![]()
Contents
[I. Introduction] [II. Study of Pseudoshasticrioceras bersaci nov. sp.]
[III. Biostratigraphy]
[IV. Conclusion]
[Bibliographic references]
[Figures] and ... [Plates]
![]()
Research in the Feraudianus Subzone of the Sartousiana Zone of the Barremian stage led to the discovery of a new species of Pseudoshasticrioceras: P. bersaci nov. sp. Its study provides evidence concerning the developments of the latest Gassendiceratinae et alii, 2006, and the relationship between the genus Pseudoshasticrioceras , 1998, and Gassendiceras et alii, 2006. In particular, this new species is derived from Pseudoshasticrioceras magnini (, 1992) by a minor revision in the processes of ontogenesis (retardation of ornamentation - neoteny). However, the evolution towards Pseudoshasticrioceras autrani , 1998, implies a "failure" in this process that may possibly be related to parallel changes in environmental conditions. On the other hand, the very closely defined stratigraphic position of Pseudoshasticrioceras bersaci nov. sp., and its position in the anagenetic lineage of Pseudoshasticrioceras, demonstrates its interest as a biostratigraphic marker: a new Bersaci Biohorizon is proposed; it is located between the Magnini and the Autrani biohorizons.
Ammonitinae; Pseudoshasticrioceras; Gassendiceratinae; Upper Barremian; Sartousiana Zone; Feraudianus Subzone; biozonation; southeastern France.
D. & G. (2009).- Pseudoshasticrioceras bersaci nov. sp. (Ammonoidea, Gassendiceratinae), and new ammonite biohorizon for the Upper Barremian of southeastern France.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2009/02 (CG2009_A02)
Description de Pseudoshasticrioceras bersaci nov. sp. (Ammonoidea, Gassendiceratinae), et nouveau biohorizon ammonitique pour le Barrémien supérieur du Sud-Est de la France.- Des recherches dans la sous-zone à Feraudianus ont permis la découverte d'une nouvelle espèce de Pseudoshasticrioceras : Pseudoshasticrioceras bersaci nov. sp. Son étude apporte des éléments concernant l'évolution des derniers Gassendiceratinae et alii, 2006, ainsi que sur les relations entres les genres Pseudoshasticrioceras , 1998, et Gassendiceras et alii, 2006. En particulier cette nouvelle espèce est issue de Pseudoshasticrioceras magnini (, 1992) par des processus d'altération de l'ontogenèse (retardation de l'ornementation - néoténie). En revanche, l'évolution vers Pseudoshasticrioceras autrani , 1998, implique une "rupture" de ce processus qui pourrait éventuellement être mise en parallèle avec les variations des conditions environnementales. D'autre part, la position stratigraphique très précise de Pseudoshasticrioceras bersaci nov. sp., ainsi que sa position dans la lignée anagénétique des Pseudoshasticrioceras, montrent son intérêt en tant que marqueur biostratigraphique : le nouveau biohorizon à Bersaci est proposé ; il est situé entre les biohorizons à Magnini et à Autrani.
Ammonitinae ; Pseudoshascicrioceras ; Gassendiceratinae ; Barrémien supérieur ; Zone à Sartousiana ; Sous-zone à Feraudianus ; biozonation ; Sud-Est de la France
The Upper Barremian was intensively researched as part of the revision of the Hemihoplitidae conducted by one of us (DB). Many ammonites were collected from levels previously reported as being poor in fossils. Some of our results have been published (, 2000; et alii, 2006, 2009). The continuation of this research in the median levels of the Feraudianus Subzone (Sartousiana Zone) have resulted in a better understanding of the dynamics of the ammonite faunas. In particular the collection of a large homogeneous sample of specimens of the genus Pseudoshasticrioceras , 1998, of which a paleontological study is presented here, provides new elements concerning the evolution of the last Gassendiceratinae et alii, 2006, and establishes a new biostratigraphic marker. The biochronological framework used for this work (Upper Barremian) is that proposed recently by et alii (2008).
We had previously recognized (, 2000; et alii, 2006, p. 181) several stages of ornamentation that recur frequently in most Hemihoplitidae , 1924, at the different stages of ontogenic development. These stages are defined as follows:
Systematics:
Order: Ammonoidea , 1884
Sub-ordrer: Ammonitina , 1889
Super-family: Ancyloceratoidea , 1871
Family: Hemihoplitidae ,
1924
Sub-family: Gassendiceratinae et alii,
2006
Genus: Pseudoshasticrioceras ,
1998
Type-species: Pseudoshasticrioceras magnini (, 1992), in (1992, Pl. 24, fig. 1).
Pseudoshasticrioceras bersaci nov. sp.
Synonymy:
? 1964. Crioceratites (Emericiceras) hoheneggeri (, 1883) - , Pl. 5, figs. 2-3.
v 1992. Emericiceras sp. gr. magnini sp. nov. - , Pl. 25, fig. 1a-d.
v 1992. Emericiceras aff. magnini sp. nov. - , Pl. 26, fig. 1; Pl. 27, fig. 1; Pl. 29, figs. 2-3.
Denomination: This species is dedicated to our colleague of Vence (France).
Holotype: Specimen n° AR76, collection
(Pl. 1 ![]() ,
figs. 1-2).
