![]()
Sommaire - Contents
[Abridged English version ![]() ]
]
[Introduction] [Upper Barremian
biozonation ...]
[Conclusions]
[Bibliographic references] and ... [Figures]
[Version française ![]() ]
]
[Introduction] [Cadre géologique et matériel]
[Méthode]
[Biozonation du Barrémien supérieur ...]
[Conclusions]
[Références bibliographiques] et ...[Figures]
Grand'rue, 04170 Saint André-les-Alpes (France) ; Biogéosciences, Université de
Bourgogne, CNRS, 6 bd Gabriel, 21000 Dijon (France)
Département des Sciences de la Terre, Université de Nice-Sophia-Antipolis, Faculté des Sciences, 28 avenue Valrose, 06108 Nice Cedex 2 (France)
Résidence le Villeneuve A, place du Grand Jardin, 06140 Vence (France)
Manuscrit en ligne depuis le 20 Mars 2008
![]()
Un nouveau découpage biozonal ammonitique du Barrémien supérieur du Sud-Est de la France est proposé. Il tient compte des renouvellements fauniques, des données séquentielles ainsi que des données historiques. Il est enrichi de quatre biohorizons nouveaux. Le Barrémien supérieur comporte à présent les trois biozones (anciennement six) à Vandenheckei, Sartousiana et Giraudi. La première contient les trois sous-zones à Uhligi, dont le statut est discuté, Sayni et Barremense, ainsi que les deux nouveaux biohorizons à Marchandi et Breistrofferi (sommet de la Sous-Zone à Barremense). La Biozone à Sartousiana admet les sous-zones à Limentinus, Provincialis et Feraudianus. Cette dernière sous-zone perd son statut de biozone par des arguments paléontologiques et stratigraphiques. Six biohorizons sont reconnus dans la Biozone à Sartousiana : Limentinus, Provincialis, Casanovai, Feraudianus, Magnini et Autrani. La Biozone à Giraudi comprend les sous-zones à Giraudi, Sarasini et Waagenoides, ainsi que trois biohorizons : Giraudi, Emerici et Puzosianum.
Crétacé inférieur téthysien ; biostratigraphie ; biozone d'intervalle ; renouvellements fauniques ; ammonites.
D., G. & S. (2008).- Nouveaux biohorizons et propositions pour le découpage biozonal ammonitique du Barrémien supérieur du Sud-Est de la France.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2008/03 (CG2008_A03)
New biohorizons and proposal for a revision of the ammonite biozonation of the Upper Barremian of southeastern France.- A new ammonite biozonal scheme is proposed for the Upper Barremian of southeastern France. It takes into account faunal changes, sequence stratigraphy and historical data. Four new biohorizons are proposed. At present, three (formerly six) are accepted for the Upper Barremian: the Vandenheckei, Sartousiana and Giraudi biozones. The first contains three subzones, the Uhligi (current status discussed), the Sayni and the Barremense subzones, and two new biohorizons, the Marchandi and Breistrofferi biohorizons (top of the Barremense Subzone). The Sartousiana Biozone now includes the Limentinus, Provincialis and Feraudianus subzones. This last one lost its status of biozone because of paleontologic and stratigraphic arguments. Six biohorizons are recognized in the Sartousiana Biozone: the Limentinus, Provincialis, Casanovai, Feraudianus, Magnini and Autrani biohorizons. The Giraudi Biozone includes the Giraudi, Sarasini and Waagenoides subzones and the Giraudi, Emerici and Puzosianum biohorizons.
Lower Tethyan Cretaceous; biostratigraphy; interval biozone; faunal turnover; ammonites.
The publications of (1990,
1997a, 1997b), et alii
(1995) and (1997), were reviewed by the IUGS Lower Cretaceous ammonite working group, the group ( & ,
1990; & ,
1993; & ,
1995; & ,
1999; & ,
2000; & ,
2003, and & ,
2006). The group recognizes 9 to 12 ammonites biozones in the Barremian stage of which 5 to 7 relate only to the upper part
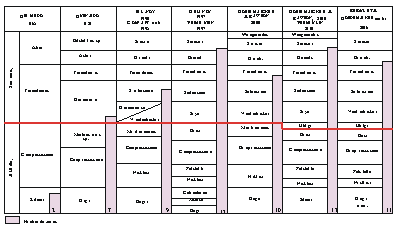
(Fig. 1 ![]() ). Unfortunately this complexity is accompanied by a great instability which has made the use of the Barremian biozonation and the placement of some biozone boundaries difficult and uncertain. Part of this is due mainly to a problem of conception and method in the definition and the use of "Zone" but also is caused in part by the as yet incomplete knowledge of of many ammonite groups.
). Unfortunately this complexity is accompanied by a great instability which has made the use of the Barremian biozonation and the placement of some biozone boundaries difficult and uncertain. Part of this is due mainly to a problem of conception and method in the definition and the use of "Zone" but also is caused in part by the as yet incomplete knowledge of of many ammonite groups.
In order to stabilize the zonation of the Upper Barremian of southeastern France, and thus to better its reproducibility and practical use, we propose to redefine the ammonitic biozonation
(Fig. 6 ![]() ). Insofar as possible we have chosen of to work with the great faunal changes
(Fig. 7
). Insofar as possible we have chosen of to work with the great faunal changes
(Fig. 7 ![]() ) while taking into account historical data (priority rules), in order to reduce to a maximum another upset of the zonal scheme. The preliminary results of this work were presented at a meeting of the French members of the Group in Digne ( et alii,
2007, unpublished).
) while taking into account historical data (priority rules), in order to reduce to a maximum another upset of the zonal scheme. The preliminary results of this work were presented at a meeting of the French members of the Group in Digne ( et alii,
2007, unpublished).
Author: biozone proposed by the Lower Cretaceous Cephalopods Working Group P.I.G.C. 262 during the Digne meeting ( & , 1990).
Index species: Toxancyloceras vandenheckei (, 1851).
Although further study is necessary, the base of the Uhligi Biozone is now considered as the lower limit of a subzone as advocated at the meeting of French members of the Group IUGS Lower Cretaceous Ammonites Working Group in
Digne
( et alii, 2007,
unpublished). The Vandenheckei Biozone is characterized by a diversification in and an important turnover of its ammonite fauna, mainly in the Ancyloceratoidea and Hemihoplitidae, with the appearance of the genera Toxancyloceras , 2003, Barrancyloceras , 2000, Gassendiceras , & ,
2006, and Camereiceras ,
1990
(Figs. 4 ![]() - 5
- 5 ![]() & 7
& 7 ![]() ). The Vandenheckei Biozone includes the Uhligi, the Sayni and the Barremense
subzones ( et alii,
2007, unpublished). The last named is now divided into two new horizons, the Breistrofferi and the Marchandi
biohorizons.
). The Vandenheckei Biozone includes the Uhligi, the Sayni and the Barremense
subzones ( et alii,
2007, unpublished). The last named is now divided into two new horizons, the Breistrofferi and the Marchandi
biohorizons.
Index species: Ancylezeiceras breistrofferi (, 1955): the index is well represented in the sections, along with the rare Gerhardtia galeatoides (, 1856) and with Gassendiceras quelquejeui , & ,
2006 (Fig. 4 ![]() ).
).
Index species: Camereiceras marchandi ( & , 2000). This species has been revised (, & ,
2006). In this horizon the last Holcodiscidae (Holcodiscus cf. uhligi , 1907) are
found (Fig. 4 ![]() ).
).
Author: biozone proposed by the Lower Cretaceous Cephalopods Working Group P.I.G.C. 262 during the Digne meeting ( & , 1990).
Index species: Gerhardtia sartousiana (d', 1841).
Because of the problems caused by anomalies in the first occurrence of Gerhardtia sartousiana (d', 1841) in the sections of the studied area, we propose to use the Limentinus Biohorizon as the base of the Sartousiana Biozone as well as the lower limit of the new Limentinus Subzone. But for nomenclatural stability we have chosen to retain the name Sartousiana Biozone for this biostratigraphical unit.
Such a choice has several advantages:
the appearance of Camereiceras limentinus (, 1979) is a step in the Tethyan evolutional development of the Camereiceras, its parent species being now well known (, & , 2006);
this index is present in abundance both in the basins and on the borders of the Vocontian platforms;
it is easy to identify, consequently more useful for all;
its appearance seems to be synchronous in all the sections that we have studied in Southeast France.
In the stratotypical area it occurs in a relatively thin succession of thick, resistant chert-bearing beds, quite visible in the
topography (Fig. 4 ![]() ), but they are not present in the Angles section.
), but they are not present in the Angles section.
Author: because of the principle of coordination, the Limentinus Subzone must be attributed to (1979), author of the Limentinus Biohorizon.
Index species: Camereiceras limentinus (, 1979).
Subdivisions: the Limentinus Subzone includes the Limentinus Biohorizon (, 1994, 1997a, 1997b) in which can be found Peirescites gygii , & , 2006, Macroscaphites ectotuberculatus , 1984, Costidiscus aff. tardus , 1984, Phyllopachyceras vergonsense & , 1995, Kotetishvilia brevicostata (, 1980), Gerhardtia galeatoides (, 1856), etc.
