![]()
Sommaire
[1. Introduction] [2. Contexte géologique et stratigraphique]
[3. Les associations d'ostracodes] [4. Remarques paléoécologiques]
[5. Implications paléobiogéographiques] [6. Systématique]
[Références bibliographiques] et ... [Planches]
Géologie des Systèmes et Réservoirs carbonatés,
E.A., 4234, Université de Provence, Centre Saint-Charles,
Case 67, Place Victor Hugo, 13331 Marseille Cedex 3 (France)
* Auteur correspondant
3 Impasse des Biroulayres, 33610 Cestas (France)
Département de Géologie, EA 4098 LPAT-GEOL, Université des Antilles et de la
Guyane, Campus de Fouillole, 97159 Pointe-à-Pitre Cedex, Guadeloupe (France)
Manuscrit en ligne depuis le 4 Avril 2009
![]()
Des associations d'ostracodes provenant de la région d'Antsiranana, ex Diego-Suarez, dans le N de Madagascar ont été étudiées au niveau systématique. Dans l'intervalle Albien-Turonien moyen, 29 espèces appartenant à 21 genres ont été reconnues, 10 nouvelles espèces, un nouveau genre (Malagasyella) et un nouveau sous-genre (Hemiglenocythere) ont été créés. L'Albien ainsi que le Cénomanien inférieur se caractérisent par des associations bien différenciées. D'un point de vue paléoécologique, la succession des faunes d'ostracodes montre une baisse progressive du niveau de la mer d'un milieu de bassin-talus à l'Albien à un environnement de plate-forme externe au Turonien. Pendant cette période les faunes d'ostracodes de Madagascar montrent de fortes affinités avec celles de l'Inde (Rajasthan) ce qui mène à proposer l'existence de province faunistique indo-malgache.
Ostracodes ; Madagascar ; Albien ; Cénomanien ; Turonien ; systématique ; paléoécologie ; paléobiogéographie.
J.-F., J.-P. & A. (2009).- Les ostracodes de l'Albien-Turonien moyen de la région d'Antsiranana (Nord Madagascar) : systématique, paléoécologie et paléobiogéographie.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2009/01 (CG2009_A01)
Albian-Middle Turonian ostracodes from the Antsiranana region (northern Madagascar): systematics, palaeoecology and palaeobiogeography.- Ostracode associations from the Antsiranana region, formerly Diego-Suarez, in northern Madagascar have been studied at a taxonomic level. In the Albian-middle Turonian interval, 29 species belonging to 21 genera have been recognized, 10 new species, one new genus (Malagasyella) and a new sub-genus (Hemiglenocythere) have been erected. From a palaeoecological stand point, the succession of ostracode faunas shows a progressive shallowing trend, from basin-slope during Albian times to outer shelf during the Turonian. During these periods, the ostracode faunas of Madagascar show strong affinities with those of India (Rajasthan). This leads to a proposal for the existence of an indo-malagasy ostracode faunal province.
Ostracodes; Madagascar; Albian; Cenomanian; Turonian; systematics; palaeoecology; palaeobiogeography.
Tsy misy mafy tsy laitra ny zoto
Avec du courage on vient à bout de tout
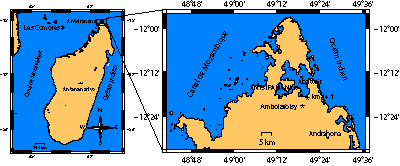
La ville d'Antsiranana (ex Diego-Suarez) est située à l'extrême N de Madagascar. Sa rade qui s'ouvre sur l'Océan Indien entre le cap Tanifotsy au N et le Cap Miné au S, développe 150 km de côte, constituée essentiellement d'affleurements d'âge crétacé, coiffés d'une couverture éocène sur les reliefs proches et miocène au sud de la passe.
Les deux sites principaux étudiés dans ce travail se situent, l'un dans la ville même d'Antsiranana dans le quartier dit de
l'Abattoir, l'autre à 40 km environ plus au SE sur le flanc sud de l'Andrahona (ex Mont Raynaud). Ils ont fourni une riche faune d'ammonites (,
1972 ; et alii, 1977),
et ont fait l'objet d'études micropaléontologiques et palynologiques (,
1982 ; et alii, 1982 ; ,
1989 ; & ,
1989). Ils sont complétés par deux prélèvements ponctuels, l'un provenant de la limite orientale de la vallée de la Betahitra
(pk14,1), l'autre de la rivière Ambolafotsy
(Fig. 1 ![]() ).
).
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 1 : Cartes de
localisation.
Figure 1: Location maps.
Les travaux concernant les ostracodes du Crétacé de Madagascar se limitent à quelques publications concernant la base du Crétacé inférieur (, 1963 ; et alii, 1970 ; , 1990) et le Maastrichtien ( et alii, 1974 ; & , 1992b ; et alii, 1996). La seule publication sur les ostracodes de l'Albien et du Cénomanien de la région étudiée dans ce travail est celle de et alii, 1979, qui regroupe des données sur les ammonites, les foraminifères planctoniques et les ostracodes. Une trentaine d'espèces, la plupart laissées en nomenclature ouverte ont été identifiées et figurées.
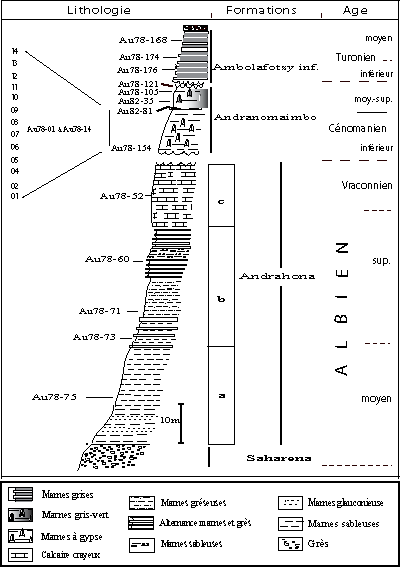
Dans la région étudiée, la série allant de l'Albien moyen au Turonien basal se subdivise en 3 formations
(Fig. 2 ![]() ).
).
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 2 : Coupe synthétique de
l'Albien moyen au Turonien moyen dans la région d'Antsiranana.
Figure 2: Composite log of Middle Albian to Middle Turonian strata in
the Antsiranana area.
La coupe de l'Andrahona (12°29'13"S : 49°26'27"E) repose sur les Grès de la Saharena (Crétacé inférieur continental). Elle montre de bas en haut une série à prédominance marneuse avec intercalations gréseuses. Les prélèvements étudiés appartiennent à la Formation éponyme dite de l'Andrahona, subdivisée en 3 membres a, b et c (, 1989), qui ont livré de riches macrofaunes (ammonites), microfaunes (foraminifères planctoniques) et nannofaunes répartis comme suit, de bas en haut :
Ammonites : Zone à Oxytropidoceras acutocarinatum et Manuaniceras jacobi de l'Albien moyen. (1972) en donne une liste plus complète.
Foraminifères planctoniques : Zone à Ticinella primula, avec présence de Favusella washitensis, Hedbergella planispira, Hedbergella rischi.
Nannofossiles calcaires : Zone à Praediscosphaera columnata ( in et alii, 1982) : membre caractérisé par l'abondance de Watznaueria barnesae et dans une moindre mesure de Zygodiscus diplogrammus, Praediscosphaera columnata, Tegumentum stradneri mais l'absence d' Eiffelithus turriseiffeli.
Palynomorphes : Prédominance du genre Classopolis ainsi que Cicatricosporites cf. australensis, Microcachryidites antarcticus, Callialasporites trilobatus (, 1982).
Dinoflagellés : présence entre autres de Cleistosphaeridium sp., Coronifera oceanica, Hystrichosphaeridium arundum, Xiphophoridium alatum, Prolixosphaeridium conulum ( et alii, 1982).
Ammonites : Zone à Hysteroceras binum avec Hysteroceras binum, Anisoceras armatum, Paraturrilites gressleyi, Phyllopachyceras whiteavesi, Puzosia mayoriana, Scaphites besairiei, Raynaudia madagascariensis.
Foraminifères planctoniques : Zone à Ticinella raynaudi avec Biticinella breggiensis, Favusella washitensis, Hedbergella planispira, ... et dans l'ordre d'apparition, Ticinella praeticinensis, Rotalipora subticinensis, puis Rotalipora ticinensis.
Nannofossiles calcaires : persistance des espèces précédentes auxquelles s'ajoutent Lithrapidites carniolensis, Cretarhabdus crenulatus ( in et alii, 1982).
Dans la partie inférieure :
Ammonites : Zone à Pervinquieria inflata de l'Albien supérieur avec : Pervinquieria inflata, Phylloceras seresitense, Paraturrilites circumtaniatus, Metahamites raynaudi.
Foraminifères planctoniques : Zone à Rotalipora praebalernaensis avec Rotalipora praebalernaensis, Rotalipora ticinensis, Ticinella raynaudi, Biticinella breggiensis, Hedbergella planispira, ...
