![]()
Sommaire
[1. Introduction] [2. Coupe
géologique ...]
[3. Résultats ...]
[4. Systématique ... Gilianelles]
[5. ... autres microproblematica]
[6. Discussion ...]
[7. Conclusion] [Références bibliographiques]
[Figures] [Planches] [Annexe 1] et ... [Annexe 2]
Unité de Géochronologie et Sédimentologie Océanique, Département
de Géologie Sédimentaire,
Université Pierre et Marie Curie, Case 119, 4 Place Jussieu,
F-75252, Paris Cedex 05 (France)
URL: http://www-sst.unil.ch/research/meta/GSO/
Manuscrit soumis le 6 Mars 2008, accepté le 10 Mars, en ligne depuis le 22 Mars
![]()
L'étude des microfossiles extraits par acétolyse des niveaux carbonatés indurés de la série campano-maastrichtienne du stratotype de limite à Tercis (Landes, France) a permis de découvrir 44 taxons attribués au groupe des gilianelles (microproblematica appartenant probablement aux Protistes, Protozoaires, Rhizopodes) ainsi que dix autres microproblematica. 281 vues au microscope électronique et 183 vues au microscope optique permettent d'illustrer, chez les gilianelles, 36 espèces, et 8 sous-espèces appartenant à 15 genres dont 14 nouveaux. Les nouveaux genres sont : Tercensella, Azymella, Numismella, Aturella, Scutellella, Corniculum, Caccabella, Orculiella, Pennigerella, Corbella, Aquilegiella, Pocillella, Coraliella, Obbella. Chez les autres microproblematica, neuf espèces et une sous-espèce sont proposées appartenant à sept genres nouveaux : Globulella, Piperella, Vasculum, Lucernellus, Cimicellus, Tubella, Pilella. Cette première revue d'ensemble avec identification des taxons a conduit à caractériser précisément un groupe dont l'identité et la cohérence sont établies d'après les caractères morphologiques du corps des tests et des expansions qu'ils portent et leur ornementation. Ces critères, utilisés de façon ordonnée permettent de proposer un classement rationnel qui pourra être confirmé par d'autres observations pour devenir une classification. L'étude de la répartition stratigraphique indique qu'ils constituent un remarquable outil de datation relative dans le site stratotypique et son voisinage immédiat oů ils permettent d'individualiser des intervalles de dépôt de l'ordre de 0,1 Ma, plus fins que la plupart de ceux obtenus d'après les nombreux groupes de microfossiles ou de macrofossiles classiques étudiés pour qualifier ce stratotype. Le repérage de la limite Campanien-Maastrichtien peut se faire grâce à la lignée des aturelles ; elle se situe au début de la zone d'existence de l'aturelle édentée (Aturella edentula).
Gilianelles ; microproblematica ; systématique ; Crétacé ; stratotype ; Campanien ; Maastrichtien ; Aquitaine.
G.S. (2008).- Description et illustration de quarante-quatre gilianelles (microproblematica) et de dix autres microproblematica du Crétacé de la coupe stratotypique de Tercis (limite Campanien-Maastrichtien), Sud-Ouest France.- Carnets de Géologie / Notebooks on Geology, Brest, Mémoire 2008/01 (CG2008_M01)
Description and illustration of forty-four gilianelles (microproblematica) and ten other microproblematica of the Cretaceous of the stratotype section at Tercis (Campanian-Maastrichtian boundary), South-West France.- Following acetolysis, microproblematica were collected from indurated carbonates sampled from the stratotype section of the Campanian-Maastrichtian boundary at Tercis (Landes, France). 44 taxa of the gilianelles group (microproblematica probably pertaining to the Protista, Protozoans, Rhizopods) as well as ten other microproblematica were discriminated. To document the taxa in this paper 281 scanning electronic microscope images and 183 optical microscope images are presented. The gilianelles encompass 36 species, and 8 subspecies referred to 15 genera of which 14 are new. The new genera are: Tercensella, Azymella, Numismella, Aturella, Scutellella, Corniculum, Caccabella, Orculiella, Pennigerella, Corbella, Aquilegiella, Pocillella, Coraliella, Obbella. Among the other microproblematica, nine species and one subspecies are proposed; they are referred to 7 new genera: Globulella, Piperella, Vasculum, Lucernellus, Cimicellus, Tubella, Pilella. This first general view uses normal taxonomic criteria to recognize and define a group of microfossils of which the identity and the consistency of occurrence are herewith established using the morphological characters of the body of the test, the presence and development of the expansions attached to that body, and its ornamentation. The use of these criteria in an ordered manner suggests that a rigorous taxonomic collocation is feasible, needing only a few other observations before becoming a formal classification. The statigraphic distribution of the taxa provides a means for establishment of a remarkable tool for relative dating in the stratotype section and its immediate vicinity ; microproblematica allow the sequence to be subdivided into intervals of deposition on the order of 0.1 Ma, a length of time shorter than the intervals obtained using the many microfossil or macrofossil groups previously employed to calibrate the stratotype. The Campanian-Maastrichtian boundary is within the limits of the genus Aturella lineage in the basal portion of the range of Aturella edentula.
Gilianelles; microproblematica; systematics; Cretaceous; stratotype; Campanian; Maastrichtian; Aquitaine.
Afin de compléter les connaissances rassemblées sur le stratotype de limite Campanien-Maastrichtien à Tercis (Aquitaine méridionale, ,
2001a) et notamment pour les niveaux indurés, une technique de dissociation par acétolyse (traitement par l'acide acétique concentré) a été expérimentée. En variant température, concentration de l'acide et durée de traitement, les 39 niveaux sélectionnés ont pu être dissociés. La plupart a livré une microfaune diversifiée de qualité. La plus remarquable observation a été celle de formes énigmatiques dans la fraction de taille comprise entre 100 et 250 µm. Des dessins en 3 dimensions ![]() de quelques-unes de ces formes furent soumises à diverses personnalités du monde micropaléontologique directement puis par le biais de la liste de diffusion PaleoNet. Aucune proposition de rattachement à un groupe connu ne correspondait aux informations rassemblées lors des multiples observations optiques. Ce sujet est devenu une de nos préoccupations principales avec deux objectifs : Contribuer à améliorer la connaissance de la biodiversité crétacée et enrichir nos outils stratigraphiques.
de quelques-unes de ces formes furent soumises à diverses personnalités du monde micropaléontologique directement puis par le biais de la liste de diffusion PaleoNet. Aucune proposition de rattachement à un groupe connu ne correspondait aux informations rassemblées lors des multiples observations optiques. Ce sujet est devenu une de nos préoccupations principales avec deux objectifs : Contribuer à améliorer la connaissance de la biodiversité crétacée et enrichir nos outils stratigraphiques.
Dans un premier temps, un examen approfondi de ces microproblematica a montré qu'une majorité présentait suffisamment de points communs -et originaux dans leur ensemble- pour proposer la création d'une nouvelle unité taxinomique de rattachement incertain (, 2007).
Une espèce type (Gilianella stellata , 2007) fut nommée et illustrée pour ce groupe baptisé gilianelles caractérisé par un "test carbonaté, uniloculaire, de taille régulièrement comprise entre 70 et 200 µm (...) composé d'un corps à symétrie axiale fondamentale, parfois modifiée en une symétrie radiaire par le développement d'expansions rayonnantes" (, 2007 ; N.B. accessible en ligne dès le 27-XI-2006, puis en version papier à partir de III-2007).
À la suite de cette proposition, la modélisation de 36 formes a été entreprise ainsi que des compléments d'examen au microscope électronique à balayage ( et , 2006 ; N.B. accessible en ligne dès le 14-XII-2006). Ces nouvelles études ont conduit à préciser la morphologie de l'ensemble des microproblematica de Tercis mais aussi à compléter la définition du groupe particulier des gilianelles parmi eux. Les gilianelles révèlent au microscope électronique une nannostructure caractéristique de la paroi du test composée d'une couche préservée unique présentant un aspect de surface "en réseau" ; ce réseau rappelle, par sa variété comme par son dessin, les mailles des napperons réalisés au crochet.
Il manquait à ce groupe une concrétisation taxinomique de ces membres. C'est la question principalement abordée dans cette troisième publication sur les microproblematica du Crétacé de Tercis. La partie systématique illustre au moyen de vues réalisés en microscopie électronique à balayage, décrit et nomme 43 nouvelles formes. Toutes ces formes sont attribuées au groupe des gilianelles tel que typifié par Gilianella stellata , 2007, dont l'illustration et la variété morphologique sont complétées ici. Afin de caractériser mieux encore ce groupe des gilianelles, 10 autres microproblematica sont illustrés, décrits et nommés qui permettent de faire ressortir les caractères propres aux gilianelles.
Le stratotype de la limite Campanien-Maastrichtien a été défini dans une carrière abandonnée située sur le territoire de la commune de Tercis, près de Dax, département des Landes, en Aquitaine méridionale, à une dizaine de km au Nord des premiers charriages nord-pyrénéens (, 2001). Ce site, devenu une référence mondiale protégée, abrite un affleurement lié à un anticlinal diapirique faisant remonter à la surface, des couches crétacées situées aux alentours sous 2 km de couverture tertiaire. Malgré cette tectonique, l'ordonnancement des couches n'a pas été atteint et une coupe continue de près de 200 m a été décrite en détail ( et , 1994). Elle représente à peu près 8 Ma de dépôts entourant la limite Campanien-Maastrichtien définie par convention à la cote 115,2 à la suite d'une approche originale et potentiellement fructueuse pour les corrélations mondiales. En effet, cette définition implique la combinaison de 12 signaux biostratigraphiques pris dans 6 groupes fossiles et entourant un événement guide accepté par convention (, 2001b).
Au Crétacé Supérieur, le site appartenait au bassin aturien ( et alii, 2001). Ce diverticule latitudinal de l'Atlantique relevait, par sa faune et son faciès, du domaine téthysien comme les faciès contemporains de Charente ( et , 2001), région oů fut choisi le stratotype historique du Campanien. Ce diverticule occupait l'emplacement des futures Pyrénées ; au Nord de ce bassin, sur le seuil landais, se déposait le faciès aturien ( et alii, 2001) composé de carbonates de plate-forme à l'abri des apports détritiques. Ce sont les calcaires à pithonelles qui caractérisent l'ensemble de la coupe échantillonnée ici ( et alii, 2001). Les pithonelles sont des restes de nature controversée ( et alii, 2001), sphériques, de 40 à 200 µm de diamètre et comprennent divers taxons. Au même moment et un peu plus au Sud, se déposaient des flyschs, produits de l'érosion de l'esquisse des Pyrénées.
Les calcaires à pithonelles sont caractérisés par la présence d'une grande variété de fossiles planctoniques, benthiques, macro- et microfossiles dont 18 groupes furent examinés et illustrés (, 2001a ; et alii, 2006, 2007) ce dernier travail note la présence des microproblematica (affiche g).
La lithologie de la coupe est schématisée dans la Figure 1 ![]() qui identifie 3 unités ( et ,
1994). L'Unité Hontarède est constituée de calcaires à silex glauconieux au sommet. L'Unité d'Avezac est une succession homogène de calcaires dont les 5 sous-unités (A1 à A5) ne se distinguent que par des teneurs en glauconie alternativement plus élevées et quasi nulle. L'Unité Les Vignes comprend à la base une sous-unité à silex clairs (V1) et, au sommet une unité à silex foncés (V2). La colonne lithologique note la présence de glauconie (pointillés), et celle de silex presque toujours en niveaux décimétriques bien séparés avec une périodicité métrique.
qui identifie 3 unités ( et ,
1994). L'Unité Hontarède est constituée de calcaires à silex glauconieux au sommet. L'Unité d'Avezac est une succession homogène de calcaires dont les 5 sous-unités (A1 à A5) ne se distinguent que par des teneurs en glauconie alternativement plus élevées et quasi nulle. L'Unité Les Vignes comprend à la base une sous-unité à silex clairs (V1) et, au sommet une unité à silex foncés (V2). La colonne lithologique note la présence de glauconie (pointillés), et celle de silex presque toujours en niveaux décimétriques bien séparés avec une périodicité métrique.
La stratigraphie en âges numériques est proposée d'après 1- des corrélations magnéto- et biostratigraphiques avec des niveaux de bentonites datés aux États Unis ( et , 2001), 2- une vitesse de sédimentation constante illustrée par une périodicité orbitale des dépôts, 3- une combinaison avec les observations liées à la présence et à la cristallinité des minéraux glauconitiques. Ceci conduit à considérer que chaque mètre de dépôt représente en moyenne 40.000 ans ( et , 2001). Une estimation de l'âge de la limite a été proposée lors de l'étude coopérative de la coupe (, 2001d) ; cette estimation a été améliorée par la suite grâce aux corrélations Europe-Amérique plus précises établies grâce à l'étude des inocérames communs aux deux ensembles ( et alii, 2002 ; et , 2003). Ces âges s'entendent, pour une limite à 71,6 Ma, avec une incertitude de ±0,6 Ma (intervalle de confiance 95 %).
Une quarantaine de niveaux ont été étudiés dans la coupe du stratotype (flèches aux cotes respectives) et en dehors dans 3 niveaux du Maastrichtien supérieur. Les acétolyses ont été adaptées en fonction du comportement des sédiments. Aucune recommandation générale ne peut être proposée pour ces traitements sinon qu'il convient de ne pas se contenter d'un essai. Après 1 ou 2 essais, le tâtonnement permet de choisir des conditions favorables de temps (quelques heures à quelques semaines), de température (20 à 90°C) et de concentration (80 à 99%) susceptibles de créer une dissociation suffisante pour isoler les restes d'origine organique sans les altérer trop fortement. Des quantités de sédiment allant de 100 à 300 g ont été traitées. La richesse en microproblematica est telle qu'il a généralement suffi d'examiner une petite portion de la fraction granulométrique favorable obtenue (100 à 250 µm) pour isoler des formes variées.
L'objectif initial de la recherche était de caractériser les niveaux de l'intervalle d'existence du foraminifère Radotruncana calcarata (, 1927) qui, dans la coupe de Tercis, s'est trouvé coďncider avec des indices de volcanisme (, 2001c). Ceci explique le nombre de prélèvements traités entre les cotes 40 et 62 oů le foraminifère a été illustré ou suspecté en lames minces ( et , 2001 ; et alii, 2001). Ensuite, des traitements ont été entrepris systématiquement tous les 20 mètres hors de cet intervalle pour vérifier la répartition des formes identifiées. Enfin, lorsque la recherche était particulièrement fructueuse (variété des formes énigmatiques), un troisième groupe de niveaux intermédiaires a été examiné pour compléter l'étude. Pour concrétiser la finesse de l'enregistrement paléontologique ainsi mis en évidence, il suffit de rappeler qu'un intervalle d'échantillonnage de 10 m représentent une durée de dépôt de l'ordre de 0,5 Ma et que les meilleurs marqueurs de temps ont une durée de vie ne dépassant pas 1 Ma ; ils peuvent alors n'être présents que dans un seul niveau examiné en détail.
La découverte tardive de ces formes énigmatiques pourrait laisser penser que leur rareté est en cause. Un résultat significatif de cette étude est qu'il n'en est
rien. Les sédiments de Tercis sont très riches dans la fraction granulométrique considérée. La
Figure 1 ![]() donne (colonne de nombres à droite notée spcmn/g) des estimations qui vont de quelques dizaines à quelques centaines d'individus observés par gramme de sédiment traité. Concrètement, une centaine d'individus peut parfois être observée dans un seul trieur de 50 cm2 saupoudré de telle sorte que les grains se touchent à
peine. L'abondance des microproblematica paraît liée à celle des pithonelles des mêmes niveaux ; elle est surtout remarquable dans les 4 sous-unités inférieures de l'Unité
d'Avezac.
donne (colonne de nombres à droite notée spcmn/g) des estimations qui vont de quelques dizaines à quelques centaines d'individus observés par gramme de sédiment traité. Concrètement, une centaine d'individus peut parfois être observée dans un seul trieur de 50 cm2 saupoudré de telle sorte que les grains se touchent à
peine. L'abondance des microproblematica paraît liée à celle des pithonelles des mêmes niveaux ; elle est surtout remarquable dans les 4 sous-unités inférieures de l'Unité
d'Avezac.
La variété (colonne de nombres à gauche notée nb. esp) suit généralement l'abondance. Dans le Maastrichtien, cependant, même une abondance élevée de 160 spcmn/g (à la cote 172) ne correspond qu'à une variété de 10 formes. Au contraire, vers la base de la coupe, au niveau -5,0 et malgré une abondance faible (30 spécimens par gramme) la variété est élevée avec 11 formes.
Le plus souvent, les codes alphanumériques sont utilisés dans ce travail car ils sont plus faciles à manipuler et à noter, ils traduisent une morphologie générale repérable dès les tris optiques et ils rappellent les désignations utilisées seules lors des publications antérieures ; on trouvera, les désignations taxinomiques correspondantes en annexe 2.
La répartition stratigraphique des 29 taxons les plus significatifs est illustrée dans la
Figure 1 ![]() (cadre de droite) le long de la
coupe principale du site de Tercis. Cette coupe étant calibrée vis à vis du temps ( et ,
2001), il est possible d'estimer la durée de vie des divers taxons.
(cadre de droite) le long de la
coupe principale du site de Tercis. Cette coupe étant calibrée vis à vis du temps ( et ,
2001), il est possible d'estimer la durée de vie des divers taxons.
Chaque apparition, ou disparition peut être considérée comme un bio-événement pratique dès lors que l'abondance du taxon correspondant est suffisante pour être aisément décelable. Ont été laissées de côté, les formes trouvées en trop petit nombre pour être d'une utilité pratique, celles pour lesquelles une portion de la répartition est fondée sur la présence occasionnelle de peu de spécimens et enfin les 7 taxons panchroniques au niveau de la coupe type : G1, A1a, A1b, A2a, A2b, D1a et D1b. Enfin, du fait de leur large contemporanéité et proximité morphologique, 3 répartitions sont des combinaisons : les taxons E3a, E3b et E3c, J1 et J2a, M2a et M2b. Il résulte de cet exercice que deux ensembles de niveaux sont riches en formes variées : entre les cotes -5 et +25 d'une part et entre les cotes 47 et 62 de l'autre. Le premier ensemble renferme à lui seul 29 des 54 taxons décrits dans ce travail dont 28 sont présents à la seule cote 11,0. La variété observée entre les cotes 47 et 62 (26 taxons) est en partie liée à une investigation plus détaillée de ces couches.
La localisation des apparitions est une autre façon d'estimer le caractère favorable du milieu ayant présidé à certains dépôts par rapport à d'autres. On constate que la majorité des apparitions est concentrée dans 2 groupes de niveaux :
entre les cotes -5 et +5 et entre les cotes 47 et 55. Les taxons laissés de côté pour simplifier la
Figure 1 ![]() ne changent rien aux observations résumées ici.
ne changent rien aux observations résumées ici.
Les dépôts du Campanien supérieur, jusqu'à la cote 80 environ, correspondent donc à un environnement favorable à la présence des gilianelles. Au dessus, les sédiments n'ont livré que 10 formes au mieux, souvent peu abondantes. Aucun facteur unique ne semble univoquement lié à ces considérations. On peut toutefois évoquer quelques facteurs. Par exemple, la présence de glauconie en quantité significative cesse dans la coupe vers la cote 80 (, 2001c, Fig. 7, p. 100). Ainsi, la présence de glauconie et celle des gilianelles seraient sous la dépendance d'un facteur commun ; toutefois, on peut observer qu'à la cote 11,3 qui sépare les sous-unités A1 et A2, il existe aussi une coupure entre des niveaux glauconieux dessous et non glauconieux au dessus ; pourtant les gilianelles sont présentes de part et d'autre.
Par exemple encore, la reconstitution des profondeurs au moyen des échinides ( et , 2001, Fig. 1, p. 615) indique que le remplacement des micrasters dominants indiquant une profondeur comprise entre 50 et 100 m par les échinocorys caractérisant une profondeur comprise entre 100 et 150 m, se fait aussi entre les cotes 70 et 80 tandis que la raréfaction des échinocorys à leur tour, indicateur d'une profondeur dépassant 150 m se fait près de la cote 125. Toujours parmi les échinodermes, il y a une nette augmentation des restes de crinoďdes (ossicules) vers la cote 80. Souvent absents en dessous, ils sont communs à abondants au dessus jusqu'à la cote 172 ( et , 2001, Fig. 1, p. 637). De même, les restes macroscopiques d'astérides (ossicules vus sur le terrain), épisodiquement observés jusqu'à la cote 80 ont été récoltés de façon quasi-continue au dessus entre les cotes 81 et 172. Ainsi, l'abondance et la variété des gilianelles peut être reliée à la présence et à l'abondance relative de divers groupes d'échinodermes connus pour être sensibles aux variations environnementales sur la plate-forme. Nous n'en tirons pas d'autre conclusion que le fait que ces microproblematica, nouvellement découverts, sont très sensibles aux changements environnementaux perçus par ces organismes benthiques que sont les échinodermes. Les gilianelles apprécieraient un bassin dont les fonds seraient favorables aux micrasters sur une plate-forme relativement peu profonde du domaine téthysien crétacé, à l'abri des apports terrigènes.
Il existe deux sortes de taxons bien distincts quant à leur durée de vie stratigraphique parmi les microproblematica de Tercis. Ceux à longue persistance stratigraphique, au nombre de 7 comme écrit plus haut, et ceux ayant une durée de vie proche du Ma, voire moins
(Fig. 1 ![]() ) ; c'est une brièveté remarquable. On peut être assuré que cette brièveté est de nature évolutive lorsque les formes plus ou moins manifestement apparentées se succèdent dans le temps. C'est le cas des formes A4a, A4b, A4b' et A6 ou encore du couple C1a - C1b. Les gilianelles évoluaient donc rapidement sur les hauts fonds de la plate-forme du bassin aturien. Elles permettent d'établir des biorepères bien plus rapprochés dans le temps que ceux obtenus avec les groupes paléontologiques contemporains étudiés en 2001 (,
2001e).
) ; c'est une brièveté remarquable. On peut être assuré que cette brièveté est de nature évolutive lorsque les formes plus ou moins manifestement apparentées se succèdent dans le temps. C'est le cas des formes A4a, A4b, A4b' et A6 ou encore du couple C1a - C1b. Les gilianelles évoluaient donc rapidement sur les hauts fonds de la plate-forme du bassin aturien. Elles permettent d'établir des biorepères bien plus rapprochés dans le temps que ceux obtenus avec les groupes paléontologiques contemporains étudiés en 2001 (,
2001e).
En se limitant aux taxons dont l'abondance est significative et la morphologie bien distincte, on peut citer une douzaine de repères vers la base de l'Unité d'Avezac. Il est à supposer que ces "apparitions" ne sont qu'apparences liées à l'absence d'investigation dessous mais aussi au caractère défavorable du faciès sous-jacent généralement très pauvre en restes fossiles.
Les multiples disparitions (N1b, E3b, C1a, C2, A7, E2b, J2a, A3) entre les cotes 15 et 25 sont significatives ; entre les cotes 43 et 47, le passage de la forme A4a à la forme A4b est graduel ; entre les cotes 47 et 50, les apparitions de H2, F1, M2 (a et b), N2b et L2 signent un niveau repère bien marqué ; entre les cotes 60 et 69 se produisent encore 3 disparitions (F1, L2 et C1b), 2 apparitions (M3 et M1) et le passage de forme entre A4b et A4b' ; H2 et M1 disparaissent au dessus de la cote 79,8 tandis que B1a n'est présent que dans ce niveau riche en formes diversifiées. C'est plus haut, aux alentours de la limite d'étage qu'apparaissent les formes A5 et A6.
Les recherches originelles sur les sédiments du Campanien et du Maastrichtien n'avaient pas pour but de découvrir des espèces, des genres, voire des familles nouvelles de microfossiles calcaires mais d'approfondir la connaissance des microfaunes des niveaux indurés de la coupe type pour la limite entre ces deux étages du Crétacé. Ces formes problématiques furent rencontrées si communément qu'elles retenaient l'attention. Elles ont été désignées par un code alphanumérique, fonction de leur morphologie générale avec le souci de ne pas perdre l'information accumulée pendant les mois passés à trier les sédiments traités. L'ordre des découvertes, en grande partie lié à l'abondance des formes, a fait que A7 (très restreint dans le temps) et plus encore A8 (une forme très discrète) ont un chiffre plus élevé que A1 (souvent commun et présent partout). Il ne faut pas attribuer une logique taxinomique rigoureuse à ces codes qui ont été conservés parce qu'ils représentent une morphologie générale, parce qu'ils se notent aisément lors des tris et parce qu'ils ont été utilisés lors des publications antérieures.
Les majuscules de ces codes sont donc liées à une morphologie générale mais parmi les restes codifiés A figurent des gilianelles telles que définies au début de 2006 (, 2007) mais aussi d'autres problematica étrangers à ce groupe homogène (voir A5 ci-dessous). De même, on discute et illustre, ci-après, les raisons qui ont fait que les tests codifiés L2 ou L3 (rares) ont été classés parmi les gilianelles tandis que la forme L1 (très commune là oů elle est présente) ne paraît pas faire partie de ce groupe bien qu'ayant une morphologie générale étonnamment similaire (convergence).
Les gilianelles sont rassemblées dans un chapitre propre ; les "autres" microproblematica sont décrites ensuite. La description de ces autres formes a été jugée utile car elles permettent, par l'observation des différences, de mieux préciser l'unité du groupe des gilianelles ; en outre, ces restes énigmatiques complètent avantageusement les outils de caractérisation biostratigraphiques de la coupe, propos central de ces recherches.
Il est encore trop tôt pour formaliser une classification des gilianelles rencontrées à Tercis. L'ordre adopté comprend 6 catégories ordonnées de telle sorte qu'une complexité croissante se dégage.
1- Considérés comme primitifs, les taxons globuleux simples sont décrits d'abord.
2- Les disques simples suivent sans ou avec des collerettes.
3- Les cupules en entonnoir sont placées ensuite ; elles sont considérées comme dérivées de la catégorie 2.
4- Les disques à bras leur succèdent avec leur symétrie radiaire et comprenant, d'une part les formes symétriques dans le plan équatorial et d'autre part les formes à face orale plus développée.
5- Les cupules à deux rangs d'expansions et face orale lisse sont décrites ensuite ; dans ces formes, le développement de la face aborale permet de distinguer un flanc et une surface péri-apicale.
6- Enfin, les cupules à 2 rangs d'expansions et face orale crêtée radiairement constituent la morphologie la plus aboutie des gilianelles avec une ornementation élaborée à partir d'une microstructure surimposée à la nannostructure couvrant tout le corps.
Une classification sera formalisée lorsque les informations nannostructurales auront été complétées et comprises mais, dès à présent, il est possible d'exposer la variété morphologique des gilianelles avec une logique embrassant l'ensemble des formes connues à ce jour.
Les autres microproblematica, ensemble certainement hétérogène dans lequel différents ordres, différentes classes voire différents règnes peuvent être impliqués, sont décrits ensuite dans un ordre qui est celui de leur intérêt stratigraphique et non taxinomique.
L'attribution des microproblematica carbonatés décrits ci-dessous a été discutée (, 2007). Les diverses possibilités envisagées (pseudarcellidés, dinoflagellés à test calcaire, microcrinoďdes, radiolaires aux tests substitués en calcite) ont été réfutées et le nom de gilianelles proposé à un rang au moins égal à la famille, un ordre n'étant pas exclu. Quelques formes présentent certes des affinités morphologiques avec l'une ou l'autre espèce de l'un ou l'autre groupe cités voire avec certains foraminifères monoloculaires ( et , 2006). D'autres remarques seront faites plus loin lors de la description de chaque taxon mais l'ensemble des formes ne peut se rattacher à aucun groupe connu. En particulier, aucun ne présente systématiquement la nannostructure en napperon avec ses différents aspects en grille, radial, concentrique, ... et qui est le point commun à tous les tests carbonatés illustrés plus loin. Au delà de cette attribution, les considérations suivantes restent hypothétiques.
La taille des tests suggère qu'ils furent réalisés par une cellule unique. Dans ce règne des Protistes, l'appartenance à l'embranchement des Protozoaires est suggérée par la réalisation systématique de tests calcaires variés peu communs chez les Protophytes sinon chez les coccolithophoridés, par exemple, dont le squelette est constitué d'un assemblage de plaques ce qui n'est jamais le cas dans nos espèces. Parmi les Protozoaires, la classe des Rhizopodes est probable. En effet, on imagine facilement la présence de protoplasme autour des tests ; le rôle nourricier des pseudopodes caractéristiques des Rhizopodes serait facilité par le fait que de nombreux tests présentent des expansions permettant d'étendre le domaine protoplasmique et le volume d'eau nourricier pour les formes planctoniques notamment. Cette reconstitution a été illustrée ( et , 2006). Les petits tests calcaires, uniloculaires rapprochent les formes décrites de l'Ordre des Foraminifères. Des pores ou foramens -caractéristique des tests de cet ordre- ont en effet été observés à plusieurs reprises en microscopie électronique. Ainsi, les gilianelles pourraient constituer un sous-ordre des foraminifères caractérisé par un test carbonaté uniloculaire à nannostructure externe en napperon mais ils peuvent tout autant constituer un ordre différent et cet aspect de la question reste à approfondir.
Sur les 54 taxons illustrés ci-dessous, 36 ont déjà été présentés sous forme de dessin ( et , 2006). Ce dernier travail utilise des désignations simples alphanumériques qui sont reprises ici pour simplifier les citations. Les modifications suivantes ont été apportées : une forme petite, proche de H2 a été désignée ici H5 ; la forme précédemment codifiée J2 s'est révélée rassembler 2 morphologies distinctes qui ont été notées J2a et J2b lors des travaux suivants. De même, la forme initialement codée N1 a été subdivisée en N1a et N1b.
Le paragraphe suivant décrit, illustre et formalise les taxons qui s'ajoutent à l'espèce type du groupe déjà désignée et baptisée (, 2007).
Les formes nommées ici seront illustrées par 4 vues désignées comme orale, de profil, aborale et cavalière pour la vue de la face montrant l'orifice, celle prise à 90°, celle opposée à la face portant l'orifice et celle vue de façon oblique respectivement. Le plus souvent, 1 à 3 vues, complètent la documentation afin d'illustrer les variations ou la constance de l'aspect d'un spécimen à l'autre. L'illustration additionnelle tend à privilégier les cas exceptionnels ; le texte des descriptions précisera donc le caractère commun ou rare de tel ou tel aspect illustré relativement à la population observée. Pour la plupart, les taxons décrits ont été récoltés en grand nombre ; le compte n'en a pas été fait en détail ; une idée des populations observées est donnée à 3 niveaux : La répartition verticale, le nombre de topotypes (choisis parmi les plus représentatifs) et préservés dans la collection de référence avec les holotypes et l'abondance donnée dans le paragraphe correspondant de chaque description.
L'ordre des descriptions a suivi une ébauche de classement qui sera rappelé. Cet exercice sera formalisé lorsque l'ensemble des taxons sera décrit. La sélection des formes présentées résulte aussi de l'état d'acquisition de la documentation photographique, tâche pour laquelle nos facilités sont restreintes.
Les descriptions suivent une terminologie (, 2007) qui distingue face orale, face aborale et expansions diverses en bras (bord de la face orale) et pieds (face aborale). Une microstructure, plus ou moins visible à l'examen optique et comprenant des reliefs radiaux ou concentriques de rides ou crêtes est distinguée d'une nannostructure seulement visible au microscope électronique. Cette nannostructure de surface constitue une des caractéristiques clé du groupe des gilianelles : La nannostructure en napperon, laquelle est variée.
Tous les spécimens proviennent de la coupe type pour la limite Campanien-Maastrichtien de Tercis et leur localisation géographique commune a été décrite et illustrée de façon répétée (, 2001a ; et , 2001 ; et alii, 2006, 2007 ; et alii, 2002). Stratigraphiquement, ils sont proches mais le détail de la provenance est important car c'est parfois en considération de leur distribution dans le temps que des formes triées ont pu être distinguées. Un holotype a été choisi pour chaque taxon parmi une sélection de spécimens de qualité raisonnable récoltés au même niveau (les topotypes). Le plus souvent, l'holotype a été coloré pour l'identifier plus sűrement. Ces types dégagés, sont disposés dans des cellules sur lesquelles le code de l'espèce et la provenance sont notés avec un numéro d'inventaire. À l'exception de 4 holotypes désignés d'après leur observation au microscope électronique, tous les holotypes, paratypes et la majorité des topotypes ont été photographiés au microscope optique. Les vues informatisées, des tirages et les spécimens sont préservés avec l'ensemble de la collection de fossiles du stratotype de Tercis actuellement à l'Université Pierre et Marie Curie. Ces archives seront rassemblées dans un local muséologique en cours de réalisation près du site géologique (environs de Dax, Landes, France).
Groupe des Gilianelles (rang incertain, famille ou supérieur)
Référence : , 2007.
Espèce type : Gilianella stellata , 2007.
Description : Test monoloculaire, carbonaté, petit (75-200 µm), composé d'une couche de cristallites radiaires ; symétrie axiale à radiaire, parfois bilatérale avec polarité avant-arrière ; orifice unique, petit, distinguant une face orale d'une face aborale parfois différenciée en flanc et surface péri-apicale ; nannostructure de surface du test en napperon dont le réseau peut s'organiser en microstructures rayonnées ou concentriques ; présence commune d'expansions (bras, pieds, ou collerettes équatoriaux, aboraux ou péri-apicaux) dont la nannostructure est différente de celle de la paroi du test ; milieu de vie de plate-forme continentale marine planctonique ou benthique selon les espèces ; âge connu : Campanien et Maastrichtien. Le suffixe "ella" termine le nom de nombreux genres créés ici ; il tend à rappeler que ces formes sont petites pour des microfossiles.
Actuellement 15 genres décrits : Tercensella, Azymella, Numismella, Aturella, Scutellella, Corniculum, Gilianella, Caccabella, Orculiella, Pennigerella, Corbella, Aquilegiella, Pocillella, Coraliella, Obbella.
Genre Tercensella nov. gen.
Étymologie : Le nom de genre dérive du latin tercensis : de Tercis.
Espèce type : Tercensella globosa nov. sp.
Description : Corps subsphérique, aplati au moins sur la face orale ; de la taille d'une pithonelle ; ornementation nannostructurale en grille concentrique sur la face orale, méridienne sur les flancs ; en microscopie optique, la première est visible sous la forme de cercles concentriques oraux ; la deuxième sous la forme d'une quarantaine de cannelures périphériques en vue polaire.
Espèces attribuées au genre : globosa, doliolum, alveare.
Tercensella globosa nov. sp.
Synonymie : Code N2b.- et ,
2006, Pl. 6 ![]() .
.
Description : Corps sub-sphérique de diamètre 125 à 135 µm, aplati sur une face (Vue 2 ![]() ) ; face orale aplatie au pôle ; orifice axial situé au fond d'une dépression de 25 µm de diamètre (Vue 4
) ; face orale aplatie au pôle ; orifice axial situé au fond d'une dépression de 25 µm de diamètre (Vue 4 ![]() ) ; rides concentriques visibles à la loupe, autour de l'orifice (Vue 1
) ; rides concentriques visibles à la loupe, autour de l'orifice (Vue 1 ![]() ) ; flancs, arrondis, paraissant cannelés en vue orale ou aborale (Vue 3
) ; flancs, arrondis, paraissant cannelés en vue orale ou aborale (Vue 3 ![]() ) et rayés de profil ; nannostructure en mailles alignées concentriquement sur la face orale mais méridiennement sur les flancs ; arrangement encore modifié sur la face aborale (Vue 3
) et rayés de profil ; nannostructure en mailles alignées concentriquement sur la face orale mais méridiennement sur les flancs ; arrangement encore modifié sur la face aborale (Vue 3 ![]() ) sans qu'une limite nette ne soit apparente entre ces 3 espaces.
) sans qu'une limite nette ne soit apparente entre ces 3 espaces.
Étymologie : Le nom d'espèce dérive du latin globosus : sphérique ; c'est la tercisiennelle sphérique.
Holotype N2b : Provenant du niveau 54,9 l'holotype a été coloré (Planche annexe 1 ![]() , vignette 1) et inventorié au Musée du site de Tercis sous la référence TCS.G032 ; il est accompagné de 9 topotypes.
, vignette 1) et inventorié au Musée du site de Tercis sous la référence TCS.G032 ; il est accompagné de 9 topotypes.
Discussion : La forme du niveau d'oů provient l'holotype (Vues 1-4 ![]() ) est petite, sub-sphérique et paraît constante dans la
coupe lors de l'observation optique ; la morphologie de ce taxon évolue peut-être le long de la coupe : la Vue 5
) est petite, sub-sphérique et paraît constante dans la
coupe lors de l'observation optique ; la morphologie de ce taxon évolue peut-être le long de la coupe : la Vue 5 ![]() montre une forme légèrement plus grande pour le niveau sommital de la carrière (cote 172) ; les critères morphologiques sont trop simples et l'abondance souvent trop faible pour tenter avec assurance de détacher des subdivisions pour ce taxon défini à la cote 54,9 oů il est contemporain du foraminifère planctonique Radotruncana calcarata ; l'aplatissement du pôle oral par rapport à une sphère régulière, a pour effet d'abaisser le centre de gravité du test vers le pôle aboral ; de nombreux tests de gilianelles montrent des adaptations de ce type qui ont conduit à suggérer une vie planctonique avec le pôle oral vers le haut (voir la discussion et une reconstitution dans et ,
2006) ; pour cette raison, les tests seront présentés dans les planches, autant que faire ce peut, dans leur position biologique ainsi reconnue, c'est à dire le pôle oral vers le haut.
montre une forme légèrement plus grande pour le niveau sommital de la carrière (cote 172) ; les critères morphologiques sont trop simples et l'abondance souvent trop faible pour tenter avec assurance de détacher des subdivisions pour ce taxon défini à la cote 54,9 oů il est contemporain du foraminifère planctonique Radotruncana calcarata ; l'aplatissement du pôle oral par rapport à une sphère régulière, a pour effet d'abaisser le centre de gravité du test vers le pôle aboral ; de nombreux tests de gilianelles montrent des adaptations de ce type qui ont conduit à suggérer une vie planctonique avec le pôle oral vers le haut (voir la discussion et une reconstitution dans et ,
2006) ; pour cette raison, les tests seront présentés dans les planches, autant que faire ce peut, dans leur position biologique ainsi reconnue, c'est à dire le pôle oral vers le haut.
On rencontre des formes voisines, plus grandes que 150 µm bien distinctes par la taille comme par l'arrangement des cellules du réseau ; leur illustration reste trop éparse pour proposer d'autres espèces ici. Ce premier taxon, très simple parmi les gilianelles, est considéré comme "primitif".
Distribution : Le taxon a été rencontré çà et là entre les cotes 49 et 172 depuis la zone d'existence de Radotruncana calcarata au Campanien supérieur jusqu'à un Maastrichtien inférieur déjà avancé. Quelques spécimens ont aussi été trouvés plus haut, 30 m sous la limite Maastrichtien-Danien qui affleure en dehors de la carrière. Son abondance est difficile à estimer (confusion avec les nombreuses pithonelles). Il est plus commun aux cotes 172 et 145 avec environ 10 spécimens par gramme de sédiment traité.
Tercensella doliolum nov. sp. (code N1b, ce travail)
Synonymie : Code N1.- et ,
2006, Pl. 6 ![]() .
.
Description : Test petit, de diamètre 125 à 130 µm ; face orale bien individualisée, sub-plane ce qui donne au test un aspect véritablement en tonnelet court ; orifice central entouré de 3-4 rides concentriques nettes ; flancs perpendiculaires avec les cellules du réseau nannostructural alignées selon les méridiens ce qui donne un aspect cannelé en vue polaire et rayé en vue de profil à la loupe binoculaire ; hauteur inférieure au diamètre sur le profil ; flancs renforcés de 2 épaulements ; l'un, moins net, sépare la face orale des flancs ; l'autre, très appuyé, ressort du flanc du fait du léger étranglement du test qui le précède vers la base du test (Vue 10 ![]() ) ; face aborale, perpendiculaire aux flancs vers l'extérieur et nettement renflée au centre.
) ; face aborale, perpendiculaire aux flancs vers l'extérieur et nettement renflée au centre.
Étymologie : L'espèce tire son nom du latin doliolum : tonnelet ; c'est la tercisiennelle tonnelet.
Holotype N1b : Provenant du niveau 11,0, l'holotype a été coloré (Planche annexe 1 ![]() , vignette 2) et inventorié au Musée du site de Tercis sous la référence TCS.G033 ; il est accompagné de 14 topotypes.
, vignette 2) et inventorié au Musée du site de Tercis sous la référence TCS.G033 ; il est accompagné de 14 topotypes.
Discussion : Le taxon codifié N1b pour le distinguer de N1a décrite dessous est membre du groupe N1 (sphère à côtes) modélisé par et (2006) ; sa morphologie est originale et aucun test pouvant être confondu n'a été rencontré. Les deux formes N2b et N1b peuvent être prises comme bases morphologiques (origine phyllogénétique?) de toutes les formes qui vont suivre. La première, sub-sphérique, conduit aux formes en disque du groupe A par simple aplatissement. Ensuite, par l'émergence d'expansions équatoriales radiaires, on aboutit aux formes du groupe B. Les formes du groupe C sont des sphères moins aplaties et avec des expansions moins étirées. De l'autre côté, on passe aisément de la forme en tonnelet (N1b), aux formes en cupule G0, J, H par l'acquisition d'expansions en collerette à la jointure des faces et des flancs. Sur ces dernières formes, l'acquisition de crêtes ou de côtes sur la face orale et les flancs conduit aux formes des groupes codifiés E2, E3, G1 et M.