Author: proposed for the first time by in 1997.
Index species: Gerhardtia provincialis (d', 1850).
Subdivisions: the Provincialis Subzone includes the Provincialis Biohorizon and the new Casanovai Biohorizon in its upper part.
Author: proposed by in 1997.
Index species: Gerhardtia provincialis (d', 1850).
This biohorizon begins with the appearance of Gerhardtia provincialis (d', 1850), followed by its acmeic phase. In the Hemihoplitinae, Hemihoplites cornagoae , & , 2006, and Pachyhemihoplites gerthi (, 1955) are well represented.
Index species: Hemihoplites casanovai , 1992 (= Hemihoplites intermedius , 2003). This species precedes Hemihoplites feraudianus (d', 1841) and occurs in all the meter-sized chert-bearing beds of the stratotypical
area (Figs. 4 ![]() - 5
- 5 ![]() ), but it doesn't exist in the Angles section.
), but it doesn't exist in the Angles section.
Author: this subzone was proposed first by (1965).
Index species: Hemihoplites feraudianus (d', 1841).
The distinction between the Sartousiana and Feraudianus biozones was founded on an "important faunal
turnover" i.e. the disappearance of Pulchellidae and the appearance of the genus Hemihoplites. New data show that Hemihoplites appears in the Sartousiana Biozone and that rare specimens of Pulchellidae (Gerhardtia aff. provincialis (d', 1850) subsist in the Feraudianus
Biohorizon
(Figs. 4 ![]() - 5
- 5 ![]() ). So it appears that the distinction between these two biozones is not adequate for defining a zonal boundary. So we propose to restrict the Feraudianus Biozone as an upper subzone of the Sartousiana Biozone.
). So it appears that the distinction between these two biozones is not adequate for defining a zonal boundary. So we propose to restrict the Feraudianus Biozone as an upper subzone of the Sartousiana Biozone.
Subdivisions: the Feraudianus Subzone includes the Feraudianus and the Magnini biohorizons created by (1997a, 1997b), and the new Autrani Biohorizon.
Author: proposed first by as a subzone (1965).
Index species: Hemihoplites feraudianus (d', 1841).
The fauna of this biohorizon is well known (see , 1997a, p. 205-206). The index species is very abundant at the base accompanied by the rare Spinocrioceras polyspinosum , 1973, and Pseudoshasticrioceras quereilhaci , & , 2006. Gassendiceras enayi , & , 2006, is present at the top of the biohorizon.
Author: proposed by in 1997a-1997b.
Index species: Pseudoshasticrioceras magnini (, 1992).
This biohorizon is defined by the appearance of its index. Gassendiceras enayi , & , 2006, is always present.
Index-species: Pseudoshasticrioceras autrani , 1999. In this biohorizon appear Heteroceras coulleti , 1994, and Heteroceras baylei , 1876. In the stratotypical area this biohorizon is confined to the last thick beds of the Feraudianus Subzone.
Author: (1955) is the author of a biozone with Heteroceras leenhardtii and Colchidites (Imerites) giraudi in the southern (former) USSR, which later became the Giraudi Biozone [see historic in (1989) and (1997a, 1997b)].
Index species: Imerites giraudi (d', 1841).
This biozone is characterized by a rise of sea level (,
2005) associated with an important faunal
turnover
(Fig. 7 ![]() ), i.e. a complete absence of the Pulchelliidae and Hemihoplitinae; proliferation of the Heteroceratidae with the genera Heteroceras d', 1850 (predominant), Rouchadzeites & , 2006, Egoianites & ,
2006, and Argvethites , 1933. The Gassendiceratinae , & ,
2006, remain well reppresented with the genus Imerites (, 1933)
(Fig. 4
), i.e. a complete absence of the Pulchelliidae and Hemihoplitinae; proliferation of the Heteroceratidae with the genera Heteroceras d', 1850 (predominant), Rouchadzeites & , 2006, Egoianites & ,
2006, and Argvethites , 1933. The Gassendiceratinae , & ,
2006, remain well reppresented with the genus Imerites (, 1933)
(Fig. 4 ![]() ). Recently collected specimens of Imerites show clearly a direct link between the last Pseudoshasticrioceras and the first Imerites. The Peirescinae , & ,
2006, are always represented by the rare genus Spinocrioceras , 1973.
). Recently collected specimens of Imerites show clearly a direct link between the last Pseudoshasticrioceras and the first Imerites. The Peirescinae , & ,
2006, are always represented by the rare genus Spinocrioceras , 1973.
In previous papers (, 1994,
1997a, 1997b; synthesis of & ,
2006), the Giraudi Biozone was followed by the Sarasini Biozone which corresponded to the stratigraphical appearance of the genus Martelites , 1989. New discoveries in several sections of the stratotypical area
(Fig. 5 ![]() ) demonstrate the presence of Martelites sp. and Martelites aff. gamkrelidzei , 1933 in the Giraudi Biozone within the Emerici Biohorizon. In addition, Heteroceras is always represented in the Sarasini Biozone by species like Heteroceras baylei , 1876 and Heteroceras fuhrae , 1997. So the boundary between the Giraudi and Sarasini
biozones (sensu ,
1997a, 1997b) is very permeable, and their distinction, based on a faunal
turnover, seems to be invalid. So we revert to the observations of
(1983, 1989) which insisted on the faunal affinities between the Giraudi and Securiformis
biozones of Georgia. For that reason we prefer to consider the Sarasini Biozone as a
subzone of the Giraudi Biozone
(Fig. 6
) demonstrate the presence of Martelites sp. and Martelites aff. gamkrelidzei , 1933 in the Giraudi Biozone within the Emerici Biohorizon. In addition, Heteroceras is always represented in the Sarasini Biozone by species like Heteroceras baylei , 1876 and Heteroceras fuhrae , 1997. So the boundary between the Giraudi and Sarasini
biozones (sensu ,
1997a, 1997b) is very permeable, and their distinction, based on a faunal
turnover, seems to be invalid. So we revert to the observations of
(1983, 1989) which insisted on the faunal affinities between the Giraudi and Securiformis
biozones of Georgia. For that reason we prefer to consider the Sarasini Biozone as a
subzone of the Giraudi Biozone
(Fig. 6 ![]() ).
).
Subdivisions: the Giraudi Biozone now includes the Giraudi Subzone, the Sarasini Subzone and the Waagenoides Subzone.
Author: proposed as a subzone by (1983) for the southeastern (former) USSR.
Index species: Imerites giraudi (, 1888).
Subdivisions: includes the Giraudi Biohorizon founded on Imerites giraudi (, 1888) and the Emerici Biohorizon founded on Heteroceras emerici d', 1842.
Author: proposed by (1997a, 1997b).
Index species: Imerites giraudi (, 1888).
In addition to the very variable index-species, the most representative ammonites are Heteroceras coulleti , 1994, and Heteroceras baylei , 1876, along with some rare taxa such as Heteroceras rawsonii & , 2006, and Spinocrioceras amadei (, 1883) of the Peirescinae , & , 2006.
Author: proposed by in 1997a-1997b.
Index species: Heteroceras emerici d', 1842.
This biohorizon is defined by the explosion of the Heteroceratidae with the group: Heteroceras emerici d', 1842, Heteroceras moriezense , 1997 and Heteroceras baylei , 1876 and the rarer Egoianites & , 2006 and Rouchadzeites & , 2006. The first Martelites (Martelites aff. gamkrelidzei , 1933) occur in this biohorizon. The Peirescinae , & , 2006, are present with Spinocrioceras trachyomphalus (, 1883).
Author: proposed first by (1990) as a biozone.
Index species: Martelites sarasini (, 1933).
Subdivisions: including the Puzosianum Biohorizon founded on Anglesites puzosianum (d', 1842).
Author: introduced first by (1965) as a subzone.
Index species: Anglesites puzosianum (d', 1842) recently revised ( & , 2007).
The base of the horizon is defined by the occurrence of numerous Anglesites puzosianum (d', 1842) accompanied by Martelites sarasini (, 1933) and Heteroceras baylei , 1876.
Authors: proposed as a biohorizon by & in 1995.
Index species: Pseudocrioceras waagenoides (, 1938).
This subzone is temporarily retained but we are disposed to consider it as a horizon developed only in shallow environments and practically unrecognizable in basin facies. This subzone is characterized by a proliferation of the genus Pseudocrioceras (1924) cohabiting with some Kutatissites and the last representatives of the Martelites.