,
figs. 1-2).
Type-locality: The LAC section, in the Méouille area, near Saint-André-les-Alpes (Alpes-de-Haute-Provence, France
Figs. 1 ![]() - 2
- 2 ![]() ) previously illustrated by et alii,
2008,
Fig. 4
) previously illustrated by et alii,
2008,
Fig. 4 ![]() .
.
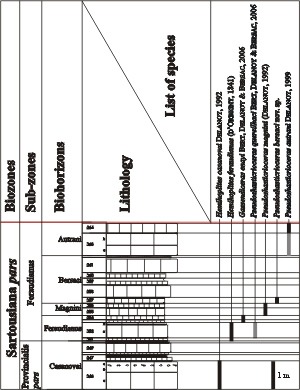
Bed-type: Bed n° 257 from LAC section (Fig. 2 ![]() ).
).
Geographic distribution: Pseudoshasticrioceras bersaci nov. sp. is known to date only in the southeastern France, in both basinal and distal platform areas.
Stratigraphic distribution: All the specimens from the Vocontian Basin come from a bed above the Magnini Biohorizon. This level is taken to be the base of the Bersaci Biohorizon (new, this work). It is located in the middle part of the Feraudianus Subzone (Sartousiana Zone) of the Upper Barremian, between the Magnini and Autrani biohorizons ( et alii, 2008).
Diagnosis: Shell weakly uncoiled; with rapid growth in height, the section always higher than wide. Ornamentation always well marked. Three successive ontogenic stages are known: (1) in the inner whorls, a sub-octagonal section ("heberti" stage); (2) after 25 mm in diameter, the whorls increase in height and the section is compressed further. Ornamentation is trituberculated main ribs, with only a few broad intermediary ribs, which join the main ribs at the top of the flanks (an alternative to the "barremense" stage); (3) up to 100 mm in diameter, compressed oval section with broader base and sides converging towards the venter. Ornamentation blunt with lateral and peri-umbilical tubercles which tend to disappear ("camereiceras" stage).
Material studied (N = 6): Of all the specimens collected, four were from the pelagic domain in the Angles historical stratotype area (Vocontien Basin, area of Saint-André-les-Alpes, Alpes-de-Haute-Provence, southeastern France -
Fig. 1 ![]() ): specimen
n° AM30 and AR76 are from that section bed LAC/257 ( collection); n° 28416 ( collection) and n° MOR 079 ( collection) were collected from the interval between beds 194B and 196 of La Grau de Moriez site (see ,
1997b, p. 233). The other two specimens come from bed 4B4 of the Saint-Laurent de l'Escarène section, Alpes-Maritimes (see ,
1992), which is on a distal platform:
n° EM 108 and EM 109 ( collection) are casts in the 's collection.
): specimen
n° AM30 and AR76 are from that section bed LAC/257 ( collection); n° 28416 ( collection) and n° MOR 079 ( collection) were collected from the interval between beds 194B and 196 of La Grau de Moriez site (see ,
1997b, p. 233). The other two specimens come from bed 4B4 of the Saint-Laurent de l'Escarène section, Alpes-Maritimes (see ,
1992), which is on a distal platform:
n° EM 108 and EM 109 ( collection) are casts in the 's collection.
| n° Specimen | D | H | E | O | H/D | E/D | O/D | E/H | O/H | N/2 | N | spiral gap | α° |
| AR76 (holotype) Pl. 1 |
c. 190 | - | - | - | - | - | - | - | - | - | - | - | |
| 158 | 59.9 | - | 59.6 | 0.38 | - | 0.38 | - | 0.99 | - | - | 0 | ||
| 121.5 | 43.3 | 27.9 | 52.7 | 0.36 | 0.18 | 0.43 | 0.47 | 1.22 | - | - | 0 | 120° | |
| 77.12 | 25.3 | 21 | 33 | 0.33 | 0.17 | 0.43 | 0.48 | 1.30 | 15 | 30 | 0 | 180° | |
| MOR 079 Pl. 3 |
73.2 | 27 | 15.5 | 33.5 | 0.37 | 0.21 | 0.46 | 0.57 | 1.24 | - | - | 2.8 | |
| 45.9 | 15.74 | 7.8 | 21.8 | 0.34 | 0.17 | 0.47 | 0.50 | 1.39 | - | - | 1.15 | 190° | |
| 28416 Pl. 6 |
80.4 | 28.7 | 18.1 | 34.5 | 0.36 | 0.23 | 0.43 | 0.63 | 1.20 | 17 | 36 | 2.8 | |
| 66.2 | 23.3 | 14.3 | 28.0 | 0.35 | 0.22 | 0.42 | 0.61 | 1.20 | 19 | 38 | 2.6 | 70° | |
| 47.8 | 18.2 | 13.1 | 23.4 | 0.38 | 0.27 | 0.49 | 0.72 | 1.29 | 18 | 38 | 1.4 | 90° | |
| EM 108 Pl. 4 Pl. 5 |
189 | 70 | - | 75.4 | 0.37 | - | 0.40 | - | 1.08 | 23 | 43 | 4 | 90° |
| 149.6 | 58.1 | 32 | 58.2 | 0.39 | 0.21 | 0.39 | 0.55 | 1.00 | 23 | 41 | 2.5 | 90° | |
| 115.2 | 43 | 27.7 | 47.2 | 0.37 | 0.24 | 0.41 | 0.64 | 1.10 | 20 | 38 | 2 | 90° | |
