete, however, warm and arid in Belgium and warm temperate and wet in Libya (,
2001).
Sediments from both localities are coeval and deposited in
very similar paleoenvironments: continental to deltaic for Ronquières and clearly deltaic for the A1-69 borehole. The
palaeoclimate of each region was discrete, however, warm and arid in Belgium and warm temperate and wet in Libya (,
2001).
![]() Cross-plot of maximum ornamentation length / maximum
diameter of the megaspores from the two areas reveal marked differences. Most of
the Libyan megaspores are located in a group characterised by large scale ornamentation
(above 50 µm) and sizes ranging from 300 to 450 µm, while in
most of the specimens of the Belgian material the length of the ornament ranges from 0 to 50 µm.
Cross-plot of maximum ornamentation length / maximum
diameter of the megaspores from the two areas reveal marked differences. Most of
the Libyan megaspores are located in a group characterised by large scale ornamentation
(above 50 µm) and sizes ranging from 300 to 450 µm, while in
most of the specimens of the Belgian material the length of the ornament ranges from 0 to 50 µm.
![]() The megaspore assemblages from
the 2 localities contain many morphotypes as yet undescribed. And they are very different from each over. Indeed, C. optivus var.
optivus is the only species present in both localities. In contrast to the
disparate megaspore assemblages, there are
many more correspondences in the respective miospore assemblages (42% of miospore species of Ronquières
were also recovered from the Libyan borehole). The coefficient of similarity (
& , 1983) is moderate to high for miospores (20.83%), but very low for megaspores (3.23%). As
the two localities are on widely separated palaeoplates, this difference in
coefficients could reflect the fact that because of their smaller size miospores are more easily transported by wind. et alii
(1992) concluded that
many Devonian heterosporous plants were probably aquatic. On the contrary,
miospore-producers lived in many different biotopes ranging from dry to humid. The fact that megaspores are
rarely transported by wind and that they may be produced by a plant living in a restricted aquatic biotope
might force a high level of endemism. The presence of processes (spinate, bifurcate processes) on most of them (81% of all specimens) may corroborate the hypothesis
regarding functional morphology proposed by et alii
(1992) for the processes of Devonian spores. They suggest
that the morphological characters of megaspores may have been developed to provide buoyancy, movement or attachment for some plants
living permanently, temporarily or for a particular phase of their life cycle in
an aquatic environment. Alternatively, these processes may have been used to
facilitate the adherence of the microspores.
The megaspore assemblages from
the 2 localities contain many morphotypes as yet undescribed. And they are very different from each over. Indeed, C. optivus var.
optivus is the only species present in both localities. In contrast to the
disparate megaspore assemblages, there are
many more correspondences in the respective miospore assemblages (42% of miospore species of Ronquières
were also recovered from the Libyan borehole). The coefficient of similarity (
& , 1983) is moderate to high for miospores (20.83%), but very low for megaspores (3.23%). As
the two localities are on widely separated palaeoplates, this difference in
coefficients could reflect the fact that because of their smaller size miospores are more easily transported by wind. et alii
(1992) concluded that
many Devonian heterosporous plants were probably aquatic. On the contrary,
miospore-producers lived in many different biotopes ranging from dry to humid. The fact that megaspores are
rarely transported by wind and that they may be produced by a plant living in a restricted aquatic biotope
might force a high level of endemism. The presence of processes (spinate, bifurcate processes) on most of them (81% of all specimens) may corroborate the hypothesis
regarding functional morphology proposed by et alii
(1992) for the processes of Devonian spores. They suggest
that the morphological characters of megaspores may have been developed to provide buoyancy, movement or attachment for some plants
living permanently, temporarily or for a particular phase of their life cycle in
an aquatic environment. Alternatively, these processes may have been used to
facilitate the adherence of the microspores.
![]() Very large megaspores, comparable
to those we collected from the Libyan borehole, are extremely rare in sediments older than Carboniferous (,
1967; , 1988)
with the exception of that reported in the Bois de Bordeaux Formation by and
(1969). TEM study of specimens will help to
determine whether or not our megaspores possess an ultrastructure indicating lycopsid affinities (,
1980; , 1981).
Very large megaspores, comparable
to those we collected from the Libyan borehole, are extremely rare in sediments older than Carboniferous (,
1967; , 1988)
with the exception of that reported in the Bois de Bordeaux Formation by and
(1969). TEM study of specimens will help to
determine whether or not our megaspores possess an ultrastructure indicating lycopsid affinities (,
1980; , 1981).
![]() Those results show that:
Those results show that:
![]() 1)
static dissolution of sediments allows isolation of some fragile large palynomorphs;
1)
static dissolution of sediments allows isolation of some fragile large palynomorphs;
![]() 2)
the biodiversity of Givetian megaspores is greater than that assumed previously.
A total of 30 different morphotypes have been recognized from Ronquières and
from the borehole A1-69 from Libya, with only one species common to both
localities;
2)
the biodiversity of Givetian megaspores is greater than that assumed previously.
