![]()
Sommaire
[Introduction] [I- Les Pseudoperisphinctinae ...]
[II- Choffatia isabellae n. sp.]
[Conclusion]
[Références bibliographiques] [Figures]
et ... [Planches]
Centre des sciences de la Terre et de l'Environnement, Université de
Bourgogne, UMR CNRS 5561, "Biogéosciences", 6 bd Gabriel, 21000 Dijon
(France)
14 rue Johannes, 49450 Villedieu-la-Blouère (France)
4 rue Parigny, 86280 Saint-Benoît (France)
Résidence Origan, Bâtiment A, 8 Avenue Sainte Claire, 06100 Nice (France) ; Centre des sciences de la Terre et de l'Environnement, Université de Bourgogne, UMR CNRS 5561, "Biogéosciences", 6 bd Gabriel, 21000 Dijon (France)
Manuscrit en ligne depuis le 17 Avril 2008
![]()
Dans la région de Montreuil-Bellay (Maine-et-Loire), de nombreuses coupes ont été réalisées au passage Callovien moyen-Callovien supérieur. Le premier banc attribué au Callovien supérieur a été daté de l'horizon à Leckenbyi. Il a fourni une très importante faune ammonitique (N=3125), dans laquelle les Perisphinctidae représentent 51% de l'effectif. À côté de formes plus ou moins bien connues comme Pseudopeltoceras leckenbyi (), Orionoides pseudorion (), Subgrossouvria famulum () et S. crassa et , on trouve une espèce qui n'a jamais été ni décrite ni figurée : cette espèce fait l'objet du présent article. Choffatia isabellae n. sp. se distingue sans aucune ambiguité des Perisphinctidae contemporains par : 1) un long stade juvénile lisse, 2) une costulation habituellement ténue, 3) l'absence de formations paraboliques et 4) la très grande fréquence des constrictions. Comme cette espèce est inconnue dans les faunes de l'extrême sommet du Callovien moyen, nous pensons qu'elle a colonisé, avec beaucoup d'autres espèces, dont Peltoceras marysae et alii, via la marge sud de la Téthys, la plate-forme nord-ouest européenne à la faveur de l'intervalle transgressif qui débute à l'extrême base du Callovien supérieur. Apparue brutalement à la base de l'horizon à Leckenbyi, cette nouvelle espèce possède son acmé dans la partie médiane de l'horizon, oů elle peut représenter la moitié des Perisphinctidae, avant de devenir rare dans la partie supérieure de l'horizon, puis très rare dans l'horizon à Athleta.
Callovien supérieur ; zone à Athleta ; horizon à Leckenbyi ; Montreuil-Bellay ; France ; Perisphinctidae ; espèce nouvelle ; dimorphisme sexuel.
A., P.-Y., P. & D. (2008).- Les Pseudoperisphinctinae (Ammonitina, Perisphinctidae) de l'horizon à Leckenbyi (Callovien supérieur, zone à Athleta) de Montreuil-Bellay (Maine-et-Loire, France) et description d'une nouvelle espèce, Choffatia isabellae.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2008/05 (CG2008_A05)
Pseudoperisphinctinae (Ammonitina, Perisphinctidae) from the Leckenbyi horizon (Upper Callovian, Athleta zone) of Montreuil-Bellay (Maine-et-Loire, France) and description of a new species, Choffatia isabellae.- Many outcrops exposing the Mid-Callovian/Upper Callovian boundary have been found near Montreuil-Bellay (Maine-et-Loire). The lowermost bed of the Upper Callovian has been dated as the Leckenbyi horizon. It contains an abundant ammonite fauna (3125 specimens), in which Perisphinctidae make up just over half of the specimens. In company with reasonably well-known species such as Pseudopeltoceras leckenbyi (), Orionoides pseudorion (), Subgrossouvria famulum () and S. crassa et , occurs a species, previously unknown, which is described in this article. Choffatia isabellae n. sp. differs markedly from contemporary Perisphinctidae, for it is distinguished by : 1) a long, smooth juvenile stage, 2) weakly developed ribbing, 3) the absence of parabolic ornamentation and 4) a high frequency of constrictions. As this species is unknown in the fauna of the uppermost Middle Callovian, along with many other species, including Peltoceras marysae et alii, it must have colonised the northwestern part of the European platform via the southern margin of the Tethys during the transgressive interval which started at the beginning of the Late Callovian. This new species appears abruptly at the base of the Leckenbyi horizon, reaches its acme in the middle of the horizon, where it probably represents half of the Perisphinctidae, before becoming rare in the upper part of the horizon, then very rare in the Athleta horizon.
Upper Callovian; Athleta zone; Leckenbyi horizon; Montreuil-Bellay; France; Perisphinctidae; new species; sexual dimorphism.
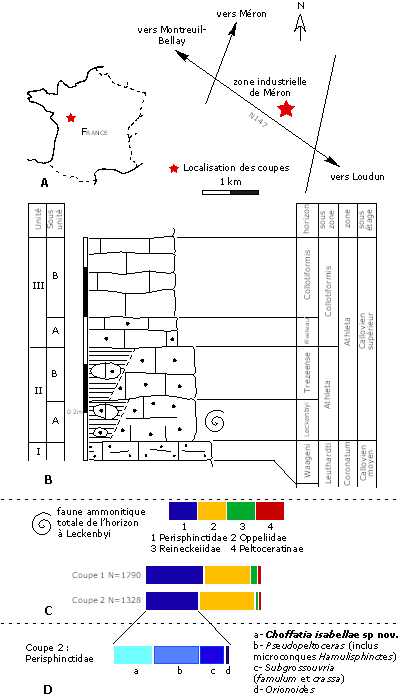
Le Callovien supérieur de Montreuil-Bellay (Maine-et-Loire) est bien connu pour ses faunes ammonitiques, et on trouvera un historique des travaux anciens dans et
(1936). Dans le cadre de la révision des Aspidoceratidae du Callovien supérieur et de l'Oxfordien inférieur d'Europe occidentale effectuée par l'un d'entre nous (,
1995), plusieurs coupes ont été réalisées dans la zone industrielle de Méron, située au sud-est de l'agglomération
(Fig. 1A ![]() ). Ces recherches ont permis, entre autre, de découvrir le plus ancien Euaspidoceratinae et le plus ancien Peltoceratinae européens connus à ce jour, respectivement Euaspidoceras raynaudi (,
1996) et Peltoceras marysae (et alii, 2005).