| 87 | 32.4 | 23 | 35 | 0.37 | 0.26 | 0.40 | 0.71 | 1.08 | 18 | 36 | 3.5 | 90° | |
| 68 | 24.1 | - | 30 | 0.35 | - | 0.44 | - | 1.24 | 18 | 36 | 2 | 90° | |
| 52.9 | 19.3 | - | 23.4 | 0.36 | - | 0.44 | - | 1.21 | 17 | - | - | ||
| EM 109 Pl. 2 Pl. 3 |
175 | 61.2 | - | 73.2 | 0.35 | - | 0.42 | - | 1.20 | 21 | 38 | 3 | |
| 138.4 | 50.5 | 34 | 55.9 | 0.36 | 0.25 | 0.40 | 0.67 | 1.11 | 20 | 36 | - | 90° | |
| 113.2 | 42.5 | 29 | 45.2 | 0.38 | 0.26 | 0.40 | 0.68 | 1.06 | 17 | - | - | 90° | |
| 88 | 31.54 | 23.5 | 35 | 0.36 | 0.27 | 0.40 | 0.75 | 1.11 | 16 | - | - | 90° | |
| Average | 0.36 | 0.23 | 0.42 | 0.61 | 1.17 | ||||||||
| Standard deviation |
0.01 | 0.04 | 0.03 | 0.09 | 0.11 | ||||||||
| Coefficient
of variation (%) |
4.09 | 16.09 | 7.07 | 15.23 | 9.16 |
Description: Three specimens are more than 175 mm in diameter, but none has an adult body chamber, all are
septate throughout. The shell is weakly uncoiled (sometimes even with sub-joined whorls) and a spiral gap generally
weak. All are less than 3.5 mm in width (average relative umbilicus of O/D = 0.42). The growth in height is rapid
(Table) with an average relative height of H/D = 0.36. In contrast, the growth in thickness remains low, even if the difference in the kind of preservation in samples from the basin and from the platform are taken into account, for the average thickness ratio is only E/D = 0.23. Nevertheless, the section remains much higher than wide (E/H = 0.61 on average) and tends to become greater with growth. In
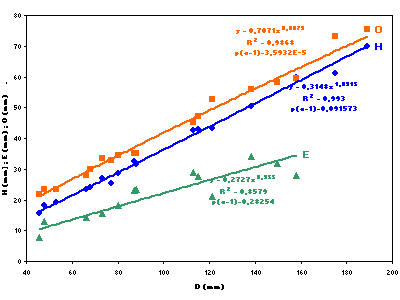
Fig. 3 ![]() , the average curves of the relations between the measured values of H, E and O in relation to the
diameter are in agreement with the allometric relationship Y = bXa. However, for H = f(D) [R2 = 0.99 with p around 0.1] and E = f(D) [R2 = 0.86 with p around 0.3], a
values are close to 1, so we may consider the growth of these characters as almost isometric. Thus relative-height and relative-thickness are fairly constant in relation to the diameter. In contrast, for O = f(D) [R2 = 0.99 and p
< 0001 -
Fig. 3
, the average curves of the relations between the measured values of H, E and O in relation to the
diameter are in agreement with the allometric relationship Y = bXa. However, for H = f(D) [R2 = 0.99 with p around 0.1] and E = f(D) [R2 = 0.86 with p around 0.3], a
values are close to 1, so we may consider the growth of these characters as almost isometric. Thus relative-height and relative-thickness are fairly constant in relation to the diameter. In contrast, for O = f(D) [R2 = 0.99 and p
< 0001 -
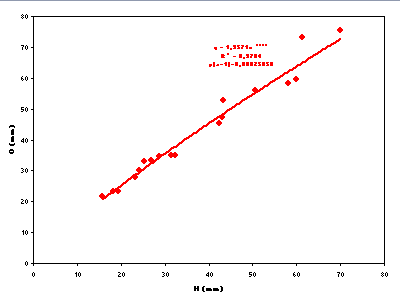
Fig. 3 ![]() ] and O = f(H) [R2 = 0.98 and p
< 0.001 -
Fig. 4
] and O = f(H) [R2 = 0.98 and p
< 0.001 -
Fig. 4 ![]() ], there is a lower value of a. This is linked to the decrease of O/H with respect to the diameter: The progression of this parameter is thus slightly allometric and the average width of the umbilicus increases in proportion with the diameter and the height of the whorl. The analysis of the dispersion parameters reveals that in all cases the clouds of points are weakly scattered around the average, homogeneous and without a breaking slope, thus indicating a harmonic growth. The coefficients of determination R² are still very high. The standard deviations
(Table) are very low and highly correlated with each other. We notice, however, that the coefficients of variation present certain disparities: around 5%, and in all are less than 10%, for reports of H and O calculations. But they are more than 15% for the relations involving E, which is quite high in comparison to values given above. This result was anticipated due to the taphonomic differences: thinner by post mortem compression in the basin and uncompressed, thicker, on the distal platform. In any case there is no bimodality in the clouds of points of these diagrams, thus demonstrating the perfect uniformity of the sample with regard to growth parameters.