Distribution : La tercisiennelle tonnelet a été observée sur une faible épaisseur depuis la cote -9,5 jusqu'à la cote 25 ; l'espèce est plus commune au niveau 11,0 oů son abondance atteint 2 à 3 individus par gramme de sédiment traité ; elle caractérise des niveaux du Campanien immédiatement sous-jacents à la zone d'existence de l'ammonite Bostrychoceras polyplocum (, 1841).
Tercensella alveare nov. sp. (code N1a, ce travail)
Description : Corps petit (diamètre oral 135 µm, aboral 110 µm, hauteur : 120 µm) en forme de tonnelet chapeauté rappelant une ruche ancienne, en paille ; face orale tronconique s'élevant nettement sur 1/3 du diamètre puis sub-plane, prolongée d'un fin bourrelet périphérique surplombant les flancs ; flancs sub-cylindriques se rétrécissant à peine vers le bas, cannelés à la loupe binoculaire ; face aborale un peu plus étroite que la face orale, d'abord plane puis avec un large dôme central dépassant vers le bas nettement sur les vues de profil (Vue 12 ![]() ) ; nannostructure en tricot bien marquée, rayonnée puis concentrique plus près de l'orifice sur la face orale (Vue 11
) ; nannostructure en tricot bien marquée, rayonnée puis concentrique plus près de l'orifice sur la face orale (Vue 11 ![]() ), méridienne sur les flancs, rayonnée puis plus désordonnée vers le centre de la face aborale (Vue 13
), méridienne sur les flancs, rayonnée puis plus désordonnée vers le centre de la face aborale (Vue 13 ![]() ).
).
Étymologie : Le nom d'espèce dérive du latin alveare : ruche, d'après sa forme générale ; c'est la tercisiennelle ruche.
Holotype N1a : L'holotype a été taché de rouge (Planche annexe 1 ![]() , vignette 3), il provient du niveau 54,9 à Tercis et est inventorié au Musée du site de Tercis sous la référence TCS.G034 avec 3 autres spécimens provenant des niveaux 62,5 et 79,8 et tous photographiés en microscopie optique (Planche annexe 1
, vignette 3), il provient du niveau 54,9 à Tercis et est inventorié au Musée du site de Tercis sous la référence TCS.G034 avec 3 autres spécimens provenant des niveaux 62,5 et 79,8 et tous photographiés en microscopie optique (Planche annexe 1 ![]() , vignette 3).
, vignette 3).
Discussion : Le taxon appartient au groupe des formes globuleuses simples ; il se distingue de Tercensella doliolum (N1b) par sa face orale élevée et par le bourrelet surplombant qui l'entoure, l'ensemble formant chapeau ; N1a était incluse dans N1 pour et (2006). Il existe au moins une autre forme (allongée, en ballon de rugby) de ce genre ; elle n'a pas été formalisée car 1 seul spécimen, parfaitement préservé, a été trouvé ; en outre, certains spécimens les plus jeunes, à la cote 172, présentent aussi un profil un peu étiré axialement qui sont inclus dans le présent taxon.
Distribution : La tercisiennelle ruche est présente de façon sporadique entre les cotes 54,9 et 172, de part et d'autre de la limite Campanien-Maastrichtien ; elle est toujours très rare, 9 spécimens ont été observés ; elle est toujours plus récente que Tercensella doliolum (N1b).
Genre Azymella nov. gen.
Étymologie : Le genre a été nommé d'après la ressemblance avec la pâte à pain non levée, latin azyma : pain azyme.
Espèce type : Azymella cannabinata nov. sp.
Description : Disque en galette simple sans flanc net, sans ornementation microstructurale concentrique ou radiaire ; nannostructure en canevas (mailles perpendiculaires) identique sur les deux faces.
Espèces attribuées au genre : cannabinata, offa.
Remarques : L'appartenance du genre aux gilianelles est discutable. La difficulté majeure est relative à l'observation d'une desquamation commune du test pour l'espèce A. offa (Vue 22 ![]() ); ce phénomène atteint une structure interprétée comme le test de moins de 5 µm d'épaisseur alors que le test des gilianelles est usuellement trois fois plus épais ( et ,
2006) sauf exception (voir plus loin gilianelle B4) et ne se desquame jamais. La nannostructure visible sur une partie des tests rapportés à A. offa ressemble à celle observée pour l'espèce A. cannabinata par l'aspect en canevas ; ainsi, les deux taxons appartiendraient au même genre d'après cette nannostructure de surface du test simple et l'organisation en galette très simple elle aussi. A. cannabinata possède un test d'épaisseur seulement légèrement inférieure à celle des autres gilianelles (près de 10 µm, Vue 15
); ce phénomène atteint une structure interprétée comme le test de moins de 5 µm d'épaisseur alors que le test des gilianelles est usuellement trois fois plus épais ( et ,
2006) sauf exception (voir plus loin gilianelle B4) et ne se desquame jamais. La nannostructure visible sur une partie des tests rapportés à A. offa ressemble à celle observée pour l'espèce A. cannabinata par l'aspect en canevas ; ainsi, les deux taxons appartiendraient au même genre d'après cette nannostructure de surface du test simple et l'organisation en galette très simple elle aussi. A. cannabinata possède un test d'épaisseur seulement légèrement inférieure à celle des autres gilianelles (près de 10 µm, Vue 15 ![]() ) ; sa nannostructure de surface est, à son tour, très proche de l'aspect visible sur le corps du test de l'espèce codifiée A3 (Scutellella pulchrevelifera, Vues 62-63
) ; sa nannostructure de surface est, à son tour, très proche de l'aspect visible sur le corps du test de l'espèce codifiée A3 (Scutellella pulchrevelifera, Vues 62-63 ![]() ) comme sur d'autres membres assurés du groupe des gilianelles. Ainsi, malgré des caractéristiques distinctes de celles de la majorité des gilianelles A. offa semble pouvoir être rattachée, de proche en proche, aux gilianelles au test plus épais, ne se desquamant pas et possèdant une nannostructure généralement plus élaborée qu'un simple canevas lequel n'est organisé que sur les flancs du test.
) comme sur d'autres membres assurés du groupe des gilianelles. Ainsi, malgré des caractéristiques distinctes de celles de la majorité des gilianelles A. offa semble pouvoir être rattachée, de proche en proche, aux gilianelles au test plus épais, ne se desquamant pas et possèdant une nannostructure généralement plus élaborée qu'un simple canevas lequel n'est organisé que sur les flancs du test.
Azymella cannabinata nov. sp.
Vues 14-18 ![]() et 19
et 19 ![]()
Synonymie : Code A7.- et ,
2006, Pl. 2 ![]() .
.
Description : Disque simple, en palet, souvent grand ; taille variant de 165 à 220 µm ; face orale avec un orifice sans lèvre, discret, de diamètre 15 µm (Vues 14 et 18 ![]() et Vue 19
et Vue 19 ![]() ), simple et lisse à la loupe binoculaire ; profil symétrique avec un aplatissement tel que la cavité du test est très réduite (Vue 15
), simple et lisse à la loupe binoculaire ; profil symétrique avec un aplatissement tel que la cavité du test est très réduite (Vue 15 ![]() ) ; nannostructure en canevas régulier aux mailles perpendiculaires.
) ; nannostructure en canevas régulier aux mailles perpendiculaires.
Étymologie : L'espèce tire son nom de l'aspect de la nannostructure de surface, latin cannabina : grosse toile, canevas ; c'est l'azymelle en canevas.
Holotype A7 : L'holotype complet provient du niveau 11,0 et a été coloré (Planche annexe 1 ![]() , vignette 4) et inventorié au Musée du site de Tercis sous la référence TCS.G035 ; il est accompagné de 28 topotypes.
, vignette 4) et inventorié au Musée du site de Tercis sous la référence TCS.G035 ; il est accompagné de 28 topotypes.
Discussion : Ŕ la loupe binoculaire, l'azymelle en canevas ne se différencie d'un débris carbonaté quelconque que par l'ombre centrale due à l'orifice présent sur une seule face et à la forme circulaire quand elle est bien préservée. S'il est difficile à déceler, le taxon ne présente aucune confusion connue en microscopie électronique.
Distribution : L'azymelle en canevas est présente entre les niveaux -5,0 et +25,0. Elle est commune dans le niveau 11,0 oů son abondance avoisine 6 spécimens par gramme de sédiment traité.
Azymella offa nov. sp. (code A8, ce travail)
Description : Corps en disque émoussé semblable à une miche de pain, petit (diamètre 150 µm, hauteur : 50 µm) sans relief ; face orale bombée, aborale concave (Planche annexe 1 ![]() , vignette 5) ; orifice très discret lorsque le test est conservé (Vue 20
, vignette 5) ; orifice très discret lorsque le test est conservé (Vue 20 ![]() ) mieux visible sur le moulage interne (Vue 22
) mieux visible sur le moulage interne (Vue 22 ![]() ) ; pas de limite marquée entre les deux faces qui se joignent sur un flanc court arrondi (Vue 22
) ; pas de limite marquée entre les deux faces qui se joignent sur un flanc court arrondi (Vue 22 ![]() ) ; nannostructure en tricot bien marquée sur la portion externe des faces et la tranche ; aspect en canevas comme pour A7 (Vue 21
) ; nannostructure en tricot bien marquée sur la portion externe des faces et la tranche ; aspect en canevas comme pour A7 (Vue 21 ![]() ). Lorsque le test est détérioré, le moulage interne présente des picots suggérant que le test est perforé.
). Lorsque le test est détérioré, le moulage interne présente des picots suggérant que le test est perforé.
Étymologie : Le nom d'espèce dérive du latin offa : miche, d'après la forme générale ; c'est l'azymelle miche.
Holotype A8 : L'holotype, provenant de la cote 79,8 à Tercis, a été coloré (Planche annexe 1 ![]() , vignette 5) et inventorié au Musée du site de Tercis sous la référence TCS.G036 ; il est accompagné de 6 topotypes choisis parmi une quinzaine de spécimens.
, vignette 5) et inventorié au Musée du site de Tercis sous la référence TCS.G036 ; il est accompagné de 6 topotypes choisis parmi une quinzaine de spécimens.
Discussion : Par la simplicité de son test, plus épais et de diamètre plus petit que celui de l'azymelle en canevas (A7) et par sa nannostructure semblable à cette dernière, A8 se rattache au même genre.
Distribution : Difficile à reconnaître à la loupe, cette forme n'a été repérée qu'au niveau 79,8 oů elle n'est pas commune ; 15 spécimens ont été identifiés.
Genre Numismella nov. gen.
Étymologie : Le genre tire son nom de la forme générale du test, latin numisma : médaille
Espèce type : Numismella tarbellica nov. sp. (espèce unique).
Description : Disque en galette, symétrie axiale parfaite comme précédemment pour Azymella ; début de collerette équatoriale (bourrelet aigu) ; ornementation microstructurale concentrique sur les deux faces.
Numismella tarbellica nov. sp.
Synonymie : Code A1a.- et ,
2006, Pl. 1 ![]() .
.
Description : Corps en disque fin comparable à une galette bretonne un peu épaisse ; taille moyenne voisine de 200 µm assez constante pour être utilisée comme échelle pour les autres formes lors des tris ; certains spécimens atteignent 220 µm, quelques rares sont petits (Vue 27 ![]() ) ; face orale avec un orifice enfoncé, sans lèvre (Vue 23
) ; face orale avec un orifice enfoncé, sans lèvre (Vue 23 ![]() ) ornée de 3 rides concentriques visibles à la loupe binoculaire sur la partie distale des tests ; profil bas (rapport diamètre/hauteur proche de 3) et symétrique à la loupe ; étroite collerette équatoriale visible au microscope électronique sur ce profil ; face aborale, au centre légèrement déprimé, avec aussi 3 rides concentriques sur la moitié distale du test de la plupart des spécimens ; ride médiane parfois estompée (Vue 26
) ornée de 3 rides concentriques visibles à la loupe binoculaire sur la partie distale des tests ; profil bas (rapport diamètre/hauteur proche de 3) et symétrique à la loupe ; étroite collerette équatoriale visible au microscope électronique sur ce profil ; face aborale, au centre légèrement déprimé, avec aussi 3 rides concentriques sur la moitié distale du test de la plupart des spécimens ; ride médiane parfois estompée (Vue 26 ![]() ) ou effacée (Vue 25
) ou effacée (Vue 25 ![]() ) ; nannostructure constante avec des cellules rectangulaires délimitées entre les rides concentriques tandis que des alignements nets rayonnants sont présents dans la partie centrale sur les deux faces ; crénelure périphérique de 70 à 80 dents parfois visible à la loupe binoculaire.
) ; nannostructure constante avec des cellules rectangulaires délimitées entre les rides concentriques tandis que des alignements nets rayonnants sont présents dans la partie centrale sur les deux faces ; crénelure périphérique de 70 à 80 dents parfois visible à la loupe binoculaire.
Étymologie : L'espèce tire son nom du peuple occupant la région dacquoise aux temps romains, les Tarbelles (latin : Tarbelli), lesquels donnèrent leur nom à la ville d'eau de Dax : Aquae tarbellicae, plus tard devenue la ville d'Acqs puis la ville de Dax ; cette espèce est ainsi la petite médaille tarbelle.
Holotype A1a : Provenant du niveau 11,0 l'holotype a été marqué d'une tache orangée (Planche annexe 1 ![]() , vignette 6) et inventorié au Musée du site de Tercis sous la référence TCS.G037 ; il est préservé avec 6 topotypes choisis parmi des centaines de spécimens.
, vignette 6) et inventorié au Musée du site de Tercis sous la référence TCS.G037 ; il est préservé avec 6 topotypes choisis parmi des centaines de spécimens.
Discussion : L'aspect aplati, la présence d'un orifice déprimé et la taille permettent de reconnaître sans ambiguďté la petite médaille tarbelle. Il existe plusieurs autres "galettes" de tailles et de profils différents dans la section. Une documentation complémentaire reste nécessaire pour formaliser un ou plusieurs taxons parmi ces formes.
Distribution : Numismella tarbellica (A1a) a été observée depuis la cote -5,0 jusqu'à la cote 172. Son abondance varie. Elle est parfois dominante ; elle constitue ainsi l'essentiel des gilianelles présentes au niveau 172 avec environ 150 spécimens par gramme de sédiment traité alors que les autres taxons ne sont représentés que par de rares individus. Ailleurs, son abondance atteint, par exemple, 40 par gramme à la cote 62,0 et 100 par gramme à la cote +5,0. L'espèce est une constante du faciès aturien de la Grande Carrière de Tercis ; elle n'a pas été trouvée dans les deux niveaux du Maastrichtien supérieur traités qui ne renferment, par ailleurs, que de rares spécimens des groupes codifiés M et N.
Genre Aturella nov. gen.
Étymologie : Le nom de genre dérive du latin Atur : nom du fleuve Adour.
Espèce type : Aturella aequilatera nov. sp.
Description : Disque haut, grand (diamètre autour de 200 µm), sans ornementation microstructurale concentrique ou radiale ; acquisition d'une symétrie bilatérale orientée.
Espèces attribuées au genre : aequilatera, altodepressa, angulata, edentula.
Aturella aequilatera nov. sp.
Vues 30-33 ![]() et 34-35
et 34-35 ![]()
Synonymie : Code A4b.- ,
2007,
Fig. 3 ; et ,
2006, Pl. 2 ![]() .
.
Description : Corps en disque haut, grand (diamètre 170-200 µm) ; face orale élevée tronconique sommée d'un plateau de diamètre moitié du corps dont se détache un col épais de 20 µm entourant l'orifice circulaire de diamètre 15 à 20 µm (Vue 30 ![]() ) ; sur certains spécimens, ce col est légèrement enfoncé à la base ; profil axial quasi-symétrique ; un léger bourrelet marque l'équateur du test (Vue 31
) ; sur certains spécimens, ce col est légèrement enfoncé à la base ; profil axial quasi-symétrique ; un léger bourrelet marque l'équateur du test (Vue 31 ![]() ) ; face aborale en dôme bas, largement enfoncé (Vue 33
) ; face aborale en dôme bas, largement enfoncé (Vue 33 ![]() ).
).
Étymologie : L'espèce tire son nom de la symétrie du test avec faces orale et aborale presque également développées par rapport à l'équateur ; du latin aequus : égal et du latin latus : côté, face ; c'est l'aturelle à faces égales.
Holotype A4b : Coloré, provenant du niveau 57,9, l'holotype (Planche annexe 1 ![]() , vignette 12) a été inventorié au Musée du site de Tercis sous la référence TCS.G038 ; il est accompagné de 13 topotypes choisis parmi la centaine de spécimens prélevés.
, vignette 12) a été inventorié au Musée du site de Tercis sous la référence TCS.G038 ; il est accompagné de 13 topotypes choisis parmi la centaine de spécimens prélevés.
Discussion : Circulaire au premier abord, le taxon présente une différenciation antéro-postérieure plus ou moins nette selon les spécimens mais les caractéristiques sont constantes (cf. A4a ci-dessous) avec un avant en "bord d'attaque" plus trapu, plus arrondi et un arrière en "bord de fuite" plus effilé ce qui est illustré par les vues de profil (Vues 31-32 ![]() ). De dessus (Vue 30
). De dessus (Vue 30 ![]() et Vue 34
et Vue 34 ![]() ), l'avant est plus régulier, l'arrière plus festonné.
), l'avant est plus régulier, l'arrière plus festonné.
Distribution : L'aturelle à faces égales se rencontre entre les cotes 47,2 et 62,5 ; elle est plus commune entre les cotes 47,2 et 57,9. La forme est dominante parmi les gilianelles dans ce dernier niveau d'oů est tiré l'holotype, niveau le plus riche en Radotruncana calcarata, ce foraminifère planctonique clé de la biostratigraphie du Campanien supérieur.
Aturella altodepressa nov. sp.
Synonymie : Code A4a.- et ,
2006, Pl. 1 ![]() .
.
Description : Corps en disque axialement dissymétrique, rappelant un casque plat, grand (diamètre 170-200 µm), souvent déformé latéralement -en béret à côtés rabattus- (Vue 37 ![]() ) ; face orale très élevée tronconique sommée d'un plateau de diamètre moitié du corps dont se détache un col épais de 20 µm entourant l'orifice circulaire de diamètre 15 à 20 µm ; une section de profil (Vue 38
) ; face orale très élevée tronconique sommée d'un plateau de diamètre moitié du corps dont se détache un col épais de 20 µm entourant l'orifice circulaire de diamètre 15 à 20 µm ; une section de profil (Vue 38 ![]() ) montre l'orifice vers le haut (interruption du test) alors que la paroi du test est continue vers le bas ; de profil, l'essentiel du volume de la cavité du test est créé par la face orale ; léger bourrelet à l'équateur du test, surtout vers l'arrière (Vues 38 et 40
) montre l'orifice vers le haut (interruption du test) alors que la paroi du test est continue vers le bas ; de profil, l'essentiel du volume de la cavité du test est créé par la face orale ; léger bourrelet à l'équateur du test, surtout vers l'arrière (Vues 38 et 40 ![]() ) oů il s'affine pour devenir transparent (Planche annexe 1
) oů il s'affine pour devenir transparent (Planche annexe 1 ![]() , vignette 11) ; face aborale, d'abord en dôme bas, puis enfoncée en son centre sur une large surface.
, vignette 11) ; face aborale, d'abord en dôme bas, puis enfoncée en son centre sur une large surface.
Étymologie : L'espèce tire son nom de la dissymétrie du profil avec sa face orale haute, sa face aborale déprimée du latin altus : haut et depressus : déprimé, surface enfoncée ; c'est l'aturelle haute-déprimée.
Holotype A4a : Provenant du niveau +5,0, l'holotype coloré a une face orale élevée, un col développé et une différenciation antéro-postérieure nette (Planche annexe 1 ![]() , vignette 11) ; il est inventorié au Musée du site de Tercis (TCS.G039) et est accompagné de 6 topotypes, très hauts avec un col plus élevé que sur les Vues 38 ou 40
, vignette 11) ; il est inventorié au Musée du site de Tercis (TCS.G039) et est accompagné de 6 topotypes, très hauts avec un col plus élevé que sur les Vues 38 ou 40 ![]() , choisis parmi quelques dizaines de spécimens prélevés.
, choisis parmi quelques dizaines de spécimens prélevés.
Discussion : Outre la dissymétrie axiale, les tests montrent communément un allongement (Vues 36 et 43 ![]() ) qui se complète d'une orientation. Les caractéristiques de cette dernière sont du même ordre que celles décrites pour la forme A4b.
Est interprété comme avant le côté plus lisse, plus régulier et plus brusquement élevé, à droite de la section (Vue 38
) qui se complète d'une orientation. Les caractéristiques de cette dernière sont du même ordre que celles décrites pour la forme A4b.
Est interprété comme avant le côté plus lisse, plus régulier et plus brusquement élevé, à droite de la section (Vue 38 ![]() ), à gauche du profil (Vue 40
), à gauche du profil (Vue 40 ![]() ). Est interprété comme arrière le rebord plus aigu, plus découpé. La différence est claire sur la Vue 43
). Est interprété comme arrière le rebord plus aigu, plus découpé. La différence est claire sur la Vue 43 ![]() , moins assurée sur la Vue 40
, moins assurée sur la Vue 40 ![]() et très subtile sur la Vue 42
et très subtile sur la Vue 42 ![]() . L'orientation se fait par assimilation avec le profil des formes adaptées au déplacement relatif par rapport à un fluide (aile d'avion, d'oiseau, aileron ou palettes natatoires d'organismes aquatiques). Sur quelques spécimens (Planche annexe 1
. L'orientation se fait par assimilation avec le profil des formes adaptées au déplacement relatif par rapport à un fluide (aile d'avion, d'oiseau, aileron ou palettes natatoires d'organismes aquatiques). Sur quelques spécimens (Planche annexe 1 ![]() , vignette 11), le col développé semble lui-même légèrement déjeté vers l'arrière comme une cheminée de paquebot, complétant cette adaptation.
, vignette 11), le col développé semble lui-même légèrement déjeté vers l'arrière comme une cheminée de paquebot, complétant cette adaptation.
Ce taxon précède A4b dans le temps ; la variation morphologique est progressive. On aurait pu faire deux sous-espèces ; la distribution distincte a suggéré que l'adoption de deux espèces n'était pas impropre car utile stratigraphiquement. Une compression liée à la compaction, de la forme A4b peut laisser penser à une forme A4a et le diagnostic peut être délicat pour des individus isolés.
Distribution : L'aturelle haute-déprimée se rencontre à la base de la section de Tercis depuis la cote -5,0 jusqu'à la cote 43. L'abondance est notable jusqu'à la cote 25,0 et surtout aux cotes +5,0 et 11,0 oů 5 à 10 spécimens sont présents par gramme de sédiment traité.
Aturella angulata nov. sp. (code A4b', ce travail)
Description : Corps en disque haut, grand (diamètre 180-200 µm le plus souvent) ; face orale élevée, tronconique avec un plateau de diamètre moitié du corps dont se détache moins nettement que pour les 2 espèces précédentes, un col épais entourant l'orifice circulaire ; profil axial assez symétrique avec un bourrelet aigu à l'équateur du test (Vues 45 et 47 ![]() ) ; face aborale (Vue 48
) ; face aborale (Vue 48 ![]() ) avec un anneau large aux contours anguleux se détachant brusquement de la surface du test ; différenciation antéro-postérieure commune (Vue 46
) avec un anneau large aux contours anguleux se détachant brusquement de la surface du test ; différenciation antéro-postérieure commune (Vue 46 ![]() ) avec l'avant plus haut et l'arrière plus effilé sur le profil (voir la modélisation Fig. 4
) avec l'avant plus haut et l'arrière plus effilé sur le profil (voir la modélisation Fig. 4 ![]() ) ; de dessus l'avant est plus régulier et l'arrière plus dentelé du fait d'un bourrelet plus étiré parfois festonné (Planche annexe 1
) ; de dessus l'avant est plus régulier et l'arrière plus dentelé du fait d'un bourrelet plus étiré parfois festonné (Planche annexe 1 ![]() , vignette 13) ; nannostructure semblable à celle des formes du genre avec, souvent, des arrangements concentriques de mailles visibles sur la face orale.
, vignette 13) ; nannostructure semblable à celle des formes du genre avec, souvent, des arrangements concentriques de mailles visibles sur la face orale.
Étymologie : Le nom d'espèce dérive du latin angulata : anguleux qui rappelle l'aspect particulier de la face aborale ; c'est l'aturelle anguleuse.
Holotype A4b' : Provenant du niveau 79,8 l'holotype coloré est bien dégagé à l'exception d'un petit débris organique collé à la face aborale (visible sur la Planche annexe 1 ![]() , vignette 13) ; il montre une différenciation antéro-postérieure ; inventorié au Musée du site de Tercis sous la référence TCS.G040, il est accompagné de 16 topotypes choisis parmi une quarantaine de spécimens.
, vignette 13) ; il montre une différenciation antéro-postérieure ; inventorié au Musée du site de Tercis sous la référence TCS.G040, il est accompagné de 16 topotypes choisis parmi une quarantaine de spécimens.
Discussion : Le volume anguleux de la face aborale distingue cette espèce de A4b (Aturella aequilatera) ; cet aspect en anneau surimposé de la face aborale est bien marqué à la loupe binoculaire ; les formes typiques sont distinctes par ce détail mais si A4b' succède clairement à A4b le passage d'une forme à l'autre est continu ; le rang de sous-espèce serait aussi justifié mais il semble utile de souligner l'intérêt stratigraphique des différences morphologiques : une évolution a modifié l'espèce précédente.
Distribution : Ŕ Tercis, cette espèce a été observée aux niveaux 69,0 et 79,8 ce qui représente une durée de vie stratigraphique de l'espèce de 0,5 Ma ; A4b' n'y est pas rare avec de l'ordre de 5 spécimens par gramme de sédiment traité ; des spécimens très proches existent dès la cote 65,4 et jusqu'à la cote 82,0 voire la cote 89,0, une durée de dépôt de l'ordre de 1 Ma cette fois.
Aturella edentula nov. sp.
Synonymie : Code A6.- et ,
2006, Pl. 2 ![]() .
.
Description : Espèce la plus grande des aturelles (diamètre souvent supérieur à 200 µm) ; profil symétrique ; face orale tronconique déprimée au centre (Vue 52 ![]() ) ; col peu marqué ou absent autour de l'orifice (Vue 49
) ; col peu marqué ou absent autour de l'orifice (Vue 49 ![]() ) ; face aborale tronconique, avec une ébauche de collerette péri-apicale épaisse, de diamètre 100 µm (Vue 50
) ; face aborale tronconique, avec une ébauche de collerette péri-apicale épaisse, de diamètre 100 µm (Vue 50 ![]() ) ; différenciation antéro-postérieure comme dans les autres membres du genre :
côté antérieur plus trapu, côté postérieur plus effilé (profil, Vue 51
) ; différenciation antéro-postérieure comme dans les autres membres du genre :
côté antérieur plus trapu, côté postérieur plus effilé (profil, Vue 51 ![]() ) ; nannostructure organisée en cercles concentriques sans relief microstructural ce qui fait paraître le test lisse à la loupe binoculaire.
) ; nannostructure organisée en cercles concentriques sans relief microstructural ce qui fait paraître le test lisse à la loupe binoculaire.
Étymologie : Le nom d'espèce est tiré du latin edentulus : édenté car l'orifice en dépression mime la bouche invaginée d'un vieillard ayant perdu ses dents ; c'est l'aturelle édentée.
Holotype A6 : Provenant du niveau 172, l'holotype légèrement teinté a un petit débris sur la face orale, côté arrière gauche ; différenciation antéro-postérieure discrète :
la moitié postérieure a un pourtour légèrement festonné (Planche annexe 1 ![]() , vignette 14) ; inventorié au Musée du site de Tercis sous la référence TCS.G041, il est accompagné de 7 topotypes choisis parmi une quinzaine de spécimens récoltés à ce niveau.
, vignette 14) ; inventorié au Musée du site de Tercis sous la référence TCS.G041, il est accompagné de 7 topotypes choisis parmi une quinzaine de spécimens récoltés à ce niveau.
Discussion : L'orifice dans une dépression différencie A. edentula des trois taxons précédents attribués au genre ; d'un autre côté, sa taille, sa forme générale et le fait qu'elle se substitue aux formes plus anciennes dans la succession stratigraphique, militent pour un rattachement au genre Aturella ; il pourrait s'agir d'une convergence de forme liée à la niche écologique occupée mais la logique de cette évolution générale est forte. C'est avec cette espèce nouvelle que se termine la lignée des aturelles connue actuellement. Cet ensemble est, à Tercis, un outil de repérage stratigraphique intéressant ; il demandera un examen plus détaillé par la suite : pour le moment, une succession morphologique est formalisée.
Distribution : Au Maastrichtien, à partir de la cote 125,5 notamment, c'est l'aturelle édentée typique (A6) qui est présente ; sa morphologie est moins caractérisée dans le Campanien terminal oů les restes ne sont pas communs ; le niveau 172 est le plus riche connu avec 1 ou 2 spécimens par gramme d'échantillon traité parmi d'autres taxons bien plus abondants (A1a livre plus de 15 spécimens par gramme de sédiment traité à la même cote).
Genre Scutellella nov. gen.
Étymologie : Le nom de genre dérive du latin scutella : soucoupe d'après la forme générale du test.
Espèce type : Scutellella crassa nov. sp
Description : Disque peu élevé avec une collerette péri-apicale en anneau développée (au contraire du genre Numismella) dont le rôle de quille peut être évoqué ; collerette seulement esquissée pour une des formes (A3) mais, dans ce cas, une face aborale avec dépression centrale joue le même rôle dans l'équilibre hydrodynamique ; ornementation microstructurale concentrique souvent présente sur la face aborale (comme Numismella) mais jamais sur la face orale (au contraire de Numismella) ; collerette équatoriale développée sauf chez l'espèce type.
Espèces attribuées au genre : crassa, pulchrevelifera, circularis, longivelifera.
Scutellella crassa nov. sp.
Synonymie : Code A1b.- et ,
2006, Pl. 1 ![]() .
.
Description : Corps en disque petit (diamètre 130-165 µm, parfois plus petits que 100 µm) aux bords tronqués ; face orale peu élevée, en dôme bas et large, aplati à légèrement déprimé au centre (Vues 56-57 ![]() ) ; orifice axial simple ; bordure externe présentant une ride équatoriale discrète (Vue 58
) ; orifice axial simple ; bordure externe présentant une ride équatoriale discrète (Vue 58 ![]() ) ; face aborale élevée, débutant par un flanc tronconique d'angle faible ; ce flanc se poursuit par un dôme bas avec plusieurs rides concentriques ; relief en anneau épais péri-apical de diamètre 75 µm, souvent peu élevé (Vues 54-55
) ; face aborale élevée, débutant par un flanc tronconique d'angle faible ; ce flanc se poursuit par un dôme bas avec plusieurs rides concentriques ; relief en anneau épais péri-apical de diamètre 75 µm, souvent peu élevé (Vues 54-55 ![]() ) mais parfois presque en collerette (Vue 59
) mais parfois presque en collerette (Vue 59 ![]() ) ; nannostructure en mailles concentriques sur la face orale (Vue 53
) ; nannostructure en mailles concentriques sur la face orale (Vue 53 ![]() ), arrangée en 4 rides sur le flanc (Vues 58-59
), arrangée en 4 rides sur le flanc (Vues 58-59 ![]() ) et 2 à 3 rides concentriques sur la face aborale (Vue 54
) et 2 à 3 rides concentriques sur la face aborale (Vue 54 ![]() ).
).
Étymologie : Le nom d'espèce dérive du latin crassus : épais d'après la caractéristique qui permet d'identifier le taxon à la loupe : l'épaisseur du corps ; c'est la scutellelle épaisse.
Holotype A1b : L'holotype provenant du niveau 79,8 a été teinté sur la face orale (Planche annexe 1 ![]() , vignette 7) ; il est préservé et inventorié au Musée du site de Tercis sous la référence TCS.G042 et accompagné de 21 topotypes.
, vignette 7) ; il est préservé et inventorié au Musée du site de Tercis sous la référence TCS.G042 et accompagné de 21 topotypes.
Discussion : A1b est équipé d'une collerette péri-apicale typique du genre mais pas de la collerette équatoriale ; la formalistion du taxon fut délicate du fait de la présence quasi permanente, avec lui, d'une autre forme de même taille petite, aux flancs plus aigus vus de profil, de même apparence en vue orale et aborale à la loupe comme au microscope électronique ; l'interprétation actuelle pour ces formes problématiques (elles furent codifiées diversement A1b' ou A2a') est que, les scutelleles à collerette (circulaire : A2a ou longue : A2b ci-dessous) usées perdent leur collerette équatoriale ce qui laisse des tests très voisins de ceux de A1b si ce n'est l'épaisseur.
Distribution : La scutellelle épaisse est présente tout au long de la coupe entre la cote -5,0 et la cote 172 avec des variations morphologiques faibles ; les planches donnent des exemples depuis la cote 11,0 (Vue 59 ![]() ) jusqu'à la cote 172 (Vue 58
) jusqu'à la cote 172 (Vue 58 ![]() ) ; dans les divers niveaux, il semble y avoir concurrence entre cette forme et la petite médaille tarbelle (A1a) ; parfois A1b domine avec près de 50 spécimens par gramme de sédiment traité, parfois elle est rare avec moins d'un spécimen par gramme et A1a domine. Nos observations sur la fréquence de l'espèce ont été en partie faussées par la question de taille : Lors de tris récents de la fraction comprise entre 80 et 100 µm, des quantités de A1b insoupçonnées dans la fraction 100-250 µm ont été récoltées.
) ; dans les divers niveaux, il semble y avoir concurrence entre cette forme et la petite médaille tarbelle (A1a) ; parfois A1b domine avec près de 50 spécimens par gramme de sédiment traité, parfois elle est rare avec moins d'un spécimen par gramme et A1a domine. Nos observations sur la fréquence de l'espèce ont été en partie faussées par la question de taille : Lors de tris récents de la fraction comprise entre 80 et 100 µm, des quantités de A1b insoupçonnées dans la fraction 100-250 µm ont été récoltées.
Scutellella pulchrevelifera nov. sp.
Synonymie : Code A3.- et ,
2006, Pl. 1 ![]() .
.
Description : Corps en disque simple, fin, lenticulaire de 120 à 160 µm de diamètre, jusqu'à 170 µm ; corps prolongé d'une collerette équatoriale grande, circulaire, transparente à la loupe binoculaire, souvent endommagée, pouvant dépasser 60 µm de large et dont l'épaisseur initiale proche de 15 µm diminue vers l'extérieur (Vue 61 ![]() ), donnant au taxon sa forme en soucoupe volante ; face orale en dôme bas, sans microstructure radiale ni concentrique ; orifice circulaire, de diamètre 15 à 20 µm sans lèvre ; profil symétrique ; face aborale, en dôme bas, un peu déprimé au pôle ; le rebord de la dépression montre des traces minimes de collerette (Vue 63
), donnant au taxon sa forme en soucoupe volante ; face orale en dôme bas, sans microstructure radiale ni concentrique ; orifice circulaire, de diamètre 15 à 20 µm sans lèvre ; profil symétrique ; face aborale, en dôme bas, un peu déprimé au pôle ; le rebord de la dépression montre des traces minimes de collerette (Vue 63 ![]() ) ; nannostructure dessinant des cellules dont l'arrangement en canevas rappelle sans le reproduire celui de Azymella ; face aborale avec un bourrelet bordant la collerette du côté proximal (Vue 62
) ; nannostructure dessinant des cellules dont l'arrangement en canevas rappelle sans le reproduire celui de Azymella ; face aborale avec un bourrelet bordant la collerette du côté proximal (Vue 62 ![]() ), bourrelet absent (Vue 60
), bourrelet absent (Vue 60 ![]() ) ou faible (Vue 64
) ou faible (Vue 64 ![]() ) face orale.
) face orale.
Étymologie : Le nom d'espèce dérive du latin pulchre : joliment et du latin velifer : pourvu d'une voile, la collerette jouant probablement ce rôle de voile pour la portance de l'espèce dans l'eau ; c'est la scutellelle joliment voilée.
Holotype A3 : Provenant du niveau 11,0 l'holotype choisi est caractérisé par une collerette marquée et de largeur régulière (Planche annexe 1 ![]() , vignette 10) ; inventorié au Musée du site de Tercis sous la référence TCS.G043, il est accompagné de 7 topotypes dont les collerettes sont moins bien préservées.
, vignette 10) ; inventorié au Musée du site de Tercis sous la référence TCS.G043, il est accompagné de 7 topotypes dont les collerettes sont moins bien préservées.
Discussion : La taille remarquable de la collerette équatoriale est le critère principal de reconnaissance pour ce représentant du genre quasi dépourvu de collerette péri-apicale ; cette acquisition adaptative par rapport aux formes en galette donne au test une flottabilité supérieure ; lorsqu'il est endommagé, le taxon reste identifiable à la loupe binoculaire par son profil symétrique et peu élevé par rapport à l'équateur, notamment par rapport au taxon suivant (A2a). La discrétion de l'anneau péri-apical peut laisser un doute sur le rattachement de l'espèce au genre Scutellella ; l'absence de rides concentriques aborales distingue aussi cette espèce de toutes les autres dans le genre mais le développement de la collerette équatoriale la rapproche de deux de ces membres décrits ci-dessous.
Distribution : La scutellelle joliment voilée a surtout été observée entre les niveaux +5,0 et 25,0 oů 30 spécimens ont été collectés. Elle est la plus abondante aux niveaux +5,0 et 11,0 avec 1 à 2 spécimens par gramme de sédiment traité ; 1 seul spécimen a été récolté à la cote 1,1.
Scutellella circularis nov. sp.
Synonymie : Code A2a.- ,
2007,
Fig. 3 ; et ,
2006, Pl. 1 ![]() .
.
Description : Corps en disque de 125-150 µm de diamètre prolongé d'une collerette équatoriale annulaire de 20 à 35 µm de large ; l'ensemble atteint 200 µm au maximum (Vue 66 ![]() ) ; face orale sans microstructure concentrique ni rayonnée (Vues 65 et 68
) ; face orale sans microstructure concentrique ni rayonnée (Vues 65 et 68 ![]() ) ; orifice circulaire, de diamètre 15 à 20 µm sans lèvre, central, au fond d'une légère dépression ; collerette équatoriale annulaire à contour entier circulaire, à structure rayonnée (Vue 70
) ; orifice circulaire, de diamètre 15 à 20 µm sans lèvre, central, au fond d'une légère dépression ; collerette équatoriale annulaire à contour entier circulaire, à structure rayonnée (Vue 70 ![]() ) apparente par transparence à la loupe binoculaire ; profil nettement dissymétrique (Vue 69
) apparente par transparence à la loupe binoculaire ; profil nettement dissymétrique (Vue 69 ![]() ) avec une face orale en dôme bas et une face aborale plus haute :
Rapport diamètre sur hauteur proche de 2 ; face aborale, en tronc de cône prolongée par une collerette cylindrique péri-apicale de diamètre moitié de celui du corps ; 3 rides concentriques constituent une microstructure nette au microscope à balayage mais peu visible à la loupe binoculaire ; nannostructure de la face orale avec 6-7 festons concentriques sur la moitié distale du corps.
) avec une face orale en dôme bas et une face aborale plus haute :
Rapport diamètre sur hauteur proche de 2 ; face aborale, en tronc de cône prolongée par une collerette cylindrique péri-apicale de diamètre moitié de celui du corps ; 3 rides concentriques constituent une microstructure nette au microscope à balayage mais peu visible à la loupe binoculaire ; nannostructure de la face orale avec 6-7 festons concentriques sur la moitié distale du corps.
Étymologie : L'espèce tire son nom du latin circularis : circulaire choisi par opposition avec le taxon suivant ; c'est la scutellelle circulaire.
Holotype A2a : L'holotype choisi a été coloré et provient du niveau 57,9 (Planche annexe 1 ![]() , vignette 8) ; inventorié au Musée du site de Tercis sous la référence TCS.G044, il est accompagné de 7 topotypes choisis parmi des dizaines récoltés à ce niveau.
, vignette 8) ; inventorié au Musée du site de Tercis sous la référence TCS.G044, il est accompagné de 7 topotypes choisis parmi des dizaines récoltés à ce niveau.
Discussion : Le taxon est reconnaissable quand la collerette équatoriale annulaire est présente ; absente, la distinction par rapport à d'autres disques repose sur la présence d'une collerette aborale cylindrique péri-apicale. Lorsque la collerette aborale est aussi usée la distinction d'avec A3 devient délicate. La présence d'une collerette péri-apicale qui déplace le centre de gravité du test vers le pôle aboral est un aspect de l'adaptation à la vie planctonique avec orientation du test la face orale vers le haut.
Distribution : La scutellelle circulaire est bien représentée entre les cotes 49,0 et 109,7. Elle est commune à divers niveaux, notamment dans l'intervalle entre les cotes 70 et 110 oů son abondance atteint 10 spécimens / g mais oů la collerette équatoriale est le plus souvent absente et qui est considérée comme détruite. En dehors de cet intervalle, des spécimens dont le corps montre une morphologie voisine existent entre les cotes -5,0 et +25,0 et jusqu'au début du Maastrichtien (cote 116) mais la collerette est soit toujours absente, ce qui en ferait un taxon distinct, soit toujours érodée ce qui rend la caractérisation délicate.
Scutellella longivelifera nov. sp.
Synonymie : Code A2b.- et ,
2006, Pl. 1 ![]() .
.