Nannofossiles calcaires : Permanence de Watznaueria barnesae mais disparition de Tegumentum stradneri. Notons l'absence partout de Eiffelithus turriseiffeli.
Dans la partie supérieure :
Ammonites : Zone à Stoliczkaia dispar de l'Albien terminal (Vraconnien) avec Stoliczkaia dispar, Neophlycticeras madagascariense.
Foraminifères planctoniques : Zone à Rotalipora appenninica avec Rotalipora evoluta, Rotalipora praebalernaensis, Favusella washitensis, Hedbergella planispira, Ticinella madecassiana, Biticinella beggiensis, de l'Albien supérieur.
La coupe de l'abattoir (12°16.35"S : 49°17.58"E) appartient à cette formation dont la base n'affleure pas. Elle montre, de bas en haut, une série homogène de marnes jaunes à cristaux de gypse secondaire, épaisse de 10 m et coiffée par une coulée basaltique cénozoïque en provenance de la Montagne d'Ambre voisine.
Ammonites : Mantelliceras suzanae, Scaphites spp., Mariella spp., Belemnites ultimus, Phylloceras spp., Scaphites spp., ... et alii (1977) datent cette formation du Cénomanien inférieur.
Foraminifères planctoniques : Zone à Rotalipora brotzeni avec, Rotalipora evoluta, Praeglobotruncana delrioensis, Praeglobotruncana stephani. Notons la présence tardive de Planomalina buxtorfi, sous sa forme bicarénée indiquant le Cénomanien basal ( & , 1989).
Nannofossiles calcaires : Zone à Eiffelithus turriseiffeli identifiée par la présence de l'espèce indice et l'absence de Vekshinella angusta. Elle marque l'Albien supérieur et le Cénomanien inférieur.
Les échantillons Au78-01 à Au78-14 proviennent de cette formation dans le quartier dit de l'Abattoir. Cette coupe est complétée par l'échantillon Au 78-154 prélevé au km 14,4 de la route de Ramena, proche du "sondage de Diego" étudié par et alii (1979). Il est situé à trois mètres environ au-dessus de la plaine de la Betahitra, site du dit sondage.
Deux échantillons datés du Cénomanien supérieur en provenance d'autres affleurements proches du secteur d'étude n'ont pas livré de faunes d'ostracodes significatives.
Cette formation montre une alternance de bancs plus ou moins indurés de marnes bleues à gypse secondaire. Les échantillons (12°22'40"S : 49°18'50"E) proviennent de l'Ambolafotsy inférieur, en aval de la chute de la rivière Ambolafotsy. Ils renferment une rare macrofaune mais une riche microfaune.
Ammonites : Rares spécimens pyriteux, dont Desmoceras s.l.
Foraminifères planctoniques : Zone à aprica du Turonien inférieur avec : Whiteinella aprica, Whiteinella baltica, Praeglobotruncana stephani, Marginotruncana (Dicarinella) hagni, Marginotrucana (Dicarinella) marianosi.
Nannofossiles calcaires : Zone à Quadrum gartneri du Turonien inférieur. Watznaueria barnesae est l'espèce dominante. L'association est marquée par la présence de Quadrum gartneri de Ahmullerella octoradiata et l'absence de Eiffelithus eximus.
Palynomorphes : La présence de Classopolis et de Xiphophoridium alatum en l'absence de Litosphaeridium siphoniphorum indiquerait un âge turonien basal ( et alii, 1982). Sont aussi présents Coronifera oceanica, Cyclonephelium vannophorum (in et alii, 1982).
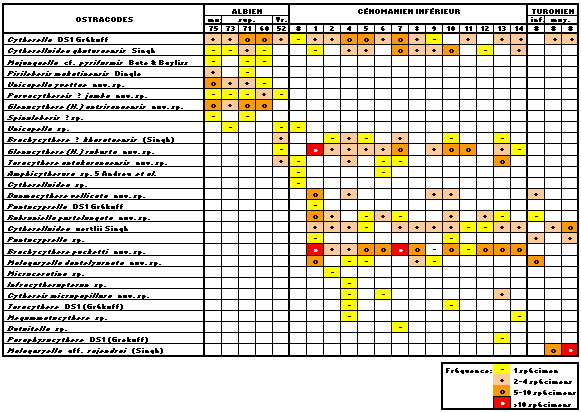
Les associations d'ostracodes se composent de 29 espèces appartenant à 21 genres. Dix nouvelles espèces, un nouveau sous-genre et un nouveau genre sont créés.
Deux ensembles sont bien différenciés : Albien moyen-supérieur et Cénomanien inférieur. Le Vraconnien est une zone de transition avec apparition de formes se développant dans le Cénomanien inférieur
(Tableau ![]() ).
).
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
Tableau :
Tableau de répartition des espèces d'ostracodes.
Table: Stratigraphic ranges of the ostracode species.
Turonien inférieur et moyen sont peu caractérisés avec persistance de quelques éléments cénomaniens.
Douze espèces appartenant à 10 genres.
Espèces caractéristiques : Majungaella cf. pyriformis & , 1969, Unicapella yvettae nov. sp., Glenocythere (Hemiglenocythere) antsirananaensis nov. subgen., nov. sp., Pirileberis makatiniensis , 1984, Paravacythereis ? jamba nov. sp.
Vingt-deux espèces appartenant à 17 genres.
Espèces caractéristiques : Brachycythere pucketti nov. sp., Brachycythere ? kharatarensis (, 1997), Taracythere antakaranaensis nov. sp., Donmacythere vellicata nov. sp., Cytherelloidea oertlii , 1997, Glenocythere (Hemiglenocythere) robusta nov. subgen., nov. sp., Malagasyella dantelyornata nov. gen., nov. sp., Robsoniella postelongata nov.sp., Megommatocythere sp.
Sept espèces appartenant à 5 genres.
Espèce caractéristique : Malagasyella aff. rajendrai (, 1997).
L'ensemble de l'ostracofaune présente les caractéristiques suivantes :
Le rapport foraminifères planctoniques/benthiques, depuis la base de la coupe montre un approfondissement progressif du milieu de sédimentation de l'Albien moyen Formation d'Andrahona) jusqu'au Cénomanien inférieur (Formation d'Andranomaimbo). Par contre, le Turonien basal (Formation d'Ambolafotsy inférieure) montre une tendance plutôt régressive (, 1989).
Des études réalisées en domaine océanique (, 1977) ont montré que l'abondance de Trachyleberididae aveugles mais aussi à architecture externe complexe étaient des indices d'un environnement profond (talus, bassin proximal). Ceci peut être corroboré par la présence simultanée de genres à test lisse et aveugles de milieu de talus et/ou de plate-forme externe ( & , 1978), ce qui serait le cas avec les Cytherellidae et les Pontocyprella bien représentés dans notre matériel (le genre Robsoniella pouvant avoir la même signification). Ces données ont été reprises par (1981) pour le Campanien-Maastrichtien d'Afrique du Sud qui montre des associations génériques proches, mais qu'il interprète en terme de moindre bathymétrie (plate-forme externe).
Dans la région étudiée les associations d'ostracodes se répartissent en deux ensembles bien différenciés. L'Albien est caractérisé par des formes aveugles et une faible diversité (au maximum 9 espèces). Au Cénomanien inférieur (avec prémices dans le Vraconnien), la diversité est nettement plus élevée (20 espèces) ; les genres aveugles persistent mais sont accompagnés de genres à fort tubercule oculaire (minoritaires). Il semble donc que la profondeur de dépôt ait été moindre, en contradiction avec les données fournies par les foraminifères), passant d'un environnement de bassin-talus à un milieu de plate-forme externe. Au Turonien ne subsistent que quelques espèces avec une disparition des formes aveugles mais augmentation du nombre d'individus à fort tubercule oculaire (Malagasyella). On peut donc en déduire une nouvelle baisse de la bathymétrie. En résumé, les ostracodes permettent d'affiner l'évolution des paléomilieux océaniques dans l'intervalle stratigraphique considéré.
Cette étude est en totale contradiction avec l'analyse effectuée par (in et alii, 1979) concernant les ostracodes de l'Albien et du Cénomanien du sondage de Diego-Suarez qui comparait ces associations essentiellement avec des taxons européens. En effet les plus grandes similarités sont avec les régions suivantes :
Arculicythere tumida , 1971, cité et illustré par (= Arculicythere DS1 in et alii, 1979) dans l'Albien du sondage de Diego-Suarez n'a pas été retrouvé dans nos échantillons. Cette espèce, bien caractéristique, a une très vaste répartition géographique sud-gondwanienne : Madagascar, Afrique du Sud, Australie, Inde, Plateau des Falklands et Argentine (, 1974 ; & , 1988, 1992a ; , 1988, 1996 ; & , 2006).
Pendant cette période (Albien-Turonien) Madagascar appartient à la "South Gondwana Fauna" (sensu , 1988), caractérisée tout particulièrement par les genres Arculicythere et Majungaella ( et alii, 1998), mais on note l'apparition dès le Cénomanien d'éléments de la "Pan Gondwana Fauna" (sensu , 1988), avec le genre Brachycythere. Or, selon (1988) et (2002), ce genre ne serait apparu dans cette province qu'à partir du Turonien (ceci est repris aussi par , 2003). La présence de Brachycythere dès le Cénomanien en Inde (Rajasthan) confirme nos observations (, 1997 ; et alii, sous presse).
