![]()
![]()
F., F., J.M. & D. (2007).- Les bioévénements de la limite Albien (Vraconnien)-Cénomanien aux marges nord et sud de la Téthys (S.E. de la France et Tunisie centrale). In : L.G., S. & D. (eds.), Relations entre les marges septentrionale et méridionale de la Téthys au Crétacé [Relations between the northern and southern margins of the Tethys ocean during the Cretaceous period].- Carnets de Géologie / Notebooks on Geology, Brest, Mémoire 2007/02, Résumé 01 (CG2007_M02/01)
![]() limite Albien-Cénomanien ; Vraconnien ; ammonites ; foraminifères planctoniques ; Téthys ; SE France ; Tunisie
limite Albien-Cénomanien ; Vraconnien ; ammonites ; foraminifères planctoniques ; Téthys ; SE France ; Tunisie
![]() Albian-Cenomanian boundary; Vraconnian; ammonites; planktonic foraminifera; Tethys; SE France; Tunisia
Albian-Cenomanian boundary; Vraconnian; ammonites; planktonic foraminifera; Tethys; SE France; Tunisia
![]() Dans le chapitre "The Cretaceous Period" ( et alii,
2004,
p. 362) du "Geologic Time Scale", il est précisé pour ce qui concerne l'étage Cénomanien
"the Cenomanian Working Group selected the lowest occurrence of the planktonic foraminifer Rotalipora globotruncanoides (equivalent to R. brotzeni of some studies) as the basal boundary criterion for the Cenomanian Stage, with the Mont Risou section in southeast France as the GSSP section (...). The GSSP was ratified in
2002."
Dans le chapitre "The Cretaceous Period" ( et alii,
2004,
p. 362) du "Geologic Time Scale", il est précisé pour ce qui concerne l'étage Cénomanien
"the Cenomanian Working Group selected the lowest occurrence of the planktonic foraminifer Rotalipora globotruncanoides (equivalent to R. brotzeni of some studies) as the basal boundary criterion for the Cenomanian Stage, with the Mont Risou section in southeast France as the GSSP section (...). The GSSP was ratified in
2002."
![]() Par rapport à cette définition, cinq aspects principaux seront l'objet de la présente contribution :
Par rapport à cette définition, cinq aspects principaux seront l'objet de la présente contribution :
![]() Le groupe des rotaliporides est généralement abondant en Tunisie dans les marnes de la Formation Fahdène où se place la limite Albien (Vraconnien)-Cénomanien. Il est représenté par une large population de morphotypes appartenant aux genres Pseudothalmanninella et Thalmanninella, alors que le genre Rotalipora n'apparaît que dans la partie moyenne du Cénomanien inférieur.
Le groupe des rotaliporides est généralement abondant en Tunisie dans les marnes de la Formation Fahdène où se place la limite Albien (Vraconnien)-Cénomanien. Il est représenté par une large population de morphotypes appartenant aux genres Pseudothalmanninella et Thalmanninella, alors que le genre Rotalipora n'apparaît que dans la partie moyenne du Cénomanien inférieur.
![]() Antérieurement, les genres Rotalipora , 1942 et Thalmanninella , 1946, qui possèdent tous deux des ouvertures ombilicales supplémentaires, ont été séparés sur l'idée que Thalmanninella n'avait pas, en plus, les ouvertures suturales si caractéristiques de Rotalipora. Cette opinion a toutefois été longuement discutée et, alternativement, les genres ont été séparés (par exemple & ,
1956) ou rassemblés (par exemple &
,
1979). Récemment, trois genres ont été de nouveau individualisés : Pseudothalmanninella, Thalmanninella et Rotalipora ( et alii,
2007).
Antérieurement, les genres Rotalipora , 1942 et Thalmanninella , 1946, qui possèdent tous deux des ouvertures ombilicales supplémentaires, ont été séparés sur l'idée que Thalmanninella n'avait pas, en plus, les ouvertures suturales si caractéristiques de Rotalipora. Cette opinion a toutefois été longuement discutée et, alternativement, les genres ont été séparés (par exemple & ,
1956) ou rassemblés (par exemple &
,
1979). Récemment, trois genres ont été de nouveau individualisés : Pseudothalmanninella, Thalmanninella et Rotalipora ( et alii,
2007).