Description : Corps en disque légèrement elliptique prolongé d'une collerette équatoriale soulignant le caractère elliptique du taxon ; longueur de l'ensemble dépassant parfois 200 µm (Vue 75 ![]() ) avec un corps plus petit que 100-125 µm ; face orale en dôme peu élevé sans microstructure nette ; profil dissymétrique, semblable à celui de A2a (Vues 72-73
) avec un corps plus petit que 100-125 µm ; face orale en dôme peu élevé sans microstructure nette ; profil dissymétrique, semblable à celui de A2a (Vues 72-73 ![]() ) ; face aborale tronconique plus élevée que la face orale et terminée par une collerette péri-apicale élevée ; nannostructure ordonnée en cellules alignées concentriquement sur la face orale et en cellules alignées radialement sur la face aborale ; collerette équatoriale communément nervurée radialement par transparence sous la loupe binoculaire ; en microscopie électronique les nervures apparaissent parfois sur la partie longue de la collerette ; sur la face orale ce sont des stries fines (Vue 77
) ; face aborale tronconique plus élevée que la face orale et terminée par une collerette péri-apicale élevée ; nannostructure ordonnée en cellules alignées concentriquement sur la face orale et en cellules alignées radialement sur la face aborale ; collerette équatoriale communément nervurée radialement par transparence sous la loupe binoculaire ; en microscopie électronique les nervures apparaissent parfois sur la partie longue de la collerette ; sur la face orale ce sont des stries fines (Vue 77 ![]() ) sur la face aborale ce sont de véritables renforts radiaires (Vue 75
) sur la face aborale ce sont de véritables renforts radiaires (Vue 75 ![]() ) parfois absents (Vue 76
) parfois absents (Vue 76 ![]() ) ; dissymétrie longitudinale, caractéristique de ce taxon :
nervures, allongement du corps et de la collerette équatoriale, ornementation microstructurale sont distinctes à l'avant et à l'arrière.
) ; dissymétrie longitudinale, caractéristique de ce taxon :
nervures, allongement du corps et de la collerette équatoriale, ornementation microstructurale sont distinctes à l'avant et à l'arrière.
Étymologie : L'espèce est nommée d'après la forme allongée de la collerette du latin longus : long et du latin velifer : pourvu d'une voile ; c'est la scutellelle à longue voile.
Holotype A2b : Provenant du niveau +5,0, l'holotype a été coloré ; il est caractérisé par une différenciation antéro-postérieure nette, l'avant de la collerette étant arrondi et l'arrière nettement plus long et anguleux (Planche annexe 1 ![]() , vignette 9) ; inventorié au Musée du site de Tercis sous la référence TCS.G045, il est accompagné de 5 topotypes choisis parmi plusieurs dizaines collectés à ce niveau.
, vignette 9) ; inventorié au Musée du site de Tercis sous la référence TCS.G045, il est accompagné de 5 topotypes choisis parmi plusieurs dizaines collectés à ce niveau.
Discussion : La forme longue distingue la scutellelle à longue voile de toutes les autres gilianelles. Ce contour allongé est largement imprimé par le développement de la collerette mais le corps lui même a un contour elliptique. L'acquisition de la symétrie bilatérale laisse supposer une adaptation hydrodynamique avec une direction privilégiée de déplacement du corps par rapport au fluide. L'observation électronique a révélé qu'il y avait un sens au déplacement relatif. La Vue 71 ![]() obtenue sur un spécimen du même niveau que l'holotype, montre ce qui sera nommé un avant, à droite, plus aigu, plus régulier et, semble-t-il, parfois renforcé sur la face aborale (à gauche sur l4 Vue 75
obtenue sur un spécimen du même niveau que l'holotype, montre ce qui sera nommé un avant, à droite, plus aigu, plus régulier et, semble-t-il, parfois renforcé sur la face aborale (à gauche sur l4 Vue 75 ![]() ). L'arrière de la collerette est plus large et plus découpé (Vues 71 et 76
). L'arrière de la collerette est plus large et plus découpé (Vues 71 et 76 ![]() à gauche). On pense encore à un bord d'attaque (avant) et un bord de fuite (arrière) comme pour une aile d'avion. L'avant du corps lui-même paraît plus long que le côté arrière (côté droit des Vues 71 et 76
à gauche). On pense encore à un bord d'attaque (avant) et un bord de fuite (arrière) comme pour une aile d'avion. L'avant du corps lui-même paraît plus long que le côté arrière (côté droit des Vues 71 et 76 ![]() ; côté gauche de la Vue 75
; côté gauche de la Vue 75 ![]() ) par rapport à l'orifice. Les Vues 74 et 77
) par rapport à l'orifice. Les Vues 74 et 77 ![]() sont plus difficiles à interpréter. On a, là encore (cf. genre Aturella), franchi l'étape d'une adaptation à la vie planctonique avec déplacement dirigé.
sont plus difficiles à interpréter. On a, là encore (cf. genre Aturella), franchi l'étape d'une adaptation à la vie planctonique avec déplacement dirigé.
Distribution : La scutellelle à longue voile (A2b) est présente dès la cote -9,5. Les niveaux de l'intervalle +5,0 / +11,0 en renferment 5 à 10 spécimens par gramme de sédiment traité. On la rencontre facilement jusqu'à la cote 57,9. Au delà, vers les couches plus récentes, les spécimens deviennent exceptionnels dans la fraction plus grande que 100 µm mais elle reste présente dans la fraction plus fine jusqu'à la cote 79,8 puis réapparaît? à la cote 125,5 ; ce taxon ne donne pas de signal biostratigraphique franc. Avec cette forme se termine la description des disques simples sans expansion radiaire.
Genre Corniculum nov. gen.
Étymologie : Le nom de genre dérive du latin corniculum : petit entonnoir, petite corne d'après la forme générale du test.
Espèce type : Corniculum sinuosum nov. sp.
Description : La morphologie du genre pourrait dériver de celle de A2a étirée axialement vers le bas. Il en résulte une collerette équatoriale en entonnoir, un corps lui-même étiré en cône vers le bas et une collerette péri-apicale allongée en ongle ou en pied. Sur ce schéma se greffe une adaptation au déplacement directionnel pour les 2 espèces du genre.
Espèces attribuées au genre : sinuosum, clausum.
Corniculum sinuosum nov. sp. (code L2, ce travail)
Description : Ŕ la loupe binoculaire, test en petit entonnoir (diamètre : 140 µm) avec un rebord transparent, de contour plus ou moins sinueux et un corps terminé par un tube large (diamètre 45 µm) développé d'un seul côté, en ongle ; au microscope électronique, corps constitué d'un cône à pointe tournée vers le bas dont l'angle est proche de 115°, extrémité large du cône fermée par une surface à peine convexe formant la face orale, de diamètre 100 µm, au centre de laquelle se trouve l'orifice (Vue 80 ![]() ) ; rebord externe de cette surface prolongé d'une collerette un peu moins large que le rayon du disque et dirigée vers le haut, dans le prolongement des flancs du cône (Vue 82
) ; rebord externe de cette surface prolongé d'une collerette un peu moins large que le rayon du disque et dirigée vers le haut, dans le prolongement des flancs du cône (Vue 82 ![]() ) ; contour de la collerette équatoriale sinueux (Vue 80
) ; contour de la collerette équatoriale sinueux (Vue 80 ![]() ) ; face aborale formée par le cône terminée par une collerette péri-apicale (Vue 78
) ; face aborale formée par le cône terminée par une collerette péri-apicale (Vue 78 ![]() ) prolongée sur un tiers du diamètre par un pied en gouttière de longueur un peu supérieure au diamètre ; sa direction est déviée de l'axe du cône ; flancs du cône inégalement développés (Vue 82
) prolongée sur un tiers du diamètre par un pied en gouttière de longueur un peu supérieure au diamètre ; sa direction est déviée de l'axe du cône ; flancs du cône inégalement développés (Vue 82 ![]() ) ; ce prolongement partiel de la collerette aborale n'est pas lié à une altération ; il est
toujours situé à l'opposé du flanc le plus long du cône ; nannostructure de la face orale voisine de celle de Scutellella circularis (A2a) avec des franges concentriques jusqu'à l'orifice (Vue 80
) ; ce prolongement partiel de la collerette aborale n'est pas lié à une altération ; il est
toujours situé à l'opposé du flanc le plus long du cône ; nannostructure de la face orale voisine de celle de Scutellella circularis (A2a) avec des franges concentriques jusqu'à l'orifice (Vue 80 ![]() ) ; un bourrelet circulaire sépare la face orale de la collerette équatoriale présentant la même nannostructure que celle de A2a avec de vagues alignements de cristaux rayonnants ; face aborale de la collerette avec des microstructures rayonnantes à peine marquées armant cette expansion qui se développe plus au niveau de ces renforts provoquant le dessin sinueux du contour (Vue 81
) ; un bourrelet circulaire sépare la face orale de la collerette équatoriale présentant la même nannostructure que celle de A2a avec de vagues alignements de cristaux rayonnants ; face aborale de la collerette avec des microstructures rayonnantes à peine marquées armant cette expansion qui se développe plus au niveau de ces renforts provoquant le dessin sinueux du contour (Vue 81 ![]() ) ; nannostructure de la face aborale en franges concentriques.
) ; nannostructure de la face aborale en franges concentriques.
Étymologie : L'espèce tire son nom du latin sinuosum : sinueux d'après le contour de la collerette équatoriale ; c'est le petit entonnoir sinueux.
Holotype L2 : Provenant du niveau 54,9 l'holotype a été taché d'orange (Planche annexe 1 ![]() , vignette 15) ; inventorié au Musée du site de Tercis sous la référence TCS.G046, il est accompagné de 4 topotypes à collerette aborale en anneau plus ou moins prolongé de l'ongle.
, vignette 15) ; inventorié au Musée du site de Tercis sous la référence TCS.G046, il est accompagné de 4 topotypes à collerette aborale en anneau plus ou moins prolongé de l'ongle.
Discussion : Difficile à rattacher à un groupe précédemment décrit (disque ou disque à expansions) ; la parenté la plus probable serait avec les disques à collerette équatoriale pourvus d'une collerette annulaire aborale (la scutellelle circulaire : Code A2a) : la face orale se serait aplatie et la face aborale allongée tandis que la collerette équatoriale se serait légèrement relevée ; le développement partiel de la collerette péri-apicale de A2a aurait conduit à l'aspect en ongle du pied de L2 ; ainsi L2 se rapprocherait (dériverait ?) des disques sans bras ; avec la déviation du cône et du tube aboral, ce taxon a acquis une symétrie bilatérale et un sens, l'arrière se situant dans la direction du pied. On peut y voir une adaptation à un déplacement directionnel.
Distribution : Entre les cotes 50,0 et 65,4, ce petit entonnoir sinueux a laissé des restes dans 5 des 9 niveaux désagrégés ; 16 spécimens ont été prélevés, parfois dans la fraction plus petite que 100 µm ; cet intervalle représente une durée de dépôt de l'ordre de 0,6 Ma à Tercis ; l'espèce est toujours rare mais plus commune à la cote 54,9.
Corniculum clausum nov. sp. (code L3, ce travail)
Description : Ŕ la loupe binoculaire, corps en petit entonnoir aux bords transparents à tube aboral discret, souvent détruit (Planche annexe 1 ![]() , vignette 16) ; au microscope électronique face orale semblable à L2 :
disque plat entouré d'un bourrelet et prolongé d'une collerette développée au contour circulaire, diamètre au niveau du bourrelet faible : 85 µm pour L3 au lieu de 100 µm pour L2 ; face aborale en cône légèrement désaxé, prolongée par une expansion aborale en pied tubulaire deux fois plus long (35 µm) que large ; nannostructure du spécimen observé au microscope électronique rappelant celle de L2 ; collerette équatoriale de même nannostructure que celles de L2, A2 ou A3 ; bourrelet séparant la face orale du corps de la collerette ; pied montrant de vagues alignements longitudinaux donnant l'aspect de cannelures.
, vignette 16) ; au microscope électronique face orale semblable à L2 :
disque plat entouré d'un bourrelet et prolongé d'une collerette développée au contour circulaire, diamètre au niveau du bourrelet faible : 85 µm pour L3 au lieu de 100 µm pour L2 ; face aborale en cône légèrement désaxé, prolongée par une expansion aborale en pied tubulaire deux fois plus long (35 µm) que large ; nannostructure du spécimen observé au microscope électronique rappelant celle de L2 ; collerette équatoriale de même nannostructure que celles de L2, A2 ou A3 ; bourrelet séparant la face orale du corps de la collerette ; pied montrant de vagues alignements longitudinaux donnant l'aspect de cannelures.
Étymologie : Le nom d'espèce dérive du latin claudere : fermer qui rappelle que l'expansion aborale est un pied clos par opposition avec le tube en ongle de L2 ; c'est le petit entonnoir fermé.
Holotype L3 : Provenant du niveau 79,8 le spécimen figuré au microscope électronique à balayage constitue l'holotype (plot VII, spécimen 21) inventorié au Musée du site de Tercis sous la référence TCS.G047, il lui manque une petite portion de la collerette cassée ; 3 topotypes sont disponibles.
Discussion : La similitude de la face orale suggère que L2 et L3 appartiennent au même genre ; créer un autre genre aurait été justifié si l'on avait accordé, comme pour les gilianelles du groupe E ci-après, un caractère cardinal à la nature de l'expansion aborale ; cependant, il est considéré ici que la collerette en tube marquée de traces longitudinales dans sa nannostructure (cas de L2), passe aisément à un pied plein par simple réduction du diamètre puis fusion (cas de L3) ; cette "évolution" possible de A2a à L2 puis de L2 à L3, n'est pas incompatible avec l'ordre dans lequel les trois taxons ont été observés dans la section de Tercis ; ce petit entonnoir fermé est original dans la mesure oů il combine 3 caractères morphologiques appartenant à 3 "groupes" distingués par ailleurs : collerette équatoriale rencontrée chez les gilianelles du groupe A, pied allongé rencontré chez les gilianelles du groupe B, corps en cupule conique rencontré chez les gilianelles du groupe E+M. L'adaptation au déplacement directionnel est la même que pour l'autre espèce du genre.
Distribution : Le petit entonnoir fermé, forme très discrète, a été observé aux niveaux 79,8 et 82,0 ; malgré sa rareté apparente (6 spécimens récoltés à ce jour) et son extension restreinte, il a paru utile de décrire et nommer ce taxon comme exemple du foisonnement des formes des gilianelles.
Genre Gilianella , 2007
Espèce type : Gilianella stellata , 2007
Description : Disque en boîte de cachou, avec des expansions, le plus souvent des bras équatoriaux, parfois un pied aboral, orifice souvent souligné d'une lèvre ; expansions constituées d'arrangements cristallins en lames (6 ou plus) allongées (Vue 102 ![]() ) donnant en coupe un aspect en rosace (Vue 98b
) donnant en coupe un aspect en rosace (Vue 98b ![]() ) avec une lumière qui s'amenuise et disparaît vers l'extrémité distale ; bord de la face orale du test souligné d'un bourrelet externe parfois aussi présent sur la face aborale ; nombre de bras souvent variable dans une espèce, pied unique quand il est présent, généralement contrebalancé par un orifice surélevé sur la face orale.
) avec une lumière qui s'amenuise et disparaît vers l'extrémité distale ; bord de la face orale du test souligné d'un bourrelet externe parfois aussi présent sur la face aborale ; nombre de bras souvent variable dans une espèce, pied unique quand il est présent, généralement contrebalancé par un orifice surélevé sur la face orale.
Espèces attribuées au genre : stellata, major, terbrachialis, tenuibrachialis.
Gilianella stellata , 2007
Synonymie : Code B2a.- et ,
2006, Pl. 2 ![]() .
.
Description : Corps en disque de diamètre 110-140 µm portant des bras développés ; face orale convexe peu élevée bordée d'un bourrelet fin précédé d'un sillon large (Vues 90-91 ![]() ) ; orifice en léger relief à lèvre irrégulièrement développée ; profil typiquement symétrique en boîte de cachou (Vue 95
) ; orifice en léger relief à lèvre irrégulièrement développée ; profil typiquement symétrique en boîte de cachou (Vue 95 ![]() ) montrant un flanc peu élevé entre face orale et face aborale ; équateur peu marqué portant des bras typiquement au nombre de 8 à la cote 50,0 d'oů est tiré l'holotype ; bras grêles, perpendiculaires à l'axe de symétrie ; face aborale présentant un même bourrelet concentrique sub-terminal précédé d'une dépression annulaire (Vues 85 et 89
) montrant un flanc peu élevé entre face orale et face aborale ; équateur peu marqué portant des bras typiquement au nombre de 8 à la cote 50,0 d'oů est tiré l'holotype ; bras grêles, perpendiculaires à l'axe de symétrie ; face aborale présentant un même bourrelet concentrique sub-terminal précédé d'une dépression annulaire (Vues 85 et 89 ![]() ) ; centre de cette face aborale en dôme très bas ; nannostructure en réseau marquée sur les deux faces avec un arrangement radial plus ordonné à la périphérie.
) ; centre de cette face aborale en dôme très bas ; nannostructure en réseau marquée sur les deux faces avec un arrangement radial plus ordonné à la périphérie.
Étymologie : Le nom d'espèce est tiré du latin stellatus : qui a la forme d'une étoile, évoque la forme en étoile du taxon ; c'est la gilianelle en étoile.
Holotype B2a : L'holotype provenant du niveau 50,0 inventorié au Musée du site de Tercis sous la référence TCS.G026 est accompagné de 5 paratypes figurés par (2007), inventoriés sous les références TCS.G027 à 031 et choisis parmi des centaines de spécimens observés.
Discussion : L'espèce type de la famille (?) des gilianelles présente une certaine variabilité instantanée et dans le temps. 91% des spécimens ont 8 bras à la cote 50,0 (sur 130 spécimens observés) pour un corps de diamètre compris entre 110 et 130 µm ; la proportion de spécimens à 9 bras (Vue 89 ![]() ), monte à 97 % à la cote 25,0 oů le diamètre moyen du corps s'élève à 135 µm. Dans le niveau type, 5,5 % des spécimens ont 9 bras, d'autres 10 bras (3,5%) ; à la cote 39,5, 50 % des spécimens ont 9 bras et 45 % ont 8 bras. Le nombre de bras ne doit pas être pris comme critère d'identification. Quant au profil, s'il est généralement symétrique à la cote 50,0 ; il est parfois plus élevé (Vue 88
), monte à 97 % à la cote 25,0 oů le diamètre moyen du corps s'élève à 135 µm. Dans le niveau type, 5,5 % des spécimens ont 9 bras, d'autres 10 bras (3,5%) ; à la cote 39,5, 50 % des spécimens ont 9 bras et 45 % ont 8 bras. Le nombre de bras ne doit pas être pris comme critère d'identification. Quant au profil, s'il est généralement symétrique à la cote 50,0 ; il est parfois plus élevé (Vue 88 ![]() ) pour un spécimen provenant aussi de la cote 50,0 tandis qu'à la cote 25,0 presque tous les spécimens ont un profil élevé (Vue 87
) pour un spécimen provenant aussi de la cote 50,0 tandis qu'à la cote 25,0 presque tous les spécimens ont un profil élevé (Vue 87 ![]() ) du fait de l'élévation en volcan de la face orale. Tout au long de la répartition, un profil sub-symétrique reste le plus commun. La section renferme encore des formes plus grandes (corps de plus de 150 µm en moyenne) aux bras généralement plus nombreux, plus épais, parfois déviés vers le haut tandis qu'un picot voire un véritable pied axial aboral se développe. Ces formes sont exclues du taxon décrit ici pour la seconde fois.
) du fait de l'élévation en volcan de la face orale. Tout au long de la répartition, un profil sub-symétrique reste le plus commun. La section renferme encore des formes plus grandes (corps de plus de 150 µm en moyenne) aux bras généralement plus nombreux, plus épais, parfois déviés vers le haut tandis qu'un picot voire un véritable pied axial aboral se développe. Ces formes sont exclues du taxon décrit ici pour la seconde fois.
Distribution : Gilianella stellata est présente de la cote -5,0 à la cote 57,9. Au delà, des formes voisines sont présentes, toujours exceptionnelles, ce qui ne permet pas de détermination précise. L'abondance atteint plusieurs dizaines de spécimens par gramme de sédiment traité au niveau 50,0.
Gilianella stellata pulvinata nov. ssp.
Synonymie : Code B2b.- et ,
2006, Pl. 2 ![]() .
.
Description : Corps en disque plat, à contour quadrangulaire sur les grands spécimens (ř : 165 µm), ce qui leur donne l'aspect d'un oreiller ; de taille moyenne (diamètre voisin de 150 µm) avec 4 bras régulièrement agencés ; face orale semblable à celle de l'espèce type ; la plupart des spécimens montrent une lèvre développée sur 1/3 seulement de la périphérie de l'orifice, curiosité qui a aussi été observée chez d'autres espèces du groupe B (Vues 90-91 et 93 ![]() ) ; profil symétrique avec bourrelet équatorial portant 4 bras (Vue 92
) ; profil symétrique avec bourrelet équatorial portant 4 bras (Vue 92 ![]() ) bien développés (longueur proche de 50 µm) ; face aborale comme l'espèce type ; nannostructure semblable à celle des autres membres du groupe B.
) bien développés (longueur proche de 50 µm) ; face aborale comme l'espèce type ; nannostructure semblable à celle des autres membres du groupe B.
Étymologie : Le nom de sous-espèce, tiré du latin pulvinatus : en forme de coussin, d'oreiller évoque l'aspect du test ; c'est la gilianelle en oreiller.
Holotype B2b : Provenant du niveau 11,0, le spécimen choisi comme holotype est le plus grand récolté, de contour bien quadrangulaire, avec 4 bras en partie érodés (Planche annexe 1 ![]() , vignette 18) ; inventorié au Musée du site de Tercis sous la référence TCS.G048. Un paratype (TCS.G049) a été coloré, plus petit, moins anguleux mais avec 4 bras mieux préservés (Planche annexe 1
, vignette 18) ; inventorié au Musée du site de Tercis sous la référence TCS.G048. Un paratype (TCS.G049) a été coloré, plus petit, moins anguleux mais avec 4 bras mieux préservés (Planche annexe 1 ![]() , vignette 18) ; ces types sont accompagnés de 6 topotypes.
, vignette 18) ; ces types sont accompagnés de 6 topotypes.
Discussion : Le nombre de bras allié à une belle taille et à un contour carré font de la gilianelle en oreiller une forme reconnaissable. Le test est morphologiquement très proche de celui de l'espèce type. Là oů elle a été observée, on a rencontré au moins un spécimen à 5 bras, ce qui a conduit à n'accorder que le rang de sous-espèce pauvre en bras de la gilianelle en étoile (B2a).
Distribution : Une vingtaine de spécimens de la gilianelle en oreiller ont été récoltés entre les niveaux +5,0 et 18,25 soit durant environ 0,5 Ma. Le niveau 11,0 est le plus riche avec un peu moins d'un spécimen par gramme de sédiment traité.
Gilianella major nov. sp. (code B3a, ce travail)
Description : Corps en disque plus grand (diamètre compris entre 140 et 190 µm) que celui de Gilianella stellata (,
2007) ; face orale souvent élevée, en dôme presque hémisphérique, avec un orifice ouvert au centre d'un rebord en relief (vue 98) ; bras équatoriaux, perpendiculaires à l'axe de symétrie, trapus, régulièrement répartis autour du corps, au nombre de 8 à 10 avec des exceptions en plus ou en moins ; face aborale aussi élevée que la face orale, en dôme, donnant un profil symétrique (Vue 98 ![]() ) ; moignon de pied souvent présent dans l'axe (Vue 99
) ; moignon de pied souvent présent dans l'axe (Vue 99 ![]() ) sans dépasser un relief conique bas ; nannostructure en napperon avec un arrangement en ride concentrique sub-terminale au rebord externe de la face orale surtout ; de ce rebord partent 1 à 3 rides basses rayonnantes qui arment les bras (Vue 97
) sans dépasser un relief conique bas ; nannostructure en napperon avec un arrangement en ride concentrique sub-terminale au rebord externe de la face orale surtout ; de ce rebord partent 1 à 3 rides basses rayonnantes qui arment les bras (Vue 97 ![]() ), les mêmes renforts existent sur la face aborale (Vue 99
), les mêmes renforts existent sur la face aborale (Vue 99 ![]() ).
).
Étymologie : L'espèce tire son nom du latin major : plus grand d'après sa taille relative à celle de l'espèce type du groupe ; c'est la grande gilianelle.
Holotype B3a : Un spécimen provenant du niveau 62,5 et pourvu de 10 bras pratiquement complets a été taché d'orange (Planche annexe 1 ![]() , vignette 19) ; cet holotype est inventorié au Musée du site de Tercis sous la référence TCS.G050 ; son corps, hors les bras, à un diamètre de 170 µm et possède un téton apical sur la face aborale, très localement dégradée ; il est accompagné de 20 topotypes provenant du même lavage dont le diamètre varie de 150 à 190 µm strictement comme pour la même espèce au niveau 54,9.
, vignette 19) ; cet holotype est inventorié au Musée du site de Tercis sous la référence TCS.G050 ; son corps, hors les bras, à un diamètre de 170 µm et possède un téton apical sur la face aborale, très localement dégradée ; il est accompagné de 20 topotypes provenant du même lavage dont le diamètre varie de 150 à 190 µm strictement comme pour la même espèce au niveau 54,9.
Discussion : B3a est un représentant du genre caractérisé par un corps en disque muni de bras ; ce disque est élevé ici ; quand le relief apical abaisse le centre de gravité du test, plus ce relief est net et porté par un dôme élevé, plus la face orale est élevée elle-même et l'orifice exhaussé par rapport au dôme oral ; le critère de taille est essentiel pour distinguer cette espèce ; ce n'est pas une variation dans une population unique ; une statistique sur 61 individus du niveau 54,9 a montré
(Fig. 2 ![]() ) une coupure nette entre deux populations ; c'est ce qui a conduit à proposer une espèce distincte ; le nombre de bras n'est pas un critère de distinction ; dans certains niveaux, B3a a souvent 8 bras, dans d'autres, les spécimens grands ont le plus souvent 10 bras, comme à la cote 79,8.
) une coupure nette entre deux populations ; c'est ce qui a conduit à proposer une espèce distincte ; le nombre de bras n'est pas un critère de distinction ; dans certains niveaux, B3a a souvent 8 bras, dans d'autres, les spécimens grands ont le plus souvent 10 bras, comme à la cote 79,8.
Distribution : La grande gilianelle se rencontre dès la cote 49,0 et jusqu'à la cote 89,0 ; elle réapparaît à la cote 172,0 ; elle succède à B2a et la remplace partiellement mais les deux espèces coexistent pendant près de 10 m soit un peu moins de 0,5 Ma ; son abondance est de 2 à 3 individus par gramme de sédiment traité.
Gilianella major heliotropium nov. ssp. (code B4, ce travail)
Description : Grands spécimens (diamètre 150 à 190 µm) voisins de la forme commune de Gilianella major ; bras nombreux, courts par rapport au corps ; expansion aborale développée (Vue 100 ![]() ) ; tests à l'apparence d'une fleur de tournesol ; face orale sans col net sur les exemplaires observés au microscope électronique à balayage ; profil symétrique, bombé sur les deux faces mais sans col pour faire le pendant du pied ; nombre de bras variant de 10 à 15, déportés vers le haut et non dans le plan équatorial ; nannostructure nette dans tous les spécimens observés ; épaisseur du test du spécimen de la cote 172 réduite à 7-8 µm (coin bas à droite de la Vue 101
) ; tests à l'apparence d'une fleur de tournesol ; face orale sans col net sur les exemplaires observés au microscope électronique à balayage ; profil symétrique, bombé sur les deux faces mais sans col pour faire le pendant du pied ; nombre de bras variant de 10 à 15, déportés vers le haut et non dans le plan équatorial ; nannostructure nette dans tous les spécimens observés ; épaisseur du test du spécimen de la cote 172 réduite à 7-8 µm (coin bas à droite de la Vue 101 ![]() ) alors que les gilianelles ont généralement des tests épais du double ( et ,
2006).
) alors que les gilianelles ont généralement des tests épais du double ( et ,
2006).
Étymologie : Le nom de la sous-espèce est tiré du latin heliotropium : tournesol, d'après l'aspect du test ; c'est la grande gilianelle tournesol.
Holotype B4 : Le spécimen illustré au microscope électronique à balayage et provenant du niveau 172 sert d'holotype (plot VII, spécimen 53) ; inventorié au Musée du site de Tercis sous la référence TCS.G051 ; il était seul à la cote 172.
Discussion : La distinction entre l'espèce type et sa sous-espèce pourvue d'un pied apical grand est renforcée par le nombre de bras élevé et leur orientation vers le haut ; avec le développement du pied, l'équilibre du test par rapport au plan équatorial (avec l'orifice vers le haut) est ainsi maintenu par cette orientation des bras ; un spécimen de la cote 50,0 (Vue 102 ![]() ) est unique par la disposition et le nombre de ses expansions ; c'est une chimère latérale dont une moitié est du type test à 8 bras et l'autre moitié du type test à 15 bras ; le pied développé montre la même organisation que les bras et place ce test dans la variété "tournesol" ; cette sous-espèce peut être considérée comme un membre extrême d'une population variable ; la coupure morphologique n'est pas assez franche pour justifier une espèce distincte.
) est unique par la disposition et le nombre de ses expansions ; c'est une chimère latérale dont une moitié est du type test à 8 bras et l'autre moitié du type test à 15 bras ; le pied développé montre la même organisation que les bras et place ce test dans la variété "tournesol" ; cette sous-espèce peut être considérée comme un membre extrême d'une population variable ; la coupure morphologique n'est pas assez franche pour justifier une espèce distincte.
Distribution : La sous-espèce en tournesol de la grande gilianelle a été rencontrée de la cote 50 à la cote 172 ; elle est ainsi contemporaine de la forme dont elle est une variété ; chaque niveau n'a livré qu'un ou deux tests pour un total observé de 8 spécimens.
Gilianella terbrachialis nov. sp. (code B3d, ce travail)
Description : Corps du test en forme de disque, grand (180-200 µm) ; face orale avec, de l'extérieur vers l'axe un rebord distal plus élevé suivi d'une légère dépression qui se relève pour porter l'orifice entouré d'un col (Vue 103 ![]() ) ; contour circulaire du corps marqué de 3 bras plutôt trapus, courts, répartis à 120° les uns des autres ; profil peu épais, symétrique (Vue 104
) ; contour circulaire du corps marqué de 3 bras plutôt trapus, courts, répartis à 120° les uns des autres ; profil peu épais, symétrique (Vue 104 ![]() ) ; face aborale peu élevée, comme la face orale, sans dôme central ni relief axial sur certains spécimens, avec pied net sur d'autres (Vue 104
) ; face aborale peu élevée, comme la face orale, sans dôme central ni relief axial sur certains spécimens, avec pied net sur d'autres (Vue 104 ![]() ) ; nannostructure en napperon particulièrement nette dans les spécimens observés au microscope électronique (Vues 103 et 105
) ; nannostructure en napperon particulièrement nette dans les spécimens observés au microscope électronique (Vues 103 et 105 ![]() ) ; rebord externe du test renforcé de rides équatoriales.
) ; rebord externe du test renforcé de rides équatoriales.
Étymologie : Le nom d'espèce est composé du latin brachialis : pourvu de bras et ter : trois fois ; c'est la gilianelle à 3 bras.
Holotype B3d : Provenant du niveau 50,0 l'holotype désigné est bien dégagé, le corps à un diamètre de 150 µm (Planche annexe 1 ![]() , vignette 20) ; inventorié au Musée du site de Tercis sous la référence TCS.G052, ce spécimen est accompagné d'un topotype plus grand, encore partiellement inclus dans sa gangue (Planche annexe 1
, vignette 20) ; inventorié au Musée du site de Tercis sous la référence TCS.G052, ce spécimen est accompagné d'un topotype plus grand, encore partiellement inclus dans sa gangue (Planche annexe 1 ![]() , vignette 20).
, vignette 20).
Discussion : La gilianelle à trois bras a une morphologie originale : le corps est toujours très grand et le tamisage à 250 µm les a souvent brisés. Le nombre de bras est constant sur ces grands exemplaires mais on ne peut rejeter l'hypothèse que ces spécimens soient de simples variants localisés dans le temps de l'espèce B3a.
Distribution : Sept spécimens de la gilianelle à trois bras ont été récoltés à Tercis à la cote 50,0 et 1 à la cote 79,8.
Gilianella terbrachialis mutata nov. ssp. (code B3d', ce travail)
Vues 106-107 ![]() et 108
et 108 ![]()
Description : Corps du test semblable à celui de l'espèce type :
disque peu épais, de taille généralement grande (170-190 µm), parfois plus petits (150 µm) ; trois grands bras "primaires" régulièrement répartis autour du test ; bras "annexes" le plus souvent rapprochés par paires et, dans ce cas, petits, d'autres fois uniques et, dans ce cas, seulement un peu plus petits que les 3 bras principaux ; nannostructure (Vue 106 ![]() ) montrant une distinction entre 3 bras primaires dont les rides radiales se branchent sur une ride concentrique du réseau en napperon de la face aborale et bras annexes simplement ajoutés sur le rebord sans s'ancrer aussi loin sur le réseau en napperon dessiné à la surface du spécimen.
) montrant une distinction entre 3 bras primaires dont les rides radiales se branchent sur une ride concentrique du réseau en napperon de la face aborale et bras annexes simplement ajoutés sur le rebord sans s'ancrer aussi loin sur le réseau en napperon dessiné à la surface du spécimen.
Étymologie : La sous-espèce tire son nom du latin mutare : modifier ; c'est la gilianelle à trois bras modifiée.
Holotype B3d' : Provenant du niveau 79,8 un spécimen a été coloré d'orange ; son corps a un diamètre de 170 µm ; entre les trois bras développés et régulièrement répartis, des moignons par paires sont présents (Planche annexe 1 ![]() , vignette 21) ; inventorié au Musée du site de Tercis sous la référence TCS.G053, il est accompagné de 5 topotypes moins complets dont les bras intermédiaires sont uniques ou par paires.
, vignette 21) ; inventorié au Musée du site de Tercis sous la référence TCS.G053, il est accompagné de 5 topotypes moins complets dont les bras intermédiaires sont uniques ou par paires.
Discussion : La différence avec l'espèce type est fondée sur la présence de bras intermédiaires ; le nombre de bras est un critère de distinction généralement peu significatif mais la présence de ces bras annexes est propre à un niveau donné ce qui a conduit à proposer ce taxon.
Distribution : La gilianelle à trois bras modifiée (8 spécimens) a été observée à la cote 79,8.
Gilianella tenuibrachialis nov. sp.
Synonymie : Code B1a.- et ,
2006, Pl. 2 ![]() .
.
Description : Corps en disque de grande taille (diamètre 180 µm), plat, muni de 6 bras peu développés ; face orale plate avec un rebord distal plus élevé suivi d'un large sillon médian peu profond dont ressort un léger dôme aplati au centre duquel se trouve l'orifice (Vue 110 ![]() ) ; profil symétrique (Vue 111
) ; profil symétrique (Vue 111 ![]() ) avec un bourrelet équatorial continu qui porte les expansions équatoriales perpendiculaires à l'axe de symétrie ; face aborale au relief semblable à celui de la face orale, sans microstructure radiale ou rayonnée (Vue 112
) avec un bourrelet équatorial continu qui porte les expansions équatoriales perpendiculaires à l'axe de symétrie ; face aborale au relief semblable à celui de la face orale, sans microstructure radiale ou rayonnée (Vue 112 ![]() ) ; nannostructure en réseau semblable sur les deux faces, semblable elle-même à celle de la scutellelle A3 sur les spécimens de taille usuelle ; la nannostructure bien dégagée chez les spécimens plus petits semble se combler chez les individus plus grands.
) ; nannostructure en réseau semblable sur les deux faces, semblable elle-même à celle de la scutellelle A3 sur les spécimens de taille usuelle ; la nannostructure bien dégagée chez les spécimens plus petits semble se combler chez les individus plus grands.
Étymologie : Le nom d'espèce est tiré du latin tenuis : menu et du latin brachialis : muni de bras ; c'est la gilianelle à bras menus.
Holotype B1a : L'holotype choisi provient du niveau 79,8 ; il a été marqué d'une tache colorée et présente 6 bras bien préservés (Planche annexe 1 ![]() , vignette 17) ; inventorié au Musée du site de Tercis sous la référence TCS.G054, il est accompagné de 10 topotypes, tous avec 6 bras ou leurs traces.
, vignette 17) ; inventorié au Musée du site de Tercis sous la référence TCS.G054, il est accompagné de 10 topotypes, tous avec 6 bras ou leurs traces.
Discussion : La gilianelle à bras menus présente de larges similitudes avec les autres membres du genre quoique l'organisation paraisse plus simple :
Bras discrets, orifice jamais porté par un col. La taille des bras illustrés est en partie sous-estimée du fait d'une détérioration partielle mais ces expansions n'atteignent jamais les dimensions de celles de B2a. Cette simplicité suggérerait une forme plus primitive mais le taxon a été observé dans un niveau plus récent que l'apparition de l'espèce type du genre. Un petit spécimen à 6 bras récolté au niveau 79,8 (Vue 113 ![]() ) révèle une nannostructure bien mieux marquée et différente :
Des cellules quadrangulaires ordonnées concentriquement à la périphérie ; ses autres caractéristiques morphologiques sont identiques par ailleurs mais il pourrait s'agir d'une autre espèce (cf. B3d', Vue 106
) révèle une nannostructure bien mieux marquée et différente :
Des cellules quadrangulaires ordonnées concentriquement à la périphérie ; ses autres caractéristiques morphologiques sont identiques par ailleurs mais il pourrait s'agir d'une autre espèce (cf. B3d', Vue 106 ![]() ).
).
Distribution : La gilianelle à bras menus a été observée à la cote 79,8 oů son abondance est de l'ordre de 1 spécimen par gramme de sédiment traité ; 13 spécimens ont été prélevés.
Genre Caccabella nov. gen.
Étymologie : Le genre tire son nom du latin caccabus : marmite d'après la forme des espèces qui rappelle celle d'un autocuiseur.
Espèce type : Caccabella spinocincta nov. sp.
Description : Test subsphérique de profil symétrique ; face orale avec orifice entouré d'un col en relief, face aborale marquée par une collerette péri-apicale de relief semblable ; 5 à 11 bras équatoriaux de même structure que ceux des gilianelles :
Section formée d'une rosace à 5-6 branches (Vues 120-121 ![]() ) sur l'essentiel de leur longueur.
) sur l'essentiel de leur longueur.
Genre monospécifique avec une sous-espèce type et une morphologiquement légèrement distincte.
Caccabella spinocincta nov. sp.
Synonymie : Code C2.- et ,
2006, Pl. 3 ![]() .
.
Description : Corps sphérique, de taille constante (diamètre 120 à 140 µm) à bras en nombre variable (5-11) dans un niveau et d'un niveau à l'autre ; face orale régulièrement hémisphérique, surmontée d'un col peu élevé entourant l'orifice (Vue 114 ![]() ) ; profil un peu dissymétrique dans la forme, non dans le développement des deux faces difficiles à distinguer à la loupe binoculaire ; les bras s'écartent au niveau de l'équateur au nombre de 7 à 10 dans le niveau d'oů provient l'holotype, plutôt 7 bras au niveau 11,0 ; des spécimens à 6 voire 5 bras existent ; d'autres, aussi rares, ont 11 bras ; face aborale présentant un diamètre plus étroit que celui de la face orale au niveau équatorial (Vue 116
) ; profil un peu dissymétrique dans la forme, non dans le développement des deux faces difficiles à distinguer à la loupe binoculaire ; les bras s'écartent au niveau de l'équateur au nombre de 7 à 10 dans le niveau d'oů provient l'holotype, plutôt 7 bras au niveau 11,0 ; des spécimens à 6 voire 5 bras existent ; d'autres, aussi rares, ont 11 bras ; face aborale présentant un diamètre plus étroit que celui de la face orale au niveau équatorial (Vue 116 ![]() ) ; elle s'élève rapidement puis marque un palier plus ou moins net au centre duquel s'élève une collerette en anneau péri-apical (Vue 120
) ; elle s'élève rapidement puis marque un palier plus ou moins net au centre duquel s'élève une collerette en anneau péri-apical (Vue 120 ![]() ).
).
Étymologie : L'espèce tire son nom du latin cingere : ceindre et du latin spina : épine pour un test caractérisé par des épines aiguës disposées en une ceinture équatoriale ; c'est la caccabelle ceinte d'épines.
Holotype C2 : L'holotype choisi est grand, possède 8 bras, une face orale hémisphérique et une face aborale anguleuse (Planche annexe 1 ![]() , vignette 22) ; il provient du niveau 18,2 et est inventorié au Musée du site de Tercis sous la référence TCS.G055 ; il est accompagné de 9 topotypes choisis parmi une cinquantaine de spécimens collectés à ce niveau.
, vignette 22) ; il provient du niveau 18,2 et est inventorié au Musée du site de Tercis sous la référence TCS.G055 ; il est accompagné de 9 topotypes choisis parmi une cinquantaine de spécimens collectés à ce niveau.
Discussion : L'aspect très anguleux illustré par le profil des Vues 117-118 ![]() est rare : la plupart des spécimens ont une face aborale seulement légèrement anguleuse (Vue 118
est rare : la plupart des spécimens ont une face aborale seulement légèrement anguleuse (Vue 118 ![]() ) ; au niveau 11,0 et là seulement, existe une variante plus symétrique décrite ci-dessous.
) ; au niveau 11,0 et là seulement, existe une variante plus symétrique décrite ci-dessous.