The new zonal scheme proposed in this paper is founded on the great faunal turnover the Upper Barremian of southeastern France
(Fig. 7 ![]() ), sometimes correlated with eustatism. The extension of this biozonation to the whole Tethyan realm should help correlation between far-distant sites, even if the pertinent index species is absent. The criteria involved can be summarized as follow:
), sometimes correlated with eustatism. The extension of this biozonation to the whole Tethyan realm should help correlation between far-distant sites, even if the pertinent index species is absent. The criteria involved can be summarized as follow:
Vandenheckei Biozone: turnover and diversification the Ancyloceratoidea;
Sartousiana Biozone: great diversification of the Hemihoplitidae to the detriment of the Pulchellidae especially in the upper part of the biozone after the acme of the genus Gerhardtia;
Giraudi Biozone: Hemihoplitinae and Pulchellidae gone, strong diversification of the Heteroceratidae more especially in the lower part of the biozone, and the appearance and local proliferation of the genera Kutatissites and Pseudocrioceras in the upper part of the biozone.
The continuation of a revision of the biostratigraphy of the rest of the Barremian stage, should in the future result in better-defined biostratigraphical subdivisions of this Cretaceous stage founded on stable, reproducible and coherent biozones.
Suite aux travaux de (1990,
1997a, 1997b), et alii
(1995) et (1997), repris lors des discussions des différentes réunions
de l'IUGS Lower Cretaceous ammonite working group, the group,
à Digne ( & ,
1990), à Mula ( & ,
1993), à Piobbico ( & ,
1995), à Londres ( & ,
1999), à Vienne ( & ,
2000), à Lyon ( & ,
2003) et dernièrement à Neuchâtel ( & ,
2006), l'étage Barrémien comprenait 9 à 12 biozones d'ammonites dont 5 à 7 pour la seule partie supérieure de cet étage
(voir Figure 1 ![]() pour un rappel historique). Malheureusement cette inflation zonale s'accompagne aussi d'une grande instabilité qui rend délicate l'utilisation de la biozonation barrémienne. Parmi les différentes causes de cette instabilité nous pouvons déplorer un problème de concept et de méthode dans la définition et l'utilisation de la "zone". Mais aussi la connaissance encore très partielle de nombreux groupes d'ammonites et de leur évolution, voire des conceptions très divergentes de la notion d'espèce et de la définition des taxons suivant les
auteurs. D'autre part l'utilisation quasi exclusive de la seule coupe du stratotype historique de la route d'Angles
(Alpes-de-Haute-Provence) dans l'établissement du schéma zonal actuel pose problème, puisque cette coupe présente d'importants manques dans les biozones à Vandenheckei et Sartousiana telles qu'admises dans ce travail. Tout cela fait que les limites de certaines biozones sont quelques fois mal caractérisées
paléontologiquement ou, en tout cas, ne correspondent pas aux événements majeurs
(renouvellements fauniques, variations du niveau marin,
etc.) que l'on est en droit d'attendre pour de telles divisions.
pour un rappel historique). Malheureusement cette inflation zonale s'accompagne aussi d'une grande instabilité qui rend délicate l'utilisation de la biozonation barrémienne. Parmi les différentes causes de cette instabilité nous pouvons déplorer un problème de concept et de méthode dans la définition et l'utilisation de la "zone". Mais aussi la connaissance encore très partielle de nombreux groupes d'ammonites et de leur évolution, voire des conceptions très divergentes de la notion d'espèce et de la définition des taxons suivant les
auteurs. D'autre part l'utilisation quasi exclusive de la seule coupe du stratotype historique de la route d'Angles
(Alpes-de-Haute-Provence) dans l'établissement du schéma zonal actuel pose problème, puisque cette coupe présente d'importants manques dans les biozones à Vandenheckei et Sartousiana telles qu'admises dans ce travail. Tout cela fait que les limites de certaines biozones sont quelques fois mal caractérisées
paléontologiquement ou, en tout cas, ne correspondent pas aux événements majeurs
(renouvellements fauniques, variations du niveau marin,
etc.) que l'on est en droit d'attendre pour de telles divisions.
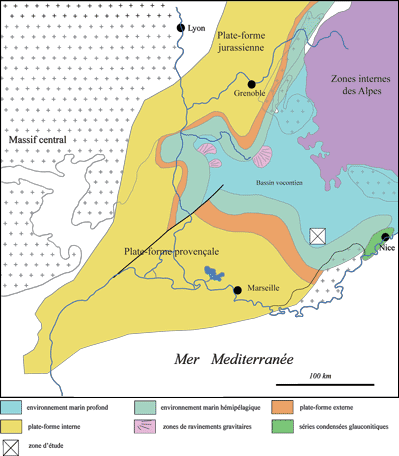
Le Sud-Est de la France est marqué au Crétacé inférieur par l'évolution d'un bassin intracratonique connu sous le nom de bassin vocontien (,
1900)
(Figs. 2 ![]() - 3
- 3 ![]() ). Sa partie
sud, dans la région du stratotype historique du Barrémien (région
d'Angles-Barrême-Castellane), est caractérisée par une sédimentation de type pélagique, généralement constituée par des alternances de marnes et de calcaires en bancs décimétriques à métriques. Cette région particulière est très favorable aux études biostratigraphiques car elle est peu affectée par les remaniements gravitaires et les effets de l'orogenèse alpine. Devant la continuité des dépôts et de l'enregistrement paléontologique, il est possible d'y suivre avec une bonne précision la succession et l'évolution des faunes
d'ammonites.
). Sa partie
sud, dans la région du stratotype historique du Barrémien (région
d'Angles-Barrême-Castellane), est caractérisée par une sédimentation de type pélagique, généralement constituée par des alternances de marnes et de calcaires en bancs décimétriques à métriques. Cette région particulière est très favorable aux études biostratigraphiques car elle est peu affectée par les remaniements gravitaires et les effets de l'orogenèse alpine. Devant la continuité des dépôts et de l'enregistrement paléontologique, il est possible d'y suivre avec une bonne précision la succession et l'évolution des faunes
d'ammonites.
Le matériel utilisé pour la réalisation de ce travail est le résultat de l'étude systématique et
rigoureuse, banc par banc, de nombreuses coupes dans la région du stratotype historique du Barrémien et des bordures de
plates-formes voisines, qui seront prochainement publiées, ainsi
que de la synthèse des données bibliographiques disponibles (,
1994,
1997a, 1997b ; ,
2002, 2005). Nous avons choisi de présenter
(Figs. 4 ![]() - 5
- 5 ![]() )
la succession des coupes de la région de Méouilles (Saint-André-les-Alpes,
Alpes-de-Haute-Provence) qui est la plus complète du Barrémien supérieur dans le secteur étudié. Les familles d'ammonites principalement étudiées pour ce schéma zonal sont celles qui présentent le plus d'intérêt biostratigraphique à ce niveau par leur évolution rapide : les
Hemihoplitidae, les Heteroceratidae, les Pulchelliidae, et les Ancyloceratidae
(Fig. 7
)
la succession des coupes de la région de Méouilles (Saint-André-les-Alpes,
Alpes-de-Haute-Provence) qui est la plus complète du Barrémien supérieur dans le secteur étudié. Les familles d'ammonites principalement étudiées pour ce schéma zonal sont celles qui présentent le plus d'intérêt biostratigraphique à ce niveau par leur évolution rapide : les
Hemihoplitidae, les Heteroceratidae, les Pulchelliidae, et les Ancyloceratidae
(Fig. 7 ![]() ).
).
D'une manière très générale, une biozone est reconnaissable par son contenu paléontologique. Il ne faut toutefois pas oublier que ce contenu est tributaire des enregistrements sédimentaires et biologiques qui peuvent être discontinus tant d'un point de vue géographique que temporel. Ainsi la durée d'existence réelle des espèces fossiles n'est jamais connue en totalité (, 1997, Fig. 49) et peut subir localement d'importantes fluctuations en fonction de nombreux paramètres paléogéographiques et paléobiogéographiques. Ainsi, en dehors de toute tentative de corrélation, la reconnaissance d'une biozone est nécessairement limitée et dépend de la répartition géographique, écologique et stratigraphique de l'index qui lui sert de référence. À la suite de & (1999) puis de & (2003) pour le Valanginien, nous avons choisi dans ce travail de définir des biozones d'intervalles au détriment, par exemple, des biozones de distribution d'un taxon. Les limites de ces dernières sont en effet trop susceptibles d'être continuellement sujettes aux modifications que pourraient leur imposer les découvertes et les révisions systématiques de leurs taxons index, ce qui entraînerait une instabilité de la biozonation. La base d'une biozone d'intervalle est définie par celle du biohorizon (ou biorepère) sélectionné, et son sommet par la base de la biozone d'intervalle suivante (, 1997).
Un biohorizon correspond à un mince intervalle caractérisé par une association paléontologique particulière et au sein duquel aucune division plus fine n'est possible (, 1976 ; , 1997). Enfin la sous-zone est définie comme une partie caractéristique d'une biozone, voire comme un regroupement de biohorizons, utile pour exprimer des précisions biostratigraphiques (, 1997, p. 77).
Afin de stabiliser la biozonation du Barrémien supérieur du Sud-Est de la France, et pour permettre sa meilleure reproductibilité et son utilisation
pratique, nous nous proposons d'en rediscuter puis d'en redéfinir les biozones ammonitiques
(Fig. 6 ![]() ). Nous avons adopté une méthode de construction du schéma zonal qui se fonde autant que possible sur les grands renouvellements
fauniques, tout en tenant compte des données historiques (règles de priorité), pour limiter au maximum un nouveau bouleversement du schéma zonal. Les résultats préliminaires de ce travail ont été présentés à la Réunion de Digne des membres français du Group
( et alii,
2007, inédit).