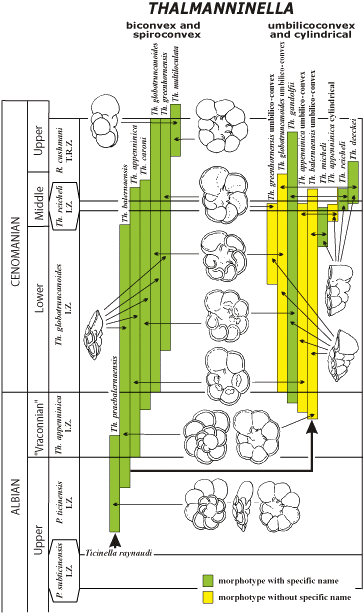
![]() Dans le genre Thalmanninella tel qu'ainsi nouvellement émendé, il semble que deux lignées morphologiques se soient différenciées dès l'Albien supérieur puisque des formes intermédiaires entre ces deux lignées sont très rares
(Fig. 1
Dans le genre Thalmanninella tel qu'ainsi nouvellement émendé, il semble que deux lignées morphologiques se soient différenciées dès l'Albien supérieur puisque des formes intermédiaires entre ces deux lignées sont très rares
(Fig. 1 ![]() ). La première lignée montre une évolution anagénétique dérivant de Ticinella raynaudi et présentant des morphotypes biconvexes et spiroconvexes, avec la succession : praebalernaensis balernaensis appenninica caroni globotruncanoides greenhornensis multiloculata. La deuxième lignée anagénétique se différencie à partir du stade balernaensis avec : balernaensis ombilicoconvexe appenninica ombilicoconvexe gandolfii globotruncanoides ombilicoconvexe greenhornensis ombilicoconvexe. Dans la partie élevée de la zone à Th. globotruncanoides et aussi dans la zone à Th. reicheli ainsi qu'au début de la zone à Rotalipora cushmani, les populations de la lignée des Thalmanninelles ombilicoconvexes différencient des formes cylindriques telles que micheli, appenninica cylindrique, reicheli et deeckei qui sont typologiquement différentes des formes ombilicoconvexes. Ce qui veut dire qu'à la limite Albien-Cénomanien de nombreuses Thalmanninelles à morphologies très variées constituent des populations successives où l'émergence de Th. globotruncanoides s'effectue très progressivement par le moyen d'innombrables intermédiaires, en particulier avec le stade prédécesseur plus primitif représenté par Th. caroni.
). La première lignée montre une évolution anagénétique dérivant de Ticinella raynaudi et présentant des morphotypes biconvexes et spiroconvexes, avec la succession : praebalernaensis balernaensis appenninica caroni globotruncanoides greenhornensis multiloculata. La deuxième lignée anagénétique se différencie à partir du stade balernaensis avec : balernaensis ombilicoconvexe appenninica ombilicoconvexe gandolfii globotruncanoides ombilicoconvexe greenhornensis ombilicoconvexe. Dans la partie élevée de la zone à Th. globotruncanoides et aussi dans la zone à Th. reicheli ainsi qu'au début de la zone à Rotalipora cushmani, les populations de la lignée des Thalmanninelles ombilicoconvexes différencient des formes cylindriques telles que micheli, appenninica cylindrique, reicheli et deeckei qui sont typologiquement différentes des formes ombilicoconvexes. Ce qui veut dire qu'à la limite Albien-Cénomanien de nombreuses Thalmanninelles à morphologies très variées constituent des populations successives où l'émergence de Th. globotruncanoides s'effectue très progressivement par le moyen d'innombrables intermédiaires, en particulier avec le stade prédécesseur plus primitif représenté par Th. caroni.
![]() Le genre Thalmanninella présente des ouvertures supplémentaires ombilicales pouvant devenir suturales entre les dernières loges du dernier tour.
Le genre Thalmanninella présente des ouvertures supplémentaires ombilicales pouvant devenir suturales entre les dernières loges du dernier tour.
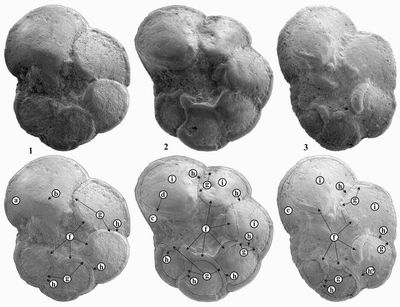
![]() Chaque loge des individus de l'espèce globotruncanoides montre une carène périphérique très nette qui se divise en deux branches, spirale et ombilicale délimitant ainsi une "anterior face" (,
1973) correspondant à une surface en forme de triangle allongé, bien visible à la partie antérieure de la dernière loge
(Pl. I
Chaque loge des individus de l'espèce globotruncanoides montre une carène périphérique très nette qui se divise en deux branches, spirale et ombilicale délimitant ainsi une "anterior face" (,
1973) correspondant à une surface en forme de triangle allongé, bien visible à la partie antérieure de la dernière loge
(Pl. I ![]() ). À la face spirale, une branche de la carène donne une suture en relief entre des loges pétaloïdes tandis qu'à la face ombilicale, l'autre branche se joint au bourrelet périombilical et donne une suture arquée à sigmoïde en relief (ressemblant ainsi à celles d'une Globotruncana, d'où son nom), mais parfois moins saillante entre les dernières loges. Par ailleurs, quand le développement d'une loge cache la branche ombilicale de la loge précédente, la présence d'une "anterior face" sur la dernière loge est certainement le meilleur critère pour séparer Th. globotruncanoides des formes plus primitives dérivées de Th. caroni
(Pl. II
). À la face spirale, une branche de la carène donne une suture en relief entre des loges pétaloïdes tandis qu'à la face ombilicale, l'autre branche se joint au bourrelet périombilical et donne une suture arquée à sigmoïde en relief (ressemblant ainsi à celles d'une Globotruncana, d'où son nom), mais parfois moins saillante entre les dernières loges. Par ailleurs, quand le développement d'une loge cache la branche ombilicale de la loge précédente, la présence d'une "anterior face" sur la dernière loge est certainement le meilleur critère pour séparer Th. globotruncanoides des formes plus primitives dérivées de Th. caroni
(Pl. II ![]() ).