). Ces recherches ont permis, entre autre, de découvrir le plus ancien Euaspidoceratinae et le plus ancien Peltoceratinae européens connus à ce jour, respectivement Euaspidoceras raynaudi (,
1996) et Peltoceras marysae (et alii, 2005).
Si la succession lithologique au passage Callovien moyen – Callovien supérieur est connue depuis très longtemps dans ses grandes lignes, on sait également qu'elle est très variée dans le détail. Des travaux récents, réalisés à grande échelle dans la zone industrielle de Méron, ont permis d'observer ces niveaux en "3D" de façon continue sur plusieurs centaines de mètres et de préciser les variations horizontales de faciès, associées ou non à des différences d'âge (travail en cours).
Le matériel étudié dans cet article provient exclusivement du banc IIA, exploité dans quatre coupes oů la totalité du banc II est carbonaté. Les raisons pour lesquelles ce banc a été daté 1) du Callovien supérieur et 2) de l'horizon à Leckenbyi ont été exposées dans un précédent article ( et alii, 2005). On peut les résumer ainsi :
En ce qui concerne plus particulièrement l'horizon à Leckenbyi, l'analyse détaillée des faunes récoltées montre que, si toutes les espèces reconnues sont présentes dans la totalité de l'horizon, leur importance quantitative peut être variable. Par exemple :
La matrice est un calcaire blanchâtre légèrement argileux à oolites ferrugineuses, avec cristallisation éventuelle en calcite translucide, blanche, orange ou verdâtre. Les ammonites sont conservées à l'état de moule interne avec un pseudotest qui rend délicate l'observation des lignes de suture et donc l'estimation du degré de maturité des individus.
Plusieurs arguments plaident en faveur d'un transport post mortem très réduit :
de très nombreux spécimens d'ammonites, toutes familles confondues, comportent le phragmocone et la chambre d'habitation et
des structures fragiles sont parfaitement conservées comme les apophyses des ammonites microconques ou le péristome des gastéropodes.
Une ammonite est considérée comme adulte lorsque ses dernières cloisons sont approximées (voire télescopées). Le péristome peut alors être semblable au péristome juvénile (cas fréquent chez les macroconques) ou différent (cas habituel chez les microconques). Les lignes de suture des spécimens étudiés ici sont rarement observables, mais dans les quelques cas oů cela a été possible, on a pu définir la morphologie adulte, en particulier les caractères de la costulation et la fréquence des constrictions.
Nous ne reviendrons pas sur la mise en évidence d'un dimorphisme attribué au sexe. La famille des Perisphinctidae ne déroge pas à cette "règle" et de très nombreux auteurs ont décrits dans le Callovien des formes microconques et des formes macroconques (dont , 1970, et , 1988). D'un point de vue taxonomique, elles sont attribuées par les auteurs soit à la même espèce (ce qui devrait être la règle), soit à deux sous-genres du même genre (pour des raisons pratiques ou historiques).
Sera considéré comme macroconque adulte tout individu de grande taille (pour l'espèce) dont les dernières cloisons sont rapprochées et dont le péristome est simple, mais aussi tout individu présentant, par comparaison, la morphologie adulte même si ses cloisons ne sont pas observables.
Sera considéré comme microconque adulte tout individu de petite taille (pour l'espèce) dont le péristome est muni d'apophyses longues, même si l'approximation des dernières cloisons n'a pas été observée.
Les Perisphinctidae de l'horizon à Leckenbyi de Montreuil-Bellay se répartissent autour de cinq grandes morphologies :
En résumé, les Perisphinctidae sont représentés dans l'horizon à Leckenbyi de Montreuil-Bellay par cinq groupes morphologiques assimilés à des bioespèces, avec leur dimorphisme sexuel et leur variabilité intraspécifique : Choffatia isabellae n. sp., Subgrossouvria famulum (), Subgrossouvria crassa et , Orionoides pseudorion (), Pseudopeltoceras leckenbyi (). Seule, la première espèce fait l'objet du présent article. La révision des autres espèces est en cours. On peut remarquer que et (1936) ne figurent, dans leur publication consacrée aux ammonites de la zone à Peltoceras athleta du centre-ouest de la France, ni Orionoides sensu stricto, ni de spécimens attribuables à Choffatia isabellae.
Aspect quantitatif
Selon les coupes, les Perisphinctidae représentent entre 43 et 55% de la faune ammonitique totale
(voir Fig. 1C ![]() pour les deux coupes ayant fourni l'essentiel du matériel). La part relative de chaque espèce varie selon les coupes, en fonction de leur localisation à l'intérieur de l'horizon
(voir Fig. 1D
pour les deux coupes ayant fourni l'essentiel du matériel). La part relative de chaque espèce varie selon les coupes, en fonction de leur localisation à l'intérieur de l'horizon
(voir Fig. 1D ![]() pour le détail de la coupe 2). En ce qui concerne Choffatia isabellae n. sp., elle est présente dès la base de l'horizon, devient plus abondante vers la partie moyenne (c'est la situation des deux coupes) et se raréfie au sommet.
pour le détail de la coupe 2). En ce qui concerne Choffatia isabellae n. sp., elle est présente dès la base de l'horizon, devient plus abondante vers la partie moyenne (c'est la situation des deux coupes) et se raréfie au sommet.
Famille Perisphinctidae
Sous-famille Pseudoperisphinctinae
Genre Choffatia
Choffatia isabellae , , et
Holotype : spécimen Mbe/n/221. Collection Pierre-Yves . Pl. 3, fig. 2 ![]() ;
Fig. 5C
;
Fig. 5C ![]() .
.
Syntypes : tous les autres spécimens figurés.
Derivatio nominis : l'espèce est dédiée à l'épouse de l'un d'entre nous (PF).
Locus typicus : Montreuil-Bellay (Maine-et-Loire, France).
Stratum typicum : Callovien supérieur, zone à Athleta, sous-zone à Trezeense, horizon à Leckenbyi.
L'espèce comprend des macroconques et des microconques.
Ombilic ouvert. Constrictions très fréquentes, proverses, présentes sur le phragmocône et sur la chambre d'habitation. Absence de formations paraboliques. Côtes simples rares. Côtes intercalaires. Côtes divisées bi-, tri- ou polyfurquées.
Taille petite à moyenne (80 à 200 mm). Chambre d'habitation en moyenne d'un peu moins d'un tour. Péristome simple. Deux pôles morphologiques avec des intermédiaires :
Pôle subserpenticone : section ovale plus ou moins haute, à épaisseur maximale assez bas sur les flancs. Flancs bombés, rarement aplatis. Stade lisse habituellement long. Côtes peu marquées. Constrictions peu profondes.