Distribution : La caccabelle ceinte d'épines est présente entre les cotes -5,0 et 32,5 dépôts représentant aux alentours de 1,5 Ma. Elle est assez commune avec 5 à 10 spécimens par gramme de sédiment traité.
Caccabella spinocincta ventriosa nov. ssp. (code C2b, ce travail)
Description : Corps sphérique, de taille constante (diamètre 120 à 140 µm) à bras équatoriaux courts comme la forme type ; profil plus symétrique que la forme type ; bras se détachant au niveau de l'équateur au nombre de 7 le plus souvent (variable de 5 à 8) ; face aborale très semblable avec seulement une très légère réduction du diamètre au niveau équatorial par rapport au diamètre de l'hémisphère oral tandis que le dôme hémisphérique s'aplatit de manière à peine perceptible au pôle (Vue 121 ![]() et 124-125
et 124-125 ![]() ) ; collerette en anneau d'apparence identique à la lèvre péri-orale autour du pôle aboral (Vue 123
) ; collerette en anneau d'apparence identique à la lèvre péri-orale autour du pôle aboral (Vue 123 ![]() ) ; distinction entre les faces délicate même au microscope électronique ; nannostructure avec un maillage régulier sans orientation préférentielle.
) ; distinction entre les faces délicate même au microscope électronique ; nannostructure avec un maillage régulier sans orientation préférentielle.
Étymologie : La sous-espèce tire son nom du latin ventriosus : pansu d'après l'aspect rebondi de la face aborale qui s'oppose à l'aspect anguleux de la forme typique ; c'est la caccabelle pansue.
Holotype C2b : Provenant du niveau 11,0, l'holotype coloré a 7 bras et la face aborale est plus étroite et plus allongée que la face orale (Planche annexe 1 ![]() , vignette 23) ; inventorié au Musée du site de Tercis sous la référence TCS.G056, 33 topotypes l'accompagnent.
, vignette 23) ; inventorié au Musée du site de Tercis sous la référence TCS.G056, 33 topotypes l'accompagnent.
Discussion : Tous les spécimens du même niveau présentent les mêmes caractéristiques de similitude entre les deux faces ; la difficulté à repérer face orale et aborale a d'abord fait croire à une espèce distincte de C2 ; il s'agit d'une variation limitée dans le temps : la forme anguleuse présente auparavant est aussi présente au dessus et le rang de sous-espèce traduit une variation localisée. Avec ce taxon, se termine la description des gilianelles à bras à section en rosace.
Distribution : La caccabelle pansue a été observée à la cote 11,0 oů elle est commune avec environ 10 spécimens par gramme de sédiment traité.
Genre Orculiella nov. gen.
Étymologie : Le nom de genre est dérivé du latin orcula : petite jarre.
Espèce type : Orculiella niduseumenei nov. sp.
Description : Forme en plat à tagine, lié au développement de la face orale avec col très haut sur un corps discoďde ; expansions équatoriales (bras) en bec de canard par 5 ou 6.
Remarques : La morphologie des 2 taxons attribués à l'espèce ne suggère pas une adaptation à la vie planctonique ; le développement du col déplace le centre de gravité vers le pôle oral et le test plongé dans un liquide aurait tendance à se renverser, l'orifice vers le bas. Le col permettrait d'éloigner l'appareil nourricier protoplasmique d'un support sur lequel serait posé le test. Le genre révèle ainsi des caractères suggérant une vie benthique de même type que pour le genre Aturella.
Genre monospécifique avec une variété type et une distincte morphologiquement et stratigraphiquement.
Orculiella niduseumenei nov. sp.
Synonymie : Code C1.- ,
2007,
Fig. 3 ; code C1b.- et ,
2006, Pl. 3 ![]() .
.
Description : Corps en disque épais de diamètre 140 -170 µm ; bras généralement par 5, parfois 6, donnant un contour sub-pentagonal, parfois hexagonal (Vue 130 ![]() ) ; face orale en dôme enflé sur lequel se greffe un col considérable (Vue 127
) ; face orale en dôme enflé sur lequel se greffe un col considérable (Vue 127 ![]() ) terminé par une lèvre péri-orale (Vue 126
) terminé par une lèvre péri-orale (Vue 126 ![]() ) ; équateur sans bourrelet marqué portant des bras souvent détériorés ; bras en épine aplatie dans le plan équatorial, l'extrémité plutôt arrondie ; face aborale régulièrement convexe.
) ; équateur sans bourrelet marqué portant des bras souvent détériorés ; bras en épine aplatie dans le plan équatorial, l'extrémité plutôt arrondie ; face aborale régulièrement convexe.
Étymologie : Le nom d'espèce dérive du latin nidus : nid et de Eumenes : eumène d'après la ressemblance étroite avec les nids de boue de certains hyménoptères solitaires du genre Eumenes, une guêpe maçonne ; c'est la petite jarre en nid d'eumène.
Holotype C1b : L'holotype sélectionné provient du niveau 54,9 ; il est légèrement coloré, grand et pourvu de 6 bras dont 4 un peu érodés (Planche annexe 1 ![]() , vignette 24) ; inventorié au Musée du site de Tercis sous la référence TCS.G057, il est accompagné de 14 topotypes.
, vignette 24) ; inventorié au Musée du site de Tercis sous la référence TCS.G057, il est accompagné de 14 topotypes.
Discussion : Le caractère épais du disque, le col développé font de ce test une forme bien particulière ; parmi la vingtaine de tests récoltés aux niveaux les plus riches (niveaux 54,9 et 62,5) aucun ne montre de dépression au centre de la face aborale comme pour le taxon suivant.
Distribution : Surtout observée entre les cotes 50,0 et 62,5, cette espèce a été trouvée par la suite à divers niveaux entre 39,5 et 69,0 mais presque toujours représentée par un seul spécimen. Son abondance n'approche jamais 1 spécimen par gramme d'échantillon trié et sa durée de vie à Tercis n'atteint pas 1,5 Ma. Cette petite jarre en nid d'eumène provient de la portion moyenne de la section de Tercis ; des spécimens ont ensuite été trouvés plus bas mais ils présentent des caractères distincts qui ont conduit à créer une sous-espèce décrite ci-après.
Orculiella niduseumenei strangulata nov. ssp.
Synonymie : Code C1a.- ,
2007 ; et ,
2006, Pl. 3 ![]() .
.
Description : Corps en disque épais proche de celui de la forme type flanqué de 6 bras courts le plus souvent ; face orale d'abord tronconique sur la moitié du diamètre (Vue 131 ![]() ) puis brusquement marquée par un relief cylindrico-conique qui projette l'orifice circulaire et large de 15 µm, loin de l'équateur du test ; profil très dissymétrique lié au col élevé qui débute par un léger étranglement basal (Vue 131
) puis brusquement marquée par un relief cylindrico-conique qui projette l'orifice circulaire et large de 15 µm, loin de l'équateur du test ; profil très dissymétrique lié au col élevé qui débute par un léger étranglement basal (Vue 131 ![]() ) ; face aborale en dôme bas, déprimé au centre sur la moitié du diamètre du test (voir la section, Vue 133
) ; face aborale en dôme bas, déprimé au centre sur la moitié du diamètre du test (voir la section, Vue 133 ![]() ) ; tous les tests observés à la cote +5,0 montrent cette dépression ; nannostructure, en napperon avec, sur la face orale, un réseau de cellules alignées concentriquement, réseau qui devient radial au niveau du col, c'est à dire longitudinal par rapport à lui (Vue 131
) ; tous les tests observés à la cote +5,0 montrent cette dépression ; nannostructure, en napperon avec, sur la face orale, un réseau de cellules alignées concentriquement, réseau qui devient radial au niveau du col, c'est à dire longitudinal par rapport à lui (Vue 131 ![]() ) ; sur la face aborale, les cellules quadrangulaires sont ordonnées concentriquement au bord distal puis deviennent plus désordonnées dans la dépression centrale ce qui rappelle la face aborale de N1b (Vue 8
) ; sur la face aborale, les cellules quadrangulaires sont ordonnées concentriquement au bord distal puis deviennent plus désordonnées dans la dépression centrale ce qui rappelle la face aborale de N1b (Vue 8 ![]() ).
).
Étymologie : La sous-espèce tire son nom du latin strangulata : resserré d'après la forme de la base du col observée sur cette petite jarre en nid d'eumène étranglée.
Holotype C1a : L'holotype est grand, a été légèrement coloré et provient du niveau +5,0 ; il présente 6 bras dont 4 un peu érodés (Planche annexe 1 ![]() , vignette 25) ; inventorié au Musée du site de Tercis sous la référence TCS.G058, cet holotype est accompagné de 7 topotypes choisis parmi 13 spécimens récoltés à ce niveau.
, vignette 25) ; inventorié au Musée du site de Tercis sous la référence TCS.G058, cet holotype est accompagné de 7 topotypes choisis parmi 13 spécimens récoltés à ce niveau.
Discussion : Les spécimens des niveaux anciens se distinguent de la forme type plus récente par la dépression de la face aborale, le col plus élevé, mieux individualisé par l'étranglement basal mais ces derniers caractères ne sont pas toujours nets et n'encouragent pas à créer une espèce. Ces deux petites jarres terminent la série des tests en disque formés d'une face orale et d'une face aborale.
Distribution : La sous-espèce étranglée de la petite jarre en nid d'eumène a été reconnue dans 4 niveaux anciens de la section de Tercis, entre -5,0 et +18,2 soit pendant environ 1 Ma. L'abondance est faible à très faible, au mieux 1 spécimen par gramme de sédiment traité ; une vingtaine de spécimens ont été récoltés.
Genre Pennigerella nov. gen.
Étymologie : Le genre tire son nom du latin penniger : pourvu d'ailes qui évoque la forme des expansions équatoriales.
Espèce type : Pennigerella palmata nov. sp.
Description : Corps subdivisé en 3 unités morphologiques au contraire de tous les précédents et comme tous les suivants : face orale et face aborale subdivisée en flanc et surface péri-apicale ; expansions aplaties en lame 1- à la périphérie de la face orale et réunies par une palmure, 2- à la périphérie de la surface péri-apicale ou 3- principalement dans un seul des deux endroits.
Remarques : Ce genre appartient à un ensemble de formes distinctes des gilianelles précédentes (codes A, B et C). Ce nouvel ensemble (comprenant G0 et les groupes informels codés J, H, M décrits plus loin) pourrait dériver d'une forme en tonnelet (de type N1b) avec des expansions aux limites entre flanc et faces. Comme pour N1b, le test se subdiviser en face orale, flanc et surface péri-apicale, cette dernière ayant un développement parfois restreint.
Espèces attribuées au genre : palmata (2 sous-espèces), castulata.
Pennigerella palmata nov. sp.
Synonymie : Code J1.- et ,
2006, Pl. 5 ![]() .
.
Description : Corps en cupule tronconique presqu'aussi haute que large ; taille du corps de l'ordre de 100 µm (diamètre : 85-105 µm) ; face orale tronconique, peu élevée, avec une ride nette entourant une dépression péri-orale au fond de laquelle se situe l'orifice lui même entouré d'une lèvre (Vue 139 ![]() ) ; collerette équatoriale marquée formant palmure reliant 4 expansions (bras aplatis en spatule :
Vues 139 et 141
) ; collerette équatoriale marquée formant palmure reliant 4 expansions (bras aplatis en spatule :
Vues 139 et 141 ![]() ) disposées en croix, à peine relevées par rapport au plan équatorial, aussi longues que le rayon du corps ; ensemble aboral essentiellement constitué d'un flanc élevé terminé par une collerette bien développée ; collerette prolongée par 2 expansions (pieds) opposées (Planche annexe 2
) disposées en croix, à peine relevées par rapport au plan équatorial, aussi longues que le rayon du corps ; ensemble aboral essentiellement constitué d'un flanc élevé terminé par une collerette bien développée ; collerette prolongée par 2 expansions (pieds) opposées (Planche annexe 2 ![]() , vignette 26), aplaties en spatule plus large à la base, allant en se rétrécissant et dirigées à environ 45° par rapport à l'axe de symétrie et de longueur atteignant la hauteur du corps ; surface péri-apicale légèrement convexe ; nannostructure formée d'un réseau sur la face orale et les flancs tandis que la surface péri-apicale est grumeleuse.
, vignette 26), aplaties en spatule plus large à la base, allant en se rétrécissant et dirigées à environ 45° par rapport à l'axe de symétrie et de longueur atteignant la hauteur du corps ; surface péri-apicale légèrement convexe ; nannostructure formée d'un réseau sur la face orale et les flancs tandis que la surface péri-apicale est grumeleuse.
Étymologie : Le nom d'espèce dérive du latin palmatus : palmé qui qualifie la disposition des bras reliés par une membrane ; c'est l'ailée palmée.
Holotype J1 : Provenant du niveau +5,0, l'holotype choisi a été coloré :
il possède 4 bras au moins en partie et 1 pied (Planche annexe 2 ![]() , vignette 26) ; inventorié au Musée du site de Tercis sous la référence TCS.G059 ; un paratype 1 a été coloré de même (TCS.G060) et possède ses deux pieds entiers ainsi que 3 des 4 bras, au moins partiellement (Planche annexe 2
, vignette 26) ; inventorié au Musée du site de Tercis sous la référence TCS.G059 ; un paratype 1 a été coloré de même (TCS.G060) et possède ses deux pieds entiers ainsi que 3 des 4 bras, au moins partiellement (Planche annexe 2 ![]() , vignette 26) ; ces types sont accompagnés de 12 topotypes, choisis parmi une vingtaine de spécimens, dont le corps, assez élevé comme sur la Vue 141
, vignette 26) ; ces types sont accompagnés de 12 topotypes, choisis parmi une vingtaine de spécimens, dont le corps, assez élevé comme sur la Vue 141 ![]() , a un diamètre proche de 100 µm.
, a un diamètre proche de 100 µm.
Discussion : En l'absence des expansions (Vue 137 ![]() ), la forme du corps permet à elle seule d'identifier le taxon caractérisé par un flanc assez élevé donnant un aspect trapu au corps. Le test comporte presque toujours au moins les racines des 4 bras. Un spécimen du même niveau, non figuré ici, possède 8 bras distincts, régulièrement répartis au pourtour de l'équateur du test et un corps trapu ; il est associé à ce taxon car le corps est haut et possède 2 pieds développés. Cette caractéristique, corps haut, a conduit à distinguer une sous-espèce alta (du latin altus : haut) distinguée de la sous-espèce suivante.
), la forme du corps permet à elle seule d'identifier le taxon caractérisé par un flanc assez élevé donnant un aspect trapu au corps. Le test comporte presque toujours au moins les racines des 4 bras. Un spécimen du même niveau, non figuré ici, possède 8 bras distincts, régulièrement répartis au pourtour de l'équateur du test et un corps trapu ; il est associé à ce taxon car le corps est haut et possède 2 pieds développés. Cette caractéristique, corps haut, a conduit à distinguer une sous-espèce alta (du latin altus : haut) distinguée de la sous-espèce suivante.
Distribution : L'ailée palmée haute est présente aux niveaux 1,1 et 5,0 oů elle est rare (1 à 2 individus/g de sédiment traité). Un spécimen probable a été récolté à la cote 14,8 (corps haut, 5 bras et 2 pieds en rame) mais, dans ce niveau, la plupart des spécimens relèvent d'une forme moins élevée décrite ci-après.
Pennigerella palmata humilis nov. ssp. (code J2a, ce travail)
Synonymie : Code J2.- et ,
2006, Pl. 5 ![]() .
.
Description : Taille voisine de la sous-espèce précédente (diamètre du corps autour de 100 µm) ; corps bien moins haut que large ; expansions équatoriales comprenant 5 à 8 bras à peine relevés par rapport au plan équatorial, nettement reliés par une palmure et disposés de façon anarchique (Vues 145 et 148-149 ![]() ), translucides à la loupe binoculaire ; collerette péri-apicale peu développée avec expansions douteuses (Vue 149
), translucides à la loupe binoculaire ; collerette péri-apicale peu développée avec expansions douteuses (Vue 149 ![]() ) peut-être au nombre de 2 et longues de 25 à 35 µm.
) peut-être au nombre de 2 et longues de 25 à 35 µm.
Étymologie : Le nom de sous-espèce dérive du latin humilis : bas, peu élevé, qui qualifie la faible hauteur du corps ; c'est l'ailée palmée basse.
Holotype J2a : Provenant du niveau 11,0, l'holotype choisi a été coloré ; il possède 6 bras disposés irrégulièrement ou leur trace (Planche annexe 2 ![]() , vignette 27) ; inventorié au Musée du site de Tercis sous la référence TCS.G061, il est accompagné de 7 topotypes aux bras bien développés et aux expansions aborales toujours peu claires choisis parmi une trentaine de spécimens récoltés à ce niveau.
, vignette 27) ; inventorié au Musée du site de Tercis sous la référence TCS.G061, il est accompagné de 7 topotypes aux bras bien développés et aux expansions aborales toujours peu claires choisis parmi une trentaine de spécimens récoltés à ce niveau.
Discussion : Dans les niveaux oů elle a été observée, l'ailée basse s'identifie aisément par l'aspect aplati du test, bien moins haut que J1 et la disposition irrégulière remarquable des bras. Dans le même niveau que le type, 1 individu resté unique, à 4 bras, a été observé (Vues 142-143 ![]() ) ; son corps très aplati suggère de l'associer plutôt à J2a. La question s'est posée de créer 1 ou 2 taxons ; l'aspect général lors des tris est cependant bien distinct (voir notamment les vignettes 26 et 27 de la Planche annexe 2
) ; son corps très aplati suggère de l'associer plutôt à J2a. La question s'est posée de créer 1 ou 2 taxons ; l'aspect général lors des tris est cependant bien distinct (voir notamment les vignettes 26 et 27 de la Planche annexe 2 ![]() ) :
Un corps plutôt en boule pour J1 et plutôt en plaquette pour J2a.
) :
Un corps plutôt en boule pour J1 et plutôt en plaquette pour J2a.
Distribution : L'ailée basse est présente entre les cotes 11,0 et 25,0 oů l'on peut en récolter 1 à 2 par gramme de sédiment traité. Des restes incertains ont été suspectés aux cotes +5,0 et 39,5 ; la durée d'existence n'a pas dépassé 1,5 Ma à Tercis.
Pennigerella castulata nov. sp. (code J2b, ce travail)
Description : Corps en cupule tronconique peu élevée ; diamètre voisin de 85 µm, hauteur d'environ 45 µm ; face orale sub-plane avec une lèvre péri-orale nette (Vue 151 ![]() ), de diamètre presque moitié de celui du corps ; collerette équatoriale forte prolongée d'expansions nombreuses (environ 10), courtes donnant une apparence festonnée plutôt que celle d'expansions bien individualisées, dirigée franchement quoique modérément vers le haut (Vue 152
), de diamètre presque moitié de celui du corps ; collerette équatoriale forte prolongée d'expansions nombreuses (environ 10), courtes donnant une apparence festonnée plutôt que celle d'expansions bien individualisées, dirigée franchement quoique modérément vers le haut (Vue 152 ![]() ) ; flanc du corps tronconique, très légèrement convexe et terminé par une puissante collerette continue à la base qui se scinde en plusieurs pans peu individualisés dont la longueur dépasse la hauteur du corps ; nannostructure en réseau indistinct sur la face orale (Vue 150
) ; flanc du corps tronconique, très légèrement convexe et terminé par une puissante collerette continue à la base qui se scinde en plusieurs pans peu individualisés dont la longueur dépasse la hauteur du corps ; nannostructure en réseau indistinct sur la face orale (Vue 150 ![]() ) tandis qu'un arrangement radial bien ordonné caractérise les flancs.
) tandis qu'un arrangement radial bien ordonné caractérise les flancs.
Étymologie : Le nom d'espèce dérive du latin castulatus : pourvu d'un jupon, d'une jupe qui évoque la forme de l'expansion aborale ; c'est l'ailée à jupon.
Holotype J2b : Provenant du niveau 11,0 l'holotype choisi a été coloré ; son corps, petit, possède de bonnes expansions avec une collerette orale découpée, étendue et un "jupon" allongé avec 2 pans nets d'un même côté (comme sur la Vue 154 ![]() , cf. Planche annexe 2
, cf. Planche annexe 2 ![]() , vignette 28) ; inventorié au Musée du site de Tercis sous la référence TCS.G062, il est accompagné de 3 topotypes.
, vignette 28) ; inventorié au Musée du site de Tercis sous la référence TCS.G062, il est accompagné de 3 topotypes.
Discussion : Le jupon formé de 2 (ou 3) lobes caractérise sans équivoque cette espèce qui possède ainsi à la fois une collerette équatoriale et une collerette péri-apicale développées ; ce taxon, le plus rare retenu ici, a été observé à 3 niveaux avec une morphologie parfaitement identifiable, parfois dans la fraction plus petite que 100 µm.
Distribution : L'ailée à jupon est présente aux cotes 1,1 et 5,0 (1 spécimen) et 11,0 (5 spécimens récoltés) soit sur une durée de dépôt de l'ordre d'1/4 de Ma.
Genre Corbella nov. gen.
Étymologie : Le nom de genre est dérivé du latin corbis : corbeille.
Espèce type : Corbella vitilis nov. sp.
Description : Corps cupulaire sub-sphérique au contraire du genre précédent ou du suivant plus tronconiques en forme de corbeille couverte; bras (5-6) à section cylindrique au contraire du genre précédent ; pieds insérés sur une collerette péri-apicale par deux et de section cylindrique passant à des rames pour les spécimens les plus récents.
Espèce unique comprenant une forme type et une sous-espèce morphologiquement et stratigraphiquement distincte.
Corbella vitilis nov. sp.
Synonymie : Code G0.- et ,
2006, Pl. 4 ![]() .
.
Description : Corps en forme de cupule sub-sphérique, petite (diamètre 110 à 120 µm) ; face orale en dôme interrompu par une dépression péri-orale de diamètre 35 µm entourée d'une lèvre peu élevée (Vue 155 ![]() ) ; orifice situé au fond de cette dépression ; équateur marqué par une légère rupture angulaire (Vues 156-157
) ; orifice situé au fond de cette dépression ; équateur marqué par une légère rupture angulaire (Vues 156-157 ![]() ) d'oů partent 6 bras (Vue 158
) d'oů partent 6 bras (Vue 158 ![]() ), parfois 5 ou 7 (Vue 155
), parfois 5 ou 7 (Vue 155 ![]() ) selon les individus ; bras cylindriques fins, un peu relevés vers le haut, translucides à la loupe binoculaire (Planche annexe 2
) selon les individus ; bras cylindriques fins, un peu relevés vers le haut, translucides à la loupe binoculaire (Planche annexe 2 ![]() , vignette 29) et régulièrement répartis autour du test, de taille atteignant 40 µm, de section circulaire ; la plupart des individus n'ont conservé que des moignons d'expansions comme illustré sur tous les tests présentés ; ensemble aboral en dôme élevé ; limite entre flanc et surface péri-apicale marquée par une modeste collerette de diamètre proche de 60 µm (Vue 158
, vignette 29) et régulièrement répartis autour du test, de taille atteignant 40 µm, de section circulaire ; la plupart des individus n'ont conservé que des moignons d'expansions comme illustré sur tous les tests présentés ; ensemble aboral en dôme élevé ; limite entre flanc et surface péri-apicale marquée par une modeste collerette de diamètre proche de 60 µm (Vue 158 ![]() ) ; cette collerette porte 2 expansions (pieds), de même longueur et forme que les bras (Vue 159
) ; cette collerette porte 2 expansions (pieds), de même longueur et forme que les bras (Vue 159 ![]() ), dirigées franchement vers le bas du test sans être parallèles à l'axe de symétrie (Vue 157
), dirigées franchement vers le bas du test sans être parallèles à l'axe de symétrie (Vue 157 ![]() ) ; surface péri-apicale (entourée de la collerette) légèrement convexe ; face orale et flanc caractérisés par une nannostructure formant un réseau régulier tandis que la surface péri-apicale est grumeleuse ; sans microstructure élevée, le test laisse deviner un arrangement concentrique des cellules du réseau à la loupe binoculaire.
) ; surface péri-apicale (entourée de la collerette) légèrement convexe ; face orale et flanc caractérisés par une nannostructure formant un réseau régulier tandis que la surface péri-apicale est grumeleuse ; sans microstructure élevée, le test laisse deviner un arrangement concentrique des cellules du réseau à la loupe binoculaire.
Étymologie : Le nom d'espèce évoque le dessin net de la nannostructure sur le corps (du latin vitilis : tressé, de la même famille que vitilia : vannerie) ; c'est la corbeille tressée.
Holotype G0 : Provenant du niveau 11,0, l'holotype coloré a conservé 6 bras et 1 pied sur 2 (Planche annexe 2 ![]() , vignette 29) ; inventorié au Musée du site de Tercis sous la référence TCS.G063, il est accompagné de 7 topotypes sur lesquels ne peuvent être observées qu'une, deux ou trois expansions complètement conservées.
, vignette 29) ; inventorié au Musée du site de Tercis sous la référence TCS.G063, il est accompagné de 7 topotypes sur lesquels ne peuvent être observées qu'une, deux ou trois expansions complètement conservées.
Discussion : Les expansions sont rarement préservées sur ce taxon ; cependant, un petit nombre de spécimens a montré quelques bras ou pieds très allongés et il n'est pas exclu que ces expansions aient dépassé les 40 µm observés jusqu'ici, sur le test initial. La forme sub-sphérique, la taille et l'absence de microstructure (si ce n'est la lèvre péri-orale) rendent ce taxon aisé à identifier bien qu'une mauvaise préservation puisse le faire confondre avec des tests du groupe J eux aussi mal préservés (cf. Vues 137-138 ![]() ). Des spécimens observés à la cote 25,0 (une dizaine) présentent déjà des pieds aplatis mais, dans l'ensemble, les expansions restent grêles et l'on en récolte environ 1 spécimen par gramme de sédiment à ce niveau.
). Des spécimens observés à la cote 25,0 (une dizaine) présentent déjà des pieds aplatis mais, dans l'ensemble, les expansions restent grêles et l'on en récolte environ 1 spécimen par gramme de sédiment à ce niveau.
Distribution : Taxon présent dans les niveaux bas du site entre les cotes 1,1 et 25,0 et notamment à la cote 11,0 oů 15 spécimens ont été récoltés, la corbeille tressée est toujours rare avec moins d'un individu par gramme de sédiment traité pour la fraction supérieure à 100 µm ; un examen à la cote 1,1 a montré que la majorité des tests passait à travers le tamis à mailles de 100 µm pour être retenu par celui de 80 µm.
Corbella vitilis biremis nov. ssp. (code G0b, ce travail)
Description : Caractéristiques générales semblables à celles de Corbella vitilis ; corps subsphérique de diamètre voisin de 100 µm (Vue 163 ![]() ) ; face orale en dôme avec un sommet couronné d'un relief annulaire plus fort que chez la forme type, de diamètre extérieur 50 µm autour d'une dépression au fond de laquelle se situe l'orifice (Vue 162
) ; face orale en dôme avec un sommet couronné d'un relief annulaire plus fort que chez la forme type, de diamètre extérieur 50 µm autour d'une dépression au fond de laquelle se situe l'orifice (Vue 162 ![]() ) ; l'anneau ne borde pas directement l'orifice ; 7 à 9 bras cylindriques plus robustes que chez la forme type (Vue 163
) ; l'anneau ne borde pas directement l'orifice ; 7 à 9 bras cylindriques plus robustes que chez la forme type (Vue 163 ![]() ) partant du rebord externe de cette surface et se dirigeant vers le haut avec un angle de 15 à 20° par rapport au plan équatorial ; flancs enflés, en forme de corbeille circulaire et terminés vers le bas, par une collerette épaisse en anneau plus nette que chez la forme type ; il s'en détache 2 pieds larges, en rames, opposés l'un à l'autre et dirigés vers le bas en s'écartant d'environ 30° par rapport à l'axe de symétrie (Vues 161 et 163
) partant du rebord externe de cette surface et se dirigeant vers le haut avec un angle de 15 à 20° par rapport au plan équatorial ; flancs enflés, en forme de corbeille circulaire et terminés vers le bas, par une collerette épaisse en anneau plus nette que chez la forme type ; il s'en détache 2 pieds larges, en rames, opposés l'un à l'autre et dirigés vers le bas en s'écartant d'environ 30° par rapport à l'axe de symétrie (Vues 161 et 163 ![]() d'un test oů n'est préservée qu'une rame) ; nannostructure en cercles concentriques de cellules allongées radialement entre la lèvre et les bras de la face orale ; flancs avec la même nannostructure évoquant un tressage.
d'un test oů n'est préservée qu'une rame) ; nannostructure en cercles concentriques de cellules allongées radialement entre la lèvre et les bras de la face orale ; flancs avec la même nannostructure évoquant un tressage.
Étymologie : Le nom de la sous-espèce est dérivé du latin biremis : pourvu de deux rames ; c'est la corbeille birème.
Holotype G0b : Provenant du niveau 54,9 l'holotype possède 7 bras dont 3 sont partiels et 2 pieds en rame ; 3 vues en sont données (Planche annexe 2 ![]() , vignette 30) ; inventorié au Musée du site de Tercis sous la référence TCS.G064.
, vignette 30) ; inventorié au Musée du site de Tercis sous la référence TCS.G064.
Discussion : Le caractère robuste des expansions de cette forme prélevée à certains niveaux, est moins marqué à d'autres niveaux et il n'a pas semblé possible de créer une autre espèce mais la distribution stratigraphique distincte a conduit à distinguer le taxon. Ŕ la loupe binoculaire, les tests de G0 et G0b peuvent être confondus avec ceux de Obbella caryophyllum (code G1, dernier taxon de ce travail) lorsque ces derniers ont perdu leur pied apical à section en croix ; l'examen du pôle aboral au microscope électronique apporte la réponse quand il y a problème ; une autre distinction est le fait que les bras du genre Corbella sont transparents à la loupe binoculaire, ce qui est lié à leur structure en tube plein alors que les expansions du genre Obella sont à section à croix; lorsque les expansions aborales sont altérées, une confusion peut avoir lieu avec H2 (l'espèce Aquilegiella varia, ci-après), l'examen de la face aborale permet d'identifier G0b par une microstructure annulaire plus distincte.
Distribution : La corbeille birème a été remarquée aux niveaux 50,0 et 54,9 (3 spécimens assurés) ; des spécimens sont probables plus bas jusqu'à la cote 39,5.
Genre Aquilegiella nov. gen.
Étymologie : Le nom de genre dérive du latin aquilegia : ancolie (venu de aquilegius : réservoir à eau, plante qui recueille de l'eau dans ses éperons).
Espèce type : Aquilegiella varia nov. sp.
Description : Corps cupulaire, en tronc de cône, face orale plus plane que le genre précédent, face aborale très élevée avec flancs presque cylindriques ou plus coniques ; expansions équatoriales (bras par 5 à 15) cylindriques mais souvent ancrées par une cornière en croix sur le corps ; expansions aborales (par 2 à 6) cylindriques insérées à la périphérie et parfois au centre de la surface péri-apicale.
Espèces attribuées au genre : varia, ericius.
Aquilegiella varia nov. sp.
Synonymie : Code H2.- ,
2007,
Fig. 3 ; et ,
2006, Pl. 4 ![]() .
.
Description : Corps cupulaire, en tronc de cône, de diamètre compris entre 80 et 95 µm ; face orale peu élevée marquée par un anneau concentrique net (Vues 164 ![]() et 173
et 173 ![]() ) de diamètre 1/3 du corps ; orifice central entouré d'une lèvre étroite sur certains spécimens (Vues 165 et 172
) de diamètre 1/3 du corps ; orifice central entouré d'une lèvre étroite sur certains spécimens (Vues 165 et 172 ![]() ) ; équateur marqué par un bourrelet supportant des bras, généralement 7, parfois 8 ou 9 (Vue 170
) ; équateur marqué par un bourrelet supportant des bras, généralement 7, parfois 8 ou 9 (Vue 170 ![]() ), seulement 6 sur un petit spécimen (Vue 169
), seulement 6 sur un petit spécimen (Vue 169 ![]() ) ; bras cylindriques, de longueur atteignant celle du rayon, dirigés vers le haut avec un angle de 30 à 45° par rapport à l'équateur ; flancs du corps se rétrécissant modérément vers la base ; le plus souvent 2 mais jusqu'à 5 pieds longs comme le corps, ancrés sur un anneau peu distinct (Vue 167
) ; bras cylindriques, de longueur atteignant celle du rayon, dirigés vers le haut avec un angle de 30 à 45° par rapport à l'équateur ; flancs du corps se rétrécissant modérément vers la base ; le plus souvent 2 mais jusqu'à 5 pieds longs comme le corps, ancrés sur un anneau peu distinct (Vue 167 ![]() ), dirigées franchement vers le bas et un peu plus développées que les bras ; pas de microstructure hormis l'anneau de la face orale ; nannostructure dessinant des rectangles alignés concentriquement et radialement sur les deux faces.
), dirigées franchement vers le bas et un peu plus développées que les bras ; pas de microstructure hormis l'anneau de la face orale ; nannostructure dessinant des rectangles alignés concentriquement et radialement sur les deux faces.
Étymologie : Le nom d'espèce dérive du latin varius : variable qui évoque le nombre d'expansions variable ; c'est la petite ancolie variable.
Holotype H2 : Provenant du niveau 54,9, l'holotype a été coloré ; il a 9 bras (2 sont détériorés) et 4 pieds (Planche annexe 2 ![]() , vignette 31) ; inventorié au Musée du site de Tercis sous la référence TCS.G065, 8 topotypes l'accompagnent.
, vignette 31) ; inventorié au Musée du site de Tercis sous la référence TCS.G065, 8 topotypes l'accompagnent.
Discussion : Le spécimen qui possédait le plus grand nombre de bras (probablement 15), montre une lèvre péri-orale nette sur un corps grand (Vues 165-166 ![]() ) ; ses pieds, 2 ou 3 dont 1 seul a été préservé, étaient très longs ; il est rattaché à la même espèce de même que le petit spécimen à 6 bras (Vue 169
) ; ses pieds, 2 ou 3 dont 1 seul a été préservé, étaient très longs ; il est rattaché à la même espèce de même que le petit spécimen à 6 bras (Vue 169 ![]() ) ; ces petits spécimens, plus délicats et au corps moins élevé, peuvent être récoltés en nombre dans la fraction inférieure à 100 µm ; ils constituent peut-être un stade de développement antérieur parmi une population au nombre d'expansions variable, lequel n'est pas fonction de la taille ainsi, l'un des spécimens de moins de 100 µm possède 8 bras et 5 pieds.
) ; ces petits spécimens, plus délicats et au corps moins élevé, peuvent être récoltés en nombre dans la fraction inférieure à 100 µm ; ils constituent peut-être un stade de développement antérieur parmi une population au nombre d'expansions variable, lequel n'est pas fonction de la taille ainsi, l'un des spécimens de moins de 100 µm possède 8 bras et 5 pieds.
Distribution : La petite ancolie variable est présente entre les cotes 50,0 et 79,8, ce qui représente une durée d'existence proche de 1,2 Ma. Apparemment peu commune, elle est plus abondante au niveau 57,9 sans atteindre 1 individu par gramme de sédiment traité ; une centaine de spécimens ont été récoltés au total dans la fraction supérieure à 100 µm. Mais dans la seule fraction 80-100 µm, l'équivalent de 7 spécimens par gramme a été récolté au niveau 74,8 lors d'un test sur un seul trieur de 50 mg ce qui laisse supposer que l'espèce pourrait être commune.
Aquilegiella ericius nov. sp. (code H5, ce travail)
Description : Corps petit, de diamètre 50 à 65 µm ; aspect de spicule multi-axones ; corps en cylindre aussi haut que large seulement distinct au microscope électronique ; face orale basse, avec un orifice de diamètre voisin de 15 µm (Vue 180 ![]() ), entouré d'une lèvre (Vue 175
), entouré d'une lèvre (Vue 175 ![]() ) ; crêtes radiales peu marquées aboutissant chacune à un bras sur lequel elles se poursuivent (Vue 182
) ; crêtes radiales peu marquées aboutissant chacune à un bras sur lequel elles se poursuivent (Vue 182 ![]() ) ; 5 à 7 bras de taille inégale, pleins avec une section en croix à la base puis circulaire distalement (Vue 180
) ; 5 à 7 bras de taille inégale, pleins avec une section en croix à la base puis circulaire distalement (Vue 180 ![]() , bras de face à droite) ; flancs dépourvus de côte ; 5 pieds forts à la base des flancs, presque perpendiculaires à l'axe de symétrie
(Vues 175-176
, bras de face à droite) ; flancs dépourvus de côte ; 5 pieds forts à la base des flancs, presque perpendiculaires à l'axe de symétrie
(Vues 175-176 ![]() ) et disposés régulièrement sur le pourtour avec un angle moyen, entre eux, de 72° (Vue 177
) et disposés régulièrement sur le pourtour avec un angle moyen, entre eux, de 72° (Vue 177 ![]() ) ; pied axial fort (Vue 179
) ; pied axial fort (Vue 179 ![]() d'un individu dont les 5 pieds des flancs sont détériorés) ; nannostructure des flancs longitudinale.
d'un individu dont les 5 pieds des flancs sont détériorés) ; nannostructure des flancs longitudinale.
Étymologie : Le nom d'espèce dérive du latin ericius : cheval de frise (dispositif multiaxe que César utilisait lors des sièges) ; c'est le petit cheval de frise.
Holotype H5 : Provenant du niveau 50,0, l'holotype coloré possède 6 bras inégaux dont 2 détériorés, 5 pieds intermédiaires bien répartis en étoile et le pied aboral, tous solides (Planche annexe 2 ![]() , vignette 32). Inventorié au Musée du site de Tercis sous la référence TCS.G066, 9 topotypes accompagnent ce spécimen.
, vignette 32). Inventorié au Musée du site de Tercis sous la référence TCS.G066, 9 topotypes accompagnent ce spécimen.
Discussion : Cette gilianelle ne se comprend que si l'on repère l'orifice qui permet d'orienter le test. Les Vues 175-181 ![]() montrent la forme usuelle. Un spécimen unique plus petit que la forme type est figuré (Vues 182-183
montrent la forme usuelle. Un spécimen unique plus petit que la forme type est figuré (Vues 182-183 ![]() ). Doté de 5 bras, la face orale ne montre pas de lèvres mais les crêtes radiales sont distinctes ; ces crêtes peuvent aussi être interprétées comme des racines des bras. Ce taxon a posé un problème de classification. Aucune gilianelle décrite jusqu'ici ne montrait une lèvre péri-orale et des crêtes radiales. Or, ce caractère morphologique marquant est commun à tout un groupe de formes décrit ci-dessous et qui constitue un ensemble homogène. Par ailleurs, H5 avait bon nombre de caractères très proches de ceux de la petite ancolie variable. Il pourrait s'agir d'une convergence imparfaite dans la mesure oů les microstructures radiales (Vue 182
). Doté de 5 bras, la face orale ne montre pas de lèvres mais les crêtes radiales sont distinctes ; ces crêtes peuvent aussi être interprétées comme des racines des bras. Ce taxon a posé un problème de classification. Aucune gilianelle décrite jusqu'ici ne montrait une lèvre péri-orale et des crêtes radiales. Or, ce caractère morphologique marquant est commun à tout un groupe de formes décrit ci-dessous et qui constitue un ensemble homogène. Par ailleurs, H5 avait bon nombre de caractères très proches de ceux de la petite ancolie variable. Il pourrait s'agir d'une convergence imparfaite dans la mesure oů les microstructures radiales (Vue 182 ![]() ) ne semblent pas se greffer effectivement sur la lèvre péri-orale. H5 a donc été rangé dans le même genre que le précédent tout en le plaçant au voisinage des gilianelles à lèvre péri-orale d'oů partent des crêtes radiales ci-dessous.
) ne semblent pas se greffer effectivement sur la lèvre péri-orale. H5 a donc été rangé dans le même genre que le précédent tout en le plaçant au voisinage des gilianelles à lèvre péri-orale d'oů partent des crêtes radiales ci-dessous.
Distribution : La distribution du petit cheval de frise reste à préciser ; elle a été bien établie entre les niveaux 50,0 et 57,9. Un curieux spicule prélevé au niveau 11,0 (il y a des spicules multi-axes tout le long de la section) s'est aussi révélé être un spécimen de H5 au microscope électronique. L'abondance de l'espèce est toujours inférieure à 1 individu par gramme de sédiment traité.
Genre Pocillella nov. gen.
Étymologie : Le nom de genre dérive du latin pocillum : tasse à boire d'après la forme générale des espèces.
Espèce type : Pocillella grandicula nov. sp.
Description : Corps en tasse plus ou moins élevée, aux flancs cylindriques, renflés ou tronconiques ; face orale plus développée que la surface péri-apicale toujours présente ; expansion de type collerette équatoriale ou simples picots à section en croix au sommet des flancs ; collerette péri-apicale réduite à la base des flancs. Les espèces se différencient par leur taille, la forme du corps depuis une sphère jusqu'à un cylindre allongé et le développement relatif des collerettes équatoriale et péri-apicale. Chaque population est morphologiquement distincte des autres malgré leur caractère contemporain.
Remarques : Ce genre inaugure la description d'un ensemble de genres caractérisés par un corps cupulaire, en tronc de cône, à face orale légèrement concave présentant un orifice entouré d'une lèvre d'oů partent des crêtes radiales (microstructure orale "en soleil") branchées ou non sur les côtes longitudinales des flancs.
Espèces attribuées au genre : grandicula, minuta, cylindrata, cochleare.
Pocillella grandicula nov. sp.
Synonymie : Code E3a.- et ,
2006, Pl. 4 ![]() .
.