Remarque : on considère ici que Th. globotruncanoides (,
1948, p. 100) est synonyme senior de Th. brotzeni ,
1948, p. 102, suivant les discussions de et (in et alii,
1994) et de et alii
(2007).
).
Remarque : on considère ici que Th. globotruncanoides (,
1948, p. 100) est synonyme senior de Th. brotzeni ,
1948, p. 102, suivant les discussions de et (in et alii,
1994) et de et alii
(2007).
![]() L'espèce prédécesseur Th. caroni ressemble beaucoup à Th. globotruncanoides sauf qu'il n'y a pas d' "anterior face" sur la dernière loge, que les sutures à la face ombilicale sont en relief seulement entre les premières loges du dernier tour, les sutures entre les dernières loges restant radiales et déprimées. On comprend ainsi que, dans une évolution anagénétique, des intermédiaires existent entre des stades successifs et que, sous la loupe binoculaire, il ne soit pas toujours commode de trancher entre l'une et l'autre espèce typologique.
L'espèce prédécesseur Th. caroni ressemble beaucoup à Th. globotruncanoides sauf qu'il n'y a pas d' "anterior face" sur la dernière loge, que les sutures à la face ombilicale sont en relief seulement entre les premières loges du dernier tour, les sutures entre les dernières loges restant radiales et déprimées. On comprend ainsi que, dans une évolution anagénétique, des intermédiaires existent entre des stades successifs et que, sous la loupe binoculaire, il ne soit pas toujours commode de trancher entre l'une et l'autre espèce typologique.
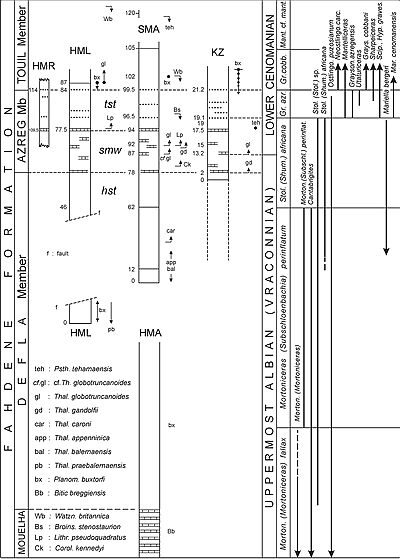
![]() La base du Cénomanien est exposée dans les trois coupes HML, SMA et KZ (dans la région de Kalaat Senan, en Tunisie Centrale), coupes qui ont fourni des foraminifères planctoniques et des ammonites
(Fig. 2
La base du Cénomanien est exposée dans les trois coupes HML, SMA et KZ (dans la région de Kalaat Senan, en Tunisie Centrale), coupes qui ont fourni des foraminifères planctoniques et des ammonites
(Fig. 2 ![]() ) ainsi que des nannofossiles
calcaires ( et alii,
1994).
) ainsi que des nannofossiles
calcaires ( et alii,
1994).
![]() Dans la coupe SMA, les premières Th. globotruncanoides typiques avec "anterior face" bien individualisée sont notées en SMA 92 et des intermédiaires avec Th. caroni (mais plus proches de Th. globotruncanoides) sont déjà fréquents en SMA 87. De ce fait, la base du Cénomanien doit se situer entre SMA 87 et 92. À noter que cette limite se trouve dans le tiers supérieur de la zone d'ammonite à Stoliczkaia (Shumarinaia) africana, essentiellement vraconnienne, et dont seulement la partie sommitale devient cénomanienne par la présence du marqueur micropaléontologique Th. globotruncanoides. En revanche, le grand changement ammonitologique s'effectue 6 à 7m plus haut, en SMA 96,50, avec les premières occurrences de Mantelliceras, Graysonites azregensis, Sciponoceras, Hypoturrilites, Mariella cenomanensi, etc.