], there is a lower value of a. This is linked to the decrease of O/H with respect to the diameter: The progression of this parameter is thus slightly allometric and the average width of the umbilicus increases in proportion with the diameter and the height of the whorl. The analysis of the dispersion parameters reveals that in all cases the clouds of points are weakly scattered around the average, homogeneous and without a breaking slope, thus indicating a harmonic growth. The coefficients of determination R² are still very high. The standard deviations
(Table) are very low and highly correlated with each other. We notice, however, that the coefficients of variation present certain disparities: around 5%, and in all are less than 10%, for reports of H and O calculations. But they are more than 15% for the relations involving E, which is quite high in comparison to values given above. This result was anticipated due to the taphonomic differences: thinner by post mortem compression in the basin and uncompressed, thicker, on the distal platform. In any case there is no bimodality in the clouds of points of these diagrams, thus demonstrating the perfect uniformity of the sample with regard to growth parameters.
The ornamentation is always well marked and increases slightly quantitatively with greater diameter
(Fig. 5 ![]() ). Two extremes
of morphology with respect to robustness (covariance) can be recognized all connected by intermediaries, but that fact does not influence the ontogenic succession. Three successive stages can be recognized during growth.
). Two extremes
of morphology with respect to robustness (covariance) can be recognized all connected by intermediaries, but that fact does not influence the ontogenic succession. Three successive stages can be recognized during growth.
The suture line is very difficult to trace in most of our specimens, especially those from the basin (partial dissolution), but it is fairly denticulate and seems to show a trifide lateral lobe, asymmetric and broad-based (n° EM 109).
Variability: (1966), used 's work
(1892) on the Sonninnidae
(Early Bajocian), that clarified in ammonites the "laws" of covariation based mainly on the relationship between the width of the section and the strength of the ornamentation. Since then, numerous studies have demonstrated that 's "First Law" can be extended to
ammonoids ranging in age from Paleozoic to Cretaceous (see ,
2003, Tab. 1 for a summary). Overall, the sample studied here is very homogeneous, especially in the succession of its ontogenic stages, but using "'s Law" in a more detailed study shows it to be quite variable. Thus, two main morphologies are recognized, based on the relative robustness of the ornamentation. But, because of differences in
both the mode of preservation and the degree of post mortem compression it is not possible
here to demonstrate a correlation between habitat and development of ornamentation as is usually the case. Variability in the
ornamentation density is quite small and very homogeneous
(Fig. 5 ![]() ). At most, the forms with a more robust ornamentation tend to have fewer ribs.
). At most, the forms with a more robust ornamentation tend to have fewer ribs.
In conclusion, the variability in shell and ornament, including all intermediaries, falls within the limits generally accepted for ammonites of the range of variability of a single species.
Differential diagnosis: The sample studied here has all the characters of the genus Pseudoshasticrioceras ,
1998. In particular it is very close to the type-species of the genus, Pseudoshasticrioceras magnini (,
1992), of which one specimen is figured here for comparison
(Pl. 6 ![]() ,
fig. 1). This species differs essentially from ours in the duration of the successive stages of ontogeny. The tuberculated stage of the type "barremense" lasts longer and exists in specimens with an average diameter of up to 100 mm rather than 50 mm (discriminated character). The "heberti" type ontogenic stage is a little longer too.
,
fig. 1). This species differs essentially from ours in the duration of the successive stages of ontogeny. The tuberculated stage of the type "barremense" lasts longer and exists in specimens with an average diameter of up to 100 mm rather than 50 mm (discriminated character). The "heberti" type ontogenic stage is a little longer too.
Compared to Pseudoshasticrioceras quereilhaci et alii, 2006, the differences from our species are more pronounced, for the tuberculated stage of P. quereilhaci is even smaller than in the type-species. In addition, the stage with a high section of the "camereiceras" type occurs later in the growth of our sample.
Pseudoshasticrioceras murphyi (, 1992) is very different from the specimens studied here, for it has no trace of the tuberculated stage of the "barremense" type, at least to when the diameter is greater than 20-25 mm. The ribs are always thinner than those of our specimens.
The adult of Pseudoshasticrioceras autrani , 1998, is smaller (around 100 mm in diameter) and the "barremense" stage, with trituberculated ribs, is smaller than in our specimens (up to 40-55 mm in diameter); also its coiling is narrower.
A comparison of the specimens studied here with other species of the genus shows that the succession of, and especially the duration of, ontogenic stages makes it original morphologically. So we propose the name Pseudoshasticrioceras bersaci nov. sp. to identify it.
More generally, when compared with species of the genus Gassendiceras et alii, 2006, and especially with Gassendiceras quelquejeui et alii, 2006, and Gassendiceras enayi et alii, 2006, the representatives of the genus Pseudoshasticrioceras , 1998, have a number of morphological and ornamentational differences which, despite undeniable affinities, allow their distinction beyond dispute:
Phyletic position and evolution: Stratigraphically (Fig. 2 ![]() ), Pseudoshasticrioceras bersaci nov. sp. immediately follows Pseudoshasticrioceras magnini (,
1992) in the Magnini Biohorizon, and it precedes Pseudoshasticrioceras autrani ,
1998, in the Autrani Biohorizon.