Description : Corps en tronc de cône d'aspect général en pot à fleur ; hauteur et grand diamètre égaux à 150 µm ou plus ; face orale légèrement concave avec un orifice entouré d'une lèvre d'oů partent 9-11 crêtes radiales terminées par de discrètes pointes au niveau d'une collerette équatoriale étroite (Vue 184 ![]() ) ; flancs du corps presque cylindriques parcourus de côtes bien visibles à la loupe binoculaire ; après un léger rétrécissement, la base des flancs se prolonge d'une collerette plus développée que la collerette équatoriale ; (Vue 186
) ; flancs du corps presque cylindriques parcourus de côtes bien visibles à la loupe binoculaire ; après un léger rétrécissement, la base des flancs se prolonge d'une collerette plus développée que la collerette équatoriale ; (Vue 186 ![]() ) le prolongement d'une partie des côtes la parcourt ; la surface péri-apicale présente des côtes radiales (vue 188) ; nannostructure en cellules rectangulaires alignées le long des flancs (vue 185) ; la présence d'un orifice sur la face orale et son absence au niveau de la surface péri-apicale est illustrée Vue 186
) le prolongement d'une partie des côtes la parcourt ; la surface péri-apicale présente des côtes radiales (vue 188) ; nannostructure en cellules rectangulaires alignées le long des flancs (vue 185) ; la présence d'un orifice sur la face orale et son absence au niveau de la surface péri-apicale est illustrée Vue 186 ![]() .
.
Étymologie : Le nom d'espèce dérive du latin grandiculus : assez grand qui distingue cette espèce des autres du même genre ; c'est la petite tasse assez grande.
Holotype E3a : Provenant du niveau 11,0, l'holotype choisi a été teinté ; le corps est petit avec collerette orale et collerette péri-apicale visibles (Planche annexe 2 ![]() , vignette 33) ; inventorié au Musée du site de Tercis sous la référence TCS.G067, il est accompagné de 9 topotypes.
, vignette 33) ; inventorié au Musée du site de Tercis sous la référence TCS.G067, il est accompagné de 9 topotypes.
Discussion : Un corps sub-cylindrique, parfois plus nettement conique, terminé par une collerette aux deux extrémités caractérise ce taxon ainsi que sa taille toujours grande pour le genre.
Distribution : La petite tasse assez grande est présente à la cote 11,0 oů elle est rare mais a été récoltée de façon répétitive avec une abondance de 0,75 spécimen par gramme de sédiment traité. Aux cotes 14,8 et 18,25 un seul spécimen a été observé malgré des recherches renouvelées.
Pocillella minuta nov. sp.
Synonymie : Code E3b.- et ,
2006, Pl. 4 ![]() .
.
Description : Corps sphérique petit (diamètre = hauteur : 115 à 120 µm, parfois moins de 100 µm), côtelé à la loupe binoculaire ; face orale à orifice entouré d'une lèvre étroite d'oů partent 8-11 crêtes radiales (Vue 189 ![]() ) aboutissant à de discrets picots périphériques ; pas de collerette équatoriale ; flancs enflés parcourus d'environ 24 côtes ; étranglement vers le bas du corps prolongé d'une collerette formant une courte jupe évasée (Vue 191
) aboutissant à de discrets picots périphériques ; pas de collerette équatoriale ; flancs enflés parcourus d'environ 24 côtes ; étranglement vers le bas du corps prolongé d'une collerette formant une courte jupe évasée (Vue 191 ![]() , forme la plus commune) parfois érodée (Vue 190
, forme la plus commune) parfois érodée (Vue 190 ![]() ), parfois plus grande que sur la plupart des spécimens illustrés (Vue 192
), parfois plus grande que sur la plupart des spécimens illustrés (Vue 192 ![]() , taille jusqu'à 1/3 de la hauteur du corps) et délimitant une surface péri-apicale grande à structures radiales discrètes ; expansion courte, crêtée, parfois visible au centre de la surface péri-apicale, (Vues 191-193
, taille jusqu'à 1/3 de la hauteur du corps) et délimitant une surface péri-apicale grande à structures radiales discrètes ; expansion courte, crêtée, parfois visible au centre de la surface péri-apicale, (Vues 191-193 ![]() ).
).
Étymologie : Le nom d'espèce dérive du latin minutus : minuscule par comparaison avec la forme précédente ; c'est la tasse minuscule.
Holotype E3b : Provenant du niveau 11,0, un petit spécimen a été choisi comme holotype et coloré ; il est caractérisé par une collerette aborale très développée mais la face orale est assez mal préservée (Planche annexe 2 ![]() , vignette 34) ; inventorié au Musée du site de Tercis sous la référence TCS.G068 ; un paratype plus grand (TCS.G069) a été teinté, sa face orale et ses flancs sont mieux conservés (Planche annexe 2
, vignette 34) ; inventorié au Musée du site de Tercis sous la référence TCS.G068 ; un paratype plus grand (TCS.G069) a été teinté, sa face orale et ses flancs sont mieux conservés (Planche annexe 2 ![]() , vignette 34) ; ces types sont accompagnés de 5 topotypes ; sur l'un d'eux, la surface péri-apicale est parcourue de côtes radiales.
, vignette 34) ; ces types sont accompagnés de 5 topotypes ; sur l'un d'eux, la surface péri-apicale est parcourue de côtes radiales.
Discussion : La tasse minuscule est une petite sphère avec une collerette péri-apicale significative.
Distribution : Cette petite espèce est présente entre les cotes -5,0 et 11,0 oů une trentaine de spécimens ont été prélevés ; elle n'est pas très rare dans les 3 niveaux ; on récolte environ 1 spécimen par gramme de sédiment traité à la cote 11,0.
Pocillella cylindrata nov. sp. (code E3c, ce travail)
Description : Corps cylindrique de 120 à 140 µm de hauteur et de 120 µm de diamètre ; parfois plus petit (diamètre 100 µm ; hauteur 95 µm) ; face orale plane, légèrement concave, marquée d'une lèvre péri-orale d'oů rayonnent environ 8 crêtes (Vue 195 ![]() ) ; ébauche de collerette orale périphérique dont le pourtour comporte des pointements au niveau de l'arrivée des crêtes ; flancs parcourus de 16-18 rides méridiennes (Vue 196
) ; ébauche de collerette orale périphérique dont le pourtour comporte des pointements au niveau de l'arrivée des crêtes ; flancs parcourus de 16-18 rides méridiennes (Vue 196 ![]() ) décelables à la loupe binoculaire ; étranglement léger suivi d'un rebord séparant les flancs de la face orale d'un côté et de la surface péri-apicale sub-plane un peu concave, de l'autre ; nannostructure en cellules alignées dont les rebords latéraux dessinent les rides méridiennes ; surface péri-apicale, souvent un peu plus réduite que la face orale, ornée de microstructures radiales peu élevées (Vue 197
) décelables à la loupe binoculaire ; étranglement léger suivi d'un rebord séparant les flancs de la face orale d'un côté et de la surface péri-apicale sub-plane un peu concave, de l'autre ; nannostructure en cellules alignées dont les rebords latéraux dessinent les rides méridiennes ; surface péri-apicale, souvent un peu plus réduite que la face orale, ornée de microstructures radiales peu élevées (Vue 197 ![]() ) souvent dissimulées.
) souvent dissimulées.
Étymologie : L'espèce a été nommée d'après sa forme en cylindre, du latin cylindratus : cylindrique ; c'est la petite tasse cylindrique.
Holotype E3c : Provenant du niveau +5,0 l'holotype a été coloré, sa face orale montre 7 angles et les flancs sont très légèrement coniques (Planche annexe 2 ![]() , vignette 35) ; inventorié au Musée du site de Tercis sous la référence TCS.G070, il est accompagné de 3 topotypes dont l'un a les flancs un peu bombés.
, vignette 35) ; inventorié au Musée du site de Tercis sous la référence TCS.G070, il est accompagné de 3 topotypes dont l'un a les flancs un peu bombés.
Discussion : La petite tasse cylindrique se distingue par un développement identique de la face orale et de la surface péri-apicale et par la discrétion des collerettes orale et aborale ; elle est distinctement plus étroite que P. grandicula (E3a) et plus élevée que P. minuta (E3b).
Distribution : 6 spécimens de cette espèce cylindrique ont été observés aux cotes +5,0 et 11,0 avec un faciès très homogène.
Pocillella cochleare nov. sp. (code E3d, ce travail)
Description : Corps tronconique, extrémité orale de diamètre compris entre 160 et 180 µm, extrémité aborale de 120 à 150 µm donnant une forme générale en coquetier ; face orale très légèrement concave, parcourue de 8 ou 9 crêtes partant d'une lèvre péri-orale (Vue 198 ![]() ) et aboutissant à une étroite collerette en formant des pointements de section cruciforme ; crêtes orales souvent en continuité avec les rides des flancs ; rides méridiennes en nombre variable (18-25) dont une douzaine prolongées sur la collerette péri-apicale (Vue 200
) et aboutissant à une étroite collerette en formant des pointements de section cruciforme ; crêtes orales souvent en continuité avec les rides des flancs ; rides méridiennes en nombre variable (18-25) dont une douzaine prolongées sur la collerette péri-apicale (Vue 200 ![]() ) de longueur atteignant presque son diamètre ; nannostructure usuelle des gilianelles du genre en alignements méridiens sur les flancs rendue discrète par le développement de la microstructure : crêtes et rides.
) de longueur atteignant presque son diamètre ; nannostructure usuelle des gilianelles du genre en alignements méridiens sur les flancs rendue discrète par le développement de la microstructure : crêtes et rides.
Étymologie : L'espèce tire son nom du latin cochleare : coquetier ; le test a une silhouette rappelant cet accessoire de table ; c'est le petit coquetier.
Holotype E3d : Un spécimen a été coloré qui provient du niveau 25,0 ; il est morphologiquement très proche du spécimen de la Vue 55 ![]() (Planche annexe 2
(Planche annexe 2 ![]() , vignette 36) ; inventorié au Musée du site de Tercis sous la référence TCS.G071, l'holotype est accompagné de 8 topotypes dont les collerettes sont plus ou moins préservées.
, vignette 36) ; inventorié au Musée du site de Tercis sous la référence TCS.G071, l'holotype est accompagné de 8 topotypes dont les collerettes sont plus ou moins préservées.
Discussion : Parmi les espèces du genre, la petite tasse en coquetier se distingue par la forme nettement conique de son test ; la distinction de ces 4 espèces a été encouragée par le fait que chacune est très homogène là oů elle a été rencontrée ; l'absence de formes intermédiaires n'interdit pas de penser que l'une ait évolué vers l'autre mais chaque population est bien définie dans les niveaux traités à la base de l'Unité d'Avezac, à Tercis.
Distribution : 16 spécimens de la plus jeune des espèces de Pocillella ont été récoltés à la cote 25,0.
Genre Coraliella nov. gen.
Étymologie : Le nom de genre dérive du latin coralium : corail qui évoque l'aspect du test.
Espèce type : Coraliella simplex nov. sp.
Description : Corps conique à face orale développée et surface péri-apicale plus réduite que pour le genre Pocillella. Depuis le cône simple, sans expansion, les différentes espèces montrent une propension à développer des collerettes équatoriales plus ou moins étendues, entières ou découpées, et péri-apicales plus ou moins longues formant tube.
Remarque : Les taxons décrits ici ne constituent qu'une sélection parmi un plus grand nombre encore mal caractérisés, notamment dans les niveaux maastrichtiens.
Espèces attribuées au genre : simplex, cantharella avec 2 formes, brachialis.
Coraliella simplex nov. sp.
Vues 201-203 ![]() et 204
et 204 ![]()
Synonymie : Code M1.- et ,
2006, Pl. 5 ![]() .
.
Description : Corps conique sans expansion, de diamètre maximal 150 à 170 µm et de hauteur 160 à 180 µm, avec des flancs convexes donnant l'aspect d'un petit corail solitaire renforcé par le fait qu'on devine les crêtes orales à la loupe binoculaire ; face orale, légèrement concave, avec une lèvre péri-orale élevée d'oů partent environ 8 crêtes élevées (9 sur la Vue 202 ![]() ) aboutissant sur la marge du test sans former de pointements ; flancs convexes de contour général circulaire, parfois vaguement découpé en portions (autour de 7) voire elliptique (Vue 204
) aboutissant sur la marge du test sans former de pointements ; flancs convexes de contour général circulaire, parfois vaguement découpé en portions (autour de 7) voire elliptique (Vue 204 ![]() ) ; flancs parcourus de 2 douzaines de rides méridiennes élevées sur la portion la plus large du cône ; certaines disparaissent vers le bas et il en reste moins de 10 au pôle aboral de diamètre inférieur à 40 µm ; surface péri-apicale petite, confuse en son centre, entourée d'un bourrelet ; nannostructure maillée cachée par le relief des microstructures (Vues 201 et 203
) ; flancs parcourus de 2 douzaines de rides méridiennes élevées sur la portion la plus large du cône ; certaines disparaissent vers le bas et il en reste moins de 10 au pôle aboral de diamètre inférieur à 40 µm ; surface péri-apicale petite, confuse en son centre, entourée d'un bourrelet ; nannostructure maillée cachée par le relief des microstructures (Vues 201 et 203 ![]() ).
).
Étymologie : Le nom d'espèce est tiré du latin simplex : simple qui souligne la simplicité de ce test dépourvu d'expansion ; c'est le petit corail simple.
Holotype M1 : Provenant du niveau 79,8 un spécimen a été coloré, sa section est circulaire en vue aborale (Planche annexe 2 ![]() , vignette 37) ; inventorié au Musée du site de Tercis sous la référence TCS.G072, il est accompagné de 12 topotypes très semblables si ce n'est cette section parfois découpée, choisis parmi une vingtaine de spécimens récoltés.
, vignette 37) ; inventorié au Musée du site de Tercis sous la référence TCS.G072, il est accompagné de 12 topotypes très semblables si ce n'est cette section parfois découpée, choisis parmi une vingtaine de spécimens récoltés.
Discussion : De petits spécimens (100 -120 µm) des cotes 69,0 et 74,8 sont rattachés à ce taxon.
Distribution : L'espèce a été récoltée dans l'intervalle 69,0 - 79,8 qui représente environ 400 ka de dépôts ; l'abondance est de 1 à 3 spécimens par gramme de sédiment traité au niveau supérieur.
Coraliella cantharella (forme type : a) nov. sp.
Synonymie : Code M2.- ,
2007,
Fig. 3 ; code M2a.- et ,
2006, Pl. 6 ![]() .
.
Description : Test paraissant grand par le développement des expansions (ensemble, expansions comprises, de diamètre 200 à 230 µm et de hauteur : 175 à 190 µm), conique, semblable au champignon dénommé girolle ou chanterelle ; face orale au contour grossièrement circulaire, fortement marquée de 7-9, rarement 10, crêtes partant d'une lèvre péri-orale (Vue 206 ![]() ) ; crêtes atteignant le rebord extérieur de la collerette qu'elles arment et renforcent prolongeant un peu la collerette à leur niveau ; face orale du corps marqué par un dôme très bas de diamètre 125-140 µm, au centre de la collerette, distinct par sa coloration blanche opposée à la transparence de la collerette à la loupe binoculaire ; profil du corps tronconique en entonnoir marqué de rides méridiennes élevées dont les principales se branchent sur les crêtes orales (Vue 207
) ; crêtes atteignant le rebord extérieur de la collerette qu'elles arment et renforcent prolongeant un peu la collerette à leur niveau ; face orale du corps marqué par un dôme très bas de diamètre 125-140 µm, au centre de la collerette, distinct par sa coloration blanche opposée à la transparence de la collerette à la loupe binoculaire ; profil du corps tronconique en entonnoir marqué de rides méridiennes élevées dont les principales se branchent sur les crêtes orales (Vue 207 ![]() ) ; corps prolongé vers le bas par une collerette en tube étroit (cf. Vue aborale 208
) ; corps prolongé vers le bas par une collerette en tube étroit (cf. Vue aborale 208 ![]() ) armé du prolongement de 5 à 8 rides, lesquelles sont élevées sur le tube et prennent l'aspect d'ailettes ; nannostructure de la face orale du corps peu distincte, en nid d'abeille (Vue 207
) armé du prolongement de 5 à 8 rides, lesquelles sont élevées sur le tube et prennent l'aspect d'ailettes ; nannostructure de la face orale du corps peu distincte, en nid d'abeille (Vue 207 ![]() ) ; la collerette montre des alignements de cristallites radiaux semblables à ceux des autres gilianelles codées A2, A3, L2, L3 ; flancs marqués de l'aspect en napperon avec un arrangement des mailles aligné le long des rides en vue de profil (Vue 207
) ; la collerette montre des alignements de cristallites radiaux semblables à ceux des autres gilianelles codées A2, A3, L2, L3 ; flancs marqués de l'aspect en napperon avec un arrangement des mailles aligné le long des rides en vue de profil (Vue 207 ![]() ).
).
Étymologie : L'espèce tire son nom de Cantharella, nom de genre de la chanterelle (champignon) lui même du grec cantharos : coupe ; c'est le petit corail chanterelle.
Holotype M2a : Provenant du niveau 57,9 l'holotype a été coloré (Planche annexe 2 ![]() , vignette 38) ; inventorié au Musée du site de Tercis sous la référence TCS.G073, il est accompagné de 7 topotypes.
, vignette 38) ; inventorié au Musée du site de Tercis sous la référence TCS.G073, il est accompagné de 7 topotypes.
Discussion : Cette espèce se rattache au même genre que l'espèce type du genre par son corps en cône, terminé par une surface péri-apicale étroite mais présente un grand développement de la collerette. Une première forme : "a" a été distinguée afin de rendre compte d'un détail morphologique souligné dans la description du taxon suivant ; ici, le nombre de crêtes est limité et elles atteignent invariablement le bord de la collerette ; ce critère est visible à la loupe binoculaire : cette forme a été distinguée de la suivante avant l'examen électronique pour les 10 spécimens photographiés.
Distribution : La forme a du petit corail chanterelle a été reconnue entre les cotes 47,2 et 109,7 ; elle est plus particulièrement commune à la base de sa distribution avec plusieurs spécimens par gramme de sédiment traité.
Coraliella cantharella (forme b) nov. ssp.
Vues 209-211 ![]() et 215
et 215 ![]()
Synonymie : Code M2b.- et ,
2006, Pl. 6 ![]() .
.
Description : Corps identique à la forme a ; face orale a contour plus régulièrement circulaire que la forme a portant un nombre de crêtes plus grand visible à l'examen optique : 12-15 crêtes donnant un ensemble plus serré que dans la forme a (Vue 209 ![]() ) ; crêtes se détachant d'une lèvre péri-orale élevée n'atteignant pas clairement le bord externe de la collerette orale ce qui donne une ornementation en forme de soleil rayonnant, la plupart des rayons de ce soleil dépassant à peine celui du corps (Vues 210-211
) ; crêtes se détachant d'une lèvre péri-orale élevée n'atteignant pas clairement le bord externe de la collerette orale ce qui donne une ornementation en forme de soleil rayonnant, la plupart des rayons de ce soleil dépassant à peine celui du corps (Vues 210-211 ![]() ) ; au microscope électronique, certaines de ces crêtes s'abaissent seulement à ce niveau et se poursuivent en s'atténuant sur certains spécimens (Vue 209
) ; au microscope électronique, certaines de ces crêtes s'abaissent seulement à ce niveau et se poursuivent en s'atténuant sur certains spécimens (Vue 209 ![]() ) ; corps en cône montrant régulièrement des rides moins élevées que pour la forme a mais en plus grand nombre (Vue 215
) ; corps en cône montrant régulièrement des rides moins élevées que pour la forme a mais en plus grand nombre (Vue 215 ![]() ) ; tube aboral généralement plus fin et moins renforcé par des ailettes pour cette forme b.
) ; tube aboral généralement plus fin et moins renforcé par des ailettes pour cette forme b.
Holotype M2b : Provenant du niveau 54,9 un spécimen a été coloré (Planche annexe 2 ![]() , vignette 39) ; inventorié au Musée du site de Tercis sous la référence TCS.G074, il est accompagné de 9 topotypes.
, vignette 39) ; inventorié au Musée du site de Tercis sous la référence TCS.G074, il est accompagné de 9 topotypes.
Discussion : La distinction entre les 2 formes est possible à la loupe binoculaire comme au microscope électronique et elles ont été formalisées pour cette raison bien que le rang de sous-espèce paraisse élevé ; il n'a pas semblé utile de proposer deux noms parce que les deux formes sont contemporaines ; des différences du même ordre : Taille du corps, développement de la collerette orale, nombre de crêtes, pied en tube plus ou moins développé existent tout au long de la section mais elles n'ont pas encore été quantifiées ; il a donc paru raisonnable de se restreindre aux 3 taxons bien reconnus du genre : C. brachialis, C. simplex et C. cantharella avec ses 2 formes, tant qu'une documentation suffisante n'aura pas été réunie pour distinguer d'autres entités.
Distribution : La forme b du petit corail chanterelle a été reconnue entre les cotes 50,0 et 125,5 ; des formes proches, souvent plus petites, existent jusque dans le Maastrichtien supérieur ; on ne sait si elles doivent ou non être rattachées à C. cantharella ; au niveau 125,5 la forme b est dominante, plusieurs dizaines ont été observés et leur fréquence est de 2-3 spécimens par gramme de sédiment traité.
Coraliella brachialis nov. sp.
Synonymie : Code M3.- et ,
2006, Pl. 6 ![]() .
.
Description : Corps en cupule côtelée de diamètre voisin de 100 µm à face orale ornée de rayons nets donnant l'aspect d'un petit corail solitaire pourvu de bras puissants ; face orale déprimée marquée d'une lèvre péri-orale nette d'oů partent 6 à 7 crêtes élevées (Vue 212 ![]() ) armant autant de bras (quelquefois 5 -Vue 218
) armant autant de bras (quelquefois 5 -Vue 218 ![]() - et, plus rarement, 8 -Vue 216
- et, plus rarement, 8 -Vue 216 ![]() -, bien détachés du corps, forts et de section en croix (Vue 219
-, bien détachés du corps, forts et de section en croix (Vue 219 ![]() ) ; bras relevés d'environ 30° par rapport au plan équatorial (Vue 217
) ; bras relevés d'environ 30° par rapport au plan équatorial (Vue 217 ![]() ), liés entre eux par une collerette continue découpée par le développement des bras ; flancs du corps en dôme ogival parcouru de côtes fortes (aspect côtelé visible à la loupe binoculaire), les unes provenant des bras tandis que 3 à 4 côtes intermédiaires naissent au niveau de la couronne de bras ; cinq de ces côtes s'élèvent fortement en ailettes radiales depuis le tiers inférieur du corps pour armer un pied en tube (Vues 213-214
), liés entre eux par une collerette continue découpée par le développement des bras ; flancs du corps en dôme ogival parcouru de côtes fortes (aspect côtelé visible à la loupe binoculaire), les unes provenant des bras tandis que 3 à 4 côtes intermédiaires naissent au niveau de la couronne de bras ; cinq de ces côtes s'élèvent fortement en ailettes radiales depuis le tiers inférieur du corps pour armer un pied en tube (Vues 213-214 ![]() ) dont la longueur peut dépasser la moitié de la hauteur du corps (Vue 217
) dont la longueur peut dépasser la moitié de la hauteur du corps (Vue 217 ![]() ) ; nannostructure en cellules rectangulaires alignées le long des flancs.
) ; nannostructure en cellules rectangulaires alignées le long des flancs.
Étymologie : Le nom d'espèce dérive du latin brachialis : pourvu de bras ; c'est le petit corail à bras.
Holotype M3 : Provenant du niveau 69,0, l'holotype choisi a été coloré (Planche annexe 2 ![]() , vignette 40) ; il possède 6 bras et un pied en tube entouré de 6 ailettes ; inventorié au Musée du site de Tercis sous la référence TCS.G075, il est accompagné de 10 topotypes possédant 6 ou 7 bras, choisis parmi une centaine de spécimens.
, vignette 40) ; il possède 6 bras et un pied en tube entouré de 6 ailettes ; inventorié au Musée du site de Tercis sous la référence TCS.G075, il est accompagné de 10 topotypes possédant 6 ou 7 bras, choisis parmi une centaine de spécimens.
Discussion : Ŕ l'origine de l'étude, la localisation du (ou des) orifice(s) n'a pas été aisée en microscopie optique : Il pouvait être au centre de la lèvre de la face déprimée ou au centre du tube opposé ou aux deux endroits. En plongeant quelques tests dans l'acide acétique dilué, il a été possible de mettre en évidence la sortie de bulles par la face déprimée seulement vérifiant ainsi la présence d'un unique orifice dans ce qui a été nommé la face orale. L'espèce se distingue de toutes les autres par la présence de bras bien individualisés.
Distribution : Le petit corail à bras est caractéristique du court intervalle de dépôt 69,0-74,8 oů il constitue l'espèce dominante de l'association de gilianelles ; près de 30 spécimens ont été observés par gramme de sédiment traité à la cote 69,0 comme à la cote 74,8.
Genre Obbella nov. gen.
Étymologie : Le nom de genre dérive du latin obba : coupe à libations se terminant en pointe à son extrémité inférieure qui rappelle la forme générale du test.
Espèce type : Obbella alatopedata nov. sp.
Description : Le genre se distingue du précédent par l'absence de surface péri-apicale ; expansions à section en croix : Bras par 6 à 9 (rarement 5) ; pied aboral axial.
Remarques : Ces formes pourraient être considérées comme la suite de l'évolution des Coraliella dont la lumière du tube se serait tellement resserrée qu'elle disparaîtrait pour laisser seulement une structure axiale en ailettes. La structure originale du pied en croix à 4 (parfois 5) ailettes a fait choisir de proposer un nouveau genre dans la mesure oů, au lieu de succéder aux formes M1, M2 et M3, l'espèce type les précède (elle ne peut pas être mise à la fin d'une suite évolutive comprenant les espèces ultérieures). Obbella est donc le genre dont l'ornementation microstructurale orale en soleil possède un pied en croix. Cette structure constitue une quille efficace pour orienter cette forme l'orifice vers le haut dans le milieu liquide ; elle est probablement planctonique.
Espèces attribuées au genre : alatopedata, caryophyllum.
Obbella alatopedata nov. sp.
Synonymie : Code E2b.- et ,
2006, Pl. 3 ![]() .
.
Description : Corps en cupule simple de diamètre égal à la hauteur (100 - 125 µm), prolongé de fortes expansions aborales qui lui donnent l'apparence générale d'une amphore à la loupe binoculaire ; face orale plane, fortement marquée par une lèvre péri-orale d'oů partent 7 ou 8 crêtes rayonnantes
(Vue 224 ![]() ), élevées, aboutissant à autant de petites expansions périphériques (bras en picots atteignant 25 µm de long) sur lesquelles elles se prolongent
(Vues 224-225
), élevées, aboutissant à autant de petites expansions périphériques (bras en picots atteignant 25 µm de long) sur lesquelles elles se prolongent
(Vues 224-225 ![]() ) ; bras relevés et de section en croix ; corps finement côtelé (~25 côtes) à la loupe binoculaire ; au microscope électronique, chaque bras, armé vers le haut d'une crête
orale, est aussi armé d'une côte sur la face opposée ; 4 (Vue 222
) ; bras relevés et de section en croix ; corps finement côtelé (~25 côtes) à la loupe binoculaire ; au microscope électronique, chaque bras, armé vers le haut d'une crête
orale, est aussi armé d'une côte sur la face opposée ; 4 (Vue 222 ![]() ) parfois 5 de ces côtes s'élèvent progressivement sur les
flancs, deviennent très hautes au pôle aboral (Vue 221
) parfois 5 de ces côtes s'élèvent progressivement sur les
flancs, deviennent très hautes au pôle aboral (Vue 221 ![]() , par exemple) et se prolongent au delà en fortes ailettes dont la longueur approche la hauteur du corps sans l'atteindre
(Vue 223
, par exemple) et se prolongent au delà en fortes ailettes dont la longueur approche la hauteur du corps sans l'atteindre
(Vue 223 ![]() ).
).
Étymologie : Le nom d'espèce dérive du latin pedatus : pourvu de pied et alatus : ailé qui traduit l'aspect de l'expansion aborale ; c'est l'obbelle à pied ailé.
Holotype E2b : Provenant du niveau 11,0, un spécimen a été teinté (Planche annexe 2 ![]() , vignette 41) ; il possède 8 bras et le pied est constitué de 4 ailettes non perpendiculaires ; inventorié au Musée du site de Tercis sous la référence TCS.G076, il est accompagné de 15 topotypes choisis parmi une trentaine.
, vignette 41) ; il possède 8 bras et le pied est constitué de 4 ailettes non perpendiculaires ; inventorié au Musée du site de Tercis sous la référence TCS.G076, il est accompagné de 15 topotypes choisis parmi une trentaine.
Discussion : Parmi les formes dont la surface orale possède une ornementation microstructurale orale "en soleil", cette espèce est caractérisée d'abord par sa petite taille et par le large pied composé de 4 à 5 ailettes.
Distribution : L'obbelle à pied ailé est présente entre les cotes 1,1 et 14,8 ; à la cote 11,0 son abondance atteint 2 à 3 exemplaires par gramme de sédiment traité.
Obbella caryophyllum nov. sp.
Synonymie : Code G.- ,
2007,
Fig. 3 ; code G1.- et ,
2006, Pl. 4 ![]() .
.
Description : Corps en cupule sphérique côtelée de diamètre compris entre 110 et 125 µm, (parfois entre 105 et 150 µm) prolongé d'un long pied aboral donnant au test l'aspect d'un clou de girofle ; face orale en cône peu élevé (Vues 228 ![]() et 233
et 233 ![]() ), terminé par une lèvre péri-orale d'oů partent 6 ou 7 crêtes (parfois jusqu'à 9, Vue 231
), terminé par une lèvre péri-orale d'oů partent 6 ou 7 crêtes (parfois jusqu'à 9, Vue 231 ![]() ) se prolongeant sur autant de bras pleins, à section en croix (Vue 232
) se prolongeant sur autant de bras pleins, à section en croix (Vue 232 ![]() ) ; bras dirigés vers le haut à 30 à 45° par rapport au plan équatorial de longueur approchant le rayon de la face orale ; flancs striés à la loupe binoculaire ; certaines côtes, légères, s'alignent avec la crête inférieure des bras tandis que 2 à 3 intermédiaires naissent à la limite entre la face orale et les flancs (Vue 228
) ; bras dirigés vers le haut à 30 à 45° par rapport au plan équatorial de longueur approchant le rayon de la face orale ; flancs striés à la loupe binoculaire ; certaines côtes, légères, s'alignent avec la crête inférieure des bras tandis que 2 à 3 intermédiaires naissent à la limite entre la face orale et les flancs (Vue 228 ![]() ) ; côtes s'effaçant vers le pôle aboral oů 4 ailettes perpendiculaires naissent sans se brancher sur la microstructure des flancs pour former le pied à section en croix (Vue 230
) ; côtes s'effaçant vers le pôle aboral oů 4 ailettes perpendiculaires naissent sans se brancher sur la microstructure des flancs pour former le pied à section en croix (Vue 230 ![]() ) de longueur atteignant voire dépassant (Vue 229
) de longueur atteignant voire dépassant (Vue 229 ![]() ) la hauteur du corps ; généralement axial, ce pied est parfois dévié (Vue 227
) la hauteur du corps ; généralement axial, ce pied est parfois dévié (Vue 227 ![]() ) ; côtes des flancs résultant d'une nannostructure en cellules alignées radialement.
) ; côtes des flancs résultant d'une nannostructure en cellules alignées radialement.
Étymologie : Le nom d'espèce dérive du latin caryophyllum : girofle ou giroflier en accord avec l'aspect en clou de girofle ; c'est l'obbelle clou de girofle.
Holotype G1 : Provenant du niveau 57,9 un spécimen a été coloré sur le pied, entier. Il possède 7 bras développés dont l'un porte un débris accolé (Planche annexe 2 ![]() , vignette 42) ; inventorié au Musée du site de Tercis sous la référence TCS.G077, il est accompagné de 7 topotypes avec un pied bien représenté et 7 ou 8 bras plus ou moins complets, choisis parmi des dizaines de spécimens récoltés dans ce niveau.
, vignette 42) ; inventorié au Musée du site de Tercis sous la référence TCS.G077, il est accompagné de 7 topotypes avec un pied bien représenté et 7 ou 8 bras plus ou moins complets, choisis parmi des dizaines de spécimens récoltés dans ce niveau.
Discussion : Sur de nombreux exemplaires, le pied est détérioré et cette espèce peut se confondre avec la précédente obbelle à pied ailé. Cette dernière a cependant un pied plus court, beaucoup plus large à la base et un corps moins nettement sphérique. L'absence de structure annulaire aborale distingue G1 et G0 quand le pied est absent.
Distribution : Cette espèce a été rencontrée depuis la cote -5,0 jusqu'à la cote 125,5 ; elle est plus commune entre les cotes 50,0 et 79,8 oů 2 à 10 spécimens peuvent être extraits par gramme de sédiment traité.
Un genre, une espèce ou une sous-espèce ont été créés en fonction du "niveau" des différences observées. Ces différences peuvent être classées en trois catégories : 1- la forme générale du test (sphère, disque, cupule sans oublier les passages de l'une à l'autre de ces géométries de base), 2- la nature et la disposition des expansions (collerettes, bras, pieds avec, là encore, des passages ou combinaisons entre bras ou pied et collerette) et 3- l'ornementation microstructurale (par exemple l'ornementation de la face orale du dernier groupe avec cette lèvre péri-orale d'oů partent des crêtes radiaires). En jouant sur ces 3 composantes morphologiques et ornementales (et non sur la seule forme générale comme nous l'avions fait lors des approches précédentes : et , 2006), il semble qu'un classement (sinon une classification) rationnel des gilianelles puisse se dégager. Nous verrons à l'usage si ce classement résiste à l'ajout de taxons nouveaux dont un certain nombre a été observé dans les niveaux de Tercis et seront recherchés en d'autres lieux.
Cet exercice exige de peser le rang lié aux différences observées et de distinguer différences entre taxons et différences entre variants d'un même taxon. La difficulté réside, entre autres, dans la comparaison de formes morphologiquement variables d'âge différent. Ŕ plusieurs reprises, les descriptions ci-dessus ont noté que le nombre d'expansions, par exemple, variait d'un niveau à l'autre sans qu'il ait été jugé justifié de créer une espèce ou une entité de rang inférieur par niveau. Cette question a obligé à examiner des niveaux en nombre suffisant pour tenter d'obtenir, pour chaque taxon, des séparats représentatifs de niveaux distincts.
L'exercice de description et de classement ci-dessus n'est pas qu'un exercice à but pédagogique ; il a contribué à mettre en évidence une cohérence, une unité, un lien entre des restes divers jusqu'ici inconnus. C'est dire qu'un groupe nouveau a été mis en évidence dont la signification est renforcée par chaque forme décrite qui s'intègre dans une logique de l'ordre des descriptions. Par ce classement, on peut passer d'une forme à la suivante par une modification simple, par exemple étirer un bourrelet équatorial pour créer une voile équatoriale (passage A1a vers A3), par exemple encore, le passage de E3a vers M3 puis vers E2b enfin vers G1 en amenuisant progressivement la face aborale tout en allongeant l'expansion aborale. La succession dans le temps des taxons est le critère qui dit si cette continuité morphologique peut traduire ou non un phénomène évolutif.
Il reste à voir si les variations d'un gisement à l'autre confortent les variations observées dans la section du stratotype de la limite Campanien-Maastrichtien de Tercis afin d'estimer le potentiel stratigraphique du groupe ; ce travail est en cours et les premières observations sont prometteuses.
Un ordonnancement morphologique et stratigraphique a été mis en évidence à l'occasion de cette étude micropaléontologique parmi des restes présents en grand nombre. Il est clair, désormais, que des êtres vivants variés, abondants, évoluant rapidement et appartenant à un groupe taxinomique homogène et nouveau : les gilianelles, existaient dans ce faciès de plate-forme téthysien du SO de la France. Cette singulière paléobiodiversité ouvre des perspectives sur le travail qui reste à faire pour faire parler des couches de dépôts auxquelles sont appliquées généralement des recherches "pratiques" qui ne laissent pas le loisir d'exercer la curiosité naturaliste source de découvertes originales. Parmi elles, les formes inconnues qui ont été détachées des gilianelles restent à décrire ici ; ceci va permettre de préciser les caractères propres à ces dernières.
Le gisement de Tercis renferme des microproblematica dont les tests ne présentent pas la nannostructure diagnostique du groupe des gilianelles dans les mêmes niveaux que ces dernières. Ces formes revêtent un moindre d'intérêt taxinomique mais il est utile d'en présenter une sélection pour l'une ou l'autre parmi diverses raisons qui seront précisées lors des discussion des descriptions par exemple : 1- leur forme est aisément reconnaissable dès lors qu'elles sont définies ; 2- leur abondance est assez grande ce qui constitue une caractéristique identifiable du faciès qui les renferme ; 3- leur répartition verticale restreinte permet de repérer des lignes de temps dans la succession de Tercis et des environs ; 4- leur test présente des similitudes avec celui de certaines gilianelles et elles pourraient leur être rattachées au premier abord, ...
Les points communs entre gilianelles et autres microproblematica décrits ci-dessous sont les suivants : test uniloculaire et carbonaté, orifice unique, symétrie axiale, taille proche. Ils se distinguent par un ou plusieurs des critères suivants : absence de nannostructure de surface, absence d'expansions présentant une nannostructure d'un des types décrits ci-dessus, structure de la paroi du test différente.
La même terminologie que pour les gilianelles sera utilisée pour l'orientation du test et la description morphologique. Pour ces microproblematica non inclus parmi les gilianelles, les figurations sont faites au grossissement x200 mais, lorsqu'une nannostructure particulière est présente, une vue de détail est proposée.
Genre Lucernellus nov. gen.
Étymologie : Le nom de genre dérive du latin lucerna : lampion, lampe, lumière.
Espèce type : Lucernellus aubouini
Description : Corps cupulaire ; expansions de la face orale constituées de diverticules de la loge : La nannostructure des expansions est la même que celle du test formé de 2 couches de cristallites. Le genre est monospécifique à Tercis, voir la description de l'espèce type.
Lucernellus aubouini nov. sp.
Synonymie : Code F.- ,
2007 ; code F1.- et ,
2006, Pl. 4 ![]() .
.
Description : Corps cupulaire ; diamètre et hauteur équivalentes (130 à 190 µm) ; aspect de lampion renversé ; face orale en dôme bas, lisse, régulier avec un orifice discret, simple, de diamètre 12 µm environ (Vue 236 ![]() ) ; bras épais, creux (Vue 235
) ; bras épais, creux (Vue 235 ![]() ), largement enracinés dans le test et se réduisant progressivement vers l'extrémité distale émoussée, régulièrement disposés autour de la face orale en se dirigeant vers le haut, à 45° par rapport au plan équatorial ; presque tous les spécimens à 5 bras à la cote 54,9 mais majorité à 6 bras, exceptionnellement 4 ou 7 à la cote 50,0 (moins de 0,2 Ma plus tard) ; face aborale en cône à flancs à peine convexes (Vue 235
), largement enracinés dans le test et se réduisant progressivement vers l'extrémité distale émoussée, régulièrement disposés autour de la face orale en se dirigeant vers le haut, à 45° par rapport au plan équatorial ; presque tous les spécimens à 5 bras à la cote 54,9 mais majorité à 6 bras, exceptionnellement 4 ou 7 à la cote 50,0 (moins de 0,2 Ma plus tard) ; face aborale en cône à flancs à peine convexes (Vue 235 ![]() ) ; apex présentant toujours ce qui est interprété comme une cicatrice dont le centre laisse deviner une lumière, une cassure correspondant soit à une attache soit à une expansion fragile ; nannostructure de la paroi du test à deux couches ; première couche fine, se détachant souvent par lambeaux (Vues 235-236
) ; apex présentant toujours ce qui est interprété comme une cicatrice dont le centre laisse deviner une lumière, une cassure correspondant soit à une attache soit à une expansion fragile ; nannostructure de la paroi du test à deux couches ; première couche fine, se détachant souvent par lambeaux (Vues 235-236 ![]() ) entraînant un aspect "déguenillé" ; seconde couche épaisse de 15 µm (Vue 238
) entraînant un aspect "déguenillé" ; seconde couche épaisse de 15 µm (Vue 238 ![]() ), formée de cristallites irréguliers orientés perpendiculairement à la paroi comme chez les gilianelles ( et ,
2006) mais sans nannostructure de surface remarquable ; paroi semblable en section au niveau du corps et des bras comme de la cicatrice aborale.
), formée de cristallites irréguliers orientés perpendiculairement à la paroi comme chez les gilianelles ( et ,
2006) mais sans nannostructure de surface remarquable ; paroi semblable en section au niveau du corps et des bras comme de la cicatrice aborale.
Étymologie : L'espèce est dédiée à Jean ; cette première forme observée de microproblematica de Tercis à attiré notre attention au cours d'un projet de recherche à finalité tectonique (les sédiments carbonatés dans lesquels elle a été trouvée incidemment furent initialement traités aux acides tamponnés pour tenter d'isoler apatite, zircon, biotite et feldspaths caractérisant un événement magmatique acide supposé lié à une subduction nord-pyrénéenne) et parce que le dédicataire a marqué plusieurs générations de géologues devenus tels sous l'influence de ses lumineuses leçons magistrales sur les chaînes de montagne ; c'est le lucernel d'.