(Fig. 2
Dans la coupe SMA, les premières Th. globotruncanoides typiques avec "anterior face" bien individualisée sont notées en SMA 92 et des intermédiaires avec Th. caroni (mais plus proches de Th. globotruncanoides) sont déjà fréquents en SMA 87. De ce fait, la base du Cénomanien doit se situer entre SMA 87 et 92. À noter que cette limite se trouve dans le tiers supérieur de la zone d'ammonite à Stoliczkaia (Shumarinaia) africana, essentiellement vraconnienne, et dont seulement la partie sommitale devient cénomanienne par la présence du marqueur micropaléontologique Th. globotruncanoides. En revanche, le grand changement ammonitologique s'effectue 6 à 7m plus haut, en SMA 96,50, avec les premières occurrences de Mantelliceras, Graysonites azregensis, Sciponoceras, Hypoturrilites, Mariella cenomanensi, etc.
(Fig. 2 ![]() ).
).
![]() Dans la coupe KZ, les premières Th. globotruncanoides, bien que rares, sont présentes à partir du niveau KZ 13,2 qui est le juste équivalent de l'intervalle SMA 87-92 tandis que le grand changement ammonitologique s'effectue environ 6m plus haut et correspond sensiblement au niveau SMA 96,50.
Dans la coupe KZ, les premières Th. globotruncanoides, bien que rares, sont présentes à partir du niveau KZ 13,2 qui est le juste équivalent de l'intervalle SMA 87-92 tandis que le grand changement ammonitologique s'effectue environ 6m plus haut et correspond sensiblement au niveau SMA 96,50.
![]() Toutefois, quand il n'existe qu'une seule coupe disponible et que l'espèce marqueur est rare ou n'est pas bien complètement différenciée de ses prédécesseurs et intermédiaires, il est nécessaire d'avoir recours aux proxies comme les ammonites, les nannofossiles (ou autres événements biologiques ou physico-chimiques) pour estimer correctement la place de la limite d'étage (cf.
Fig. 3
Toutefois, quand il n'existe qu'une seule coupe disponible et que l'espèce marqueur est rare ou n'est pas bien complètement différenciée de ses prédécesseurs et intermédiaires, il est nécessaire d'avoir recours aux proxies comme les ammonites, les nannofossiles (ou autres événements biologiques ou physico-chimiques) pour estimer correctement la place de la limite d'étage (cf.
Fig. 3 ![]() ).
).
![]() Ici également des intermédiaires existent entre Th. globotruncanoides et ses prédécesseurs mais la base du Cénomanien a bien été prise aux premières formes typiques de l'espèce (figs. 10-12 de la fig. 8 de in et alii,
2004, où, par contre, les figs. 4-6 nommées "gandolfii" morphotype qui est l'équivalent ombilicoconvexe de caroni- pourraient peut-être justement illustrer la forme caroni par le fait que le spécimen paraît biconvexe et que seules ses premières sutures ombilicales sont arquées et en relief).
Ici également des intermédiaires existent entre Th. globotruncanoides et ses prédécesseurs mais la base du Cénomanien a bien été prise aux premières formes typiques de l'espèce (figs. 10-12 de la fig. 8 de in et alii,
2004, où, par contre, les figs. 4-6 nommées "gandolfii" morphotype qui est l'équivalent ombilicoconvexe de caroni- pourraient peut-être justement illustrer la forme caroni par le fait que le spécimen paraît biconvexe et que seules ses premières sutures ombilicales sont arquées et en relief).
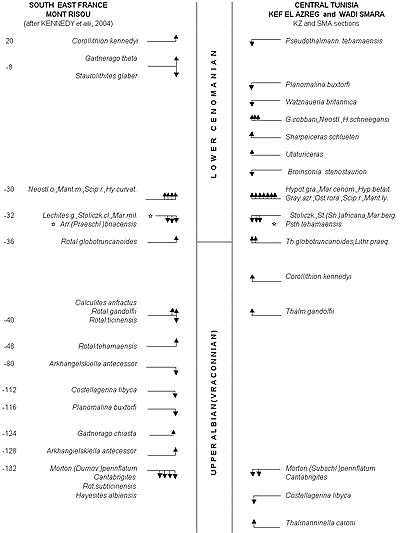
![]() La Figure 4
La Figure 4 ![]() présente une comparaison des bioévénements à proximité de la limite Albien (Vraconnien)-Cénomanien entre le Mont Risou (Téthys Nord) et la coupe KZ en Tunisie Centrale (Téthys Sud). On y remarque que les concordances dans la position des divers bioévénements pour deux régions séparées autrefois par l'Océan téthysien sont très nombreuses
(cf. paragraphe 3).
présente une comparaison des bioévénements à proximité de la limite Albien (Vraconnien)-Cénomanien entre le Mont Risou (Téthys Nord) et la coupe KZ en Tunisie Centrale (Téthys Sud). On y remarque que les concordances dans la position des divers bioévénements pour deux régions séparées autrefois par l'Océan téthysien sont très nombreuses
(cf. paragraphe 3).