). Nous avons adopté une méthode de construction du schéma zonal qui se fonde autant que possible sur les grands renouvellements
fauniques, tout en tenant compte des données historiques (règles de priorité), pour limiter au maximum un nouveau bouleversement du schéma zonal. Les résultats préliminaires de ce travail ont été présentés à la Réunion de Digne des membres français du Group
( et alii,
2007, inédit).
Auteur : biozone proposée par le Working Group Cephalopods du Crétacé inférieur du P.I.G.C. 262 à Digne ( & , 1990).
Espèce index : Toxancyloceras vandenheckei (, 1851).
Statut : en l'attente de nouvelles données, la limite inférieure et les subdivisions de la
Biozone à Vandenhekei sont celles proposées lors de la réunion des biostratigraphes français du Group IUGS Lower Cretaceous Ammonites Working Group à Digne-les-Bains ( et alii,
2007, inédit). Nous préconisons ainsi d'éviter l'utilisation de la
"Biozone" à Uhligi dans la mesure où son espèce index n'a pas encore fait l'objet d'une
révision. Cette forme reste mal connue tant paléontologiquement que
stratigraphiquement, et des exemplaires affines ont été récoltés au sommet de la
Biozone à Vandenheckei. Enfin, cette subdivision est trop brève pour être élevée au rang de biozone. En accord avec nos collègues français du Group IUGS Lower Cretaceous Ammonites Working Group ( et alii,
2007, inédit), nous plaçons provisoirement la "Biozone" à Uhligi en tant que
sous-zone à la base de la Biozone à Vandenheckei que nous considérons donc comme l'unité basale du Barrémien supérieur
(Fig. 6 ![]() ).
Les apparitions de Toxancyloceras vandenheckei (, 1851) et d'Holcodiscus uhligi , 1907, semblent presque simultanées ( et alii,
2007, inédit ; , 2005). Cette position a également l'avantage de rapprocher la base de la
Biozone à Vandenheckei de la limite historique (, 1965) du Barrémien
inférieur/supérieur, ainsi que d'une limite séquentielle majeure (,
2005).
).
Les apparitions de Toxancyloceras vandenheckei (, 1851) et d'Holcodiscus uhligi , 1907, semblent presque simultanées ( et alii,
2007, inédit ; , 2005). Cette position a également l'avantage de rapprocher la base de la
Biozone à Vandenheckei de la limite historique (, 1965) du Barrémien
inférieur/supérieur, ainsi que d'une limite séquentielle majeure (,
2005).
La Sous-Zone à Barremense est maintenue dans l'attente d'une véritable révision de l'espèce index qui fait l'objet d'interprétations très contradictoires suivant les auteurs ( et alii,
1995,
2008 ; , 2005), bien que ceux-ci semblent paradoxalement s'accorder sur sa position
stratigraphique. Le foisonnement des Barrancyloceras et des formes
apparentées, le plus souvent connues d'après des échantillons très
incomplets, dans la partie supérieure de la zone à Vandenheckei
(Fig. 7 ![]() )
est sans aucun doute à l'origine de ces divergences.
)
est sans aucun doute à l'origine de ces divergences.
D'autre part, une meilleure connaissance des faunes d'ammonites des niveaux supérieurs de la
Biozone à Vandenheckei
(Figs. 4 ![]() - 5
- 5 ![]() ), telle qu'admise dans ce travail,
nous permet de proposer à son sommet deux nouveaux biohorizons ammonitiques très bien
caractérisés. Le premier est basé sur l'utilisation d'Ancylezeiceras breistrofferi (, 1955) dont la position stratigraphique est à présent bien établie
(ce travail
: Fig. 4
), telle qu'admise dans ce travail,
nous permet de proposer à son sommet deux nouveaux biohorizons ammonitiques très bien
caractérisés. Le premier est basé sur l'utilisation d'Ancylezeiceras breistrofferi (, 1955) dont la position stratigraphique est à présent bien établie
(ce travail
: Fig. 4 ![]() ). Le second est basé sur l'utilisation de l'espèce index Camereiceras marchandi ( & , 2000) récemment révisée ( et alii,
2006).
). Le second est basé sur l'utilisation de l'espèce index Camereiceras marchandi ( & , 2000) récemment révisée ( et alii,
2006).
Assemblage faunique : la Biozone à Vandenheckei se caractérise essentiellement par une diversification et un renouvellement faunique
importants, en particulier des Ancyloceratidae et des Hemihoplitidae
(Fig. 7 ![]() ). On observe, entre
autres, l'apparition des genres Toxancyloceras , 2003,
Barrancyloceras , 2000, Gassendiceras , & ,
2006, et Camereiceras ,
1990.
). On observe, entre
autres, l'apparition des genres Toxancyloceras , 2003,
Barrancyloceras , 2000, Gassendiceras , & ,
2006, et Camereiceras ,
1990.
Espèce index : Ancylezeiceras breistrofferi (, 1955).
Statut : ce biohorizon est défini par l'apparition de son espèce index, et sa limite supérieur est actuellement fixée à la base du Biohorizon à Marchandi.
Assemblages fauniques : Ancylezeiceras breistrofferi (, 1955) est en général bien présent dans les coupes à défaut
d'être très
abondant, et associé aux premières Gerhardtia galeatoides (, 1856)
extrêmement rares
(Fig. 4 ![]() )
ainsi qu'à des Camereiceras sp., Gassendiceras quelquejeui , & ,
2006, Acantholytoceras gr. tenuiscostatum (, 1964) et des Rugacrioceras gr. monopujae (, 1955).
)
ainsi qu'à des Camereiceras sp., Gassendiceras quelquejeui , & ,
2006, Acantholytoceras gr. tenuiscostatum (, 1964) et des Rugacrioceras gr. monopujae (, 1955).
Espèce index : Camereiceras marchandi ( & , 2000).
Statut : ce biohorizon est défini par l'apparition de son espèce index, et sa limite supérieur est fixée à la base du Biohorizon à Limentinus.
Assemblage faunique : Camereiceras marchandi (, 2000)
(microconque et macroconque) est en général assez bien représenté ; les derniers Holcodiscidae (H.
cf. uhligi , 1907), qui disparaissent dans cet horizon, sont en revanche
extrêmement rares
(Fig. 4 ![]() ).
).
Auteur : zone proposée par le Working Group Cephalopods du Crétacé inférieur du P.I.G.C. 262 à Digne ( & , 1990).
Espèce index : Gerhardtia sartousiana (d', 1841) sensu , 2002.
Statut : bien que la base de cette biozone soit mal caractérisée par son espèce index, il nous parait néanmoins nécessaire de conserver cette unité biostratigraphique dans un
souci de
stabilité. En effet, en suivant les recommandations du Comité Français de Stratigraphie (,
1997), ainsi que celles du Group ( et alii,
2007, inédit), il n'est pas nécessaire qu'une biozone soit forcément concomitante de la chronozone
homonyme qui est, dans ce cas, une biozone d'intervalle. En pratique, on désigne souvent une zone d'intervalle par le nom d'un taxon unique bien
représenté, mais pas nécessairement restreint à cette zone,
même si les limites de la zone sont choisies sur la base de l'apparition ou de la disparition d'un autre taxon (,
1997, p. 71-72). Dans le cas présent, la
Biozone à Sartousiana débute par le Biohorizon à
Limentinus. On y observe la forte diversification des Hemihoplitidae, ainsi que la raréfaction progressive des Pulcheliidae
(Figs. 4 ![]() et 7
et 7 ![]() ).
).
Auteur : par principe de coordination, la Sous-Zone à Limentinus doit être attribuée à (1979), auteur d'un Biohorizon à Limentinus.
Espèce index : Camereiceras limentinus (, 1979).
Statut : l'espèce Gerhardtia sartousiana (d', 1841) pose actuellement certains problèmes d'utilisations en tant qu'index pour caractériser la limite inférieure de sa biozone homonyme. En effet, outre son relatif manque d'abondance dans la plupart des coupes, son apparition stratigraphique dans le bassin vocontien n'a encore jamais été très clairement établie (, 2002). C'est pourquoi nous proposons l'utilisation de la Sous-Zone à Limentinus basée sur l'espèce index Camereiceras limentinus (, 1979) pour caractériser la partie inférieure de la Biozone à Sartousiana. Un tel choix présente plusieurs avantages :
l'apparition de cet index s'effectue dans le cadre évolutif des Camereiceras , 1992, dont l'espèce mère, Camereiceras marchandi ( & , 2000), vient d'être révisée ( et alii, 2006) et est à présent mieux connue ;
Camereiceras limentinus (, 1979) est aussi bien présent en abondance tant dans le bassin que sur ses bordures de plates-formes ;
d'utilisation aisée, cette espèce est très facilement identifiable, même par des non-spécialistes, et
son apparition est synchrone dans toutes les coupes que nous avons étudiées dans le Sud-Est de la France.