Holotype F1 : Provenant du niveau 54,9 l'holotype a été coloré (Planche annexe 2 ![]() , vignette 48) ; inventorié au Musée du site de Tercis sous la référence TCS.X083, il est accompagné de 29 topotypes à 5 bras ; un petit nombre de spécimens présentent 4 ou 6 bras dans ce niveau mais ils paraissent tératologiques (moignons pour les bras supplémentaires, espace vide quand 4 bras sont seuls développés).
, vignette 48) ; inventorié au Musée du site de Tercis sous la référence TCS.X083, il est accompagné de 29 topotypes à 5 bras ; un petit nombre de spécimens présentent 4 ou 6 bras dans ce niveau mais ils paraissent tératologiques (moignons pour les bras supplémentaires, espace vide quand 4 bras sont seuls développés).
Discussion : L'absence de nannostructure de surface en napperon, la présence d'une paroi à deux couches, les expansions constituées de simples diverticules du test (même nannostructure que ce dernier) ont fait séparer ce taxon des gilianelles. Le lucernel d' fait penser à la coque des pseudarcellidés (microproblematica du Paléogène : , 1989 ; , 1990 ; , 2000) ; dans le genre Conicarcella par exemple, la cupule sans expansion de 160 µm de diamètre est de taille semblable et sans nannostructure externe ; cependant, l'orifice de la face plane est large et atteint le tiers du diamètre de cette face ; ce large orifice est commun aux pseudarcellidés illustrés par ces auteurs.
Distribution : Cette forme énigmatique qui a conduit à approfondir la question des microproblematica et à la découverte des gilianelles bien que n'en faisant pas partie elle-même, a été trouvée dans les mêmes niveaux que les minéraux pyroclastiques (, 2001c) : dans la zone d'extension du foraminifère Radotruncana calcarata ; elle a été récoltée dans les 7 niveaux traités entre les cotes 47,2 et 57,9, soit durant un intervalle de dépôt de l'ordre de 0,4 Ma, ce qui fait de ce taxon, à Tercis, un substitut possible du foraminifère planctonique beaucoup plus rare ; sans être commune, l'espèce a fourni 1 à 5 restes pour 10 g de sédiment traité ; c'est un précieux marqueur de temps.
Genre Tubella nov. gen.
Étymologie : Le genre tire son nom du latin tuba : trompette droite qui décrit la forme générale en cône ouvert.
Espèce type : Tubella hunzikeri nov. sp.
Description : Corps en entonnoir de morphologie convergente avec celle des gilianelles L2 et L3 ; test à paroi formée de 2 couches ( et ,
2006) ; la première fine (2 à 3 µm), sans arrangement particulier ; la seconde, épaisse de 15 µm, avec des arrangements cristallins remarquables qui peuvent faire penser à l'aspect de surface en napperon des gilianelles (Vue 245 ![]() ) et sont différents dans le pied (cristallites allongés axialement), sur les flancs du corps (alignements de cristallites courts) et sur la face orale (arrangements en croix).
) et sont différents dans le pied (cristallites allongés axialement), sur les flancs du corps (alignements de cristallites courts) et sur la face orale (arrangements en croix).
Remarque : Les cristallites de 2 µm de long, arrangés en trame perpendiculaire, ont fait penser à la nannostructure des Schizosphaerella (nannofossiles incertae sedis du Trias et du Jurassique) mais ces restes sont composés de 2 valves, leur paroi est bien plus fine et leur taille inférieure (15-30 µm, , 2001). Le genre est monospécifique ; voir la description de l'espèce type.
Tubella hunzikeri nov. sp.
Synonymie : Code L.- ,
2007, Fig. 3 ; code L1.- et ,
2006, Pl. 5 ![]() .
.
Description : Corps en cône prolongé d'un pied, l'ensemble ayant l'aspect d'un entonnoir ou d'une trompe à grand pavillon ; face orale de contour circulaire, d'abord peu concave puis peu convexe (Vue 234 ![]() ) ; orifice discret (Vues 234 et 246
) ; orifice discret (Vues 234 et 246 ![]() ), quelquefois plus net au centre de cette face (Vues 240-241
), quelquefois plus net au centre de cette face (Vues 240-241 ![]() ) sans structure pour le mettre en évidence ; à l'équateur, une fissure sur le pourtour du test (Vue 243
) sans structure pour le mettre en évidence ; à l'équateur, une fissure sur le pourtour du test (Vue 243 ![]() ) pourrait faire penser à un couvercle posé sur le reste du corps ; face aborale en cône aux flancs soit droits (Vue 246
) pourrait faire penser à un couvercle posé sur le reste du corps ; face aborale en cône aux flancs soit droits (Vue 246 ![]() ) soit un peu renflés (Vue 243
) soit un peu renflés (Vue 243 ![]() ) ; pied fort, plein et, le plus souvent, dévié par rapport à l'axe de symétrie (Vue 241
) ; pied fort, plein et, le plus souvent, dévié par rapport à l'axe de symétrie (Vue 241 ![]() ) ; par la déviation du pied, le test acquiert une symétrie bilatérale ; nannostructure de la paroi présentant 2 couches. La Vue 245
) ; par la déviation du pied, le test acquiert une symétrie bilatérale ; nannostructure de la paroi présentant 2 couches. La Vue 245 ![]() laisse voir, sur le côté droit, un reste de la couche extérieure du test, fine.
laisse voir, sur le côté droit, un reste de la couche extérieure du test, fine.
Étymologie : L'espèce est dédiée à Johannes Christopher (prononcer ountsicoeur) géologue alpin, géochimiste et environnementaliste ; ce collègue suisse a influé sur les recherches de l'auteur en lui communiquant son expertise géochronologique tout en mettant largement à sa disposition les facilités de recherche dont il disposait dans son pays ouvert aux coopérations internationales ; c'est la tubelle de .
Holotype L1 : Provenant du niveau 11,0 un spécimen a été coloré (Planche annexe 2 ![]() , vignette 51) ; il présente un pied axial et un orifice visible à la loupe binoculaire ; inventorié au Musée du site de Tercis sous la référence TCS.X086, 7 topotypes l'accompagnent.
, vignette 51) ; il présente un pied axial et un orifice visible à la loupe binoculaire ; inventorié au Musée du site de Tercis sous la référence TCS.X086, 7 topotypes l'accompagnent.
Discussion : Comme le lucernel d', la tubelle de est une espèce commune ; sa distinction d'avec les gilianelles du genre Corniculum est difficile en microscopie optique ; l'examen au microscope électronique fait apparaître 1- que la nannostructure de surface du test diffère, 2- que la paroi est constituée de deux couches ; 3- que la similitude avec un cône à collerette est due à une convergence adaptative ; chez la tubelle, ce n'est pas une collerette pleine formée de cristallites disposés radiairement, c'est un repli de la paroi du test qui s'affine vers l'extérieur tellement que la moindre érosion fait apparaître la cavité interne du test ; il s'agit ici d'une convergence (même forme, même taille et même adaptation au même milieu) entre deux taxons de position taxinomique peut-être éloignée ; il était important de rapporter et décrire cette intéressante espèce.
Distribution : Le taxon a été observé dans tous les niveaux traités entre les cotes -9,5 et 50,25 dans la section principale de la Grande Carrière de Tercis ; il est toujours absent au dessus ; partout oů il a été observé, il est commun à extrêmement abondant avec des récoltes de près de 100 spécimens par gramme de sédiment traité ; c'est le marqueur le plus facile à repérer de toute la section de Tercis ; sa disparition brutale (totalement absent aux cotes 52,2 puis 54,9 puis 57,9) est surprenante dans une série sédimentaire homogène et continue.
Genre Vasculum nov. gen.
Étymologie : Le genre tire son nom de la forme en vase du test, du latin vasculum : petit vase.
Espèce type : Vasculum laevis nov. sp.
Description : La cupule est lisse ; la paroi de cet objet, observée sur un individu brisé lors de sa mise en place sur le plot est complexe : une couche fine, (1-2 µm d'épaisseur) non structurée en surface suivie d'une couche plus épaisse (environ 5 µm) formée d'un rang de tubes aplatis accolés par leur côté étroit, d'environ 6 µm de diamètre maximum. Le genre est monospécifique ; voir la description de l'espèce type
Vasculum laevis nov. sp.
Synonymie : Code E1.- et ,
2006, Pl. 3 ![]() .
.
Description : Corps cupulaire allongé, en vase rétréci vers le bas, variable, souvent en forme de casque italo-grec ou de pain de sucre pointe en bas, d'apparence lisse à la loupe binoculaire ; diamètre et hauteur de l'ordre de 150 µm ; face orale en cône volcanique sommé d'un relief annulaire de diamètre 40 µm, l'anneau lui-même étant très étroit (Vue 252 ![]() ) ; l'orifice au centre (Vues 247 et 250
) ; l'orifice au centre (Vues 247 et 250 ![]() ) reste supposé (Vue 252
) reste supposé (Vue 252 ![]() ) ; bord de la face orale parfois marqué d'un relief perpendiculaire, étroit, prolongeant les flancs de la cupule (Vue 248
) ; bord de la face orale parfois marqué d'un relief perpendiculaire, étroit, prolongeant les flancs de la cupule (Vue 248 ![]() ) ; flancs coniques ou renflés (Vue 248
) ; flancs coniques ou renflés (Vue 248 ![]() ), d'autre fois concaves (Vue 253
), d'autre fois concaves (Vue 253 ![]() ) ; pôle aboral en pointe émoussée (Vue 251
) ; pôle aboral en pointe émoussée (Vue 251 ![]() ) souvent détruit (Vues 249 et 253
) souvent détruit (Vues 249 et 253 ![]() ) ; apparence lisse à la loupe binoculaire ; pas de nannostructure de surface en napperon mais un réseau irrégulier se devine au microscope électronique, sur la face orale (Vues 250-253
) ; apparence lisse à la loupe binoculaire ; pas de nannostructure de surface en napperon mais un réseau irrégulier se devine au microscope électronique, sur la face orale (Vues 250-253 ![]() ).
).
Étymologie : Le nom d'espèce fait allusion au caractère lisse du test du latin laevis : lisse ; c'est le petit vase lisse.
Holotype E1 : Provenant du niveau 11,0 l'holotype a été coloré ; sa forme est en pain de sucre et
l'orifice (?) en relief est porté par un col annulaire, net (Planche annexe 2 ![]() , vignette 47) ; inventorié au Musée du site de Tercis sous la référence TCS.X082, il est accompagné de 11 topotypes dont les flancs sont bombés à coniques.
, vignette 47) ; inventorié au Musée du site de Tercis sous la référence TCS.X082, il est accompagné de 11 topotypes dont les flancs sont bombés à coniques.
Discussion : Ce taxon a posé problème par la variabilité de sa forme, par l'absence de nannostructure nette si ce n'est le réseau indistinct de la face orale et par sa répartition disjointe ; les 4 spécimens soumis à l'examen électronique ont révélé des caractéristiques semblables qui ne correspondent pas à celles des gilianelles et suggèrent une variabilité de la forme du test qui est unique parmi les microproblematica examinés dans ce travail de même que la nannostructure de la paroi du test.
Distribution : Le petit vase lisse a été trouvé dans les niveaux +5 à +18,2 d'une part et à la cote 50,0 d'autre part ; les observations au microscope n'ont pas révélé de différences entre ces deux ensembles ; à la cote 11,0 l'abondance est autour de 2 spécimens par gramme de sédiment traité.
Genre Cimicellus nov. gen.
Étymologie : Le nom de genre dérive du latin cimex : punaise d'après la forme des tests en punaise à fixer les documents.
Espèce type : Cimicellus nudatus nov. sp.
Description : Corps en forme de punaise avec orifice sur une face orale presque plane ; orifice surtout marqué par le relief qu'il forme au centre de la face orale sur les moulages internes ; face aborale en cône à flancs concaves ; paroi du test très fine souvent absente, sans nannostructure en napperon ; les moulages internes qui sont les restes observés le plus souvent, sont carbonatés, parfois glauconieux.
Espèces attribuées au genre : nudatus, clavellus.
Cimicellus nudatus nov. sp.
Synonymie : Code K.- ,
2007,
Fig. 3 ; code K1.- et ,
2006, Pl. 5 ![]() .
.
Description : Corps grand (diamètre = hauteur = 150-200 µm) ; face orale de contour circulaire, modérément convexe avec un très léger changement de pente au tiers extérieur de la face (Vue 256 ![]() ) ; orifice axial mal défini, probablement un léger relief péri-oral (Vue 258
) ; orifice axial mal défini, probablement un léger relief péri-oral (Vue 258 ![]() ) avec l'orifice au centre ; test très fin se détachant du moulage interne notamment à l'endroit de l'orifice (Vue 255
) avec l'orifice au centre ; test très fin se détachant du moulage interne notamment à l'endroit de l'orifice (Vue 255 ![]() ) et brouillant son aspect ; profil dissymétrique (Vues 254 et 256
) et brouillant son aspect ; profil dissymétrique (Vues 254 et 256 ![]() ) avec une importante face aborale conique, à la pointe parfois courbée, aux flancs concaves ; corps terminé par une pointe souvent altérée (Vues 256
) avec une importante face aborale conique, à la pointe parfois courbée, aux flancs concaves ; corps terminé par une pointe souvent altérée (Vues 256 ![]() et 258
et 258 ![]() ) ; face aborale sans ornementation (Vue 257
) ; face aborale sans ornementation (Vue 257 ![]() ) ; test sans nannostructure organisée ; moule interne régulièrement piqueté en relief en l'absence du test (partie desquamée en haut de la Vue 255
) ; test sans nannostructure organisée ; moule interne régulièrement piqueté en relief en l'absence du test (partie desquamée en haut de la Vue 255 ![]() ) laissant supposer la présence de pores dans le test ; généralement carbonaté dans le niveau de l'holotype, les moulages internes sont parfois glauconitisés (Vue 259
) laissant supposer la présence de pores dans le test ; généralement carbonaté dans le niveau de l'holotype, les moulages internes sont parfois glauconitisés (Vue 259 ![]() avec la trace des pores en relief).
avec la trace des pores en relief).
Étymologie : Le nom d'espèce fait allusion à l'absence usuelle du test, du latin nudatus : dépouillé ; c'est la petite punaise dépouillée.
Holotype K1 : Provenant du niveau 79,8 l'holotype est teinté d'orange, de taille assez grande, avec un cône aboral légèrement courbé, le site de l'orifice est un peu en relief (Planche annexe 2 ![]() , vignette 49) ; inventorié au Musée du site de Tercis sous la référence TCS.X084, il est accompagné de 11 topotypes dont l'un est exceptionnellement petit (Planche annexe 2
, vignette 49) ; inventorié au Musée du site de Tercis sous la référence TCS.X084, il est accompagné de 11 topotypes dont l'un est exceptionnellement petit (Planche annexe 2 ![]() , vignette 49) choisis parmi des dizaines de spécimens récoltés.
, vignette 49) choisis parmi des dizaines de spécimens récoltés.
Discussion : Ces restes communs dans un seul échantillon traité, sont restés mystérieux tant qu'il n'a pas été reconnu qu'il s'agissait de moulages internes ; les spécimens illustrés ont été choisis parmi les rares ayant un test au moins partiellement préservé, très fin (il a fallu examiner 7 spécimens au microscope électronique pour en trouver 1 seul avec test entièrement préservé) ; la finesse de ce qui parait être le test pose problème : il est trop fin pour offrir une résistance telle que de nombreux restes soient fossilisés.
Distribution : L'essentiel des restes carbonatés, moulage interne ou test rempli, a été observé à la cote 79,8 oů leur abondance est significative : Environ 7 spécimens par gramme de sédiment traité ; de rares spécimens ont été récoltés aux cotes 65,4 et 74,8 tandis que des moulages internes glauconitiques ont été récoltés à divers niveaux entre les cotes 54,9 et 89,0 ; avec ce taxon complémentaire, la diversité des microproblematica est étonnante à la cote 79,8 oů sont actuellement dénombrées 24 formes attestées.
Cimicellus clavellus nov. sp.
Synonymie : Code K2a.- et ,
2006, Pl. 5 ![]() .
.
Description : Corps en pain de sucre renversé, haut de près de 200 µm et de diamètre 125 µm ; ensemble avec la même symétrie axiale et la même absence de test mais avec une forme plus élancée que le taxon précédent ; face orale débordant les flancs (Vue 260 ![]() ) mimant la présence d'une collerette équatoriale formée par une extension du test replié ; cône aboral lisse et sans nannostructure n'ayant jamais montré de test ; reste interprété comme la forme précédente :
un moulage interne d'un test à la paroi très fine.
) mimant la présence d'une collerette équatoriale formée par une extension du test replié ; cône aboral lisse et sans nannostructure n'ayant jamais montré de test ; reste interprété comme la forme précédente :
un moulage interne d'un test à la paroi très fine.
Étymologie : Le nom d'espèce fait allusion à l'étroitesse du corps rappelant la forme du clou ; du latin clavellus : petit clou ; c'est le petit clou.
Holotype K2a : Provenant du niveau +5,0 l'holotype a été légèrement teinté d'orange ; il possède une ébauche de collerette plus large que les autres spécimens ; inventorié au Musée du site de Tercis sous la référence TCS.X085, il est accompagné des 3 topotypes (Planche annexe 2 ![]() , vignette 50) restant après l'étude de 2 spécimens au microscope électronique.
, vignette 50) restant après l'étude de 2 spécimens au microscope électronique.
Discussion : Cette forme est décrite pour souligner que l'espèce précédente n'est pas unique de sa sorte : Elle appartient au même genre ; d'autres microproblematica voisins ou à symétrie bilatérale (provoquée par un aplatissement latéral ?) ont aussi été observés dans divers niveaux ; leur distribution ni leur abondance ne sont parlantes et ils sont omis ici.
Distribution : Le petit clou a été récolté à la cote +5,0 en petit nombre sans être recherché.
Genre Piperella nov. gen.
Étymologie : Le genre tire son nom du latin piper : poivre, d'après l'aspect en grain de poivre ridé.
Espèce type : Piperella dercourti nov. sp.
Description : Tests sphériques montrant une ornementation en cellules de type nid d'abeille plus ou moins serrée. Pas de différenciation en face orale et aborale ; orifice situé au centre d'un arrangement spécifique de cellules avec un col de développement variable depuis l'absence complète jusqu'au long tube ; paroi du test plus épaisse (20 à 30 µm) que celle des gilianelles (10-15 µm) d'après et (2006).
Remarques : C'est ce genre qui ressemble le plus aux dinokystes calcaires. (1984) illustre le genre Calciodinellum qui mime étonnamment l'aspect de D1a ci-dessous ; cependant, il n'y a pas ces "crêtes disposées de manière à diviser, par une série équatoriale (correspondant à un sillon transversal) la coque en deux parties "épithèque et hypothèque" rappelées par la citation de la diagnose de ; de plus, le diamètre de Calciodinellum est plus petit (illustré autour de 25 à 35 µm) que celui de nos piperelles. Une autre illustration de Heptasphaera michaeli du Crétacé inférieur ( et , 1994, Fig. 8, n° 9) rappelle encore D1a mais le diamètre plus faible (45 µm au lieu de 150-200 µm) et l'absence de cette organisation en plaques propre aux kystes de dinoflagellés indiquent encore une fois qu'il s'agit d'une convergence de forme. L'observation clé des travaux de (1979), pour D1a comme pour les autres piperelles, est la présence de kystes complets et de kystes ouverts à la suite de la sortie de l'organisme ; ceci n'a pas été observé pour les microproblematica de Tercis qui ne sont donc pas des formes de résistance de Dinoflagellés.
On peut encore comparer la forme épineuse de D1a avec le taxon Thoracosphaera spinosa illustré par (1979) avec les mêmes réserves (taille et organisation du test en plaques) que ci-dessus.
Autre ressemblance, celle qui nous fut signalée par Jean-Paul avec le genre Bolboforma (Protophytes, incertae sedis, Tertiaire). Ces formes du Miocène présentent aussi des enveloppes carbonatées en panneaux de ballon cousu avec, parfois, une ornementation plus ou moins hérissée ( et , 1991) très convergente avec celle de nos piperelles de même qu'une certaine variabilité de l'ornementation pour un taxon donné ; l'âge est différent, la paroi n'est jamais monocristalline dans nos échantillons mais ces deux raisons n'excluent pas complètement ce groupe fossile pour le genre Piperella.
Espèces attribuées au genre : dercourti, camelopardalis.
Piperella dercourti nov. sp.
Synonymie : Code D1a.- ,
2007,
Fig. 3 ; et ,
2006, Pl. 3 ![]() .
.
Description : Corps sphérique de diamètre 170 à 220 µm ayant l'aspect d'un grain de poivre ridé ; ornementation découpant toute la sphère en panneaux comme ceux d'un ballon de cuir cousu ; orientation repérée par la présence d'un orifice porté par un col parfois aussi élevé que son diamètre (Vue 264 ![]() ) ; hémisphère oral présentant, autour du col, une rangée latitudinale de 6 à 7 cellules à 5 ou 6 côtés (Vue 262
) ; hémisphère oral présentant, autour du col, une rangée latitudinale de 6 à 7 cellules à 5 ou 6 côtés (Vue 262 ![]() ) ; sur les "flancs", 2 rangées succèdent à cette première série ; côté aboral, une dernière rangée entoure 3 à 4 cellules aborales souvent légèrement en dépression (Vue 2645
) ; sur les "flancs", 2 rangées succèdent à cette première série ; côté aboral, une dernière rangée entoure 3 à 4 cellules aborales souvent légèrement en dépression (Vue 2645 ![]() ) ; relief des cellules plus ou moins marqué avec, parfois, au carrefour entre les cellules, des excroissances donnant un aspect hérissé (Vue 266
) ; relief des cellules plus ou moins marqué avec, parfois, au carrefour entre les cellules, des excroissances donnant un aspect hérissé (Vue 266 ![]() ) masquant les cellules ; pas de bonne nannostructure en napperon ; paroi épaisse avec des arrangements cristallins complexes très variés autour des cellules :
anneaux, lattes concentriques, ... qui demanderaient une étude particulière :
il peut s'agir d'une nannostructure originelle ou résultant d'une recristallisation.
) masquant les cellules ; pas de bonne nannostructure en napperon ; paroi épaisse avec des arrangements cristallins complexes très variés autour des cellules :
anneaux, lattes concentriques, ... qui demanderaient une étude particulière :
il peut s'agir d'une nannostructure originelle ou résultant d'une recristallisation.
Étymologie : L'espèce est dédiée à Jean , un maître apprécié, qui a maintenu la qualification de son unité "laboratoire de stratigraphie" à une époque oů cette science de synthèse cardinale de la géologie était déjà jugée dévalorisante dans les sphères administratives de la recherche ; hommage est ici rendu à sa contribution au développement de cette discipline dans son aboutissement paléogéographique ; c'est la piperelle de .
Holotype D1a : Provenant du niveau 79,8 le spécimen coloré présente, outre celle entourant l'orifice avec un col bien dessiné, 4 rangées de cellules polygonales (Planche annexe 2 ![]() , vignette 44) ; inventorié au Musée du site de Tercis sous la référence TCS.X079, il est accompagné de 9 topotypes dont l'un est hérissé.
, vignette 44) ; inventorié au Musée du site de Tercis sous la référence TCS.X079, il est accompagné de 9 topotypes dont l'un est hérissé.
Discussion : La variation de l'ornementation avec contours de cellules plus ou moins élevés, parfois renforcés de pointes donnant un aspect hérissé à la sphère est continue et ce n'est pas une question de préservation ; la forme hérissée appartient à la même population que la forme type de ce taxon ; la tentation de rattacher la piperelle de aux gilianelles (, 2007, Fig. 3) a été contrariée par l'absence d'expansions et une épaisseur du test inhabituelle ( et , 2006) ; la nannostructure en napperon n'est jamais manifeste mais un doute demeure sur une possible variante.
Distribution : Cette piperelle est présente dans toute la section y compris jusque près de la limite Crétacé-Paléogène avec une abondance très variable de rare à près de 10 spécimens par gramme de sédiment traité (notamment aux cotes 11,0 et 79,8).
Piperella dercourti rugatissimus nov. ssp. (code D1b, ce travail)
Description : Corps sphérique de taille parfois plus grande que l'espèce type (jusqu'à 230 µm) ; nombre de cellules élevé avec une organisation latitudinale moins rigoureuse ; orifice parfois entouré d'une vague structure annulaire (Vue 267 ![]() ) mais sans col ; nannostructure présentant des arrangements complexes en chaînette de tubes perpendiculaires à la surface (spécimen de la Vue 269
) mais sans col ; nannostructure présentant des arrangements complexes en chaînette de tubes perpendiculaires à la surface (spécimen de la Vue 269 ![]() ) qui ne rappelle aucun des aspects décrits chez les gilianelles.
) qui ne rappelle aucun des aspects décrits chez les gilianelles.
Étymologie : La sous-espèce tire son nom du latin rugatus : ridé ; c'est la piperelle très ridée.
Holotype D1b : Provenant du niveau 79,8 un grand spécimen a été coloré ; il présente 7 rangs de cellules petites et l'orifice est sans col (Planche annexe 2 ![]() , vignette 45 ; inventorié au Musée du site de Tercis sous la référence TCS.X080, il est accompagné de 5 topotypes dont l'un est modérément hérissé.
, vignette 45 ; inventorié au Musée du site de Tercis sous la référence TCS.X080, il est accompagné de 5 topotypes dont l'un est modérément hérissé.
Discussion : Il existe des spécimens intermédiaires entre les formes illustrées de D1b et celles de D1a mais l'aspect général très différent à laissé penser qu'il était utile de proposer une désignation particulière à cette sous-espèce ; les holotypes des deux formes ont été choisis dans le même niveau pour illustrer le fait qu'ils étaient membres d'un même ensemble présentant deux morphologies.
Distribution : Le plus souvent 10 fois plus rare que la forme type contemporaine, la sous-espèce très ridée a été observée dans le Campanien entre les cotes +5,0 et 79,8 essentiellement ; exceptionnellement (cote 18,2), elle est plus commune que D1a avec une abondance de deux spécimens par gramme de sédiment traité ; cette absence de parallélisme entre les abondances respectives confirme la distinction de deux taxons ou suggère une adaptation distincte.
Piperella camelopardalis nov. sp. (code D3, ce travail)
Description : Corps sphérique, petit (diamètre 140 µm) prolongé d'un long col (65 µm, Vue 270 ![]() ) ; corps recouvert d'une microstructure dessinant des cellules polygonales visibles au microscope électronique, moins nettes à la loupe binoculaire ; excroissances plus marquées au carrefour des cellules ; holotype présentant un aspect moins hérissé que le topotype oů ces excroissances sont fortes ; nannostructure évoquant des mailles de type "napperon" sur la sphère (agrandissement :
Vue 272
) ; corps recouvert d'une microstructure dessinant des cellules polygonales visibles au microscope électronique, moins nettes à la loupe binoculaire ; excroissances plus marquées au carrefour des cellules ; holotype présentant un aspect moins hérissé que le topotype oů ces excroissances sont fortes ; nannostructure évoquant des mailles de type "napperon" sur la sphère (agrandissement :
Vue 272 ![]() ) ; le col est constitué de nannostructures longitudinales, parallèles à l'axe de symétrie (Vue 270
) ; le col est constitué de nannostructures longitudinales, parallèles à l'axe de symétrie (Vue 270 ![]() ).
).
Étymologie : L'espèce tire son nom du latin camelopardalis : girafe qui fait référence à l'ampleur du col ; c'est la piperelle girafe.
Holotype D3 : Provenant du niveau 79,8, l'holotype est le spécimen figuré au microscope électronique (plot XI, spécimen n° 2) qui est inventorié au Musée du site de Tercis sous la référence TCS.X081 ; un topotype a été récolté, incomplet mais présentant toutes les caractéristiques de l'espèce (Planche annexe 2 ![]() , vignette 46).
, vignette 46).
Discussion : Malgré la rareté de ses représentants, l'espèce est formalisée ici pour illustrer la question de la nannostructure de la paroi ; D3 présente des caractéristiques d'ensemble similaires à celles de D1a avec le même corps sphérique portant une microstructure en cellules : L'espèce est donc rattachée au même genre ; cette microstructure est discrète dans le spécimen figuré ; elle recouvre apparemment une nannostructure en réseau qui rappelle celle des gilianelles ; par ailleurs, d'autres taxons de gilianelles ont révélé que les petits spécimens (supposés plus jeunes) laissent mieux percevoir la nannostructure que les formes plus grandes ; une interprétation de l'absence de nannostructure en napperon pour le genre piperelle serait alors que celle-ci est cachée par des surcroissances postérieures et, si c'était le cas, les piperelles devraient être rattachées aux gilianelles, ce qui n'est pas admis ici.
Distribution : Les deux spécimens de piperelle girafe récoltés proviennent du niveau 79,8.
Genre Globulella nov. gen.
Étymologie : Le nom du genre dérive du latin globulus : petite boule, bille, d'après l'aspect sphérique du test (en vue orale ou aborale dans lesquelles il se dispose toujours).
Espèce type : Globulella opprimata nov. sp.
Description : Le corps globulaire est plus ou moins aplati, sans expansion ; la paroi du test montre 2 couches avec une nannostructure de la couche interne spécifique ; le genre est monospécifique pour le moment ; il y a probablement divers taxons possédant des caractéristiques morphologiques proches mais une distribution stratigraphique différente, voir la description de l'espèce type.
Globulella opprimata nov. sp. (code A5, ce travail)
Description : Corps en sphère lisse, partiellement aplatie en lentille, de diamètre 160 à 200 µm ; aplatissement dissymétrique par rapport à un équateur parfaitement circulaire (Vue 274 ![]() ) ; face orale en dôme plus bas avec un orifice (?) discret (Vue 273
) ; face orale en dôme plus bas avec un orifice (?) discret (Vue 273 ![]() ) ; équateur presque anguleux ; face aborale en dôme plus haut, hémisphérique (Vue 274
) ; équateur presque anguleux ; face aborale en dôme plus haut, hémisphérique (Vue 274 ![]() ) ; paroi du test formée de deux couches ; test le plus souvent dépouillé partiellement (Vue 273
) ; paroi du test formée de deux couches ; test le plus souvent dépouillé partiellement (Vue 273 ![]() ) ou totalement (Vue 274
) ou totalement (Vue 274 ![]() ) de la pellicule grumeleuse formée par la couche externe fine peu adhérente ; couche interne piquetée de cristaux à extrémité carrée (Vue 275
) de la pellicule grumeleuse formée par la couche externe fine peu adhérente ; couche interne piquetée de cristaux à extrémité carrée (Vue 275 ![]() ), épaisse de 8 à 10 µm.
), épaisse de 8 à 10 µm.
Étymologie : Le nom d'espèce est dérivé du latin opprimere : écraser, aplatir pour qualifier le fait que le test semble aplati axialement ; c'est la boulette aplatie.
Holotype A5 : Provenant du niveau 109,7 un spécimen grand avec une tache noire sur la face plus conique a été coloré (Planche annexe 2 ![]() , vignette 43) ; cet holotype est inventorié au Musée du site de Tercis sous la référence TCS.X078 ; il est accompagné de 13 topotypes de taille variée choisis parmi un grand nombre de spécimens.
, vignette 43) ; cet holotype est inventorié au Musée du site de Tercis sous la référence TCS.X078 ; il est accompagné de 13 topotypes de taille variée choisis parmi un grand nombre de spécimens.
Discussion : Tous les spécimens répondant précisément à la description ci-dessus proviennent de niveaux entourant la limite Campanien-Maastrichtien ; 119 m plus haut, dans le Maastrichtien élevé, des restes très proches ont été récoltés ; ils présentent le même aplatissement et la même nannostructure ; une vue cavalière (Vue 276 ![]() ) illustre la face supposée orale avec, au centre, ce qui est interprété comme la trace d'un orifice et, sur le côté droit, les restes de la couche extérieure du test largement desquamée ailleurs ; le profil (Vue 277
) illustre la face supposée orale avec, au centre, ce qui est interprété comme la trace d'un orifice et, sur le côté droit, les restes de la couche extérieure du test largement desquamée ailleurs ; le profil (Vue 277 ![]() ) montre un aplatissement plus sensible que pour les spécimens du niveau 109,7. Tous ces restes sont rassemblés dans le même taxon ; A5 montre une nannostructure de la paroi en 2 couches, bien définie qu'il a semblé utile de rapporter car elle est propre à une des multiples formes lenticulaires observées dans les sédiments ; ces diverses lentilles plus ou moins épaisses n'ont pu être caractérisées et distinguées par manque de fait notable au contraire de A5 et ne seront pas décrites ici.
) montre un aplatissement plus sensible que pour les spécimens du niveau 109,7. Tous ces restes sont rassemblés dans le même taxon ; A5 montre une nannostructure de la paroi en 2 couches, bien définie qu'il a semblé utile de rapporter car elle est propre à une des multiples formes lenticulaires observées dans les sédiments ; ces diverses lentilles plus ou moins épaisses n'ont pu être caractérisées et distinguées par manque de fait notable au contraire de A5 et ne seront pas décrites ici.
Distribution : La boulette aplatie typique a été récoltée en nombre (environ 25 spécimens par gramme de sédiment traité à la cote 109,7) entre les cotes 109,7 et 125,5 ; les spécimens plus petits et écrasés proviennent de l'affleurement complémentaire entourant la limite Crétacé-Paléogène, précisément 29 m (représentant environ 1,2 Ma) sous celle-ci.
Genre Pilella nov. gen.
Étymologie : Le nom de genre dérive du latin pila : balle, pelote, d'après l'aspect sphérique du test.
Espèce type : Pilella reticulata nov. sp.
Description : Corps sphérique ; taille commune avec celle des pithonelles ( et alii, 2001) ; test à nannostructure de surface réticulée, en cannage ; microproblematica sphérique distinct des pithonelles (et calcisphères) ; en effet, les nannostructures des calcisphères illustrées par (1975, 1977) sont différentes de celles vues pour Pilella ; le genre est monospécifique pour le moment, voir la description de l'espèce type.
Pilella reticulata nov. sp. (code N4, ce travail)
Description : Corps sphérique, de diamètre 175 µm ; face orale distincte à la loupe binoculaire avec un orifice bien dessiné en creux, de 25 µm de diamètre (Vue 280 ![]() ) ; de profil, la sphère est à peine aplatie au pôle oral (Vue 278
) ; de profil, la sphère est à peine aplatie au pôle oral (Vue 278 ![]() ) ; nannostructure très caractéristique formant un réseau d'aspect similaire au cannage des sièges (Vue 279
) ; nannostructure très caractéristique formant un réseau d'aspect similaire au cannage des sièges (Vue 279 ![]() ).
).
Étymologie : Le nom d'espèce est dérivé du latin reticulatus : réticulé pour qualifier le dessin de la nannostructure ; c'est la petite pelote réticulée.
Holotype N4 : L'holotype est le spécimen figuré Vues 278-280 ![]() (plot XI, spécimen 03) qui est inventorié au Musée du site de Tercis sous la référence TCS.G087 il provient du niveau 79,8.
(plot XI, spécimen 03) qui est inventorié au Musée du site de Tercis sous la référence TCS.G087 il provient du niveau 79,8.
Discussion : La nannostructure caractéristique du genre n'est visible qu'au microscope électronique à balayage ; c'est pour rendre compte de celle-ci que le taxon a été présenté ; une réticulation semblable mais plus large a été rencontrée sur un autre spécimen de même forme et taille ; il n'est pas rangé dans l'espèce décrite pour le moment ; des examens plus systématiques permettraient probablement de décrire divers taxons.
Distribution : Les deux spécimens ayant mis en évidence la réticulation remarquable, très fine, proviennent des niveaux 62,0 et 79,8 ; la réticulation plus large a été observée dans un spécimen de la cote +5,0 ; l'abondance de ces restes n'est pas connue car leur reconnaissance n'est pas possible en microscopie optique.
Les taxons inclus parmi les gilianelles dans ce travail sont ceux pour lesquels une série de caractéristiques a été retenue y compris la nannostructure de surface en napperon. Pour une espèce donnée, cette nannostructure de surface est parfois masquée sur une proportion plus ou moins grande des individus.
Chez les taxons classés dans le genre Tercensella, la nannostructure paraît toujours très bien visible (Vues 1-13 ![]() ) mais il convient de rappeler que c'est précisément parce que la nannostructure était évidente sur ces globules, autrement peu distincts des pithonelles à l'observation optique, que les spécimens ont été prélevés pour l'illustration au microscope électronique. Pour d'autres taxons reconnus et sélectionnés d'après leur forme générale et non d'après une nannostructure évidente à l'observation optique (Vues 48
) mais il convient de rappeler que c'est précisément parce que la nannostructure était évidente sur ces globules, autrement peu distincts des pithonelles à l'observation optique, que les spécimens ont été prélevés pour l'illustration au microscope électronique. Pour d'autres taxons reconnus et sélectionnés d'après leur forme générale et non d'après une nannostructure évidente à l'observation optique (Vues 48 ![]() , 109
, 109 ![]() , 126
, 126 ![]() , par exemple) l'aspect en napperon est moins distinct voire rarement observé. Quelques exemples ont montré que les formes plus petites -plus jeunes- montrent une nannostructure remarquablement nette alors que pour les formes plus grandes -adultes-, la nannostructure est masquée, seules les microstructures étant claires. La nannostructure pourrait donc être une caractéristique liée au stade de développement, un colmatage survenant à l'état adulte.
, par exemple) l'aspect en napperon est moins distinct voire rarement observé. Quelques exemples ont montré que les formes plus petites -plus jeunes- montrent une nannostructure remarquablement nette alors que pour les formes plus grandes -adultes-, la nannostructure est masquée, seules les microstructures étant claires. La nannostructure pourrait donc être une caractéristique liée au stade de développement, un colmatage survenant à l'état adulte.
L'exemple de la forme D3 incite à la prudence : il est difficile d'interpréter ce que l'on voit sur les vues du spécimen illustré (Vues 270-271 ![]() ) et il se peut que certains taxons exclus des gilianelles soient re-associés par la suite. Mais pour la plupart des "autres" restes décrits ici, les observations ont conduit à mettre en évidence des nannostructures différentes de celles des gilianelles et il est peu probable qu'elles soient proches des gilianelles.
) et il se peut que certains taxons exclus des gilianelles soient re-associés par la suite. Mais pour la plupart des "autres" restes décrits ici, les observations ont conduit à mettre en évidence des nannostructures différentes de celles des gilianelles et il est peu probable qu'elles soient proches des gilianelles.
Par ailleurs, la connaissance actuelle n'a pas de perspective et rien ne permet d'affirmer que des organismes de même généalogie que les gilianelles n'aient pas perdu ce caractère visible de la nannostructure en napperon voire n'aient pas développé une paroi à deux couches comme celle observée parmi les "autres" microproblematica.
La conception actuelle des gilianelles est localisée dans le temps et l'espace et une connaissance plus large serait désirable. Pourtant, il a paru important, après 3 années d'observations de faire le point de ce qui était connu et de formaliser des taxons. C'est dans cette optique qu'il a été jugé nécessaire d'examiner les formes qui ne semblaient pas appartenir au groupe cible des observations mais qui pourraient être confondues : les gilianelles en ressortent plus précisément homogènes.
Cet aspect descriptif de la biodiversité crétacée n'a pas été dissocié de l'aspect pratique :
utilité pour le géologue qui désire caractériser le sédiment qu'il tente de faire parler. Ainsi, l'examen de la distribution et de la fréquence des taxons apporte d'utiles informations qu'il s'agisse des gilianelles ou des autres microproblematica. Le tableau de répartition stratigraphique de cette étude est basé sur une observation de suffisamment longue haleine pour être considéré comme caractérisant une présence significative dans la succession étudiée (espèces rassemblant au moins une quinzaine de récoltes sélectionnées pour la
Fig. 1 ![]() ). Les distributions rapportées indiquent qu'une forte proportion des taxons du groupe des gilianelles (mais aussi certains autres taxons) présentent une extension verticale limitée dans le temps à l'échelle de la succession de Tercis. Ceci dénote une vitesse d'évolution rapide de ces micro-organismes dans ce diverticule de l'Océan Atlantique. Elle se révèle aussi rapide, voire plus rapide que celle des meilleurs marqueurs de temps utiles à la biostratigraphie de cet intervalle de temps fini-Crétacé. Dans cet exercice, la mise en évidence de variations morphologiques continues est un excellent témoignage de cette rapidité et plusieurs exemples en ont été rapportés : évolution des aturelles (A4-A6), des gilianelles (B), des petites jarres (C1).
). Les distributions rapportées indiquent qu'une forte proportion des taxons du groupe des gilianelles (mais aussi certains autres taxons) présentent une extension verticale limitée dans le temps à l'échelle de la succession de Tercis. Ceci dénote une vitesse d'évolution rapide de ces micro-organismes dans ce diverticule de l'Océan Atlantique. Elle se révèle aussi rapide, voire plus rapide que celle des meilleurs marqueurs de temps utiles à la biostratigraphie de cet intervalle de temps fini-Crétacé. Dans cet exercice, la mise en évidence de variations morphologiques continues est un excellent témoignage de cette rapidité et plusieurs exemples en ont été rapportés : évolution des aturelles (A4-A6), des gilianelles (B), des petites jarres (C1).
Lors de la recherche de microfaune dans les niveaux indurés du stratotype de la limite Campanien-Maastrichtien en Aquitaine méridionale, des restes carbonatés originaux et variés ont été mis en évidence. Parmi eux, un ensemble présente des caractéristiques communes permettant de constituer le groupe des gilianelles ; 44 taxons appartenant à ce groupe sont illustrés ici ; 43 sont nouveaux ce qui nous a conduit à proposer 35 nouvelles espèces, 8 sous-espèces, appartenant à 15 genres, en complément de l'espèce type du groupe des gilianelles préalablement désignée et nommée (, 2007).