En conclusion on peut affirmer que pendant l'intervalle Albien-Turonien, les faunes d'ostracodes malgaches montrent de très fortes affinités avec celles du NW de l'Inde (Rajasthan : travaux de , 1997, et et alii, sous presse) ce qui pourrait définir une province faunistique indo-malgache.
La classification adoptée utilise celle proposée par et alii (2002) pour les taxons jusqu'à la famille. Pour les taxons de rang inférieur, la classification de (1974) a été suivie.
L'ensemble du matériel et les types figurés sont déposés dans les Collections du Centre de Sédimentologie et Paléontologie de l'Université de Provence, Centre Saint-Charles, Marseille.
Classe Ostracoda ,
1802
Sous-classe Podocopa G.W. , 1894
Ordre Platycopida , 1866
Super-famille Cytherelloidea , 1866
Famille Cytherellidae , 1866
Genre Cytherella , 1849
Cytherella DS1 , 1979
(Pl. 1 ![]() ,
figs. 1-3)
,
figs. 1-3)
1979. Cytherella DS1 et alii, p. 234 , Pl. 4, fig. 5a-b.
Répartition : Albien moyen à Turonien moyen de Madagascar.
Genre Cytherelloidea ,
1929
Cytherelloidea ghotaruensis , 1997
(Pl. 1 ![]() ,
figs. 4-8)
,
figs. 4-8)
1979. Cytherelloidea DS2 et alii,
p. 236, Pl. 5, fig. 3.
1997. Cytherelloidea ghotaruensis : p.
21, Pl. 13, figs. 2, 7-9, 11-13 ; Pl. 14, fig. 1 ; Fig. 2.
sous-presse. Cytherelloidea ghotaruensis et alii,
Pl. 1, figs. 13-17.
Remarques : À noter une certaine affinité avec Cytherelloidea ndumuensis , 1984, du Cénomanien d'Afrique du Sud (Zululand) ; mais, cette dernière en diffère par son bord dorsal à apex médian plus marqué et bord postérieur plus pointu.
Répartition : Albien moyen à Cénomanien inférieur de Madagascar.
Espèce décrite dans le Coniacien de l'Inde (Rajasthan) (, 1997 ; et alii, sous presse).
Cytherelloidea oertlii , 1997
(Pl. 1 ![]() ,
figs. 9-14)
,
figs. 9-14)
1979. Cytherelloidea DS1 et alii,
p. 235, Pl. 4, fig. 11a-b.
1997. Cytherelloidea oertlii nov. sp.- ,
p. 23, Pl. 14, figs. 8-9, 13-14, 16, 20-22.
sous-presse. Cytherelloidea oertlii et alii,
Pl. 2, figs. 3-7.
Répartition : Albien terminal (Vraconnien) à Turonien moyen de Madagascar.
Espèce décrite dans le Coniacien-Santonien de l'Inde (Rajasthan) (, 1997 ; et alii, sous presse).
Cytherelloidea sp.
(Pl. 1 ![]() , fig. 15)
, fig. 15)
Remarques : Un seul individu assez proche de Cytherelloidea oertlii, mais avec un court bourrelet ventral supplémentaire. Le test apparaît finement ponctué.
Répartition : Cénomanien inférieur de Madagascar.
Ordre Podocopida ,
1894
Sous-ordre Cytherocopina , 1850
Super-famille Cytheroidea , 1850
Famille Cytheridae , 1850
Genre Majungaella , 1963 emend. & ,
1996
Majungaella cf. pyriformis & ,
1969
(Pl. 1 ![]() ,
figs. 16-18)
,
figs. 16-18)
1969. Majungaella pyriformis nov. sp.- & ,
p. 133, Pl. 5, fig. 13.
1979. Majungaella pyriformis & et alii,
p. 237, Pl. 5, fig. 10a-b.
Remarques : Seuls trois spécimens ont été trouvés. Cette espèce diffère de celle décrite dans l'Albien de Tanzanie par l'absence de ponctuation. Le spécimen figuré par (in et alii, 1979) dans l'Albien du sondage de Diego présente de nettes ponctuations.
Répartition : Albien moyen-supérieur de Madagascar.
Espèce décrite dans l'Albien de Tanzanie ( & , 1969).
Genre Paraphysocythere ,
1969
Paraphysocythere DS1 (, 1979)
(Pl. 1 ![]() , fig. 19)
, fig. 19)
1979.? Physocythere DS1 et alii, p. 231, Pl. 4, fig. 3a-b.
Remarques : Espèce assez proche de Paraphysocythere thomsoni , 1969, du Santonien d'Afrique du Sud. En diffère essentiellement par un contour des valves plus étiré chez cette dernière.
Répartition : Albien terminal (Vraconnien) à Cénomanien inférieur de Madagascar.
Famille Cytherideidae ,
1925
Sous-famille Schulerideinae , 1959
Genre Pirileberis , 1963
Pirileberis makatiniensis , 1984
(Pl. 1 ![]() ,
figs. 20-22)
,
figs. 20-22)
1979. Habrocythere DS1 et alii,
p. 237, Pl. 5, fig. 5a-b.
1984. Pirileberis makatiniensis nov. sp.- ,
p. 170-174, figs. 31D-F, 32, 33A.
Répartition : Albien moyen à supérieur de Madagascar.
Cette espèce a été décrite dans l'Albo-Aptien du Zululand en Afrique du Sud. Des formes proches ont été signalées dans le Cénomanien des Indes (Rajasthan), sous l'appellation Isohabrocythere aff. DS1 ( et alii, sous presse).
Famille Cytheruridae G.W. ,
1894
Sous-famille Eucytherurinae , 1974 emend. & ,
1987
Genre Microceratina , 1980
Microceratina sp.
(Pl. 1 ![]() ,
figs. 23, 23a)
,
figs. 23, 23a)
Remarques : Espèce très proche de Microceratina bhannesensis , & , 2005 (in et alii, 2005) du Jurassique supérieur du Liban.
Répartition : Base du Cénomanien inférieur de Madagascar.
Sous-famille Cytheropterinae ,
1957
Genre Infracytheropteron , 1964
Infracytheropteron sp.
(Pl. 1 ![]() , fig. 24)
, fig. 24)
Remarques : Forme proche de l'espèce Indet. sp. 327/18 , 1984, de l'Albien du Plateau des Falklands (Atlantique Sud).
Répartition : Cénomanien inférieur de Madagascar.
Famille Schizocytheridae ,
1960
Genre Amphicytherura & ,
1957
Amphicytherura sp. 5 , &
(sous presse)
(Pl. 1 ![]() ,
figs. 25-26)
,
figs. 25-26)
sous presse. Amphicytherura sp. 5 - et alii, Pl. 7, figs. 5-7.
Répartition : Cénomanien inférieur de Madagascar.
Espèce signalée dans le Cénomanien de l'Inde (Rajasthan) ( et alii, sous presse).
Famille Trachyleberididae ,
1948
Sous-famille Trachyleberidinae ,
1948
Genre Cythereis , 1849
Cythereis micropapillata nov. sp.
(Pl. 2 ![]() ,
figs. 1-3)
,
figs. 1-3)
Origine du nom : Présence de papilles dans les mailles du réticulum.
Holotype : Une carapace (Pl. 2 ![]() ,
fig. 1), HCE 19.
,
fig. 1), HCE 19.
Paratypes : PCE 19/1, PCE 19/2 (Pl. 2 ![]() ,
figs. 2-3).
,
figs. 2-3).
Localité-type : Antsiranana, Madagascar, coupe de l'Abattoir, Formation de l'Andranomaimbo, Cénomanien inférieur.
Diagnose : Espèce du genre Cythereis caractérisée par la présence de micropapilles dans les mailles du réticulum.
Dimensions : Holotype : L = 0,750 mm ; h = 0,400 mm
Paratypes : L = 0,660-0,680 mm ; h = 0,350-0,400 mm
Remarques : Cette espèce présente une ornementation très particulière sous forme de mailles circulaires régulièrement
disposées, contenant une dizaine de micropapilles apparemment creuses (Pl. 2 ![]() , fig. 1b). Les tubercules oculaire et musculaire sont bien
différenciés, ce dernier étant entouré d'une rosette de pore-conuli.
, fig. 1b). Les tubercules oculaire et musculaire sont bien
différenciés, ce dernier étant entouré d'une rosette de pore-conuli.
Une espèce proche est Cythereis sp. et alii (sous presse), de l'Albien de l'Inde (Rajasthan).
Age et distribution : Cénomanien inférieur de Madagascar.
Genre Megommatocythere & ,
1982
Megommatocythere sp.
(Pl. 2 ![]() ,
figs. 4-5)
,
figs. 4-5)
Remarques : Le tubercule oculaire très en relief ainsi que la forme de la carapace justifie l'attribution au genre Megommatocythere (voir et alii, 1982), jusqu'à présent connu du Campanien au Paléocène.