Pôle serpenticone : section subcirculaire avec la plus grande épaisseur au milieu des flancs. Stade lisse habituellement court. Côtes fortes. Constrictions profondes.
Petite taille (25 à 85 mm). Chambre d'habitation en moyenne entre 3/4 de tour et un tour. Péristome à apophyses longues. Enroulement subserpenticone à section ovale. Flancs et ventre arrondis. Costulation discrète.
Sur N=25, la taille adulte varie de 80 mm à 200 mm (moyenne 115 mm). Cette importante amplitude de taille n'est pas habituelle mais on peut remarquer 1) qu'il n'existe dans l'échantillon qu'un seul spécimen de 200 mm, les autres grands macroconques mesurant plutôt entre 140 et 160 mm et 2) qu'une amplitude proche est connue par ailleurs chez les Perisphinctaceae, par exemple chez Peltoceras athleta, avec une taille comprise entre 130 et 300 mm. On pourrait avancer l'hypothèse de plusieurs espèces successives, avec augmentation de la taille. Mais cette hypothèse ne résiste pas aux données de terrain, puisque les quelques spécimens les plus récents ne se distinguent pas des spécimens du milieu de l'horizon. Quelques spécimens adultes, de taille plus petite, peuvent être considérés comme nains. D'autres spécimens sont certainement adultes, mais incomplets et donc non reconnaissables comme tels. La longueur de la chambre d'habitation varie de 300 à 430° (moyenne 340° pour N=19, soit un peu moins d'un tour).
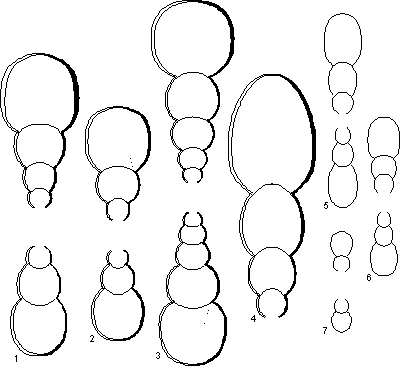
On peut définir deux groupes morphologiques : morphologie subserpenticone à section subcirculaire puis ovale
(Fig. 2.4 ![]() ), rarement à flancs aplatis dans leur partie inférieure et morphologie plus serpenticone à section restant subcirculaire
(Fig. 2.2 et 2.3
), rarement à flancs aplatis dans leur partie inférieure et morphologie plus serpenticone à section restant subcirculaire
(Fig. 2.2 et 2.3 ![]() ). Il semblerait que les spécimens de ce groupe soit en moyenne plus petits que les autres. Tous les intermédiaires existent
(Fig. 2.1
). Il semblerait que les spécimens de ce groupe soit en moyenne plus petits que les autres. Tous les intermédiaires existent
(Fig. 2.1 ![]() ), si bien que ces groupes sont interprétés comme de simples pôles de variabilité. La région ventrale est toujours arrondie. L'ombilic est toujours ouvert. La fin de la chambre d'habitation est caractérisée par une dernière constriction, habituellement très proverse et qui perturbe la costulation lorsque celle-ci persiste. Le péristome est simple, proverse, sans aucune expansion.
), si bien que ces groupes sont interprétés comme de simples pôles de variabilité. La région ventrale est toujours arrondie. L'ombilic est toujours ouvert. La fin de la chambre d'habitation est caractérisée par une dernière constriction, habituellement très proverse et qui perturbe la costulation lorsque celle-ci persiste. Le péristome est simple, proverse, sans aucune expansion.
Le stade lisse est toujours bien individualisé, mais de longueur variable. Il est plus long chez les spécimens à costulation ténue, et plus court chez les spécimens à costulation forte, bien que des exceptions existent
(Pl. 2, fig. 1 ![]() ).
).
Les côtes sont généralement proverses, un peu arquées, très rarement flexueuses ; à noter un spécimen qui possède des côtes flexueuses sur un seul flanc. Les côtes sont divisées, avec un mode de division variable : il existe des côtes bifurquées, trifurquées, voire polyfurquées. Il n'y a pas de côtes simples, mais les côtes intercalaires sont plus ou moins fréquentes. Les côtes secondaires sont plus fines que les côtes primaires et la différence s'accentue au cours de la croissance, surtout lorsque les côtes primaires sont fortes. Les côtes secondaires et les côtes intercalaires passent sur la région ventrale sans s'interrompre et sans changer de trajectoire. Les adultes de petite taille gardent des côtes secondaires jusqu'à la fin de la chambre d'habitation, alors qu'elles s'estompent, ainsi que les primaires, chez les adultes de grande taille.
Les constrictions sont particulièrement fréquentes, présentes à la fois sur le phragmocône et sur la chambre d'habitation. On en dénombre entre 10 et 16 au total (moyenne 12 pour N=43) et entre 1 et 5 sur le dernier tour, soit approximativement la chambre d'habitation (moyenne 2 pour N=97). Dans les tours initiaux, la présence de nombreuses constrictions est corrélée avec un enroulement "polygonal", de triangulaire à quadrangulaire, par courtes sections à la limite du déroulement
(Fig. 3 ![]() ). Les constrictions sont en moyenne plus profondes chez les spécimens à section arrondie. Leur trajectoire est habituellement la même que celle des côtes, mais elle peut être un peu plus proverse. Aucune formation parabolique n'a été observée.
). Les constrictions sont en moyenne plus profondes chez les spécimens à section arrondie. Leur trajectoire est habituellement la même que celle des côtes, mais elle peut être un peu plus proverse. Aucune formation parabolique n'a été observée.
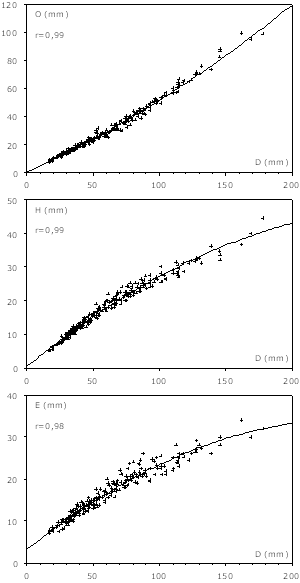
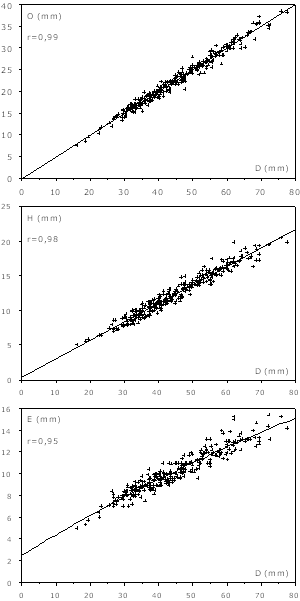
Les croissances de l'ombilic, de la hauteur et de l'épaisseur sont très fortement corrélées avec le diamètre (r vaut respectivement 0,99, 0,99 et 0,98), ce qui vérifie l'homogénéité de l'échantillon. La croissance s'effectue sans rupture évidente, et l'ombilic s'ouvre très progressivement en même temps que la hauteur et l'épaisseur diminuent de la même façon.