![]() Six zones sont actuellement reconnues en Tunisie Centrale avec une limite Albien-Cénomanien se situant vers la base du tiers supérieur de la zone à St. (Sh.) africana, soit, du haut vers le bas :
Six zones sont actuellement reconnues en Tunisie Centrale avec une limite Albien-Cénomanien se situant vers la base du tiers supérieur de la zone à St. (Sh.) africana, soit, du haut vers le bas :
Mantelliceras cf. mantelli IZ |
|
Graysonites cobbani TRZ |
|
Graysonites azregensis IZ |
CÉNOMANIEN INFÉRIEUR |
Stoliczkaia (Shumarinaia) africana PRZ |
------------------------------- |
Mortoniceras (Subschloenbachia) perinflatum TRZ |
ALBIEN SUP. SUP. (VRACONNIEN) |
Mortoniceras (Mortoniceras) fallax IZ |
|
![]() Cette zone est définie comme l'intervalle entre la dernière occurrence de M. (S.) perinflatum et la première occurrence de Graysonites
(Pl. III
Cette zone est définie comme l'intervalle entre la dernière occurrence de M. (S.) perinflatum et la première occurrence de Graysonites
(Pl. III ![]() ), Hypoturrilites, etc. Cet intervalle où il n'y a plus ni Mortoniceras ni Cantabrigites a été indexé dans le Sud-Est de la France par
(1973) avec l'ammonite Arraphoceras (Praeschloenbachia) briacensis. Malheureusement, Arraphoceras est un Hoplitide restreint à la Province Nord-Européenne du domaine boréal et, par là, non utilisable dans la Téthys méridionale. Le choix de l'espèce Stoliczkaia (Shumarinaia) africana , 1907 comme index de la même zone pour la marge sud de la Téthys est justifié par deux raisons : l'espèce est fréquente et elle est cosmopolite (connue en Tunisie, au Nigéria, en Angleterre). Elle est illustrée dans la
Planche IV
), Hypoturrilites, etc. Cet intervalle où il n'y a plus ni Mortoniceras ni Cantabrigites a été indexé dans le Sud-Est de la France par
(1973) avec l'ammonite Arraphoceras (Praeschloenbachia) briacensis. Malheureusement, Arraphoceras est un Hoplitide restreint à la Province Nord-Européenne du domaine boréal et, par là, non utilisable dans la Téthys méridionale. Le choix de l'espèce Stoliczkaia (Shumarinaia) africana , 1907 comme index de la même zone pour la marge sud de la Téthys est justifié par deux raisons : l'espèce est fréquente et elle est cosmopolite (connue en Tunisie, au Nigéria, en Angleterre). Elle est illustrée dans la
Planche IV ![]() .
.
![]() Ces deux genres d'ammonites n'avaient jamais été cités auparavant en Afrique du Nord dans les associations du Cénomanien inférieur alors que l'un ou l'autre sont connus du Texas aux Indes en passant par la Californie, le Mexique, le Brésil, l'Espagne, Madagascar et le Japon. Ils sont décrits et illustrés dans les planches (Graysonites : cf.
Pl. III
Ces deux genres d'ammonites n'avaient jamais été cités auparavant en Afrique du Nord dans les associations du Cénomanien inférieur alors que l'un ou l'autre sont connus du Texas aux Indes en passant par la Californie, le Mexique, le Brésil, l'Espagne, Madagascar et le Japon. Ils sont décrits et illustrés dans les planches (Graysonites : cf.
Pl. III ![]() ).
).
![]() Le Mont Risou est situé dans la Drôme, au milieu du Bassin Vocontien, dans un contexte sédimentaire de marnes bleues pélagiques très épaisses et apparemment sans discontinuités. C'est là qu'a été choisi le GSSP (Global boundary Stratotype, Section and Point) de la base du Cénomanien ( et alii,
2004).
Le Mont Risou est situé dans la Drôme, au milieu du Bassin Vocontien, dans un contexte sédimentaire de marnes bleues pélagiques très épaisses et apparemment sans discontinuités. C'est là qu'a été choisi le GSSP (Global boundary Stratotype, Section and Point) de la base du Cénomanien ( et alii,
2004).
![]() La Figure 4
La Figure 4 ![]() rapporte les principaux bioévénements mis en évidence aux environs de la limite Albien-Cénomanien au Mont Risou et dans la coupe KZ de la Tunisie Centrale. Elle appelle et suggère plusieurs commentaires.
rapporte les principaux bioévénements mis en évidence aux environs de la limite Albien-Cénomanien au Mont Risou et dans la coupe KZ de la Tunisie Centrale. Elle appelle et suggère plusieurs commentaires.
![]() Le terme "Vraconnien" a été introduit par en
1868 pour individualiser une faune intermédiaire entre celles de l'Albien et du Cénomanien typiques. Utilisé en tant que sous-étage de l'Albien par en
1936 et considéré alors comme un équivalent de la zone à Stoliczkaia dispar, le Vraconnien a été exclu des étages crétacés au Colloque sur le Crétacé inférieur à Lyon en 1963 en raison de sa très faible épaisseur dans la région type et, par là, non cartographiable (,
1965).