), Pseudoshasticrioceras bersaci nov. sp. immediately follows Pseudoshasticrioceras magnini (,
1992) in the Magnini Biohorizon, and it precedes Pseudoshasticrioceras autrani ,
1998, in the Autrani Biohorizon.
As in many other groups of ammonites, the duration and the appearance of the different stages of ontogeny may be determinants in evolutionary trends ( et alii, 1986). Thus, during time, there is a retardation (neoteny) of the stages of ornamentation between Pseudoshasticrioceras quereilhaci et alii, 2006, Pseudoshasticrioceras magnini (, 1992) and Pseudoshasticrioceras bersaci nov. sp., caused mainly by a lengthening of the trituberculated stage. It should be noted that the process appears to be reversed in the Gassendiceras where there is a reduction in the duration of the "barremense" stage over time (unpublished data).
Although data are still incomplete for these Pseudoshasticrioceras species and for Pseudoshasticrioceras autrani , 1998, there is an abrupt reduction in duration of the "barremense" and "camereiceras" stages, and a sudden decrease in the adult size. This "failure" may parallel some extrinsic elements like the contemporary drop in sea level (sequence boundary SbB4 in , 2005) which preceded the marine transgression at the base of the Giraudi Biozone. These changes caused the morphological distinctions known among the representatives of the genus Imerites , 1933, now classified as Gassendiceratinae et alii, 2006 ( et alii, 2008). New specimens ( et alii, 2009) clearly show a direct link between the latest Pseudoshasticrioceras and the first Imerites.
Bersaci Biohorizon (new): Pseudoshasticrioceras bersaci nov. sp. has a very limited and precise stratigraphic position as the successor, by anagenetic processes, of the index-species Pseudoshasticrioceras magnini (,
1992); so in the stratotype area (Vocontian Basin, southeastern France) its stratigraphic appearance is in conjunction with its biological appearance. This
position and the necessity of establishing a complete high resolution biozone for the Barremian ( et alii,
2006, 2007; et alii,
2008) lead us to propose Pseudoshasticrioceras bersaci nov. sp. as a new biostratigraphic marker, a biohorizon, in the Vocontian Basin of southeastern France (Fig. 6 ![]() ). Pseudoshasticrioceras bersaci nov. sp. characterizes the strata immediately above those of the Magnini Biohorizon. It should be noted that these beds are not distinguishable as such in the historical section of the stratotype near Angles, where deposits of this portion of the Upper Barremian are abnormal (gaps, faunal concentration,
etc. - see et alii, 2008). As already noted by
(1997b), the thickness of the deposits of the Feraudianus
Subzone increases in the direction of Saint-André-les-Alpes, then toward Moriez and Barrême. In the sections near Saint-André-les-Alpes, the first appearance of Pseudoshasticrioceras bersaci nov. sp. closely precedes the major beds of the Autrani Biohorizon ( et alii,
2008), at the top of the Feraudianus Subzone (Sartousiana
Biozone), where Pseudoshasticrioceras autrani ,
1998, and the first recognized Heteroceras coulleti ,
1994, occur.
). Pseudoshasticrioceras bersaci nov. sp. characterizes the strata immediately above those of the Magnini Biohorizon. It should be noted that these beds are not distinguishable as such in the historical section of the stratotype near Angles, where deposits of this portion of the Upper Barremian are abnormal (gaps, faunal concentration,
etc. - see et alii, 2008). As already noted by
(1997b), the thickness of the deposits of the Feraudianus
Subzone increases in the direction of Saint-André-les-Alpes, then toward Moriez and Barrême. In the sections near Saint-André-les-Alpes, the first appearance of Pseudoshasticrioceras bersaci nov. sp. closely precedes the major beds of the Autrani Biohorizon ( et alii,
2008), at the top of the Feraudianus Subzone (Sartousiana
Biozone), where Pseudoshasticrioceras autrani ,
1998, and the first recognized Heteroceras coulleti ,
1994, occur.
Index species: Pseudoshasticrioceras bersaci nov. sp.
Status: This biohorizon is defined by the first appearance of its index-species (bed n° 257 in the LAC section,
Fig. 2 ![]() ), and its upper limit is currently set at the base of the Autrani Biohorizon (bed n° 263 in the LAC section). It is also present in the GRY section, near Angles.
), and its upper limit is currently set at the base of the Autrani Biohorizon (bed n° 263 in the LAC section). It is also present in the GRY section, near Angles.
Faunal assemblages: The index-species, characterized in this work, is generally fairly well represented in the sections. It is associated with the "classic" fauna of the Feraudianus Subzone (, 1997a, 1997b; et alii, 2008): Macroscaphites yvani () macro and microconch, Protetragonites crebisulcatus (), Eulytoceras phestus (), Silesites seranonis (d'), Barremites difficilis (d'), etc.
The discovery of Pseudoshasticrioceras bersaci nov. sp. at a specific level of the Ferraudianus Subzone provides a biostratigraphic marker which can serve as a new biohorizon for the Upper Barremian of southeastern France. This new biohorizon is between the Magnini and the Autrani biohorizons. It is the result of the need for the establishment of the finest possible biostratigraphic framework, to further the study of the different ammonites groups in the Upper Barremian, and especially the Hemihoplitidae (, in progress).