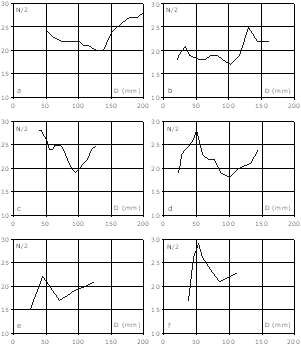
Chez la grande majorité des spécimens, la densité de la costulation (nombre de côtes par demi-tour) évolue de façon caractéristique :
Chez quelques grands spécimens, la densité costale diminue à nouveau avant le péristome.
Elle a été dégagée à l'acide sur deux spécimens :
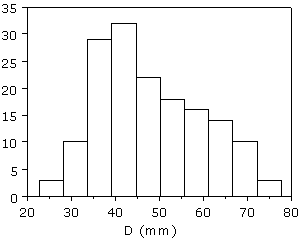
La taille adulte est comprise entre 23 et 85 mm avec une moyenne de 48 mm (N=158).
La légère dissymétrie (médiane = 42 mm) de l'histogramme
(Fig. 7 ![]() )
est due à l'excès de formes adultes de petite taille, mais elle ne remet aucunement en cause l'homogénéité de l'échantillon. Une telle variabilité n'est pas rare ; elle a été observée dans d'autres groupes de Perisphinctaceae, entre autres les Peltoceratinae chez lesquels par exemple le microconque de Peltoceras athleta a une taille comprise entre 30 et 105 mm (,
1993, 1995). La longueur de la chambre d'habitation est comprise entre 240 et 350 degrés, avec une moyenne de 300° (N=15). Le stade adulte est marqué par une dernière constriction, suivie d'un bourrelet habituellement peu marqué. Les apophyses latérales sont longues, légèrement courbes et non spatulées
(Fig. 8
)
est due à l'excès de formes adultes de petite taille, mais elle ne remet aucunement en cause l'homogénéité de l'échantillon. Une telle variabilité n'est pas rare ; elle a été observée dans d'autres groupes de Perisphinctaceae, entre autres les Peltoceratinae chez lesquels par exemple le microconque de Peltoceras athleta a une taille comprise entre 30 et 105 mm (,
1993, 1995). La longueur de la chambre d'habitation est comprise entre 240 et 350 degrés, avec une moyenne de 300° (N=15). Le stade adulte est marqué par une dernière constriction, suivie d'un bourrelet habituellement peu marqué. Les apophyses latérales sont longues, légèrement courbes et non spatulées
(Fig. 8 ![]() ). Des apophyses courtes sont interprétées comme des apophyses en cours de construction. Nous n'avons jamais observé d'apophyses transitoires longues.
). Des apophyses courtes sont interprétées comme des apophyses en cours de construction. Nous n'avons jamais observé d'apophyses transitoires longues.
L'enroulement est subserpenticone, avec un recouvrement constamment très faible. Il est rythmé par la présence des constrictions, particulièrement fréquentes : de 7 à 18, moyenne 11, pour l'ensemble de la coquille dont 1 à 4 sur le dernier tour, c'est-à-dire la chambre d'habitation (N=153, moyenne 2). Cela fait en moyenne 2 constrictions par tour, et souvent 3 dans les tours initiaux. Les constrictions sont corrélées dans les tours initiaux avec le même type d'enroulement "polygonal" que chez les macroconques
(Fig. 9 ![]() ).
).
Les constrictions sont bien marquées, profondes, proverses ou flexueuses, quelquefois répétées (Pl. 8, fig. 10 ![]() , et
Pl. 10, fig. 2
, et
Pl. 10, fig. 2 ![]() ).
).
La section est subcirculaire dans les tours internes puis, soit elle se comprime de plus en plus chez les adultes les plus grands
(Fig. 2.5 et 2.6 ![]() ), soit elle reste subcirculaire chez les petits adultes
(Fig. 2.7
), soit elle reste subcirculaire chez les petits adultes
(Fig. 2.7 ![]() ). E/H vaut en moyenne 0,82 juste avant l'ouverture (N=140). La croissance en épaisseur est faible, si bien que l'ombilic n'est jamais profond. Il est par contre très ouvert : O/D vaut en moyenne 0,50.
). E/H vaut en moyenne 0,82 juste avant l'ouverture (N=140). La croissance en épaisseur est faible, si bien que l'ombilic n'est jamais profond. Il est par contre très ouvert : O/D vaut en moyenne 0,50.
Il n'y a pas de formations paraboliques si ce n'est, chez un unique exemplaire, la présence de deux petites excroissances localisées juste avant une constriction et qui peuvent être interprétées comme des paraboles très discrètes
(Fig. 11 ![]() ).
).
Le stade lisse est long, pouvant atteindre un diamètre de 20 mm. Puis, d'une manière générale, la costulation est faible. Les côtes sont fines, proverses, bi- ou trifurquées, polyschizotomes, avec d'éventuelles côtes intercalaires et de rares côtes simples. La division des côtes s'effectue au milieu des flancs ou un peu audessus. Les côtes intercalaires et les côtes secondaires passent sur la région ventrale sans s'interrompre et sans dessiner de sinus. L'indice de division varie de 1,7 à 2,9 avec une moyenne de 2,3 (N=68).
Les croissances de l'ombilic, de la hauteur et de l'épaisseur sont très bien corrélées avec le diamètre (r vaut respectivement 0,99, 0,98 et 0,95), ce qui vérifie l'homogénéité de l'échantillon. La croissance de l'ombilic et de la hauteur peut être considérée comme harmonieuse, si bien qu'on n'observe pas l'habituel déroulement en fin de croissance. La croissance en épaisseur diminue progressivement, ce qui correspond au passage d'une section subcirculaire à une section légèrement comprimée.
Elle est plus simple que celle des macroconques et présente avec elle des points communs et des différences :
Elle met en évidence des points communs fondamentaux :
mais aussi des différences :
Quelques très rares macroconques (Pl. 2, fig.