Le terme "Vraconnien" a été introduit par en
1868 pour individualiser une faune intermédiaire entre celles de l'Albien et du Cénomanien typiques. Utilisé en tant que sous-étage de l'Albien par en
1936 et considéré alors comme un équivalent de la zone à Stoliczkaia dispar, le Vraconnien a été exclu des étages crétacés au Colloque sur le Crétacé inférieur à Lyon en 1963 en raison de sa très faible épaisseur dans la région type et, par là, non cartographiable (,
1965).
![]() Depuis, l'étude de nombreuses coupes dans le SE de la France, à Madagascar, en Californie et en Tunisie a révélé de fortes épaisseurs de Vraconnien (334m à Marcoule, 145m au Mont Risou, 260m à Kalaat Senan, 157m à Diego
Suarez, ... cf. ,
2002). Ces épaisseurs sédimentaires sont souvent beaucoup plus grandes que celles de l'Albien sensu stricto. Par ailleurs, le Vraconnien coïncide avec un événement eustatique de 3ème ordre superposé à un pic de transgression de 2ème ordre au cours desquels les biotas marins se développent énormément.
Depuis, l'étude de nombreuses coupes dans le SE de la France, à Madagascar, en Californie et en Tunisie a révélé de fortes épaisseurs de Vraconnien (334m à Marcoule, 145m au Mont Risou, 260m à Kalaat Senan, 157m à Diego
Suarez, ... cf. ,
2002). Ces épaisseurs sédimentaires sont souvent beaucoup plus grandes que celles de l'Albien sensu stricto. Par ailleurs, le Vraconnien coïncide avec un événement eustatique de 3ème ordre superposé à un pic de transgression de 2ème ordre au cours desquels les biotas marins se développent énormément.
![]() La durée du Vraconnien étant estimée à 2,4 Ma ± 0,2 (voisine de celles du Santonien ou du Coniacien), une réhabilitation de l'étage permettrait de diminuer d'autant celle de l'Albien qui atteint presque 13 Ma. L'étage Vraconnien serait alors composé de trois zones : à Mortoniceras (M.) fallax à la base, puis à Mortoniceras (S.) perinflatum et, au-dessus, à Stoliczkaia (Shumarinaia) africana (dont la partie tout à fait sommitale est d'âge cénomanien par la présence des premières Thalmanninella globotruncanoides).
La durée du Vraconnien étant estimée à 2,4 Ma ± 0,2 (voisine de celles du Santonien ou du Coniacien), une réhabilitation de l'étage permettrait de diminuer d'autant celle de l'Albien qui atteint presque 13 Ma. L'étage Vraconnien serait alors composé de trois zones : à Mortoniceras (M.) fallax à la base, puis à Mortoniceras (S.) perinflatum et, au-dessus, à Stoliczkaia (Shumarinaia) africana (dont la partie tout à fait sommitale est d'âge cénomanien par la présence des premières Thalmanninella globotruncanoides).
![]() À noter que la distribution verticale de Stoliczkaia (St.) dispar est en réalité restreinte à la zone à Mortoniceras (S.) perinflatum et que la "Zone à
Dispar" auct. ne peut en aucun cas être synonyme du Vraconnien.
À noter que la distribution verticale de Stoliczkaia (St.) dispar est en réalité restreinte à la zone à Mortoniceras (S.) perinflatum et que la "Zone à
Dispar" auct. ne peut en aucun cas être synonyme du Vraconnien.
![]() Dans la note sont également discutées des comparaisons avec les coupes de Marcoule (vallée du Rhône), de Hameima (Tunisie Centrale), de Diego Suarez (Madagascar) et de Dry Creek (Californie) où la limite Albien-Cénomanien est bien exposée. Un paragraphe résume aussi les relations entre les changements biologiques et les séquences de dépôt.
Dans la note sont également discutées des comparaisons avec les coupes de Marcoule (vallée du Rhône), de Hameima (Tunisie Centrale), de Diego Suarez (Madagascar) et de Dry Creek (Californie) où la limite Albien-Cénomanien est bien exposée. Un paragraphe résume aussi les relations entre les changements biologiques et les séquences de dépôt.
F. (2002).- Plaidoyer pour un étage Vraconnien entre l'Albien sensu stricto et le Cénomanien (système Crétacé).- Académie Royale de Belgique, Publication de la Classe des Sciences, Bruxelles, (3e série), t. IV, 128 p.
M. (1936).- Les subdivisions du Vraconien dans le Sud-Est de la France.- Bulletin de la Société géologique de France, Paris, (5e série), t. 6, fasc. 1-3, p. 63-68.
P. & N.K. (1956).- Taxonomy of the Globotruncanidae.- Eclogae Geologicae Helvetiae, Basel, vol. 48, p. 503-561.
M. (1965).- Rapport sur l'étage Albien. In : Colloque sur le Crétacé inférieur (Lyon, septembre 1963).- Mémoires du Bureau de Recherches Géologiques et Minières, Orléans, n° 34, p. 313-317.