Répartition : Cénomanien inférieur.
Genre Donmacythere ,
1976
Donmacythere vellicata nov. sp.
(Pl. 3 ![]() ,
figs. 1-6)
,
figs. 1-6)
1979. ? Batavocythere DS1 et alii, p. 231, Pl. 4, fig. 1.
Origine du nom : Du latin vellicare (pincé), en raison de l'aspect pincé de la valve entre le tubercule musculaire et la côte postéro-médiane.
Holotype : Une carapace (Pl. 3 ![]() , fig. 1), HCE21
, fig. 1), HCE21
Paratypes : PCE21/1 à PCE21/10
Localité-type : Antsiranana, Madagascar, coupe de l'Abattoir, Formation de l'Andranomaimbo, Cénomanien inférieur.
Diagnose : Espèce attribuée au genre Donmacythere , 1976, à tubercule musculaire et costule médiane peu développée, reliés par un pont formant pincement entre deux dépressions adjacentes.
Remarques : Cette espèce diffère de l'espèce-type, Donmacythere damottae (, 1974) du Cénomanien du S de la France, qui possède une très longue côte médiane lisse oblique reliée à l'extrémité postérieure de la côte dorsale. Donmacythere inferangulata ( & , 1972) du Cénomanien du SE de la France possède trois côtes longitudinales étroites et lisses. Donmacythere hafsuni , 1984, du Turonien inférieur d'Espagne, présente des bourrelets ventral et dorsal lisses, ce dernier débordant largement le bord ventral.
Dimensions : Holotype : L = 0,800 mm ; h = 0,470 mm
Paratypes : L = 0,660-0,750 mm ; h = 0,400-0,460 mm
Répartition : Cénomanien inférieur et base du Turonien inférieur de Madagascar.
Genre Parvacythereis ,
1973
Parvacythereis ? jamba nov. sp.
(Pl. 3 ![]() ,
figs. 7-10)
,
figs. 7-10)
1979. Cythereis DS6 et alii, p. 238, Pl. 5, fig. 11a-b
Origine du nom : Du malgache jamba, aveugle.
Holotype : Une carapace femelle (Pl. 3 ![]() , fig. 10), HAL2.
, fig. 10), HAL2.
Paratypes : PAL2/1-PAL2/3.
Localité-type : Flanc S de l'Andrahona, 40 km SE d'Antsiranana, Madagascar, Membre b de la Formation Andrahona, Albien supérieur.
Diagnose : Espèce attribuée sous réserve au genre Parvacythereis caractérisée par une absence de tubercule oculaire et une réticulation primaire et secondaire.
Dimensions : Holotype : L = 0,680 mm ; h = 0,400mm
Paratypes : L = 0,650-0,750 mm ; h = 0,410-0,460 mm
Remarques : En raison de sa forme générale et de sa petite taille cette espèce est provisoirement attribuée au genre Parvacythereis. L'absence de tubercule oculaire ne permet pas de justifier pleinement cette attribution générique, d'où notre réserve. Des formes proches ont été attribuées à ce genre par , notamment Parvacythereis monziensis , 1981, du Campanien-Maastrichtien du Zululand et Parvacythereis spinosa (, 1971) du Maastrichtien du Banc des Agulhas, Afrique du Sud. Cette dernière espèce est aussi aveugle. Une autre espèce aussi très proche est Isocythereis ? sp. 3 , 1998, de l'Albien d'Argentine, espèce aussi aveugle.
Répartition : Albien moyen à supérieur de Madagascar.
Genre Spinoleberis ,
1966
Spinoleberis ? sp.
(Pl. 4 ![]() ,
figs. 7-8)
,
figs. 7-8)
Remarques : Seulement 2 spécimens tentativement attribués au genre Spinoleberis.
Répartition : Albien moyen-supérieur de Madagascar
Genre Glenocythere ,
1979
Sous-genre Hemiglenocythere nov. subgen.
Origine du nom : En raison de la charnière hemiamphidonte.
Diagnose : Sous-genre de Glenocythere caractérisé par une charnière hemiamphidonte.
Remarques : La diagnose originale du genre Glenocythere (, 1979) mentionne une charnière amphidonte hétérodonte avec sillon médian très nettement crénelé et dent distale lisse.
Espèce-type : Glenocythere (Hemiglenocythere) robusta nov. sp.
Autres espèces rattachées à ce genre : Glenocythere (Hemiglenocythere) antsirananaensis nov. sp. de l'Albien de Madagascar et Glenocythere sp. 1 et alii (sous presse) du Cénomanien de l'Inde (Rajasthan).
Glenocythere (Hemiglenocythere) robusta nov. sp.
(Pl. 3 ![]() ,
figs. 11-17 ; Pl. 4
,
figs. 11-17 ; Pl. 4 ![]() , fig. 1)
, fig. 1)
1979. ? Batavocythere DS 1 et alii, p. 231, Pl. 4, fig. 1a-b.
Origine du nom : D'après la grande taille et robustesse des carapaces.
Holotype : une carapace (Pl. 3 ![]() , fig. 11), HCE22.
, fig. 11), HCE22.
Paratypes : PCE22/1 à PCE22/20.
Localité-type : Antsiranana, Madagascar, coupe de l'Abattoir, Formation de l'Andranomaimbo, Cénomanien inférieur.
Diagnose : Espèce du sous-genre Hemiglenocythere de grande taille, à test lisse et côte dorsale courte et aigue.
Dimensions : Holotype : L = 0,950 mm ; h = 0,600 mm
Paratypes : L = 0,80-0,980 ; h = 0,480-0,620 mm
Remarques : Les valves sont très renflées en leur milieu, donnant cet aspect de lourdeur
caractéristique, plus aplaties le long du bord antérieur. Tubercule musculaire inexistant. Bord dorsal rectiligne caché par une côte étroite et aplatie en position médiane et inclinée vers l'avant. Côte
ventrale, très régulièrement arquée et débordant largement le bord ventral. Présence de quelques épines sur les bords postéro- et antéro-ventraux. La charnière
(Pl. 3 ![]() ,
figs. 15-17) est de type hemiamphidonte avec à la valve droite une dent
antérieure, un sillon médian lisse et une dent postérieure lobée (3-4 denticules).
,
figs. 15-17) est de type hemiamphidonte avec à la valve droite une dent
antérieure, un sillon médian lisse et une dent postérieure lobée (3-4 denticules).
Glenocythere sp. 1 et alii (sous presse), du Cénomanien de l'Inde (Rajasthan) est morphologiquement très proche mais la charnière n'a pas pu être observée.
Répartition : Albien terminal (Vraconnien) Cénomanien inférieur de Madagascar.
Glenocythere (Hemiglenocythere) antsirananaensis nov. sp.
(Pl. 4 ![]() ,
figs. 2-6)
,
figs. 2-6)
1979. ? Batavocythere DS2 et alii, p. 238, Pl. 5, fig. 14.
Origine du nom : De la ville d'Antsiranana (Madagascar).
Holotype : Une carapace (Pl. 4 ![]() , fig. 4), HAL3.
, fig. 4), HAL3.
Paratypes : PAL3/1 à PAL3/15.
Localité-type : Flanc S de l'Andrahona, 40 km SE d'Antsiranana, Madagascar, Membre a de la Formation Andrahona, Albien supérieur.
Diagnose : Espèce du sous-genre Hemiglenocythere à carapace allongée subrectangulaire, lisse. Côte dorsale convexe ; côte ventrale épaisse surélevée par rapport au bord ventral.
Dimensions : Holotype : L = 0,730 mm ; h = 0,420 mm
Paratypes : L = 0,660 0,780 mm ; h = 0,380 0,440 mm
Remarques : Cette espèce diffère de Glenocythere (Hemiglenocythere) robusta par une taille moindre et une carapace nettement plus allongée et sub-rectangulaire. La charnière est hemiamphidonte, conforme à la diagnose générique. Le dimorphisme sexuel est bien marqué, les mâles étant plus allongés.
Répartition : Albien moyen et supérieur de Madagascar.
Sous-famille Unicapellinae ,
1981
Genre Unicapella , 1980
Unicapella yvettae nov. sp.
(Pl. 2 ![]() ,
figs. 6-9)
,
figs. 6-9)
1979. Cythereis DS4 et alii, p. 236, Pl. 5, fig. 4a-b
Origine du nom : En hommage posthume à notre collègue et amie ostracodologiste Yvette (Toulouse).
Holotype : une carapace (Pl. 2 ![]() , fig. 6), HAL1.
, fig. 6), HAL1.
Paratypes : PAL1/1-PAL1/10 (Pl. 2 ![]() ,
figs. 7-8).
,
figs. 7-8).
Localité-type : Flanc S de l'Andrahona, 40 km SE d'Antsiranana, Madagascar, Membre a de la Formation Andrahona, Albien moyen.
Diagnose : Espèce du genre Unicapella présentant un lobe cardinal ("hinge ear"), très développé sur la valve gauche ; une côte dorsale étroite et sinueuse et un tubercule musculaire peu développé.