5 ![]() , par exemple) ne sont pas sans rappeler Alligaticeras alligatum (), en particulier l'holotype, refiguré par
(1988) ; mais cette espèce est beaucoup plus récente (zone à Lamberti). D'autres sont proches, par leur tour externe, d'Alligaticeras sp. aff. Raguini ( et ) in
(1983,
Pl. fig. 22), mais les tours internes de ce dernier sont inconnus et il s'agit d'une forme probablement plus récente. D'autres macroconques enfin évoquent Alligaticeras rotifer () in
(1988), mais il s'agit là encore de formes plus récentes, à constrictions beaucoup moins nombreuses, dont la croissance en épaisseur est plus forte (à l'origine d'une section déprimée) et qui présentent des côtes franchement proverses dans les tours internes et des côtes secondaires fortes sur la chambre d'habitation.
, par exemple) ne sont pas sans rappeler Alligaticeras alligatum (), en particulier l'holotype, refiguré par
(1988) ; mais cette espèce est beaucoup plus récente (zone à Lamberti). D'autres sont proches, par leur tour externe, d'Alligaticeras sp. aff. Raguini ( et ) in
(1983,
Pl. fig. 22), mais les tours internes de ce dernier sont inconnus et il s'agit d'une forme probablement plus récente. D'autres macroconques enfin évoquent Alligaticeras rotifer () in
(1988), mais il s'agit là encore de formes plus récentes, à constrictions beaucoup moins nombreuses, dont la croissance en épaisseur est plus forte (à l'origine d'une section déprimée) et qui présentent des côtes franchement proverses dans les tours internes et des côtes secondaires fortes sur la chambre d'habitation.
Certains microconques montrent une ressemblance superficielle avec certaines formes réunies dans le sous-genre Grossouvria (en toute logique puisqu'il s'agit des microconques de Choffatia), mais ils s'en distinguent immédiatement par la fréquence des constrictions et l'absence de formations paraboliques.
C'est donc la réunion de quatre caractères qui fait l'originalité de Choffatia isabellae : stade lisse long, suivi d'une costulation discrète, absence de formation parabolique et fréquence élevée des constrictions. Pris séparément ou deux à deux, ces caractères sont reconnus chez certains Perisphinctidae calloviens ou même oxfordiens, mais Choffatia isabellae est la seule espèce oů ils se trouvent réunis.
Les caractéristiques spécifiques de Choffatia isabellae – grande fréquence des constrictions, absence de formations paraboliques et ornementation atténuée – ne sont jamais réunies chez les ammonites d'Europe occidentale datées du sommet du Callovien moyen. Il est donc très peu probable que Choffatia isabellae se soit individualisée à partir d'une espèce européenne.
Par ailleurs, on constate à l'extrême base du Callovien supérieur de la région de Montreuil-Bellay, et dans une certaine mesure dans toutes les régions oů affleure cette tranche de temps, un renouvellement faunique important, particulièrement net chez les ammonites, avec l'apparition de taxons caractéristiques du biota téthysien (Peltoceras, Orionoides s.s., Pseudopeltoceras, Subgrossouvria famulum, etc.). Or, la base du Callovien supérieur correspond à l'augmentation de la tranche d'eau sur les plates-formes carbonatées d'Europe occidentale, puisque la surface d'inondation maximale de la dernière séquence du Callovien (Ca5) est datée de la base de la zone à Athleta ( et , 1997). Comme Choffatia isabellae est absente des dépôts contemporains du Dijonnais (, 1995 ; , 2000), du Châtillonnais ( et , 1998 ; , 2000), du Jura français ( et alii, 1989 ; et alii, 1998) et du Jura suisse (, 1951, et observations personnelles). Il est donc envisageable que Choffatia isabellae s'enracine dans un groupe, actuellement non reconnu, colonisant la plate-forme nord-ouest européenne plutôt via la marge sud de la Téthys.
Choffatia isabellae n. sp. est encore présente de façon certaine dans l'horizon à Athleta de Montreuil-Bellay, mais elle y est beaucoup plus rare. Dans la partie supérieure de la zone à Athleta (sous-zone à Collotiformis), il est difficile de rattacher les Perisphinctidae connus à Choffatia isabellae, en particulier à cause d'une conservation nettement moins favorable à l'observation des tours initiaux. Mais il n'est pas impossible que certaines formes attribuées, à tort le plus souvent, au genre Orionoides, puissent être considérées comme des descendants potentiels de Choffatia isabellae. Dans l'horizon à Collotiformis du Dijonnais on a constaté, grâce à une conservation plus favorable, que le stade lisse à enroulement "polygonal" est encore présent, mais réduit à sa plus simple expression. Dans les niveaux plus récents (sommet du Callovien supérieur à base de l'Oxfordien inférieur), des formes attribuées au genre Alligaticeras rappellent clairement certains variants de Choffatia isabellae, mais la filiation directe ne peut être démontrée à partir du seul matériel de Montreuil-Bellay. De même, on peut constater que l'ornementation polygonale des tours initiaux se retrouvera chez des Perisphinctidae oxfordiens, sans y voir une relation particulière avec Choffatia isabellae.
Choffatia isabellae n. sp. apparaît à Montreuil-Bellay à la base de l'horizon à Leckenbyi, atteint son acmé dès le milieu de l'horizon puis devient rare au sommet de l'horizon et plus encore dans l'horizon à Athleta. Dans la mesure ou elle représente jusqu'à 10% de la faune ammonitique totale, il peut paraître surprenant que cette espèce n'ait jamais été ni décrite ni figurée. Plusieurs hypothèses peuvent être avancées, la plus plausible étant qu'elle n'a pas été reconnue comme taxon nouveau par les précédents auteurs, tout comme Peltoceras marysae et alii récemment décrite (2005), même si cette dernière espèce est par ailleurs beaucoup plus rare.
Les macroconques et les microconques possèdent des caractères communs qui, réunis, permettent de les distinguer sans difficulté de tous les autres Perisphinctidae contemporains : long stade initial lisse puis ornementation peu puissante, absence de formations paraboliques et grande fréquence des constrictions. Les macroconques sont variables par la force de leur ornementation et par leur morphologie, avec un pôle subserpenticone à section ovale comprimée et côtes peu marquées et un pôle plus serpenticone à section subcirculaire et côtes fortes. Les microconques sont moins variables par l'absence du pôle serpenticone. La chambre d'habitation mesure entre trois-quart de tour et un tour.