Enfin, dans la région stratotypique le Biohorizon à Limentinus correspond à un bon repère lithologique formé d'une petite succession de gros bancs à silex bien visibles dans la topographie
(Fig. 4 ![]() ),
bien qu'inexistants dans la coupe du stratotype historique d'Angles
(lacune probable).
),
bien qu'inexistants dans la coupe du stratotype historique d'Angles
(lacune probable).
Auteur : ce biohorizon a été proposé pour la première fois par en 1979.
Espèce index : Camereiceras limentinus (, 1979).
Statut : établi initialement dans la Biozone à Feraudianus (,
1979), le
Biohorizon à Limentinus a été situé ensuite au sein de la Biozone à Sartousiana (,
1997a, 1997b), puis au sommet de la
Biozone à Sayni (=
Vandenheckei) par (2002). Il caractérise à présent la base de la
Biozone à Sartousiana sensu et alii
(ce travail : Fig. 6 ![]() ).
).
Assemblages fauniques : l'espèce index présente un développement quantitatif important en comparaison avec les autres faunes d'ammonites des
mêmes niveaux. Bien que déjà très épisodiquement présentes, c'est dans le Biohorizon à Limentinus que commencent vraiment à s'implanter les Gerhardtia avec Gerhardtia galeatoides (, 1856). Cette dernière se différenciera avant la
sous-zone suivante en la chrono-espèce Gerhardtia sartousiana (d', 1841). D'autres formes sont également intéressantes à signaler
(Fig. 4 ![]() ), en particulier Peirescites gygii , & ,
2006, Macroscaphites ectotuberculatus , 1984, Costidiscus
aff. tardus , 1984, Phyllopachyceras vergonsense & , 1995, etc. (,
1994,
1997a, 1997b), ainsi que les derniers représentants de Kotetishvilia brevicostata (, 1980).
), en particulier Peirescites gygii , & ,
2006, Macroscaphites ectotuberculatus , 1984, Costidiscus
aff. tardus , 1984, Phyllopachyceras vergonsense & , 1995, etc. (,
1994,
1997a, 1997b), ainsi que les derniers représentants de Kotetishvilia brevicostata (, 1980).
Auteur : cette sous-zone a été proposée pour la première fois par en 1997.
Espèce index : Gerhardtia provincialis (d', 1850).
Statut : cette sous-zone débute avec le Biohorizon à
Provincialis, défini par l'apparition de Gerhardtia provincialis (d', 1850), et est essentiellement caractérisée par l'important développement des Hemihoplitinae
(Fig. 7 ![]() ).
Une fois le
Biohorizon basal à Provincialis passé, on observe une raréfaction progressive des
Pulchelliidae. Par ailleurs, de nouvelles récoltes au sommet de la Sous-Zone à Provincialis permettent de préciser la position stratigraphique d'Hemihoplites casanovai , 1992
(Figs. 4
).
Une fois le
Biohorizon basal à Provincialis passé, on observe une raréfaction progressive des
Pulchelliidae. Par ailleurs, de nouvelles récoltes au sommet de la Sous-Zone à Provincialis permettent de préciser la position stratigraphique d'Hemihoplites casanovai , 1992
(Figs. 4 ![]() - 5
- 5 ![]() ). Son abondance nous fait proposer l'utilisation d'un nouveau
Biohorizon à
Casanovai.
). Son abondance nous fait proposer l'utilisation d'un nouveau
Biohorizon à
Casanovai.
Auteur : ce biohorizon a été proposé par en 1997.
Espèce index : Gerhardtia provincialis (d', 1850).
Assemblages fauniques : ce biohorizon débute avec l'apparition de Gerhardtia provincialis
(d', 1850), rapidement suivie par sa pullulation
brutale, et se termine actuellement par la fin de sa phase acméique. Gerhardtia provincialis
(d', 1850) est issue par un processus anagénétique de Gerhardtia sartousiana
(d', 1841) dont elle est principalement différenciée par l'apparition d'un sillon ventral. Chez les
Hemihoplitinae, Hemihoplites cornagoae , & ,
2006, ainsi que Pachyhemihoplites gerthi (, 1955) sont bien représentés
(Fig. 4 ![]() ).
).
Espèce index : Hemihoplites casanovai , 1992 (= Hemihoplites intermedius , 2003). De nombreux spécimens de cette espèce ont été récoltés dans plusieurs coupes du Sud-Est de la France (bassin vocontien et bordures de ses plates-formes), où elle est assez fréquente.
Assemblages fauniques : Hemihoplites casanovai , 1992, est caractéristique des niveaux antérieurs à l'apparition d'Hemihoplites feraudianus
(d', 1841). Sur le plan lithologique ces niveaux sont principalement
représentés, dans la région stratotypique, par un banc à silex d'épaisseur métrique
(Figs. 4 ![]() - 5
- 5 ![]() )
inexistant dans la coupe du stratotype historique. La présence des Gerhardtia
est devenue anecdotique, tandis que se mettent en place les faunes déjà décrites et reconnues dans le
Biohorizon à Feraudianus (,
1994,
1997a, 1997b).
)
inexistant dans la coupe du stratotype historique. La présence des Gerhardtia
est devenue anecdotique, tandis que se mettent en place les faunes déjà décrites et reconnues dans le
Biohorizon à Feraudianus (,
1994,
1997a, 1997b).
Auteur : cette sous-zone a pour la première fois été proposée par lors du Colloque sur le Crétacé inférieur (1965).
Espèce index : Hemihoplites feraudianus (d', 1841).
Statut : au Barrémien supérieur téthysien, la limite entre les biozones à Sartousiana et à Feraudianus sensu & , 2006, était habituellement argumentée par les auteurs sur la base d'un "important renouvellement faunique". On y constate l'explosion quantitative des Hemihoplitinae et en particulier d'Hemihoplites feraudianus (d', 1841) qui caractérise très bien la base de sa biozone (ici sous-zone), alors que la disparition des Pulchelliidae était reconnue pour être effective dès la partie supérieure de la Sous-Zone à Provincialis. Cependant, de nouvelles observations viennent nuancer ces données.
D'une part, les Pulchelliidae disparaissent en réalité de manière plus progressive et plus
tardive. En effet, en 1995, & ont signalé la présence du genre Heinzia (sic) dans la
Biozone à Giraudi (plus vraisemblablement Sous-Zone à Feraudianus) de la coupe du Rio Argos (Caravaca, SE Espagne),.
Cette observation a été reprise sans argumentation nouvelle par
(2004, p. 10).
Depuis, et alii (2004) ont signalé en Espagne
(région d'Alicante), la présence de spécimens du genre Heinzia (= Gerhardtia) dans des niveaux qu'ils ont datés de la
Biozone à
Feraudianus. En parallèle, nos propres recherches dans le bassin Vocontien (SE de la France) nous ont permis de récolter plusieurs exemplaires de Gerhardtia
aff. provincialis (d', 1850) dans des niveaux très clairement datés du
Biohorizon à Feraudianus
(Figs. 4 ![]() - 5
- 5 ![]() ).
).
D'autre part, l'explosion quantitative des Hemihoplitinae à la base du Biohorizon à Feraudianus est nuancée par la présence abondante d'autres espèces d'Hemihoplites dans les niveaux plus anciens. En effet, Hemihoplites casanovai , 1992, est à présent connue au milieu de la Biozone à Sartousiana (Sous-Zone à Provincialis) sensu et alii (ce travail), ainsi qu'Hemihoplites cornagoae , & , 2006, dans le Biohorizon à Provincialis ( et alii, 2006).
Il apparaît donc que la limite jusqu'à présent admise entre les biozones à Sartousiana et à Feraudianus sensu & (2006) n'est en réalité pas pertinente :
en ce qui concerne les Pulchelliidae puisqu'ils subsistent au-delà de celle-ci avec Gerhardtia provincialis (d', 1850) et
pour les Hemihoplites sensu stricto, dont l'apparition est à présent connue dans la Biozone à Sartousiana.
S'il semble bien exister un renouvellement faunique à la limite inférieure de la
Sous-Zone à Feraudianus, celui-ci est majoritairement induit par la prolifération de son espèce index. L'interface de ces deux unités biostratigraphiques
ne nous paraît pas adéquate en terme de limite de zone. C'est
pourquoi, en prenant compte ces arguments paléontologiques et
biostratigraphiques, nous proposons de restreindre l'ancienne Biozone à Feraudianus en tant que
sous-zone au sommet de la Biozone à Sartousiana. Ainsi la nouvelle limite entre les biozones à Sartousiana sensu et alii
(ce travail) et à Giraudi sus-jacente serait effectivement caractérisée par un important renouvellement faunique
(Fig. 7 ![]() ). Elle aurait également l'avantage de correspondre à un changement séquentiel et lithologique important, puisque la base de la
Biozone à Giraudi correspond à une phase de transgression marine (,
2005,
Fig. 7).