Dimensions : Holotype : L = 0,670 mm ; h = 0,360 mm
Paratype (PAL1/1) : L = 0,630 ; h = 0,330 mm
Remarques : La carapace présente une très fine réticulation sous forme d'un bandeau le long du bord antérieur, et des mailles de réticulum étirées verticalement sous le bord dorsal. Le lobe cardinal possède en son centre un pore conulus. La côte dorsale est étroite et sinueuse et le tubercule musculaire peu développé, ce qui différencie cette espèce des autres espèces du genre Unicapella et en particulier de l'espèce-type Unicapella sacsi , 1980 (sensu , 1981) du Campanien-Maastrichtien d'Afrique du Sud. Selon (1981, 1985), l'importance du lobe cardinal est très variable à l'intérieur d'une même espèce. (2002), crée le genre Hiltermanniana à partir de matériel de l'Aptien d'Égypte, pour des formes à très grand lobe cardinal, mais dans sa discussion il ne compare pas avec le genre Unicapella dont la création est nettement antérieure. Il attribue l'espèce Cythereis DS4 à ce nouveau genre.
Une espèce très proche, mais qui diffère par la présence d'une crête longitudinale en arrière du tubercule musculaire, a été décrite comme Golcocythere oertlii par (1978) dans l'Albien de l'Inde (Tamil Nadul).
Répartition : Albien moyen-supérieur de Madagascar.
Unicapella sp.
(Pl. 2 ![]() , fig. 10)
, fig. 10)
Remarques : Espèce très proche d'Unicapella yvettae nov. sp., mais qui en diffère par un alignement de nodules sur la côte ventrale et un bord dorsal plus rectiligne.
Répartition : Albien moyen-supérieur à base du Cénomanien inférieur de Madagascar.
Genre Dutoitella ,
1981
Dutoitella sp.
(Pl. 2 ![]() , fig. 11)
, fig. 11)
Remarques : Espèce proche de Dutoitella dutoiti , 1981, du Maastrichtien d'Afrique du Sud, et qui en diffère essentiellement par l'absence de tubercule musculaire et de relief postéro-médian.
Répartition : Cénomanien inférieur de Madagascar.
Genre Taracythere ,
1995
Taracythere antakaranaensis nov. sp.
(Pl. 2 ![]() ,
figs. 12-19)
,
figs. 12-19)
1979. Cythereis DS3 et alii, p. 236, Pl. 4, fig. 14a-b.
Origine du nom : D'après l'ethnie Antakarana du N de Madagascar.
Holotype : Une valve gauche, mâle (Pl. 2 ![]() , fig. 13), HCE20.
, fig. 13), HCE20.
Paratypes : PCE20/1 à PCE20/7.
Localité-type : Antsiranana, Madagascar, coupe de l'Abattoir, Formation de l'Andranomaimbo, Cénomanien inférieur.
Diagnose : Espèce attribuée au genre Taracythere caractérisée par une carapace sub-rectangulaire, une extrémité postérieure triangulaire non relevée et une ornementation complexe.
Dimensions : Holotype L = 0,800 mm ; h = 0,430 mm
Paratypes : L = 0,710-0,780 mm ; h = 0,410-0,460 mm.
Remarques : Le genre Taracythere a été décrit par (1995) pour des espèces de Trachyleberididae sans tubercule oculaire ni lobe cardinal de l'Éocène de Nouvelle Zélande. Ultérieurement, & (2003) ont révisé ce genre à partir de matériel actuel. Dans cette étude ils démontrent que ce genre n'est pas restreint à la Nouvelle Zélande et que d'autres espèces pouvant y être rattachées sont connues dès le Paléogène de l'Océan Indien, notamment Actinocythereis orientalis orientalis , 1985, et Actinocythereis orientalis rete , 1993 (voir aussi & , 1992). Taracythere antakaranaensis nov. sp., par sa morphologie générale (extrémité postérieure non relevée, présence d'une nodosité musculaire, et crêtes ventrale et dorsale bien individualisées et noduleuses), est beaucoup plus proche des formes du Paléogène de l'Océan Indien.
L'ornementation de base est en accord avec la diagnose générique : réticulation primaire et secondaire affectant toute la surface des valves. Une caractéristique diagnostique de cette espèce est l'abondance de pore conuli élevés aux intersections des mailles du réticulum primaire. La forme générale, l'absence de tubercule oculaire et l'ornementation complexe nous amènent à ranger ce genre dans la sous-famille des Unicapellinae , 1981.
Répartition : Albien terminal (Vraconnien) à Cénomanien inférieur de Madagascar.
Taracythere DS1 (, 1979)
(Pl. 4 ![]() , fig. 9)
, fig. 9)
1979. Cythereis DS1 et alii, p. 231 , Pl. 4, fig. 2a-b.
Remarques : Cette espèce possède un réticulum primaire bien différencié et des pores conuli de petite taille.
Répartition : Albien terminal (Vraconnien) à Cénomanien inférieur de Madagascar.
Sous-famille Brachycytherinae ,
1954
Genre Brachycythere , 1929 emend. ,
2002
Brachycythere ? kharatarensis (, 1997)
(Pl. 5 ![]() ,
figs. 4-8)
,
figs. 4-8)
1979. ? Veenia DS1 et alii,
p. 235, Pl. 4, fig. 13a-b.
1997. Veenia kharatarensis : p.
20, Pl. 11, figs. 5-11.
sous presse. Brachycythere kharatarensis () et alii,
Pl. 3, figs. 14-16.
Remarques : L'élément postérieur de la charnière de la valve droite
(Pl. 5 ![]() , fig. 7), n'est pas allongé et denticulé comme dans le genre Brachycythere sensu stricto (,
2002). L'attribution générique n'est donc pas justifiée. Faute de matériel bien
conservé, nous gardons provisoirement cette appellation. La charnière étant de type
amphidonte, l'attribution au genre Arculicythere proposée par &
(2006) ne peut être
retenue, ni sa synonymie avec Arculicythere defluxa ,
1963, du Valanginien de Madagascar. Brachycythere sp. 1 ,
1991, de l'Albien de la région d'Essaouira (Maroc), pourrait aussi appartenir à ce
genre, ainsi qu'une espèce similaire de l'Albien du Venezuela ( & ,
1996).
, fig. 7), n'est pas allongé et denticulé comme dans le genre Brachycythere sensu stricto (,
2002). L'attribution générique n'est donc pas justifiée. Faute de matériel bien
conservé, nous gardons provisoirement cette appellation. La charnière étant de type
amphidonte, l'attribution au genre Arculicythere proposée par &
(2006) ne peut être
retenue, ni sa synonymie avec Arculicythere defluxa ,
1963, du Valanginien de Madagascar. Brachycythere sp. 1 ,
1991, de l'Albien de la région d'Essaouira (Maroc), pourrait aussi appartenir à ce
genre, ainsi qu'une espèce similaire de l'Albien du Venezuela ( & ,
1996).
Répartition : Albien terminal (Vraconnien) Cénomanien inférieur de Madagascar.
Cénomanien à Coniacien de l'Inde (Rajasthan).
Brachycythere pucketti nov. sp.
(Pl. 5 ![]() ,
figs. 9-14)
,
figs. 9-14)
1979. ? Sphenocytheridea DS1 et alii, p. 234, Pl. 4, fig. 6.
Origine du nom : En hommage à notre collègue Terry M. (University of Alabama, U.S.A.) auteur d'une récente révision des Brachycytherinae.
Holotype : carapace (Pl. 5 ![]() , fig. 9), HCE24.
, fig. 9), HCE24.
Paratypes : PCE24/1-PCE24/5.
Localité-type : Antsiranana, Madagascar, coupe de l'Abattoir, Formation de l'Andranomaimbo, Cénomanien inférieur.
Diagnose : Espèce du genre Brachycythere, de taille moyenne, carapace lisse triangulaire allongée avec des denticules postéro- et antéro-ventraux. Sillon d'accommodation très large et profond dans la valve gauche.
Dimensions : Holotype : L = 0,850 mm, h = 0,470 mm
Paratypes : L = 0,730-0,840 mm ; h = 0,420-0,460 mm
Remarques : La charnière de la valve droite diffère de celle du genre Brachycythere (type du genre : Brachycythere crenulata , 1965) par une dent postérieure crénelée mais courte et massive.
Répartition : Albien terminal (Vraconnien) - Cénomanien inférieur de Madagascar.
Genre Malagasyella nov. gen.
Origine du nom : De la langue malagasy, parlée à Madagascar.
Diagnose : Genre de Trachyleberididae à carapace triangulaire allongée, renflée, sans costulation ni tubercule musculaire et avec tubercule oculaire très proéminent. Ornementation avec réticulum primaire et secondaire sur toute la surface des valves. Charnière hemiamphidonte.
Remarques : Ce genre diffère du genre Opimocythere , 1968, du Danien des États-Unis, par son contour plus triangulaire, son ornementation complexe, la présence de fort tubercule oculaire. La charnière hemiamphidonte est en parfait accord avec l'attribution à la sous-famille des Brachycytherinae (, 2002).