L'illustration présentée permet de caractériser des taxons qui pourront être reconnus par la suite sans difficulté. Une attention particulière a été portée à la préparation de types choisis comme il est d'usage pour assurer la validité de la description originale (l'holotype éventuellement accompagné de paratypes) mais aussi, dans le cas particulier, une gamme plus large de matériel (topotypes) qui devrait faciliter d'une part, la mise à la disposition de ces formes nouvelles pour d'éventuels curieux et d'autre part, la mise en pratique de la taxinomie élaborée grâce à des restes illustrant une variété de caractères propres à une population en plus de ceux particuliers à l'individu de définition.
Les couches crétacées de Tercis ont ainsi révélé une biodiversité encore plus grande que celle déjà largement illustrée au cours des études de qualification du stratotype pour lesquelles 18 groupes avaient été considérés. Il s'y ajoute aujourd'hui les formes attribuées aux gilianelles, ainsi que d'autres microproblematica pour lesquels 7 genres renfermant 9 espèces et 1 sous-espèce sont décrites.
Cette énumération confirme que les microproblematica rassemblent des populations variées ayant trouvé dans le bassin aturien, au nord des Pyrénées, des conditions de vie favorables au Crétacé terminal. Pour situer cette variété, rappelons que pour 54 formes illustrées ici, plusieurs études sur les ammonites par exemple n'avaient totalisé que deux douzaines de taxons avec un nombre d'individus parfois très restreint tandis que les foraminifères planctoniques considérés comme communs en livraient environ 125 formes (, 2001a) après une longue étude ayant impliqué 4 experts.
Un bon nombre de gilianelles, comme des autres microproblematica présentés révèlent une durée de vie stratigraphique courte avec des tests présents dans des ensembles de niveaux ne dépassant pas 1 Ma comme ordre de grandeur. C'est une durée courte par comparaison avec celle de la grande majorité des formes de microfossiles connues au Crétacé. Les restes étudiés ici permettent d'élaborer des unités stratigraphiques de l'ordre de 0,1 Ma ; par comparaison, la plupart des "zones" de microfossiles (foraminifères, nannofossiles calcaires, kystes de dinoflagellés) comme de macrofossiles (ammonites) de la fin du Crétacé ont des durées de l'ordre du Ma.
La connaissance des gilianelles a maintenant une base solide. Il reste pourtant beaucoup à faire dans quatre domaines : taxinomie, stratigraphie, anatomie, biologie.
Dans l'optique taxinomique, il reste à compléter l'illustration et comprendre une douzaine de gilianelles actuellement codifiées A1c (très petite galette ornée concentriquement), B2c (gilianelle en étoile à 5 bras), B3b (grande gilianelle à 4 bras), E3d' (petit coquetier à microstructure orale en soleil à multiple rayons courts), I1 (très petite ancolie à bras autour et sur la face orale), M4 (petit corail en petit mortier à piler le mil), M5 (petit corail en ombrelle japonaise déjà modélisé par et , 2006), M6 (très petit petit corail), N5 (tercisiennelle en forme de wok), N6 (tercisiennelle en ballon de rugby), N7 (tercisiennelle à quille), ... Mais l'objet prioritaire sera de mettre au point une classification des gilianelles en intégrant les formes restant à définir.
Dans l'optique stratigraphique, il reste à préciser les distributions des espèces rares ou connues d'un seul niveau ; par ailleurs la présence et la distribution des gilianelles dans le Maastrichtien supérieur et jusqu'à la limite Crétacé-Paléogène reste à illustrer. Un second objectif prioritaire sera d'établir l'extension géographique des restes déjà identifiés. Notre souhait est que la présente illustration attirera des curieux qui s'investiront dans cette recherche. L'étude de cette extension et la comparaison des distributions permettra de préciser l'intérêt éventuel des microproblematica du Crétacé terminal pour une application corrélative.
Dans l'optique anatomique, il reste à préciser certains critères de distinction générique basés sur la nannostructure des expansions. C'est une des clés pour comprendre les relations taxinomiques entre les espèces décrites et nous tenterons de suivre le modèle que nous a donné (1975, 1977) pour mettre en évidence les caractères propres aux restes de gilianelles vis à vis des autres groupes de microfossiles.
Dans l'optique biologique, il reste à préciser la signification environnementale locale des gilianelles, notamment leur caractère planctonique ou benthique déjà abordé par ailleurs ( & , 2006) ; leur signification régionale ne pourra être précisée que par des études comparatives en dehors du site de Tercis, objet unique du présent travail.
Ce travail fut rédigé en 2006 et au début de l'année 2007 et divers points soumis à l'attention de divers experts que nous tenons à remercier vivement. Agnès , professeur de lettres classiques, au Collège , Aulnay-sous-bois, a bien voulu nous faire part de remarques linguistiques sur les noms initialement proposés. Jean-Pierre , Correspondant du Muséum National d'Histoire Naturelle, Département systématique et Évolution, nous a fait bénéficier de son expérience dans la pratique des termes taxinomiques en paléontologie et particulièrement sur la question des adjectifs ou des noms en apposition. Agnès , attachée honoraire au Muséum National d'Histoire Naturelle, Département Histoire de la terre a parachevé l'examen des noms et nous a fait bénéficier de sa maîtrise des dictionnaires et des déclinaisons. Pascal nous a précisé quelques points du Code International de Nomenclature Zoologique. Claire , Muséum National d'Histoire Naturelle, Laboratoire d'entomologie, nous a précisé le nom de l'hyménoptère solitaire dont les nids en forme de gilianelle (groupe C1) peuvent s'observer sur les poutres des maisons. Jean-Paul nous a signalé l'existence des Bolboforma et celle de noms génériques qui auraient pu être confondus avec nos propositions antérieures. Paulian n'a pas rechigné à relire deux versions de ce travail lors de la première soumission; Jean-Michel nous a suggéré diverses possibilités de complément de l'étude de ces micro-organismes mais l'objet de ce travail a dű être restreint à la taxinomie. D'abord soumis à une revue de micropaléontologie en Avril 2007, les délais excessifs de prise en considération de notre travail nous ont conduit à solliciter le présent support. Alexandre a mis en page les vues de microscopie électronique et réalisé l'animation. Omar a apporté tous ses soins à la réalisation des prises de vues électroniques. Un grand merci à Nestor grâce à qui les portions en anglais de cette contribution utilisent réellement cette langue. Le lecteur habitué aux Carnets de Géologie reconnaîtra la patte du rédacteur en chef, Bruno , dans la mise en page de la version PDF et dans l'aisance d'accès aux références de la version HTML entièrement de sa main.
P.-Y., G.S., E., & J.-M. (2001).- Microfaciès des sédiments du Campanien et du Maastrichtien de Tercis les Bains (Landes, France). In : G.S. (réd., 2001a), chap. B1b, p. 113-119.
G. (1989).- Les pseudarcellidés, groupe original de microfossiles incertae sedis du Paléogène.- Revue de Micropaléontologie, Paris, vol. 31, n° 4, p. 211-224.
G. (2001).- Introduction à la micropaléontologie.- Gordon & Breach Science Publishers, 258 p.
M., G.S. & B. (2001).- Geology and Late Cretaceous palaeogeography of the geological site at Tercis les Bains (Landes, France). In : G.S. (réd., 2001a), chap. A4, p. 47-59.
M. & J.-P. (2001).- Place of the Charente in the Tethyan domain. In : G.S. (réd., 2001a), chap. A4, note ajoutée sur épreuves, p. 59.
J. & G.S. (2001).- Planktonic Foraminifera from the Campanian-Maastrichtian at Tercis les Bains (Landes, France). In : G.S. (réd., 2001a), chap. C5c, p. 349-370.
J.W.M. & G.S. (2001).- Campanian-Maastrichtian crinoids (Echinodermata) from Tercis les Bains (Landes). In : G.S. (réd., 2001a), chap. D7, p. 635-644.
H. (1979).- Lower Cretaceous Calcisphaerulidae and their relationship to calcareous dinoflagellate cysts.- Bulletin des Centres de Recherche Exploration-Production Elf-Aquitaine, Pau, vol. 3, n° 2, p. 651-663.
H. (1984).- Revision der kalkigen Dinoflagellaten-Zysten G. , 1948.- Paläontologisches Zeitschrift, Stuttgart, vol. 58, n° 1-2, p. 9-31.
H. & J. (1994).- Calcareous phytoplankton from the Barremian/Aptian boundary interval from NW Germany.- Cretaceous Research, London, vol. 15, n° 6, p.739-763.
Y., J., M., E. & G.S. (2001).- Campanian-Maastrichtian continental-marine connection in the Aquitaine-Pyrenees-Provence area (S France). In : G.S. (réd., 2001a), chap. D10, p. 657-665.
Z. & G.S. (2001).- Magnetostratigraphy across the Campanian-Maastrichtian boundary at Tercis les Bains in comparison with northern Germany, the Apennines (Central Italy) and North America; biostratigraphical and geochronological constraints. In : G.S. (réd., 2001a), chap. B2d, p. 175-183.
D. & G.S. (2001).- Echinoids from the geological site at Tercis les Bains (Landes, France). In : G.S. (réd., 2001a), chap. D6, p. 612-634.
G.S. (rédacteur, 2001a).- The Campanian - Maastrichtian stage boundary. Characterisation at Tercis les Bains (France) and correlation with Europe and other continents.- IUGS Special Publication (monograph) Series, 36; Developments in Palaeontology and Stratigraphy Series, 19, Elsevier Sciences Publ. Amsterdam, xxviii + 881 p.
G.S. (2001b).- The Campanian-Maastrichtian boundary: definition at Tercis (Landes, SW France) principle, procedure, and proposal. In : G.S. (réd., 2001a), chap. F3, p. 820-833.
G.S. (2001c).- Descriptive lithostratigraphy of the Campanian-Maastrichtian succession at Tercis les Bains (SW France). In : G.S. (réd., 2001a), chap. B1a, p. 85-109.
G.S. (2001d).- Numerical age calibration of the Campanian-Maastrichtian succession at Tercis les Bains (Landes, France) and in the Bottaccione Gorge (Italy). In : G.S. (réd., 2001a), chap. E5c, p. 775-782.
G.S. (2001e).- Les fossiles du site géologique de Tercis les Bains (Landes, France). In : G.S. (réd., 2001a), chap. B1d, p. 134-160.
G.S. (2007).- Sur des microfossiles calcaires problématiques du stratotype de la limite Campanien-Maastrichtien de Tercis (Sud-Ouest, France) : Les gilianelles.- Comptes rendus Palévol, Paris, vol. 6, n° 3, p. 181-188.
G.S., C., A. & B. (2006).- Présentation du site géologique de Tercis (Landes, France) ; 13 affiches pédagogiques informatisées.
a- Le site de la Grande Carrière de Tercis ; haut lieu géologique, témoin des 10 Ma de la fin du Crétacé.
b- Les grands fossiles de la carrière de Tercis; une richesse et une diversité remarquable en macrofossiles.
c- Les bivalves de Tercis ; des fossiles marqueurs de l'environnement et un groupe marqueur de temps (les inocérames).
d- Les ammonites de Tercis ; un groupe disparu depuis 65 Ma, proche des nautiles et des seiches, très utile pour dater les sédiments de plate-forme.
e- Les échinides de Tercis.
f- Les astérides de Tercis; des fossiles communs sur la plate-forme continentale et utiles à la datation relative.
g- Les petits fossiles de la carrière de Tercis.
h- Les nannofossiles calcaires à Tercis.
i- Les kystes de dinoflagellés ; outil stratigraphique de datation relative excellemment illustré à Tercis.
j- Les foraminifères de la Grande Carrière de Tercis; une présence suffisante pour une datation relative acceptable.
l- Datation comparée Continent-Océan à Tercis.
m- Les sites préhistoriques à Tercis ; une grande carrière sur le plus grand site de plein air du sud-ouest de la France.
n- Le site de Tercis, stratotype de référence mondial ; définition du calendrier géologique et application à la limite d'étage Campanien-Maastrichtien.
En ligne sur internet (depuis le 14-VI-06): http://paleopolis.rediris.es/odin/
G.S., C., A. & B. (2007).- Gilianelles et autres microfossiles problématiques : Contribution à l'étude de la paléobiodiversité et à la biostratigraphie crétacée dans le stratotype de la limite Campanien-Maastrichtien à Tercis (Sud Aquitaine, France).- Journée Hommage à Gérard , Paris 06-XII-2007.
En ligne sur internet (depuis le 21-XI-07): http://paleopolis.rediris.es/odin/images/poster-14.png
G.S. & A. (2001).- Interpretative reading of the Campanian-Maastrichtian deposits at Tercis les Bains: sedimentary breaks, rhythms, accumulation rate, sequences. In : G.S. (réd., 2001a), chap. B1c, p. 120-131.
G.S., J.-A., M., J. & E. (2001).- Campanian-Maastrichtian planktonic foraminifera at Tercis les Bains (Landes, France); synthetic view and potential for global correlation. In : G.S. (réd., 2001a), chap. C5d, p. 379-395.
G.S. & M.A. (2001).- The global Campanian-Maastrichtian stage boundary at Tercis les Bains, Landes, SW France.- Episodes, Beijing, vol. 24, n° 4, p. 229-238.
G.S. & A. (2006).- Nouvelles études micropaléontologiques sur le stratotype de la limite Campanien-Maastrichtien à Tercis (SO France) : Les gilianelles (microproblematica) extraites par acétolyse.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2006/05 (CG2006_A05), 28 p., 3 videos.
G.S. & P. (1994).- Première description d'une coupe dans la localité aturienne de Tercis (Landes) : Contribution à l'étude de la limite Campanien-Maastrichtien.- Géologie de la France, Orléans, n° 2 (1994), p. 31-37.
G.S. & I. (2003).- Sur les inocérames de Tercis (Landes, France) : Le meilleur outil corrélatif entre Europe et Amérique du Nord autour de la limite Campanien-Maastrichtien / Inoceramids of the site of Tercis (Landes, France): the best correlative tool between Europe and North America across the Campanian-Maastrichtian boundary.- Comptes rendus Géosciences, Paris, vol. 335, n° 2, p. 239-246.
A. (1990).- Nouveaux microproblematica : Yvonniellina parva n. sp. ; Voorthuyseniella bearnensis n. sp. et Pseudarcella sp. dans l'Éocène d'Aquitaine (Sud-Ouest de la France).- Revue de Micropaléontologie, Paris, vol. 33, n° 2, p. 115-122.
M. (2001).- The site at Tercis les Bains (Landes), stratotype of the Campanian-Maastrichtian limit : insights from tectonic studies. In : G.S. (réd., 2001a), chap. A6, p. 68-81.
D. & C.H. (1991). A stratigraphic and taxonomic atlas of Bolboforma (Protophytes, incertae sedis, Tertiary).- Journal of Foraminiferal Research, Lawrence, vol. 21, n° 2, p. 126-158.
K. (2000).- Les foraminifères de l'Eocène du Bassin de l'Adour (Aquitaine, France) : biostratigraphie et taxinomie.- Revue de Micropaléontologie, Paris, vol. 43, n° 1, p. 71-172, 23 pl.
J.-M. (1975).- Calcisphaerulidae (incertae sedis) du Crétacé supérieur du Limbourg (Pays-Bas) et d'autres régions.- Palaeontographica, Stuttgart, Abt. 149, p. 193-242.
J.M. (1977).- Les calcisphaerulidae : architectures, calcification de la paroi et phylogenèse.- Palaeontographica, Stuttgart, Abt. 159, p. 139-177.
I., G.S. & A.V. (2002).- Inoceramids from the Upper Campanian and Lower Maastrichtian of the Tercis section (SW France), the Global Stratotype Section and Point for the Campanian - Maastrichtian boundary; taxonomy, biostratigraphy and correlation potential.- Acta Geologica Polonica, Warsaw, vol. 52, n° 3, p. 269-305.
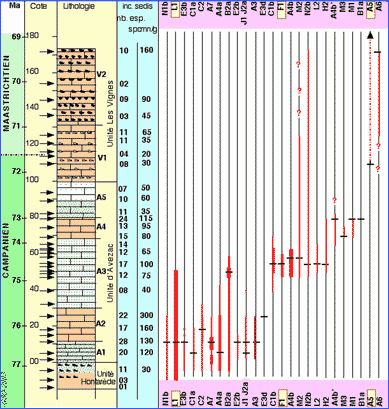
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 1 : Distribution des microproblematica les plus significatifs au plan stratigraphique dans la Grande Carrière de Tercis (Aquitaine, stratotype de limite Campanien-Maastrichtien), gilianelles et autres microproblematica (codes encadrés). La localisation stratigraphique est donnée en millions d'années le long de la colonne des étages d'après et
(2003). Les niveaux traités sont notés par une flèche à leur cote respective à gauche de la lithologie. Le nombre de formes reconnues est indiqué (nb. esp.) pour la plupart des niveaux, suivi d'une estimation minimale du nombre total de spécimens par gramme de sédiment traité (spcmn/g). La répartition verticale de 25 taxons choisis (identifiés par leur code alphanumérique) est indiquée à droite. L'abondance absolue est soulignée par un trait plus épais. Un tiret horizontal localise le niveau dans lequel l'holotype a été désigné.
Distribution of the taxa with the most significance stratigraphically (Grande Carrière at Tercis, Aquitaine, Campanian-Maastrichtian boundary stratotype). The stratigraphic age in millions of years is on the left of the columnar section (after and ,
2003). Arrows at the left of the lithology show the respective locations of the treated samples. The lithology column shows glaucony
(dotted green) grey flint (open signs), and dark flint (black signs). The two columns to the right of the named units give respectively (left and right) the number of identified species (nb. esp.) for most levels and an estimate of the minimum total number of specimens per gramme of treated sediment (spcmn/g). The distribution of 25 selected taxa identified by their code (top and bottom: gilianelles codes bare; other microproblematica framed) is shown to the right. The greater abundance is indicated by a thicker bar. A horizontal dash on the range bar indicates the level from which the holotype was selected.
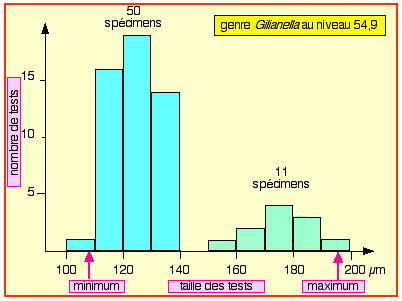
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 2 : Diamètre du corps des Gilianella par classes de 10 en 10 µm au niveau 54,9 de la Grande Carrière de Tercis (SO France) ; tri de 61 spécimens ; les tailles de la plus petite et de la plus grande sont indiquées (flèches). Deux populations sont présentes : B2a Gilianella stellata est plus petite que Gilianella major sp.
nov.
Bar graph: diameter of the body of Gilianella per 10 µm class from 100 to 200 µm at level 54.9 from the Grande Carrière at Tercis (SO France). 61 specimens measured, the sizes of the smallest and largest are indicated by arrows. Two populations are present: B2a (Gilianella stellata) is distinctly smaller than B3a (Gilianella major sp.
nov.).

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Cliquer sur l'image pour découvrir l'animation / Click on image to play the video file (flv = 1,550 KB)
ou cliquer sur l'icone pour télécharger la vidéo au format avi / or click on the icon ![]() to download another version of video file (avi = 1,323 KB)
to download another version of video file (avi = 1,323 KB)
ou cliquer sur l'icone pour ouvrir une nouvelle fenêtre avec l'animation au format mpg / or click on the icon ![]() to open and play another version of video file (mpg = 827 KB)
to open and play another version of video file (mpg = 827 KB)
ou cliquer sur l'icone pour télécharger la vidéo au format wmv / or click on the icon ![]() to download another version of video file (wmv = 629 KB)
to download another version of video file (wmv = 629 KB)
Figure 3 : Reconstitution en 3 dimensions de Aquilegiella ericius
(H5), réalisation A. .
Three dimensional reconstruction of Aquilegiella ericius (H5) by A. . The organisation of the spines on the body of the test is constant.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 4 : Modélisation de la gilianelle Aturella angulata (A4b'). Noter que, par comparaison avec les premières modélisations publiées pour le genre ( & ,
2006), ces représentations mettent en évidence une symétrie bilatérale dans les vues orale et aborale et une direction dans les vues de profil et cavalière (réalisation A. ).
A three-dimensional model of the gilianelle Aturella angulata (A4b'). Note that when compared with earlier 3-D models of the genus ( & ,
2006), these new representations show a bilateral symmetry in oral and aboral views and indicate an antero-posterior differentiation (drawing A. ).

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 1 : Gilianelles vues au microscope électronique, grossies 240 fois ; vues de profil avec l'orifice vers le haut sauf indication contraire.
Vues 1-5, gilianelle Tercensella globosa (N2b) : 1- vue cavalière orale, niveau 54,9 ; 2- profil, orifice vers le haut, niveau 54,9 ; 3- vue aborale, niveau 54,9 ; 4- vue cavalière orale, même spécimen que 2 ; 5- vue cavalière orale, niveau 172.
Vues 6-10, gilianelle Tercensella doliolum (N1b) : 6- vue orale, niveau 11,0 ; 7- profil, niveau 11,0 ; 8- vue aborale, niveau 11,0 ; 9- vue cavalière orale, niveau 11,0 ; 10- profil, même spécimen que 9.
Vues 11-13, gilianelle Tercensella alveare (N1a), même spécimen du niveau 54,9 : 11- vue cavalière orale ; 12- profil ; 13- vue cavalière
aborale.
Vues 14-18, gilianelle Azymella cannabinata (A7) : 14- vue orale, niveau 11,0 ; 15- vue cavalière aborale, orifice vers le bas, bord manquant vers l'observateur, niveau 11,0 ; 16- vue aborale, niveau 11,0 ; 17- vue aborale, niveau 11,0 ; 18- vue orale, niveau +5,0.
Plate 1: Gilianelles enlarged 240 times; profile views with the aperture upward in all but view 15.
Images 1-5, gilianelle Tercensella globosa (N2b): 1- oral oblique view, level 54.9; 2- profile, level 54.9; 3- aboral view, level 54.9; 4- oral oblique view, same specimen as view 2; 5- oral oblique view, level 172.
Images 6-10, gilianelle Tercensella doliolum (N1b): 6- oral view, level 11.0; 7- profile, level 11.0; 8- aboral view, level 11.0; 9- oral oblique view, level 11.0; 10- profile, same spacimen as 9.
Images 11-13, gilianelle Tercensella alveare (N1a): one specimen, level 54.9; 11- oral oblique view; 12- profile; 13- aboral oblique view.
Images 14-18, gilianelle Azymella cannabinata (A7): 14- oral view, level 11.0; 15- aboral oblique view, aperture at bottom, damaged specimen shows the structure of the wall, level 11.0; 16- aboral view, level 11.0; 17- aboral view, level 11.0; 18- oral view, level +5.0.
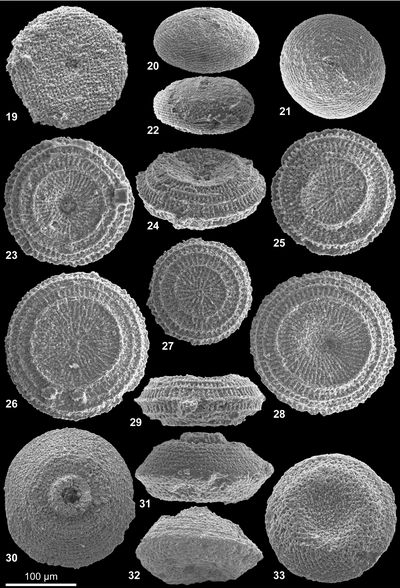
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 2 : Gilianelles vues au microscope électronique, grossies 240 fois ; vues de profil avec l'orifice vers le haut.
Vue 19, gilianelle Azymella cannabinata (A7) : vue orale, niveau -5,0.
Vues 20-22, gilianelle Azymella offa (A8), niveau 79,8 : 20- vue cavalière orale ; 21- vue
orale, même spécimen ; 22- vue cavalière orale, test incomplet.
Vues 23-29, gilianelle Numismella tarbellica (A1a) : 23- vue orale, niveau 11,0 ; 24- vue cavalière
orale, niveau 172 ; 25- vue aborale, niveau 172 ; 26- vue aborale, niveau 54,9 ; 27- vue
aborale, petit spécimen, niveau +5,0 ; 28- vue orale, niveau 54,9 ; 29-
profil, orifice vers le haut, même spécimen que 24.
Vues 30-33, gilianelle Aturella aequilatera (A4b) : 30- vue orale, niveau 54,9 ; 31-
profil, avant à gauche, niveau 54,9 ; 32- vue en profil cavalier, avant à gauche, niveau 39,5 ; 33- vue
aborale, niveau 54,7).
Plate 2: Gilianelles enlarged 240 times; profile views with the aperture upward.
Image 19, gilianelle Azymella cannabinata (A7): oral view, level -5.0.
Images 20-22, gilianelle Azymella offa (A8): Level 79.8; 20- oral oblique view; 21- oral view, same specimen; 22- oral oblique view, test partially removed.
Images 23-29, gilianelle Numismella tarbellica (A1a): 23- oral view, level 11.0; 24- oral oblique view, level 172; 25- aboral view, level 172; 26- aboral view, level 54.9; 27- aboral view, small specimen, level +5.0; 28- oral view, level 54.9; 29- profile, same specimen as 24.
Images 30-33, gilianelle Aturella aequilatera (A4b): 30- oral view, level 54.9; 31- profile, front to the left, level 54.9; 32- oblique profile, front to the left, level 39.5; 33- aboral view, level 54.7.
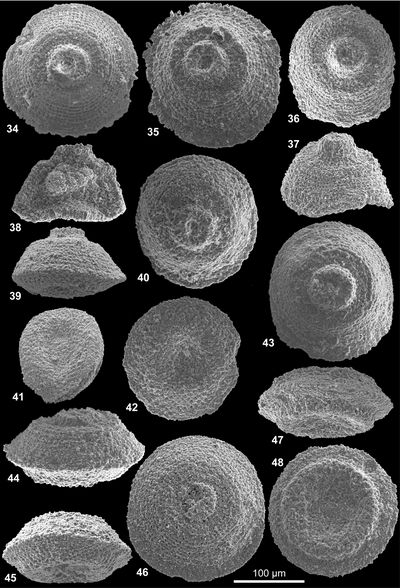
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 3 :
Gilianelles vues au microscope électronique, grossies 240 fois ; vues de profil avec l'orifice vers le haut ; avant vers le haut lorsqu'il est identifiable.
Vues 34-35, gilianelle Aturella aequilatera (A4b) : 34- vue orale, niveau 57,9 ; 35- vue
orale, niveau 47,2.
Vues 36-43, gilianelle Aturella altodepressa (A4a) : 36- vue orale, niveau 5,0 ; 37- vue de profil arrière gauche, même individu ; 38- section de
profil, niveau +5,0 ; 39- profil, avant à gauche, niveau -5,0 ; 40- vue
orale, niveau 11,0 ; 41- vue aborale, petit individu, niveau +5,0 ; 42- vue
aborale, niveau 11,0 ; 43- vue orale, niveau 18,2.
Vues 44-48, gilianelle Aturella angulata (A4b') : 44- profil oral, niveau 69,0 ; 45- profil aboral du précédent ; 46- vue
orale, niveau 79,8 ; 47- profil aboral, niveau 79,8 ; 48- vue aborale du précédent.
Plate 3: Gilianelles enlarged 240 times; profile views with the aperture upward; front upward when recognizable.
Images 34-35, gilianelle Aturella aequilatera (A4b): 34- oral view, level 57.9; 35- oral view, level 47.2.
Images 36-43, gilianelle Aturella altodepressa (A4a): 36- oral view, level 5.0; 37- profile from left back side, same specimen; 38- profile section, level +5.0; 39- profile, front -anterior of the test- to the left, level -5.0; 40- oral view, level 11.0; 41- aboral view of small specimen, level +5.0; 42- aboral view, level 11.0; 43- oral view, level 18.2.
Images 44-48, gilianelle Aturella angulata (A4b'): 44- oral profile, level 69.0; 45- aboral profile of the same; 46- oral view, level 79.8; 47- profile
aboral, level 79.8; 48- aboral view of the same.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 4 :
Gilianelles vues au microscope électronique, grossies 240 fois ; vues de profil avec l'orifice vers le haut ; avant vers le haut lorsqu'il est identifiable.
Vues 49-52, gilianelle Aturella edentula (A6) du niveau 172 : 49- vue orale ; 50- vue
aborale, avant vers le haut ; 51- profil, avant à gauche ; 52- profil oral, avant à
droite.
Vues 53-59, gilianelle Scutellella crassa (A1b) : 53- vue orale, niveau 89,0 ; 54- vue
aborale, niveau 79,8 ; 55- vue cavalière aborale, même spécimen que 54 ; 56- vue cavalière
orale, niveau 89,0 ; 57- profil oral, même spécimen que 56 ; 58-
profil, niveau 172 ; 59- profil, niveau 11,0.
Vues 60-64, gilianelle Scutellella pulchrevelifera (A3) : 60- vue
orale, niveau +5,0 ; 61- profil, orifice vers le bas, niveau 11,0 ; 62- vue cavalière
aborale, orifice vers le bas, même spécimen que 61 ; 63- vue aborale, niveau 11,0 ; 64- vue
orale, niveau 5,0.
Plate 4: Gilianelles enlarged 240 times; profile views with the aperture upward in all except 61; front upward when visible.
Images 49-52, gilianelle Aturella edentula (A6) of level 172: 49- oral view; 50- aboral view of an oriented specimen; 51- profile, front to the left; 52- profile oral, front to the right.
Images 53-59, gilianelle Scutellella crassa (A1b): 53- oral view, level 89.0; 54- aboral view, level 79.8; 55- aboral oblique view, same specimen; 56- oral oblique view, level 89.0; 57- oral profile, same specimen; 58- profile, level 172; 59- profile, level 11.0.
Images 60-64, gilianelle Scutellella pulchrevelifera (A3): 60- oral view, level +5.0; 61- profile, aperture to the bottom, level 11.0; 62- aboral oblique view, same specimen; 63- aboral view, level 11.0; 64- oral view, level 5.0.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 5 :
Gilianelles vues au microscope électronique, grossies 240 fois ; vues de profil avec l'orifice vers le haut.
Vues 65-70, gilianelle Scutellella circularis (A2a) : 65- vue orale, niveau 54,9 ; 66- vue
aborale, niveau 54,9 ; 67- vue aborale, niveau 79,8 ; 68- vue orale, niveau 57,9 ; 69- profil cavalier
aboral, niveau 79,8 ; 70- vue orale, niveau 54,9.
Vues 71-77, gilianelle Scutellella longivelifera (A2b) : 71- vue
orale, avant à droite, niveau +5,0 ; 72- profil court, niveau +5,0 ; 73- profil long, cavalier
aboral, même spécimen que 72 ; 74- vue orale, niveau 54,9 ; 75- vue
aborale, avant à gauche, niveau +5,0 ; 76- vue aborale, avant à
droite, niveau 50,0 ; 77- vue orale, niveau 57,9.
Vues 78-80, gilianelle Corniculum sinuosum (L2) : 78- vue aborale, niveau 50,0 ; 79-
profil, niveau 62,5 ; 80- vue cavalière orale, niveau 57,9.
Plate 5: Gilianelles enlarged 240 times; profile views with the aperture upward.
Images 65-70, gilianelle Scutellella circularis (A2a): 65- oral view, level 54.9; 66- aboral view, level 54.9; 67- aboral view, level 79.8; 68- oral view, level 57.9; 69- aboral oblique profile, level 79.8; 70- oral view, level 54.9.
Images 71-77, gilianelle Scutellella longivelifera (A2b): 71- oral view, front to the right, level +5.0; 72- short profile, level +5.0; 73- long oblique aboral profile, same specimen; 74- oral view, level 54.9; 75- aboral view, front to the left, level +5.0; 76- aboral view, front to the right, level 50.0; 77- oral view, level 57.9.
Images 78-80, gilianelle Corniculum sinuosum (L2): 78- aboral view, level 50.0; 79- profile, level 62,5; 80- oral oblique view, level 57.9.
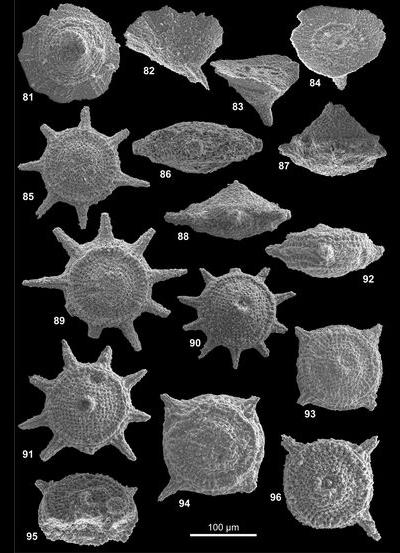
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 6 : Gilianelles vues au microscope électronique, grossies 240 fois ; vues de profil avec l'orifice vers le haut.
Vues 81-82, gilianelle Corniculum sinuosum (L2) : 81- vue aborale, niveau 57,9 ; 82- profil
aboral, du même.
Vues 83-84, gilianelle Corniculum clausum (L3), holotype du niveau 79,8 : 83- profil oral ; 84- vue cavalière
orale.
Vues 85-91, gilianelle Gilianella stellata (B2a) : 85- vue aborale, spécimen exceptionnel à 7 bras, niveau 50,0 ; 86-
profil, niveau 50,0 ; 87- profil, niveau 25,0 ; 88- profil, niveau 50,0 ; 89- vue
aborale, spécimen à 9 bras, niveau 54,9 ; 90- vue orale, spécimen rare à 10 bras, niveau 54,9 ; 91- vue
orale, spécimen à 9 bras dont 1 brisé, niveau 54,9.
Vues 92-96, gilianelle Gilianella stellata pulvinata (B2b) : 92-
profil, niveau 11,0 ; 93- vue orale, niveau 11,0 ; 94- vue aborale, niveau 18,2 ; 95- vue cavalière
orale, spécimen cassé montrant la section, niveau 11,0 ; 96- vue
orale, niveau +5,0.
Plate 6: Gilianelles enlarged 240 times; profile views with the aperture upward.
Images 81-82, gilianelle Corniculum sinuosum (L2): 81- aboral view, level 57.9; 82- aboral profile of the same.
Images 83-84, gilianelle Corniculum clausum (L3): holotype of level 79.8; 83- oral profile; 84- oral oblique view.
Images 85-91, gilianelle Gilianella stellata (B2a): 85- aboral view, unusual specimen with 7 arms, level 50.0; 86- profile, level 50.0; 87- profile, level 25.0; 88- profile, level 50.0; 89- aboral view, specimen with 9 arms, level 54.9; 90- oral view, unusual specimen with 10 arms, level 54.9; 91- oral view, specimen with 9 arms, one lacking, level 54.9.
Images 92-96, gilianelle Gilianella stellata pulvinata (B2b): 92- profile, level 11.0; 93- oral view, level 11.0; 94- aboral view, level 18.2; 95- oral oblique view, broken specimen showing the structure of the wall, level 11.0; 96- oral view, level +5.0.
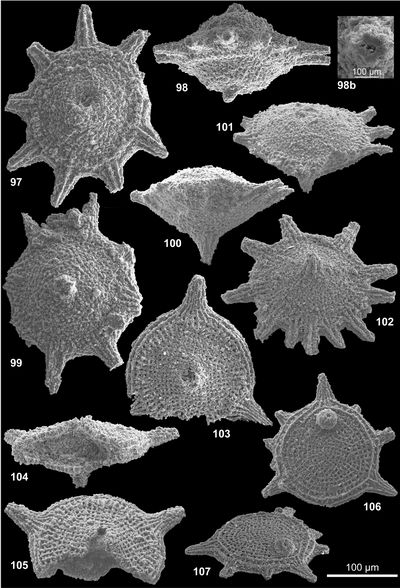
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 7 :
Gilianelles vues au microscope électronique, grossies 240 fois ; vues de profil avec l'orifice vers le haut sauf vues 102 et 107.
Vues 97-99, gilianelle Gilianella major (B3a) : 97- vue orale, niveau 50,0 ; 98-
profil, niveau 79,8 ; 98b- section proximale d'un bras agrandie ; 99- vue
aborale, niveau 54,9.
Vues 100-102, gilianelle Gilianella major heliotropium (B4) : 100- profil de
l'holotype, niveau 172 ; 101- vue cavalière orale de l'holotype ; 102- vue cavalière
aborale, niveau 50,0.
Vues 103-105, gilianelle Gilianella terbrachialis (B3d) : 103- vue
orale, partie basse endommagée, niveau 50,0 ; 104- profil, test ouvert, niveau 50,0 ; 105- vue orale du
même.
Vues 106-107, gilianelle Gilianella terbrachialis mutata (B3d') : 106- vue
aborale, niveau 79,8 ; 107- profil aboral du précédent, orifice vers le bas.
Plate 7: Gilianelles enlarged 240 times; profile views with the aperture upward except in views 102, 107.
Images 97-99, gilianelle Gilianella major (B3a): 97- oral view, level 50.0; 98- profile, level 79.8; 98b- enlarged proximal section of an arm; 99- aboral view, level 54.9.
Images 100-102, gilianelle Gilianella major heliotropium (B4): 100- profile of the
holotype, level 172; 101- oral oblique view of the holotype; 102- aboral oblique view, aperture to the bottom, level 50.0.
Images 103-105, gilianelle Gilianella terbrachialis (B3d): 103- oral view, lower portion damaged, level 50.0; 104- profile, open test, level 50.0; 105- oral view of the same.
Images 106-107, gilianelle Gilianella terbrachialis mutata (B3d'): 106- aboral view, level 79.8; 107- aboral profile of the same, aperture to the bottom (note the presence of a sticked micro-sphere at five o'clock).

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 8 :
Gilianelles vues au microscope électronique, grossies 240 fois ; vues de profil avec l'orifice vers le haut sauf indication contraire.
Vue 108, gilianelle Gilianella terbrachialis mutata (B3d') : vue
orale, niveau 79,8.
Vues 109-113, gilianelle Gilianella tenuibrachialis (B1a) du niveau 79,8 : 109- vue aborale ; 110- vue orale ; 111- profil ; 112- vue cavalière aborale du même ; 113- vue
aborale, unique spécimen très petit (peut-être une autre espèce).
Vues 114-120, gilianelle Caccabella spinocincta (C2) : 114- vue sub-orale, niveau 11,0 ; 115-
profil, niveau 18,2 ; 116- profil, niveau +5,0 ; 117- vue cavalière aborale du précédent ; 118- vue cavalière
aborale, même spécimen que 115 ; 119- vue cavalière orale, niveau 18,25 ; 120- vue
aborale, niveau +5,0.
Vues 121-122, gilianelle Caccabella spinocincta ventriosa (C2b) du niveau 11,0 : 121- profil ; 122- vue
orale.
Plate 8: Gilianelles enlarged 240 times; profile views with the aperture upward.
Image 108, gilianelle Gilianella terbrachialis mutata (B3d'): oral view, level 79.8.
Images 109-113, gilianelle Gilianella tenuibrachialis (B1a) of level 79.8: 109- aboral view; 110- oral view; 111- profile; 112- aboral oblique view of the same; 113- aboral view, unique very small specimen (possibly another species).
Images 114-120, gilianelle Caccabella spinocincta (C2): 114- oblique oral view, level 11.0; 115- profile, level 18.2; 116- profile, level +5.0; 117- aboral oblique view of the same; 118- aboral oblique view, same specimen as 115; 119- oral oblique view, level 18.25; 120- aboral view, level +5.0.
Images 121-122, gilianelle Caccabella spinocincta ventriosa (C2b) of level 11.0: 121- profile; 122- oral view;

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 9 : Gilianelles vues au microscope électronique, grossies 240 fois ; vues de profil avec l'orifice vers le haut.
Vues 123-125, gilianelle Caccabella spinocincta ventriosa (C2b) du niveau 11,0 : 123- vue aborale ; 124 et 125- 2 spécimens en vue cavalière
aborale.
Vues 126-130, gilianelle Orculiella niduseumenei (C1b) : 126- vue cavalière
orale, niveau 54,9 ; 127- profil du précédent ; 128- vue
orale, niveau 50,0 ; 129- vue cavalière aborale, niveau 39,5 ; 130- vue
aborale, niveau 50,05.
Vues 131-136, gilianelle Orculiella niduseumenei strangulata (C1a) : 131-
profil, niveau 11,0 ; 132- vue cavalière aborale du précédent ; 133-
profil, spécimen cassé avec section montrant la face aborale déprimée, niveau +5,0 ; 134- vue
orale, niveau 5,0 ; 135- profil du précédent ; 136- vue orale partielle du
col, grossi 2 fois plus, niveau +5,0.
Vues 137, 139-141, gilianelle Pennigerella palmata alta (J1) : 137- vue
orale, niveau +5,0 ; 139- vue cavalière orale, niveau +5,0 ; 140- vue
aborale, niveau +5,0 ; 141- vue cavalière aborale du précédent.
Vue 138, gilianelle Corbella vitilis (G0) : vue orale, niveau 11,0.
Plate 9: Gilianelles enlarged 240 times; profile views with the aperture upward.
Images 123-125, gilianelle Caccabella spinocincta ventriosa (C2b) of level 11.0: 123- aboral view; 124 and 125- aboral oblique views of 2 specimens.
Images 126-130, gilianelle Orculiella niduseumenei (C1b): 126- oral oblique view, level 54.9; 127- profile of the same; 128- oral view, level 50.0; 129- aboral oblique view, level 39.5; 130- aboral view, level 50.05.