). Elle aurait également l'avantage de correspondre à un changement séquentiel et lithologique important, puisque la base de la
Biozone à Giraudi correspond à une phase de transgression marine (,
2005,
Fig. 7).
La Sous-Zone à Feraudianus est définie par l'apparition de son espèce index (qui précède immédiatement sa phase acméique), et est caractérisée par la disparition des derniers Pulchelliidae, par la présence du genre Pseudoshasticrioceras , 1998, et la présence des premiers Heteroceras d', 1850 connus dans sa partie sommitale ( 1994, 1997a, 1997b ; & , 2006). Le genre Gassendiceras , & , 2006, est toujours présent avec l'espèce enayi , & , 2006.
Auteur : proposé pour la première fois en tant qu'horizon par (1994).
Espèce index : Hemihoplites feraudianus (d', 1841).
Assemblages fauniques : la faune de ce biohorizon a déjà été citée, et outre l'espèce index majoritaire, elle est composée par des formes classiques pour le Barrémien supérieur (voir , 1997a, p. 205-206), ainsi que par les plus rares Spinocrioceras polyspinosum , 1973, et Pseudoshasticrioceras quereilhaci , & , 2006. Gassendiceras enayi , & , 2006, est déjà présent à son sommet.
Auteur : proposé pour la première fois par en 1997a-1997b.
Espèce index : Pseudoshasticrioceras magnini (, 1992).
Assemblages fauniques : ce biohorizon est caractérisé à sa base par l'apparition de son index ; on observe également la présence de Gassendiceras enayi , & , 2006.
Espèce index : Pseudoshasticrioceras autrani , 1999.
Assemblages fauniques : ce biohorizon est toujours représenté dans le bassin vocontien par une série de gros bancs où les Heteroceras coulleti , 1994, et du groupe baylei , 1876 sont bien représentés.
Auteur : (1955) est l'auteur d'une Biozone à Heteroceras leenhardtii et Colchidites (Imerites) giraudi pour l'ex-URSS méridionale, qui deviendra ensuite la Biozone à Giraudi [voir historique dans (1989) et (1997a, 1997b)].
Espèce index : Imerites giraudi (d', 1841).
Statut et assemblages fauniques : cette biozone est caractérisée par un très grand renouvellement faunique
(Fig. 7 ![]() )
associé à une importante phase de transgression marine (,
2005). Ce dernier est concrétisé par l'absence totale de Pulchelliidae et des
Hemihoplitinae, toutefois les Gassendiceratinae , & ,
2006, restent bien présents avec le genre Imerites (, 1933)
(Fig. 5
)
associé à une importante phase de transgression marine (,
2005). Ce dernier est concrétisé par l'absence totale de Pulchelliidae et des
Hemihoplitinae, toutefois les Gassendiceratinae , & ,
2006, restent bien présents avec le genre Imerites (, 1933)
(Fig. 5 ![]() ). En
effet, de nouveaux spécimens de ce genre en cours d'étude montrent clairement un lien direct entre les derniers Pseudoshasticrioceras et les premiers Imerites. Les Peirescinae , & ,
2006, sont toujours rares avec le genre Spinocrioceras , 1973. Ce renouvellement faunique est toutefois surtout remarquable par la prolifération des Heteroceratidae qui deviennent prépondérants : on observe la forte présence du genre Heteroceras d', 1850, et dans une moindre mesure de Rouchadzeites & ,
2006, Egoianites & ,
2006, et Argvethites , 1933.
). En
effet, de nouveaux spécimens de ce genre en cours d'étude montrent clairement un lien direct entre les derniers Pseudoshasticrioceras et les premiers Imerites. Les Peirescinae , & ,
2006, sont toujours rares avec le genre Spinocrioceras , 1973. Ce renouvellement faunique est toutefois surtout remarquable par la prolifération des Heteroceratidae qui deviennent prépondérants : on observe la forte présence du genre Heteroceras d', 1850, et dans une moindre mesure de Rouchadzeites & ,
2006, Egoianites & ,
2006, et Argvethites , 1933.
Dans les travaux antérieurs sur le Sud-Est de la France (,
1994,
1997a, 1997b), et dans la synthèse de & (2006), la
Biozone à Giraudi était surmontée par la Biozone à Sarasini en réalité caractérisée dans la pratique par l'apparition du genre Martelites , 1989. Or des découvertes très récentes dans plusieurs coupes de la région stratotypique
(Fig. 5 ![]() )
montrent la présence de Martelites sp. et Martelites aff. gamkrelidzei , 1933 dans la
Biozone à Giraudi et au sein du Biohorizon à
Emerici, soit bien avant l'apparition du taxon sarasini , 1933 avec lequel on faisait habituellement surgir le genre Martelites , 1989. Parallèlement le genre Heteroceras est encore représenté par un petit nombre d'espèces dans la
Biozone à Sarasini (Heteroceras baylei , 1876, Heteroceras fuhrae , 1997). Ainsi la limite entre les "biozones" à Giraudi et Sarasini sensu &
(2006) est très
perméable, et leur distinction basée sur un renouvellement faunique est aujourd'hui beaucoup moins
fondée. Déjà en 1983 et 1989, avait noté les affinités entre les faunes des biozones à Giraudi et à Securiformis
(sensiblement équivalente à la Biozone à Sarasini) de Géorgie. Il avait ainsi proposé de les réunir en
sous-zones d'une seule biozone. C'est cette position que nous avons retenue ici
(Fig. 6
)
montrent la présence de Martelites sp. et Martelites aff. gamkrelidzei , 1933 dans la
Biozone à Giraudi et au sein du Biohorizon à
Emerici, soit bien avant l'apparition du taxon sarasini , 1933 avec lequel on faisait habituellement surgir le genre Martelites , 1989. Parallèlement le genre Heteroceras est encore représenté par un petit nombre d'espèces dans la
Biozone à Sarasini (Heteroceras baylei , 1876, Heteroceras fuhrae , 1997). Ainsi la limite entre les "biozones" à Giraudi et Sarasini sensu &
(2006) est très
perméable, et leur distinction basée sur un renouvellement faunique est aujourd'hui beaucoup moins
fondée. Déjà en 1983 et 1989, avait noté les affinités entre les faunes des biozones à Giraudi et à Securiformis
(sensiblement équivalente à la Biozone à Sarasini) de Géorgie. Il avait ainsi proposé de les réunir en
sous-zones d'une seule biozone. C'est cette position que nous avons retenue ici
(Fig. 6 ![]() ).
).
Auteur : proposée en tant que sous-zone pour la première fois par (1983) pour le Sud-Est de l'ex-URSS.
Espèce index : Imerites giraudi (, 1888).
Auteur : proposé pour la première fois par en 1997a-1997b.
Espèce index : Imerites giraudi (, 1888).
Assemblages fauniques : présence, en plus de l'espèce index, d'Heteroceras coulleti , 1994, et Heteroceras baylei , 1876, ainsi que de formes plus rares comme Heteroceras rawsonii & , 2006. On notera aussi la présence des Peirescinae avec Spinocrioceras amadei (, 1883).
Auteur : proposé pour la première fois par en 1997a-1997b.
Espèce-index : Heteroceras emerici d', 1842.
Statut et assemblages fauniques : ce biohorizon est défini par l'explosion morphologique des Heteroceratidae avec l'apparition d'Heteroceras emerici d', 1842, ainsi que par des formes de petite taille du groupe d'Heteroceras moriezense , 1997, et Heteroceras baylei , 1876. D'autres taxons plus rares appartenant aux genres Egoianites & , 2006, et Rouchadzeites & , 2006, sont présents, ainsi que les tous premiers Martelites (Martelites aff. gamkrelidzei , 1933). On notera la persistance des Peirescinae , & , 2006, avec Spinocrioceras trachyomphalus (, 1883). Le Biohorizon à Emerici se termine actuellement par la fin de la phase acméique de son espèce index.
Auteur : proposée pour la première fois par (1990) en tant que biozone.
Espèce-index : Martelites sarasini (, 1933).
Statut et assemblages fauniques : cette sous-zone débute par le Biohorizon à Puzosianum et est donc définie par l'apparition d'Anglesites puzosianum (d', 1842). Elle est concrétisée par la prédominance des Martelites par rapport aux Heteroceras. Le genre Kutatissites , 1970, apparaît dans la partie supérieure de la sous-zone. D'une manière générale, on assiste en milieu de bassin à un appauvrissement qualitatif et quantitatif des faunes d'ammonites.
Auteur : introduit pour la première fois en tant que biozone par (1965).
Espèce index : Anglesites puzosianum (d', 1842), très récemment révisé ( & , 2007).
Assemblages fauniques : la base de ce biohorizon est définie par la présence de nombreux Anglesites puzosianum (d', 1842). Il est accompagné de Martelites sarasini (, 1933) et d'Heteroceras baylei , 1876. Le Biohorizon à Puzosianum se termine actuellement par la fin de la phase acméique de son espèce index.
Auteur : cette unité a été proposée comme biohorizon par & en 1995.
Espèce index : Pseudocrioceras waagenoides (, 1938).