Espèce-type : Malagasyella dantelyornata nov. sp.
Autres espèces rattachées à ce genre : Majungaella rajendrai , 1997 (Albien-Coniacien de l'Inde), Majungaella ? sp. 3 et alii, sous presse (Cénomanien de l'Inde), Brachycythere batei , 1975 (Coniacien) de l'Inde, Majungaella ? sp. 1 et alii, sous presse (Coniacien de l'Inde), Majungaella ? sp. 2 et alii, sous presse (Cénomanien supérieur de l'Inde). Des formes proches mais sans tubercule oculaire proéminent ont été figurées par & (1992) sous l'appellation Brachycythere sp. 1 dans le Paléocène supérieur du Pakistan.
Malagasyella dantelyornata nov. sp.
(Pl. 4 ![]() ,
figs. 10-13)
,
figs. 10-13)
1979. ? Velarocythere DS1 et alii, p. 235, Pl. 4, fig. 10.
Origine du nom : De dantely, dentelle en malgache, en raison de l'ornementation.
Holotype : Carapace (Pl. 4 ![]() , fig. 10), HCE23.
, fig. 10), HCE23.
Paratypes : PCE23/1-PCE23/3.
Localité-type : Antsiranana, Madagascar, coupe de l'Abattoir, Formation de l'Andranomaimbo, Cénomanien inférieur.
Diagnose : Celle du genre.
Dimensions : Holotype : L = 0,780 mm ; h = 0,420 mm
Paratypes : L = 0,730-0,760 mm ; h = 0,400-0,420 mm
Répartition : Cénomanien inférieur et Turonien inférieur de Madagascar.
Malagasyella aff. rajendrai (, 1997)
(Pl. 4 ![]() ,
figs. 14-15 ; Pl. 5
,
figs. 14-15 ; Pl. 5 ![]() ,
figs. 1-3)
,
figs. 1-3)
aff. 1997. Majungaella rajendrai ,
p. 15-16, Pl. 8, figs. 4-9 ; Pl. 9, fig. 1.
aff. sous presse. Majungaella ? rajendrai et alii,
Pl. 9, figs. 7-10, 13-17.
Remarques : Cette espèce diffère de Malagasyella dentelyornata essentiellement par une ornementation formée d'alvéoles étirés verticalement.
Répartition : Turonien inférieur et moyen (Formation Ambolafotsy) de Madagascar.
En Inde, cette espèce a été décrite dans le Coniacien et retrouvée jusque dans l'Albien supérieur du Rajasthan ( et alii, sous presse).
Sous-ordre Bairdiocopina ,
1866
Super-famille Bairdioidea , 1888
Famille Bythocyprididae , 1969
Genre Robsoniella , 1956
Robsoniella postelongata nov. sp.
(Pl. 5 ![]() ,
figs. 15-17)
,
figs. 15-17)
Origine du nom : En raison de l'allongement de la partie postérieure de la carapace.
Holotype : Une carapace (Pl. 5 ![]() , fig. 16), HCE25.
, fig. 16), HCE25.
Paratypes : PCE25/1, PCE25/2.
Localité-type : Antsiranana, Madagascar, coupe de l'Abattoir, Formation de l'Andranomaimbo, Cénomanien inférieur.
Diagnose : Espèce du genre Robsoniella caractérisée par une carapace relativement allongée.
Dimensions : Holotype : L = 0,830 mm ; h = 0,480 mm.
Paratypes : L = 0,780-0,800 mm ; h = 0,360-0,410 mm.
Remarques : Cette espèce diffère de l'espèce-type Robsoniella obovata ,
1956 (voir illustrations in ,
1961), de l'Aptien d'Azerbaïdjan et de Robsoniella falklandensis ,
1984, de l'Albien du Plateau des Falklands et d'Australie par l'allongement de la carapace et une extrémité postérieure plus acuminée. Robsoniella longa
1961, de l'Aptien d'Azerbaïdjan est encore plus allongée mais possède une extrémité postérieure large et arrondie. Les fines crénulations ainsi que la disposition des empreintes musculaires sont caractéristiques de ce genre
(Pl. 5 ![]() ,
figs. 17, 17a).
,
figs. 17, 17a).
Répartition : Cénomanien inférieur et Turonien inférieur de Madagascar.
Sous-ordre Cypridocopina ,
1945
Super-famille Pontocypridoidea G.W. , 1894
Famille Pontocyprididae G.W. , 1894
Genre Pontocyprella , 1955
Pontocyprella DS 1 , 1979
(Pl. 5 ![]() , fig. 18)
, fig. 18)
1979. Pontocyprella DS1 et alii, p. 234, Pl. 4, fig. 8.
Répartition : Albien moyen à Cénomanien inférieur de Madagascar.
Pontocyprella sp.
(Pl. 5 ![]() , fig. 19)
, fig. 19)
Répartition : Cénomanien inférieur-Turonien moyen de Madagascar.
Les auteurs remercient G. (Université de Provence, Marseille), pour son contrôle des datations (notamment en ce qui concerne le Turonien), A. et R. pour les prises de vue (Appareil S XL 30 ESEM), A. pour le montage définitif des planches, ainsi que les rapporteurs, nos collègues M.C. (Lisbonne, Portugal) et J. (Bilbao, Espagne), pour leurs judicieuses corrections.
S. (1979).- Glenocythere, a new ostracode genus from the Hamadi Formation (Cretaceous) of Kuwait.- Journal of Paleontology, Tulsa, vol. 53, n° 4, p. 920-930.
B. (1991).- Les ostracodes du Crétacé Moyen (Barrémien à Turonien) le long d'une transversale Agadir-Nador.- Strata, Toulouse, (Sér. 2), vol. 14, 765 p.
B., J.-P. & J. (sous-presse).- Cretaceous (Albian to Coniacian) ostracodes from the subsurface of the Jaisalmer Basin, Rajasthan, India.- Micropaleontology, New-York.
M.A. (1995).- Late Eocene ostracoda (Crustacea) from the Waihao District, South Canterbury, New Zealand.- Journal of Paleontology, Tulsa, vol. 69, n° 5, p. 897-921.
J.-F. & J.-P. (1988).- Paleobiogeography of Tethyan Cretaceous marine ostracodes. In : T., N. & K. (eds.), Evolutionary Biology of Ostracodes. Its Fundaments and Applications.- Elsevier-Kodansha Ltd., Tokyo-Amsterdam, p. 625-641.
J.-F. & J.-P. (1992a).- Marine ostracode provincialism in the Late Cretaceous of the Tethyan realm and the Austral province.- Palaeogeography, Palaeoclimatology, Palaeoecology, Amsterdam, vol. 92, n° 3-4, p. 283-293.
J.-F. & J.-P. (1992b).- On Arcacythere aurani n. sp.- A Stereo-Atlas of Ostracod Shells, London, vol. 19, n° 2, p. 91-94.
J.-F., J.-P. & A. (1996).- Les ostracodes du Maastrichtien et du Danien du Bassin de Mahajanga, Madagascar : données préliminaires. In : S., I. & J.-P. (eds.), Géologie de l'Afrique et de l'Atlantique Sud.- Bulletin des Centres de Recherche Exploration-Production Elf-Aquitaine, Mémoire 16, Pau, p. 6.
S.C. (1998).- Ostrácodos del Albiano de la Cuenca austral, Argentina.- Ameghiniana, Buenos Aires, vol. 35, n° 4, p. 355-377.
S.C., D.I. & R. (1998).- The ostracod genus Majungaella in Argentina.- Revista Geologica de Chile, Santiago, vol. 25, n° 1, p. 45-55.
S. & R. (2006).- The Mesozoic ostracod genus Arculicythere : further evidence for the southern Gondwana seaway.- Cretaceous Research, London, vol. 27, n° 6, p. 728-734.
M.A.A. (2002).- Middle Cretaceous (Aptian-Early Turonian) ostracoda from Sinai, Egypt.- Neue Paläontologische Abhandlungen, Dresden, Bd. 5, p. 1-123.
R.H. & D.D. (1969).- An outline account of the Cretaceous and Tertiary foraminifera and of the Cretaceous ostracods of Tanzania.- Proceedings of the Third African Micropaleontological Colloquium, Cairo, p. 113-164.
R.H. (1977).- The Cenozoic ostracode faunas of the Sao Paulo Plateau and the Rio Grande Rise (DSDP Leg 39, Sites 566 and 357). In : P.R., K. et alii, Initial Reports of the Deep Sea Drilling Project, Washington, vol. 39, p. 869-883.
H. (1972).- Géologie de Madagascar. I Les terrains sédimentaires.- Annales de Géologie de Madagascar, Antananarivo, n° 35, 463 p.
E. & S.F. (1992).- Biostratigraphic and paleoecologic analysis of ostracode assemblages from Late Paleocene and Early Eocene sedimentary rocks, core UAK-5, Ganjo Takkar Inlier, Sindh Province, Pakistan.- U.S. Geological Survey Open-File Report 92-517, Denver, 78 p.