The description of this new species also provided a better understanding of the genus Pseudoshasticrioceras , 1998, and of its differences from the genus Gassendiceras et alii, 2006, morphologically close, and from which it is very probably derived. The stages of ornamentation in these genera do indeed have the same ontogenic organization with the succession of "heberti", "barremense" and "camereiceras" stages. But there are a number of morphological differences (coiling, structure of the shell) and ornamentation (rarer and less differentiated interlayer ribs, and "V" shaped peri-umbilical bifurcations) in the successive stages, especially in the "barremense" type.
This contribution, and the recognition of certain ontogenic heterochronies in its development, has furnished a better understanding of the evolution of Pseudoshasticrioceras. They are an intermediate between the Gassendiceras, with which they occur jointly in the Feraudianus Subzone ( et alii, 2006, 2008), and the Imerites that distinguish the more recent Giraudi Biozone. Further research is necessary to complete this evolutionary canvas, particularly to clarify certain points relationship between P. bersaci nov. sp. and P. autrani , 1998, as well as the precise derivation of the Pseudoshasticrioceras among the Gassendiceras.
We would especially like to thank Mr. Stéphane for all the help he has provided, and Mr. Émile for the loan of their specimens. We also want to sincerely thank François (University of Lyon) and José (University of Granada) for their constructive remarks, and especially Nestor who kindly corrected our English in the final version.
H. (2005).- The South-East France Basin (SFB) and its Mesozoic evolution.- Géologie Alpine, Grenoble, (série spéciale "Colloques et Excursions"), n° 7, p. 5-28.
D. & G. (2000).- Considérations nouvelles sur quelques représentants barrémiens de Puchelliidae , 1890 et des Hemihoplitidae , 1924 (Ammonoidea).- Annales du Muséum d'Histoire Naturelle de Nice, t. XV, p. 63-89.
D., G. & S. (2006).- Descriptions de représentants nouveaux ou peu connus de la Famille des Hemihoplitidae , 1924 (Barrémien supérieur, Sud-Est de la France) : conséquence taxinomique et phylétiques.- Annales du Muséum d'Histoire Naturelle de Nice, t. XXI, p. 179-253.
D., G. & S. (2008).- Propositions pour un nouveau découpage bio-zonal ammonitique, et nouveaux bio-horizons pour le Barrémien supérieur.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2008/03 (CG2008_A03), 18 p.
D., G. & L. (2009).- L'origine des Imerites , 1933 : résultat d'une innovation chez les Gassendiceratinae , 2006 (Ammonoidea, Ancyloceratina).- Annales de Paléontologie, Paris, vol. 95, p. 21-35.
S.S. (1887-1907).- A monograph of the ammonites of the Inferior Oolite Series.- Palaeontological Society, London, 456 p.
G. (1990).- Camereiceras nov. gen. (Ammonoidea, Ancyloceratina) du Barrémien supérieur du Sud-Est de la France.- Geobios, Villeurbanne, n° 23, fasc. 1, p. 71-93.
G. (1992).- Les ammonites du Barrémien supérieur de Saint-Laurent de l'Escarène (Alpes-Maritimes, Sud-Est de la France).- Annales du Muséum d'Histoire Naturelle de Nice, t. IX, 148 p.
G. (1994).- Les biozones à Feraudianus, Giraudi et Sarasini du Barrémien supérieur de la région stratotypique d'Angles-Barrême-Castellane (Sud-Est de la France).- Géologie Alpine, Grenoble, Mémoire H.S., n° 20, p. 279-319.
G. (1997a).- Biostratigraphie des faunes d'Ammonites à la limite Barrémien-Aptien dans la région d'Angles-Barrême-Castellane. Étude particulière de la Famille des Heteroceratidae (Ancyloceratina, Ammonoidea).- Annales du Muséum d'Histoire Naturelle de Nice, t. XII, 270 p.
G. (1997b).- Biostratigraphie haute résolution du Barrémien supérieur du Sud-Est de la France.- Comptes Rendus de l'Académie des Sciences, Paris, (Série II, Sciences de la Terre et des Planètes), t. 325, p. 689-694.
G. (1998).- Pseudoshasticrioceras gen. nov. : un nouveau genre d'ammonite hétéromorphes du Barrémien supérieur du Sud-Est de la France.- Annales du Muséum d'Histoire Naturelle de Nice, t. XIII, p. 431-439.
J.-L., B. & D. (1986).- Les relations ontogenèse-phylogenèse : application paléontologiques.- Geobios, Villeurbanne, n° 19, fasc. 3, p. 335-356.
E. (1884) - Sur un gisement crétacé fossilifère des environs de la gare d'Èze (Alpes-Maritimes).- Bulletin de la Société Géologique de France, Paris, (3ème Série), t. XII, p. 289-300, Pl. IX.
W. & F. (1895).- Sur le Néocomien des environs de Moustiers Ste-Marie (Basses-Alpes).- Bulletin de la Société géologique de France, Paris, (3ème Série), t. XXIII, p. 970-981.
A. & J. (2003).- Ontogeny and covariation in the Toarcian genus Osperleioceras (Ammonoidea).- Bulletin de la Société géologique de France, Paris, t. 174, n° 6, p. 607-615.