D.L. (1973).- Phylogeny of the late Cenomanian planktonic foraminifera Anaticinella multiloculata ().- Journal of Foraminiferal Research, Washington D.C., vol. 2, p. 184-190.
A.S., W.J., J.A., M. & H.E. (1996).- The Late Albian to Early Cenomanian succession at Mont Risou near Rosans (Drôme, SE France): an integrated study (ammonites, inoceramids, planktonic foraminifera, nannofossils, oxygen and carbon isotopes).- Cretaceous Research, Amsterdam, vol. 17, n° 5, p. 515-606.
J.M., D. & F. (2007).- The rotaliporids, a polyphyletic group of Albian-Cenomanian planktonic foraminifera. Emendation of genera.- Journal of Foraminiferal Research, Washington D.C., vol. 37, n° 2, p. 175-186.
W.J., A.S., J.A. & M. (2004).- The GSSP for the base of the Cenomanian Stage, Mont Risou, Hautes-Alpes.- Episodes, Beijing, vol. 27, p. 21-32.
V., M. & H.C. (2004).- Albian high-resolution biostratigraphy and isotope stratigraphy: the Coppa della Nuvola pelagic succession of the Gargano Promontory (Southern Italy).- Eclogae Geologicae Helvetiae, Basel, vol. 97, p. 77-92.
J.G., P.P. & F.M. (2004).- The Cretaceous Period. In: F.M. et alii (eds.), A geologic time scale.- Cambridge University Press, p. 344-383.
M.R. & B.T. (2006).- Biostratigraphy and taxonomy of Late Albian planktonic foraminifera from ODP Leg 171B (Western North Atlantic).- Journal of Foraminiferal Research, Washington D.C., vol. 36, p. 166-190.
E. (1868).- Notices géologiques et paléontologiques sur les Alpes Vaudoises et les régions environnantes. V : Complément de la faune de Cheville.- Bulletin de la Société vaudoise des Sciences naturelles, Lausanne, t. 9, n° 58, p. 389-482.
F. & M. (eds., 1979).- Atlas de foraminifères planctoniques du Crétacé moyen (mer Boréale et Téthys).- Cahiers de Micropaléontologie, Paris, vol. 1, première partie, 185 p. + vol. 2, deuxième partie, 181 p.
F., M., F., C., J., J.M., D. & S. (1994).- Le Cénomanien de la région de Kalaat Senan (Tunisie Centrale) : litho-biostratigraphie et interprétation séquentielle.- Revue de Paléobiologie, Genève, vol. 12, n° 2 (1993), p. 351-505.
F., F., J.M. & D. (soumis).- The Albian (Vraconnian)-Cenomanian boundary at the North and South margins of the Tethyan Ocean (SE France, Tunisia).- Bulletin de la Société Société géologique de France, Paris.
G. (1973).- Sur l'âge de la faune d'ammonites au Château près de St-Martin-en-Vercors (Drôme) et quelques considérations sur l'évolution des Turrilitidés et des Hoplitidés vracono-cénomaniens.- Géologie Alpine, Grenoble, t. 49, p. 119-129.
J. (1948).- Note sur les genres de foraminifères Rotalipora (1942) et Thalmanninella, famille des Globorotaliidés.- Revue de l'Institut français du Pétrole, Rueil-Malmaison, vol. III, n° 4, p. 95-103.
J. (1977).- Essai de zonation du Crétacé méditerranéen à l'aide des foraminifères planctoniques.- Géologie méditerranéenne, Marseille, t. 4, n° 2, p. 99-108.
Cliquer sur la miniature pour agrandir l'image.
Figure 1 : Évolution anagénétique de deux lignées de Thalmanninella dans l'Albien supérieur-Cénomanien, l'une à formes biconvexes à spiroconvexes, l'autre à formes ombilicoconvexes à cylindriques (selon et alii, 2007).

Cliquer sur la miniature pour agrandir l'image.
Figure 2 : La coupe SMA dans la région de Kalaat Senan (et les collectes significatives dans les coupes voisines SMX, HML et HMR) : distribution verticale des principales espèces de foraminifères planctoniques et d'ammonites.
Cliquer sur la miniature pour agrandir l'image.
Figure 3 : Principaux bioévénements autour de la limite Albien-Cénomanien dans les régions de Kalaat Senan et Jérissa (Tunisie Centrale).
Cliquer sur la miniature pour agrandir l'image.
Figure 4 : Comparaison des principaux bioévénements autour de la limite Albien-Cénomanien entre la marge nord de la Téthys (Mont Risou, SE France) et la marge sud (coupe KZ de Kef el Azreg, Tunisie Centrale).
Cliquer sur la miniature pour agrandir l'image.