J.-P. (1974).- Contribution à l'étude des ostracodes du Crétacé supérieur de Dordogne.- Geobios, Villeurbanne, n° 7, fasc. 1, p. 19-42.
J.-P., B., J. & R. (2005).- Sur l'origine du genre Microceratina , 1980 (Ostracoda, Eucytherurinae) et sa présente téthysienne dès le Jurassique supérieur.- Revue de Micropaléontologie, Paris, vol. 48, n° 1, p. 15-24.
J.-P. & J.-F. (1996).- Preliminary account of ostracodes from the Aptian-Albian of Venezuela. In : M.C. (ed.), Proceedings of the 2nd European Ostracodologists Meeting.- British Micropalaeontological Society, London, p. 29-34.
M., J. & N. (1979).- L'Albien et le Cénomanien du sondage de Diego (Madagascar) et ses faunes d'ammonites, foraminifères planctoniques et ostracodes.- Eclogae Geologica Helvetiae, Basel, vol. 72, n° 1, p. 215-249.
R.V. (1969).- Upper Senonian ostracods from the coast of Pondoland, South Africa.-Transactions of the Royal Museum of South Africa, Cape Town, vol. 38, n° 4, p. 347-385.
R.V. (1971).- Some Cretaceous ostracodal assemblages from the Agulhas Bank (South African continental margin).- Transactions of the Royal Museum of South Africa, Cape Town, vol. 40, part 4, p. 393-418.
R.V. (1980).- Marine Santonian and Campanian ostracods from a borehole at Richards Bay, Zululand.- Annals of the South African Museum, Cape Town, vol. 82, part 1, p. 1-70.
R.V. (1981).- The Campanian and Maastrichtian ostracoda of South-East Africa.- Annals of the South African Museum, Cape Town, vol. 85, part 1, p. 1-101.
R.V. (1984).- Mid-Cretaceous ostracoda from southern Africa and the Falkland Plateau.- Annals of the South African Museum, Cape Town, vol. 93, part 3, p. 97-211.
R.V. (1985).- Turonian, Coniacian, and Santonian ostracoda from south-east Africa.- Annals of the South African Museum, Cape Town, vol. 96, part 5, p. 123-239.
R.V. (1988).- Marine ostracode distributions during the early breakup of southern Gondwanaland.- In : T., N. & K (eds.), Evolutionary Biology of Ostracoda. Its Fundamentals and Applications.- Elsevier-Kodansha, Amsterdam-Tokyo, p. 841-854.
R.V. (1996).- Cretaceous ostracoda of the SE Atlantic and SW Indian Ocean, a stratigraphical review and atlas. In : S., I. de & J.-P. (eds.), Géologie de l'Afrique et de l'Atlantique Sud.- Bulletin des Centres de Recherche Exploration-Production Elf-Aquitaine, Mémoire 16, Pau, p. 1-17.
P., J.-P., R., H.J., J.-P. & R. (1982).- Les ostracodes du Campanien terminal à l'Éocène inférieur de la coupe du Kef, Tunisie nord-occidentale.- Bulletin des Centres de Recherche Exploration-Production ElfAquitaine, Pau, vol. 6, n° 2, p. 273-335.
P. & B. (1972).- Les ostracodes de la sous-famille des Trachyleberidinae dans quelques coupes de référence du Cénomanien du Sud-Est de la France.- Revista Española de Micropaleontologia, Madrid, vol. 4, n° 3, p. 355-376.
N. (1963).- Contribution à l'étude des ostracodes du Mésozoïque moyen (Bathonien-Valanginien) du Bassin de Majunga, Madagascar.- Revue de l'Institut Français du Pétrole, Paris, vol. 18, n° 12, p. 709-762.
C. (1985).- Ostracodes paléogènes de quelques sites "D.S.D.P." de l'Océan Indien (Legs 22 et 23).- Revue de Paléobiologie, Genève, vol. 4, n° 2, p. 279-295.
C. (1993).- Ostracodes du Plateau d'Exmouth (Océan Indien) : remarques systématiques et évolution des environnements océaniques profonds au cours du Cénozoïque.- Geobios, Villeurbanne, n° 26, fasc. 3, p. 345-360.
C. & B. (1992).- Data Report: preliminary reports on the ostracodes of Leg 122 (Exmouth Plateau, Indian Ocean).- In : U. von, B.U. et al, Proceedings of the Ocean Drilling Program, Scientific Results, College Station, vol. 122, p. 835-837.
G. & H.S. (1974).- Summary of neontological and paleontological classification of ostracoda.- Mitteilungen aus dem Hamburgischen Zoologischen Museum und Institut, Hamburg, Bd. 70, p. 7-73.
J.E. (1968).- Ostracodes from the Brighseat Formation (Danian) of Maryland.- Journal of Paleontology, Tulsa, vol. 42, n° 1, p. 100-142.
G.F.W. (1982).- Palynological analysis of an additional series of Cretaceous samples from Madagascar.- Report n° 2152 A, 4 p.
G.F.W., A., & J.W. (1982).- Micropaleontology of Albian to Danian strata in Madagascar.- Micropaleontology, New-York, vol. 28, n° 1, p. 97-109.
D.J., A. & K. (2002). Taxonomy, morphology and biology of Quaternary and living ostracoda. In : J.A. & A.R. (eds.), The ostracoda: Applications in Quaternary Research.- Geophysical Monograph 131, American Geophysical Union, Washington D.C., p. 5-36.
S.P. (1975).- Ostracoda from the Bagh Beds (Upper Cretaceous) of Madhya Pradesh.- Geophytology, Lucknow, vol. 5, n° 2, p. 188-212.
S.P. (1978).- Further ostracoda from the Kallakkudi Limestone (Albian) Tiruchirapalli district, Tamil Nadu, India.- Neues Jahrbuch für Geologie und Paläontologie Monatshefte, Stuttgart, Heft 8, p. 502-512.
T. & K.M. (2003).- Report on the taxonmy, biogeography and phylogeny of mostly living benthic ostracoda (Crustacea) from deep-sea samples (intermediate water depths) from the Challenger Plateau (Tasman Sea) and Campbell Plateau (Southern Ocean), New Zealand.- Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, Frankfurt am Main, n° 558, 329 p.
Y., H. & K. (1977).- Lower Cenomanian mollusks from Diego-Suarez, northern Madagascar.- Bulletin of the National Science Museum, Tokyo, (Ser. C), vol. 3, n° 2, p. 107-132.
Z.V. (1956).- New ostracode species and varieties from the Lower Cretaceous of northeastern Azerbaidjan.- Trudy Azerbaydzhanskogo Nauchno-Issledovatelskogo Instituta du Dobyche Nefti, Baku, p. 49-70 (en russe).
Z.V. (1961).- Ostracodes from Cretaceous deposits of Azerbaidjan and their stratigraphical importance.- Baku, 149 p. (en russe).
P. (2003).- Paleobiogeography of late Early Cretaceous to Early Paleocene marine ostracoda in Arabia and North to Equatorial Africa.- Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 196, n° 3-4, Amsterdam, p. 319-342.
H.-J. (1974).- Lower Cretaceous and Jurassic ostracods from DSDP Leg 27 a preliminary account.- Initial Reports of the Deep Sea Drilling Project, vol. 27, p. 947-965.
T.M. (2002).- Systematics and paleobiogeography of brachycytherine ostracoda.- Micropaleontology, New-York, vol. 48, (suppl. 2), 878 p.
A.H (1990).- Les ostracodes du Jurassique Moyen Néocomien du Bassin de Majunga (Madagascar).- Geobios, Villeurbanne, n° 23, fasc. 4, p. 415-443.
A. (1989).- Étude stratigraphique et micropaléontologique des bassins sédimentaires malgaches situés au nord de la Betsiboka, de l'Albien moyen à la limite Crétacé-Tertiaire.- Thèse Doctorat ès Sciences 1986, Université d'Aix-Marseille I.- Impr. Petroconsultants S.A., Genève, 248 p.
A. & R. (1989).- La lignée Hedbergella wondersi nov. sp.- Planomalina buxtorfi () (foraminifères planctoniques) dans l'Albo-Cénomanien du Bassin d'Antsiranana (nord de Madagascar).- Geobios, Villeurbanne, n° 22, fasc. 6, p. 803-823.
R.A. (1984).- Upper Cretaceous ostracoda of North Central Spain.- Bulletin of the Geological Institute of the University of Uppsala, Uppsala, (n.s.), vol. 10, p. 67-110.
A. & A. (1978).- A preliminary note on recent ostracodes from shelf to rise sediments off north-east Africa. Meteor Forschungsergebnisse, Stuttgart, (Reihe C), n° 29, p. 14-20.
J., N., J.-F. & M. (1974).- Microfaune des dépôts maastrichtiens de la côte orientale de Madagascar. Comparaison avec celle du forage 239 du Glomar Challenger.- Service Géologique de Tananarive, Antananarivo, p. 1-6.
J., N., N.P., A & M. (1970).- Sur l'âge et les affinités "gondwaniennes" de microfaunes (foraminifères et ostracodes) malgaches, indiennes et chiliennes au sommet du Jurassique et à la base du Crétacé.- Comptes Rendus de l'Académie des Sciences, Paris, t. 271, (Série D), p. 24-27.