Images 131-136, gilianelle Orculiella niduseumenei strangulata (C1a): 131- profile, level 11.0; 132- aboral oblique view of the same; 133- profile, broken specimen showing the section and the depressed aboral face, level +5.0; 134- oral view, level 5.0; 135- profile of the same; 136- oral view of the aperture, enlarged twice more, level +5.0.
Images 137, 139-141, gilianelle Pennigerella palmata alta (J1): 137- oral view, level +5.0; 139- oral oblique view, level +5.0; 140- aboral view, level +5.0; 141- aboral oblique view of the same.
Image 138, gilianelle Corbella vitilis (G0): oral view, level 11.0.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 10 :
Gilianelles vues au microscope électronique, grossies 240 fois ; vues de profil avec l'orifice vers le haut.
Vues 142-149, gilianelle Pennigerella palmata humilis (J2a) : 142- vue
orale, niveau 11,0 ; 143- profil du précédent, noter le test peu élevé ; 144-
profil, niveau 11,0 ; 145- vue orale du précédent ; 146- vue cavalière
orale, niveau 11,0 ; 147- vue orale du précédent ; 148- vue
aborale, niveau 11,0 ; 149- vue aborale, niveau 11,0.
Vues 150-154, gilianelle Pennigerella castulata (J2b) : 150- vue cavalière
orale, niveau 11,0 ; 151- vue cavalière orale, niveau +5,0 ; 152- profil du précédent ; 153 et 154- 2 vues cavalières aborales du même spécimen.
Vues 155-159, gilianelle Corbella vitilis (G0) : 155- vue orale, niveau 18,25 ; 156 et 157- 2 profils d'un même
individu, niveau 11,0 ; 158- vue aborale, niveau 11,0 ; 159- vue cavalière aborale du précédent.
Vues 160-163, gilianelle Corbella vitilis biremis (G0b) : 160- vue
aborale, niveau 50,0 ; 161- vue cavalière aborale, niveau 54,9 ; 162- vue orale du précédent ; 163- vue cavalière orale du
même.
Vue 164, gilianelle Aquilegiella varia (H2) : vue cavalière
orale, spécimen incomplètement dégagé, niveau 54,9.
Plate 10: Gilianelles enlarged 240 times; profile views with the aperture upward.
Images 142-149, gilianelle Pennigerella palmata humilis (J2a): 142- oral view, level 11.0; 143- profile of the same, note the low height of the test; 144- profile, level 11.0; 145- oral view of the same; 146- oral oblique view, level 11.0; 147- oral view of the same; 148- aboral view, level 11.0; 149- aboral view, level 11.0.
Images 150-154, gilianelle Pennigerella castulata (J2b): 150- oral oblique view, level 11.0; 151- oral oblique view, level +5.0; 152- profile of the same; 153 et 154- 2 aboral oblique views of the same.
Images 155-159, gilianelle Corbella vitilis (G0): 155- oral view, level 18.25; 156 et 157- 2 profiles of a same specimen, level 11.0; 158- aboral view, level 11.0; 159- aboral oblique view of the same.
Images 160-163, gilianelle Corbella vitilis biremis (G0b): 160- aboral view, level 50.0; 161- aboral oblique view, level 54.9; 162- oral view of the same; 163- oral oblique view of the same.
Image 164, gilianelle Aquilegiella varia (H2): oral oblique view, specimen with adhering matrix, level 54.9.
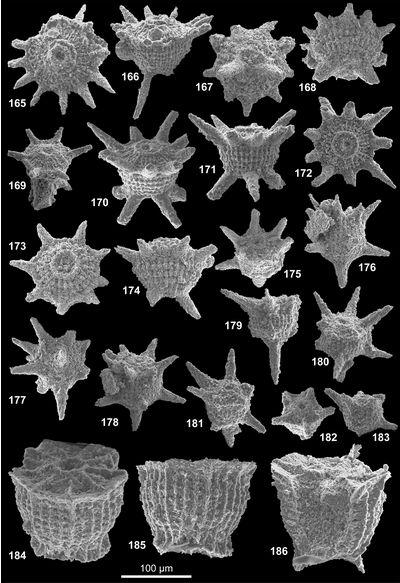
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 11 :
Gilianelles vues au microscope électronique, grossies 240 fois ; vues de profil avec l'orifice vers le haut.
Vues 165-174, gilianelle Aquilegiella varia (H2) : 165- vue orale, niveau 54,9 ; 166- profil du précédent ; 167- vue
aborale, niveau 50,0 ; 168- vue cavalière aborale du précédent ; 169- vue cavalière orale d'un petit
individu, niveau 79,8 ; 170- vue cavalière orale, niveau 57,9 ; 171- profil du précédent ; 172- vue orale du même spécimen ; 173- vue
orale, niveau 50,0 ; 174- profil du précédent.
Vues 175-183, gilianelle Aquilegiella ericius (H5) du niveau 50,0 : 175- vue cavalière orale ; 176- profil ; 177- vue
aborale, pied axial vers l'observateur ; 178- profil cavalier oral, même spécimen que 176 ; 179-
profil, tous les pieds latéraux brisés ; 180 & 181- 2 profils cavaliers oraux du même spécimen tourné de 90° ; 182- vue cavalière orale d'un petit individu de la
même (?) espèce ; 183- profil du précédent.
Vues 184-186, gilianelle Pocillella grandicula (E3a) du niveau 11,0 : 184- profil cavalier oral ; 185- profil du précédent ; 186- profil d'un spécimen
ouvert.
Plate 11: Gilianelles enlarged 240 times; profile views with the aperture upward.
Images 165-174, gilianelle Aquilegiella varia (H2): 165- oral view, level 54.9; 166- profile of the same; 167- aboral view, level 50.0; 168- aboral oblique view of the same; 169- oral oblique view of a small specimen, level 79.8; 170- oral oblique view, level 57.9; 171- profile of the same; 172- oral view of the same; 173- oral view, level 50.0; 174- profile of the same.
Views 175-183, gilianelle Aquilegiella ericius (H5) of level 50.0: 175- oral oblique view; 176- profile; 177- aboral view, axial foot in front; 178- oblique oral profile, same specimen as 176; 179- profile, all lateral feet broken; 180 & 181- 2 oblique oral profiles of the same specimen; 182- oblique oral view of a small specimen possibly of the same species; 183- profile of the same.
Images 184-186, gilianelle Pocillella grandicula (E3a) of level 11.0: 184- oblique oral profile; 185- profile of the same; 186- profile of a broken specimen.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 12 :
Gilianelles vues au microscope électronique, grossies 240 fois ; vues de profil avec l'orifice vers le haut.
Vues 187-188, gilianelle Pocillella grandicula (E3a) du niveau 11,0 : 187- profil cavalier aboral ; 188- vue aborale du précédent.
Vues 189-194, gilianelle Pocillella minuta (E3b) : 189- vue orale, niveau 11,0 ; 190- profil cavalier oral du précédent ; 191- vue
aborale, niveau -5,0 ; 192- profil cavalier aboral, niveau +5,0 ; 193-
profil, niveau -5,0 ; 194- profil cavalier oral, niveau 11,0.
Vues 195-197, gilianelle Pocillella cylindrata (E3c) : 195- vue cavalière
orale, niveau +5,0 ; 196- profil d'un autre spécimen, niveau 11,0 ; 197- vue cavalière aborale du
même.
Vues 198-200, 3 gilianelles Pocillella cochleare (E3d) du niveau 25,0 : 198- vue orale ; 199- vue aborale ; 200-
profil.
Vues 201-203, gilianelle Coraliella simplex (M1) du niveau 79,8 : 201- vue cavalière aborale ; 202- vue cavalière orale du précédent ; 203-
profil.
Plate 12: Gilianelles enlarged 240 times; profile views with the aperture upward.
Images 187-188, gilianelle Pocillella grandicula (E3a) of level 11.0: 187- oblique aboral profile; 188- aboral view of the same.
Images 189-194, gilianelle Pocillella minuta (E3b): 189- oral view, level 11.0; 190- oblique oral profile of the same; 191- aboral view, level -5.0; 192- oblique aboral profile, level +5.0; 193- profile, level -5.0; 194- oblique oral profile, level 11.0.
Images 195-197, gilianelle Pocillella cylindrata (E3c): 195- oblique oral view, level +5.0; 196- profile, level 11.0; 197- oblique aboral view of the same.
Images 198-200, 3 gilianelles Pocillella cochleare (E3d) of level 25.0: 198- oral view; 199- aboral view; 200- profile.
Images 201-203, gilianelle Coraliella simplex (M1) of level 79.8: 201- oblique aboral view; 202- oblique oral view of the same; 203- oblique oral profile of another specimen.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 13 :
Gilianelles vues au microscope électronique, grossies 240 fois ; vues de profil avec l'orifice vers le haut.
Vue 204, gilianelle Coraliella simplex (M1) : vue aborale.
Vues 205-208, gilianelle Coraliella cantharella forme a (M2a) : 205-
profil, niveau 57,9 ; 206- vue orale, niveau 50,0 ; 207- vue cavalière
orale, niveau 50,0 ; 208- vue aborale, niveau 54,9.
Vues 209-211, gilianelle Coraliella cantharella forme b (M2b) : 209- vue cavalière
orale, niveau 47,2 ; 210- profil oral, niveau 54,9 ; 211- vue cavalière orale du
même.
Vues 212-214, gilianelle Coraliella brachialis (M3) du niveau 69,0 : 212- vue orale ; 213- profil cavalier aboral ; 214- vue aborale du précédent.
Plate 13: Gilianelles enlarged 240 times; profile views with the aperture upward.
Image 204, gilianelle Coraliella simplex (M1): aboral view.
Images 205-208, gilianelle Coraliella cantharella form a (M2a): 205- profile, level 57.9; 206- oral view, level 50.0; 207- oblique oral view, level 50.0; 208- aboral view, level 54.9.
Images 209-211, gilianelle Coraliella cantharella form b (M2b): 209- oblique oral view, level 47.2; 210- oblique oral profile, level 54.9; 211- oblique oral view of the same.
Images 212-214, gilianelle Coraliella brachialis (M3) du level 69.0: 212- oral view; 213- oblique aboral profile; 214- aboral view of the
same.
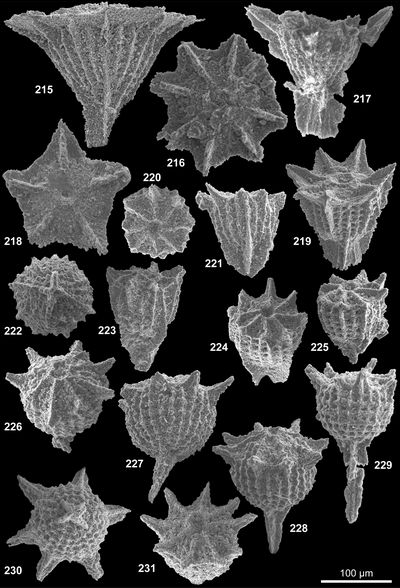
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 14 : Gilianelles vues au microscope électronique, grossies 240 fois ; vues de profil avec l'orifice vers le haut.
Vues 215, gilianelle Coraliella cantharella form b (M2b) : profil, niveau 50,0.
Vues 216- 219, gilianelle Coraliella brachialis (M3) : 216- vue
orale, spécimen rare à 8 bras ; 217- profil ; 218- vue orale, spécimen rare à 5 bras ; 219- profil cavalier oral du spécimen 212
(forme commune à 7 bras).
Vues 220-225, gilianelle Obbella alatopedata (E2b) du niveau 11,0 : 220- vue orale ; 221- profil ; 222- vue aborale ; 223- profil ; 224- profil cavalier oral ; 225- profil cavalier oral, même spécimen que 220.
Vues 226-231, gilianelle Obbella caryophyllum (G1) : 226- vue cavalière
orale, niveau 57,9 ; 227- profil, niveau 57,9 ; 228- profil cavalier oral, niveau 54,9 ; 229-
profil, spécimen à long pied, niveau 69,0 ; 230- vue aborale, niveau 79,8 ; 231- vue cavalière
orale, niveau 50,0.
Plate 14: Gilianelles enlarged 240 times; profile views with the aperture upward.
Images 215, gilianelle Coraliella cantharella form b (M2b): profile, level 50.0.
Images 216- 219, gilianelle Coraliella brachialis (M3): 216- oral view, unusual specimen with 8 arms; 217- profile; 218- oral view, unusual specimen with 5 arms; 219- oblique oral profile of specimen 212 (usual form with 7 arms).
Images 220-225, gilianelle Obbella alatopedata (E2b) of level 11.0: 220- oral view; 221- profile; 222- aboral view; 223- profile; 224- oblique oral profile; 225- oblique oral profile, same specimen as 220.
Images 226-231, gilianelle Obbella caryophyllum (G1): 226- oblique oral view, level 57.9; 227- profile, level 57.9; 228- oblique oral profile, level 54.9; 229- profile, level 69.0; 230- aboral view, level 79.8; 231- oblique oral view, level 50.0.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 15 :
Gilianelles et autres microproblematica vus au microscope électronique, grossis 240 fois en haut, 160 fois en bas
(sauf vue 245) ; vues de profil avec l'orifice vers le haut.
Vues 232-233, gilianelle Obbella caryophyllum (G1) : 232- profil cavalier oral, niveau 11,0 ; 233- profil cavalier oral, niveau 57,9.
Vue 234, microproblematica Tubella hunzikeri (L1), vue cavalière
orale, niveau 11,0.
Vues 235-239, microproblematica Lucernellus aubouini (F1) : 235- profil cavalier oral, niveau 54,9 ; 236- vue cavalière
orale, niveau 47,2 ; 237- vue cavalière aborale, niveau 47,2 ; 238- test ouvert vu de
profil, niveau 54,9 ; 239- vue aborale, niveau 47,2.
Vues 240-246, microproblematica Tubella hunzikeri (L1) : 240- vue
orale, niveau 50,0 ; 241- vue cavalière orale du même ; 242- vue
orale, niveau 11,0 ; 243- profil, niveau +5,0 ; 244- profil cavalier oral, niveau -5,0 ; 245- grossissement du même ; 246- vue cavalière orale du spécimen de la vue 242.
Vues 247-253, microproblematica Vasculum laevis (E1) : 247- vue cavalière
orale, niveau 50,0 ; 248- profil du même ; 249- vue aborale, niveau 11,0 ; 250- vue
orale, niveau 11,0 ; 251- profil cavalier oral, niveau 11,0 ; 252- vue
orale, niveau 11,0 ; 253- profil cavalier oral du même.
Vues 254-256, microproblematica Cimicellus nudatus (K1), niveau 79,8 : 254- vue cavalière
orale, test présent ; 255- vue orale, test partiellement présent ; 256-
profil, test présent sur les flancs.
Plate 15: Gilianelles and other microproblematica. x 240 above horizontal division, below it
x 160 (except view 245); profile views with the aperture upward.
Images 232-233, gilianelle Obbella caryophyllum (G1): 232- oblique oral profile, level 11.0; 233- oblique oral profile, level 57.9.
Image 234, microproblematica Tubella hunzikeri (L1), oblique oral view, level 11.0.
Images 235-239, microproblematica Lucernellus aubouini (F1): 235- oblique oral profile, level 54.9; 236- oblique oral view, level 47.2; 237- oblique aboral view, level 47.2; 238- profile of an open test, level 54.9; 239- aboral view, level 47.2.
Images 240-246, microproblematica Tubella hunzikeri (L1): 240- oral view, level 50.0; 241- oblique oral view of the same; 242- oral view, level 11.0; 243- profile, level +5.0; 244- oblique oral profile, level -5.0; 245- enlargement of the same; 246- oblique oral profile of specimen 242.
Images 247-253, microproblematica Vasculum laevis (E1): 247- oblique oral view, level 50.0; 248- profile of the same; 249- aboral view, level 11.0; 250- oral view, level 11.0; 251- oblique oral profile, level 11.0; 252- oral view, level 11.0; 253- oblique oral profile of the same.
Images 254-256, microproblematica Cimicellus nudatus (K1), level 79.8: 254- oblique oral view, test of specimen complete; 255- oral view, test partially removed; 256- profile, test removed on the oral face and aboral end.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 16 :
Autres microproblematica vus au microscope électronique, grossis 160 fois sauf détails plus grossis ; vues de profil avec l'orifice vers le haut.
Vues 257-259 microproblematica Cimicellus nudatus (K1), niveau 79,8 : 257- vue
aborale, test présent en partie ; 258- vue cavalière orale, test présent ; 259- vue cavalière
orale, moule interne glauconitisé.
Vues 260-261, microproblematica Cimicellus clavellus (K2a) du niveau +5,0 : 260-
profil, moule interne ; 261- vue cavalière orale du même.
Vues 262-266, microproblematica Piperella dercourti (D1a) : 262- vue
orale, niveau 89,0 ; 263- profil cavalier oral du même ; 264- profil, niveau 79,8 ; 265- vue cavalière
aborale, niveau 79,8 ; 266- vue aborale d'un spécimen hérissé, niveau 79,8.
Vues 267-269, microproblematica Piperella dercourti rugatissima (D1b) : 267- vue
orale, niveau 79,8 ; 268- profil, niveau 79,8 ; 269- vue orale, niveau 54,9.
Vues 270-272, microproblematica Piperella camelopardalis (D3) du niveau 79,8
(holotype de l'espèce) : 270- profil ; 271- profil cavalier oral du même ; 272- nannostructure du
même.
Vues 273-277, microproblematica Globulella opprimata (A5) : 273- vue
orale, spécimen avec tout le test, niveau 109,7 ; 274- profil, test sans sa couche
externe, niveau 109,7 ; 275- nannostructure externe de la couche interne du test, même spécimen ; 276- profil cavalier, niveau B-29 ; 277-
profil, niveau B-29.
Vues 278-280, microproblematica Pilella reticulata (N4): 278- profil de
l'holotype, niveau 79.8; 279- nannostructure du même; 280- vue orale, niveau 62.0.
Plate 16: Other microproblematica; profile views with the aperture upward. x
160 times except 272, 275 and 279 enlarged as indicated.
Images 257-259 microproblematica Cimicellus nudatus (K1), level 79.8: 257- aboral view, test damaged; 258- oblique oral view, test present; 259- oblique oral view, glauconitised internal cast.
Images 260-261, microproblematica Cimicellus clavellus (K2a) of level +5.0: 260- profile, internal cast; 261- oblique oral view of the same.
Images 262-266, microproblematica Piperella dercourti (D1a): 262- oral view, level 89.0; 263- oblique oral profile of the same; 264- profile, level 79.8; 265- oblique aboral view, level 79.8; 266- aboral view of a spiky specimen, level 79.8.
Images 267-269, microproblematica Piperella dercourti rugatissima (D1b): 267- oral view, level 79.8; 268- profile, level 79.8; 269- oral view, level 54.9.
Images 270-272, microproblematica Piperella camelopardalis (D3) of level 79.8
(holotype of the species): 270- profile; 271- oblique oral profile of the same; 272- nannostructure of the same.
Images 273-277, microproblematica Globulella opprimata (A5): 273- oral view, specimen its whole test, level 109.7; 274- profile, test without the external layer, level 109,7; 275- external nannostructure of the internal layer of the test, same specimen; 276- oblique profile, level B-29; 277- profile, level B-29.
Images 278-280, microproblematica Pilella reticulata (N4): 278- profile of the
holotype, level 79.8; 279- nannostructure of the same; 280- oral view, level 62.0.
Les spécimens figurés dans les planches réalisées à partir des vues prises au microscope électronique à balayage sont collés et recouverts d'un film conducteur sur douze plots numérotés III à XIV. Chaque plot, comportant 60 à 70 spécimens numérotés, a été photographié et un tirage annexé aux plots préservés dans la collection des fossiles de Tercis actuellement (Avril-2007) déposée à l'Université Pierre & Marie Curie (adresse de l'auteur). La collection sera transférée au Musée du site géologique de Tercis dans les environs de Dax (Landes) dès l'achèvement des travaux et consultable sur place. La liste des figurés, ci-après, donne la localisation sur les plots et le numéro d'ordre des spécimens, éventuellement le niveau de prélèvement. Toutes les vues sont archivées individuellement sous forme électronique.
Planche 1
N2b : 1- IV44 ; 2- IV39 ; 3- IV40 ; 4- IV39 ; 5- VII11 ;
N1b : 6- VII14 ; 7- XI61 ; 8- V05 ; 9 & 10- V04 ;
N1a : 11 à 13- IX-17 ;
A7 : 14- XII03 ; 15- VII17 ; 16- XII09 ; 17- VI40 ; 18- VI65 ;
Planche 2
A7 : 19- VIII21 ;
A8 : 20 & 21- XIII-23 (79,8) ; 22- XI-10 (79,8) ;
A1a : 23- VI47 ; 24 & 29- VI05 ; 25- VI04 ; 26- IV30 ; 27- V52 ; 28- VII58 ;
A4b : 30- IX19 ; 31- IV34 ; 32- IX13 ; 33- XI60 (54,7) ;
Planche 3
A4b : 34- VI56 ; 35- III63 ;
A4a : 36 & 37- XIII08 ; 38- V59 ; 39- VIII23 ; 40- VI49 ; 41- XIV60 ; 42- V27 ; 43- XII27 ;
A4b' : 44 & 45- VII-29 (69,0) ; 46- X-47 (79,8) ; 47 & 48- XI-58 (79,8) ;
Planche 4
A6 : 49- VI02 (172) ; 50- VI01 (172) ; 51- VI10 (172) ; 52- VI51 (172) ;
A1b 53- IX-39 (89,0) ; 54 & 55- IX16 (79,8) ; 56 & 57- IX37 (89,0) ; 58- VI06 (172) ; 59- V16 (11,0) ;
A3 : 60- VIII40 ; 61 & 62 VII18 ; 63- XII22 ; 64- XIV61 ;
Planche 5
A2a : 65- IV32b ; 66- VIII54 ; 67- V33 ; 68- VI57 ; 69- VII23 ; 70- VII59 ;
A2b : 71- VIII33 ; 72 & 73 VIII31 ; 74- VII61 ; 75- VIII28 ; 76- IX47 ; 77- XII65 ;
L2 : 78-X-11 (50,0) ; 79- X20 (62,5) ; 80- VII57 (57,9) ;
Planche 6
L2 : 81 & 82- XII63 (57,9) ;
L3 : 83 & 84- VII21 (holotype, 79,8) ;
B2a : 85- IX50 ; 86- III19b ; 87- IX01 ; 88- IX09 ; 89- IX16 ; 90- VIII01 ; 91- VIII02 ;
B2b : 92- XII17 ; 93- XIV54 ; 94- XII36 ; 95- VI41 ; 96- VIII38 ;
Planche 7
B3a : 97- III18 (50,0) ; 98 et 98b- X65 (79,8) ; 99- VIII58 (54,9) ;
B4 : 100 & 101- VII53 (holotype, 172) ; 102- IX39 (50,0) ;
B3d : 103- X16 (50,0) ; 104 & 105- XIII56 (50,0) ;
B3d' : 106 & 107 XI23 (79,8) ;
Planche 8
B3d' : 108- XI22 (79,8) ;
B1a : 109- V41 ; 110- V46 ; 111 & 112- XII56 ; 113- V35 ;
C2 : 114- VI46 ; 115 & 118- IX38 ; 116 & 117- VIII52 ; 119- IV50 ; 120- VIII43 ;
C2b : 121- XI62 (11,0) ; 122- XI64 (11,0) ;
Planche 9
C2b : 123- V30 (11,0) ; 124- XI65 (11,0) ; 125- XI64 ;
C1b : 126 & 127- IV41 ; 128- IX46b ; 129- IX12 ; 130- X12b ;
C1a : 131- VI56 ; 132- VI51 ; 133- V58 ; 134 & 135- XIV55 ; 136- VIII50 (p.p.) ;
J1 : 137- VIII41 ; 138- V22 ; 139- XIII09 ; 140 & 141- V61 ;
Planche 10
J2a : 142 & 143- XII10 ; 144 & 145- XII20 ; 146 & 147- VI43 ; 148- VI44 ; 149- XII05 ;
J2b : 150- XII21 ; 151 à 154- VIII49 ;
G0 : 155- XII35 ;156 & 157- XII18 ; 158 & 159- V17 ;
G0b : 160- X-07 (50,0) ; 161 à 163- XII-60 (54,9) ;
H2 : 164- IV43 ;
Planche 11
H2 : 165 & 166- VIII57 ; 167 & 168- IX53 ; 169- V49 ; 170 à 172- XII58 ; 173 & 174- IX43 ;
H5 : 175- IX48 ; 176 & 178- IX41 ; 177- IX51 ; 179- XII-40 ; 180 & 181- XII41 ; 182 & 183- XII49 ;
E3a : 184 & 185- VI62 ; 186- XII22 ;
Planche 12
E3a : 187 & 188- XII08 ;
E3b :189 & 190- XII15 ; 191- XIV56 (-5,0) ; 192- V50 ; 193- VIII19 ; 194- V29 ;
E3c : 195- XIII05 (+5,0) ; 196 & 197- XII19 (11,0) ;
E3d : 198- XIII36 (25,0) ; 199- IX04 (25,0) ; 200- IX06 (25,0) ;
M1 : 201 & 202- X5 (79,8) ; 203- X40 (79,8) ;
Planche 13
M1 : 204- V43 (79,8) ;
M2a : 205- VI52 (57,9) ; 206- IX62 (50,0) ; 207- X13 (50,0) ; 208- IV42 (54,9) ;
M2b : 209- X15 (50,0) ; 210 & 211- IV62 (54,9) ;
M3 : 212- XII55 ; 213 & 214- VI25 ;
Planche 14
M2b : 215- X-14 (50,0) ;
M3 : 216- VI24 ; 217- VI29 ; 218- VI28 ; 219- XII55 ;
E2b : 220- XII13 ; 221- XIII-44 ; 222- V20 ; 223- V23 ; 224- XII14 ; 225- XII13 ;
G1 : 226- XII62 ; 227- XII64 ; 228- IV45 ; 229- VI17 ; 230- XI24 (79,8) ; 231- IX42 ;
Planche 15
G1 : 232- V25 ; 233- XII64 ;
L1 : 234-V-3 (11,0) ;
F1 : 235- XII57 (54,9) ; 236- II49 (47,2) ; 237- II50 (47,2) ; 238- IX30 (54,9) ; 239- II48 (47,2) ;
L1 : 240 & 241- IX44 (50,0) ; 242 & 246- V09 (11,0) ; 243- V56 (+5,0) ; 244 & 245- VIII16 (-5,0) ;
E1 : 247 & 248- IX65 (50,0) ; 249- VII65 (11,0) ; 250- V08 (11,0) ; 251- XIII46 ; 252 & 253- VII12 (11,0) ;
K1 : 254- X46 (79,8) ; 255- VII22 (79,8) ; 256- IV60 (79,8) ;
Planche 16
K1 : 257- VII19 (79,8) ; 258- V36 (79,8) ; 259- VII24 (79,8) ;
K2a : 260 & 261- VI67 (+5,0) ;
D1a : 262 & 263- IV59 (89,0) ; 264- X48 (79,8) ; 265- XI25 (79,8) ; 266- IV61 (79,8) ;
D1b : 267 & 268- IX35 (79,8) ; 269- IV33 (54,9) ;
D3 : 270 à 272- XI02 (holotype, 79,8) ;
A5 : 273- IV23 (109,7) ; 274 & 275- IV-24 (109,7) ; 276 & 277- VII04 (B-29) ;
N4 : 278 & 279- XI03 (holotype, 79,8) ; 280- X30 (62,0).
Hormis les 4 holotypes désignés d'après des spécimens illustrés au microscope électronique (cf. ci-dessus : L3, B4, D3, N4), tous les holotypes, paratypes et quelques topotypes d'accompagnement des espèces nouvelles de ce travail ont été photographiés au microscope optique (deux planches jointes à cette annexe). Holotypes et paratypes ont été colorés pour être repérés plus aisément dans les cellules de préservation.
Comme noté plus haut dans le texte, les holotypes ont été, aussi systématiquement que possible, accompagnés de spécimens de morphologie identique provenant des mêmes niveaux (topotypes) au cas oů la fragilité des tests serait mise à l'épreuve par le temps.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche annexe 1 : Vues des gilianelles types au microscope optique.
L'échelle commune (vignette 5) est de 200 µm ; l'holotype est systématiquement disposé en haut à gauche de chaque vignette. 1- Tercensella globosa (N2b), holotype en vue cavalière orale et 4 topotypes ; 2- Tercensella doliolum (N1b), holotype de profil et 4 topotypes ; 3- Tercensella alveare (N1a), holotype en vue de profil et 3 topotypes ; 4- Azymella cannabinata (A7), holotype en face orale et 1 topotypes ; 5- Azymella offa (A8), holotype et 2 topotypes ; 6- Numismella tarbellica (A1a), holotype en face aborale et 3 topotypes ; 7- Scutellella crassa (A1b), holotype en vue aborale et 3 topotypes en profil, face orale et face aborale ; 8- Scutellella circularis (A2a), holotype en face aborale et 1 topotype ; 9- Scutellella longivelifera (A2b), holotype en face aborale et 2 topotypes ; 10- Scutellella pulchrevelifera (A3), holotype en face orale ; 11- Aturella altodepressa (A4a), holotype en face orale et 3 topotypes ; 12- Aturella aequilatera (A4b), holotype en face aborale, orientation avant vers le haut des 3 spécimens ; 13- Aturella angulata (A4b'), holotype en vue orale et topotype en vue aborale ; 14- Aturella edentula (A6), holotype en vue orale et topotype en vue aborale ; 15- Corniculum sinuosum (L2), holotype en vues aborale et de profil et 3 topotypes ; 16- Corniculum clausum (L3), topotype en vues aborale et cavalière orale et 2ème topotype ; 17- Gilianella tenuibrachialis (B1a), holotype en face orale et 2 topotypes ; 18- Gilianella stellata pulvinata (B2b), holotype en face orale, paratype dessous, à gauche en vue aborale et 2 topotypes ; 19- Gilianella major (B3a), holotype en vue orale et 1 topotype ; 20- Gilianella terbrachialis (B3d), holotype en face orale et 1 topotype mal dégagé ; 21- Gilianella terbrachialis mutata (B3d'), holotype en face aborale, (un microfossile est collé sur la face orale) et 2 topotypes ; 22- Caccabella spinocincta (C2), holotype en vue cavalière aborale et 4 topotypes ; 23- Caccabella spinocincta ventriosa (C2b), holotype en vue cavalière orale et 3 topotypes ; 24- Orculiella niduseumenei (C1b), holotype de profil et 4 topotypes ; 25- Orculiella niduseumenei strangulata (C1a), holotype en face orale et 3 topotypes.
Appendix plate 1: Images of type specimens of gilianelles through an optical microscope.
The common scale is 200 µm (vignette 5); the holotype is at the upper left of each vignette. 1- Tercensella globosa (N2b), oblique oral view of the
holotype and 4 topotypes ; 2- Tercensella doliolum (N1b), profile of the
holotype and 4 topotypes ; 3- Tercensella alveare (N1a), profile of the
holotype and 3 topotypes ; 4- Azymella cannabinata (A7), oral view of the
holotype and 1 topotypes ; 5- Azymella
offa (A8), holotype and 2 topotypes ; 6-
Numismella tarbellica (A1a), aboral view of the holotype and 3 topotypes ; 7-
Scutellella crassa (A1b), aboral view of the holotype and 3 topotypes with profile, oral, and aborale views; 8-
Scutellella circularis (A2a), aboral view of the holotype and 1 topotype ; 9-
Scutellella longivelifera (A2b), aboral view of the holotype and 2 topotypes; 10-
Scutellella pulchrevelifera (A3), oral view of the holotype and 1 topotype; 11-
Aturella altodepressa (A4a), oral view of the holotype and 3 topotypes ; 12- Aturella aequilatera (A4b), aboral view of the
holotype and 2 topotypes all of them oriented with the front upward; 13- Aturella angulata (A4b'), oral view of the
holotype and aboral view of a
topotype; 14- Aturella edentula (A6), oral view of the holotype and aboral view of a
topotype; 15- Corniculum sinuosum (L2), aboral and profile views of the holotype and 3
topotypes; 16- Corniculum clausum (L3), aboral and oblique oral views of a and second
topotype; 17- Gilianella tenuibrachialis (B1a), oral view of the holotype and 2
topotypes; 18- Gilianella stellata pulvinata (B2b), oral view of the holotype, aboral view of the paratype below, and 2 topotypes; 19- Gilianella major (B3a), oral view of the
holotype and 1
topotype; 20- Gilianella terbrachialis (B3d), oral view of the holotype and 1 topotype with matrix; 21- Gilianella terbrachialis mutata (B3d'), aboral view of the
holotype and 2 topotypes ; 22- Caccabella spinocincta (C2), oblique aboral view of the
holotype and 4 topotypes ; 23- Caccabella spinocincta ventriosa (C2b), oblique oral view of the
holotype and 3 topotypes ; 24- Orculiella niduseumenei (C1b), profile of the
holotype and 4 topotypes ; 25- Orculiella niduseumenei strangulata (C1a), oral view of the
holotype and 3
topotypes.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche annexe 2 : Vues des gilianelles et autres microproblematica types au microscope optique.
L'échelle commune (vignette 46) est de 200 µm ; l'holotype est disposé en haut de chaque vignette. 26- Pennigerella palmata alta (J1), holotype en profil oral, paratype en haut à droite en vue aboral et 3 topotypes ; 27- Pennigerella palmata humilis (J2a), holotype en profil oral et 3 topotypes ; 28- Pennigerella castulata (J2b), holotype de profil sur les deux vues du haut (coloré et non coloré) et 2 topotypes ; 29- Corbella vitilis (G0), holotype en profil oral et 3 topotypes ; 30- Corbella vitilis biremis (G0b), 3 vues de l'holotype, en haut, face aborale mise au point sur les pieds, au milieu, même disposition mise au point sur les bras, en bas, profil oral ; 31- Aquilegiella varia (H2), holotype de profil et 3 topotypes ; 32- Aquilegiella ericius (H5), holotype en profil oral et 4 topotypes ; 33- Pocillella grandicula (E3a), holotype en profil oral et 2 topotypes ; 34- Pocillella minuta (E3b), holotype en profil aboral comme le paratype en haut à droite et 3 topotypes ; 35- Pocillella cylindrata (E3c), holotype en vues aborale et de profil et 2 topotypes ; 36- Pocillella cochleare (E3d), holotype en profil oral et 1 topotype ; 37- Coraliella simplex (M1), holotype en vue aborale et 3 topotypes ; 38- Coraliella cantharella forme a (M2a), holotype en vue aborale et 1 topotype ; 39- Coraliella cantharella forme b (M2b), holotype en vue cavalière orale et 2 topotypes ; 40- Coraliella brachialis (M3), holotype en vue aborale et 3 topotypes ; 41- Obbella alatopedata (E2b), holotype de profil et 5 topotypes ; 42- Obbella caryophyllum (G1), holotype en vue aborale et 2 topotypes ; 43- Globulella opprimata (A5), holotype en vue orale et 3 topotypes ; 44- Piperella dercourti (D1a), holotype en profil oral ; topotype en face aborale au centre, topotype "hérissé" en profil oral en bas ; 45- Piperella dercourti rugatissima (D1b), holotype en vue orale et 2 topotypes ; 46- Piperella camelopardalis (D3), topotype de profil ; 47- Vasculum laevis (E1), holotype en profil oral et 4 topotypes ; 48- Lucernellus aubouini (F1), holotype en profil oral et 2 topotypes ; 49- Cimicellus nudatus (K1), holotype en vues de profil et aborale et 2 topotypes ; 50- Cimicellus clavellus (K2a), holotype de profil et 3 topotypes ; 51- Tubella hunzikeri (L1), holotype en vues aborale et de profil et 3 topotypes.
Appendix plate 2: Images of type specimens of gilianelles and other microproblematica through an optical microscope.
The common scale is 200 µm (vignette 46); the holotype is at the upper left of each vignette. 26- Pennigerella palmata alta (J1), oral profile of the holotype, aboral view of the paratype (top right) and 3 topotypes ; 27- Pennigerella palmata humilis (J2a), oral profile of the holotype and 3 topotypes ; 28- Pennigerella castulata (J2b), two profiles of the holotype (top views after and before colouring) and 2 topotypes ; 29- Corbella vitilis (G0), oral profile of the holotype and 3 topotypes ; 30- Corbella vitilis biremis (G0b), 3 views of the holotype, aboral, aboral, and oral profile from top to bottom; 31- Aquilegiella varia (H2), profile of the holotype and 3 topotypes ; 32- Aquilegiella ericius (H5), oral profile of the holotype and 4 topotypes ; 33- Pocillella grandicula (E3a), oral profile of the holotype and 2 topotypes; 34- Pocillella minuta (E3b), aboral profile of the holotype and of the paratype (top right) and 3 topotypes; 35- Pocillella cylindrata (E3c), aboral and profile views of the holotype and 2 topotypes; 36- Pocillella cochleare (E3d), oral profile of the holotype and 1 topotype; 37- Coraliella simplex (M1), aboral view of the holotype and 3 topotypes ; 38- Coraliella cantharella form a (M2a), aboral view of the holotype and 1 topotype ; 39- Coraliella cantharella form b (M2b), oblique oral view of the holotype and 2 topotypes ; 40- Coraliella brachialis (M3), aboral view of the holotype and 3 topotypes ; 41- Obbella alatopedata (E2b), profile of the holotype and 5 topotypes ; 42- Obbella caryophyllum (G1), aboral view of the holotype and 2 topotypes ; 43- Globulella opprimata (A5), oral view of the holotype and 3 topotypes ; 44- Piperella dercourti (D1a), oral profile of the holotype; aboral view of a topotype -centre-, oral profile of a spicky topotype -bottom-; 45- Piperella dercourti rugatissima (D1b), oral view of the holotype and 2 topotypes ; 46- Piperella camelopardalis (D3), profile of a topotype; 47- Vasculum laevis (E1), oral profile of the holotype and 4 topotypes ; 48- Lucernellus aubouini (F1), oral profile of the holotype and 2 topotypes ; 49- Cimicellus nudatus (K1), profile and aboral views of the holotype and 2 topotypes ; 50- Cimicellus clavellus (K2a), profile of the holotype and 3 topotypes ; 51- Tubella hunzikeri (L1), aboral and profile views of the holotype and 3 topotypes.
Numismella tarbellica : La médaille tarbelle, code A1a
Scutellella crassa : La scutellelle épaisse, code A1b
Scutellella circularis : La scutellelle circulaire, code A2a
Scutellella longivelifera : La scutellelle à longue voile, code A2b
Scutellella pulchrevelifera : La scutellelle joliment voilée, code A3
Aturella altodepressa : L'aturelle haute-déprimée, code A4a
Aturella aequilatera : L'aturelle à faces égales, code A4b
Aturella angulata : L'aturelle anguleuse, code A4b'
Aturella edentula : L'aturelle édentée, code A6
Azymella cannabinata : L'azymelle en canevas, code A7
Azymella offa : L'azymelle miche, code A8
Gilianella tenuibrachialis : La gilianelle à bras menus, code B1a
Gilianella stellata : La gilianelle en étoile, code B2a
Gilianella stellata pulvinata : La gilianelle en oreiller, code B2b
Gilianella major : La grande gilianelle, code B3a
Gilianella terbrachialis : La gilianelle à 3 bras, code B3d
Gilianella terbrachialis mutata : La gilianelle à 3 bras modifiée, code B3d'
Gilianella major heliotropium : La gilianelle tournesol, code B4
Orculiella niduseumenei : La petite jarre en nid d'eumène, code C1b
Orculiella niduseumenei strangulata : La petie jarre étranglée, code C1a
Caccabella spinocincta : La caccabelle ceinte d'épines, code C2
Caccabella spinocincta ventriosa : La caccabelle pansue, code C2b
Obbella alatopedata : L'obbelle à pied ailé, code E2b
Pocillella grandicula : La petite tasse assez grande, code E3a
Pocillella minuta : La tasse minuscule, code E3b
Pocillella cylindrata : La petite tasse cylindrique, code E3c
Pocillella cochleare : Le petit coquetier, code E3d
Corbella vitilis : La corbeille tressée, code G0
Corbella vitilis biremis : La corbeille birème, code G0b
Obbella caryophyllum : L'obbelle clou de girofle, code G1
Aquilegiella varia : La petite ancolie variable, code H2
Aquilegiella ericius : Le petit cheval de frise, code H5
Pennigerella palmata alta : L'ailée palmée haute, code J1
Pennigerella palmata humilis : L'ailée palmée basse, code J2a
Pennigerella castulata : L'ailée à jupon, code J2b
Corniculum sinuosum : Le petit entonnoir sinueux, code L2
Corniculum clausum : Le petit entonnoir fermé, code L3
Coraliella simplex : Le petit corail simple, code M1
Coraliella cantharella a : Le petit corail chanterelle var a, code M2a
Coraliella cantharella b : Le petit corail chanterelle var b, code M2b
Coraliella brachialis : Le petit corail à bras, code M3
Tercensella alveare : La tercisiennelle ruche, code N1a
Tercensella doliolum : La tercisiennelle tonnelet, code N1b
Tercensella globosa : La tercisiennelle sphérique, code N2b
Lucernellus aubouini : Le lucernel d', code F1
Tubella hunzikeri : La tubelle de , code L1
Vasculum laevis : Le petit vase lisse, code E1
Cimicellus nudatus : La petite punaise dépouillée, code K1
Cimicellus clavellus : La petite punaise en clou, code K2a
Piperella dercourti : La piperelle de , code D1a
Piperella dercourti rugatissima : La piperelle très ridée, code D1b
Piperella camelopardalis : La piperelle girafe, code D3
Pilella reticulata : La petite pelote réticulée, code N4
Globulella opprimata : La boulette aplatie, code A5