Statut : cette unité a été élevée au rang de biozone par & (2000), et plus récemment en tant que sous-zone par & (2006). Nous avions proposé à Digne ( et alii, 2007, inédit) de la ramener à son statut originel pour les raisons suivantes :
répartition très localisée du genre en liaison avec des contraintes écologiques très fortes (prolifération sur les pourtours du bassin vocontien et quasi-absence en domaine pélagique) ;
présence conjointe des premiers Pseudocrioceras avec les derniers Martelites (, & , 1998).
Toutefois, par consensus avec nos collègues du Group ( et alii,
2007, inédit), et dans l'attente d'une meilleure connaissance des niveaux terminaux du Barrémien dans le bassin
vocontien, nous acceptons ici le positionnement de cette unité au rang de sous-zone
(Fig. 6 ![]() ).
).
Assemblages fauniques : prolifération sur les pourtours du bassin vocontien du genre Pseudocrioceras , 1924 où il est représenté par de nombreux taxons. Dans le domaine pélagique les Pseudocrioceras sont quasiment absents, et la Sous-Zone à Waagenoides n'y a jusqu'à présent encore jamais été mise en évidence. Toutefois cette unité pourrait éventuellement être matérialisée en bassin par quelques représentants du genre Kutatissites , 1970, mais ce genre est très mal connu tant au plan paléontologique que stratigraphique.
Le nouveau schéma zonal proposé dans ce travail s'appuie essentiellement sur les renouvellements fauniques mis en évidence dans le Barrémien supérieur du Sud-Est de la France
(Fig. 7 ![]() ), et souvent associés avec les variations
eustatiques. L'extension de cette biozonation au domaine téthysien devrait en
être facilitée ainsi que les corrélations à plus grande distance,
même en l'absence des espèces index. Ces renouvellements fauniques peuvent
être résumés comme
suit :
), et souvent associés avec les variations
eustatiques. L'extension de cette biozonation au domaine téthysien devrait en
être facilitée ainsi que les corrélations à plus grande distance,
même en l'absence des espèces index. Ces renouvellements fauniques peuvent
être résumés comme
suit :
Biozone à Vandenheckei (base du Barrémien supérieur) : renouvellement et diversification des Ancyloceratoidea ;
Biozone à Sartousiana : forte diversification des Hemihoplitidae au détriment des Pulchellidae au sommet de la biozone après l'acmé du genre Gerhardtia ;
Biozone à Giraudi : les Hemihoplitinae et les Pulcheliidae ont disparu, forte diversification des Heteroceratidae notamment dans la partie inférieure de la biozone, puis apparition et prolifération locales des genres Kutatissites et Pseudocrioceras dans la partie terminale de la biozone.
La poursuite de cette démarche de révision de la biostratigraphie du Barrémien devra se faire en relation avec les révisions nécessaires des faunes d'ammonites qui lui servent de support, et en tenant bien entendu compte des données historiques. D'une manière générale, si la recherche d'une meilleure précision dans l'écoulement du temps est une nécessité pour les recherches paléobiologiques (entre autres), elle devrait se faire dans le cadre des biohorizons fauniques, et non par la multiplication des biozones qui doivent rester des unités de références stables, reproductibles et cohérentes.
Nous tenons à exprimer notre gratitude à nos collègues français du Group, ainsi qu'aux relecteurs, et en particulier à Stéphane , Miguel et Robert pour leurs précieux conseils dans l'élaboration de ce travail. Nous remercions également la Réserve Géologique de Haute Provence en la personne de sa directrice Myette pour nous avoir autorisé l'accès aux coupes étudiées. Enfin, nous ne saurions oublier dans ces remerciements Nestor qui a significativement contribué à l'amélioration de la version anglaise abrégée.
H. (2005).- The South-East France Basin (SFB) and its Mesozoic evolution.- Géologie Alpine, Grenoble, (Série spéciale "Colloques et Excursions"), n° 7, p. 5-28.
D., G. & S. (2006).- Descriptions de représentants nouveaux ou peu connus de la Famille des Hemihoplitidae , 1924 (Barrémien supérieur, Sud-Est de la France) : conséquence taxinomique et phylétiques.- Annales du Muséum d'Histoire Naturelle de Nice, t. XXI, p. 179-253.
R. (1965).- Rapport sur l'étage Barrémien. In: Colloque sur le Crétacé inférieur (Lyon, septembre 1963).- Mémoires du Bureau de Recherches Géologiques et Minières, Orléans, n° 34, p. 161-169.
R. (1984).- Crétacé inférieur. 1.3.1. Ammonites. In : Synthèse géologique du Sud-Est de la France.- Bureau de Recherches Géologiques et Minières, Mémoire, Orléans, n° 125, p. 292-294.
M., J. & J. M. (1995).- Lower Barremian ammonite biostratigraphy in the Subbetic Domain (Betic Cordillera, southern Spain).- Cretaceous Research, London, vol. 16, n° 2-3, p. 243-256.
M., J. & J.M. (2004).- El Barremiense de Foncalent y L'Alcoraia (Alicante).- Geo-Temas, Madrid, vol. 7, p. 217-221.
M., J., J.M., M. & M. (2008).- Barremian ammonite faunas from the western High Atlas, Morocco Biostratigraphy and palaeobiogeography.- Cretaceous Research, London, vol. 29, n° 1, p. 9-26.
G. (1990).- Précisions nouvelles sur "Emericiceras" collignoni (Ammonoidea) et proposition d'un nouveau schéma zonal dans le Barrémien supérieur du Sud-Est de la France.- Comptes Rendus de l'Académie des Sciences, Paris, (Série II), t. 311, p. 254-264.
G. (1994).- Les biozones à Feraudianus, Giraudi et Sarasini du Barrémien supérieur de la région stratotypique d'Angles-Barrême-Castellane (Sud-Est de la France).- Géologie Alpine, Grenoble, Mémoire H.S., n° 20, p. 279-319.
G. (1997a).- Biostratigraphie des faunes d'Ammonites à la limite Barrémien-Aptien dans la région d'Angles-Barrême-Castellane. Étude particulière de la Famille des Heteroceratidae (Ancyloceratina, Ammonoidea).- Annales du Muséum d'Histoire Naturelle de Nice, t. XII, 270 p.
G. (1997b).- Biostratigraphie haute résolution du Barrémien supérieur du Sud-Est de la France.- Comptes Rendus de l'Académie des Sciences, Paris, (Série II, Sciences de la Terre et des Planètes), t. 325, p. 689-694.
G. & D. (2006).- Nouvelles unités taxinomiques de la Famille des Heteroceratidae , 1924.- Annales du Muséum d'Histoire Naturelle de Nice, t. XXI, p. 255-295.
G. & R. (2007).- Anglesites gen. nov. (Ammonoidea, Ancyloceratina), un nouveau genre d'ammonites hétéromorphes du Barrémien supérieur du Sud-Est de la France.- Geobios, Villeurbanne, vol. 40, n° 6, p. 801-807.
M.S. (1955).- Nizhnemelovaja fauna Gruzii (Lower Cretaceous fauna of Georgia).- Institut Geologii i Mineralogii Akademii Nauk G.S.S.R., Monografii, Tbilisi, 6, p. 1-224 (en russe).
J. (1976).- Le Toarcien à Thouars et dans le Centre-Ouest de la France. Biostratigraphie. Évolution de la faune (Harpoceratinae, Hildoceratinae).- Série "Les stratotypes français", vol. 3, Éditions du Centre National de la Recherche Scientifique, Paris, 217 p.
P.J. (2004).- On the Barremian-lower Albian stratigraphy of Colombia.- Scripta Geologica, Leiden, n° 128, p. 3-15.
P.J. & L. (reporters), E., R., M., G., M., E., J., I., J.L., L., P.F., J., J.M., J.-P., G., Z. & J. (1990).- Preliminary ammonite zonation for the lower Cretaceous of Mediterranean region (Lower Cretaceous Cephalopod team, Working Group of IGCP, Project 262).- Géologie Alpine, Grenoble, t. 66, p. 123-127.
P.J. & F. (reporters), E., M., G., E., M., P., M., G., A., L., G., P.F., P., J., J.M. & Z. (1995).- Report on the 3rd International Workshop on the standard Lower Cretaceous ammonite zonation of the Mediterranean region.- Memorie Descrittive della Carta Geologica d'Italia, Roma, vol. LI, p. 213-215.
P.J. & M. (reporters), M.B., E., T.N., L., L., F., G., M., L., H., P.F., J., J.M., J.-P., S.Z. & Z. (1993).- Ammonites zonation for the Lower Cretaceous of the Mediterranean region; basis for the stratigraphic correlation within IGCP-Project 262.- Revista Española de Paleontologia, Madrid, vol. 8, n° 1 , p. 117-120.
P. & H. (1995).- Biostratigraphy and sequence stratigraphy of the Berriasian-lowest Aptian (Lower Cretaceous) of the Rio Argos succession, Caravaca, SE Spain.- Cretaceous Research, London, vol. 16, n° 2-3, p. 169-230.