S. (reporter), F., D., L., R., G. & J. (2007, unpublished).- Zonation Hauterivien Barrémien.- Compte rendu de la réunion des biostratigraphes français du Group (IUGS Lower Cretaceous Ammonites Working Group), Digne-les-Bains (2 Mai 2007), 14 p.
S. & P.J. (reporters), M.B., P., F., E.Y., M., G., Y., J. J.L., A., V., F.A., I., N., P., J., J.M., Z., J., H., B. & I. (2006).- Report on the second international meeting of the IUGS Lower Cretaceous Ammonite Working Group, the " Group" (Neuchâtel, Switzerland, 8 September 2005).- Cretaceous Research, London, vol. 27, n° 5, p. 712-715.
S. & J. (reporters), R., M., C., A., S.N., J., O., J.M., Z. & J. (2009).- Report on the 3rd International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the " Group" (Vienna, Austria, 15th April 2008).- Cretaceous Research, London, vol. 30, n° 2, p. 496-502.
J. (1933).- Les ammonites aptiennes de la Géorgie occidentale.- Bulletin de l'Institut géologique de Géorgie (1932), Tbilisi, vol. 1, n° 3, p. 165-273.
L.F. (1924).- On the ammonites of the Speeton Clay and the subdivisions of the Neocomian.- Geological Magazine, Cambridge, vol. 61, n° 716, p. 73-89.
G. (1964).- Contribution à la connaissance des céphalopodes crétacés du Sud-Est de la France. Note sur les ammonites déroulées du Crétacé inférieur vocontien.- Mémoires de la Société géologique de France, Paris, (Nouvelle Série), t. XXXVIII, Mémoire n° 101, 80 p.
V. (1887).- Ueber neocom Fossilien vom Gardenazza in Südtirol nebst einen Anhang über das Neocom von Ischl.- Jahrbuch der kaiserlich-königlichen geologischen Reichsanstalt, Wien, Band 37, Heft 1, p. 69-108.
G.E.G. (1966).- Covariation and taxonomy of the Jurassic ammonite Soninia adicra ().- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, Band 124, Heft 3, p. 289-312.
Click on thumbnail to enlarge the image.
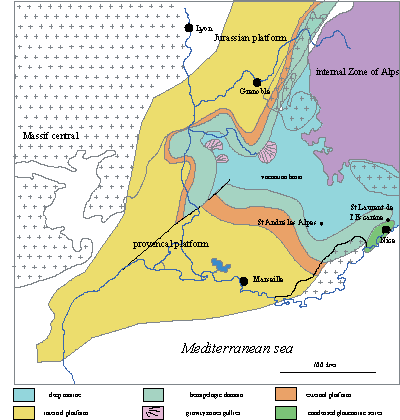
Figure 1: Barremian paleogeography of southeastern France and location of the area studied (from ,
2005).
Figure 1 : Cadre paléogéographique du Sud-Est de la France et localisation du secteur d'étude (d'après ,
2005).
Click on thumbnail to enlarge the image.
Figure 2: Distribution of the ammonites in the LAC section pars
(Méouille area, Saint-André-les-Alpes, Alpes-de-Haute-Provence). In grey are the specimens found in stratigraphically equivalent strata (outside the main section).
Figure 2 : Répartition des faunes dans la coupe LAC pars (secteur de Méouille, Saint-André-les-Alpes, Alpes-de-Haute-Provence). En grisé, les espèces trouvées en place latéralement dans d'autres coupes, mais dans des niveaux corrélables banc par banc.
Click on thumbnail to enlarge the image.
Figure 3: Representation of H, E and O in function of the diameter (D) for Pseudoshasticrioceras bersaci
nov. sp.
Figure 3 : Représentation de H, E et O en fonction du diamètre (D) pour Pseudoshasticrioceras bersaci nov. sp.
Click on thumbnail to enlarge the image.
Figure 4:
Representation of the umbilicus (O) in function of whorl height (H) for Pseudoshasticrioceras bersaci
nov. sp.
Figure 4 : Représentation de l'ombilic (O) en fonction du de la hauteur du tour (H) pour Pseudoshasticrioceras bersaci nov. sp.
Click on thumbnail to enlarge the image.
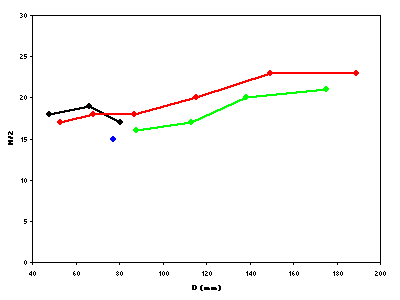
Figure 5:
Representation in the number of ribs per half-turn of whorl (N/2) depending on the diameter (D) for Pseudoshasticrioceras bersaci
nov. sp.
Figure 5 : Représentation du nombre de côtes par demi-tour de spire (N/2) en fonction du diamètre (D) pour Pseudoshasticrioceras bersaci nov. sp.
Click on thumbnail to enlarge the image.