L'horizon à Leckenbyi, c'est-à-dire l'extrême base du Callovien supérieur, est caractérisé par un renouvellement important de la faune ammonitique. Parmi les taxons nouveaux, Choffatia isabellae est accompagné de taxons d'affinités nettement téthysiennes. On peut supposer que Choffatia isabellae fait partie des espèces qui, comme Peltoceras marysae et alii, ont colonisé la plate-forme nord-ouest européenne à la faveur de son "ennoiement" généralisé à la base du Callovien supérieur.
Enfin, si le devenir de Choffatia isabellae est difficile à connaître sur le seul matériel de Montreuil-Bellay, on peut quand même constater que la morphologie et l'ornementation de certains variants (plutôt marginaux) se retrouveront chez des formes plus récentes, attribuées par certains auteurs au genre Alligaticeras et que l'enroulement initial polygonal se retrouvera chez de nombreux Perisphinctidae oxfordiens.
Nous tenons à remercier toutes les personnes qui ont autorisé le travail de terrain : M. , Maire de Montreuil-Bellay, M. , directeur de la Covipac (zone industrielle de Méron), M. (société Giuntoli), MM. et (Société E.H.T.P.), MM. et à Méron, M. à la Motte-Bourbon. Que soient également remerciés M. à la pelle mécanique, ainsi que Didier, Yvon, Philippe et Louis pour leur participation aux fouilles, et les ouvriers de la zone industrielle, pour leur accueil souvent bienveillant. Nous sommes redevables à Carmela et à Nestor pour la version anglaise du résumé. Enfin, nous remercions les rapporteurs, Francois , Robert et József , pour leurs remarques et conseils judicieux qui ont fait progresser le manuscrit.
P.-A. (1999).- Biochronologie et dynamique évolutive des Distichoceratinae (Ammonitina, Callovien).- Thèse nouveau Doctorat, Université de Poitiers, inédit, 324 p.
A. (1993).- Les Peltoceratinae (Ammonoidea) de la sous-zone à Trezeense (Zone à Athleta, Callovien supérieur) en Côte d'Or (France).- Géobios, Villeurbanne, vol. 36, n° 2, p. 135-160.
A. (1995).- Les Aspidoceratidae d'Europe occidentale au Callovien supérieur et à l'Oxfordien inférieur.- Thèse nouveau Doctorat, Université Dijon, inédit, 487 p.
A. (1996).- Découverte d'une nouvelle espèce du genre Euaspidoceras (Ammonitina, Aspidoceratidae) dans l'horizon à Collotiformis (Callovien supérieur, zone à Athleta, sous-zone à Collotiformis) de Montreuil-Bellay (France, Maine-et-Loire).- Annales de Paléontologie, Paris, vol. 82, n° 2, p. 117-139.
A., P.-Y. & P. (2005).- Peltoceras marysae, une nouvelle espèce de Peltoceratinae (Ammonitina, Aspidoceratidae) dans l'horizon à Leckenbyi (Callovien supérieur, zone à Athleta) de Montreuil-Bellay (Maine-et-Loire, France).- Revue de Paléobiologie, Genève, vol. 24, n° 1, p. 75-95.
J. & D. (1968).- Données nouvelles sur la stratigraphie et la paléogéographie du Callovien du Jura franc-comtois.- Bulletin de la Société géologique de France, Paris, (7e Série), t. X, n° 3, p. 362-369.
P.-Y. (2000).- Environnements, géochimie et processus de formation de séries condensées au Callovo-Oxfordien : du Bassin de Paris à des considérations globales.- Thèse nouveau Doctorat, Université Dijon, inédit, 282 p.
D., A. & D. (1989).- Le passage Dogger-Malm en bordure de la chaîne de l'Heute (Jura français).- Annales scientifiques de l'Université de Besançon, (4e Série), n° 9, p. 17-25.
P. & A. (1998).- Faunes ammonitiques et biochronologie de la zone à Athleta et de la base de la zone à Lamberti (Callovien supérieur) de la Côte de Meuse (France). Intérêt des nouvelles faunes d'Aspidoceratidae.- Revue de Paléobiologie, Genève, vol. 17, n° 2, p. 307-346.
P., A., P.-Y., D. & D. (1998).- Coupures morphologiques et biochronologie chez les Kosmoceratinae de l'Est de la France (Callovien inférieur pp. à Callovien supérieur pp.).- Comptes rendus de l'Académie des Sciences, Paris, t. 327, n° 10, p. 685-691.
B.-M. (1988).- English Callovian (Middle Jurassic) Perisphinctid ammonites: Part I.- Monograph of the Palaeontographical Society, London, n° 575, vol. 140, 54 p.
C. & H. (1936).- Les ammonites de la zone à Peltoceras athleta du Centre-Ouest de la France.- Mémoires de la Société géologique de France, Paris, n° 29, (N.S.), 100 p.
A. (1951).- Stratigraphie und Paläontologie des oolithischen Eisenerzlagers von Herznach und seiner Umgebung.- Beiträge zur Geologie der Schweiz, Bern, (Geotechnische Serie), Lieferung XIII-5, 240 p.
Z. (1983).- Upper Callovian Ammonites and Middle Jurassic history of the Middle East.- Geological Survey of Israël, Bulletin, Jerusalem, vol. 76, 56 p.
C. (1970).- Les Perisphinctidae (Ammonitina) du Jura méridional au Bathonien et au Callovien.- Documents des laboratoires de géologie de la Faculté des Sciences de Lyon, Villeurbanne, vol. 41, n° 2, 246 p.
D. & J. (1997). - Enregistrements des variations morphologiques et de la composition des peuplements d'ammonites durant le cycle régressif/transgressif de 2e ordre Bathonien inférieur-Oxfordien inférieur en Europe occidentale-. Bulletin de la Société géologique de France, Paris, t. 168, n° 2, p. 121-132.
Cliquer sur la vignette pour agrandir l'image.
Figure 1 : Données géographique, lithologique, stratigraphique et faunique des coupes de la zone industrielle de Méron (Maine-et-Loire). A : localisation des coupes ; B : lithologie et biostratigraphie ; C : faune ammonitique totale ; D : les Perisphinctidae dans la faune ammonitique totale.
Cliquer sur la vignette pour agrandir l'image.
Figure 2 : Sections de spécimens macroconques et microconques adultes (x1). Macroconques 1 : spécimen Mbe/n/221 (holotype) ; 2 : spécimen I20 ; 3 : spécimen C40 ; 4 : Mbe/n/223 ; Microconques 5 : spécimen Mbe8700 ; 6 : spécimen Mbe0135 ; 7 : spécimen L53.

Cliquer sur la vignette pour agrandir l'image.