P. (1997).- Ostracoda from the subsurface Cretaceous strata of Manhera Tibba well-1 and Shagarh well-B, Jaisalmer Basin, Rajasthan, India with special remarks on foraminiferids.- Geoscience Journal, Lucknow, vol. 18, n° 1, p. 1-27.

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
C = carapace ; VD = valve droite ; VG = valve gauche
1-3. Cytherella DS1 , 1979
1. C, vue latérale gauche (78.06)
2. VG (78.02)
3. C, vue dorsale (78.11)
4-8. Cytherelloidea ghotaruensis , 1997
4. VG (78.05)
5. VD (78.04)
6. VG, juvénile (78.04)
7. VD (78.10)
8. C, vue dorsale (78.04)
9-14. Cytherelloidea oertlii , 1997
9. C, vue latérale gauche (78.07)
10. C, vue latérale droite (78.12)
11. C, vue latérale droite (78.08)
12. VG (78.02)
13. VG (78.09)
14. C, vue dorsale (78.02)
15. Cytherelloidea sp.
C désarticulée (78.154)
16-18. Majungaella cf. pyriformis & , 1969
16. C, vue latérale gauche (78.71)
17. C vue latérale droite (78.60)
18. VG, vue interne (78.75)
19. Paraphysocythere DS1 (, 1979)
C vue latérale droite (78.13)
20-22. Pirileberis makatiniensis , 1984
20. C, vue latérale droite (78.71)
21. VG, vue interne (78.75)
22. C, vue latérale droite (78.75)
23, 23a. Microceratina sp.
23. C, vue latérale droite (78.02)
23a. Idem, détail de l'ornementation zone dorsale
24. Infracytheropteron sp.
C, vue latérale droite (78.04)
25-26. Amphicytherura sp. 5 , & (sous presse)
25. C, vue latérale gauche (78.06)
26. C, vue latérale droite (78.154)

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
C = carapace ; VD = valve droite ; VG = valve gauche
1-3. Cythereis micropapillata nov. sp.
1. C, mâle, vue latérale droite, holotype HCE19 (78.13)
1a. Idem détail ornementation, zone médio-postérieure
1b. idem, détail du réticulum
2. C, femelle, vue latérale gauche, paratype PCE19/1 (78.04)
3. VG, femelle, paratype PCE19/2 (78.06)
4-5. Megommatocythere sp.
4. C, vue latérale gauche (78.04)
5. C, vue latérale droite (78.14)
6-9. Unicapella yvettae nov. sp.
6. C, vue latérale gauche, holotype HAL1 (78.75)
6a. Idem à fort grossissement
7. C, vue latérale gauche, paratype PAL1/1 (78.75)
8. C, vue latérale droite, paratype PAL1/2 (78.71)
9. C, vue latérale droite, paratype PAL1/3 (78.73)
10. Unicapella sp.
C, vue latérale droite (78.73)
11. Dutoitella sp.
C, vue latérale gauche (78.07)
12-19. Taracythere antakaranaensis nov. sp.
12. C, femelle, vue latérale droite, paratype PCE20/1 (78.04)
13. VG, mâle, holotype HCE20 (78.13)
14. C, femelle, vue latérale droite, paratype PCE20/2 (78.13)
15. VG, vue interne, mâle, paratype PCE20/3 (78.13)
16. C, femelle, vue latérale droite, PCE20/4 (78.06)
17. VG, femelle, paratype PCE20/5 (78.13)
18. C, mâle, vue latérale droite, paratype PCE20/6 (78.154)
19. C, vue dorsale, femelle, paratype PCE20/7 (78.13)

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
C = carapace ; VD = valve droite ; VG = valve gauche
1-6. Donmacythere vellicata nov. sp.
1.C ;, vue latérale gauche, holotype HCE21 (78.01)
2. C, vue latérale droite, paratype PCE21/1 (78.01)
3. VD, paratype PCE21/2 (78.01)
4. VG, vue interne, paratype PCE21/3 (78.04)
5. C, vue dorsale, paratype PCE21/4 (78.01)
6. VD, vue interne, paratype PCE21/5 (78.01)
7-10. Parvacythereis ? jamba nov. sp.
7. C, femelle, vue latérale droite, paratype HAL2/1 (78.75)
8. C, mâle, vue latérale droite, paratype HAL2/2 (78.71)
9. C, femelle, vue latérale droite, paratype HAL2/3 (78.60)
10.C, femelle, vue latérale gauche, holotype HAL2 (78.60)
11-17. Glenocythere (Hemiglenocythere) robusta nov. subgen., nov. sp.
11. C, vue latérale droite, holotype HCE22 (78.01)
12. VD, paratype PCE22/1 (78.01)
13. C, vue dorsale, paratype PCE22/2 (78.01)
14. VG, paratype PCE22/3 (78.11)
15. VD, vue interne, paratype PCE22/4 (78.13)
16. VG, vue interne, paratype PCE22/5 (78.01)
16a, Idem, détail des empreintes musculaires
17. VD, vue interne, paratype PCE22/6 (78.11)

Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
C = carapace ; VD = valve droite ; VG = valve gauche
1. Glenocythere (Hemiglenocythere) robusta nov. subgen., nov. sp.
VG, paratype PCE22/7 (78.52)
2-6. Glenocythere (Hemiglenocythere) antsirananaensis nov. subgen., nov. sp.
2. C, femelle, vue latérale droite, paratype PAL3/1 (78.60)
3. Carapace, mâle, vue latérale gauche, paratype PAL3/2 (78.75)
4. C, mâle, vue latérale droite, holotype HAL3 (78.71)
5. C, femelle, vue latérale gauche, paratype PAL3/3 (78.75)
6. VG, vue interne, paratype PAL3/4 (78.75)
7, 8. Spinoleberis ? sp.
7. C, femelle, vue latérale gauche (78.71)
8. C, mâle, vue latérale gauche (78.75)
9. Taracythere DS1 (, 1979)
C, vue latérale gauche (78.10)
10-13. Malagasyella dantelyornata nov. gen, nov. sp.
10. C, vue latérale gauche, holotype HCE23 (78.176)
10a. Idem, détail de l'ornementation, zone postérieure
11. VG, paratype PCE23/1 (78.09)
12. C, vue latérale droite, paratype PCE23/2 (78.176)
12a. idem, détail de la zone antéro-dorsale
13. C, vue dorsale, paratype PCE23/3 (78.176)
14-15. Malagasyella aff. rajendrai (, 1997)
14. C, vue latérale droite (78.176)
14a. Idem, détail de l'ornementation, zone médio-postérieure
15. C, vue dorsale (78.176)
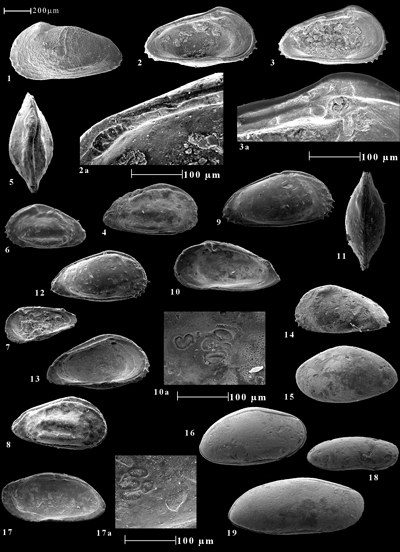
Cliquer sur la vignette pour agrandir l'image.
Click on thumbnail to enlarge the image.
C = carapace ; VD = valve droite ; VG = valve gauche
1-3. Malagasyella aff. rajendrai (, 1997)
1. VG (78.174)
2. VG, vue interne (78.174)
2a. Idem, détail de l'élément cardinal postérieur
3. VG, vue interne (78.168)
3a. Idem, détail de l'élément cardinal antérieur
4-8. Brachycythere ? kharatarensis (, 1997)
4. C, vue latérale droite (78.52)
5. C, vue dorsale (78.75)
6. C, vue latérale gauche (78.05)
7. VD, vue interne (78.02)
8. C, vue latérale droite (78.75)
9-14. Brachycythere pucketti nov. sp.
9. C, vue latérale droite, holotype HCE24 (78.14)
10. VD, vue interne, paratype PCE24/1 (78.14)
10a. Idem, détail des empreintes musculaires
11. C, vue dorsale, paratype PCE24/2.
12. C, vue latérale droite, paratype PCE24/3 (78.07)
13. VG, vue interne, paratype PCE24/4 (78.14)
14. VG, paratype PCE24/5 (78.07)
15-17. Robsoniella postelongata nov. sp.
15. VG, paratype PCE25/1 (78.01)
16. C, vue latérale droite, holotype PCE25 (78.12)
17. VD, vue interne, paratype PCE25/2 (78.13)
17a. Idem, détail des empreintes musculaires
18. Pontocyprella DS1 , 1979
C, vue latérale droite (78.01)
19. Pontocyprella sp.
C, vue latérale droite (78.176)