A total of 30 different morphotypes have been recognized from Ronquières and
from the borehole A1-69 from Libya, with only one species common to both
localities;
![]() 3) evolution
in the size of megaspores occured earlier and took place more rapidly than
expected;
3) evolution
in the size of megaspores occured earlier and took place more rapidly than
expected;
![]() 4)
process morphology could be either an adaptation to an aquatic environment ( et alii,
1992) and/or
due to climatic factors.
4)
process morphology could be either an adaptation to an aquatic environment ( et alii,
1992) and/or
due to climatic factors.
![]() We thank M.
for discussion on the SEM micrographs, M. from Aramco (Saudi Arabia) for corrections and comments, and M.
(Liège) for her help with the
processing of our samples.
We thank M.
for discussion on the SEM micrographs, M. from Aramco (Saudi Arabia) for corrections and comments, and M.
(Liège) for her help with the
processing of our samples.
S. (1981).- On a lycopsid cone with winged spores.- Botany Gazette, University of Chicago, vol. 142, n° 2, p. 294-304.
P. & L. (2002). Devonian lithostratigraphic units (Belgium). In: P. & L. (eds.), Lithostratigraphic scale of Belgium.- Geologica Belgica, Brussels, p. 39-69.
W.G. (1967).- Spores and land plants evolution.- Review of Palaeobotany and Palynology, Amsterdam, vol. 1, p. 83-93.
W.G. (1980).- Morphological and ultrastructural features of Nikitinsporites canadensis , a Devonian megaspore from the Frasnian of Canada.- Review of Palaeobotany and Palynology, Amsterdam, vol. 30, n° 1-2, p. 89-99.
D.L. & E.W. (1983).- Paleoenvironmental factors and the distribution of conodonts in Lower Triassic of Svalbard and Nepal.- Fossils and Strata, Oslo, n° 15, p. 171-175.
D.L., R.K. & M.E. (1992).- The functional biology of Devonian spores with bifurcate processes a hypothesis.- Palaeobotanist, Lucknow, vol. 41, p. 67-74.
P., B., M., M. & P. (2004).- Runcaria, a Middle Devonian Seed Plant Precursor.- Science, Washington, vol. 306, n° 5697, p. 856-858.
M. & B. (2002).- Carte géologique de Wallonie 1/25.000 - 39/5-6 - Braine-le-Comte - Feluy.- Ministère de la Région wallonne, Direction générale des Ressources naturelles et de l'Environnement (DGRNE), Service Ressources du Sous-Sol, Bruxelles, 63 p.
S., P., M. & D. (1992).- Biostratigraphie par miospores du Dévonien inférieur à supérieur du sondage MG-1 (Bassin d'Hammadah, Tunisie) - Comparaison avec les données des faunes.- Review of Palaeobotany and Palynology, Amsterdam, vol. 74, p. 193-205.
S. & M. (1989).- Middle-Upper Devonian miospores from the Ghadamis Basin (Tunisia-Libya) : Systematics and Stratigraphy.- Review of Palaeobotany and Palynology, Amsterdam, vol. 58, p. 173-196.
D. (1988, unpublished).- Paléozoïque de Libye occidentale : Stratigraphie et paléogéographie.- Thèse de Doctorat, Université de Nice, 2 vol., 514 p.
C.R.
(2001).- Atlas of Earth
History.- PALEOMAP Project, Arlington, 52 p.
http://www.scotese.com/earth.html
F. & M. (1969).- Une mégaspore de grande taille au sommet du Givétien, à Sart-Dame-Avelines.- Annales de la Société Géologique de Belgique, Liège, vol. 92, n° 1, p. 47-50.
M., M., P., S. & P. (1990).- Lower and Middle Devonian miospore-based stratigraphy in Libya and its relation to the megafloras and faunas.- Review of Palaeobotany and Palynology, Amsterdam, vol. 66, n° 3-4, p. 229-242.
M., K., S., W. & P. (1987).- Spore stratigraphy and correlation with faunas and floras in the type marine Devonian of the Ardenne-Rhenish regions.- Review of Palaeobotany and Palynology, Amsterdam, vol. 50, p. 211.
A. (1988).- Paleopalynology.- Allen and Unwin Inc., Winchester, 600 p.
de F. de (2005).- Reconstitution paléo-environnementale des couches de dépôts du Bois de Bordeaux, Dévonien moyen, bord nord du Synclinorium de Namur.- Mémoire, Université de Liège, 63 p.
Click on thumbnail to enlarge the image.
Figure 1: Biostratigraphic extension of selected species observed in the Ronquières and A1-69 borehole sections. The green and yellow rectangles represent the stratigraphic range of the samples containing megaspores. Biostratigraphic scale after et alii (1987).
Click on thumbnail to enlarge the image.
Plate 1: Some selected miospores from the Ronquières and A1-69 borehole
sections (scale bars = 100 µm):
figure 1. Contagisporites optivus ( 1959) var. optivus 1971. Libyan
specimen;
figure 2. Heliotriletes longispinosus
et 1981. Libyan specimen;
figure 3. Biharisporites sp. Belgian specimen;
figure 4. Contagisporites optivus ( 1959) var.
optivus 1971. Belgian specimen;
figure 5. Corystisporites multispinosus 1965. Belgian
specimen;
figure 6. Undetermined specimen showing a large gula. Libyan specimen.