Figure 3 : Enroulement et ornementation initiaux de Choffatia isabellae macroconque. Barre = 1 cm. 1-2 : Stade lisse très long puis costulation initiale discrète – Enroulement initial peu perturbé (1 : spécimen Mbe9721 ; 2 : spécimen A11). 3-4 : Stade lisse court, puis costulation forte – Enroulement initial fortement perturbé (3 : spécimen I11 ; 4 : spécimen Mbe0217).
Cliquer sur la vignette pour agrandir l'image.
Figure 4 : Courbes de croissance des macroconques. O : ombilic ; H : hauteur ; E : épaisseur ; D : diamètre ; r : coefficient de corrélation.
Cliquer sur la vignette pour agrandir l'image.
Figure 5 : Courbes de costulation des macroconques. a : spécimen Mbe/n/223 ; b : spécimen C40 ; c : spécimen Mbe/n/221 (holotype) ; d : spécimen Mbe9411 ; e : spécimen H6 ; f : spécimen Mbe/n/2218. N/2 : nombre de côtes par demi-tour ; D : diamètre.
Cliquer sur la vignette pour agrandir l'image.
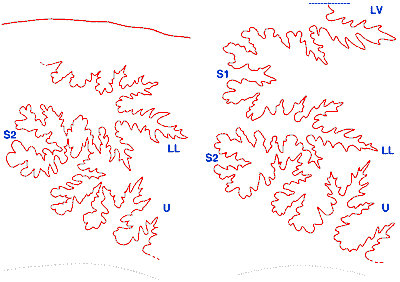
Figure 6 : Lignes de suture des macroconques (d'après photos numériques). a : spécimen I20, ligne de suture complète. Trait bleu = axe de la région ventrale ; b : spécimen Zi/70/n/139, ligne de suture pars. LV : lobe ventral ; LL : lobe latéral ; U : lobe suspensif ; S1 : première selle latérale ; S2 deuxième selle latérale.
Cliquer sur la vignette pour agrandir l'image.
Figure 7 : Répartition du diamètre (D) des microconques adultes (N=158).

Cliquer sur la vignette pour agrandir l'image.
Figure 8 : Péristome et apophyse (spécimen Mbe9201).

Cliquer sur la vignette pour agrandir l'image.
Figure 9 : Enroulement et ornementation initiaux de Choffatia isabellae microconque. Observer l'enroulement triangulaire (1, 2, 4) ou subquadrangulaire (3) rythmé par les constrictions, et la faiblesse de l'ornementation, avec potentiellement des côtes ventrales sans côtes ombilicales (2). 1 : spécimen Mbe9900 ; 2 : spécimen Mbe0229 ; 3 : spécimen Mbe0135 ; 4 : spécimen Mbe9814.
Cliquer sur la vignette pour agrandir l'image.
Figure 10 : Courbes de croissance des microconques. O : ombilic ; H : hauteur ; E : épaisseur ; D : diamètre ; r : coefficient de corrélation.

Cliquer sur la vignette pour agrandir l'image.
Figure 11 : Paraboles ventrales.
Cliquer sur la vignette pour agrandir l'image.
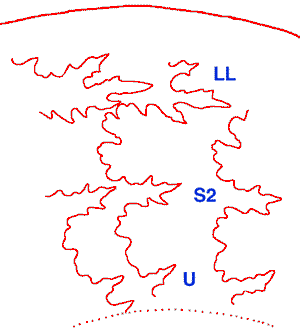
Figure 12 : Ligne de suture du microconque (d'après photo numérique). LL : lobe latéral ; U : lobe suspensif ; S2 deuxième selle latérale.
Prises de vue et montage des planches photos : A. .
L'holotype et les spécimens référencés UBGD sont conservés au Centre des Sciences de la Terre de Dijon.
Les spécimens des collections Pierre-Yves (PYB) et Patrice (PF) sont conservés et visibles chez les auteurs.
Tous les spécimens sont d'âge Callovien supérieur : zone à Athleta, sous-zone à Trezeense, horizon à Leckenbyi. Coupes de la zone industrielle de Méron (Maine-et-Loire).

![]()
Cliquer sur la vignette pour agrandir l'image.
Fig. 1 : Subgrossouvria famulum () macroconque adulte. Observer les côtes radiales, l'écartement et l'épaississement progressifs des côtes, le stade lisse précédant le péristome, les constrictions peu fréquentes sur le phragmocône et absentes sur la chambre d'habitation. Spécimen A16, collection PF. D = 121 mm.
Fig. 2 : Subgrossouvria famulum () microconque adulte avec les apophyses. Observer la similitude de l'ornementation des tours initiaux et l'absence de constrictions sur la chambre d'habitation. Spécimen I2, collection PF. D = 49 mm.
Fig. 3 : Orionoides pseudorion (), nucléus. Observer le recouvrement fort, l'ombilic profond, les côtes dichotomes ou polyschizotomes flexueuses, les constrictions rares et flexueuses. Spécimen UGBD276401. D = 48 mm.
Fig. 4 : Subgrossouvria crassa et . Observer le changement brutal de style ornemental après une constriction, les côtes et les constrictions radiales, la forte croissance en épaisseur. Spécimen Mbe9329, collection PYB. D = 97 mm.

![]()
Cliquer sur la vignette pour agrandir l'image.
Planche 2 : Choffatia isabellae n. sp. (Macroconques).
Fig. 1 : Macroconque évolute probablement adulte, à tours internes graciles peu ornés, puis à costulation plus forte. Spécimen Mbe9721, collection PYB. D = 82 mm.
Fig. 2 : Nucléus à costulation gracile. Spécimen C24, collection PF. D = 67 mm.
Fig. 3 : Macroconque évolute probablement adulte, à stade lisse court et côtes primaires fortes sur la chambre d'habitation. Spécimen Mbe0217, collection PYB. D = 90 mm.
Fig. 4 : Juvénile. Spécimen I17, collection PF. D = 52 mm.
Fig. 5 : Nucléus évolute à stade lisse court et à costulation forte. Observer les premiers tours à enroulement subquadrangulaire. Spécimen I11, collection PF. D = 46 mm.
Fig. 6 : Macroconque probablement adulte, à croissance en hauteur plus forte et à costulation gracile. Spécimen G20, collection PF. D = 90 mm.
Fig. 7 : Macroconque à costulation fine, proverse et régulière. Spécimen A11, collection PF. D = 71 mm.

![]()
Cliquer sur la vignette pour agrandir l'image.
Planche 3 : Choffatia isabellae n. sp. (Macroconques).