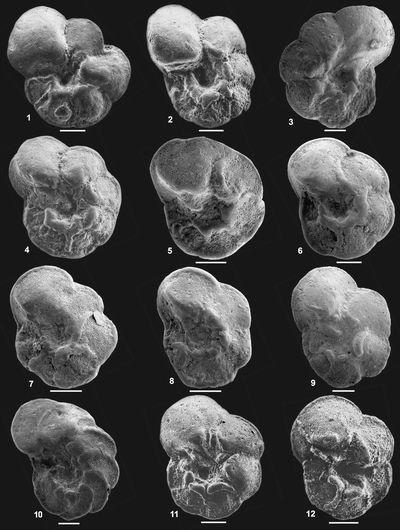
Planche I : Morphologie de l'espèce index Thalmanninella globotruncanoides () et de ses formes intermédiaires avec son ancêtre Th. caroni (). Échelle : barre = 0,5mm.
1-2 : formes intermédiaires entre Th. caroni et Th. globotruncanoides ; 3 : Th. globotruncanoides.
a : "anterior face" non différenciée ; b :bourrelet périombilical faiblement développé ; c : "anterior face" faiblement développée ; d : ligne d'inflexion entre l' "anterior face" et la face ombilicale ; e : "anterior face" ; f : bourrelet périombilical ; g : branche ombilicale de la carène ; h : suture ; i : loge recouvrant la branche ombilicale de la loge précédente.
Cliquer sur la miniature pour agrandir l'image.
Planche II : Les foraminifères planctoniques Thalmanninella caroni (), Th. globotruncanoides () et leurs intermédiaires dans les coupes SMA et HML, et Planomalina buxtorfi (). Échelle : barre = 0,1mm.
1 : Thalmanninella caroni, SMA 81.- 2 : Th. caroni, SMA 87.- 3 : Forme intermédiaire entre Th. caroni et Th. globotruncanoides ou Th. globotruncanoides : l' "anterior face" est développée, mais la branche ombilicale de la carène n'est pas différenciée ; SMA 87.- 4 : Th. globotruncanoides, SMA 92.- 5 : Th. globotruncanoides, SMA 93.- 6 : Th. globotruncanoides, SMA 93,5.- 7 : Th. globotruncanoides, SMA 93,5.- 8 : Th. globotruncanoides, SMA 97.- 9 : Th. globotruncanoides, SMA 97.- 10 : Planomalina buxtorfi, SMA 102.- 11 : Forme intermédiaire entre Th. caroni et Th. globotruncanoides : l' "anterior face" et la branche ombilicale de la carène ne sont pas différenciées ; HML 87.- 12 : Th. globotruncanoides, HML 87.

Cliquer sur la miniature pour agrandir l'image.
Planche III : a-d : Graysonites azregensis (, 1994), deux spécimens provenant du Cénomanien basal de Kef el Azreg au niveau KZ 20, Formation Fahdène, dans l'IZ à G. azregensis. Échelle : barre = 10cm.
a-b : coll. SG2, spécimen adulte où se développent de petites cornes sur la première moitié de la loge d'habitation ; c-d : coll. SG1, fragment phosphatisé sur lequel les longues côtes deviennent progressivement plus épaisses dans le dernier tour du phragmocône. SG : collection Stijn .
Cliquer sur la miniature pour agrandir l'image.
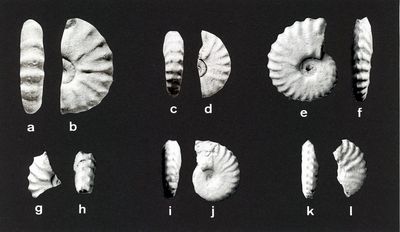
Planche IV : L'espèce index de la dernière zone d'ammonite de l'Albien supérieur (Vraconnien) dans la Téthys méridionale : a-l, Stoliczkaia (Shumarinaia) africana , 1907. Échelle : barre = 5cm.
a-b : lectotype, collection , de l'Albien supérieur du Djebel Mrhila en Tunisie Centrale, copie de & , 1994, p. 553, fig. 3 g-h ; c-d : paralectotype, collection , des mêmes localité et horizon que l'holotype, copie de & , 1994, p.553, fig. 3 v-w ; e-f : collection F. , de l'oued Smara, au niveau SMA 25-30, Formation Fahdène, Albien terminal (Vraconnien), dans la TRZ à Mortoniceras (Subschloenbachia) perinflatum ; g-h : coll. F. , de l'oued Hammadja, au niveau HMR 48, Formation Fahdène, dans la TRZ à M. (S.) perinflatum ; i-j : coll. F. , de l'oued Smara, au niveau SMA 94,50, Formation Fahdène, dans l'IZ à S. (Sh.) africana ; k-l : coll. F. , de l'oued Hammadja, au niveau HML 64, Formation Fahdène, dans l'IZ à St.(Sh). africana. La collection F. est maintenant conservée au Muséum d'Histoire naturelle de Lille (France).