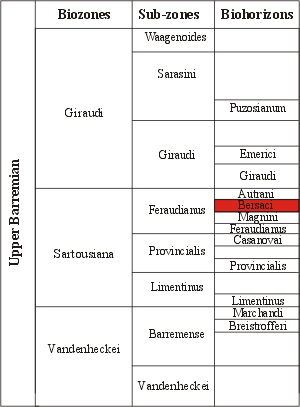
Figure 6:
Proposed biozonal scheme, after et alii,
2008, and et alii,
2009, amended. In red the Bersaci Biohorizon (new).
Figure 6 : Biozonation proposée, d'après et alii,
2008, et et alii,
2009, modifié. En rouge le
Biohorizon à Bersaci (nouveau).

![]()
Click on thumbnail to enlarge the image.
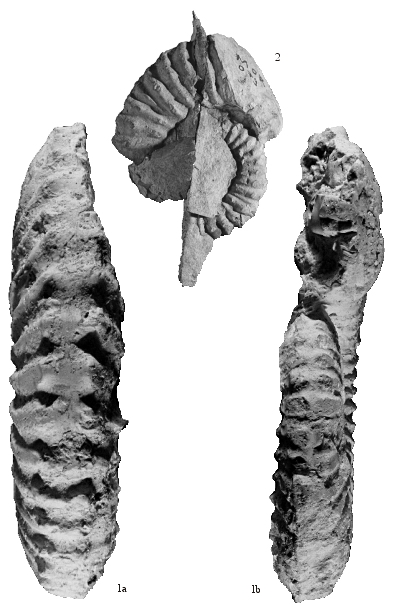
Pseudoshasticrioceras bersaci nov. sp.
Figs. 1-2: Specimen n° AR76 (holotype), collection, from the bed LAC/257, Méouille near Saint-André-les-Alpes (Alpes-de-Haute-Provence, France).
Planche 1 : Figs. 1-2 : Spécimen n° AR76 (holotype), collection , banc LAC/257 de Méouille près de Saint-André-les-Alpes (Alpes-de-Haute-Provence, France).

![]()
Click on thumbnail to enlarge the image.
Pseudoshasticrioceras bersaci nov. sp.
Fig. 1: Specimen n° EM 109, collection, from the bed 4B4 of the deposit of Saint-Laurent de l'Escarène (Alpes-Maritimes, France).
Planche 2 : Fig. 1 : Spécimen n° EM 109, collection , banc 4B4 de Saint-Laurent de l'Escarène (Alpes-Maritimes, France).
![]()
Click on thumbnail to enlarge the image.
Pseudoshasticrioceras bersaci nov. sp.
Fig. 1a-b: Specimen n° EM 109, collection, from the bed 4B4 of the deposit of Saint-Laurent de l'Escarène (Alpes-Maritimes, France).
Fig. 2: Specimen n° MOR 079, collection, from the gap of beds 194B to 196 of the deposit of La Grau de Moriez (Alpes-de-Haute-Provence, France).
Planche 3 : Fig. 1a-b : Spécimen n° EM 109, collection , banc 4B4 de Saint-Laurent de l'Escarène
(Alpes-Maritimes, France).
Fig. 2 : Spécimen n° MOR 079, collection , de l'intervalle de bancs 194B à 196 de La Grau de Moriez (Alpes-de-Haute-Provence, France).

![]()
Click on thumbnail to enlarge the image.
Pseudoshasticrioceras bersaci nov. sp.
Fig. 1: Specimen n° EM 108, collection, from the bed 4B4 of the deposit of Saint-Laurent de l'Escarène (Alpes-Maritimes, France).
Planche 4 : Fig. 1 : Spécimen n° EM 108, collection , banc 4B4 de Saint-Laurent de l'Escarène (Alpes-Maritimes, France).

![]()
Click on thumbnail to enlarge the image.
Pseudoshasticrioceras bersaci nov. sp.
Fig. 1a-b: Specimen n° EM 108, collection, from the bed 4B4 of the deposit of Saint-Laurent de l'Escarène (Alpes-Maritimes, France).
Fig. 2: Specimen n° 28416, collection, from the gap of beds 194B to 196 of the deposit of La Grau de Moriez (Alpes-de-Haute-Provence, France).
Planche 5 : Fig. 1a-b : Spécimen n° EM 108, collection , banc 4B4 de Saint-Laurent de l'Escarène
(Alpes-Maritimes, France).
Fig. 2 : Spécimen n° 28416, collection , de l'intervalle de bancs 194B à 196 de La Grau de Moriez (Alpes-de-Haute-Provence, France).

![]()
Click on thumbnail to enlarge the image.
Plate 6:
Planche 6 :
Pseudoshasticrioceras magnini (, 1992)
Fig. 1: Specimen n° LAC 032, collection, from bed LAC/256, Méouille near Saint-André-les-Alpes (Alpes-de-Haute-Provence, France).
Fig. 1 : Spécimen n° LAC 032, collection , banc LAC/256 de Méouille près de Saint-André-les-Alpes (Alpes-de-Haute-Provence, France).
Pseudoshasticrioceras bersaci nov. sp.
Fig. 2a-b: Specimen n° 28416, collection, from the gap of beds 194B to 196 of the deposit of La Grau de Moriez (Alpes-de-Haute-Provence, France).
Fig. 2a-b : Spécimen n° 28416, collection , de l'intervalle de bancs 194B à 196 de La Grau de Moriez (Alpes-de-Haute-Provence, France).