Fig. 1 : Macroconque adulte avec son péristome. Observer le stade lisse, les côtes secondaires qui persistent jusqu'à la fin de la croissance, la rareté des constrictions dans les tours moyen et externe. Spécimen H6, collection PF. D = 124 mm.
Fig. 2 : Macroconque adulte avec son péristome. Holotype. Observer les côtes relativement fortes et l'effacement de la costulation secondaire après la dernière constriction. Spécimen Mbe/n/221, collection PYB. D = 129 mm.

![]()
Cliquer sur la vignette pour agrandir l'image.
Planche 4 : Choffatia isabellae n. sp. (Macroconques).
Fig. 1 : Macroconque adulte avec son péristome. Observer la fréquence des constrictions et la costulation moyenne persistant jusqu'à la fin de la croissance. Spécimen Mbe/n/217, collection PYB. D = 107 mm.
Fig. 2 : Macroconque adulte avec son péristome. Observer le long stade lisse suivi d'une costulation ténue, mais qui persiste jusqu'à la fin de la croissance. Spécimen Mbe/n/218, collection PYB. D = 114 mm.

![]()
Cliquer sur la vignette pour agrandir l'image.
Planche 5 : Choffatia isabellae n. sp. (Macroconque).
Fig. 1 : Macroconque adulte avec son péristome (visible sur l'autre face). Le plus grand spécimen de l'échantillon (200 mm). Spécimen Mbe/n/223, collection PYB. D = 207 mm.

![]()
Cliquer sur la vignette pour agrandir l'image.
Planche 6 : Choffatia isabellae n. sp. (Macroconques).
Fig. 1 : Macroconque adulte. Observer la croissance en hauteur faible, la fréquence des constrictions dans les tours internes et moyens et la costulation assez forte qui s'estompe avant l'ouverture. Spécimen Mbe9411, collection PYB. D = 146 mm.
Fig. 2 : Macroconque adulte de petite taille. Observer la costulation forte qui s'interrompt au niveau d'une forte constriction et la dernière constriction précédant immédiatement le péristome. Spécimen UGBD276402. D environ 130 mm.

![]()
Cliquer sur la vignette pour agrandir l'image.
Planche 7 : Choffatia isabellae n. sp. (Macroconque).
Fig. 1 : Macroconque adulte avec son péristome. Observer l'enroulement très serpenticone (croissance faible en hauteur), la costulation forte, la disparition de la costulation secondaire suite à une constriction et la dernière constriction précédent de peu le péristome. Spécimen C40, collection PF. D = 158 mm.

![]()
Cliquer sur la vignette pour agrandir l'image.
Planche 8 : Choffatia isabellae n. sp. (Microconques).
Fig. 1 : Microconque adulte de très petite taille avec les apophyses complètes. Spécimen J4. Collection PF. D = 23 mm.
Fig. 2 : Microconque adulte avec les apophyses en construction. Observer la faiblesse de la costulation primaire et l'ornementation ventrale très ténue. Spécimen Mbe0137, collection PYB. D = 26 mm.
Fig. 3 : Microconque adulte avec les apophyses presque complètes. Observer l'ornementation primaire et secondaire forte. Spécimen G3, collection PF. D = 26,5 mm.
Fig. 4 : Microconque adulte avec les apophyses complètes. Observer l'ornementation primaire ténue et les côtes secondaires plus marquées. Spécimen J33, collection PF. D = 31 mm.
Fig. 5 : Microconque adulte mince à ornementation primaire très faible et côtes secondaires fortes. Spécimen C11, collection PF. D = 42 mm.
Fig. 6 : Microconque adulte avec les apophyses complètes. Observer l'ornementation primaire moyennennement forte, dichotome, et l'enroulement légèrement scaphitoďde de la chambre d'habitation. Spécimen A4, collection PF. D = 39 mm.
Fig. 7 : Microconque adulte avec les apophyses en construction. Observer l'ornementation presqu'inexistante. Spécimen Mbe0218, collection PYB. D = 34 mm.
Fig. 8 : Microconque adulte avec les apophyses en construction. Observer la fréquence des constrictions, la costulation primaire presqu'inexistante dans les tours internes. Spécimen Mbe0229, collection, PYB. D = 42 mm.
Fig. 9 : Microconque adulte avec les apophyses presque complètes. Observer la costulation moyennement forte. Spécimen Mbe2011, collection PYB. D = 43,5 mm.
Fig. 10 : Microconque adulte avec les apophyses cassées. Observer la costulation primaire très faible et l'affaiblissement de la costulation secondaire après une constriction "multiple". Spécimen L53, collection PF. D = 43,5 mm.

![]()
Cliquer sur la vignette pour agrandir l'image.
Planche 9 : Choffatia isabellae n. sp. (Microconques).
Fig. 1 : Microconque adulte avec les apophyses presque complètes. Observer la fréquence des constrictions, le long stade lisse et l'ornementation ténue. Spécimen Mbe9900, collection PYB. D = 50 mm.
Fig. 2 : Microconque adulte avec les apophyses complètes. Observer l'affaiblissement de l'ornementation en fin de croissance. Spécimen G18, collection PF. D = 57 mm.
Fig. 3 : Microconque adulte avec les apophyses complètes. Observer la costulation. Spécimen Mbe9814, collection PYB. D = 66 mm.
Fig. 4 : Microconque adulte avec les apophyses complètes. Observer les côtes légèrement flexueuses, un premier affaiblissement de la costulation après l'avant-dernière constriction et un second affaiblissement après la dernière constriction. Spécimen Mbe0135, collection PYB. D = 69 mm.

![]()
Cliquer sur la vignette pour agrandir l'image.
Planche 10 : Choffatia isabellae n. sp. (Microconques).
Fig. 1 : Microconque adulte avec les apophyses cassées. Observer l'affaiblissement de la costulation après la dernière constriction. Spécimen Mbe/n/57, collection PYB. D = 69 mm.
Fig. 2 : Microconque adulte avec les apophyses cassées. Observer la faible épaisseur et la constriction "double" à la fin de la loge d'habitation, après laquelle la costulation reste assez forte. Spécimen F17, collection PF. D = 69 mm.
Fig. 3 : Microconque adulte avec les apophyses cassées. Observer l'affaiblissement de la costulation sur la fin de la chambre d'habitation. Spécimen UGBD276403. D = 77,5 mm.
Fig. 4 : Microconque adulte avec les apophyses cassées. C'est le plus grand spécimen de l'échantillon (85 mm). Observer l'enroulement légèrement scaphitoďde de la chambre d'habitation. Spécimen Mbe8700, collection PYB. 84,5 mm.