P.J. & P.F. (reporters), M.B., T.N., I., P., C., M., J., A., N., O. & Z. (2000).- Report on the 5th International Workshop of the Lower Cretaceous Cephalopod team (Vienna, 5 September 2000).- Cretaceous Research, London, vol. 21, n° 6, p. 857-860.
P.J. & S. (reporters), M.B., P., M., F., R., M., C., J., A., I., N., P.F., P., Z., J. & M.G.E. (2003).- Report on the 1st International Workshop of the IUGS Lower Cretaceous Ammonite Working Group, the ' Group' (Lyon, 11 July 2002).- Cretaceous Research, London, vol. 24, n° 1, p. 89-94 ; erratum, vol. 24, n° 6, p. 805.
M.V. (1983).- On the Hauterivian-Barremian correlations between the South of the USSR and certain southern and northern regions of Europe. In : 2. Symposium Kreide. München 1982.- Zitteliana, München, (Reihe B: Abhandlungen der Bayerischen Staatssammlung für Paläontologie und Geologie), 10, p. 501-508.
M.V. (1989).- The Barremian biostratigraphical subdivisions of Georgia and comparison with some western mediterranean regions. In : J. (ed.), Cretaceous of the Western Tethys.- Proceedings 3rd International Cretaceous Symposium, Tubingen (1987), E. Schweizerbart'sche Verlagsbuchandlung, Stuttgart, p. 551-560.
M.V. & E. (1995).- New data on the Upper Barremian biostratigraphy of the Georgian region (Caucasus). In : F. (ed.), Proceeding of 3rd Workshop on Early Cretaceous Cephalopods.- Memorie Descrittive della Carta Geologica d'Italia, Roma, vol. LI, p. 103-109.
M.V. (1900).- Recherches géologiques dans le Diois et les Baronnies orientales.- Travaux du Laboratoire de Géologie de la Faculté des Sciences de l'Université de Grenoble, t. 5, p. 149-556.
P.F & P.J. (reporters), M.B, E., M., S.R.A., J., E., H.G., P., M.R.A., M. & Z. (1999).- Report on the 4th International Workshop of the Lower Cretaceous Cephalopod team (IGPC-Project 362).- Scripta Geologica, Leiden, Special Issue 3, p. 313.
S. (reporter), F., D., L., R., G. & J. (2007, inédit).- Zonation Hauterivien Barrémien.- Compte rendu de la réunion des biostratigraphes français du Group (IUGS Lower Cretaceous Ammonites Working Group), Digne-les-Bains (2 Mai 2007), 14 p.
S. & F. (1999).- Comments and proposals about the Valanginian-Lower Hauterivian ammonite zonation of south-eastern France.- Eclogae Geologicae Helvetiae, Basel, vol. 92, n° 2, p. 183-197.
S. & P.J. (reporters), M.B., P., F., E.Y., M., G., Y., J. J.L., A., V., F.A., I., N., P., J., J.M., Z., J., H., B. & I. (2006).- Report on the second international meeting of the IUGS Lower Cretaceous Ammonite Working Group, the " Group" (Neuchâtel, Switzerland, 8 September 2005).- Cretaceous Research, London, vol. 27, n° 5, p. 712-715.
P., R. & G. (1998).- Le genre Pseudocrioceras dans les couches de passage du Barrémien supérieur / Bédoulien inférieur de Cassis-La Bédoule (SE France).- Géologie Méditerranéenne, Marseille, vol. XXV, n° 3-4, p. 85-123.
J.-P. (1979).- Maheronites limentinus n. sp. (Ammonoidea) espèce-type d'un horizon-repère Barrémien supérieur du Vercors méridional (massif subalpin français).- Geobios, Villeurbanne, Mémoire spécial, n° 3, p. 305-317.
J. (1997).- La biostratigraphie, du taxon à la biobiozone et aux échelles biozonales. In : J. (coord.), Stratigraphie. Terminologie française.- Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine, Mémoire, Pau, n° 19, p. 63-89.
J. (1997).- Biozonation ammonitique dans le Barrémien du Sud-Est de la France (de la Biozone à Hugii à la Biozone à Sartousiana).- Géologie Alpine, Grenoble, t. 73, p. 99-117.
J. (2002).- Étude stratigraphique et paléontologique de la Famille des Pulchellidae (Ammonoidea, Ammonitina, Endemoceratidae).- Géologie Alpine, Grenoble, Mémoire H.S., t. 42, p. 1-332.
J. (2005).- Boundaries, ammonite fauna and main subdivisions of the stratotype of the Barremian.- Géologie Alpine, Grenoble, (Série spéciale "Colloques et Excursions"), n° 7, p. 147-173.
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 1: Corrélations des principales biozonations du Barrémien depuis
1965.
Correlation of the main Barremian biozonations since 1965.
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 2: Cadre paléogéographique du Sud-Est de la France et localisation du secteur d'étude
(d'après , 2005).
Barremian paleogeography of southeastern France and location of the area studied (from ,
2005).
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
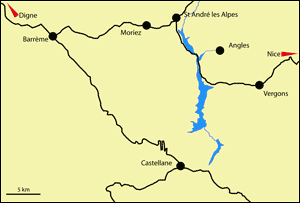
Figure 3:
Localisation géographique de l'aire d'étude (Alpes-de-Haute-Provence, Sud-Est de la France).
Geographic location of the area studied (Alpes-de-Haute-Provence, southeastern France).
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
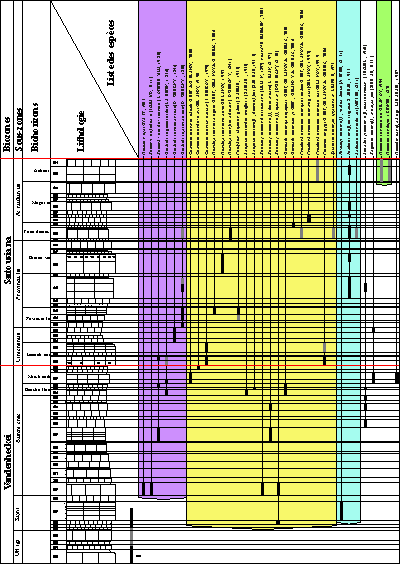
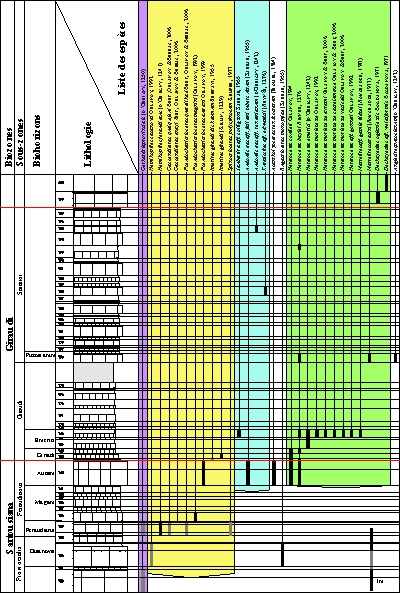
Figure 4: Répartition des faunes dans la coupe LAC
(secteur de Méouille, Saint-André-les-Alpes, Alpes-de-Haute-Provence). En grisé, les espèces trouvées en place
latéralement.
Distribution of faunas in the LAC section (Méouille area, Saint-André-les-Alpes,
Alpes-de-Haute-Provence). In grey,
the species found in stratigraphically equivalent strata (outside the main section).
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 5:
Répartition des faunes dans la coupe MEO (secteur de Méouille,
Saint-André-les-Alpes, Alpes-de-Haute-Provence). En grisé, les espèces trouvées en place
latéralement.
Distribution of faunas in the MEO section (Méouille area, Saint-André-les-Alpes,
Alpes-de-Haute-Provence). In grey,
the species found in stratigraphically equivalent strata (outside the main
section).
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
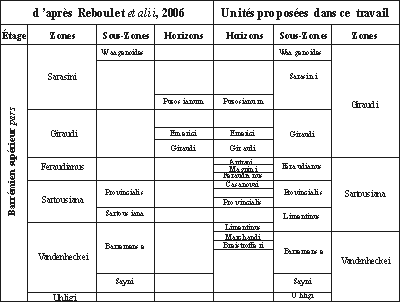
Figure 6:
Comparaison des schémas zonaux issus de la réunion de l'IUGS Group
(à Neuchâtel en 2005 : & , 2006) et de ce travail.
Chart comparing the zonation of the IUGS Group meeting
(Neuchâtel, 2005: & , 2006)
with that proposed in this work.
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
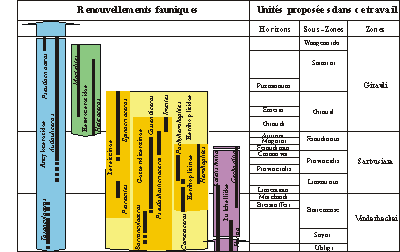
Figure 7:
Schéma des renouvellements fauniques majeurs au cours du Barrémien supérieur et nouveau schéma zonal
associé.
Chart of the major faunal turnovers during the Late Barremian and the new zonal
succession associated with it.